Embed Size (px)
Citation preview

PRIRODOSLOVNO-MATEMATIČKI FAKULTET BIOLOŠKI ODSJEK
Slavko Kepec
EKOTOKSIKOLOŠKA
I MIKROBIOLOŠKA KARAKTERIZACIJA
PROČIŠĆENIH OTPADNIH VODA
GRADA VIROVITICE
DOKTORSKI RAD
Zagreb, 2016.

FACULTY OF SCIENCE DEPARTMENT OF BIOLOGY
Slavko Kepec
ECOTOXICOLOGICAL AND
MICROBIOLOGICAL
CHARACTERIZATION OF TREATED
WASTEWATER OF THE CITY OF
VIROVITICA
DOCTORAL THESIS
Zagreb, 2016.

Ovaj je doktorski rad izraĎen u Institutu RuĎer Bošković u Zagrebu, Laboratoriju za
biotehnologiju u akvakulturi (stari naziv: Laboratorij za ihtiopatologiju – biološke
materijale), Biološkom odsjeku Prirodoslovno-matematičkog fakulteta u Zagrebu i poduzeću
Virkom d.o.o. Virovitica, pod vodstvom mentorice dr.sc. Natalije Topić Popović i mentora
prof.dr.sc. Gorana Klobučar, u sklopu Sveučilišnog poslijediplomskog doktorskog studija
biologije.

Zahvale
Prije svega, najiskrenije se zahvaljujem mojoj mentorici dr.sc. Nataliji Topić Popović i
mojem mentoru prof.dr.sc. Goranu Klobučaru, bez čijeg strpljenja i velike pomoći, kroz cijelo
vrijeme, sve ovo ne bih mogao privesti kraju.
Zahvaljujem se ponaosob svima u Laboratoriju za biotehnologiju u akvakulturi Instituta
RuĎer Bošković u Zagrebu na pomoći u stručnom dijelu, na razumijevanju i podršci u
svakom smislu.
Ogromnu zahvalnost dugujem mojoj dragoj suradnici iz Tvornice šećera Virovitica, Kseniji
Zidar, dipl.ing. zbog svega što je učinila kako bi se istraţivanje moglo provesti na pravi
način.
Zahvaljujem mojim dragim kolegicama Blaţenki Čeleš, dipl.ing. i Ljiljani Jarčov,
mag.ing.univ.spec. iz Zavoda za javno zdravstvo Bjelovarsko bilogorske ţupanije, Mirjani
Špehar, mag.med.biochem. iz Zavoda za javno zdravstvo „Sveti Rok“ Virovitičko-podravske
ţupanije, dr.sc. Ljiljani Čačić, dr.sc. Ţeljki Romić i Antoniji Kezerle, dipl.ing. iz Vodovoda
Osijek, na pomoći u stručnom dijelu i moralnoj podršci. Hvala Draţenki Stipaničev, dipl.ing.
i Siniši Repecu, dipl.ing. iz Glavnog vodnogospodarskog laboratorija Hrvatskih voda na
njihovom doprinosu.
Posebno se zahvaljujem mojim prijateljima Denisu Balaţu, dipl.ing. i Marijanu Mlinariću iz
Tvornice šećera, Draţenu Viljevcu iz Brezovice, mojem nećaku Ivanu, i ostalima, na pomoći u
izlovu riba.
Zahvaljujem se direktorima Virkom-a, Zdravku Peru, dipl.ing. na pomoći u početku
doktorskog studija i Damiru Mareniću, dipl.ing. na razumijevanju i velikoj podršci u svakom
smislu. Hvala i ostalima iz Virkom-a koji su u mene vjerovali.
Zahvaljujem i onima koje nisam naveo imenom, a koje sam u poslovnoj suradnji, kroz dugi
niz godina, susretao i zavolio.
Hvala cijelom mom obiteljskom krugu na inspiraciji i strpljenju.

Sveučilište u Zagrebu Doktorski rad
Prirodoslovno-matematički fakultet
Biološki odsjek
EKOTOKSIKOLOŠKA I MIKROBIOLOŠKA KARAKTERIZACIJA PROĈIŠĆENIH
OTPADNIH VODA GRADA VIROVITICE
SLAVKO KEPEC
Prirodoslovno-matematički fakultet, Biološki odsjek
Analiza pročišćenih otpadnih voda koje se ispuštaju u vodene prijemnike uglavnom
podrazumijeva tek utvrĎivanje koncentracije organskih tvari (kemijske i biološke potrošnje
kisika) i nutrijenata (dušika i fosfora). Stoga je u ovom radu ispitan i učinak efluenta na
bioindikatorske organizme (ribe (Carassius gibelio)) i gujavice (Eisenia fetida, Oligochaeta))
i stupanj rezistentnosti bakterijskih izolata na antimikrobne lijekove podrijetlom iz otpadnih
voda, s ciljem utvrĎivanja štetnog djelovanja na ţiva bića u vodi, a posredno i na zdravlje
ljudi. U tom smislu provedena su ekotoksikološka ispitivanja na bioindikatorskim modelnim
organizmima, izvršeno je mjerenje mehanizma multiksenobiotičke otpornosti i vrijednosti
parametara oksidacijskog stresa. Citogenetska oštećenja utvrĎivana su mikronukleus testom a
mutageni učinci Ames testom. Izvršena je komparativna mikrobiološka analiza standardnim
mikrobiološkim testovima i MALDI-TOF spektrometrom masa, te su provedena hematološka
i histopatološka istraţivanja. Rezultati navedenih istraţivanja pokazuju da pročišćene otpadne
vode unošenjem ksenobiotika mijenjaju okolišne parametre recipijenta, dovode do porasta
otpornosti bakterija na antibiotike, histopatoloških promjena i oksidacijskog stresa u riba.
Dobiven je cjeloviti prikaz utjecaja pročišćenih otpadnih voda na ključne prarametre kakvoće
i ekološkog stanja vode i biote.
115 stranica/ 36 slika/ 17 tablica/ 165 literaturnih navoda/ jezik izvornika hrvatski
Ključne riječi: ureĎaj za biološko pročišćavanje, efluent, bakterije, ribe, genotoksičnost,
mehanizma multiksenobiotičke otpornosti, mikronukleus, oksidacijski stres
Mentori: dr.sc. Natalija Topić Popović, znanstvena savjetnica, IRB, Zagreb
prof.dr.sc. Goran Klobučar, PMF, Zagreb
Ocjenjivači:
1. Dr.sc. Ivančica Strunjak-Perović, viša znanstvena savjetnica, IRB, Zagreb
2. Prof.dr.sc. Mirjana Pavlica, PMF, Zagreb
3. Izv.prof.dr.sc. Jasna Hrenović, PMF, Zagreb
4. Zamjena: Dr.sc. Rozelindra Čoţ-Rakovac, znanstvena savjetnica, IRB, Zagreb
Rad prihvaćen:

University of Zagreb Doctoral thesis
Faculty of Science
Department of Biology
ECOTOXICOLOGICAL AND MICROBIOLOGICAL CHARACTERIZATION OF
TREATED WASTEWATER OF THE CITY OF VIROVITICA
SLAVKO KEPEC
Faculty of Science, Department of Biology
Currently, analysis of treated wastewater discharged from the wastewater treatment plants
(WWTP) inflowing into recipients involves determination of concentrations of organic
substances (chemical and biological oxygen demand) and nutrients (nitrogen and
phosphorus). In this study, the impact of effluent on organismic bioindicators and degree of
resistance of bacterial isolates to antimicrobial drugs originating from wastewater has been
evaluated with the aim to determine toxic effects on aquatic (fish, Carassius gibelio) and
terrestrial (earthworm, Eisenia fetida, Oligochaeta) organisms, and indirectly on human
health. The ecotoxicological testings on bioindicator model organisms were conducted,
measuring mechanisms of multixenobiotic resistance and parameters of oxidative stress.
Cytogenetic damages were determined by micronucleus test, and mutagenic effects with
Ames test. Comparative microbiological analyses were made with conventional methods and
mass spectrometry analyser. Results of this study show that treated wastewater (leaving
WWTP) changed environmental parameters by introduction of xenobiotics, contributed to
microbial contamination and antimicrobial resistance, histopathological tissue changes and
oxidative stress of examined organisms. Hencewith, our approach encompassed an integrated
overview of the impact of treated wastewater on key environmental and organismal
parameters.
115 pages/ 36 figures/ 17 tables/ 165 references/ original in Croatian
Keywords: wastewater treatment plant, effluent, bacteria, fish, genotoxicity, mechanism of
multixenobiotic resistance, micronuclei, oxidative stress
Supervisors:
Natalija Topić Popović, Ph.D., Senior Scientist, RBI, Zagreb
Goran Klobučar, Ph.D., Full Professor, PMF, Zagreb
Reviewers:
1. Ivančica Strunjak-Perović, Ph.D., Senior Scientist, RBI, Zagreb
2. Mirjana Pavlica, Ph.D., Full Professor, PMF, Zagreb
3. Jasna Hrenović, Ph.D., Associate Professor, PMF, Zagreb
Replacement: Rozelindra Čoţ-Rakovac, Ph.D., Senior Scientist, RBI, Zagreb
Thesis accepted:

SADRŢAJ
SADRŢAJ ..............................................................................................................................7
1. UVOD ...........................................................................................................................1
1.1. Cilj i značaj istraţivanja............................................................................................3
2. LITERATURNI PREGLED ..........................................................................................6
2.1. UreĎaji za biološko pročišćavanje otpadnih voda ......................................................6
2.2. Fizikalno-kemijske analize .......................................................................................7
2.3. Biološke analize utjecaja onečišćenja na okoliš ........................................................7
2.4. Mikrobiološka istraţivanja ..................................................................................... 10
2.5. Histološke pretrage ................................................................................................. 11
2.6. Biokemijske i hematološke pretrage krvi ................................................................ 12
2.7. Ekotoksikološki testovi na modelnim organizmima ................................................ 13
2.8. Mehanizam multiksenobiotičke otpornosti .............................................................. 13
2.9. Mikronukleus test ................................................................................................... 14
2.10. Oksidacijski stres ................................................................................................ 15
2.11. AMES test .......................................................................................................... 16
3. MATERIJALI I METODE .......................................................................................... 18
3.1. UreĎaj za biološko pročišćavanje otpadnih voda i lokacije za uzimanje uzoraka ..... 18
3.2. OdreĎivanje fizikalno-kemijskih pokazatelja .......................................................... 22
3.3. Analitika vode i sedimenta ..................................................................................... 23
3.4. Ţivotinjske vrste i obrada tkiva ............................................................................... 24
3.4.1. Ribe ................................................................................................................ 24
3.4.1.1. Ribe, pregled i uzimanje tkiva za analize .................................................. 25
3.4.1.2. Histološke analize .................................................................................... 28
3.4.1.3. Analize krvi riba....................................................................................... 28
3.4.1.4. Istraţivanje genotoksičnosti ...................................................................... 28

3.4.2. Gujavice .......................................................................................................... 29
3.4.2.1. Gujavice, predtretman: Test kontakta na filtar papiru................................ 29
3.4.2.2. Mjerenje MXR inhibitornog potencijala gujavica nakon izlaganja otpadnim
vodama i otpadnom mulju ......................................................................................... 29
3.4.2.3. Mjerenje oksidacijskog stresa nakon izlaganja gujavica otpadnim vodama i
otpadnom mulju ........................................................................................................ 30
3.5. Ames test ............................................................................................................... 31
3.6. Mikrobiološke analize vode, mulja i tkiva riba........................................................ 32
3.6.1. Maldi TOF MS ................................................................................................ 34
3.6.2. OdreĎivanje osjetljivosti izolata bakterija prema antimikrobnim lijekovima .... 35
3.7. Statistička obrada rezultata ..................................................................................... 35
4. REZULTATI ............................................................................................................... 36
4.1. Analiza uzoraka otpadnih i površinskih voda .......................................................... 36
4.1.1. Rezultati mjerenja temperature vode, suspendirane tvari, pH i
elektroprovodljivosti ..................................................................................................... 36
4.1.2. Rezultati mjerenja otopljenog kisika, kemijske potrošnje kisika (KPKCr i
KPKMn) i biološke potrošnje kisika (BPK-5) ................................................................. 37
4.1.3. Rezultati mjerenja amonijaka, nitrita, nitrata i ukupnog dušika ........................ 38
4.1.4. Rezultati ispitivanja ortofosfata i ukupnog fosfora ........................................... 40
4.1.5. Rezultati mjerenja teških metala ...................................................................... 41
4.1.6. Rezultati mjerenja farmaceutika ...................................................................... 42
4.2. Ţivotinjske vrste i obrada tkiva ............................................................................... 44
4.2.1. Ribe ................................................................................................................ 44
4.2.1.1. Zdravstveni status riba .............................................................................. 44
4.2.1.2. Histologija škrga, bubrega, jetre i slezene ................................................. 44
4.2.1.3. Hematologija, biokemijski parametri u krvi riba ....................................... 48
4.2.1.4. Procjena genotoksičnosti (ENA i MN test) ............................................... 51
4.2.2. Gujavice .......................................................................................................... 52

4.2.2.1. Gujavice: Test kontakta na filtar papiru .................................................... 52
4.2.2.2. Mjerenje lipidne peroksidacije u gujavicama nakon izlaganja otpadnim
vodama i otpadnom mulju ......................................................................................... 54
4.2.2.3. Razina inhibicije MXR proteina uslijed izlaganja otpadnom mulju ........... 56
4.2.2.4. Razina oksidacijskog stresa uslijed izlaganja otpadnom mulju .................. 58
4.3. AMES test .............................................................................................................. 60
4.4. Mikrobiološke analize vode, mulja i tkiva riba........................................................ 64
4.5. Osjetljivost izdvojenih sojeva prema antimikrobnim lijekovima ............................. 76
5. RASPRAVA ............................................................................................................... 83
6. ZAKLJUČAK ............................................................................................................. 94
7. POPIS SIMBOLA I KRATICA ................................................................................... 95
8. LITERATURA ............................................................................................................ 97
9. ŢIVOTOPIS .............................................................................................................. 111
10. POPIS SLIKA I TABLICA ....................................................................................... 113

1
1. UVOD
Voda je opće dobro od osobitog povijesnog, gospodarskog, ekološkog značenja i u sluţbi
zaštite ljudskog zdravlja. Komunalne i industrijske otpadne vode opasne su po okoliš s
obzirom na to da mogu sadrţavati toksične i štetne tvari organskog i anorganskog podrijetla,
koje pak mogu negativno djelovati na vode u koje se ulijevaju, ugroţavajući biotu koja u
njima ţivi narušavajući njihove mikrobiološke, fizikalno-kemijske i biološke osobitosti.
Onečišćivači organskog podrijetla posebice mogu narušiti ravnoteţu ekosustava. U cilju
sprečavanja onečišćenja kopnenih voda koriste se ureĎaji za pročišćavanje otpadnih voda na
kojima se provodi preliminarno, primarno, sekundarno i tercijarno pročišćavanje otpadnih
voda. Preliminarno pročišćavanje predstavlja uklanjanje krupnijih plivajućih tvari, šljunka,
pijeska, ulja i masti, kao i ujednačavanje sastava daljnjeg toka vode. Primarnim
pročišćavanjem uklanjaju se raspršene zrnate plivajuće tvari i suspendirane čestice, te se u
slučaju odstupanja pH vrijednosti provodi neutralizacija. Sekundarno (biološko) pročišćavanje
je uklanjanje otopljenih biorazgradivih organskih tvari gdje mikroorganizmi aerobnim,
anoksičnim i anaerobnim postupcima (biooksidacija, metanska fermentacija, nitrifikacija,
denitrifikacija, biološko uklanjanje fosfora) razgraĎuju sastojke sa ugljikom, dušikom i
fosforom iz otpadne vode (Glancer i sur. 2005). Tercijarno pročišćavanje predstavlja
uklanjanje iz otpadnih voda posebno opasnih tvari kao što su npr. otopljene soli,
mikroorganizmi, pesticidi, deterdţenti, toksične i radioaktivne tvari. Upotreba aktivnog mulja
najčešći je način pročišćavanja otpadnih voda, koji se pokazao vrlo učinkovit stvarajući
relativno kvalitetan efluent (Naidoo i Olaniran 2014). Ispuštanjem pročišćenih otpadnih voda
u prirodne vodotoke mijenjaju se prirodne vrijednosti parametara sastava vode, kako u
kvantitativnom, tako i u kvalitativnom smislu. Ispuštanje pročišćenih otpadnih voda još uvijek
opterećenih organskim tvarima, nutrijentima i različitim zagaĎivalima dovodi do
eutrofikacije, a procesi samopročišćavanja ostatka organskog opterećenja troše kisik otopljen
u vodi i dovode do njegova manjka. TakoĎer dolazi i do akumulacije onečišćivača u
površinskim vodama koje sluţe kao recipijent (Bhatia i Goyal 2013). UreĎaji za biološko
pročišćavanje otpadnih voda projektirani su za smanjenje biokemijske potrošnje kisika,
ukupne suspendirane tvari, dušika i fosfora iz otpadnih voda, dok se uklanjanju patogenih
mikroorganizama poklanjalo manje paţnje (Lucas i sur. 2014.) Iako se primarnim i
sekundarnim pročišćavanjem iz otpadnih voda ukloni i do 99 % bakterija fekalnog porijekla
(Lucas i sur. 2014; Servais i sur. 2007), to još uvijek nije dovoljno smanjenje da bi se

