Embed Size (px)
Citation preview

Formation of Complex ExtrachromosomalT-DNA Structures in Agrobacteriumtumefaciens-Infected Plants1[C][W][OA]
Kamy Singer*, Yoel M. Shiboleth, Jianming Li, and Tzvi Tzfira2
Department of Molecular, Cellular, and Developmental Biology, University of Michigan, Ann Arbor, Michigan48109–1048
Agrobacterium tumefaciens is a unique plant pathogenic bacterium renowned for its ability to transform plants. The integration oftransferred DNA (T-DNA) and the formation of complex insertions in the genome of transgenic plants during A. tumefaciens-mediated transformation are still poorly understood. Here, we show that complex extrachromosomal T-DNA structures form inA. tumefaciens-infected plants immediately after infection. Furthermore, these extrachromosomal complex DNA molecules cancircularize in planta. We recovered circular T-DNA molecules (T-circles) using a novel plasmid-rescue method. Sequencinganalysis of the T-circles revealed patterns similar to the insertion patterns commonly found in transgenic plants. The patternsinclude illegitimate DNA end joining, T-DNA truncations, T-DNA repeats, binary vector sequences, and other unknown “filler”sequences. Our data suggest that prior to T-DNA integration, a transferred single-stranded T-DNA is converted into a double-stranded form. We propose that termini of linear double-stranded T-DNAs are recognized and repaired by the plant’s DNAdouble-strand break-repair machinery. This can lead to circularization, integration, or the formation of extrachromosomalcomplex T-DNA structures that subsequently may integrate.
The soil pathogen Agrobacterium tumefaciens geneti-cally transforms plants. This natural and unique abilityof transkingdom gene transfer has made A. tumefaciens-mediated transformation one of the most importanttools in plant biology research and the preferredmethod for producing transgenic plants. The trans-formation process is initiated by the transfer of asingle-stranded (ss) DNA segment derived from theA. tumefaciens tumor-inducing (Ti) plasmid into theplant cell nucleus. This transferred DNA (T-DNA) isdelimited by two 25-bp direct imperfect repeats knownas the right border (RB) and left border (LB), which arerequired for its transfer. The process also requires a setof bacterial chromosomal (chv) and Ti plasmid viru-lence (vir) genes as well as various host genes (for
review, see McCullen and Binns, 2006; Citovsky et al.,2007; Gelvin, 2010; Tzfira and Citovsky, 2010). Innatural A. tumefaciens strains, the T-DNA carries thegenes responsible for the formation of tumor growthon plants, known as the “crown gall” disease. In dis-armed laboratory strains, the native T-DNA is re-moved from the Ti plasmid. Instead, a gene of interestcan be placed between the two T-DNA borders and betransferred from a smaller binary plasmid (for review,see Gelvin, 2003; Tzfira and Citovsky, 2006).
To study the last step in the transformation process,T-DNA integration into the plant genome, T-DNA in-sertion events have been extensively characterizedby sequencing of the T-DNA/chromosome junctions.These studies have shown that T-DNAs are insertedrandomly throughout the plant genome (Forsbach et al.,2003; Kim et al., 2007). They have also shown that theintegration is “illegitimate” (i.e. not sequence specific)but may include overlapping microhomologies of ap-proximately 2 to 7 bp (Gheysen et al., 1991; Mayerhoferet al., 1991). Furthermore, T-DNA integration can resultin complex structures. These may include truncations ofT-DNA ends and multicopy T-DNAs arranged asinverted or direct repeats (Kwok et al., 1985; Spielmannand Simpson, 1986). Complex structures may also in-clude non-T-DNA bacterial sequences (Martineau et al.,1994; Ulker et al., 2008), DNA from an unknown source(“filler”), and plant sequence duplications (Gheysenet al., 1987, 1991; Mayerhofer et al., 1991). More recentstudies have further characterized these integrationpatterns under different experimental settings, thusproviding more insight into the transformation process(Kumar and Fladung, 2002; Meza et al., 2002; Stahl
1 The work was supported by the U.S.-Israel Binational Agricul-tural Research and Development Fund (grant no. BARD US–4150–08to T.T.) and the U.S.-Israel Binational Science Foundation (grant no.2007140 to T.T.).
2 Present address: Department of Life Sciences, Ben-Gurion Uni-versity of the Negev, Beer Sheva 84105, Israel.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Tzvi Tzfira ([email protected]).
[C] Some figures in this article are displayed in color online but inblack and white in the print edition.
[W] The online version of this article contains Web-only data.[OA] Open Access articles can be viewed online without a subscrip-
tion.www.plantphysiol.org/cgi/doi/10.1104/pp.112.200212
Plant Physiology�, September 2012, Vol. 160, pp. 511–522, www.plantphysiol.org � 2012 American Society of Plant Biologists. All Rights Reserved. 511 www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from
Copyright © 2012 American Society of Plant Biologists. All rights reserved.

et al., 2002; Forsbach et al., 2003; Kim et al., 2003;Windels et al., 2003; Thomas and Jones, 2007; Zhanget al., 2008; De Buck et al., 2009). However, the mecha-nism of T-DNA integration is still poorly understood (forreview, see Tzfira et al., 2004; Ziemienowicz et al.,2010).
Although complex T-DNA insertions are undesired intransgenic plants for commercial or research purposes,they are a relatively frequent outcome of transformation(Windels et al., 2003, 2010; Zhang et al., 2008). Under-standing how complex T-DNA insertions form is im-portant to better understand the mechanism behindT-DNA integration. De Neve et al. (1997) proposed thatcomplex T-DNA structures, such as those that involveT-DNA repeats, formwhen two or more double-stranded(ds) T-DNA intermediates ligate in the plant nucleusprior to integration. In contrast, other models proposethat T-DNA repeats form via ss T-DNA intermediatesthat ligate during (Krizkova and Hrouda, 1998) or priorto (Stahl et al., 2002) integration. Whether T-DNA inte-grates as an ss or ds intermediate is a fundamentalquestion related to T-DNA integration. According to themodel of Tinland (1996), conversion of ss T-DNA into dsT-DNA occurs only during its incorporation to the ge-nome. Similar ss-based models have been proposed(Brunaud et al., 2002; Kumar and Fladung, 2002; Mezaet al., 2002; Thomas and Jones, 2007; Teo et al., 2011).Conversely, the ds T-DNA model suggests that con-version to ds T-DNA occurs in plants prior to integra-tion (Mayerhofer et al., 1991). This is supported byevidence that at least some of the nonintegrating “free”T-DNA molecules in infected plants are ds (Janssen andGardner, 1990; Offringa et al., 1990; Narasimhulu et al.,1996). Also supporting the ds T-DNA model are datasuggesting that ds T-DNA integrates into genomicdouble-stranded breaks (DSBs; Salomon and Puchta,1998; Chilton and Que, 2003; Tzfira et al., 2003).
While a common approach to studying the mecha-nism of T-DNA integration is to characterize patternsof postintegration events in plants, analyzing T-DNAtransfer events in plants prior to integration offers aneffective alternative approach. Bakkeren et al. (1989)described a virus-based infection system to recovertransfer events indirectly. That method is based on thehypothesis that ds T-DNAs occasionally circularize inplanta. However, the experimental setup of that indi-rect virus-based system, which may have resultedfrom recombination by viral components within theT-DNA molecules, did not allow the detection of po-tential complex T-DNA structures.
To study the formation of complex T-DNA struc-tures in plants, we developed a novel method to di-rectly isolate transfer events from infected plants. Themethod adopts a plasmid-rescue approach to recoverds T-DNA molecules that have undergone in plantacircularization. When sequencing these molecules, wediscovered various complex circular structures withpatterns similar to those previously reported in trans-genic plants. This suggests that complex extrachro-mosomal ds T-DNA structures may form in plants
prior to integration via ligation between ds DNA in-termediates. We propose that some of these ds DNAextrachromosomal complex structures may then inte-grate into the genome.
RESULTS
The Experimental Approach
To test the extrachromosomal T-DNA ligation modelfor the formation of complex T-DNA structures (De Neveet al., 1997), we hypothesized that extrachromosomalcomplex T-DNA molecules will occasionally circularizevia the same DNA-repair pathway responsible for theirproposed formation. To recover complex circular T-DNAstructures potentially forming in A. tumefaciens-infectedplants, we developed a direct plasmid-rescue method inwhich circular T-DNA molecules (T-circles) are capturedas plasmids in Escherichia coli.
