Embed Size (px)
Citation preview

N E W S A N D V I E W S
NATURE GENETICS VOLUME 35 | NUMBER 4 | DECEMBER 2003 303
oocyte expression system, Kahle et al.3 nowshow that WNK4 also inhibits ROMK activityby stimulating clathrin-dependent endocytosisof ROMK, a mechanism distinct from WNK4inhibition of NCCT. Notably, the mutations inPRKWNK4 that cause PHAII further stimulatethe retrieval of ROMK from the cell surface,which may help explain the impaired potas-sium secretion and consequent hyperkalemiaobserved in PHAII. Together, these observa-tions establish the reciprocal effect of muta-tions in PRKWNK4 that cause PHAII on therouting of NCCT and ROMK to the plasmamembrane. Thus, WNK4 is a novel dual regu-lator that balances both sodium reabsorptionand potassium secretion in the distal part ofthe nephron, a site where the final compositionof the urine is determined.
A WNK and a nodIn light of the identified actions of wild-typeand mutant WNK4, we may now be able toexplain at the molecular level the nephron’sability to control independently sodium-chloride reabsorption and potassium secre-tion in response to varying conditions.Several physiologically important roles cannow be envisioned (Fig. 1). Aldosterone is aprimary regulator of both sodium-chloridereabsorption and potassium secretion. Thebaseline condition resembles a situation of lowaldosterone levels in which WNK4 continu-ously suppresses the routing of both trans-porters, resulting in only a modest number ofactive transport proteins at the plasma mem-brane. In the presence of substantial levels of
aldosterone, two extreme situations can occur.In hypovolemic states (reduced blood vol-ume), the inhibition of NCCT by WNK4 isrelieved, whereas the routing of ROMK to theplasma membrane is abolished, allowing max-imal sodium-chloride reabsorption. Similarly,during conditions of hyperkalemia, the inhibi-tion of ROMK is lessened, whereas the routingof NCCT is blocked, maximally stimulatingpotassium secretion. Thus, this model predictsa differentiating role of WNK4 in aldosterone-regulated sodium and potassium homeostasis.
Intriguing questionsSeveral outstanding issues can now beaddressed experimentally to improve ourunderstanding of the bifunctional role ofWNK4 in general and in PHAII in particular.For instance, the subcellular localization ofWNK4 to intercellular junctions and cyto-plasmic compartments hints at additional,unknown functions along the nephron. Theclinical features of PHAII are chloride-dependent. Infusion of sulfate as an anionreplacing chloride corrects the elevated levelsof potassium in the urine (hyperkaliurea)observed in PHAII (ref. 10). Together withthe conspicuous localization of WNK4 in thetight junction complex, this suggests thatWNK4 could regulate the paracellular chlo-ride conductance. In addition, the dominantmode of inheritance of PHAII is difficult toreconcile with the experimental data obtainedso far, suggesting that the situation is morecomplex than presently envisioned or that theX. laevis oocyte expression system may not
reproduce an effect that occurs in the humankidney. Thus, further studies in polarizedepithelial cells are certainly warranted to fur-ther substantiate the (patho)physiologicalrole of WNK4.
Finally, many details of the mechanism ofaction of WNK4 have yet to be addressed.WNK4 contains several potential regulatorysites that illuminate the signaling pathwaysthat can now be explored. The catalytic kinasedomain in WNK4 indicates that phosphoryla-tion of downstream target proteins could havean important regulatory role. Initial studiesindeed suggest that WNK4 inhibits NCCT in akinase-dependent manner, but the inhibitionof ROMK is kinase-independent, suggestingadditional pathways3,8. Thus, further work isnecessary to determine conclusively all thefunctions of the regulatory domains inWNK4. The outlined studies again illustratethe importance of unraveling the moleculardefects of rare genetic diseases, such as PHAII,to establish new concepts in the physiology ofrenal salt and water homeostasis.
1. Giebisch, G.H. Kidney Int. 62, 1498–1512 (2002).2. Rossier, B.C. J. Clin. Invest. 111, 947–950 (2003).3. Kahle, K.T. et al. Nat. Genet. 35, 372–376 (2003).4. Xu, B. et al. J. Biol. Chem. 275, 16795–16801
(2000).5. Verissimo, F. & Jordan, P. Oncogene 20, 5562–5569
(2001).6. Manning, G. et al. Science 298, 1912–1914 (2002).7. Wilson, F.H. et al. Science 293, 1107–1112 (2001).8. Wilson, F.H. et al. Proc. Natl. Acad. Sci. USA 100,
680–684 (2003).9. Yang, C.L. et al. J. Clin. Invest. 111, 1039–1045
(2003).10. Schambelan, M. et al. Kidney Int. 19, 716–727
(1981).
