Embed Size (px)
Citation preview

101
Failure of Impulse Propagation in a MathematicallySimulated Ischemic Border Zone: Influence of Direction
of Propagation and Cell-to-Cell Electrical Coupling
BARBARA J. MULLER-BORER, PH.D., TIMOTHY A. JOHNSON, PH.D.,
LEONARD S. GETTES, M.D., atid WAYNE E. CASCIO, M.D.
From the Division of Cardiology. Department of Medicine.The University of North Carolina at Chapel Hill. Chapel Hill. North Carolina
Propagation in a Modeled Ischemic Border Zone. Introduction: It is suggested thatheterogeneous extracellular potassium concentration, cell-to-cell coupling, and geometricnonuniformities of the ischemic border zone contribute to the incidence of unidirectional blockand subsequent development of lethal ventricular arrhythmias.
Method and Results: A discrete electrical network was used to model a single cardiac fiberwitb a [K 1 gradient characteristic of an ischemic border zone. Directional differences inpropagation were evaluated by creating discrete regions with increased gap junctional resis-tance within the [K J gradient. Furthermore, the effect of homogeneity/heterogeneity of celllength on impulse propagation through the [K ] . gradient in the presence of increased gapjunctional resistance was evaluated. The results indicate that failure of impulse propagationoccurs at the junction between partially uncoupled and normally coupled cells. Furthermore,propagation failure was more likely to occur as the impulse propagated from a region of high[K' \^ to low [K ] . Heterogeneity in cell length contributes to tbe variability in tbe occurrenceof unidirectional and bidirectional block.
Conclusions: The onset of cellular uncoupling in an iscbemic border zone may interact witbtbe inberent [K ] gradient leading to unidirectional conduction block. Tbis mechanism may beimportant for the generation of reentrant arrbytbmias at the iscbemic border zone. (J Cardio-vasc ElectrophysioL Vol. 6. pp. 1101-1112, December 1995)
conduction, heart, cetl-to-cell conduction, discontinuous electrical propagation, discontinuous cablemodel, action potential propagation, numerical simulations, ischemic boundary, arrhythmia
Introduction
The accumulation of extracellular potassiumand associated membrane depolarization may bethe single most important determinant of impulsepropagation failure iti the ischemic heart.' Yet. theinteractions of cellular K" loss, changes in cellu-lar coupling, and cellular geometry on impulse
Supported by Grants ROI-HL48769. RO1-HL498I8, and POl-HL27430 from the National Institutes of Health.
Address for correspondence: Barbara J, Muller-Borer. Ph,D,.CB#7075. 354 Burnett-Woniack Building. The University ofNorth Carolina at Chapel Hill. Chapel Hill. NC 27599-7075, Fax:9l9-%6-1743.
Manuscript received 7 March 1995; Accepted for publication 28September 1995.
propagation in ischemic myocardium are not wellunderstood. Duting early ischemia, extracellularpotassium concentration ([K+]J increases and fol-lows a triphasic titne course charactetized by arapid initial rise of LK ] ., followed by a secondor plateau phase, which is terminated by a thirdand final phase of continuous cellular K' loss.Along the ischetnic boundary, the second phaseof cellular K^ loss is characterized by a [K l gra-dient with tnaximal concentrations rarely exceeding10 mmol/L and a small change in extracellularpH." Such ionic changes, by themselves, are notlikely to produce failure of impulse propagation^due to the large safety factor for longitudinal con-duction.'*- The failure of impulse propagation thatoccurs under these ionic conditions is likely to re-

1102 Journal of Cardiovascular Electrophysiology Vot. 6, No. 12. December 1995
late to changes in passive properties (i.e., cell-to-cell electrical uncoupling) and enhanced resis-tive discontinuities. This hypothesis is supportedby the experimental observation that failure of itn-pulse propagation during the terminal portion ofthe second phase of cellular K^ loss in ischemicrabbit^ and porcine hearts^ corresponds to theonset of cell-to-cell electrical uncoupling. The rel-evance of this relationship to arrhythmogenesis issuggested hy the observation that cell-to-cell elec-trical uncoupling, evidenced by a secondary risein the whole tissue resistance, has heen shown torelate temporally to the onset of ventricular ar-rhythmias and fibrillation during the lb phase ofventricular arrhythmias in regionally ischemic insitu porcine heart.'
The objective of this study was to evaluate theloss of cell-to-cell electrical interaction within theischemic border zone, simulated by a [K']^ andmembrane potential gradient, on the developmentof unidirectional block at ischemic boundaries. Thiswas accomplished hy studying, on a microscopicscale, the influence of increased gap junctional re-sistance within discrete areas of a modeled isch-emic border zone on failure of impulse propaga-tion. It was hypothesized that the asymmetry ofexcitatory current along the length of a modeledventricular fiber created by the [K^ l,. gradient con-tributes to direction-dependent propagation failurewhen modeled fiber lengths are separated by asmall region of decreased electrical cell-to-cell cou-pling. Experimental data ** and modeling results'"support the role of asymmetry of the preparationin determining directional differences in propaga-tion. Asymmetries created by geometric differencesin fiber length,** membrane areas," and electrotonictransmission'"" may result in propagation disor-ders. Consequently, the role of asymmetry on thedirection of impulse propagation and failure wasinvestigated by introducing a region of increasedgap junction resistance within the [K^]^ gradient.For the purpose of this investigation, a mathe-maticiU model of a single cardiac fiber that includedthe contribution of [K l in the membrane actionpwtential response was used. This model simulatesventricular tissue, incorporafing ionic, resistive, andstructural parameters within physiologic limits, un-like previous modeling studies of the ischemic bor-der zone.'- A simplified ischemic border zone wasmodeled as a [K+]^ gradient spanning 4 to 12mmol/L, distributed over 13 mm. The rationaleto utilize only a [K * ] gradient relates to the factthat the membrane responsiveness and conductionare most strongly determined by the [K^]^ during
ischemia. Finally, the sensitivity of impulse prop-agation to partial cell-to-cell electrical uncouplingand unequal cell lengths was evaluated. Inhomo-geneity in ceil length, as measured by Sommer andScherer,'-* was introduced in the modeled fiber. Thisallowed us to study the effects of geometric andelectrotonic inhomogeneities on failure of impulsepropagation.