To this end, we constructed AMP-ORI and KAN-ORI T-DNAs containing a pBR322 origin of replication(ori) for E. coli and a bacterial selectable marker, eitherthe ampicillin resistance gene (AmpR; bla gene) or thekanamycin resistance gene (KanR; aph gene; Fig. 1A;Supplemental Fig. S1). A KAN T-DNA construct thatdoes not contain an ori was also prepared for ad-ditional experiments (Fig. 2). The T-DNA constructswere cloned into a pRCS2 binary plasmid (Tzfiraet al., 2005). The backbone of this binary plasmid con-tains the bacterial spectinomycin and streptomycinresistance gene (SpR/SmR; aadA gene) and the pVS1ori for DNA replication in A. tumefaciens and E. coli(Fig. 1B, 1). The AMP-ORI and KAN-ORI constructswere introduced separately into plant leaves viaagroinfiltration (Fig. 1B, 2). At 4 to 8 d post agro-infiltration, we extracted DNA from the infected leaves(Fig. 1B, 3) and transformed E. coli cells with thisDNA (Fig. 1B, 4). Colonies resistant to either kana-mycin or ampicillin were isolated on medium sup-plemented with the corresponding antibiotics (Fig. 1B,5). In order to distinguish between E. coli colonies thatacquired binary plasmids from the A. tumefaciens inthe infected plant tissue and E. coli colonies that poten-tially acquired T-circles from plant cells, each colonywas tested for spectinomycin/streptomycin resistance(Fig. 1B, 6). As the SpR/SmR sequence is located onthe binary backbone, outside the T-DNA borders, col-onies sensitive to spectinomycin/streptomycin wereruled out as containing binary plasmids and thuswere further analyzed (Fig. 1B, 7). The frequency ofspectinomycin/streptomycin-sensitive colonies amongampicillin- or kanamycin-resistant colonies varied inagroinfiltration experiments using different plant species.While 5% to 10% of the colonies were spectinomycin/streptomycin sensitive when using tobacco (Nicotianatabacum; Supplemental Fig. S2), more than 30% ofthe colonies were spectinomycin/streptomycin sensi-tive when using Nicotiana benthamiana (see additionaldata below).
512 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

Restriction Digestion Reveals T-Circles of Various Sizesand Structures
The plasmids recovered from spectinomycin/streptomycin-sensitive colonies (T-circles) were ana-lyzed by restriction digestion and gel electrophoresis.A ScaI site is present within the AmpR gene of theAMP-ORI construct (Fig. 1A). Thus, the T-circles con-ferring E. coli resistance to ampicillin are expected tobe digested at least once by ScaI. A second ScaI site ispresent in the pRCS2 binary backbone sequence 113 bpfrom the T-DNA’s right border (Fig. 1A). Thus, theT-circles digested twice with ScaI may contain an ad-ditional sequence that may be either a second T-DNAfragment or a binary backbone sequence. The latter mayderive from incorrect processing of T-DNAmolecules inthe bacterium (Kononov et al., 1997; Wenck et al., 1997).A random selection of six T-circles isolated from
tobacco agroinfiltrated with the AMP-ORI construct isshown in Figure 2A (T-1–T-6). Out of the six treatedwith ScaI, four T-circles generated a single band in asize ranging between 2.2 and 3.4 kb, suggesting thatthey consisted of a single copy of T-DNA with somedeletions or additions of DNA (T-1–T-4). The T-circlesT-5 and T-6 produced two bands after ScaI digestion,suggesting a complex structure in these T-circles.
Using Different T-DNA Constructs Simultaneously viaCoagroinfiltration Generates Complex T-Circles ThatContain Both T-DNAs
Cotransforming plants with different T-DNA con-structs simultaneously often results in the integrationof different T-DNAs at the same location (De Neveet al., 1997; Radchuk et al., 2005). To test the extra-chromosomal T-DNA ligation model, we hypothe-sized that when different T-DNA constructs are
delivered to plants simultaneously, they ligate to eachother, prior to integration, and occasionally generateT-circles. To this end, we introduced the AMP-ORIconstruct and either the KAN-ORI or the KAN con-struct simultaneously into plants by “coagroinfiltra-tion.” The different A. tumefaciens strains containingeach of the T-DNA constructs were mixed immedi-ately before coagroinfiltration.
Following coagroinfiltration, we isolated T-circlesfrom the E. coli colonies resistant to both ampicillin andkanamycin. A ClaI site is present within the KanR genein KAN-ORI and KAN (Fig. 1A) but is not present inthe binary backbone itself. Accordingly, T-circles weredigested at least once with ClaI in addition to ScaI.Figure 2B shows a sample of T-circles digested withScaI and ClaI. Overall, the T-circles ranged in size be-tween 5 and 15 kb. KAN was coagroinfiltrated withAMP-ORI in T-7, T-8, T-10, and T-11, while KAN-ORIwas coagroinfiltrated with AMP-ORI in T-9 and T-12to T-16. Digestion of T-7 and T-8 with either ScaI orClaI revealed a single site for each enzyme and pro-duced a band of about 5 kb. This represents the ap-proximate fragment size expected from a ligation ofAMP-ORI and KAN. Sequencing analysis confirmedthe ligation between the two different T-DNA con-structs (Fig. 2C shows a schematic diagram of one ofthe junction regions of T-7 based on the sequencinganalysis shown in Supplemental Fig. S3). Digestion ofT-9 and T-12 also revealed single ScaI and ClaI sites.These T-circles were notably larger than expected fromthe joining of two T-DNAs and also conferred a re-sistance to spectinomycin/streptomycin, suggestingthat they included the aadA (SpR/SmR) sequence fromthe binary backbone. Similarly, T-15 and T-16 alsoconferred resistance to spectinomycin/streptomycin.Digestion of T-10, T-11, and T-13 to T-16 with ScaIrevealed two to three sites, while digestion of T-10 and
Figure 1. Constructs and experimental procedurefor the isolation of T-circles. A, Schematic dia-gram of the three T-DNA constructs used through-out the experiments: AMP-ORI, AmpR with ori;KAN-ORI, KanR with ori; KAN, KanR without ori. B,Illustration of the experimental procedure: 1,transformation of A. tumefaciens with a pRCS2binary plasmid harboring one of the three T-DNAconstructs (AMP-ORI is shown); 2, agroinfiltrationof plant leaves; 3, extraction of DNA from theagroinfiltrated leaves (4–8 d after infiltration); 4,transformation of E. coli with the DNA; 5, selectionof colonies resistant to ampicillin (or kanamycinif KAN-ORI or KAN is used); 6, detection ofspectinomycin/streptomycin-sensitive colonies;7, isolation of plasmids from each colony andanalysis by restriction digestion and DNA se-quencing. amp, Ampicillin; kan, kanamycin;spec, spectinomycin; strep, streptomycin. [Seeonline article for color version of this figure.]
Plant Physiol. Vol. 160, 2012 513
Complex Extrachromosomal T-DNA Structures
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

T-16 with ClaI revealed two sites (note the nearlyidentical size of the two ClaI bands in T-16), thussuggesting complex T-circles that contain multipleDNA fragments.