Free energy lights the path toward more effective RNAiJose M Silva, Ravi Sachidanandam & Gregory J Hannon
Silencing gene expression by RNA interference (RNAi) has become a daily activity for mammalian cell biologists.Those who often use this approach have come to appreciate its occasional frustrations and potential limitations. Apair of recent papers in Cell provides rules for designing short interfering RNAs (siRNAs) that are both validated atthe practical level and understood mechanistically.
The discovery that double-stranded RNA canelicit RNAi, a specific, potent gene silencingresponse, began a genetic revolution in those
organisms for which genetic approaches werelargely unavailable or inconvenient. Generally,the interference process initiates with the pro-cessing of a double-stranded RNA by Dicer(reviewed in ref. 1). This RNaseIII-classenzyme creates siRNAs, which compriseduplexes of ∼ 19 nucleotides (nt) bounded by2-nt 3′ overhangs. The siRNAs become incor-porated into the RNA-induced silencing com-plex (RISC), which uses such sequences as
guides to the selection of its substrates byWatson-Crick base-pairing interactions.Thus, siRNAs must be unwound for exposureto potential targets, which is thought to bemediated by an ATP-dependent RNA heli-case2. Single-stranded versions of siRNAs canbe used to trigger RISC assembly in vitro inboth Drosophila melanogaster and mam-malian cell extracts, although they are less effi-cient triggers than double-stranded siRNAs3,4.
Jose M. Silva, Ravi Sachidanandam andGregory J. Hannon are at Cold Spring HarborLaboratory, Watson School of BiologicalSciences, 1 Bungtown Road, Cold SpringHarbor, New York 11724, USA.e-mail: [email protected]
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reg
enet
ics
N E W S A N D V I E W S
304 VOLUME 35 | NUMBER 4 | DECEMBER 2003 NATURE GENETICS
Tuschl and colleagues5 and Caplen andcoworkers6 first found that siRNAs them-selves could be used to trigger RNAiresponses in mammalian cells. In practice,the approach can be made to work well,achieving reduction of target mRNAs by>90% after transfection of cells with syn-thetic siRNAs. But achieving this goal oftenrequires screening many siRNAs beforetruly effective inhibitors are discovered.Two papers published recently in Cellpromise to improve the success rate of theapproach through the discovery of real,mechanism-based guidelines for siRNAselection7,8.
Eliminate free energy!Zamore and colleagues arrived at theirinsights into siRNA design by solving a mys-tery that emerged from their ongoing in vitrostudies of RISC biochemistry8. They firstcompared single-stranded and double-stranded siRNAs and their respective abilitiesto direct the cleavage of either sense- or anti-sense-oriented luciferase target RNAs. Bothstrands of the siRNA worked equally wellwhen presented to the extract as singlestrands. With double-stranded molecules,however, there was profound strand bias inability to direct target cleavage. Thus, the twostrands did not differ significantly in their
intrinsic efficacy but the ability of individualstrands to silence, in vitro, differed if thestrands were presented in double-strandedform. In a novel RISC-capture assay, strandasymmetry was apparent, with the antisensestrand (with respect to the coding orienta-tion of the luciferase mRNA) showing bothbetter incorporation into RISC and moreefficient cleavage for this particular siRNA.
The authors’ examination of the sequenceof the siRNA identified one immediatelyobvious difference between the two terminiof this molecule. The 5′ end of the strandthat was incorporated into RISC more effi-ciently had a lower stability and began withan A-U pair, whereas the less effectivelyincorporated strand had a G-C terminus.This led to the hypothesis that RISC prefer-entially accepts the strand of the siRNA thatpresents the less stable 5′ end. To test thisnotion, Zamore and colleagues madenumerous alterations to a series of siRNAsthat target sod1 in vitro. By altering the freeenergy of base pairing at the ends (by creat-ing a mismatch or just changing the bases),and also throughout the neighboring threenucleotides by mismatches, the effectivestrand of the siRNA could be manipulatedin a predictable manner.