2
površinske vode koje su recipijent s ureĎaja za pročišćavanje otpadnih voda mogle koristiti za
kupanje i rekreaciju. Promjene u vodenom okolišu mogu povećati ranjivost organizama koji
ţive u vodi na toksične kemikalije zbog promjene kapaciteta organizma za prepoznavanje i
popravak oštećenja izazvanih toksikantima ili promjenom u ponašanju ţivotinja kao što su
migracije ili predatorstvo (Couillard i sur. 2008). MeĎutim, vrlo je vaţno naglasiti da se sastav
otpadnih (komunalnih i industrijskih) voda tijekom proteklih desetljeća promijenio. Povećala
se količina endokrinih modulatora, genotoksikanata, antimikrobnih lijekova, antikoroziva i
organofosfata u otpadnim vodama, vodotocima, pa i u vodi za piće. Njihov nalaz u
vodotocima zavisi od postupaka obrade otpadnih voda jer ih svi pročistači otpadnih voda ne
uklanjaju u cijelosti, tako da njihovo otpuštanje u vodotoke moţe čak biti kontinuirano
(Esteban i sur. 2014). Tvari strane organizmu (ksenobiotici) mogu utjecati na sposobnost
akvatičnih vrsta za prilagodbu na promjene uzrokovane različitim stresorima ili patogenim
bakterijama u okolišu. Ribe su vrlo osjetljive na promjene okolišnih uvjeta tako da njihovo
fiziološko stanje moţe posluţiti kao jedan od najranijih indikatora specifičnog zdravstvenog
statusa ekosustava (Kaur i Dua 2014).
U Virovitici je 1989. g. izgraĎen ureĎaj za biološko pročišćavanje (UBP) otpadnih voda grada
i industrije te Tvornice šećera Virovitica (TŠ). Pročistač je sekundarnog tipa pročišćavanja i
pomoću mehaničkih i bioloških metoda pročišćava komunalne i tehnološke otpadne vode.
Gradske otpadne vode mješovite su vode iz kućanstava i industrije te oborinske vode.
Industrijske otpadne vode prije ispuštanja u sustav gradske kanalizacije, zavisno od prirode
procesa, prolaze različite predtretmane. Odvodni sustav otpadnih voda završava na
mehaničkom i biološkom ureĎaju za pročišćavanje otpadnih voda koji je tehnološki
koncipiran tako da se na njemu zajedno pročišćavaju otpadne vode grada Virovitice i
tehnološke otpadne vode TŠ-a na anaerobno-aerobnom principu, tehnologijom aktivnog
mulja. Na tom ureĎaju otpadne vode nakon primarnog čišćenja (rešetka, pjeskolov, aerobni
mastolov) ulaze u bioaeracijske bazene i nakon toga se u sekundarnim taloţnicama obavlja
odvajanje aktivnog mulja i bistre faze. Konačni recipijent nakon pročišćavanja otpadnih voda
je kanal Manteč, Ţupanijski kanal i dalje rijeka Drava.
Radi dobivanja reprezentativnih uzoraka vode, sedimenta, mulja i riba razvijena je strategija
prikladna za UBP na kojem se čiste komunalne, šećeranske i bolničke otpadne vode. Prednost
zajedničkog pročišćavanja ovih vrsta otpadnih voda je razrjeĎenje organski visoko
opterećenih šećeranskih otpadnih voda komunalnim otpadnim vodama radi čega ne dolazi do
izlijevanja nepročišćenih otpadnih voda zbog gubitaka kroz propusni odvodni sustav ili
prelijevanjem u okoliš. Za vrijeme provoĎenja ovog istraţivanja odvijala se proizvodnja

3
šećera, u proljeće iz sirovog šećera dobivenog od šećerne trske, a u jesen iz šećerne repe.
Šećeranske otpadne vode, u odnosu na druge industrije koje ispuštaju otpadne vode u sustav
za njihovo sakupljanje i odvodnju, daju značajan doprinos hidrauličkom i netoksičnom
organskom opterećenju ureĎaja (Ingaramo i sur. 2009).
1.1. Cilj i znaĉaj istraţivanja
Odnos izmeĎu specifičnog doprinosa efluenta s ureĎaja za pročišćavanje otpadnih voda i
promjena u organizmima koji obitavaju u efluentu ili su mu izloţeni u prijemnicima
pročišćenih otpadnih voda ili mulju, uvelike je bio neistraţen. Isto tako, nedovoljno je
evaluirana učinkovitost uklanjanja iz otpadnih voda bakterija i neorganskih onečišćivača
(posebice antimikrobnih lijekova). Zbog toga je cilj ovog rada ispitati hipotezu da 1) efluent s
UBP-a uzrokuje biološke učinke na indikatorskim organizmima; 2) u pročišćenoj vodi i mulju
postoje bakterije rezistentne na antimikrobne lijekove koje mogu biti patogene za ribe i ljude,
a u sedimentu brojni antimikrobni lijekovi. Kako bi se ove hipoteze potvrdile, proveden je niz
testova kojima se dokazivao utjecaj na zdravstveni status riba koje obitavaju nizvodno od
ulijevanja prijemnika pročišćenih otpadnih voda. OdreĎivana je njihova mikrobiološka
kontaminacija, struktura tkiva, biokemijski profil krvi, oksidacijski stres, genotoksični učinci,
te mehanizam multiksenobiotičke otpornosti na gujavicama (MXR; engl. multixenobiotic
resistance), a za odreĎivanje mutagenosti tvari iz vode i mulja koristio se Ames test: (i)
provjeren je zdravstveni status riba, provedena je histopatološka obrada tkiva (za hipotezu 1);
(ii) fenotipskim testovima i masenom spektrometrijom - matricom potpomognutom laserskom
desorpcijom/ionizacijom i analizatorom vremena leta (MALDI-TOF MS, engl. Matrix-
assisted laser induced desorption ionization-time of flight-mass spectrometry) istraţen je veći
broj bakterijskih izolata uzoraka voda, riba i mulja (za hipoteze 1, 2); (iii) u krvi riba
istraţivani su biokemijski parametri i parametri oksidacijskog stresa (za hipotezu 1); (iv)
odreĎivana je učestalost nepravilnosti graĎe stanične jezgre; (v) nakon izlaganja testnih
organizama (gujavica) nepročišćenoj i pročišćenoj otpadnoj vodi i mulju odreĎivan je stupanj
staničnog detoksikacijskog mehanizma posredovanog aktivnošću ABC transportnih proteina
koji na sebe veţu i iz stanice izbacuju ksenobiotike, uključujući antropogeno stvorene
spojeve, sprečavajući na taj način potencijalno toksičan učinak u stanici (za hipotezu 1); (vi)
izvršeno je odreĎivanje koncentracije antimikrobnih lijekova u uzorcima voda i sedimenta (za
hipotezu 2).

4
Naglasak je stavljen na istraţivanje utjecaja pročišćenih otpadnih voda na ribe i gujavice kao
testne organizme, naročito u pogledu utjecaja svih korisnika javnog odvodnog sustava
(otpadne vode iz kućanstava, šećeranske i bolničke otpadne vode) što je jedinstven slučaj u
proučavanju učinkovitosti rada ureĎaja za pročišćavanje otpadnih voda.
U ovom se radu, dakle, istraţivao odnos čimbenika koji utječu na fizikalno-kemijske,
mikrobiološke, genotoksične i mutagene značajke voda koje ulaze i izlaze iz UBP-a, odnosno
djelovanje pročistača na smanjenje toksičnog utjecaja otpadnih voda. OdreĎivan je sastav
populacija riba koje obitavaju u neposrednim recipijentima (kanal Manteč i Ţupanijski kanal
prije utoka u Dravu), njihovo zdravstveno stanje, identificirane su mikrobiološke prijetnje
njihovom zdravlju, odreĎeni su hematološki pokazatelji citotoksičnosti i biokemijski profil
krvi. Na uzorcima vode i u aktivnom mulju obavljena je mikrobiološka pretraga, kao i
odreĎivanje mutagenog potencijala i prisutnost MXR inhibitora. Na taj način dobivena je
cjelovita slika utjecaja pročišćenih otpadnih voda grada Virovitice na ključne parametre
kakvoće i ekološkog stanja, a poglavito na mikrobiološke i toksikološke značajke vode i biote.
Takvo sveobuhvatno istraţivanje, uzimajući u obzir granične vrijednosti emisija
onečišćujućih tvari u otpadnim vodama (NN 80/13, NN 43/14, NN 27/15, NN 3/16),
poglavito fizikalno-kemijske, ekotoksikološke, organske i anorganske pokazatelje, do sada
nije provoĎeno.
Značaj ovog istraţivanja ogleda se u činjenici da je svaki ţivi organizam podloţan fiziološkim
i patološkim promjenama koje je moguće sagledati kroz kvalitativne i kvantitativne
pokazatelje na različitim razinama biološke organizacije, a koje su inducirane abiotičkim
(kakvoća vode, antibiotici i kemoterapeutici, tragovi metala) i biotičkim (vrsta, starost, spol,
mikroorganizmi) čimbenicima. UtvrĎivanje fiziološkog statusa te biokemijskih/kemijskih
profila tkiva (krvi) riba i njihovo korištenje kao bioloških senzora od iznimne je vaţnosti jer
oni mogu sluţiti kao dijagnostičke metode za procjenu zdravstvenog statusa i stupnja
ekološke ugroţenosti vodenog sustava (Affonso i sur. 2002; Čoţ-Rakovac i sur. 2008; Ferri i
sur. 2011; Topić Popović i sur. 2008). Prethodne studije i citološka istraţivanja pokazali su da
okoliš bitno utječe na morfologiju stanice i njezine regulatorne mehanizme. Dokazana je
korelacija izmeĎu pojave mikronukleusa i eritrocitnih nuklearnih abnormalnosti, te utjecaj
ekoloških faktora (temperatura, kisik, salinitet) na pojavu morfoloških promjena jezgri
(nepravilan oblik, jezgre s vakuolama, binuklearni eritrociti) riba (Polard i sur. 2011;
Strunjak-Perović i sur. 2009). Korištenjem novih kliničko-biokemijskih dijagnostičkih metoda
(raspon referentnih vrijednosti biokemijskih i kemijskih parametara tkiva), te modifikacijom

5
postojećih mikrobioloških dijagnostičkih metoda (brzi biokemijski dijagnostički i fenotipski
testovi) uz usporedbu s MALDI-TOF MS rezultatima mikrobiološke identifikacije skraćuje
se vrijeme do donošenja konačne dijagnoze i procjene onečišćenosti okoliša (Biswas i Rolain
2013; Braga i sur. 2012; El-Bouri i sur. 2012). OdreĎivanje mikrobioloških parametara
opisano je u brojnim istraţivanjima kao presudno za kakvoću vode (Cabral 2010; Garrido i
sur. 2014; Varela i Manaia 2013) i zdravlje riba (Novotny i sur. 2004; Stoskopf 1993), a oni
izravno ili neizravno utječu na zdravlje korisnika tih voda.

6
2. LITERATURNI PREGLED
2.1. UreĊaji za biološko proĉišćavanje otpadnih voda
Bez obzira na to što arheološki pronalasci upućuju da su sustavi sakupljanja i odvodnje
otpadnih voda graĎeni već 4 000 godina pr.n.e., prvi ureĎaji za pročišćavanje otpadnih voda
su se pojavili tek sredinom XIX stoljeća. Veći dio XIX stoljeća sanitarne otpadne vode, kao i
sve veće količine industrijskih otpadnih voda, ispuštale su se izravno u mora, u rijeke
nizvodno od naselja, jezera ili u tlo, često u neposrednoj blizini zdenaca iz kojih se zahvaćala
voda za piće. Za vrijeme epidemije kolere u Londonu sir John Snow prvi je ukazao na vezu
izmeĎu otpada ljudskog podrijetla i oboljenja vezanih uz vodu. Tema rasprava Kraljevskog
instituta Velike Britanije (Royal Institution of Great Britain) 1868. bila je onečišćenje vode za
piće sa slivnog područja planina Cader Idris i Plyalimmon (sjeverni Wales) nezdravim
„klicama“ i kemikalijama. U to vrijeme Fankland u Velikoj Britaniji i Finkener i Zinreck u
Njemačkoj započinju s uzorkovanjem voda iz zagaĎenih rijeka i njihovim analizama i
utvrĎuje se proces samopročišćavanja nizvodno od mjesta zagaĎenja (Wiesmann i sur. 2007).
ProvoĎenjem pokusa aeracijom otpadne vode utvrĎeno je da do smanjenja koncentracije
organske tvari dolazi tek nakon nekoliko tjedana. Smatra se da je Alexander Müller bio prvi
koji je 1869. godine naslutio da smanjenju organske tvari u otpadnoj vodi doprinose
mikrobiološki procesi (Wiesmann i sur. 2007). Veliki doprinos u razumijevanju procesa
pročišćavanja otpadnih voda dao je Winogradsky (1890) koji je ukazao na procese
denitrifikacije. On je ustvrdio da, za razliku od bakterija koje kao izvor energije koriste ugljik
iz organske tvari, postoje kemolitičko-autotrofne nitrificirajuće bakterije koje energiju
dobivaju oksidacijom i
a ugljik dobivaju redukcijom (Wiesmann i sur. 2007).
Razvoj industrije povećavao je onečišćenje rijeka i nametao potrebu ţurnog rješavanja ovog
problema. U Berlinu je s tim ciljem Hobrecht izgradio jedinstveni sustav za odvodnju
otpadnih i sakupljanje oborinskih voda, koje su se gravitacijski slijevale u središnju najniţu
točku, iz koje se ova voda koristila za navodnjavanje poljoprivrednih površina izvan grada.
Procjedna voda se sakupljala i odvodila keramičkim drenaţnim cijevima u kanale i rijeke.
Analizama je utvrĎeno da ova voda sadrţi znatno manje suspendirane i organske tvari, manju
vrijednost potrošnje i amonijaka, uz manju redukciju otopljenih i anorganskih tvari.
Dr. Angus Smith je objavio da se aeracijom otpadne vode moţe stabilizirati organska tvar a
1913. god. na Lawrence Experiment Station (SAD) utvrĎeno je da mikroorganizmi povezani s