A. tumefaciens T-DNA Transfer Is Required forT-Circle Formation
To further study the origin of the T-circles, we testedthe effect of using a mutant A. tumefaciens strain defi-cient in T-DNA transfer. If the T-circles are producedin plant cells, then they should not be generated usingthis mutant. We used a virB92 A. tumefaciens mutantstrain that does not produce galls due to a defect in theT-DNA transfer apparatus (Beaupré et al., 1997). Wefirst tested whether T-DNA transfer can be detected inan agroinfiltration assay using this mutant strain. AT-DNA construct with the GUS sequence interruptedby an intron (GUS-int) was used to examine transientexpression in plants as an indication for T-DNA
transfer (Ohta et al., 1990). As expected, the virB92
strain did not induce GUS expression in N. benthamianaleaves (Fig. 3A), confirming that it is transfer deficientunder our experimental conditions. To complementthis strain, we used the pED37 plasmid, which con-tains virB9 under virB promoter control (Ward et al.,1991). GUS activity was restored in virB92/pED37,confirming that T-DNA transfer is restored by thisplasmid (Fig. 3A). Next, we transformed strainsvirB92, virB92/pED37, and control EHA105 with theAMP-ORI construct and agroinfiltrated plants in or-der to test the formation of T-circles (Fig. 3B). Whenthe virB92 strain was used, all of the E. coli coloniesselected on ampicillin (n = 675) were spectinomycin/streptomycin resistant, indicating that T-circles werenot generated. When the virB92/pED37 strain wasused, 44% of the colonies selected on ampicillin (n =107) were spectinomycin/streptomycin sensitive, anindication that a large proportion of the colonies con-tained T-circles. Similarly, the EHA105 control strainproduced 61% spectinomycin/streptomycin-sensitive
Figure 2. Restriction digestion reveals T-circles ofvarious sizes and structures. A, Agroinfiltration oftobacco with AMP-ORI (T-1–T-6). B, Coagroinfil-tration of N. benthamiana with AMP-ORI andKAN (T-7, T-8, T-10, and T-11) or AMP-ORI andKAN-ORI (T-9 and T-12–T-16). T-circles are shownon agarose gels as uncut (U; left lane), ScaI treated(S; middle lane), or ClaI treated (C; right lane). EachT-circle is marked by + or 2 (below the gel) toindicate the resistance or sensitivity (respectively)to an antibiotic (left side) it confers to E. coli. Thebinary plasmids AMP-ORI and KAN-ORI areshown (last two on bottom right). C, Schematicdiagram of T-7 shows a junction fragment of AMP-ORI and KAN. Numbers in parentheses representthe positions within the T-DNA relative to the LB(0 point) to indicate the deleted regions from eachside. [See online article for color version of thisfigure.]
514 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.
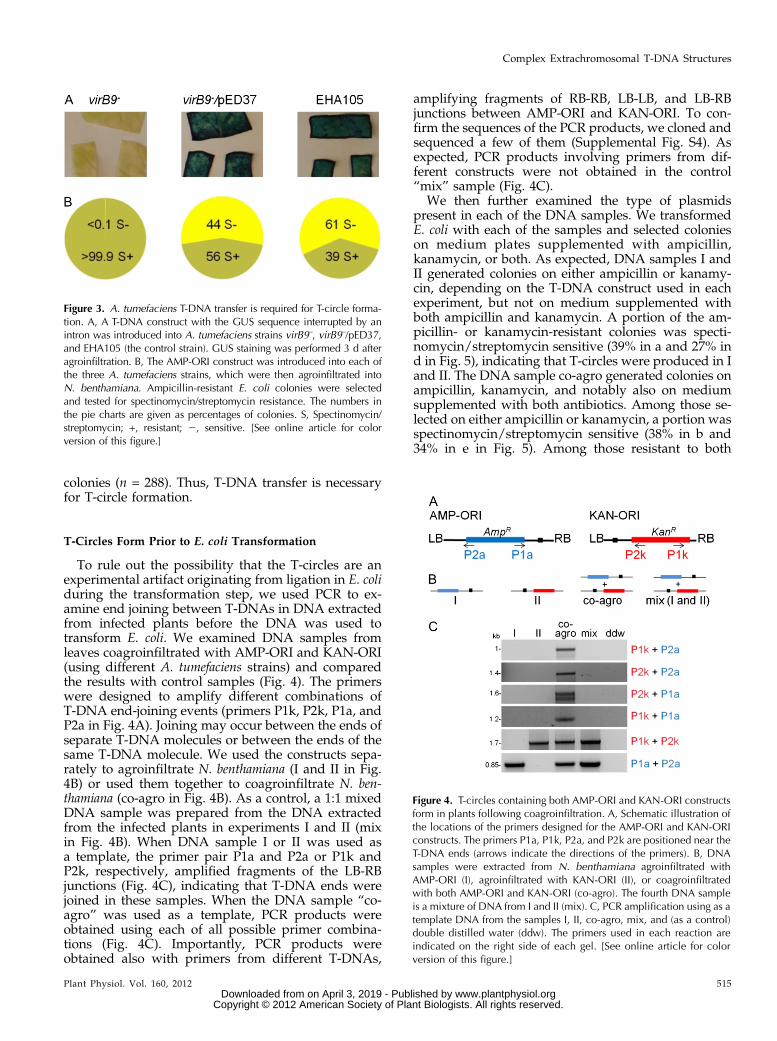
colonies (n = 288). Thus, T-DNA transfer is necessaryfor T-circle formation.
T-Circles Form Prior to E. coli Transformation
To rule out the possibility that the T-circles are anexperimental artifact originating from ligation in E. coliduring the transformation step, we used PCR to ex-amine end joining between T-DNAs in DNA extractedfrom infected plants before the DNA was used totransform E. coli. We examined DNA samples fromleaves coagroinfiltrated with AMP-ORI and KAN-ORI(using different A. tumefaciens strains) and comparedthe results with control samples (Fig. 4). The primerswere designed to amplify different combinations ofT-DNA end-joining events (primers P1k, P2k, P1a, andP2a in Fig. 4A). Joining may occur between the ends ofseparate T-DNA molecules or between the ends of thesame T-DNA molecule. We used the constructs sepa-rately to agroinfiltrate N. benthamiana (I and II in Fig.4B) or used them together to coagroinfiltrate N. ben-thamiana (co-agro in Fig. 4B). As a control, a 1:1 mixedDNA sample was prepared from the DNA extractedfrom the infected plants in experiments I and II (mixin Fig. 4B). When DNA sample I or II was used asa template, the primer pair P1a and P2a or P1k andP2k, respectively, amplified fragments of the LB-RBjunctions (Fig. 4C), indicating that T-DNA ends werejoined in these samples. When the DNA sample “co-agro” was used as a template, PCR products wereobtained using each of all possible primer combina-tions (Fig. 4C). Importantly, PCR products wereobtained also with primers from different T-DNAs,
amplifying fragments of RB-RB, LB-LB, and LB-RBjunctions between AMP-ORI and KAN-ORI. To con-firm the sequences of the PCR products, we cloned andsequenced a few of them (Supplemental Fig. S4). Asexpected, PCR products involving primers from dif-ferent constructs were not obtained in the control“mix” sample (Fig. 4C).
We then further examined the type of plasmidspresent in each of the DNA samples. We transformedE. coli with each of the samples and selected colonieson medium plates supplemented with ampicillin,kanamycin, or both. As expected, DNA samples I andII generated colonies on either ampicillin or kanamy-cin, depending on the T-DNA construct used in eachexperiment, but not on medium supplemented withboth ampicillin and kanamycin. A portion of the am-picillin- or kanamycin-resistant colonies was specti-nomycin/streptomycin sensitive (39% in a and 27% ind in Fig. 5), indicating that T-circles were produced in Iand II. The DNA sample co-agro generated colonies onampicillin, kanamycin, and notably also on mediumsupplemented with both antibiotics. Among those se-lected on either ampicillin or kanamycin, a portion wasspectinomycin/streptomycin sensitive (38% in b and34% in e in Fig. 5). Among those resistant to both
Figure 4. T-circles containing both AMP-ORI and KAN-ORI constructsform in plants following coagroinfiltration. A, Schematic illustration ofthe locations of the primers designed for the AMP-ORI and KAN-ORIconstructs. The primers P1a, P1k, P2a, and P2k are positioned near theT-DNA ends (arrows indicate the directions of the primers). B, DNAsamples were extracted from N. benthamiana agroinfiltrated withAMP-ORI (I), agroinfiltrated with KAN-ORI (II), or coagroinfiltratedwith both AMP-ORI and KAN-ORI (co-agro). The fourth DNA sampleis a mixture of DNA from I and II (mix). C, PCR amplification using as atemplate DNA from the samples I, II, co-agro, mix, and (as a control)double distilled water (ddw). The primers used in each reaction areindicated on the right side of each gel. [See online article for colorversion of this figure.]