Using this model, we show that the onset of cel-lular uncoupling in the ischemic border zone in-teracts with the inherent [K+1 gradient and leadsto unidirectional conduction block. In our model,failure of impulse propagation always occurs atthe junction of partially uncoupled and normallycoupled cells, suggesting that an impedance mis-match at this site contributes to conduction block.Heterogeneity of cellular structure further con-tributes to the occurrence of unidirectional blockunder these modeled conditions. This mechanismmay be important for the formation of reentrantventricular arrhythmias within ischemic boundariesduring ischemia.
Methods
A single cardiac fiber was modeled with the dis-crete electrical network shown in Figure la. Thediscrete electrical network was assembled from abasic membrane unit. The ionic and resistive pa-rameters were scaled to equal the !0-^m lengthand \O-fim diameter of the membrane unit. Themembrane unit consisted of an intracellular resis-tive element, (r ^ , 255 kQ), connected to a non-linear electrical circuit (m) defining the active mem-brane response. The electrical response of the non-linear electrical circuit is defined mathematicallyin Equations 1 through 3.
'ion — ^Na "*• hi + AT "•" ^K!(T)
= C^dVJdt
(1)
(2)
(3)
where I^ = the transmembrane current, !•„„ = theionic current, and I ,,, = the capacitive current.The sodium and calcium currents (I .,, I ,) werepreviously established.'-"'^ The potassium currents(\K^ ^KIIT) represent recent changes that incorpo-rate the [K^Jj. in characterizing the ventricular ac-tion potential waveform."" This allows the [K+lto be set to a given value within the modeled fiber.Currents are given in //A/cm- and voltages in mV.C^ is the specific membrane capacitance (1.0

Mutler-Borer, et al. Propagation in a Modeled Ischemic Border Zone 103
a. Periodic Fiber
m r,
13 mm
gapCnyo
b. Non-periodic Fiber
Figure 1. Schematic diagram of the periodic (a) and non-periodic (b) discrete eiectricat fiber. The modeted cells of the peri-odic ftlyer are WO /im in length (10 membrane units) and !0 fim in diameter. Ttie modeled celts ofttie non-periodic fiber arerandomly assigned lengths from 30 to 130 fim. normally distributed. Each metnbrane unit is composed of a myoplasmic resis-tance (r^,^.J and membrane circuit (m). The modeled celts are connected ttirough a gap junction resistance (r^^p) to form 13-mm tong. modeled fibers.
To form cells, the membrane units are connectedend to end via an intracellular resistor (r ,,,). Eachcell is connected through a gap junction resistor (r^,1275 kU) to form a modeled fiber The standardvalue of v^^ was selected to be approximately fivetimes the value of r , ,,. This resulted in conductionvelocities within physiologic bounds. For the pur-pose of this study, it was assumed that the fiber wassurrounded by an extensive extracellular mediumwith an extracellular resistance of zero. Two fiberswere modeled for this study. The first, a periodicfiber with each cell measuring lCX) /im in length (10membrane units) and 10 ^m in diameter, is illus-trated in Figure 1 a. The inodeled cells are connectedend to end to form a modeled cardiac fiber 13 mmin totid overall length. To introduce geometric in-homogeneities occurring due to unequal cell length.
a second fiber was created as illustrated in Figurelb. This nonperi(xiic fiber with cells ranging in lengthfrom 30 ^m to 130 ^jm, normally distributed, is also13 mm in length. A discrete stepwise [K^]^. gradi-ent was created along each fiber by fixing the IK^l^parameter for each membrane unit. This was ac-complished by incorporating a value of IK^l^. intothe mathematical equations that defined the activemembrane response. The [K^ l gradient ranged ftx)m4.0 to 12.0 mmol/L over the 13 mm length. Each[K^lg section along the gradient was approximately1.3 mm in length with the end sections 2.5 mm (Fig.2). All increments in [K^]^ were made at the cellboundaries and not within a modeled cell. Thisconfiguration resulted in an approximate [K^l^gradient of 8 mmoI/L-cm when measured over the10-mm region of interest. This gradient favorably
2.5 mm l..*3 mm13 miTi
4.0 5.14 6.28 7.42 8.56 9.7 U).S4 12.0
4.0 >.
antejiradestimulu.s
7.42 8.56 9.7 10.84 12.0
mobileresistivebarrier
stimulu.s
Figure 2. Schematic diagram of the tnodeted fiber. The top figure illustrates the extracellular potassium gradient along thefiber tength and approximate length of each extracellular potassium region. The mobile resistive barrier and locations of theantegrade and retrograde stimuli are noted in the bottom figure.

1104 Journal of Cardiovascular Electrophysiology Vot. 6. No. 12. December 1995
compiirss to the 8.3 mmol/L-cm [K+\ gradient, mea-sured at the ischemic border zone.' '**
As it was the intention of this study to evalu-ate directional differences in propagafion, a sin-gle stimulus applied to a resting fiber was deter-mined sufficient to evoke an action potential re-sponse (i.e., rather than the use of premature beatsused to evaluate reentry or reflection). A 15.0-nApulse stimulus was applied for 1.0 msec to a 0.1-mm region to initiate action potential propagationthrough the fiber. Two stimulus sites were lo-cated on the fibers 0.50 to 0.59 mm from eachfiber end (Fig. 2). All measurements were recordedbeginning 1.5 mm distal to the stimulus site. Theregion of interest evaluated and reported includesa total length of approximately 10 mm. This al-lowed action potential propagation to be evaluatedin the modeled liber without stimulus artifact orboundary effects. The direcfion of action potentialpropagation was termed anterograde when the fiberswere stimulaled in the region with [K^]^ = 4mmol/L. When the modeled fibers were sfimulatedin the region with [K^]^ = 12 mmol/L, the direc-tion of propagation was termed retrograde. Onlyone stimulus site was used per simulation. Table1 lists action potential waveform characteristicsmeasured 2 mm beyond the stimulus site and be-tween 2 and 5 mm for conduction velocity.These measurements were acquired from 13-mmlong, modeled periodic fibers with fixed (K ] . val-ues (i.e., no [K^]^ gradient). It should be noted thatshort action potential durations (APDs) and elec-trotonic transmission were observed in the fiberwith a fixed [KM^ of 12 mmol/L and block rapidlyoccurred. For this reason, conduction velocity isnot provided.