7
uronjenim pločicama škriljevca poboljšavaju stabilizaciju organske tvari (Wiesmann i sur.,
2007). Na bazi ovog pronalaska britanski inţenjeri Edward Arden i William T. Lockett su
došli do zaključka da se višekratnom recirkulacijom krute tvari koja se akumulira u bačvama
za vrijeme aerobne stabilizacije, u trajanju od nekoliko sati, povećava stupanj stabilizacije
organske tvari i prvi put uveli pojam „aktivni mulj“. U narednim godinama ovaj proces
primijenjen je na model s kontinuiranim protokom uz korištenje aeracijskog bazena, bazena
za taloţenje aktivnog mulja i sistema za njegovu reciklaciju. Nakon više pokusa na pilot-
ureĎajima u Velikoj Britaniji i SAD-u, prvi ureĎaj za pročišćavanje otpadnih voda
tehnologijom aktivnog mulja izgraĎen 1920. godine u gradu Sheffeld u Velikoj Britaniji koji
je započinjao dugim meandriranim kanalom zbog aeracije. Na ureĎaju u Indianapolisu (SAD)
aeracija se vršila utiskivanjem zraka u cijevi perforirane pri dnu, a prvi površinski aerator s
vertikalnom osovinom pomoću kojeg se dopremao u otpadnu vodu kisik iz zraka i vršilo
miješanje aktivnog mulja izgradio je Bolton 1921. godine. U Njemačkoj su prvi ureĎaji za
obradu otpadnih voda izgraĎeni 1926. u Essen-Rellinghausenu i u Stahnsdorfu pokraj Berlina
izmeĎu 1929 i 1931. godine (Wiesmann i sur. 2007).
2.2. Fizikalno-kemijske analize
Provedba zaštite voda u Hrvatskoj regulirana je (Zakonom o vodama, NN 153/09, 130/11,
56/13, 14/14), Strategijom upravljanja vodama (NN 91/08), Drţavnim planom za zaštitu voda
(NN 8/99), Pravilnikom o graničnim vrijednostima emisije otpadnih tvari (NN 80/13, NN
43/14, NN 27/15, NN 3/16) i Uredbom o standardu kakvoće voda (NN73/13). Dosadašnja
istraţivanja kakvoće vode Ţupanijskog kanala provoĎena su u razdoblju od 1983. do 2003.
godine sa ciljem uporabe vode za navodnjavanje. Vrijednosti za KPKMn u tom razdoblju
kretale su se od 3,48 do 9,00 mgO2/L, BPK-5 od 2,16 do 24,17 mgO2/L a otopljenog kisika od
1,63 do 9,2 mg/L . Koncentracija amonijaka kretale su se u rasponu od 0,10 do >1,50 mgN/L
a nitrita od <0,01 do 0,20 mgN/L. Broj koliformnih bakterija bio je u rasponu od <5x102 do
105/100 mL a broj aerobnih bakterija na 22 °C od od <10
3 do 10
5/mL (Šimunić i sur. 2006).
2.3. Biološke analize utjecaja oneĉišćenja na okoliš
Sve do 1980. godine osnovni cilj pročišćavanja otpadnih voda bio je udovoljiti estetskim i
okolišnim kriterijima, nakon čega se, zbog rastućeg onečišćenja otpadnih voda otrovnim

8
tvarima, sve veća pozornost daje brizi za ljudsko zdravlje i bioti vodenih sustava kao
prijemnika pročišćenih otpadnih voda. UreĎaji za pročišćavanje otpadnih voda projektirani su
na način da iz njih uklanjaju suspendirane tvari, organsku tvar i nutrijente, a nacionalnim
propisima utvrĎeni su kriteriji za ispuštenu otpadnu vodu u toj razini čišćenja. Nacionalni
propisi za ispuštanje organskih mikrozagaĎivala i polarnih spojeva u vode ne postoje, a
najčešće se mjerenja ovih spojeva u otpadnim vodama niti ne provode. Uz mjerenje
anorganskih (metali, slabi metali-metaloidi, nemetali) i organskih toksikanata (ugljikovodici,
pesticidi, detergencije, klorofenoli, TBT) u središte istraţivanja dolaze nova onečišćivala (npr.
antibiotici, antiepileptici, analgetici i protuupalni spojevi, regulatori lipida, beta-blokatori,
diuretici, kontrastna sredstva, mirisi, psihostimulansi, dezinficijensi, antidepresivi,
plastifikatori (Ašperger i sur. 2013). Značaj sve više dobivaju toksikološka i ekotoksikološka
istraţivanja. Dok toksikologija proučava štetne učinke kemijskih tvari na ţive organizme,
pozornost pridaje ciljanim organima i promjenama u njima uzorkovanim kemikalijama,
proučava pretvorbu toksikanata u organizmu, tvorbu metabolita i zaštitne mehanizme,
ekotoksikologija se bavi istraţivanjem toksičnih učinaka onečišćivača prirodnog i
antropogenog porijekla na sastavnice ekosustava, odnosno ekosustav u cjelini (Truhaut 1977).
U najnovije vrijeme umjesto pojma ekotoksikologija, sve se češće navodi pojam ekologija
stresa zbog neprijeporne činjenice da su, uz izravno djelovanje kemikalija na organizme u
okolišu, prisutni i drugi, abiotički i biotički aspekti negativnog utjecaja na okoliš (van Stralen
2003). Utjecaj na ţivi svijet velikog broja kemikalija koje dospijevaju u okoliš, zbog njihove
meĎusobne interakcije uvjetovane nizom fizikalno-kemijskih čimbenika, ne moţe se utvrditi
mjerenjem samo kemijskih pokazatelja, odnosno u turbulentnim medijima kao što je voda to
bi moglo navesti na pogrešne zaključke. Interakcije meĎu kemikalijama mogu biti sinergijske,
antagonističke, potencirajuće ili aditivne (Stepić 2010). Zbog toga se rezultati toksikoloških
ispitivanja kojima se procjenjuje utjecaj kemikalije na testni organizam u laboratorijskim
uvjetima ne mogu izjednačiti s utjecajem na ţiva bića u uvjetima okoliša. Biološka
istraţivanja uključuju sve analize koje ukazuju na interakciju izmeĎu biološkog sustava i
potencijalno štetnog kemijskog, fizikalnog ili biološkog djelovanja (WHO 1993). Često se
negativni utjecaj zagaĎivala na ţivi svijet uočava tek nakon incidentnog zagaĎenja, ili ako je
onečišćenje bilo prisutno kroz duţe vrijeme. U takvim slučajevima potreban je dugi period,
nekad i deseci godina, da se ekosustav oporavi, a nekad ugibanjem ţivih bića nestaje dio
genetičke informacije, smanjuje se biološka raznolikost i mogućnost oporavka ekosustava.
Radi sprečavanja štetnog utjecaja zagaĎenja na okoliš razvile su se biološke metode i
uspostavio biološki nadzor (biomonitorig) kojim se utvrĎuje utjecaj zagaĎivala na različitim

9
razinama biološke organizacije. Ekološki pokazatelji promjena mogu se utvrditi na razini
ekosustava, zajednice ili populacije korištenjem ekoloških indikatora kao što su analize
promjena u populacijama, brojnost jedinki, mortalitet, natalitet, starosna struktura, omjer
spolova, gustoća i raspored jedinki u prostoru. Prate se i promjene u sastavu zajednica
kvantitativnim i kvalitativnim istraţivanjem. Kvantitativnim ispitivanjem se odreĎuju vrste
ţivih bića koje nastanjuju sustav uvaţavajući činjenicu da svaka vrsta u drugačijoj mjeri
podnosi promjene uzrokovane onečišćenjem. Razvijen je saprobni sustav s osnovnim
kategorijama kopnenih voda (Kolkwitz i Marsson 1902) gdje je kriterij bio koncentracija
organske tvari, količina otopljenog kisika i raznolikost flore i faune (oligosaprobna, β-
mezosaprobna, α-mezosaprobna i polisaprobna voda s najmanje kisika zbog visoke
koncentracije organske tvari i najmanjom raznolikosti flore i faune). Biološko odreĎivanje
kvalitete vode zasniva se na indikatorskim vrstama (bioindikatorima) čiji pronalazak ili
izostanak upućuje na organsko onečišćenje sustava. Prvi popisi obuhvaćali su oko 800 biljnih
i ţivotinjskih indikatorskih vrsta (Kolkwitz i Marsson 1908; Kolkwitz i Marsson 1909).
Kvantitativna istraţivanja zajednica podrazumijevaju odreĎivanje vrsta i procjenu njihove
brojnosti, a za prikaz ovih istraţivanja uveden je saprobni indeks (Liebman 1962; Pantle i
Buck 1955; Sladeček 1973). Kasnije se za prikaz kvantitativnih istraţivanja koristi indeks
raznolikosti (npr. Shannon-Wienerov ili Shannonov indeks) za čiji izračun je potrebno
ustanoviti broj pronaĎenih vrsta i ukupan broj naĎenih jedinki svake vrste. Primjenom metoda
biotičkog indeksa koriste se indikatorske vrste, najčešće makrozoobentos, za koje se utvrĎuje
prisutnost ili odsutnost, a svakoj se biotičkoj skupini pridaju bodovi koji su u korelaciji s
njihovom osjetljivošću prema onečišćenju. Ţivotinjama koje su više osjetljive na onečišćenje i
pronalaze se u nezagaĎenim vodama dodaje se veća brojčana vrijednost i obrnuto.
Istraţivanjima utjecaja zagaĎenja na razini zajednice i populacije kao ekološkim indikatorima
najčešće se otkrivaju već poodmakle promjene. UtvrĎivanjem promjena na razini organizma,
organa, tkiva, stanice i molekule, tj. biomarkerima najranije se mogu dobiti informacije o
mogućem pogoršanju zdravlja promatrane jedinke, populacije i biocenoze u cijelini (Koeman
i sur. 1993). Pokazatelji promjene na organizmima kao bioindikatorima su morfološke i
bioenergetske prirode i time izdvojeni iz skupine biomarkera (Van Gastel i Van Brummelen
1994). Biomarkeri predstavljaju histopatološke, imunološke, fiziološke, detoksikacijsko-
enzimatske, biokemijske i molekularno-genetičke promjene na tkivu, organu, stanici ili
molekuli.

10
2.4. Mikrobiološka istraţivanja
Voda, posebice ako je opterećena organskim tvarima, okoliš je u kojemu uspješno opstaju
pripadnici mnogih bakterijskih rodova. Mikrobiološki sastav otpadnih voda odreĎuju bakterije
podrijetlom iz probavnog sustava kraljeţnjaka. Najčešće se radi o bakterijama Bacteroides
spp, Bifidobacterium spp., Chlostridium perfingens, Enterobacter aerogenes, Escherichia
coli, Lactobacillus spp. i Enteroococcus fecalis. Budući da je otpadna voda jednim dijelom
porijeklom iz domaćinstava, ljudski patogeni mogu se naći i u muljevima pročistača otpadnih
voda. Najčešće bakterije ove skupine su Campilobacter spp., Leptospira spp., Salmonella
paratyphi, Salmonella typhi, Salmonella typhimurium, Shigella dysenteriae i Vibrio cholerae.
Osim navednih, u otpadnim vodama nalaze se i bakterije rodova Cytophaga, Micrococcus,
Pseudomonas, Bdellovibrio, Chromobacterium, Aeromonas, Rhodispirillium i druge vrste. Od
protozoa načešće se u otpadnim vodama mogu naći Entamoeba histolytica i Giardia
intestinalis a od virusa adenovirusi, enterovirusi (koksaki virusi, echovirusi i poliovirusi),
parvovirusi (hepatitis A virus) i reovirusi (Pejić 2014.).
Dokazano je da je normalna bakterijska flora riba odraz bakterijske flore vode u kojoj plivaju.
Ipak, postoje neke bakterijske vrste koje su obligatne ribljim domaćinima i bez njih ne mogu
opstati (Frerichs i Roberts 1989). S obzirom na to da se ribe trajno nalaze u vodenoj suspenziji
mikroorganizama, njihova je vanjska površina sa njima u stalnom doticaju (Topić Popović
1997). No, prema Austin i Austin-u (1987), tek naseljavanje tih bakterija u ribi je od posebnog
značaja, a ono moţe biti sljedeće:
a- bakterije se mogu naseliti na vanjsku površinu ribe i tu postati dijelom rezidentne mikroflore
koja inhibira kolonizaciju drugim mikroorganizmima
b- bakterije mogu biti privučene ili slučajno dospjeti na mjesto ozljede (oštećenje koţe,
nedostatak ljusaka) i tu se razmnoţiti
c- bakterije se mogu zaustaviti na škrgama i tu postati dijelom rezidentne mikroflore
d- bakterije mogu u ribu ući kroz usta, vodom ili česticama hrane, i dospjeti u probavni sustav.
Pojedini sojevi postanu dijelom normalne mikroflore, neki budu uništeni ili postaju patogeni, a
drugi prolaze kroz probavni sustav i izluče se izmetom.
Općenito uzevši, broj aerobnih heterotrofnih bakterija koje se nalaze na površini ribe odgovara
njihovom broju u okolnoj vodi. Predstavnici bakterijske populacije na površini slatkovodnih riba
najčešće su vrste Acinetobacter spp., Aeromonas hydrophila, Alcaligenes piechaudii,
Enterobacter aerogenes, E. coli, Flavobacterium spp., Micrococcus luteus, Moraxella spp.,

11
Pseudomonas fluorescens i Vibrio fluvialis (Allen i sur. 1983; Christensen 1977; Southgate
1993). Velika populacija mikroorganizama nalazi se u crijevima riba, gdje je broj bakterija
daleko veći nego u okolnoj vodi, ukazujući na to da u probavnom sustavu vladaju povoljni uvjeti
za mikroorganizme. Nedvojbeno je da veliki broj tih mikroorganizama potječe iz hrane, u kojoj
se većinom nalaze gram-pozitivni mikroorganizmi (Ringo i Strom, 1994). Bakterije koje se
nalaze u unutrašnjim organima riba, gdje neke mogu uzrokovati bolesti, su Lactobacillus spp.,
Mycobacterium spp., Nocardia spp., Renibacterium salmoninarum, vrste iz roda Aeromonas,
vrste iz porodice Enterobacteriaceae, poput Edwardsiella ictaluri, Yersinia ruckeri, zatim
Flavobacterium spp., Pseudomonas anguilliseptica, Pseudomonas fluorescens, Vibrio spp.,
Vibrio anguilarum, Acinetobacter spp., Photobacterium damselae i druge vrste. Stresori
(mikrobiološko, kemijsko, organsko onečišćenje) mogu narušiti zdravlje riba. Osim toga,
izlaganje stresorima u okolišu moţe smanjiti postotak preţivljavanja riba, smanjiti njihov
reproduktivni uspjeh sa mogućim ozbiljnim posljedicama na preţivljavanje cijele populacije riba
u vodotoku (Topić Popović 1997).
2.5. Histološke pretrage
S obzirom na to da onečišćenje okoliša moţe dovesti do odstupanja od normalne strukture tj.
manjih ili većih histopatoloških promjena, histološke metode se sve češće koriste kako bi se
bolje odredio zdravstveni status riba u biomonitoringu njihove ţivotne sredine. Prednost
histološkom pregledu organa riba leţi i u činjenici da se ribe u vodenim ekosustavima nalaze
na vrhu trofičke piramide, a zbog učinaka biokoncentracije i bioakumulacije dobri su
indikatori zagaĎenja vodenih ekosustava (Dallinger i sur. 1987; Polak-Juszczak 2012). Zbog
izravnog doticaja s vodom, u riba su djelovanju toksikanata najizloţenije škrge, a štetni se
učinci pojavljuju u vidu hiperplazije i hipertrofije epitela, te odizanja epitela kao regresivne
promjene. U cilju utvrĎivanja utjecaja zagaĎivala provode se histološke pretrage organa koji
su pridruţeni probavnom sustavu (npr. jetre), slezene i bubrega kao krvotvornih i ekskretornih
organa, a često i reproduktivnih organa.