Figure 3. A. tumefaciens T-DNA transfer is required for T-circle forma-tion. A, A T-DNA construct with the GUS sequence interrupted by anintron was introduced into A. tumefaciens strains virB9–, virB9–/pED37,and EHA105 (the control strain). GUS staining was performed 3 d afteragroinfiltration. B, The AMP-ORI construct was introduced into each ofthe three A. tumefaciens strains, which were then agroinfiltrated intoN. benthamiana. Ampicillin-resistant E. coli colonies were selectedand tested for spectinomycin/streptomycin resistance. The numbers inthe pie charts are given as percentages of colonies. S, Spectinomycin/streptomycin; +, resistant; 2, sensitive. [See online article for colorversion of this figure.]
Plant Physiol. Vol. 160, 2012 515
Complex Extrachromosomal T-DNA Structures
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

ampicillin and kanamycin, a higher proportion wasspectinomycin/streptomycin sensitive (75% [6% from8%] in b and 80% [8% from 10%] in e in Fig. 5). Thecontrol mixture did not generate resistant colonieson medium supplemented with both ampicillin andkanamycin, only on ampicillin or kanamycin sepa-rately. Thus, our experiments confirm that T-circles arepresent in the DNA prior to E. coli transformation andfurther support that they form in plants.
DNA Sequencing of T-DNA Junctions Reveals Details ofComplex T-Circles
To further analyze T-circle structures at the sequencelevel, we sequenced the T-DNA junctions in T-1 to T-9,T-11, and T-12 (Supplemental Fig. S3). According tothe junction sequences and the sizes of the bands afterrestriction digestion, we predicted the completestructure of the T-circles T-1 to T-5 (Fig. 6A). In T-1, an82-bp fragment from the LB side of the T-DNA wasmissing. However, the RB side was complete: it in-cluded the first three nucleotides of the RB sequence(Fig. 6A; Supplemental Fig. S3). We refer to a borderending at the third position of the 25-bp direct im-perfect repeat as a “precise” ending because it is thesite at which the VirD1-VirD2 complex is known tonick the lower T-DNA strand in A. tumefaciens (Wanget al., 1987). In T-2, both the LB and RB had preciseendings (Fig. 6A; Supplemental Fig. S3). On the otherhand, T-3 was missing 598 and 595 bp from the LB and
RB sides, respectively, and was attached to a 1.4-kbsequence derived from the binary plasmid (Fig. 6A;Supplemental Fig. S3). This binary fragment, whichdid not include SpR/SmR, was not a “read-through” ofthe binary sequence adjacent to T-DNA from its LB orRB side (Kononov et al., 1997; Wenck et al., 1997;Kim et al., 2003). T-9 also had a T-DNA with a non-read-through binary plasmid sequence. In this case,the binary backbone sequence that is immediately nextto the LB was attached to the RB side of a T-DNAin the T-circle (Supplemental Fig. S3). On the otherhand, in T-11 and T-12, the binary plasmid sequencewas a read-through of the T-DNA RB and LB, re-spectively (Supplemental Fig. S3). In T-4, the borderjunction included 5 bp of microhomology (GCGCC);thus, the exact ending point of each side can be any-where within this microhomology. The LB and RBsides lost from 363 to 368 bp and from 596 to 601 bp,respectively (Fig. 6A; Supplemental Fig. S3). T-5 con-tained two T-DNA fragments arranged in an invertedrepeat configuration. One T-DNA fragment includedthe AmpR gene and the ori. It was missing 612 bp of theLB side, whereas the RB was precise. The secondfragment was missing 1,088 and 1,465 bp from theLB and RB sides, respectively (Fig. 6A; SupplementalFig. S3). T-6 produced two fragments after digestionwith ScaI (Fig. 2A), BsaXI, or AatII (Fig. 6B). The sizesof the bands suggested that two relatively completeT-DNAs had joined in an inverted configuration. Toconfirm this, we cloned the junction fragments pro-duced by AatII and BsaXI (Fig. 6B) into the EcoRVsite of pBluescript KS1 (after blunt ending the frag-ments) and sequenced them. Sequencing the AatIIfragment showed LB-LB region joining (Fig. 6C, top;Supplemental Fig. S3). The junction included 3 bp ofmicrohomology (TAA). One LB side was missing be-tween 21 and 24 bp, whereas the other was missingbetween 261 and 264 bp. The BsaXI fragment includedRB-RB region joining (Fig. 6C, bottom; SupplementalFig. S3). It had a patchwork of unknown filler se-quence (“scrambled”) interrupted by short T-DNAsequences. The filler sequences from both ends of theinverted T-DNAs started at similar positions near theirBsaXI sites (2,640): positions 2,658 and 2,686. From theleft side, the T-DNA ending at 2,658 was interruptedby 225 bp of scrambled filler sequence. After this filler,there appeared a 37-bp sequence from a RB T-DNAregion (2,885–2,922). Interestingly, this short T-DNAsequence started 227 bp away from the position atwhich the previous T-DNA was interrupted by the225-bp filler, and it was in the same orientation. Afterthis T-DNA sequence, another scrambled filler DNAsequence began. We were not able to sequence theinternal region of the fragment, possibly due to theformation of secondary structures. Sequencing fromthe right side revealed 137 bp of scrambled filler DNAfollowed by an inverted T-DNA RB region fragment(greater than 199 bp). Additional sequences from T-7,T-8, T-9, T-11, and T-12 are shown in SupplementalFigure S3.
Figure 5. Antibiotic resistance of E. coli colonies transformed withDNA I, II, co-agro, and mix (the DNA sample types are indicatedabove the pie charts). Colonies were initially selected on either am-picillin or kanamycin (indicated to the left of the pie charts) and thentested for spectinomycin/streptomycin, ampicillin, and kanamycinresistance. The numbers in the pie charts are given as percentages ofcolonies. The numbers of colonies tested in each group are follows:a, n = 96; b, n = 478; c, n = 383; d, n = 120; e, n = 473; f, n = 477.A, Ampicillin; K, kanamycin; NC, no colonies; S, spectinomycin/streptomycin; +, resistant, 2, sensitive. [See online article for colorversion of this figure.]
516 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

DISCUSSION
To our knowledge, extrachromosomal complexT-DNA structures have not been reported previouslyin plants or A. tumefaciens. To study T-DNA integra-tion and the formation of complex T-DNA insertions inplants, we developed a method to isolate complexT-DNA structures in the form of T-circles directly fromA. tumefaciens-infected plants. The T-circles resembledstructures commonly found in stable transgenic plants,which suggests that similar linear ds DNA structuresare present in plants prior to integration. If not inte-grated, ds DNA linear structures may circularize.
The Formation of Complex T-Circles in Plants
We first confirmed that our isolated T-circles areformed in the plant cells, because T-circles can poten-tially also form in A. tumefaciens during the infection.Although the circularization of a T-DNA in A. tume-faciens is a rare event with no apparent role in the chain
of events leading to transformation, a T-circle may beoccasionally released from its parent plasmid throughhomologous recombination between two direct re-peats that flank the T-DNA (Koukoliková-Nicola et al.,1985; Machida et al., 1986; Timmerman et al., 1988).Sequencing of end-joining junctions in our T-circlesrevealed that their ends had none or only a few basepairs of overlapping microhomology (SupplementalFig. S3), thus indicating that our T-circles did not formthrough homologous recombination, as occurs in A.tumefaciens, but through illegitimate end joining, whichis typical of the circularization of T-DNAs in infectedplants (Bakkeren et al., 1989; Zhao et al., 2003).Moreover, T-circles containing T-DNAs from differentA. tumefaciens strains were generated following co-agroinfiltration (Figs. 4 and 5), emphasizing the re-quirement of T-DNA transfer out of the bacteria priorto circularization. Finally, the requirement of T-DNAtransfer for T-circle formation was also demonstratedusing an A. tumefaciens strain deficient in the ability to
Figure 6. Schematic illustration of the T-circles T-1 to T-6 based on DNA sequencing. A, T-1 to T-5 are shown in a linear form,opened at the position of a junction, and aligned below a complete AMP-ORI construct. B, T-6 after digestion with BsaXI (leftlane) or AatII (right lane). The bands that include the junctions are indicated by arrows. C, Schematic illustration of the AatIIfragment (top; the LB-LB region junction of T-6) and the BsaXI fragment (bottom; the RB-RB region junction of T-6). [See onlinearticle for color version of this figure.]