To evaluate direcfional effects of partial cell-to-eell uncoupling on signal conduction, a 1.0-mm
TABLE ICharacteristics of Propagating Action Potentials Measured
in Five Individual, Periodic, 13-mni Long Fibers
3 5.4 10 12
RMP(mV) -95.6 -84.0 -78.4 -70.0 -65.4APA(mV) 109.0 100.0 93.0 76.0 43.5V,n. (V/sec) 157.0 162.0 145.0 76.5 2.0APDw (msec) 493 375 311 226 L5.4B (cnVsec) 41.4 43.8 45.0 34.4 —
The IK*] . of the cells in each fiber was set to a single value.All measurements were recorded 1.5 !o 4.5 mm beyond thestimulus site. Action potential propagation was observed inall fibers except in the fiber with a [K*"] of 12 mmol/L. Inthis fiber, decremental conduction and electrotonie trans-mission were observed.
long barrier with increased r p was introduced intoeach modeled cardiac fiber. The barrier was cre-ated by increasing the r..^ within a range of 2 to12 times the standard rj.,,,,.'" The resistive barrierwa.s located in the center of one potassium regionwithin each fiber per simulation as shown in Fig-ure 2. The characteristics of anterograde and ret-rograde propagation were evaluated. In addition,the amount of uncoupling that produced unidirec-tional iind bidirectional block was determined. ITiisprotocol was followed for both the periodic andnonperiodic fibers.
The circuit analyzer with macromodeling(CAzM)^" was used to compute the action poten-tial response as the signal propagated through theelectrical circuit. CAzM, a general purpose circuitsimulator, requires that the circuit be representedin the form of a system of differential equationsa.s previously descrihcd.- ' Each ionic componentis defined as a muhiterminal nonlinear deviceand written as a subcircuit of the total membranecircuit (m). CAzM uses a variable time step (At)during integrafion to provide a small At (~ 30
during depolarization and lai^er At ( - 600during repolarization of the acfion potential.
Results
Periodic Fiber with [K'l^ Gradient
No directional differences in action potentialpropagation were observed in the periodic mod-eled fiber with a [K+1 . gradient. Figure 3 illustratesthe changes in action potential characteristics asthe signal passed through the periodic fiber in theanterograde direcfion. The acfion potenfiiils are sep-iirated by approximately 1.25 mm. As shown inthe figure, as the electrical signal traveled from a[K^l, of 4.0 mmoI/L to a [K+\. of 12.0 mmol/L,the resting membrane potentials (RMPs) becameless negative, the action potential amplitudes (APAs)decrea.sed, and the APDs decreased as expected.Furthermore, the conduction velocity respondedbiphasically. Conduction velocity gradually in-creased from 45 cm/sec to a maximum of 54cm/sec at a (K'l,. of 8.56 mmol/L. The conduc-tion velocity gradually decreased to a minimum of25 cm/sec at a [K | . of 12.0 mmoI/L. During ret-rograde propagation, as the impulse propagatedfrom a region of [K ] . of 12.0 mmol/L to a [K^]^of 4.0 mmol/L, the RMPs became more negative,and the APAs and APDs increa.sed. Overall con-duction velocities were approximately equal dur-ing anterograde and retrograde propagation.

Muller-Borer. et al. Propagation in a Modeled Ischemic Border Zone ! 105
4.0
5.14
6.28
7.42
8.56
9.7
10.84
12.0
450Time (ms)
Figure 3. Action potential waveforms of ihe propagatingsignal through the extracellular potassium gradient. As theaction potential propagated through the modeted fiber, theaction potentiat durations and action potentiat timptitudesdecreased white the resting membrane potential becameless negative.
In the modeled [K+]^ gradient, the electricalsignal propagated through (anterograde) andaway (retrograde) from the IK + J region of 12mmoi/L. This region of the fiber was consid-ered to be at the boundary of the ischemic bor-der zone and ischemic region (i.e., constant high[K^l^). Conduction through this area was un-like that observed in the fiber with a constant[K^]g of 12 mmol/L where electrotonic trans-mission and block were observed. It was ex-pected that conduction block would occur asthe action potential propagated from an areaof low [K^], to high [K*],, In the modeled [K^],gradient, it is likely that distal end effects anda short segment (2.5 mm) of modeled fiber con-taining the [K^] . of 12 mmol/L contributed tothis phenomena. Indeed, when the ends of themodeled fiber were extended to represent re-gions of constant levels of [K" ] , as measuredin normal and ischemic tissue, action potential
propagation blocks as the electrical signal prop-agates into regions of constant high [K+1 . (i.e.,the end of the fiber was extended to include a5.0-mm region with a constant [K + J of 12mmol/L).
Periodic Fiber with
Barrier
Gradient and Resistive
When a high resistive barrier was placed withinthe modeled periodic fiber, directional differencesin action potential propagation were observed. Fig-ure 4a shows a sequence of action potential up-strokes as the electrical impulse propagated thaiugha high resistive barrier in the anterograde directionof the periodic fiber. A section of the modeled fiberis shown. The action potentials are separated by
6.28 7.42 8.56
20
Time (ms)
Figure 4. (a) Action potential up.ttrotces. ittustrating con-duction, during anterograde propagation through a re-sistive barrier, (b) Action potentiat upstrokes, illustratingconduction block, during retrograde propagation. Thesignals illustrate propagation through a {K^}^ of 6.28mM. a IK^j^ of 7.42 mM with r ,,, , increased by 8 times
the baseline r . and .i of 8.56 mM with basetine

1106 Journal of Cardiovascular Electrophysiology Vot. 6, No. 12. December 1995
0.11 mm. These waveforms were observed whena resistive barrier with r ^ = 8 times standard wasplaced in a region of [K" ] = 7.42 mmol/L. Thewaveforms illustrate several action potential char-acteristics common to all simulafions with con-duction through a high resistive barrier, both an-terograde and retrograde: (I) As the electrical sig-nal approached the high resistive barrier, theconducfion velocity increa.sed as evidenced by theclose spacing of the individual action potential up-strokes; (2) As the electrical signal propagatedthrough the high resistive barrier, conduction ve-locity decreased. This is clearly shown by the in-creased distance between the action potential up-strokes; and (3) Upon exiting the resistive barrier,conducfion velocity was restored to the expectedvalue within the [K' \ gradient. In the exampleshown in Figure 4a, the conduction velocitychanged from 55 to 24 cm/sec and back to 47cm/sec in this region.
When the same fiber illustrated in Figure 4a wasstimulated in the region of [K" ] = 12 mmol/L,retrograde propagation was observed and resultedin conducfion block as shown in Figure 4b. TTiewaveforms are characteristic of all simulations re-sulting in conduction block: (1) conduction ve-locity increased as the electrical signal approachedthe resistive bamer; (2) conduction velocity de-creased as the electrical signal propagated throughthe resistive barrier; and (3) slower conduction andelectrotonic transmission were observed in tlie areawith increased gap junction resistance. Beyond the
resistive barrier only electrotonic transmission isobserved.