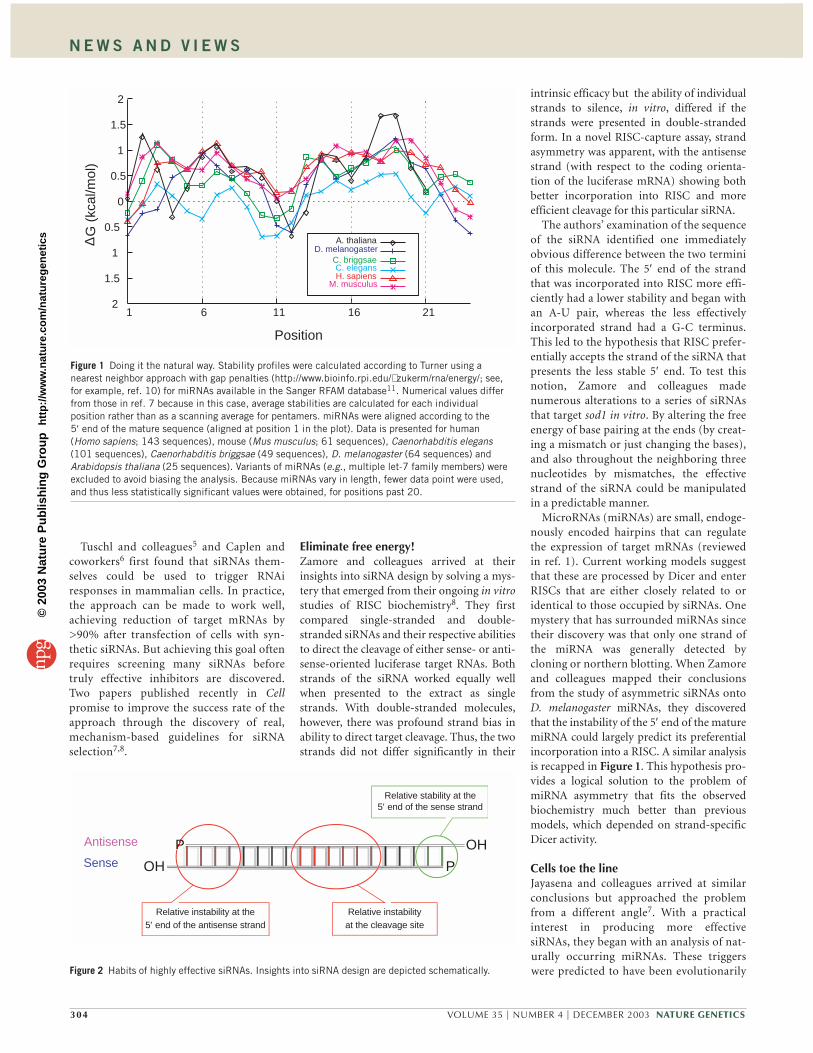
MicroRNAs (miRNAs) are small, endoge-nously encoded hairpins that can regulatethe expression of target mRNAs (reviewedin ref. 1). Current working models suggestthat these are processed by Dicer and enterRISCs that are either closely related to oridentical to those occupied by siRNAs. Onemystery that has surrounded miRNAs sincetheir discovery was that only one strand ofthe miRNA was generally detected bycloning or northern blotting. When Zamoreand colleagues mapped their conclusionsfrom the study of asymmetric siRNAs ontoD. melanogaster miRNAs, they discoveredthat the instability of the 5′ end of the maturemiRNA could largely predict its preferentialincorporation into a RISC. A similar analysisis recapped in Figure 1. This hypothesis pro-vides a logical solution to the problem ofmiRNA asymmetry that fits the observedbiochemistry much better than previousmodels, which depended on strand-specificDicer activity.
Cells toe the lineJayasena and colleagues arrived at similarconclusions but approached the problemfrom a different angle7. With a practicalinterest in producing more effectivesiRNAs, they began with an analysis of nat-urally occurring miRNAs. These triggerswere predicted to have been evolutionarily
Figure 1 Doing it the natural way. Stability profiles were calculated according to Turner using anearest neighbor approach with gap penalties (http://www.bioinfo.rpi.edu/∼ zukerm/rna/energy/; see,for example, ref. 10) for miRNAs available in the Sanger RFAM database11. Numerical values differfrom those in ref. 7 because in this case, average stabilities are calculated for each individualposition rather than as a scanning average for pentamers. miRNAs were aligned according to the5′ end of the mature sequence (aligned at position 1 in the plot). Data is presented for human(Homo sapiens; 143 sequences), mouse (Mus musculus; 61 sequences), Caenorhabditis elegans(101 sequences), Caenorhabditis briggsae (49 sequences), D. melanogaster (64 sequences) andArabidopsis thaliana (25 sequences). Variants of miRNAs (e.g., multiple let-7 family members) wereexcluded to avoid biasing the analysis. Because miRNAs vary in length, fewer data point were used,and thus less statistically significant values were obtained, for positions past 20.