Plant Physiol. Vol. 160, 2012 517
Complex Extrachromosomal T-DNA Structures
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

transfer T-DNA to plants (Fig. 3). However, it is im-possible to rule out completely that a small number ofT-circles are produced in A. tumefaciens. For example,T-2 had a precise border joining (Fig. 6); thus, poten-tially, it may have formed in A. tumefaciens throughhomologous recombination between the borders. Thenext possibility that we considered was that theT-circles are an artifact of the experimental procedure,forming through random ligation in E. coli during thetransformation step. We ruled out this possibility, first,because PCR of DNA extracted from coagroinfiltratedplants (before the DNA was used for E. coli transfor-mation) confirmed end joining between the AMP-ORIand KAN-ORI constructs (Fig. 4C; Supplemental Fig.S4). Second, T-circles containing both AMP-ORI andKAN-ORI could be isolated in E. coli only if trans-formed with DNA extracted from coagroinfiltratedplants, not in E. coli transformed with a DNA mixturefrom separate agroinfiltration experiments (Fig. 5).Thus, we conclude that the T-circles formed in theplants cells.
T-DNA Integration and the Formation of ComplexT-DNA Insertions
Our results support a model of integration thatpredicts that T-DNAs integrate as ds DNA molecules(Mayerhofer et al., 1991) and that complex transgenicstructures form when ds DNA fragments ligate ex-trachromosomally prior to integration (De Neve et al.,1997). Our conclusion is based on the finding of ex-trachromosomal ds T-DNA complex structures similarto complex patterns commonly found integrated intransgenic plants. The complex patterns that weidentified include T-DNA repeats, binary plasmid se-quences, filler DNA sequences, microhomologies, andT-DNA truncations.
T-DNA repeats are a frequent pattern found at in-sertion sites following A. tumefaciens-mediated trans-formation (Cluster et al., 1996; Krizkova and Hrouda,1998; Wolters et al., 1998; De Paepe et al., 2009).Moreover, following cotransformation using differentT-DNA constructs, cointegration of the T-DNAs oftenoccurs into the same site (De Buck et al., 2000; Radchuket al., 2005). Similarly, following agroinfiltration andcoagroinfiltration, we recovered T-circles with morethan one T-DNA of the same type or different types,respectively (Figs. 2, 5, and 6). The joining between theT-DNAs can be at their RB-LB, RB-RB, or LB-LB sides.Joining at the RB-RB or LB-LB side is particularly dif-ficult to explain by the ss model, because the ss T-DNAis always 59 at its RB terminus and 39 at its LB ter-minus. Thus, further supporting the idea that theT-DNAs in our T-circles were ds DNA molecules priorto their extrachromosomal ligation, sequencing re-vealed T-DNAs joined from their RB-RB and LB-LBsides (Fig. 6; Supplemental Fig. S3).
Binary plasmid sequences adjacent to the insertedT-DNAs is a pattern that has been reported in 30% to
70% of transgenic plant lines (Martineau et al., 1994;Kononov et al., 1997; Wenck et al., 1997; Kim et al.,2003; Lange et al., 2006). Similarly, our results indicatethat binary sequences were present in more than one-quarter of the T-circles containing both AMP-ORIand KAN-ORI, because one-quarter of them also con-ferred spectinomycin/streptomycin resistance (Fig. 5).A read-through transfer of a T-DNA together with anadjacent binary sequence occurs frequently when theVirD1-VirD2 endonuclease supposedly skips the pro-cessing of one of the borders in A. tumefaciens duringthe release of the T-DNA from its parent binary plas-mid (Kononov et al., 1997; Wenck et al., 1997). Becausethe SpR/SmR gene sequence in our system is located onthe binary backbone near the LB (approximately 0.5 kbaway), a read-through transfer of SpR/SmR sequencewith the T-DNA can be expected to occur frequently.For example, a read-through of the LB occurred in T-12(Supplemental Fig. S3), which also conferred specti-nomycin/streptomycin resistance (Fig. 2B). In addi-tion, other binary sequences that do not include SpR/SmR may have been present in other structures butwere not detected in the resistance assay. For example,the binary sequence adjacent to the T-DNA in T-3 didnot include SpR/SmR and was not a read-through fromeither side of the T-DNA (Fig. 6; Supplemental Fig. S3).Because complex T-DNA structures have never beenreported in A. tumefaciens, and in agreement with thereport of Kononov et al. (1997) that binary sequencesin plants can integrate independently of the T-DNA,the binary sequence in T-3 most likely entered theplant cell independently as ss DNA, then converted tods DNA, and finally ligated to a ds T-DNA.
Filler DNA is DNA sequence from an unknownsource that often flanks T-DNA insertions. Filler DNAis a pattern also found at genomic DSB-repair sitesin plants. It is believed to form through a modifiedsynthesis-dependent strand-annealing mechanism thatinvolves random template “switches” of the DNA poly-merase, resulting in a patchwork of different fillersequences (Gorbunova and Levy, 1997; Salomon andPuchta, 1998). Patterns of filler DNA at T-DNA inser-tion sites often consist of short sequences of T-DNA,plant genome, or scrambled DNA (Gheysen et al.,1991; Mayerhofer et al., 1991; Krizkova and Hrouda,1998; Forsbach et al., 2003; Windels et al., 2003). Wefound a similar pattern of filler DNA in T-6 (Fig. 6C;Supplemental Fig. S3). The RB-RB junction consisted ofa patchwork of scrambled filler DNA interrupted byshort sequences homologous to the RB region. Onefiller sequence had a length almost identical (225 bp) tothe T-DNA sequence it intercepted (227 bp). This mayindicate that a template switch of DNA polymerasehad occurred at this region. Alternatively, end joiningbetween “free-floating” ds DNA fragments may po-tentially result in patchworks.
Truncations of T-DNA ends and microhomologiesof 2 to 7 bp at plant/T-DNA and T-DNA/T-DNA in-sertion junctions are common in transgenic plants(Tinland, 1996; Brunaud et al., 2002; Windels et al.,
518 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

2003). They are also associated with the repair of ge-nomic DSBs in plants (Gorbunova and Levy, 1997;Salomon and Puchta, 1998). Similarly, T-DNA trun-cations were found in most of our T-circles (Fig. 6;Supplemental Fig. S3). Microhomologies of between1 and 5 bp were found at T-circles T-4, T-6, T-8, andT-9. In T-2, a precise junction contained 12 bp of theoverlapping border direct repeats. In T-11, AMP-ORIand KAN T-DNA were joined within an 85-bp over-lapping region (Supplemental Fig. S3). Truncationsand microhomology were also detected in AMP-ORI/KAN-ORI junctions amplified by PCR (SupplementalFig. S4).Because the ligation patterns in the T-circles de-
scribed above resemble the ligation patterns of re-paired DNA DSBs, it is likely that T-DNA integration,the formation of complex T-DNA structures, and cir-cularization share the same DNA-repair mechanism.That we isolated only circular but not linear ds DNAstructures is due only to the prerequisite of our de-tection system; circular configurations suggest thatsimilar linear configurations also exist in the plant cell.In support of the idea that termini of ds T-DNA arerecognized as DSBs, it has been shown previously thatgenomic DSBs may be “hot spots” for ds T-DNA inte-gration (Salomon and Puchta, 1998; Chilton and Que,2003; Tzfira et al., 2003). The major pathway to repairDSBs in plants is the nonhomologous end joining(NHEJ) DNA-repair pathway (for review, see Bleuyardet al., 2006). Supporting a role for the NHEJ pathwayin T-DNA integration, two components of the NHEJpathway, LigIV and KU80, have been reported to berequired for T-DNA integration (Friesner and Britt,2003; Li et al., 2005). Conversely, reports by Gallegoet al. (2003) that KU80 is dispensable for T-DNA in-tegration and by Park et al. (2011) that KU80 and other