To continue with this evaluation, the gap junc-tion resistance was further increased to detennineat what level of partial uncoupling bidirectionalblock would be observed. The levels of increasedr p necessary to produce unidirectional and bidi-rectional block in the modeled periodic cardiacfiber were determined for each |K'l^. region (Fig.5). The y-axis is nonnalized to standard r .. val-ues. The value 1 indicates a tightly coupled fiberand 12 indicates one twelffh of the baseline cel-lular coupling. The x-axis is the [K' \. level in thefiber where the resistive barrier was placed duringthe simulation. In the modeled periodic fiber, uni-direcfional block always occurred when the signalpropagated in the retrograde direction. This graphsuggests that in the mid-levels of [K^]^ where ven-tricular fibrillation is shown to occur experiinen-tally (i.e., between a [K"J, of 6.28 and 9.7 mmol/L),a significant amount of uncoupling must take placebefore unidirectional block occurs. Furthennore,within this mid-range of [K" ] , an equal increasein partial cell-to-cell uncoupling occurs as condi-fions progress from unidirectional to bidirectionalblock.
Changes in APA and the maximum change inslope of the action potential upstroke (Y,,;,,) dur-ing anterograde propagation, through the periodicfiber with a [K']^ gradient, both with and withouta resistive barrier are shown in Figures 6a and 6b.In the modeled fiber with no resistive barrier, APA
PS n
•- 5
ilii
12 J1 1 •
10 -
9 i87 - •
6 • •
5 • •
4 -
3 - •
2 - •
1 - -
5.14
Periodic Fiber
O bidirectional block
* unidirectional block
1
6.28 7.42 8.56 9.7 10.84
[K 1 of Resistive Barrier (mM)
Figure 5. Graph illustrating the level of increased r ,,,, of the resistive barrier that produced unidirectional and bidirectionalblock in the periodic fiber. The shaded area represents a window of opportunity when unidirectional block occurs. This win-dow is a constant width for the periodic fiber.
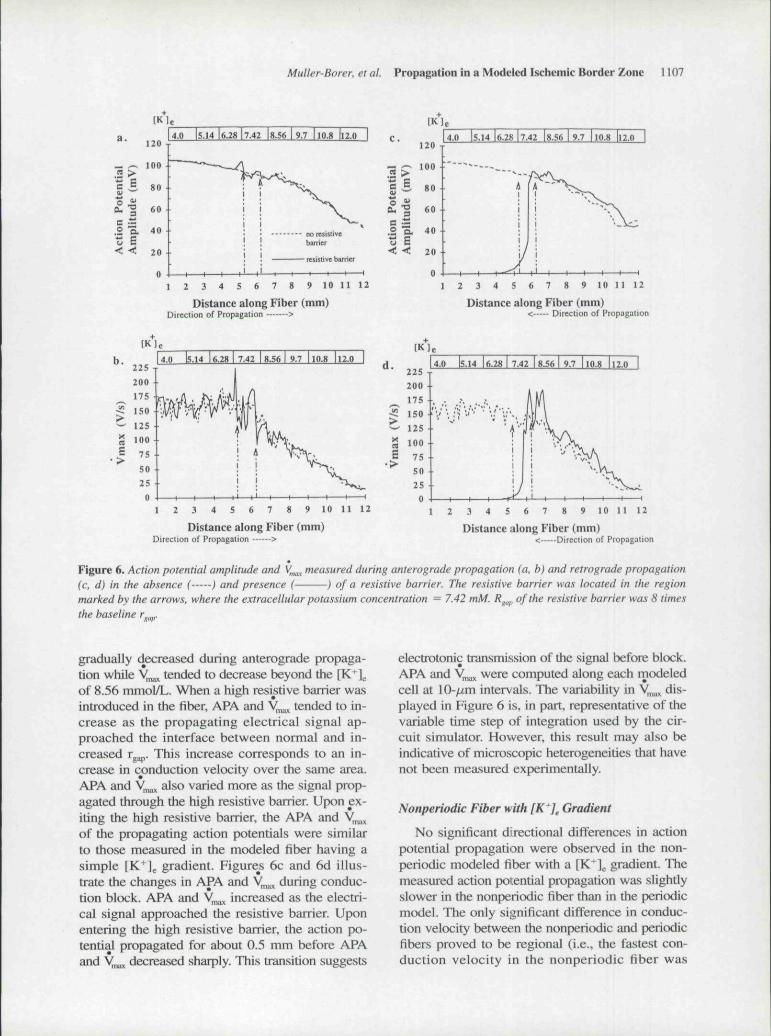
Muller-Borer, et aL Propagation in a Modeled Ischemic Border Zone 1107
4.0 5.14 6.28 7.42 8.56 9.7 10.8 12.0
.2 >
.2 D,t3 E
liO 1
100
80
60
40
20
0
4.0
1—
5.14
• - - - -
—1
6.28
" ~ V
lAl 8.56 9.7 10.8 12.0
\
1 --U;i
j \1 [1 1 1 1 1 1 1
9 10 I I 12
Distance along Fiber (mm)Direclion of Propagation >
3 4 5 6 7 8 9 10 11 12
Distance along Fiber (mm)< Direclion of Propagation
IK l,b.
225
200
175
150
125
100
75
SO
25
0
4.0 15.14
Ww
—1—1—
6.28 7.42
! f
1
i
-1 h* l-l
8.56
;,
1—1—
9.7 10.8 12.0
-1—1—h——(—1
1 2 3 4 5 6 7 8 9 10 11 12
Distance along Fiber (mm)Direction of Propagation >
10 11 12
Distance along Fiber (mm)< Direction of Propagation
Figure 6. Action potential amplitude and V ,, measured during anterograde propagation (a, b) and retrograde propagation(c. d) in the absence ( ) and presence (- ) of a resistive barrier. The resistive barrier was located in the regionmarked by the arrows, where the extracellular potassium concentration = 7.42 mM. R^^^, of the resistive barrier was 8 timesthe baseline r ,.
gradually decreased during anterograde propaga-tion while y,^ tended to decrease beyond the [K ]of 8.56 mmol/L. When a high resistive barrier wasintroduced in the fiber, APA and y^^ tended to in-crease as the propagating electrical signal ap-proached the interface between normal and in-creased rggp. This increase corresponds to an in-crease in conduction velocity over the same area.APA and y,,^ also varied more as the signal prop-agated through the high resistive barrier. Upon ex-iting the high resistive barrier, the APA and V ,of the propagating action potentials were similarto those measured in the modeled fiber having asimple [K*l . gradient. Figures 6c and 6d illus-trate the changes in APA and y,^ during conduc-tion block. APA and y,^ increased as the electri-cal signal approached the resistive barrier Uponentering the high resistive barrier, the action po-tential propagated for about 0.5 mm before APAand y ^ decreased sbarply. This transition suggests
eiectrotonic transmission of the signal before block.APA and y^^ were computed along each modeledcell at 10- ,m intervals. Tbe variability in V^^ dis-played in Figure 6 is, in part, representative of tbevariable time step of integration used by the cir-cuit simulator. However, tbis result may also beindicative of microscopic heterogeneities that havenot been measured experimentally.