12
2.6. Biokemijske i hematološke pretrage krvi
Analize krvi u humanoj medicini, kao i kod viših kraljeţnjaka (sisavci, ptice) vrlo su vaţne
metode utvrĎivanja fizioloških i patoloških stanja organizama, a u proučavanju zdravstvenog
statusa riba postaju sve značajniji čimbenik. Proteini krvne plazme su globularni proteini, a
plazma sadrţi više desetaka različitih proteina od kojih najveći dio dolazi u tragovima.
Proteini su i sastavni dio svake stanice. Stvaraju kontraktilne elemente i enzime koji
oslobaĎaju energiju potrebnu za odrţavanje ţivota. Nalaze se u krvi gdje imaju funkciju
transporta lijekova, hormona, vitamina i elektrolita koji se veţu na proteine. Pojedini proteini
imaju specifične funkcije, uključujući i zaštitnu funkciju (enzimi, hormoni, inhibitori enzima,
faktori koagulacije, imunoglobulini) (Čolak, 2011). Utječu na koloidno-osmotski tlak i time
na raspodjelu vode izmeĎu vaskularnog i meĎustaničnog prostora (Karlson 1988; Štraus
1988). Lipidi su, osim kao izvor energije neophodni za rast, razmnoţavanje, sastavni dio
stanične membrane, i sastavni su dio steroidnih hormona (Cooper i Hausman 2010). Lipidi
krvne plazme prelaze u tkiva i iz tkiva. Predstavljaju najvaţniju grupu masnih tvari sa gledišta
energetskih potreba organizma.
Glukoza je jednostavni šećer monosaharid, koji se nalazi u hrani, krvi i vaţan je izvor energije
za organizme. Ona je završni produkt probave ugljikohidrata. Oksidacijom glukoze proizvodi
se energija za tjelesne stanice. Ureja je diamid karbonske kiseline koji je glavni metabolički
produkt dušikovih tvari. U metabolizmu se razgraĎuje odreĎena količina proteina, a dušik se
izlučuje kao mokraćevina putem urina. Metabolizmom aminokiselina nastaje u amonijak,
NH3, odnosno NH4+. Dnevna količina izlučenog kreatinina nalazi se pod neznatnim utjecajem
mišićnog rada ili volumena izlučenog urina. Njihov laboratorijski nalaz moţe biti
upotrijebljen kao mjera funkcije ekskretornih organa. Kreatinin se normalno proizvodi u
ujednačenim količinama kao rezultat cijepanja fosfokreatina i izlučuje se u urinu. Enzimi,
poput alkalne fosfataze, kreatin kinaze, transaminaze, gama glutamil transpeptidaze, laktat
dehidrogenaze, nalaze se u staničnim membranama i jezgrama i kataliziraju hidrolizu
fosfatnih estera, prijenos fosfatne grupe iz fosfokreatina na ATP, proces provoĎenja amino
grupe iz jedne molekule na drugu, odnosno proces provoĎenja aminokiseline u keto kiselinu
transaminacijom, interkonverziju laktata i piruvata, a nalaze se u gotovo svim tkivima riba.
(Čolak, 2011).

13
2.7. Ekotoksikološki testovi na modelnim organizmima
Kao modelni organizam u ekotoksikološkim testovima često se koriste gujavice. Gujavice
pripadaju razredu Oligochaeta su dvospolci i jedna su od najvaţnijih i najbrojnijih terestričkih
porodica. Karakterizira ih bilateralna simetrija te jasna kolutičavost. Tijelo je zaštićeno
kutikulom ispod koje se nalazi jednoslojni epiderm te sloj prstenastih i uzduţnih mišića koji
im omogućuju kretanje. Tjelesna šupljina (celom) ispunjena je celomskom tekućinom koja
sluţi kao hidrostatski skelet te direktno utječe na pokretanje te ispruţanje tijela. Opt jecajni
sustav je zatvoren te se disanje odvija isključivo preko koţe. Apsorpcija se odvija preko
probavnog sustava, ali i preko koţe.
Njihova uloga u terestričkim zajednicama je višestruka – miješajući slojeve tla doprinose
poboljšavanju kvalitete tla te prozračivanju, povećavajući propusnost tla utječu na hidraulička
svojstva te sudjeluju u mineralizaciji organske tvari čime direktno utječu na kemiju tla
(Edwards i Bohlen, 1996).
Gujavica Eisenia fetida (Savigny, 1826) epigejna je vrsta koja s obzirom na mogućnost
apsorpcije preko kutikule i probavne površine moţe biti izloţena širokom spektru
potencijalno toksičnih onečišćivača. Karakterizira je kratak ţivotni ciklus, brzi reprodukcijski
potencijal, laka dostupnost, jednostavnost za uzgoj te odrţavanje. Upravo iz tih razloga,
prema OECD protokolu (1984) preporučena je kao izvrstan modelni organizam za
ekotoksikološka ispitivanja tla.
2.8. Mehanizam multiksenobiotiĉke otpornosti
Mehanizam multiksenobiotičke otpornosti (MXR; engl. multixenobiotic resistance) kao
pojam u znanosti uveo je akademik Branko Kurelec (1992) te ga je okarakterizirao kao
mogućnost obrane organizma od okolišnih potencijalno štetnih spojeva. MXR omogućuje
zaštitu stanice (organizma) od toksičnih tvari. MXR kao obrambeni mehanizam zasniva se na
aktivnosti ABC transportnih proteina čija je prisutnost utvrĎena u gotovo svim organizmima
od bakterija pa sve do kraljeţnjaka (Kurelec 1992; Litman i sur. 2001; Sauerborn Klobučar i
sur. 2010). ABC transportni proteini (engl. ATP-binding cassette) najveća su porodica dosad
otkrivenih transmembranskih proteinskih porodica. Glavni predstavnici ABC transportera

14
uključeni u MXR mehanizam su: P-glikoprotein (P-gp), MRP (engl. multidrug resistance-
associated protein), BCRP (engl. breast cancer resistance protein) (Bard i sur. 2000; Litman i
sur. 2001). Neki okolišni spojevi, bilo prirodnog ili antropogenog podrijetla, mogu inhibirati
ABC proteine tj. MXR mehanizam. Takve spojeve nazivamo MXR inhibitorima. Oni mogu
djelovati pojedinačno kao jedan spoj ili u sinergiji, kada blokirajući transportnu aktivnost
ABC proteina onemogućuju izbacivanje drugog potencijalno štetnog spoja iz stanice (Epel i
sur. 2008). U konačnici to moţe dovesti do akumulacije štetnih spojeva unutar same stanice,
što predstavlja potencijalnu opasnost za organizam. Upravo zato, mjerenje aktivnosti MXR
mehanizma moţe posluţiti kao izvrstan biomarker za biomonitoring zagaĎenja okoliša MXR
inhibitorima.
Princip samog testa zasniva se na mjerenju fluorescentnog supstrata (u našem slučaju
Rodamin123) akumuliranog u tkivu. Mjerenjem fluorescentnih vrijednosti u cijelom
organizmu dobivamo jasan uvid u stupanj inhibicije MXR mehanizma (Epel i sur. 2008). U
slučaju inhibicije ABC transportera, dolazi do zaostajanja fluorescentnog supstrata u stanici i
njegove akumulacije u organizmu. Ukoliko testiran spoj ili smjesa spojeva nema MXR
inhibitorsko svojstvo, transportni proteini će učinkovito izbacivati fluorescentni supstrat izvan
stanice (organizma), te na taj način sprječavati potencijalno štetan učinak na organizam.
2.9. Mikronukleus test
Onečišćenje okoliša moţe dovesti do pojave citogenetičkih oštećenja, a kao česta metoda za
odreĎivanje genotoksičnog učinka, zbog svoje jednostavnosti i učinkovitosti, koristi se
mikronukleus test. Mikronukleus nastaje zbog oštećenja kromosoma (npr. loma kromosoma)
ili poremećaja funkcije diobenog vretena pa se pojedini dijelovi ili cijeli kromosomi ne uspiju
integrirati u jezgru stanice nastale diobom. Mikronukleus se pod svjetlosnim mikroskopom
moţe vidjeti kao mala jezgra čiji promjer ne prelazi 1/3 promjera glavne jezgre (Fenech 2000;
Fenech 2003). Za razliku od artefakata nastalih bojanjem preparata ne reflektira svjetlost, nije
povezan s jezgrom, boji se jednako kao i stanična jezgra (Fenech 2000; Jiraungkooskul i sur.
2007). Kod riba koje nisu bile izlagane genotoksičnim agensima eritrociti s mikronukleusom
se relativno rijetko pojavljuju te njihova spontana pojava varira ovisno o vrsti ribe. Učestalost
spontano nastalih razina MN-a kreće se od 0,017 % kod orade (Sparus aurata) (Strunjak-
Perović i sur. 2009), 0,02-0,03 % kod šarana (Cyprinus carpio) (Klobučar i sur. 2010), 0,03 %
kod vrste Puntius altus (Jiraungkoosskul i sur. 2007) do 0,06 % kod vrste Trematomus

15
newnesii (van Ngan i sur. 2006). Kod tilapije (Oreochromis niloticus) MN se pojavljuje
prosječno 0,04 % (de Campos Ventura i sur. 2007) s najmanjom frekvencijom u kasnu jesen
(0,0067 %) a najviše u ljeto (0,034 %) (da Silva Souza i Fontanetti 2006). Pojava eritrocita s
mikronukleusima povezana je s utjecajem genotoksikanata u vodi, čak i kad su oni kronično
prisutni i u manjim koncentracijama. Učestalost pojave MN veća je s povećanjem
koncentracije genotoksikanata (Carola i sur. 2014). Izlaganje teškim metalima (bakar i
kadmij) moţe dovesti do pojave jezgrinih nepravilnosti (ENA; eng. erythrocytic nuclear
abnormalities) u eritrocitima riba (Günner 2011). ENA podrazumijevaju morfološke
promjene oblika jezgre crvenih krvnih zrnaca. In vivo i in vitro istraţivanja genotoksičnog
potencijala olova nisu utvrdila povećan broj mikronukleusa u odnosu na ostale nepravilnosti
jezgri, tako da se u istraţivanju Monteiro i sur. (2011) ENA pokazao kao bolji biomarker za
ispitivanje genotoksičnog utjecaja olova. Poredbena istraţivanja stupnja genotoksičnih
učinaka teških metala ( , , , i ) u nezagaĎenoj vodi i vodi zagaĎenoj
nepročišćenim komunalnim otpadnim vodama, te u vodama intenzivne poljoprivredne
proizvodnje ukazuju na mogući genotoksični potencijal i uvjetovanost pojave MN-a i ENA-e
nakon izloţenosti teškim metalima (Omar i sur. 2012).
2.10. Oksidacijski stres
Do oksidacijskog stresa moţe doći na razini stanica, tkiva ili čitavog organizma, što za
posljedicu ima lipidnu peroksidaciju s reaktivnim aldehidima kao krajnjim proizvodima.
Nusproizvod lipidne peroksidacije su reaktivne tvari tiobarbiturne kiseline (eng.
thiobarbituric acid reactive substances; TBARS) i često se koriste kao biomarker
oksidacijskog stresa (Ribera i sur. 2001).
Uzrok oksidacijskom stresu mogu biti patološka stanja kao što su razne ozljede tkiva, šok,
upala, sepsa, ishemija i reperfuzija, tumori, metaboličke bolesti i imunološki poremećaji.
(Ţarković i sur. 2001). Prilikom oksidacijskog stresa dolazi do pomaka ravnoteţe u staničnim
osidacijsko-redukcijskim reakcijama, što dovodi do prekomjernog stvaranja slobodnih
radikala, tj. atoma ili molekula koji u vanjskoj ljusci imaju nespareni elektron (Poli i Parola
1997; Sies 1985). Na taj se način narušava ravnoteţa slobodnih radikala i mogućnost stanice
da ih neutralizira. Kako je redukcija kisika nepotpuna, dolazi do stvaranja niza reaktivnih
kisikovih spojeva (Reactive Oxygen Species, ROS). ROS je zajednički naziv za radikale
kisika kao i za njegove reaktivne neradiklane derivate kao što su superoksidni anion

16
perhidroksilni radikal hidroksilni radikal ( vodikov peroksid ,
hipokloritna kiselina (HClO) i drugi. Uz reaktivne kisikove spojeve, veliki značaj imaju i
reaktivni dušikovi spojevi (RNS; engl. reactive nitrogen species) u koje se ubrajaju dušikovi
slobodni radikali kao što su dušikov(II)oksid ( i dušikov(IV)oksid ( , te spojevi i
molekule kao peroksinitrit ( ) i nitrozilni kation ( ). Neki se reaktivni spojevi mogu
oksidoredukcijskim reakcijama prevesti u još agresivnije oblike. Npr. superoksid moţe
reagirati s vodikovim peroksidom čime nastaje vrlo toksičan hidroksilni radikal (1), a u
reakciji s dušik(II)oksidom (NO) nastaje izuzetno agresivan oblik, peroksinitrit (2).
•- • -
2 2 2 2O + H O OH + O + OH (1)
•- -
2O + NO ONOO (2)
Izvori slobodnih radikala mogu biti endogeni i egzogeni. Endogeni slobodni radikali u
organizmu mogu nastati tijekom metabolizma kisika, fagocitoze, kemotaksije, apoptoze,
koagulacije, hipoksije ili hiperoksije. Egzogeni izvor slobodnih radikala mogu biti dim
cigareta, lijekovi, prehrana, pesticidi, radioaktivno zračenje ili UV-zračenje (Šmuc, 2009) .
Uslijed oksidacijskog stresa najčešće dolazi do oštećenja DNA koja mogu izazvati promjenu
(mutaciju) ili smrt stanice. ROS uzrokuju oštećenja proteina sa sulfhidrilnom skupinom a
lipidna peroksidacija dovodi do kroničnih poremećaja koji mogu ugroziti ţivot jedinke
(Ţarković 2000). Mjerenje razine ovih spojeva antioksidacijske obrane koristi se kao
biomarker toksičnog djelovanja onečišćenja (Ašperger i sur. 2013).
2.11. AMES test
AMES test, kao metodu detekcije mutagena, u znanost je po prvi puta 1973. g. uveo Bruce
Ames. S obzirom na to da većina mutagenih/kancerogenih tvari oštećuje DNA molekulu i
izaziva mutacije, princip AMES testa zasniva se na utvrĎivanju genske mutacije kao odgovora
na djelovanje pojedinog spoja. Test se provodi pomoću posebno konstruiranih test-sojeva
bakterije vrste Salmonella typhimurium koja nosi mutaciju u genu za sintezu histidina, tj.
auksotrofa za histidin (his-). U istraţivanjima se najčešće koriste 2 soja bakterije S.
typhimurium: TA 100 i TA 98. TA 100 je oznaka za uzgojen mutantni soj u genu za histidin
G46. TA 98 je oznaka za uzgojen mutantni soj u genu za histidin D3052. Oba soja su mutanti
u genu za liposaharidnu ovojnicu i genu za popravak DNA. Ovi sojevi dodatno sadrţe R
faktor plazmid (i otpornost na ampicilin) koji ih čini izuzetno osjetljivim na velik broj

17
kancerogena u smislu stvaranja povratnih mutacija, te posebno otpornim na toksičnost
njihovih meĎuprodukata.
Dodatak mutagena u hranjivu podlogu s navedenim sojevima izazvat će mnogo mutacija, a
njihov mali dio predstavljat će i reverziju prvobitne mutacije u genu za biosintezu histidina, te
će u tako mutiranih bakterija ponovo doći do sinteze histidina. Ti se revertanti razmnoţavaju
bez dodatka histidina u hranjivoj podlozi, ubrzo izrastu vidljive kolonije koje se mogu brojat i,
a broj revertanata predstavlja kriterij za mutagenost (ISO 162420 2005; Mortelmans i Zeiger
2000; Pavlica 2012). Ukoliko je pojedini spoj mutagen, vjerojatnost da je istovremeno i
kancerogen iznosi 90 %. Druga vaţna značajka jest da se testiranje spojeva Ames testom
izvodi dvojako: sa i bez metaboličke aktivacije. Naime, enzimski sustavi u jetri i drugim
tkivima sisavaca mogu neke potencijalne karcinogene pretvoriti u njihove aktivne oblike.
Kako bakterije S. typhimurium nemaju tih enzima oni se prilikom testa moraju dodati u
hranjivu podlogu kako bi se mogli detektirati i tzv. premutageni, odnosno prekarcinogeni.
Kako bi rezultati dobiveni AMES testom bili primjenjivi i na eukariotske organizme, kao
aktivacijski metabolički sustav u in vitro testovima na mutagenost uvodi se
postmitohondrijska frakcija (S9) homogenata jetre štakora tretiranih s jednom od tvari koje
induciraju enzimski sustav oksigenaza miješanih funkcija ovisnih o citokromu P450 (Sinal i
sur. 1999; ISO 162420 2005). Direktni mutageni mogu se dokazati u opisanom postupku i bez
dodatka S9 frakcije, te se takav test naziva AMES test bez aktivacije. Najveća prednost
AMES testa u usporedbi sa ţivotinjskim testom na kancerogenost je mogućnost testiranja
sloţenih uzoraka, kako se oni u okolišu jedino i nalaze (voda, zrak, hrana i aditivi, nafta,
pesticidi, kozmetika, lijekovi i dr.).