2
1.5
1
0.5
0
0.5
1
1.5
2
1 6 11 16 21
D. melanogasterC. briggsae
M. musculusH. sapiensC. elegans
Position
G (
kcal
/mol
)∆ A. thaliana
Figure 2 Habits of highly effective siRNAs. Insights into siRNA design are depicted schematically.
Relative instability at the5′ end of the antisense strand
Relative stability at the5′ end of the sense strand
Relative instabilityat the cleavage site
Antisense
SensePPPP
OH
OH
P
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reg
enet
ics

N E W S A N D V I E W S
through asymmetry in what might superfi-cially appear as a symmetric siRNA. In thefinal tally, the reports reviewed here make aclear case that with RNAi, the best way towin the game is to play by the rules.
1. Hannon, G.J. Nature 418, 244–251 (2002).2. Nykanen, A., Haley, B. & Zamore, P.D. Cell 107,
309–321 (2001).3. Schwarz, D.S., Hutvagner, G., Haley, B. & Zamore,
P.D. Mol. Cell 10, 537–548 (2002).4. Martinez, J., Patkaniowska, A., Urlaub, H.,
Luhrmann, R. & Tuschl, T. Cell 110, 563–574(2002).
5. Elbashir, S.M. et al. Nature 411, 494–498 (2001).6. Caplen, N.J., Parrish, S., Imani, F., Fire, A. &
Morgan, R.A. Proc. Natl. Acad. Sci. USA 98,9742–9747 (2001).
7. Khvorova, A., Reynolds, A. & Jayasena, S.D. Cell 115,209–216 (2003).
8. Schwarz, D.S. et al. Cell 115, 199–208 (2003).9. Lee, Y. et al. Nature 425, 415–419 (2003).10. Turner, D.H. Curr. Opin. Struct. Biol. 6, 299–304
(1996).11. Griffiths-Jones, S., Bateman, A., Marshall, M.,
Khanna, A. & Eddy, S.R. Nucleic Acids Res. 31,439–441 (2003).
NATURE GENETICS VOLUME 35 | NUMBER 4 | DECEMBER 2003 305
selected for the ability to regulate targetseffectively. These investigators noted thatthe average stability of the miRNA was notrandom along its length (Fig. 1). MaturemiRNAs were unstable at their 5′ ends.Internal stability was also lower near thecenter of the miRNA (around positions9–11). Notably, an examination of 37siRNAs targeting several different genesshowed that effective sequences (16 of 37)matched the miRNA-derived patternswhereas ineffective sequences (21 of 37) didnot. They obtained additional consistentdata with a wider panel of 180 siRNAs tar-geting a region of firefly luciferase.
The bias in strand-specific cleavage byRISC was not tested in vivo. But siRNAsmatching the predicted profiles accumulatedpreferentially from an expressed, long dou-ble-stranded RNA trigger in tobacco. Thisobservation matches the proposal that
strand-specific incorporation into RISCdetermines both the stability and, to a largedegree, the efficacy of siRNAs, irrespective ofwhether they are synthetic RNAs that areexperimentally introduced or are generatedby Dicer in vivo.
The combination of in vitro and in vivostudies provide complementary insightsinto the design of effective siRNAs (sum-marized in Fig. 2). These insights will prob-ably also guide the design of more effectiveexpressed silencing triggers, which enter theRNAi pathway by mimicking miRNAs. Ofcourse, this achievement will depend on adetailed understanding of how syntheticmiRNAs give rise to specific siRNAsequences in vivo through the actions ofDicer and Drosha9. Finally, the studieshighlighted here shed light on the mecha-nisms of RISC assembly, suggesting that in-built structural cues create specificity
©20
03 N
atu
re P
ub
lish
ing
Gro
up
h
ttp
://w
ww
.nat
ure
.co
m/n
atu
reg
enet
ics







![Drosophila: RNAi and Non-RNAi - biblio.ugent.be · subsequently in Drosophila [3] and Caenorhabditis elegans [4]. In Drosophila, the major RNAi pathway involved in antiviral immunity](https://img.dokumen.tips/doc/110x75/5ccc3ff188c99335448bc823/drosophila-rnai-and-non-rnai-subsequently-in-drosophila-3-and-caenorhabditis.jpg)





