Nonperiodic Fiber with [K*]^ Gradient
No significant directional differences in actionpotential propagation were observed in the non-periodic modeled fiber with a \K^\ gradient. Themeasured action potential propagation was slightlyslower in the nonperiodic fiber than in the periodicmodel. The only significant difference in conduc-tion velocity betwetin the nonperiodie and periodicfibers proved to be regional (i.e., the fastest con-duction velocity in the nonperiodic fiber was
1108 Journal of Cardiovascular Electrophysiology VoL 6, No. 12. December 1993
recorded in the region where the fK"]^ = 7.42mmol/L while the fastest conduction velocity inthe periodic fiber was recorded in the region wherethe [K J = 8.56 mmol/L).
Nonperiodic Fiber withBarrier
Gradient and Resistive
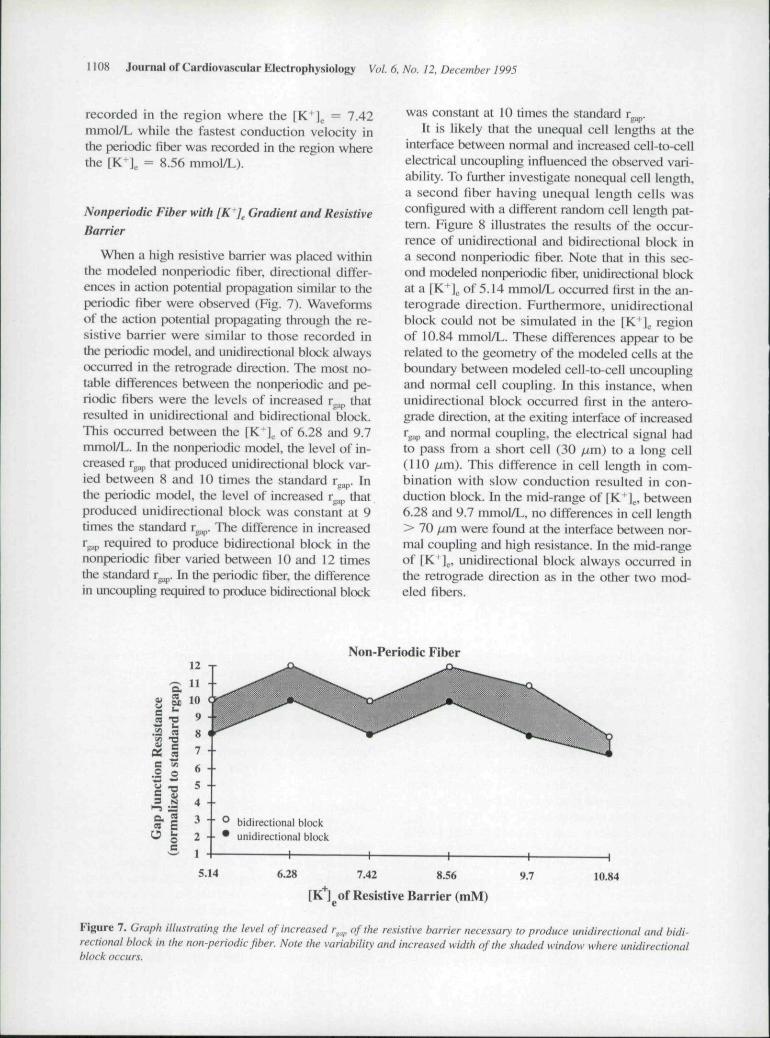
When a high resistive barrier was placed withinthe modeled nonperiodic fiber, directional differ-ences in action potential propagation similar to theperiodic fiber were observed (Fig. 7). Waveformsof the action potential propagating through the re-sistive barrier were similar to those recorded inthe periodic mtxiel, imd unidirectional block alwaysoccurred in the retrograde direction. The most no-table differences between the nonperiodic and pe-riodic fibers were the levels of increased r that
gap
resulted in unidirectional and bidirectional block.This occurred between the [K" | of 6.28 and 9.7mmoI/L. Ill the nonperiodic model, the level of in-creased r^ that produced unidirectional block var-ied between 8 and 10 times the standard r p. Inthe peritxlic model, the level of increased r^^ thatproduced unidirectional block was constant at 9times the standiu-d r p. The difference in increasedT^^^ required to produce bidirectional hlock in thenoaperiodic fiber varied between 10 and 12 timesthe standard r ,, ,. In ihe periodic fiber, the differencein uncoupling requited to produce bidirectional block
was constant at 10 times the standard rg .It is likely that the unequal cell lengths at the
interface between nonnal and increased cell-to-cellelectrical uncoupling influenced the observed vari-ability. To further investigate nonequal cell length,a second fiber having unequal length cells wasconfigured with a different random cell length pat-tern. Figure 8 illustrates the results of the occur-rence of unidirectional and bidirectional block ina second nonperiodic fiher. Note that in this sec-ond modeled nonperiodic fiber, unidirectional blockat a [K l . of 5.14 mmol/L occurred first in the an-terograde direction. Furthermore, unidirectionalblock could not be simulated in the fK+] regionof 10.84 mmoI/L. These differences appear to berelated to the geometry of the modeled cells at theboundary between modeled cell-to-cell uncouplingand normal cell coupling. In this instance, whenunidirectional hlock occurred first in the antero-grade direction, at the exiting interface of increasedr 3,, and normal coupling, the electrical signal hadto pass from a short cell (30 fim) to a long cell(110 fim). This difference in cell length in com-bination with slow conduction resulted in con-duction block. In the mid-range of [K J ., between6.28 and 9.7 mmol/L, no differences in cell length> 70 fjm were found at the interface hetween nor-mal coupling and high resistance. In the mid-rangeof [K+] , unidirectional hlock always occurred inthe retrograde direction as in the other two mod-eled fibers.
tanc
e
.i2f^
ion
net
3
aa
O
,,^anS£
L.M
T3B
o•a
,Sn£uo
1211
10
9
87
6
5
43
2
1
5 14
Non-Periodic Fiber
' bidirectional block- • * unidirectional block
7.42 8.56 9.7
1
10.84
[K ] of Resistive Barrier (mM)
Figure 7. Graph illustrating the level of increased r^^,^ of the resi.\iive barrier neces.sary to produce unidirectional and bidi-rectional block in the non-periodic fiber. Note the variability and increased width of the shaded window where unidirectionalblock occurs.