18
3. MATERIJALI I METODE
3.1. UreĊaj za biološko proĉišćavanje otpadnih voda i lokacije za uzimanje
uzoraka
Na ureĎaju za biološko pročišćavanje (UBP) otpadnih voda grada Virovitice godišnje se
pročisti 1 900 000 otpadnih voda iz grada Virovitice s prigradskim naseljima i 250 000
otpadnih voda Tvornice šećera Viro (TŠ), a aglomeracija koja pripada ureĎaju broji
otprilike 21 000 stanovnika. Projektirani kapacitet UBP-a je 41 000 E.S. (1 E.S.=60gBPK-
5/dan). Hidrauličkom i organskom opterećenju sustava za sakupljanje i odvodnju otpadnih
voda koje pristiţu na UBP od gospodarskih subjekata najviše pridonosi Opća bolnica
Virovitica s 300 postelja u bolničkoj zdravstvenoj zaštiti, 53 postelje u dnevnoj bolnici i 655
zaposlenika (Izvješće financijske revizije za 2011. godinu Drţavnog ureda za reviziju).
Godišnje se u sustav za sakupljanje i odvodnju otpadnih voda isporuči 46 938 m3 bolničkih
otpadnih voda (4,9 % od otpadne vode svih korisnika javne kanalizacije) s korekcijskim
koeficijentom k1=1,68 (korekcijski koeficijent k1 izraţava sastav otpadnih voda kroz
pokazatelje onečišćenja voda prisutnost opasnih i drugih onečiščujućih tvari u otpadnim
vodama, NN 82/10) i prosječnim dnevnim opterećenjem od 40 kg BPK-5. Virkom d.o.o.
Virovitica ispušta godišnje 26 695 m3 otpadne vode (o.v.) od pranja pješčanih filtara za
pripremu vode za piće, Silosi d.d. Virovitica ispuštaju 7 251 m3 o.v. s k1=2,70 i 19,07 kg
BPK-5/dan, Hrvatski duhani d.d. 7 426 m3
o.v. s k1=1,17 i 2,53 kg BPK-5/dan , R.S. Metali
d.d. Pogon Rapid Virovitica 4 121 m3 o.v. s k1=0,29 i 0,11 kg BPK-5/dan, TVIN d.d. 2 346
m3 o.v. s k1=1,08 i 2,98 kgBPK-5/dan i Vitrex d.d. 907 m
3 o.v. s k1=0,51 i 0,11 kg BPK-
5/dan. Na ureĎaj dolazi 30-50 % drugih voda poput procjednih i oborinskih voda. Godišnji
doprinos ostalih gospodarskih subjekata manji je od 900 m3 (Kepec S, 2015). Slika 1.
prikazuje shemu projektiranog i izgraĎenog UBP-a grada Virovitice. Primarno pročišćavanje
otpadnih voda odvija se na automatskoj rešetki s udaljenošću izmeĎu štapova od 5 cm,
uzduţno aeriranom pjeskolovu i mastolovu ukupnog kapaciteta 390 L/sek i volumena od 120
m3. Sekundarno pročišćavanje odvija se u dva bioaeracijska bazena ukupnog volumena 8 000
m3 opremljenih plivajućim turbinama za aeraciju, dvije sekundarne taloţnice sa zgrtačem
aktivnog mulja ukupnog volumena 1 160 m3 i crpilištem aktivnog mulja iz kojeg se dio mulja
(povratni mulj) vraća u bioaeracijske bazene, a višak mulja u lagune šećeranskih otpadnih
voda.

19
Slika 1. Projektirani i izgraĎeni UreĎaj za biološko pročišćavanje otpadnih voda grada
Virovitice (UBP)
IzgraĎeno:
1: Taloţne lagune za karbonatni mulj, 2: Taloţne lagune za zemljani mulj, 3: Kompenzacijska
laguna, 4: Rezervna laguna, 5: Crpilište tehnološke otpadne vode TŠ, 6: Izmjenjivač topline,
7: Anaerobni fermentor, 8: Otplinjač, 9: Lamelni separator, 10: Crpilište anaerobnog mulja,
11: Bioaeracijski bazen, 12: Sekundarna taloţnica, 13: Crpilište aerobnog mulja, 14:
Plinosprem, 15: Baklja za spaljivanje bio-plina, 16: Crpilište komunalne otpadne vode, 17:
Automatska fina rešetka, 18: Aerirani pjeskolov-mastolov, 19: Mjerni ţlijeb
Nije izgraĎeno:
20: Primarni zguščivač mulja, 21: Izmjenjivač topline, 22: Digestor za anaerobnu razgradnju
mulja, 23: Sekundarni zguščivač mulja, 24: Strojna dehidracija mulja, 25: Plinosprem, 26:
Baklja za spaljivanje bio-plina.
11 11
19 18
IZ GRADA
ODVOZ NA DEPONIJU
16
17
7
8 9
5
15
14
6
10
ODVOZ MULJA
1
4
1
2
3
2
Manteč 13
26
22 25
24 23 20
21
O
12 12

20
Kanal Manteč je recipijent pročišćenih otpadnih voda UBP-a, kao i voda Biljnog ureĎaja
Lukač (BU) na kojem se pročišćuju otpadne vode naselja Lukač s 443 stanovnika. Godišnje se
na ovom ureĎaju pročisti 5 000 otpadne vode. Kanal Manteč ulijeva se u Ţupanijski kanal
kod naselja Budrovac, a Ţupanijski kanal ulijeva se u Dravu kod naselja Sopje (Slika 2).
Uzimanje uzoraka za istraţivanja provedeno je u proljeće, ljeto i jesen 2014. godine (Tablica
1, Slika 2.). Na taj je način obuhvaćen reţim rada UBP-a u kojem se pročišćuju komunalne
otpadne vode sa i bez tehnoloških otpadnih voda TŠ-a.
Tablica 1. Prikaz uzoraka prema lokacijama uzorkovanja. Datum i broj uzorkovanih jedinki i
uzoraka prema lokacijama uzorkovanja: a) Ribe - patološko-anatomski pregled,
mikrobiološke, genotoksikološke, histopatološke analize b) Voda - fizikalno-kemijske
analize, analize teških metala i mikrobiološke analize, c) Aktivni mulj za fizikalno-kemijske
analize, analize teških metala i mikrobiološke analize, d) Voda za analitičko-kemijske analize,
e) Sediment za analitičko-kemijske analize. Uzorci vode pod d) su kompozitni uzorci
volumena 1 000 mL uzimani u 08:30, 14:00 i 20:30 sati, dok su uzorci mulja i sedimenta
trenutni, uzimani od 07:00 do 10:00 sati volumena 1 000 mL.
Tablica 1. Prikaz uzoraka prema lokacijama uzorkovanja
a) Ribe b) Voda c) Mulj d) Voda e) Sediment
Oznaka
lokacije
uzorkovanja
prema Slici
2.
22.0
4.2
014.
08.0
7. 2014.
14.1
0. 2014.
22.0
4. 2014.
01.0
7. 2014.
26.0
9. 2014.
22.0
4. 2014.
01.0
7. 2014.
26.0
9. 2014.
22.0
4. 2014.
08.0
7. 2014.
14.1
0. 2014.
22.0
4. 2014.
1 5 4 25 1 1 1
2 1 1 1 3 3 3
3 1 1 1
4 1 1 1 3 3 3 1
5 1 1 1 1
6 1 1 1
7 1 1 1
8 5 6 11 1 1 1
9 20 9 19 1 1 1 1
10 1 1 1
11 1 1 1

21
Slika 2. Prikaz lokacija uzorkovanja
Lokacije uzimanja uzoraka su bile: 1. potok OĎenica prije grada Virovitice, kao vodotok za
koji se pretpostavlja da nije recipijent industrijskih otpadnih voda niti je pod utjecajem
degradacije kakvoće vode zbog poljoprivredne djelatnosti, 2. ulaz gradskih otpadnih voda na
UBP, 3. ulaz tehnoloških otpadnih voda TŠ-a na UBP, 4. izlaz pročišćenih otpadnih voda iz
preljevnog kanala sekundarne taloţnice UBP-a, 5. kanal Manteč nakon prihvata pročišćenih
otpadnih voda sa UBP-a prije BU-a, 6. kanal Manteč nakon BU-a, 7. kanal Manteč
neposredno prije utoka u Ţupanijski kanal, 8. Ţupanijski kanal nakon utoka kanala Manteč, 9.
Ţupanijski kanal nizvodno prema rijeci Dravi u naselju Brezovica, 10. mjesto izljeva
povratnog aktivnog mulja u bioaeracijski bazen UBP-a, 11. mulj s deponije viška aktivnog
mulja i zemlje od pranja šećerne repe. Uzorci vode i mulja uzimani su u sterilne staklene boce
i polipropilenske vrećice, hlaĎeni za vrijeme transporta u laboratorij, a analize su obavljene
odmah nakon dostave u roku od najviše 3 sata.

22
3.2. OdreĊivanje fizikalno-kemijskih pokazatelja
Temperatura vode i otopljeni kisik u vodi mjereni su prilikom uzimanja uzoraka
multiparametarskim mjeračem (HQ40d, Hach Lange, Njemačka) na 10-15 cm dubine.
Koncentracija vodikovih iona mjerena je u laboratoriju pH metrom (MP 220 k, Mettler
Toledo, Njemačka), koji je umjeravan certificiranim otopinama pufera (Mettler Toledo).
Elektroprovodljivost je odreĎivana konduktometrijski (MC226k, Mettler Toledo), a
instrument je umjeren certificiranom puferskom otopinom od 1,413 µS/cm. Suspendirana tvar
odreĎivana je gravimetrijski iz 500 mL homogeniziranog uzorka (ili alikvota) koji se filtrirao
preko filtar papira promjera 47 mm, veličina pora 1,1 µm (Glass-Microfibre disc, Watman 11,
Sigma-Aldrich, SAD). Membranska filtracija uzorka kroz filtar papir, prethodno sušen na 105
°C, 2h, obavljena je vakuum crpkom (MZ 2, Wacuubrand, Njemačka), nakon čega je sušen
na 105 °C, min. 2h (UNE 400, Memmert, Njemačka) do konstantne teţine. Nakon svakog
sušenja i hlaĎenja u eksikatoru, filtar papir je vagan (Ohaus Discovery, Švicarska). Umnoţak
razlike odvaga i razrjeĎenja daje vrijednost suspendirane tvari (mg/L).
Kemijska potrošnja kisika uz korištenje kao oksidansa (KPKCr) odreĎena je
spektrofotometrijski (UV/VIS, HACH DR 5000). Za eliminaciju klorida dodaje se , a
za digestiju 3 mL otopine koja se dobije miješanjem otopine i otopine u 98
%-tnoj . Digestija je provedena pri 148 °C kroz 2h u termoreaktoru (TR 300, Merck,
Njemačka). KPKCr je očitana spektrofotometrijski (Hach DR 5000, Njemačka) i baţdarnim
pravcem postavljenim u deset koncentracijskih vrijednosti (0 do 1 000 mgO2/L pri 620 nm).
Za pripremu standardne otopine KPK korišten je .
Potrošnja (KPKMn) odreĎena je kuhanjem 100 mL (ili alikvota) uzorka uz dodatak 5
mL razrjeĎenja 1+3 i 15 mL 0,002 mol/L (0,01 N) . Nakon kuhanja od 10
minuta dodano je 15 mL 0,005 mol/L i titrirano s 0,002 mol/L , a rezultat je
izraţen u mg .
Biokemijska potrošnja kisika u pet dana (BPK-5) odreĎena je tako da se, uvaţavajući
izmjeren KPK i/ili KPKMn, izračunalo razrjeĎenje uzorka otpadne ili površinske vode. Uzorci
su razrijeĎeni aeriranom otopinom za razrjeĎivanje, tj. demineraliziranom vodom u koju je
dodana fosfatna pufer otopina. Iz razlike u vrijednostima otopljenog kisika prvog i petog

23
dana, i uz uvaţavanje stupnja razrjeĎenja uzorka, izračunata je potrošnja kisika za biološku
oksidaciju organske tvari u uzorku u pet dana u mg /L.
Za odreĎivanje amonijaka u vodi korištena je spektrofotometrijska metoda (655 nm) sa
salicilatom nakon dodavanja salicilat-citratne otopine i otopine pripremljene od NaOH i
(izosan G) u 40 mL uzorka (ili alikvota). Nitriti su odreĎivani
spektrofotometrijski (520 nm) metodom s Za odreĎivanje nitrata korištena je
spektrofotomerijska metoda u UV području (220 nm) uz dodatak 1 N HCl. Ortofosfati su
odreĎivani spektrofotometrijski (690 nm) metodom s . Ukupni dušik
odreĎivan je korištenjem kivetnog testa Hach Lange LCK 238. Za odreĎivanje ukupnog
fosfora korišteni su Hach Lange kivetni testovi LCK 348 i LCK 349. OdreĎivanje ukupnog
dušika i fosfora provedeno je zagrijavanjem na 100 °C (Hach DRB 200).
Za odreĎivanje koncentracije teških metala uzorak mulja je osušen i homogeniziran, a 2-3 g
uzorka razoreno je koncentriranom (Merck). Sadrţaj kadmija, bakra, olova, cinka,
kroma i nikla u vodi i mulju odreĎen je grafitnom tehnikom (AA 600, Perkin Elmer, SAD),
dok je sadrţaj cinka odreĎivan induktivno-spregnutom plazmom (ICP-OES 2100 DV, Perkin
Elmer). Kalibracijske krivulje napravljene su u četiri točke i to za Cd (raspon od 0,05-2 µg/L),
Cr (raspon 5-50 µg/L), Cu (raspon od 10-100 µg/L), Ni (raspon od 5-50 µg/L), Pb (raspon od
1-20 µg/L), Zn (raspon od 5-100 µg/L). Sadrţaj ţive odreĎen je hidridnom tehnikom (FIAS
100, Perkin Elmer). Kalibracijske krivulje za ţivu napravljene su u četiri točke u rasponu od
1-10 µg/L. Svi osnovni standardi su od 1 000 mg/L, single element, Merck. Ţeljezo je
odreĎeno spektrofotometrijskom Hachovom metodom na valnoj duljini od 510 nm. Svaki
uzorak analiziran je u tri replike.
3.3. Analitika vode i sedimenta
Uzorci vode i sedimenta uzimani su u polikarbonatne boce od 250 mL koje su do analize
drţane na temperaturi od 4 °C. Uzorci vode u 1/3 od ukupnog volumena uzimani su u tri
vremenska termina (08:00, 14:00 i 20:30 sati) sa dva lokaliteta: ulaz komunalnih otpadnih
voda na UreĎaj (lokalitet 2) i izlaz pročišćenih otpadnih voda (4). Uzorkovanje sedimenta
izvršeno je ujutro na tri lokacije: izlaz pročišćenih otpadnih voda (4), kanal Manteč kao
prijemnik pročišćenih otpadnih voda (5) i Ţupanijski kanal (9). Analiza vode obavljena je