NHEJ components limit integration suggest thatthe absolute role of the NHEJ pathway for T-DNAintegration is still inconclusive. It is possible that al-ternative DSB-repair pathways, such as the micro-homology-mediated end joining DNA-repair pathway(McVey and Lee, 2008), play a role in T-DNA inte-gration. However, the process of DSB repair in plantsis still not fully understood, underscoring the difficultyin understanding the mechanism behind T-DNA in-tegration. Nonetheless, given the high frequency ofcomplex T-DNA structures usually found in transgenicplants and our detection of similar complex ds DNAstructures prior to integration, we suggest that themajor route for integration is through the ds T-DNAand the DSB DNA-repair pathway. According to thismodel (Fig. 7), ss T-DNA molecules are first trans-ferred into the plant nucleus via the type IV secretionsystem (for review, see McCullen and Binns, 2006;Citovsky et al., 2007; Gelvin, 2010; Tzfira and Citovsky,2010). In addition to T-DNA molecules, other bacterialDNA may enter the plant cell, including binary plasmidDNA (Kononov et al., 1997; Wenck et al., 1997) and A.tumefaciens chromosomal DNA (Ulker et al., 2008).Once inside the nucleus, ss DNA molecules are con-verted into ds DNA molecules (Janssen and Gardner,1990; Offringa et al., 1990; Narasimhulu et al., 1996).The plant’s DSB DNA-repair machinery recognizes assubstrate for repair the exposed termini of the ds DNAmolecules. This repair can lead to end joining betweenfree-floating ds DNA molecules, thus creating circularand complex extrachromosomal molecules. Alterna-tively, the DSB DNA-repair machinery may also me-diate integration of the ds DNA molecules into theplant genome. VirD2, which is covalently attached tothe RB of the transferred ss T-DNAs, may also remainattached to the ds T-DNAs and thus play a role in the
Figure 7. A proposed model for T-DNA integration, the formation of complex T-DNA structures, and T-circles. 1, In A.tumefaciens, the T-DNA borders are nicked by the VirD1-VirD2 endonuclease complex and an ss T-DNA (T-strand) is excised(the T-DNAs are marked in red). A VirD2 protein remains attached to the 59 terminus of the T-strand (the RB). 2, The T-strand istransferred into the plant cell nucleus. Additional ss DNA molecules (derived from the A. tumefaciens binary plasmid orchromosomal DNA) may occasionally be transferred independently or together with a T-DNA by read-though linkage. 3, In thenucleus, ss DNAs are converted into ds DNAs (only ds T-DNAs are shown). 4, ds DNA may then integrate into the genome (firstoutcome) via VirD2 and the plant’s DSB-repair pathway. 5, Termini of a ds DNAmay ligate to each other (second outcome) andgenerate T-circles. 6, Termini of different ds DNA molecules may ligate to each other (third outcome) and form complexstructures. These structures may then integrate or form complex circular molecules. [See online article for color version of thisfigure.]
Plant Physiol. Vol. 160, 2012 519
Complex Extrachromosomal T-DNA Structures
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

integration process. It is important to note that T-DNAmolecules may also integrate as ss molecules (Tinland,1996), yet the process by which such molecules mayform complex T-DNA structures may differ from ourproposed model.
The Aspects of in Planta T-DNA Circularization
Because “free” ends are not available in T-circles,such molecules may be mostly “dead-end” productsthat do not integrate. Therefore, the extent of ds T-DNAcircularization is a relevant question when consideringthe efficiency of T-DNA integration. Previous studieshave shown that linearized plasmids recircularize inprotoplasts (Gorbunova and Levy, 1997; Gallego et al.,2003). However, circularization of ds T-DNAs in plantswas considered a relatively rare event until now, be-cause evidence for circularization was indirect andrequired a viral amplification system (Bakkeren et al.,1989; Zhao et al., 2003). Our ability to detect T-circlesdirectly from infected plants using a plasmid-rescueapproach suggests that circularization of ds T-DNAsmay not be as rare as previously thought. The rela-tionship between circularization and integration is anespecially interesting question, considering that dsT-DNAs seem to be abundant in infected plants. Theexistence of free ds T-DNAs in infected plant cellsimmediately after infection is evident from the earlyand broad transient gene expression in infected planttissues (Janssen and Gardner, 1990), from transientexpression using a nontemplate strand (Narasimhuluet al., 1996), from experiments involving homologousrecombination (Offringa et al., 1990), and from theisolation of extrachromosomal T-DNAs in the formof ds plasmids (this work). Further highlighting thisquestion, the majority the free ds T-DNA mole-cules do not integrate (Janssen and Gardner, 1990;Narasimhulu et al., 1996; Maximova et al., 1998; DeBuck et al., 2000), but their fate is unclear becausetransient gene expression that can be used to monitorthem is silenced usually a few days after infection(Johansen and Carrington, 2001).
Any outcome that is not integration of the deliveredT-DNA is probably unfavorable for the A. tumefaciens.Thus, it is tempting to speculate that A. tumefaciensevolved mechanisms that favor T-DNA integration overextrachromosomal ligation. It might also explain why A.tumefaciens-mediated transformation is more efficientwhile producing relatively fewer complex insertions com-pared with transformation by the particle-bombardmentmethod (Makarevitch et al., 2003; Travella et al., 2005).A potential candidate that may be involved in amechanism that prefers integration over extrachromo-somal ligation is VirD2, because it is bound to the 59 endof the RB (Herrera-Estrella et al., 1988) and has beensuggested to play a role, albeit unknown, in T-DNAintegration (Tinland et al., 1995; Mysore et al., 1998).
Understanding how complex T-DNA insertions formmay help identify ways to develop plants with single
perfect insertions, as generally desired for crop im-provements. Our DNA recovery system may be use-ful in future related studies. Furthermore, additionalstudies are needed to uncover the spectrum of factorsfrom plants and A. tumefaciens involved in T-DNA in-tegration and their specific roles in the process.
MATERIALS AND METHODS
Plant Material and Agroinfiltration
Tobacco (Nicotiana tabacum) and Nicotiana benthamiana plants were grownat 20°C under long-day conditions (14 h of light). Plants were grown on SuperGermination mix (Fafard) and agroinfiltrated when about 4 to 8 weeks old.Agroinfiltration was performed as described by Sparkes et al. (2006) withmodifications. Infected leaves were harvested and DNA extracted using amodified cetyl-trimethyl-ammonium bromide protocol as described by Doyleand Doyle (1987). For complete DNA extraction and agroinfiltration protocols,see Supplemental Texts S1 and S2, respectively.
Constructs, Bacterial Strains, Transformation, andTransient T-DNA Assay
The AMP-ORI T-DNA construct was prepared by cloning the backbone ofthe pSAT6 vector (Tzfira et al., 2005) into the pRCS11 binary plasmid (Zeeviet al., 2008). The KAN-ORI and KAN T-DNA constructs were prepared bycloning pET28c backbone sequences into the pRCS2 vector (Tzfira et al., 2005).The primers used for PCR and cloning the KanR gene are listed in SupplementalText S3. Cloning was performed using standard procedures. Binary vectors wereintroduced into EHA105, a disarmed stain of Agrobacterium tumefaciens (excep-tions are experiments involving the virB9– mutant [strain Ax42], which is derivedfrom the A348 background; Beaupré et al., 1997). To recover T-circles, we usedthe X-cell electroporation system (Bio-Rad) with DH5a, Top10 (Invitrogen), or10-b (New England Biolabs) Escherichia coli competent cells. The following an-tibiotic concentrations in Luria-Bertani medium were used for E. coli: ampicillin(100 mg mL21), kanamycin (50 mg mL21), and spectinomycin/streptomycin(50 mg mL21 each). A. tumefaciens cells were grown on yeast extract peptonemedium with the following antibiotic concentrations: spectinomycin/streptomycin(200 mg mL21 each) and rifampicin (10 mg mL21). After agroinfiltration with aGUS-expressing vector, leaves were stained using 5-bromo-4-chloro-3-indolyl-b-glucuronic acid as described by Li et al. (2005).
T-Circle Analysis
Molecular genetics methods for T-circle analysis, including the primersused for sequencing, are detailed in Supplemental Text S3.
Supplemental Data
The following materials are available in the online version of this article.
Supplemental Figure S1. DNA Sequences of the T-DNA constructs AMP-ORI, KAN-ORI, and KAN.
Supplemental Figure S2. Frequency of spectinomycin/streptomycin sen-sitive colonies from agroinfiltrated tobacco plants.
Supplemental Figure S3. Sequencing of T-DNA junctions in T-circles T-1to T-9, T-11, and T-12.