.X O
Muller-Borer. et al. Propagation in a Modeled Ischemic Border Zone 1109
Non-Periodic Fiber12 T
— 5 - •
N 4 - •
® bidirectional block• unidirectional block
5.14 6.28 742 8.56 9.7
[K ]_of Resistive Barrier (mM)
Figure 8. The level of increased r^^ that resulted in unidirectional and bidirectiotial block in a non-periodic fiber with a differ-ent random urrungement of unequal length celts. In this modeled fiber, unidirectional hlock was observed first in the antero-grade direction in the region of[K'J^ = 5.14 mM (noted by the "X"). Furtheimore, unidirectional block was not obsen-ed inthe region of[K^]^ = 10.84 mM. The width of the shaded window has decreased but remains variable as in Figure 7.
Discussion
This article describes a method to investigatedirectional differences in impulse propagationthrough a modeled cardiac fiber. Computer mod-els have established that inhomogeneities in cell-to-cell coupling, tissue excitability, and tissue struc-ture produce unidirectional block of propagatingaction potentials.'"'^^ This study is unique as itattempts to combine the active and passive mem-brane property characteristics of the ischemic bor-der zone and examine their interaction, thus sim-ulating the changing ionic environment within phys-iologic bounds. Heterogeneous changes in IK^],.,cell-to-eell coupling, and cell length were of par-ticular consideration. The simulations present foursignificant effects. First, the failure of action po-tential propagation through a modeled excitablecardiac fiber with a [^'1^ gradient of 8 mmol/Lwas not directionally dependent. That is, despitegraduiil changes in conduction velocity, APD, APA,and V,,,., , no significant differences were ob-served in action potential propagation throughthe simulated border zone. However, when the endsof the modeled border zone were extended to sim-ulate normal and ischemic tissue, conduction blockwas observed as expected. Second, inhomogene-ity in cell length, which contributes to geometricinhomogeneity, did not result in impulse propa-gation failure through a tighdy coupled fiber witha [K^]g gradient. Third, when a region of par-
tially uncoupled cells was introduced in a modeledfiber with a IK^l^ gradient, impulse propagationfailure was shown to be directionally dependent.Finally, inhomogeneity in cell length appeared tocontribute only to the variability in the increase inr yp needed to produce conduction block.
Slow Conduction
The first two findings suggest that when themodeled fiber is fightly coupled, the gradual changein IK l and geometric inhomogeneity together donot result in propagation failure across an ischemieborder zone. Other factors in addition to hetero-geneous [K^]e and cell length must interact. How-ever, this model adequately represents conductionvelocity and V ^ changes in action potential prop-agation as measured experimentally in regionallyischemic in situ porcine hearts.-^ As the actionpotential propagated from a IK ] of 4 to 8 mmol/L,conduetion velocity increased and y ^ showed lit-tle change. As the action potential propagated be-yond the region with 8 mmol/L, conduction ve-locity decreased, as did V ^ . These changes cor-respond with an increase in RMP.
When eell-to-cell electrical uncoupling was su-perimposed on the IK ] gradient, uniditiectional andbidirectional block were observed. In addition, in-homogeneities in eell length appear to contribute tothe degree of cell-to-cell eiectrieal uncoupling re-sulting in conduction block. These two factors are

10 Journal of Cardiovascular Electrophysiology Vot. 6, No. 12. December 1995
associated with slow conduction and spatial dis-continuities that contribute to arThythmogenic be-havior. Kleber et al.-^ have shown experimentallythat, during ischetnia, the whole tissue resistance in-creases, marking the onset of cellular uncoupling.The increase in whole tissue resistance is shown tooccur between 10 and 20 minutes following the on-set of ischemia. Furthermons, Smith et al. concludedthat an increase in whole tissue resistance measuredin the ischetnic zone coincided with increased lev-els of extracellular potassium during the lb phase,resulting in an increased incidence in ventricular ar-rhythmias. While there is no experimental evidenceto indicate whether cellular uncoupling is homoge-neous or heterogeneous in the ischemic border zone,oitr simplitied approach in evaluating directional dif-ferences of conduction demonstrates that propaga-tion failure results when [K+1^ nonuniformity existsin the presence of cell-to-cell uncoupling (i.e., slowconduction). Unidirectional block occuned most of-ten within the physiologic [K+]^ range of 6 to 10mmol/L, corresponding to [K^]^ levels where ven-tricular fibrillation and block occur in experimen-tal preparations.-'' Furthermore, a significant amountof cell-to-cell uncoupling was necessary to produceunidirecdonai block, which relates temporally to thelb phase. Increasing celi-to-cell uncoupling by 8%to 16% beyond the level of unidirectional blockresulted in bidirectional block. This increase in cell-to-cell uncoupling created a zone (shown in Figs.5, 7, and 8) in which inhomogeneities in cell-to-cellcoupling provided favorable conditions for the de-velopment of reentrant circuits.
It was the intent of this study to evaluate slowconduction due to cell-to-^cell electrical uncouplingin an ischemic border zone. Yet, geometric het-erogeneity at branching sites and anatomic het-erogeneities of collagenous and injured tissue mayinfluence propagation and wavefront curvature inischemic tissue. While it was not within the scopeof this study to evaluate two-dimensional propa-gation, it is evident that factors which influenceslow conduction in an ischemic border zone con-tribute to propagation disorders.