24
nakon filtriranja uzorka kroz PTEF filtar s porama od 0,2 µm. Usporedno s analizom uzoraka
vode izvršena je i analiza ultra čiste vode i sedimenta. Za analize uzoraka sedimenta korištena
je metoda prema Darwano i sur. (2014). Za odreĎivanje farmaceutika u vodi i sedimentu
korišteni su ultra čisti reagensi LC MS čistoće. OdreĎivanje farmaceutika izvršeno je
tekućinskom kromatografijom ultra visoke djelotvornosti s kvadrupolnom time-of-flight
(TOF) masenom spektrometrijom (UHPLC-QTOF-MS). Istraţivanja su provedena na 1290
UHPLC sistemu (Agilent Technologies, SAD) pri čemu je korišten automatski uzorkivač
G4226A, binarna crpka G4220B i termostatirana kolona G1316C. Razdvajanje ispitivanih
analita izvršeno je pomoću kolone RP ACQUITY UPLC, HSST3 (15 mm x 2,1 mm, 1,8 µm).
Volumen injektiranja bio je 100 µL a temperatura komore kolone postavljena je na +50 °C. U
slučaju ESI (ESI(+)), mobilna faza sastojala se od otopine A (mM ) i otopine
B (100 % ). Korišten je gradijent eluacije protoka 0,4 mL/min. Analiti su odreĎivani
korištenjem instrumenta 6550 i-Funnel Q-TOF-LC/MS (Agilent Technologies) u 4 GHz
ratom detektora s moći razlučivanja 40 000 i <2 ppm točnosti. Radilo se u pozitivnom (ESI+)
ionskom modu QTOF masenog spektrometra (Agilent 6550) a ioni su generirani pomoću
dualnog izvora iona AJS ESI (Agilent Jet Stream). Baţdarni pravci izraĎeni su direktnim
injektiranjem standardnih otopina sedam različitih koncentracijskih vrijednosti s
koeficijentom korelacije >0,999 kao prihvatljivim kriterijem linearnosti. Testiranjem vršnih
vrijednosti slijepe probe i tri koncentracijske vrijednosti izračunata je preciznost, te je
evaluirana u četiri replike. Uvjeti rada bili su slijedeći: temperatura plina poklopca +375 °C,
temperatura plina 125 °C, vrući plin 12 L , kapilarni napon 3 500 V, fragmentor 400
V i raspršivač 2,413 bara. Korekcija prilikom mjerenja za svako očitanje na osi mase bila je
automatska. Analize su izvedene u MS i MS/MS modulu s fiksnom energijom sudara i
rasponom mase 50 do 1 000 m/z.
3.4. Ţivotinjske vrste i obrada tkiva
3.4.1. Ribe
Rukovanje ţivotinjama obavljeno je u skladu s Direktivom Vijeća 86/609/EEZ i Pravilnikom
o zaštiti ţivotinja koje se koriste u pokusima ili u druge znanstvene svrhe (NN 47/11).

25
Ribe su lovljene mreţama i udicama u potoku OĎenica na mjestu gdje voda nije pod
utjecajem otpadnih voda, na mjestu gdje se kanal Manteč koji je recipijent pročišćenih
otpadnih voda ulijeva u Ţupanijski kanal, i nizvodno iz Ţupanijskog kanala kod naselja
Brezovica (Slika 2.). Gdje god je to bilo moguće, ribe su u mreţi drţane u vodi na mjestu
ulova do prijevoza u laboratorij. Ribe su prevezene u laboratorij u posudama s vodom uz
stalnu aeraciju do trenutka razudbe.
Odrasle spolno zrele gujavice (Eisenia fetida) sa razvijenim klitelumom odabrane su na
uzgajalištu Eršek u Donjoj Bistri te su dopremljene u laboratorij gdje su odrţavane u
kontroliranim uvjetima (tama, ± 21 °C), u tlu nepromjenjenog sastava (kompost, ilovača i
treset jednakih udjela). Gujavice su hranjene jednom tjedno organskom hranom iz
kontroliranog uzgoja bez pesticida uz redovito vlaţenje tla.
3.4.1.1. Ribe, pregled i uzimanje tkiva za analize
Najbrojnija ulovljena vrsta bila je babuška (Carassius gibelio), zatim slijedi somić (Ictalurus
nebulosus), klen (Squalius cephalus), deverika (Abramis brama), bjelica (Rutilus rutilus),
uklija (Alburnus alburnus) i linjak (Tinca tinca). Od svih ulovljenih riba 35 % je bilo
muţjaka, a 42 % ţenki, dok za 23 % riba spol nije odreĎen. Rezultati analiza koje se odnose
na ribe u ovom radu prikazani su samo za babušku zbog najvećeg broja ulovljenih jedinki
ove vrste.
Slika 3. Babuška (Carassius gibelio), ulovljena 08.07.2014. u Ţupanijskom kanalu nizvodno,
muţjak, duţine 202 mm, teţine 223 g.

26
Ribama je odreĎena teţina (g) i duţina od vrha glave do račvanja repne peraje (mm), te im je
iz repne arterije/vene izvaĎena krv. Za biokemijske analize krv je ubrizgana u tubice
presvučene antikoagulansom litij-heparinom, centrifugirana na 12 000 x g kroz 90 s (StatSpin,
Idexx, SAD), a dobivena plazma pohranjena je na -20 °C do analiza. Heparinizirane
mikrohematokritske kapilare punjene su krvlju, zatvorene punilom i centrifugirane na 12 000
x g kroz 120 s najkasnije do 20 minuta po vaĎenju krvi. Hematokrit (Hct) je odreĎen kao
volumni postotak crvenih krvnih zrnaca u krvi (Wedemeyer and Yasutake, 1977). Za
MN/ENA test tri krvna razmaza po ribi osušena su na zraku, fiksirana 95 %-tnim metanolom
kroz 3 minute i obojena MayGrünwald/Giemsa bojanjem.
Ribe odreĎene za pregled ţrtvovali smo prekomjernom dozom anestetika MS-222 (Sigma-
Aldrich). Zdravstveni pregled riba obuhvaćao je opći klinički pregled, mikroskopski pregled
nativnih preparata obrisaka koţe, peraja i škrga, te patološko-anatomsku pretragu. Materijal
škrga, bubrega, jetre i slezene nacijepljen je na Tryptone Soya Agar (Oxoid, UK), krvni agar
(Certifikat d.o.o, Osijek, Hrvatska), Mac-Conkey Agar (Oxoid). Materijal škrga i koţe
nacijepljen je i u 0,09 % NaCl za dodatnu mikrobiološku pretragu.
27
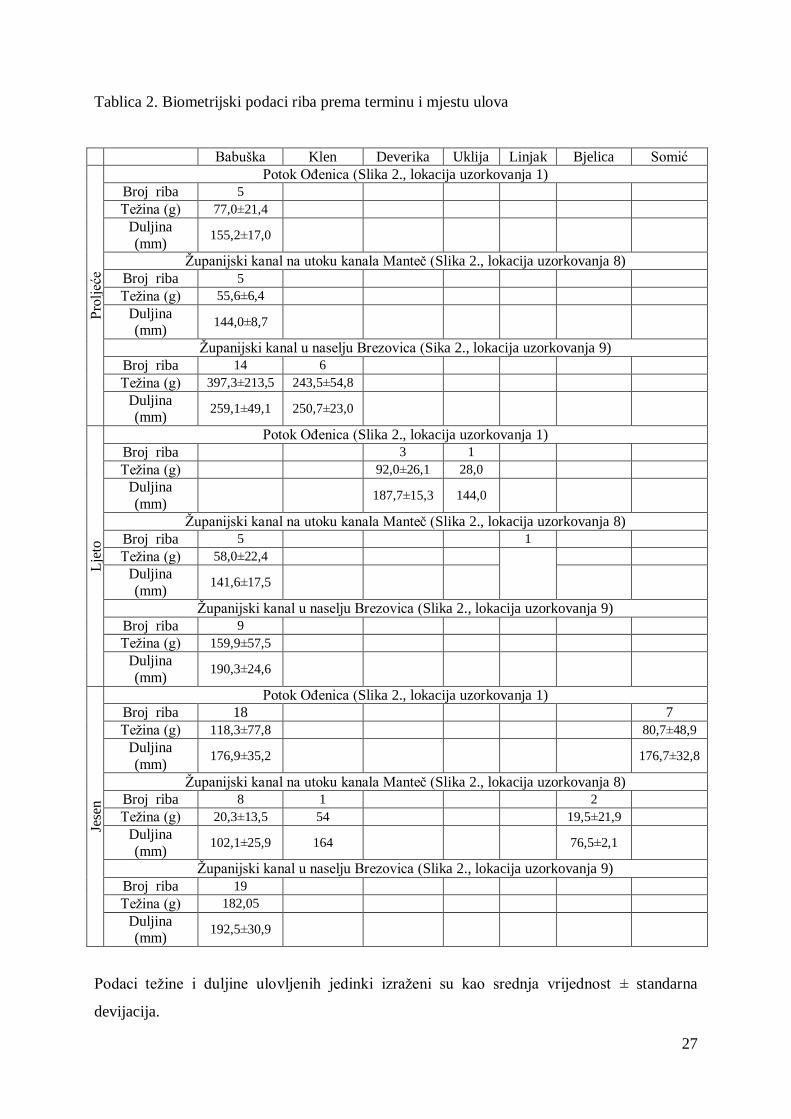
Tablica 2. Biometrijski podaci riba prema terminu i mjestu ulova
Babuška Klen Deverika Uklija Linjak Bjelica Somić
Pro
ljeć
e
Potok OĎenica (Slika 2., lokacija uzorkovanja 1)
Broj riba 5
Teţina (g) 77,0±21,4
Duljina
(mm) 155,2±17,0
Ţupanijski kanal na utoku kanala Manteč (Slika 2., lokacija uzorkovanja 8)
Broj riba 5
Teţina (g) 55,6±6,4
Duljina (mm)
144,0±8,7
Ţupanijski kanal u naselju Brezovica (Sika 2., lokacija uzorkovanja 9)
Broj riba 14 6
Teţina (g) 397,3±213,5 243,5±54,8
Duljina (mm)
259,1±49,1 250,7±23,0
Lje
to
Potok OĎenica (Slika 2., lokacija uzorkovanja 1)
Broj riba 3 1
Teţina (g) 92,0±26,1 28,0
Duljina
(mm) 187,7±15,3 144,0
Ţupanijski kanal na utoku kanala Manteč (Slika 2., lokacija uzorkovanja 8)
Broj riba 5 1
Teţina (g) 58,0±22,4
Duljina
(mm) 141,6±17,5
Ţupanijski kanal u naselju Brezovica (Slika 2., lokacija uzorkovanja 9)
Broj riba 9
Teţina (g) 159,9±57,5
Duljina
(mm) 190,3±24,6
Jese
n
Potok OĎenica (Slika 2., lokacija uzorkovanja 1)
Broj riba 18 7
Teţina (g) 118,3±77,8 80,7±48,9
Duljina
(mm) 176,9±35,2 176,7±32,8
Ţupanijski kanal na utoku kanala Manteč (Slika 2., lokacija uzorkovanja 8)
Broj riba 8 1 2
Teţina (g) 20,3±13,5 54 19,5±21,9
Duljina
(mm) 102,1±25,9 164 76,5±2,1
Ţupanijski kanal u naselju Brezovica (Slika 2., lokacija uzorkovanja 9)
Broj riba 19
Teţina (g) 182,05
Duljina (mm)
192,5±30,9
Podaci teţine i duljine ulovljenih jedinki izraţeni su kao srednja vrijednost ± standarna
devijacija.

28
Za pripremu histoloških preparata izdvojena su tkiva škrga, slezene, bubrega i jetre koja su do
analiza fiksirana u 10 %-tnom neutralnom formalinu.
3.4.1.2. Histološke analize
Nakon fiksacije, tkiva su uklopljena u parafinske blokove koja su zatim rezana na rezove
debljine 3 µm. Dobiveni rezovi deparafinirani su ksilolom i padajućim koncentracijama
etanola, obojeni hemalaunom i eozinom, isprani i dehidrirani rastućim koncentracijama
etanola i ksilola te uklopljeni u kanadski balzam (Švob 1974). Parafinski blokovi korišteni su
za bojanje PAS metodom po McManusu (Pearse 1968) za prikaz lokalizacije mukoznih
stanica i glikogena. Metoda se zasniva na oksidaciji polisaharida s perjodnom kiselinom uz
nastanak aldehidnih skupina, a nakon tretiranja sa Schiffovim reagensom dodani bazični
fuksin veţe se duţ lanca polisaharida dajući crvenu do purpurnu boju. Histološki preparati
pregledani su na Olympus BX51 svjetlosnom mikroskopu. Patološko-histološke alteracije
mjerene su pomoću računalnog programa Microsoft AnalySIS Imaging System.
3.4.1.3. Analize krvi riba
Koncentracije glukoze (GLU), uree (URE), kreatinina (CRE), kolesterola (CHOL),
triglicerida (TRIG), ukupnih bjelančevina (TP), albumina (ALB), aktivnosti alanin
aminotransferaze (ALT), alkalne fosfataze (ALP), gama-glutamil transferaze (GGT)
odreĎivane su komercijalnim testovima Beckman Coulter (Olympus Life and Material
Science Europe, Irska) i biokemijskim analizatorom Olympus AU 640 (Olympus, Japan).
Aktivnost superoksid dizmutaze (SOD) i glutation peroksidaze (GSH-Px) odreĎivane su
Randox komercijalnim reagensima na Olympus AU 640 biokemijskom analizatoru. Aktivnost
paraoksonaze (PON 1) u plazmi testirana je modificiranom metodom hidrolize paraoxona
prema Charlton-Menys i sur. (2006) na Olympus AU 640.
3.4.1.4. Istraţivanje genotoksiĉnosti
Istraţivanje genotoksičnosti provedeno je na eritrocitima babuški primjenom mikronukleus
testa (MN test) i analizom nepravilnosti jezgre eritrocita (ENA test). Nakon sušenja krvnih