Supplemental Figure S4. Sequencing of PCR products.
Supplemental Text S1. DNA extraction protocol for T-circle isolation.
Supplemental Text S2. Agroinfiltration protocol.
Supplemental Text S3. T-circle analysis.
ACKNOWLEDGMENTS
We thank all members of the Tzfira laboratory for their help. We thankAndrew Binns (University of Pennsylvania) for providing strain Ax42 and
520 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

plasmid pED37. We thank Lyle Simmons and Tariq Akhtar (University ofMichigan) for helpful discussions and critical reading of the manuscript
Received May 13, 2012; accepted July 12, 2012; published July 13, 2012.
LITERATURE CITED
Bakkeren G, Koukolíková-Nicola Z, Grimsley N, Hohn B (1989) Recoveryof Agrobacterium tumefaciens T-DNA molecules from whole plants earlyafter transfer. Cell 57: 847–857
Beaupré CE, Bohne J, Dale EM, Binns AN (1997) Interactions betweenVirB9 and VirB10 membrane proteins involved in movement of DNAfrom Agrobacterium tumefaciens into plant cells. J Bacteriol 179: 78–89
Bleuyard JY, Gallego ME, White CI (2006) Recent advances in under-standing of the DNA double-strand break repair machinery of plants.DNA Repair (Amst) 5: 1–12
Brunaud V, Balzergue S, Dubreucq B, Aubourg S, Samson F, Chauvin S,Bechtold N, Cruaud C, DeRose R, Pelletier G, et al (2002) T-DNA in-tegration into the Arabidopsis genome depends on sequences of pre-insertion sites. EMBO Rep 3: 1152–1157
Chilton MD, Que Q (2003) Targeted integration of T-DNA into the tobaccogenome at double-stranded breaks: new insights on the mechanism ofT-DNA integration. Plant Physiol 133: 956–965
Citovsky V, Kozlovsky SV, Lacroix B, Zaltsman A, Dafny-Yelin M, VyasS, Tovkach A, Tzfira T (2007) Biological systems of the host cell in-volved in Agrobacterium infection. Cell Microbiol 9: 9–20
Cluster PD, O’Dell M, Metzlaff M, Flavell RB (1996) Details of T-DNAstructural organization from a transgenic Petunia population exhibitingco-suppression. Plant Mol Biol 32: 1197–1203
De Buck S, De Wilde C, Van Montagu M, Depicker A (2000) Determi-nation of the T-DNA transfer and the T-DNA integration frequenciesupon cocultivation of Arabidopsis thaliana root explants. Mol Plant Mi-crobe Interact 13: 658–665
De Buck S, Podevin N, Nolf J, Jacobs A, Depicker A (2009) The T-DNAintegration pattern in Arabidopsis transformants is highly determinedby the transformed target cell. Plant J 60: 134–145
De Neve M, De Buck S, Jacobs A, Van Montagu M, Depicker A (1997)T-DNA integration patterns in co-transformed plant cells suggest thatT-DNA repeats originate from co-integration of separate T-DNAs. PlantJ 11: 15–29
De Paepe A, De Buck S, Hoorelbeke K, Nolf J, Peck I, Depicker A (2009)High frequency of single-copy T-DNA transformants produced by floraldip in CRE-expressing Arabidopsis plants. Plant J 59: 517–527
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for smallquantities of fresh leaf tissue. Phytochem Bull 19: 11
Forsbach A, Schubert D, Lechtenberg B, Gils M, Schmidt R (2003) Acomprehensive characterization of single-copy T-DNA insertions in theArabidopsis thaliana genome. Plant Mol Biol 52: 161–176
Friesner J, Britt AB (2003) Ku80- and DNA ligase IV-deficient plants aresensitive to ionizing radiation and defective in T-DNA integration. PlantJ 34: 427–440
Gallego ME, Bleuyard JY, Daoudal-Cotterell S, Jallut N, White CI (2003)Ku80 plays a role in non-homologous recombination but is not requiredfor T-DNA integration in Arabidopsis. Plant J 35: 557–565
Gelvin SB (2003) Agrobacterium-mediated plant transformation: the biologybehind the “gene-jockeying” tool. Microbiol Mol Biol Rev 67: 16–37
Gelvin SB (2010) Plant proteins involved in Agrobacterium-mediated ge-netic transformation. Annu Rev Phytopathol 48: 45–68
Gheysen G, Montagu MV, Zambryski P (1987) Integration of Agro-bacterium tumefaciens transfer DNA (T-DNA) involves rearrangements oftarget plant DNA sequences. Proc Natl Acad Sci USA 84: 6169–6173
Gheysen G, Villarroel R, Van Montagu M (1991) Illegitimate recombina-tion in plants: a model for T-DNA integration. Genes Dev 5: 287–297
Gorbunova V, Levy AA (1997) Non-homologous DNA end joining in plantcells is associated with deletions and filler DNA insertions. NucleicAcids Res 25: 4650–4657
Herrera-Estrella A, Chen ZM, Van Montagu M, Wang K (1988) VirDproteins of Agrobacterium tumefaciens are required for the formation of acovalent DNA-protein complex at the 59 terminus of T-strand molecules.EMBO J 7: 4055–4062
Janssen BJ, Gardner RC (1990) Localized transient expression of GUS inleaf discs following cocultivation with Agrobacterium. Plant Mol Biol 14:61–72
Johansen LK, Carrington JC (2001) Silencing on the spot: induction andsuppression of RNA silencing in the Agrobacterium-mediated transientexpression system. Plant Physiol 126: 930–938
Kim SI, Veena , Gelvin SB (2007) Genome-wide analysis of AgrobacteriumT-DNA integration sites in the Arabidopsis genome generated undernon-selective conditions. Plant J 51: 779–791
Kim SR, Lee J, Jun SH, Park S, Kang HG, Kwon S, An G (2003) Transgenestructures in T-DNA-inserted rice plants. Plant Mol Biol 52: 761–773
Kononov ME, Bassuner B, Gelvin SB (1997) Integration of T-DNA binaryvector ‘backbone’ sequences into the tobacco genome: evidence formultiple complex patterns of integration. Plant J 11: 945–957
Koukoliková-Nicola Z, Shillito R, Hohn B, Wang K, Van Montagu M,Zambryski P (1985) Involvement of circular intermediates in the transferof T-DNA from Agrobacterium tumefaciens to plant cells. Nature 313:191–196
Krizkova L, Hrouda M (1998) Direct repeats of T-DNA integrated intobacco chromosome: characterization of junction regions. Plant J 16:673–680
Kumar S, Fladung M (2002) Transgene integration in aspen: structuresof integration sites and mechanism of T-DNA integration. Plant J 31:543–551
Kwok WW, Nester EW, Gordon MP (1985) Unusual plasmid DNA orga-nization in an octopine crown gall tumor. Nucleic Acids Res 13: 459–471
Lange M, Vincze E, Møller MG, Holm PB (2006) Molecular analysis oftransgene and vector backbone integration into the barley genomefollowing Agrobacterium-mediated transformation. Plant Cell Rep 25:815–820
Li J, Vaidya M, White C, Vainstein A, Citovsky V, Tzfira T (2005) In-volvement of KU80 in T-DNA integration in plant cells. Proc Natl AcadSci USA 102: 19231–19236
Machida Y, Usami S, Yamamoto A, Niwa Y, Takebe I (1986) Plant-inducible recombination between the 25 bp border sequences of T-DNAin Agrobacterium tumefaciens. Mol Gen Genet 204: 374–382
Makarevitch I, Svitashev SK, Somers DA (2003) Complete sequenceanalysis of transgene loci from plants transformed via microprojectilebombardment. Plant Mol Biol 52: 421–432
Martineau B, Voelker TA, Sanders RA (1994) On defining T-DNA. PlantCell 6: 1032–1033
Maximova SN, Dandekar AM, Guiltinan MJ (1998) Investigation ofAgrobacterium-mediated transformation of apple using green fluorescentprotein: high transient expression and low stable transformation suggestthat factors other than T-DNA transfer are rate-limiting. Plant Mol Biol37: 549–559
Mayerhofer R, Koncz-Kalman Z, Nawrath C, Bakkeren G, Crameri A,Angelis K, Redei GP, Schell J, Hohn B, Koncz C (1991) T-DNA inte-gration: a mode of illegitimate recombination in plants. EMBO J 10:697–704
McCullen CA, Binns AN (2006) Agrobacterium tumefaciens and plant cellinteractions and activities required for interkingdom macromoleculartransfer. Annu Rev Cell Dev Biol 22: 101–127
McVey M, Lee SE (2008) MMEJ repair of double-strand breaks (director’scut): deleted sequences and alternative endings. Trends Genet 24:529–538
Meza TJ, Stangeland B, Mercy IS, Skårn M, Nymoen DA, Berg A,Butenko MA, Håkelien AM, Haslekås C, Meza-Zepeda LA, et al (2002)Analyses of single-copy Arabidopsis T-DNA-transformed lines showthat the presence of vector backbone sequences, short inverted repeatsand DNA methylation is not sufficient or necessary for the induction oftransgene silencing. Nucleic Acids Res 30: 4556–4566
Mysore KS, Bassuner B, Deng XB, Darbinian NS, Motchoulski A, ReamW, Gelvin SB (1998) Role of the Agrobacterium tumefaciens VirD2 proteinin T-DNA transfer and integration. Mol Plant Microbe Interact 11:668–683
Narasimhulu SB, Deng XB, Sarria R, Gelvin SB (1996) Early transcriptionof Agrobacterium T-DNA genes in tobacco and maize. Plant Cell 8:873–886
Offringa R, de Groot MJ, Haagsman HJ, Does MP, van den Elzen PJ,Hooykaas PJ (1990) Extrachromosomal homologous recombination andgene targeting in plant cells after Agrobacterium mediated transforma-tion. EMBO J 9: 3077–3084
Ohta S, Mita S, Hattori T, Nakamura K (1990) Construction and expressionin tobacco of a b-glucuronidase (GUS) reporter gene containing an in-tron within the coding sequence. Plant Cell Physiol 31: 805–813
Plant Physiol. Vol. 160, 2012 521
Complex Extrachromosomal T-DNA Structures
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.