Asymmetry
The asymmetry in [K* ] on either side of the re-sistive barrier appeared to play a significant rolein determining conduction block. Ptiopagadon tendedto block most often in the retrograde direction (i.e.,from high to low {K^\) whether the membrane ar-eas on either side of the resistive barrier were equalor asymmetric. The interface where block occurred
represented a step decrease in cell-to-cell electricaluncoupling and a decrease in [K+J^. Conductionblock is expected with an increase in [K^ J or r jp.These findings illustrate the interaction of asym-metric depression of excitability and slow conduc-tion. Although [K' \ decreased along the modeledfiber at discrete intervals defined by the model struc-ture, changes in RMP and APD were gradual asthe ionic changes were infiuenced by neighboringelements. These gradual changes are illustrated inFigure 4a. Action potendal characteristics measuredat each end of a fK^]^ region were not equal, rep-resendng an asymmetry in tissue excitability, i.e.,a tK" ]e change with each 10-yum step.
While the [K^]^. gradient in the presence of cell-to-cell uncoupling infiuenced die direcdon of acdonpotential block, inhomogeneity in cell length alsocontributed to the variability of propagation failure.Yet, this factor was not significant in determiningpropagation failure except when the difference incell size was > 70 ^,m at the boundary betweenpartially coupled and dghtly coupled regions. Thisresult, while not significant in this evaiuadon, doessuggest that geometric or electrotonic asymmetrymay play a role in conducdon block. Jalife and Moe.^showed experimentally that asymmetry in fiberlengths separated by a resisdve barrier would resultin unidirecdonai block. That is, propagadon acrossa resisdve bamer was more likely to fail when prop-agating from a short to long segment rather thanfrom a long to short segment. These results werefurther demonstrated in a single fiber model byJoyner et al.,'" who showed that asymtnetry in mem-brane area contributes to unidirectional block acrossa resisdve barrier. More recently, Joyner et al.^ in-vesdgated impulse conducdon between asymmetriccell pairs and the effect of increased intercellular re-sistance. Again, this study agrees widi findings that,for equal levels of cell coupling, signal conductionis more likely to block in the direcdon of short tolong cells. As the length of a cell decreases, its in-put impedance increases and less current is requiredto bring it to threshold. Conversely, as the length ofa cell increases, its input impedance decreases andmore current is required to bring it to threshold.Indeed, at the cellular level, asymmetry in cell lengthoffers an explanation for the variability in cell-to-cell electrical uncoupling t^esulting in acdon poten-dal block. In the modeled periodic fiber, the inputimpedance was equal for each cell and an equalamount of current was requit^ to bring each cellto threshold. However, in the modeled nonperiodicfiber, unequal cell size created regional diiferencesin the electrical load at the modeled cell boundaries.

Muller-Borer. et aL Propagation in a Modeled Ischemic Border Zone 111
These differences in eiectrotonic lengths may alsobe present at branching sites and within fiber bun-dle geometries. Indeed, the eiectrotonic length ratherthan membrane properties is thought to attribute todifferential conduction block at axonal bifurcations."
Reflection
It has been suggested that an inci^eased resistiveor [K^]^ barrier may produce reflection in cardiactissue.-^ Reflection in experimental preparations andcomputer models is often shown to occur betweenproximal and distal regions with nornial membranecharacteristics connected by a region of depressedconduction.^'-^ For reflection to occur between tworegions, the conduction delay must be longenough for the proximal fibers to recover from therefractory state before the stimulus arrives from thedistal region. It is unlikely that this model of theischemic border zone would exhibit reflection. Thisis due to the modeled fiber length of 10 mm andthe I-mm long, resistive barrier. As shown in Fig-ure 3, the action potentials generated in the regionof [K^]^. = 4 mmol/L were in an early phase ofrepolarization when the signal nsached the distal por-tion of the modeled fiber. Simulations with a singlefiber F*urkinje modeP suggest that the occurrenceof reflection depends on the magnitude of the de-lay and lengths of the proximal and distal segmentsseparated by the delay. Based on our simulation re-sults, the lengths of the proximal and distal seg-ments and the conduction delays established by in-creased cell-to-cell uncoupling either did not allowthe proximal region to recover sufficiently or thedistal region was not able to re-excite the proximalregion to yield reflection. This was true for both an-terograde and retrograde conduction. While it is pos-sible to show reflection through microcireuits withpathways of only a few millimeters in modeledfibers,-** extracellular recordings from the ischemicborder in whole tissue prepiirations have not recordedthis activity.-**"^ It is likely thai if the region of in-creased cell-to-ceil uncoupling were letigthened. inthe [K^]g gradient model, the proxitnal-distal delaywould be sufficient to cause reflection. However,reflection was not the ftKus of this study, and fur-ther research is necessary to evaluate differencesin tissue excitability of the proximal and distal re-gions and the affect on arrhythmogenesis.
Limitations
This single fiber model does not allow for mul-tidimensional analysis, yet, these results are
in understanding propagation through the ischemicborder zone. Cardiac tissue is composed of bun-dles of cardiac fibers with frequent branching ofthe bundles. An elecdiciU signal propagatitig throughthe tissue will encounter geometric and eiectrotonicheterogeneities such as areas of increased/decreaseddiameter, frequency, and magnitude changes in r pand different branching configurations. In the isch-emic border zone, these changes will be superim-posed on a [K" ] gradient as well as other ionic in-homogeneities. This model is an inifial attempt tounderstand how some of these factors interact. Toadd to tliis complexity, extracellular resistance, whichis known to affect impulse propagation in ventric-ular muscle"' and modeled with the bidornain ap-proach,^' is not addressed in this model. The ex-tracellular resistance is important, as anisotropieswill result in wavefonn changes that are difficultto predict with a single fiber model. In the singlefiber model, adding a homogeneous extracellularresistance would slow impulse conduction overalland may reduce the amount of cell-to-cell uncou-pling necessary for unidirectional and bidirectionalblock. Adding a heterogeneous extracellular resis-tance would also affect impulse conduction, pos-sibly altering directional differences in conducfionblock. Presently, we are not able to add this levelof complexity to tbe model. Finally, the mathe-matical model uses the slow inward current de-scribed by Beeler-Reuter'** and does not includeother dynamic changes in ionic concentrations (i.e.,Na-Ca exchange), L- and T-type calcium currents,or the role of intracellular calcium concentration ininacfivation.
In conclusion, the single fiber model providesa simplified method to investigate the effects ofcell-to-cell coupling and cell geometry on electri-cal impulse conduction in a mcxieled ischemic bor-der zone. The results suggest that heterogeneitiesmodeled by a [K^]^ gradient and modifications incell-to-cell coupling and tissue structure interact toalter the direction of propagation in modeledventricular tissue. Furthermore, it is shown that acotnbination of factors contributes to differentialconduction block in the formation of lethal ven-triculai' arrhythtnias.