29
razmaza na zraku, fiksirani su 95 %-tnim metanolom (3 minute), a zatim obojani May-
Grünwald/Giemsa bojama. Svjetlosnim mikroskopom (Olympus BX51, povećanje 1000x)
pretraţeno je 1000 eritrocita po jednom krvnom razmazu. Morfološke promjene eritrocita
pretraţivane su prema Carrasco i sur. (1990) i Carrola i sur. (2014), a uključivale su
polimorfne jezgre (nepravilna i nekonzistentna oblika), segmentirane jezgre, jezgre
bubreţastog oblika, jezgre s vakuolama, jezgre s pupovima, jezgre s invaginacijama i
binuklearne eritrocite. Sve navedene kategorije morfoloških promjena prikazane su zbirno kao
ukupne nepravilnosti eritrocitnih jezgri, a rezultati su prikazani u ‰.
3.4.2. Gujavice
3.4.2.1. Gujavice, predtretman: Test kontakta na filtar papiru
Gujavice (190-270 mg) rasporeĎene su u staklenke obloţene navlaţenim filtar papirom kako
bi u periodu od 24 h očistile sadrţaj probavila. Nakon faze čišćenja, gujavice su stavljene na
predtretman u trajanju od 8 h, tijekom čega su izlagane trima koncentracijama otpadnih voda;
originalnom uzorku, uzorku razrijeĎenom 100x te 1000x. Predtretman je obuhvaćao stavljanje
gujavica u Petrijeve zdjelice (promjera 90 mm) dna obloţenog filtar papirom te navlaţenog sa
1,2 mL testirane koncentracije uzorka vode. Kontrolna skupina preko filtar papira izlagana je
volumenu od 1,2 mL destilirane vode.
3.4.2.2. Mjerenje MXR inhibitornog potencijala gujavica nakon izlaganja otpadnim
vodama i otpadnom mulju
Nakon 8-satnog predtretmana započet je tretman (24 h) u kojem je filtar papir navlaţen sa 1,2
mL mješavine testne otopine te 10 µM fluorescentnog supstrata rodamina (R123, Sigma-
Aldrich, Njemačka). Negativna kontrolna skupina izlagana je 10 µM R123, dok je pozitivna
kontrolna skupina izlagana mješavini 50 µM modelnog MXR inhibitora - MK571 te 10 µM
R123. Nakon završenog tretmana svaka gujavica pojedinačno isprana je destiliranom vodom
kako na kutikuli ne bi zaostao fluorescentni supstrat, izvagana je te homogenizirana (Ultra-
Turray T18. IKA, Njemačka) u 1 mL hladne destilirane vode. Homogenat je centrifugiran
Eppendorf 5417C centrifugom (Njemačka) pri brzini od 9 000 x g kroz 10 min. Po 200 µL

30
dobivenog supernatanta u duplikatima preneseno je u crne mikroploče (Nunc, Thermo Fisher
Scientific, Njemačka). Uz pomoć spektrofotofluorimetra FLUOstar OPTIMA Platereader
(BMG Labtech, UK) izmjerene su fluorescentne vrijednosti pomoću kojih smo dobili uvid u
aktivnost MXR mehanizma. Valna duljina korištena prilikom mjerenja ekscitacije R123
iznosila je 490 nm, dok je za emisiju ta vrijednost bila 544 nm. Tijekom svake faze uzorci su
drţani u tami kako bi se spriječila fotodegradacija R123. Svaka koncentracija testirana je sa
po 7 gujavica (n = 224 gujavice po ekspoziciji). Dobivene jedinice fluorescence izraţene su u
odnosu na masu svake gujavice pojedinačno te pretvorene u postotke.
Ručnim miješanjem sa tlom prikupljenim sa uzgajališta Eršek, napravljene su dvije
koncentracije aktivnog mulja: 30% i 70% te su raspodijeljene u dvije staklenke od 3 L.
Odrasle jedinke gujavica sa razvijenim pojasom izdvojene su te nasumično rasporeĎene u
staklenke sa otpadnim muljem. Gujavice su uzorkovane nakon 24 h, 48 h te 7-dnevnog
izlaganja mješavini tla, kada su uklonjene iz staklenke, isprane destiliranom vodom kako bi se
uklonio višak zemlje, te su stavljene na filtar papir navlaţen sa 1,2 mL 10 µM R123. Nakon
24-satnog izlaganja fluorescentnom supstratu, gujavice su isprane destiliranom vodom kako bi
se višak fluorescentnog supstrata uklonio sa kutikule. Pokus je proveden na isti način kao i za
otpadnu vodu.
3.4.2.3. Mjerenje oksidacijskog stresa nakon izlaganja gujavica otpadnim vodama i
otpadnom mulju
U svrhu odreĎivanja oksidacijskog stresa uzrokovanog otpadnim vodama grada Virovitice,
mjerili smo koncentraciju malondialdehida (MDA) kao pokazatelja lipidne peroksidacije.
Korištena metoda zasniva se na reakciji tiobarbiturne kiseline (TBA) sa MDA uslijed čega
nastaje crveni pigment. Svakoj gujavici pojedinačno odreĎena je masa, nakon čega je
homogenizirana u hladnom fosfatnom puferu (PBS) čiji je volumen iznosio tri udjela
volumena mase. Homogenat je zatim centrifugiran na 9 000 x g kroz 10 min. Supernatant
(250 µL) razrijeĎen je sa 500 µL smjese 10 % trikloroctene kiseline (TCA) te 0.01 %
butiliranog hidroksitoluena (BHT). Uzorci su vorteksirani na ledu te hlaĎeni 15 min na 4 °C.
Nakon hlaĎenja ponovno su centrifugirani. Odvojeno je 700 µL uzorka koji je razrijeĎen sa
500 µL TBA. Smjesa je zatim grijana 30 min na 99 °C. Kako bi se reakcija zaustavila, uzorci
su ohlaĎeni na 4 °C te je 200 µL supernatanta u duplikatima preneseno u crne mikroploče
kako bi se uz pomoć spektrofotofluorimetra izmjerila apsorbancija. Valna duţina prilikom

31
mjerenja apsorbancije postavljena je na 535 nm. Svaka koncentracija, uključujući i kontrolnu
skupinu izlaganu destiliranoj vodi, testirana je sa po 5 gujavica (n = 160 gujavica po
ekspoziciji). Vrijednost apsorbancije izraţena je u odnosu na kalibracijsku krivulju odreĎenu
na temelju prireĎenih standardnih otopina MDA poznatih koncentracija. Jedinice apsorbancije
izraţene su u odnosu na masu svake gujavice pojedinačno te pretvorene u postotke.
Nakon 24 h, 48 h i 7 dana izlaganja otpadnom mulju, gujavice su nasumično uzorkovane iz
zemlje, isprane destiliranom vodom s ciljem uklanjanja zemlje sa kutikule te su u periodu od
3 h stavljene na filtar papir navlaţen sa 1,2 mL destilirane vode kako bi se ispraznilo
probavilo. Izlaganje je provedeno na isti način kao i za otpadnu vodu.
3.5. Ames test
Kako bi se pripremila minimalna podloga za uzgoj sojeva TA 98 i TA 100, autoklaviralo se
15 g agara i 930 mL destilirane vode. Po završetku autoklaviranja otopini agara ohlaĎenoj na
oko 70°C doda se 20 ml sterilne otopine Vogel-Bonner medija (10 g MgSO4 x 7 H2O; 100 g
limunske kiseline x H2O; 500 g K2HPO4 i 175 g NaHNH4 PO4 x 4 H2O) i 50 mL sterilne
40% otopine glukoze. Nakon temeljitog miješanja, minimalna podloga se ravnomjerno
raspodjelila u Petrijeve zdjelice.
Dan prije testa hranjivi medij (0,8 % Bacto bogati medij i 0,5 % NaCl) sa bakterijskim
kulturama stavljen je u inkubatorsku tresilicu (frekvencija 250 pomaka/min, 37 °C). Nakon
12-15 sati inkubacije prekida se dioba bakterija uranjanjem posude s kulturom u ledenu kupelj
ili friţider (+4 °C) gdje se čuva do upotrebe. Koncentracija bakterija od 1 do 2 x 109 bakterija
u 1 ml medija podesila se mjerenjem turbiditeta pomoću spektrofotofluorimetra FLUOstar
OPTIMA platereader (BMG Labtech, Aylesbury, UK) na valnoj duţini 650 nm.
Prilikom priprave tzv. S9 smjese, dodano je 8 mM MgCl2, 33 mM KCl, 5 mM glukoza-6-
fosfat, 4 mM NADP, 100 mM natrij fosfatni pufer pH 7,4. 100 µL uzorka koji ispitujemo
(prethodno profiltriranog 0.2 µm filterom), 0,1 mL bakterijske suspenzije (1 – 2 x 109
bakterija) i 2,0 mL površinskog agara (0,6 % agar, 0,5 % NaC1, termostatirano na 45 °C)
pomiješa se u mikrobiološkoj epruveti i izlije (u duplikatu) u sterilne Petrijeve zdjelice s 20
ml minimalne podloge. Negativne kontrole su prirodni-spontani broj revertanata (TA 100 od
100 - 200; TA 98 od 20 - 50). Pozitivne kontrole (dijagnostički mutageni) koriste se u potvrdi
svojstva povećanja broja revertanata (His-
u His+) odreĎenog bakterijskog soja S.
typhimurium koja je istovremeno povezana s djelotvornošću (bioaktivacijskim potencijalom)

32
S9 frakcije u S9 mix. Za TA 100 koristi se benzo(a)piren (BaP), a za TA 98 koristi se 2-
aminofluoren (2-AF). Kod ispitivanja mutagenog potencijala direktnih mutagena tj. tvari za
koje nije potrebna metabolička aktivacija (- S9 mix) kao pozitivne kontrole koriste se direktni
mutageni. To su za TA 100 metil metansulfonat (MMS), a za TA 98 NOPD. Nakon
inkubacije 48 h (u mraku) na temperaturi od 37 °C broje se kolonije, koje su dobro vidljive
na ploči. Brojanje kolonije izvodi se ručno ili na automatskom brojaču kolonija. Broj
revertanata na ploči s ispitivanim spojem iznad broja spontanih mutanata mjerilo je
mutagenosti testiranog spoja. Ako je broj induciranih mutacija dvostruko veći od kontrole i
ako se unutar toga povećanja moţe utvrditi doza-odgovor, smatra se da je ispitivani spoj
mutagen. Za očekivati je da će broj revertanata u pozitivnoj kontroli biti povećan najmanje
10-puta u odnosu na negativnu kontrolu.
Pri interpretaciji rezultata bitno je znati da je temeljni kriterij u Ames testu broj pozitivnih
(His+) kolonija (revertanata) na ploči, koji u slučaju mutagene tvari mora biti najmanje
dvostruko veći od broja spontanih revertanata (kontrole). U skladu s tim kriterijem modelni
mutageni, odnosno premutageni koji se koriste kod kontrole postupka, moraju rezultirati s
brojem revertanata koji je uvjerljivo viši od broja spontanih revertanata.
3.6. Mikrobiološke analize vode, mulja i tkiva riba
Mikrobiološke analize provedene su prema metodama potank o opisanim u slijedećim
normama i pravilnicima: HRN EN ISO 6222:2000; HRN EN ISO 9308-1:2000/Corr.1:2008;
HRN EN ISO 7899-2:2000; HRN EN ISO 16266:2008; HRN EN ISO 6579:2003/Corr.
1:2008; HR EN ISO 11290-1:1999/A1:2008; HR EN ISO 11290-1:1999/A1:2008; HRN EN
ISO 26461-2:2008; HRN EN ISO 7954:2002, ISO 21527-1:2008.
Za odreĎivanje ukupnog broja bakterija u otpadnim i površinskim vodama korišten je Tryptic
Glucose Yeast agar (Biolife, Italija). Bakterije su inkubirane na 37 °C (Memmert, tip BE 400
i INB 500, Njemačka) i na 22 °C (Memmert, tip IPP 400). Porasle bakterijske kolonije koje su
se inkubirale na 37 °C brojane su nakon 48 sati, a bakterije koje su inkubirane na 22 °C
nakon 72 sata. Broj koliformnih bakterija odreĎivan je metodom membranske filtracije uz
pomoć mikrobiološke podloge m-Les Endo Agar (Biolife), inkubacija je izvršena na 37 °C
kroz 24 sata, a za potvrdne testove korišteni su Triple Sugar Iron agar (Biolife), Mac Conkey
Broth Purple (Biolife) i Bactident Oxydase trakice (Merck). Fekalni koliformi i E. coli
odreĎivani su membranskom filtracijom na selektivnoj mikrobiološkoj podlozi m-Faecal

33
coliform agar (Biolife), inkubacija je izvršena na 44 °C kroz 24 sata, a za potvrdne testovi
korišteni su Triple Sugar Iron agaru USP (Biolife), Mac Conkey Broth Purple Brothu
(Biolife), Simmons Citrate agaru (Biolife), Bacitiden Oxidase trakicama (Merck), DEV
Tryptophan Brothu (Merck), Kovačevim reagensom (Biolife) i bojenjem po Gramu. Osim
navedenih testova, koliformne bakterije i E. coli su u završnom dijelu istraţivanja ispitivani
po Colilert enzimatskoj metodi (Idexx, SAD) za odreĎivanje najvjerojatnijeg broja
koliformnih bakterija i E. coli, uz inkubaciju na 37 °C kroz 24 sata. Koliformne bakterije
metaboliziraju hranjivi indikator i uzorak postaje ţut, a E. coli metabolizira drugi hranjivi
indikator tako da uzorak pod UV svjetlom postaje fluorescentan. Crijevni enterokoki su
odreĎivani metodom membranske filtracije pomoću selektivne mikrobiološke podloge Slanetz
Bartley Agar (Biolife) koji sadrţi 2,3,5-trifeniltetrazolium klorida (TTC). Inkubacija je
izvršena na 37 °C kroz 48 sati. Za potvrĎivanje enterokoka korištene su podloge Esculin Iron
Agar (Biolife), Triple Sugar Iron Agar i katalaza test. Za utvrĎivanje Pseudomonas
aeruginosa korištena je metoda membranske filtracije i mikrobiološka podloga Pseudomonas
Selective Agar (Biolife), inkubacija je izvršena na 37 °C kroz 24 sata, a za potvrĎivanje je
korišten Tryple Sugar Iron Agar, oksidaza i katalaza test, DEV Tryptophan Broth u kojem se
utvrĎivao nastanak piocijanina dodatkom kloroforma, te Simmons Citrate Agar. Za
odreĎivanje Salmonella spp. nakon umnaţanja bakterija koje su zadrţane na sterilnom
membranskom filtru u puferiranoj peptonskoj vodi korištene su podloge Salmonella Shigela
Agar (Biolife), Rappaport Vassiliadis Soy Broth (Biolife), Xylose-Lysine-Deoxycholate
(XLD) Agar (Biolife), Triple Sugar Iron Agar, Lisine Iron Agar (Biolife) uz biokemijsku
identifikaciju (laktoza, glukoza, urea) i serološku tipizaciju. Sulfitoreducirajuće
klostridije odreĎivane su pomoću SPS Agara (Liofilchem, Italija). Kvasci i plijesni dokazivani
su porastom na Sabouraud Dextrose Agar s kloramfenikolom (Biolife). Prisutnost Listeria
monocytogenes odreĎivana je nakon primarnog i sekundarnog obogaćivanja u bujonu Fraser
Broth (Biolife) na 30 °C kroz 24 sata, korištenjem podloga Agar Listeria Ottaviani Agosti
(Biolife), Rapid L.Mono (Bio-Rad, SAD), krvnog agara Columbia Agar Base (Biolife), a za
identifikaciju vrsta iz roda Listeria API Listeria (bioMérieux, SAD).
Biološki materijal na kojem su izvršena mikrobiološka ispitivanja bili su jetra, slezena, škrge i
bubreg riba sa kojih je izvršena inokulacija na Tryptone Soya Agar (Oxoid), Blood Agar
(Certifikat d.o.o., Hrvatska) i Mac Conkey Agar (Oxoid). Nakon inkubacije na 22 °C kroz 48-
72 sata izolirane su reprezentativne kolonije kolonije i reinkubirane na svjeţim podlogama.
Čiste kolonije bojane su po Gramu i utvrĎivana su njihova morfološka, fiziološka i