Park S, Vaghchhipawala ZE, Vasudevan B, Waterworth WM, West CE,Gelvin SB, Mysore KS (2011) “Classical” non-homologous end-joiningproteins limit T-DNA integration into the plant genome. In 32th Inter-national Crown Gall Conference 18.
Radchuk VV, Van DT, Klocke E (2005) Multiple gene co-integration inArabidopsis thaliana predominantly occurs in the same genetic locus aftersimultaneous in planta transformation with distinct Agrobacterium tu-mefaciens strains. Plant Sci 168: 1515–1523
Salomon S, Puchta H (1998) Capture of genomic and T-DNA sequencesduring double-strand break repair in somatic plant cells. EMBO J 17:6086–6095
Sparkes IA, Runions J, Kearns A, Hawes C (2006) Rapid, transient ex-pression of fluorescent fusion proteins in tobacco plants and generationof stably transformed plants. Nat Protoc 1: 2019–2025
Spielmann A, Simpson R (1986) T-DNA structure in transgenic tobaccoplants with multiple independent integration sites. Mol Gen Genet 205:34–41
Stahl R, Horvath H, Van Fleet J, Voetz M, von Wettstein D, Wolf N (2002)T-DNA integration into the barley genome from single and doublecassette vectors. Proc Natl Acad Sci USA 99: 2146–2151
Teo CH, Ma L, Kapusi E, Hensel G, Kumlehn J, Schubert I, Houben A,Mette MF (2011) Induction of telomere-mediated chromosomal trun-cation and stability of truncated chromosomes in Arabidopsis thaliana.Plant J 68: 28–39
Thomas CM, Jones JD (2007) Molecular analysis of Agrobacterium T-DNAintegration in tomato reveals a role for left border sequence homology inmost integration events. Mol Genet Genomics 278: 411–420
Timmerman B, Van Montagu M, Zambryski P (1988) vir-induced re-combination in Agrobacterium: physical characterization of precise andimprecise T-circle formation. J Mol Biol 203: 373–384
Tinland B (1996) The integration of T-DNA into plant genomes. TrendsPlant Sci 1: 178–184
Tinland B, Schoumacher F, Gloeckler V, Bravo-Angel AM, Hohn B (1995)The Agrobacterium tumefaciens virulence D2 protein is responsiblefor precise integration of T-DNA into the plant genome. EMBO J 14:3585–3595
Travella S, Ross SM, Harden J, Everett C, Snape JW, Harwood WA (2005)A comparison of transgenic barley lines produced by particle bom-bardment and Agrobacterium-mediated techniques. Plant Cell Rep 23:780–789
Tzfira T, Citovsky V (2006) Agrobacterium-mediated genetic transformationof plants: biology and biotechnology. Curr Opin Biotechnol 17: 147–154
Tzfira T, Citovsky V, editors (2010) Agrobacterium: From Biology to Bio-technology. Springer, New York
Tzfira T, Frankman LR, Vaidya M, Citovsky V (2003) Site-specific inte-gration of Agrobacterium tumefaciens T-DNA via double-stranded inter-mediates. Plant Physiol 133: 1011–1023
Tzfira T, Li J, Lacroix B, Citovsky V (2004) Agrobacterium T-DNA inte-gration: molecules and models. Trends Genet 20: 375–383
Tzfira T, Tian GW, Lacroix B, Vyas S, Li J, Leitner-Dagan Y, KrichevskyA, Taylor T, Vainstein A, Citovsky V (2005) pSAT vectors: a modularseries of plasmids for autofluorescent protein tagging and expression ofmultiple genes in plants. Plant Mol Biol 57: 503–516
Ulker B, Li Y, Rosso MG, Logemann E, Somssich IE, Weisshaar B (2008)T-DNA-mediated transfer of Agrobacterium tumefaciens chromosomalDNA into plants. Nat Biotechnol 26: 1015–1017
Wang K, Stachel SE, Timmerman B, van Montagu M, Zambryski PC(1987) Site-specific nick in the T-DNA border sequence as a result ofAgrobacterium vir gene expression. Science 235: 587–591
Ward JE Jr, Dale EM, Binns AN (1991) Activity of the AgrobacteriumT-DNA transfer machinery is affected by virB gene products. Proc NatlAcad Sci USA 88: 9350–9354
Wenck A, Czakó M, Kanevski I, Márton L (1997) Frequent collinearlong transfer of DNA inclusive of the whole binary vector duringAgrobacterium-mediated transformation. Plant Mol Biol 34: 913–922
Windels P, De Buck S, Depicker A (2010) Patterns of T-DNA integrationinto the host genome. In T Tzfira, V Citovsky, eds, Agrobacterium: FromBiology to Biotechnology. Springer, New York, pp 441–481
Windels P, De Buck S, Van Bockstaele E, De Loose M, Depicker A (2003)T-DNA integration in Arabidopsis chromosomes: presence and origin offiller DNA sequences. Plant Physiol 133: 2061–2068
Wolters AA, Trindade LM, Jacobsen E, Visser RGF (1998) Fluorescence insitu hybridization on extended DNA fibres as a tool to analyse complexT-DNA loci in potato. Plant J 13: 837–847
Zeevi V, Tovkach A, Tzfira T (2008) Increasing cloning possibilities usingartificial zinc finger nucleases. Proc Natl Acad Sci USA 105: 12785–12790
Zhang J, Cai L, Cheng J, Mao H, Fan X, Meng Z, Chan KM, Zhang H, Qi J,Ji L, et al (2008) Transgene integration and organization in cotton(Gossypium hirsutum L.) genome. Transgenic Res 17: 293–306
Zhao X, Coats I, Fu P, Gordon-Kamm B, Lyznik LA (2003) T-DNA re-combination and replication in maize cells. Plant J 33: 149–159
Ziemienowicz A, Tzfira T, Hohn B (2010) Mechanisms of T-DNA inte-gration. In T Tzfira, V Citovsky, eds, Agrobacterium: From Biology toBiotechnology. Springer, New York, pp 395–440
522 Plant Physiol. Vol. 160, 2012
Singer et al.
www.plantphysiol.orgon April 3, 2019 - Published by Downloaded from Copyright © 2012 American Society of Plant Biologists. All rights reserved.





