References
1. Morena H, Janse MJ, Fiolet JWT, et al: Comparison ofthe effects of regional ischemia, hypoxia. hyperkalemiaand acidosis on intracellular and extracellular poten-tials and metabolism in the isolated porcine heart. CircRes 1980:46:634-646.

1112 Journal of Cardiovascular Electrophysiology VoL 6. No. 12, December 1995
2. Hill JL. Gettes LS: Effect of acute coronary artery oc-clusion on local myocLirdial extracellular K' activity inswine. Circulation 19S0;61:768-778.
3. Kagiyama Y. Hill JL, Gettes LS: Interaction of acido-sis and increased extracellular potassium on action po-tential characteristics and conduction in guinea pigventricular muscle. Circ Res 1982:51:614-623.
4. Delgado C. Steinhaus B. Delmar M. et al: Directionaldifference in excitability and margin of safety for prop-agation in sheep ventricular epicardial muscle. CircRes 1990:67:97-110.
5. Vorperian VR, Wisialowski TA, Deegan R, et al: Ef-fect of hypercapnic acidemia on anisotropic propaga-tion in canine ventricle. Circulation I994;9O:456-461.
6. Cascio WE. Yan G-X, Kleber AG: The relationshipamong aetive and passive electrical properties, me-chanical activity and extracellular potas.sium in arteri-ally perfused and ischemic rabbit ventricular muscle.Effects of calcium entry blockade or hypocalcemia.Circ Res I99O;66:I46M473.
7. Smith WT, Fleet WF. Johnson TA, et al: The I-B phaseof ventricular arrhythmias in ischemic in situ porcineheart is related to changes in cell-to-cell electrical un-coupling. Circulation 1994:90:1-524.
8. Jalife J. Moe GK: Excitation, conduction and retlectionof impulses in isolated bovine and canine cardiac Pur-kinje tibers. Circ Res 1981:49:233-247.
9. Joyner RW. Sugiura H, Tan RC: Unidirectional blockbetween isolated rabbit ventricular cells coupled by avariable resistance. Biophys J 1991:60:1038-1045.
10. Joyner RW. Veenstra R. Rawling D. et al: Propagationthrough electrically coupled ceils: Effects of a resistivebarrier. Biophys J I9«4;45:1017-1024.
11. Stockbridge N, Stockbridge LL: Differential conduc-tion at axonal bifurcations. I. Effect of eiectrotoniclength. J Neurophysioi 1988:59:1277-1285.
12. Sahakian AV. Myers GA. Maglaveras N: Unidirec-tional block in cardiac tibers: Effects of discontinuitiesin coupling resistance and spatial changes in restingmembrane potential in a computer simulation study.IEEE Trans Biomed Eng 1992:39:510-522.
13. Sommer JR. Scherer B: Geometry of cell and bundleapposition in cardiac muscle: Light microscopy. Am JPhysiol 1985:248:H792-H803,
14. Ebihara L. Johnson EA: East sodium current in cardiacmuscle: A quantitative description. Biophys J 1980;.12:779-790.
15. Beeler GW. Reuter H: Reconstruction of the action po-tential of ventricular myocardial (ibres. J Physiol(Lond) 1977:268:177-210.
16. Luo C. Rudy Y: A model of the ventricular cardiac ac-tion potential depolarization, repolarization and theirinteraction. Circ Res 1991:68:1501 -1526.
17. Coronel R, Eiolet J. Wilms-Schopman FJ. et al: Distri-bution of extracellular potassium and its relation to
electrophysiologic changes during acute myocardialischemia in the isolated perfused porcine heart. Circu-lation 1988;77:1125-1138.
18. Johnson TA, Engle CL, Boyd LM, et al: Magnitudeand time course of extracellular potassium inhomo-geneities during acute ischemia in pigs, effect of verap-amil. Circulation 1991:83:622-634.
19. Israel D, Edell DJ. Mark RG: Time delays in propaga-tion of cardiac action potentials. Am J Physiol1990:258:H!906-HI917.
20. Erdman DJ, Hooke N. Muller-Borer BJ: Simulation ofcardiac action potentials using a table based circuitsimulator. Technical Report tr9O-IO. MicroelectronicsCenter of North Carolina. 1990.
21. Muller-Borer BJ. Erdman DJ. Buchanan JW: Electricalcoupling and impulse propagation in anatomicallymodeled ventricular tissue, IEEE Trans Biomed Eng1994:41:445-454.
22. Quan R. Rudy Y: Unidirectional block and reentry ofcardiac excitation: A model study. Circ Res 1990;66:367-382.
23. Gettes LS, Buchanan JW. Tomoaki S: Studies con-cerned with slovi' conduction. In Zipes DP. Jalite J.eds: Cardiac Electrophysiology and Arrhythmias.Grune and Stratton, Inc., Orlando. 1985. pp. 81-87.
24. Kleber AG. Riegger CB. Janse MJ: Electrical uncou-pling and increase of extracellular resistance after in-duction of ischemia in isolated, arterially perfused rab-bit papillary muscle. Circ Res 1987:61:271-279.
25. Fleet WF, Johnson TA. Graebner CA. et al: Effects ofverapamil on ischemia-induced changes in extracellu-lar K'. pH and local activation in the pig. Circulation1986:73:837-846.
26. Antzelevitch C. Jaiife J, Moe GK: Characteristics ofreflection as a mechanism of reentrant iurhythmias andits relationship to parasystole. Circulation 1980;61:182-191.
27. Cabo C. Barr R: Retlection after delayed excitation in acomputer model ot" a single fiber. Circ Res 1992:71:260-270.
28. Janse MJ. Cinca J. Morena H. et al: The "border zone"in myocardial ischemia: An electrophysiological. meta-bolic and histochemical correlation in the pig heart.Circ Res 1979:44:576-588.
29. Janse MJ. van Capelle FJL. Morsink H. et al: Flow of"injury" current and patterns of excitation during earlyventricular arrhythmias in acute regional myocardialischemia in isolated porcine and canine hearts: Evi-dence for two different arrhythmogenic mechanisms.Circ Res 1980:47:151-165.
30. Kieber AG, Riegger CB: Electrical constants of arteri-ally perfused rabbit papillary muscle. J Physiol1987:385:307-324.
31. Henriquez C. Plonsey R: Simulating the electrical be-havior of cardiac tissue using the bidomain model. CritRev Biomed Eng 1993;21:1 -77.






