34
biokemijska svojstva. Izolati su podvrgnuti su standardiziranom identifikacijskom sustavu za
Enterobacteriaceae i druge Gram-negativne štapiće, API 20E (bioMérieux, Francuska) i
Bruker Biotyper MALDI-TOF MS analizama (Bruker Daltonics, Billerica, SAD).
Mikroepruvetice API 20E traka sa dehidriranim supstratima inokulirane su sa bakterijskom
suspenzijom koja je reagirala sa medijem. Tijekom inkubacije metabolizam je proizvodio
promjene u boji koje su bile spontane ili potaknute dodatkom reagenasa. Reakcije su
očitavane prema tablici uz upute, a identifikacija je dobivena prema Indeksu analitičkog
profila putem identifikacijskog programa (apiweb, bioMérieux). API 20E testovi provedeni su
u skladu s uputama proizvoĎača uz nekoliko prilagodbi za bakteriološka ispitivanja riba:
vrijeme inkubacije povećano je na 48-72 sata uz temperaturu od 22 °C, te je u ugljikohidratne
mikroepruvetice dodano sterilno mineralno ulje (Topić Popović i sur., 2007).
3.6.1. Maldi TOF MS
Bakterijski izolati (jedna eza bakterijske kulture) naneseni su kao tanki film na čelične ploče
(Bruker Daltonics) sa 24 mjesta, u dvije replike, te osušeni na sobnoj temperaturi. Zatim je na
koloniju naneseno 2 µL MALDI matrice (zasićena otopina α-cijano-4-hidroksicinamične
kiseline u 50 % acetonitrila i 2,5 % trifluoroctene kiseline) i osušeno u digestoru. Ioni su
generirani sa 337–nm dušikovim laserom i hvatani u pozitivan linearni mod u masenom
rasponu od 2 do 20 kDa. Pozitivni ioni ekstrahirani su akceleriranim naponom od 20 kV u
linearnom modu. Svaki spektar je zbroj iona dobivenih laserskim snimkama iz pet različitih
područja istog izvora. Uhvaćeni spektri analizirani su korištenjem MALDI Biotyper ureĎajem
i Bruker Biotyper 2.0 softverom (Bruker Daltonics). MALDI Biotyper baza podataka sadrţi
3 740 spektara sa 319 rodova i 1 946 vrsta mikroorganizama. Za svaku od 24 ploče bio je
uključen bakterijski standardni test za kalibraciju i validaciju instrumenta.
Kriterij za identifikaciju bio je kako slijedi: vrijednost od 2,300 do 3,000 ukazuje na visoku
sigurnost identifikacije bakterija, vrijednost od 2,000 do 2,299 ukazuje na sigurnu
identifikaciju roda sa vjerojatnom identifikacijom vrsta, vrijednost 1,700 do 1,999 ukazuje na
vjerojatnu identifikaciju roda, a vrijednost < 1,700 se smatra nepouzdanom. Nazočnost ili
odsustvo specifičnih peak-ova smatra se očitanjem za svaki izolat. Identifikacija izolata
korespondira sa vrstama referentnog soja koji se najbolje podudara sa bazom podataka.

35
3.6.2. OdreĊivanje osjetljivosti izolata bakterija prema antimikrobnim
lijekovima
Osjetljivost izolata bakterija prema antimikrobnim lijekovima odreĎivana je pomoću
kvalitativne Kirby-Bauer disk difuzijske metode na Mueller Hinton agaru (Oxoid). Na
površinu hranjivog agara inokuliranog bakterijskom suspenzijom postavljeni su diskovi
natopljeni standardiziranom količinom (µg) slijedećih antibiotika: oksitetraciklin (OTC30,
amoksicilin (AMC30), oksolinska kiselina (OA2), eritromicin (E15), sulfametoksazol (SX50),
florfenikol (FFC30), norfloksacin (NOR10) i flumekvin (UB30). Inokulum je suspendiran u 5
mL sterilnog 0,85 % medija (bioMerieux) uz MacFarland standard 0,5. Za očitavanje
promjera zone inhibicije i izraţavanje rezultata korištene su standardizirane tablice
proizvoĎača. Osjetljivost na antibiotike izraţavana je kao: osjetiljiv na antibiotik (susceptible,
S), umjereno osjetljiv (intermediate, I) i otporan na antibiotik (resistant, R).
3.7. Statistiĉka obrada rezultata
Za sve prikupljene rezultate provodila se standardna deskriptivna statistika pomoću SigmaStat
i SigmaPlot Statistical softvera verzija 11.0 (Jandel Corp., SAD). Razina statističke
značajnosti odreĎena je na p < 0,05. Normalnost raspodjela provjeravana je Shapiro-Wilk
testom, dok je statistička značajnost razlike u srednjim vrijednostima pojedinih parametara
analizirana Kruskal-Wallis testom. Analize i korelacije izmeĎu parametara krvi odreĎivane su
pomoću t-testa, Mann-Whitney Rank Sum Testa i Spearman Rank Order Correlation testa.
Povezanost pojedinih parametara sa obiljeţjima sezone analizirana je i Pearsonovim
koeficijentom korelacije. Statističke obrade obavljene su u paketima SPSS 13.0 for Windows
(SPSS Inc., SAD) i Microsoft Excel 2007. Analiza glavnih komponenti (PCA) korištena je za
opis glavnih izvora varijacija i odnosa izmeĎu varijabli (SPSS 13.0 for Windows).
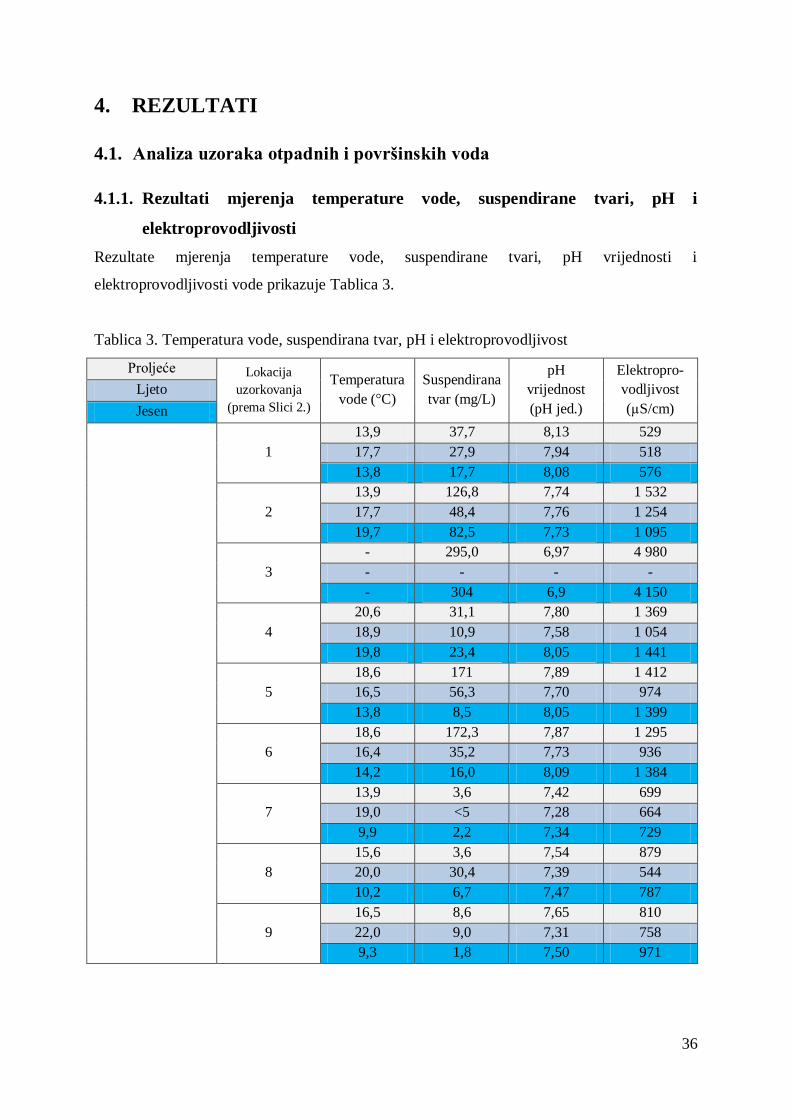
36
4. REZULTATI
4.1. Analiza uzoraka otpadnih i površinskih voda
4.1.1. Rezultati mjerenja temperature vode, suspendirane tvari, pH i
elektroprovodljivosti
Rezultate mjerenja temperature vode, suspendirane tvari, pH vrijednosti i
elektroprovodljivosti vode prikazuje Tablica 3.
Tablica 3. Temperatura vode, suspendirana tvar, pH i elektroprovodljivost
Proljeće Lokacija
uzorkovanja
(prema Slici 2.)
Temperatura
vode (°C)
Suspendirana
tvar (mg/L)
pH
vrijednost
(pH jed.)
Elektropro-
vodljivost
(µS/cm)
Ljeto
Jesen
1
13,9 37,7 8,13 529
17,7 27,9 7,94 518
13,8 17,7 8,08 576
2
13,9 126,8 7,74 1 532
17,7 48,4 7,76 1 254
19,7 82,5 7,73 1 095
3
- 295,0 6,97 4 980
- - - -
- 304 6,9 4 150
4
20,6 31,1 7,80 1 369
18,9 10,9 7,58 1 054
19,8 23,4 8,05 1 441
5
18,6 171 7,89 1 412
16,5 56,3 7,70 974
13,8 8,5 8,05 1 399
6
18,6 172,3 7,87 1 295
16,4 35,2 7,73 936
14,2 16,0 8,09 1 384
7
13,9 3,6 7,42 699
19,0 <5 7,28 664
9,9 2,2 7,34 729
8
15,6 3,6 7,54 879
20,0 30,4 7,39 544
10,2 6,7 7,47 787
9
16,5 8,6 7,65 810
22,0 9,0 7,31 758
9,3 1,8 7,50 971

37
Vrijednosti suspendirane tvari komunalnih, šećeranskih i pročišćenih otpadnih voda pokazuju
visoku učinkovitost uklanjanja suspendirane tvari iz otpadnih voda. Koncentracija
suspendirane tvari voda u pročišćenim otpadnim vodama koje se prelijevaju iz sekundarnih
taloţnica u Kanal Manteč kao njihov recipijent, sukladna je sa Pravilnikom o graničnim
vrijednostima emisije otpadnih tvari (NN 80/13, NN 43/14). Zakiseljenost tehnoloških
otpadnih voda Tvornice šećera (TŠ) veća je od zakiseljenosti komunalnih otpadnih voda, a
zbog manjeg učešća u ukupnom hidrauličkom opterećenju UreĎaja (UBP), pH vrijednost
pročišćenih otpadnih voda bila je od 7,58 do 8,05. pH vrijednost na svim mjernim postajama
prema kriterijima Uredbe o standardu kakvoće voda (NN 73/13) bila je vrlo dobra ili dobra.
4.1.2. Rezultati mjerenja otopljenog kisika, kemijske potrošnje kisika (KPKCr i
KPKMn) i biološke potrošnje kisika (BPK-5)
U potoku OĎenica, kao vodotoku za koji se drţi da nije pod utjecajem otpadnih voda, kako
pokazuje Tablica 4, prilikom svakog je mjerenja izmjerena visoka koncentracija otopljenog
kisika u vodi. Uzimanje uzorka otpadne vode koja dolazi iz grada Virovitice na UBP
izvršeno je nakon podizanja otpadne vode puţnim crpkama i nakon uklanjanja krupnih
nečistoća pomoću automatske rešetke. Do obogaćivanja otpadne vode kisikom iz zraka došlo
je na puţnim crpkama. Prema izvršenim ispitivanjima, pročišćena otpadna voda u pravilu je
sadrţavala niske vrijednosti otopljenog kisika (< 0,5 mg /L). Uklanjanje ugljika na UBP-u
bilo je visoko, a pročišćena otpadna voda bila je sukladna sa Pravilnikom o graničnim
vrijednostima emisije otpadnih tvari (NN 80/13, NN 43/14). Zajedničko pročišćavanje
komunalnih i šećeranskih otpadnih voda provodilo se u travnju za vrijeme razdoblja
proizvodnje šećera iz šećerne trske, i u rujnu kada se šećer proizvodio iz šećerne repe.
Uvaţavajući hidrauličko opterećenje komunalnih i šećeranskih otpadnih voda u terminima
provoĎenja istraţivanja, prosječni stupanj redukcije KPK vrijednosti iznosio je 93,64 % a
BPK-5 93,75 %. Tablica 4. pokazuje i izmjerene vrijednosti KPKMn i BPK-5 u uzorcima iz
potoka OĎenica i vodotoka koji su recipijent pročišćenih otpadnih voda. Vrijednosti ovih
parametara bile su najveće na prvom mjernom mjestu nakon ispuštanja pročišćenih otpadnih
voda, dok su nizvodno zbog procesa autopurifikacije ove vrijednosti bile niţe.

38
Tablica 4. Otopljeni kisik, kemijska potrošnja kisika (KPKCr i KPKMn) i biološka potrošnja
kisika (BPK-5)
Proljeće Lokacija
uzorkovanja
(prema Slici 2.)
Otopljeni kisik
(mg /L)
KPKCr KPKMn BPK-5 Ljeto
Jesen
1
9,66 43,16 15,63 19,85
8,14 42,69 15,70 19,00
8,91 - 9,18 10,75
2
5,93 369,91 69,66 310
5,28 339,32 53,00 254
4,96 174,34 - 117,45
3
- 7.536 1.678 6.153
- - - -
0,9 4.418 - 3.980
4
0,39 42,63 20,63 36,75
0,28 53,66 27,86 41,22
4,05 63,63 - 39,33
5
2,24 57,55 30,64 48,26
5,11 64,16 31,43 41,66
1,23 - 18,14 17,5
6
2,72 71,96 31,64 58,23
6,07 52,24 30,17 35,20
2,32 43,08 16,68 34,7
7
4,86 32,49 8,85 25,00
1,83 17,83 3,56 2,18
3,91 19,49 6,36 3,84
8
1,91 22,2 16,93 15,07
1,07 96,6 13,00 14,20
2,48 24,15 10,96 7,85
9
9,29 18,49 11,93 12,84
4,37 23,67 10,09 4,73
2,48 18,77 11,64 7,60
4.1.3. Rezultati mjerenja amonijaka, nitrita, nitrata i ukupnog dušika
UreĎaj za biološko pročišćavanje (UBP) nije tehnološki izveden za tercijarno pročišćavanje
otpadnih voda. Do uklanjanja dušika iz otpadnih voda ipak dolazi, ali ne u dovoljnoj mjeri da
bi se njegova vrijednost dovela ispod 10 mg/L kako je propisano Pravilnikom o graničnim
vrijednostima emisije otpadnih tvari (NN 80/13, NN 43/14). Na Slici 3. prikazane su
vrijednosti dušikovih spojeva za sve ispitane uzorke. Najniţe vrijednosti izmjerene su u
uzorcima iz potoka OĎenica.

39
Slika 4. Koncentracija amonijaka, nitrita, nitrata i ukupnog dušika u pročišćenoj otpadnoj vodi
i vodotocima (Slika 2., Lokacije uzorkovanja 1-9)
0
10
20
30
40
50
60
70
1 2 3 4 5 6 7 8 9
mgN
H4
+-N
/L
Amonijak
proljeće ljeto jesen
0
0,5
1
1,5
2
2,5
1 2 3 4 5 6 7 8 9
mgN
O2- -
N/L
Nitriti
proljeće ljeto jesen
0
5
10
15
20
25
30
1 2 3 4 5 6 7 8 9
mg
NO
3- -
N/L
Nitrati
proljeće ljeto jesen
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9
mgN
/L
Ukupni dušik
proljeće ljeto jesen

40
Usporedbom vrijednosti dušikovih spojeva u uzorcima otpadnih voda koje dolaze na ureĎaj i
pročišćenih otpadnih voda moţe se pratiti u kojoj mjeri se mijenjaju koncentracije amonijaka,
nitrita, nitrata i ukupnog dušika i utvrditi stupanj procesa nitrifikacije i denitrifikacije.
4.1.4. Rezultati ispitivanja ortofosfata i ukupnog fosfora
Temeljem Uredbe o standardu kakvoće voda (NN 73/13), potok OĎenica (Lokacija
uzorkovanja 1) po sadrţaju ortofosfata i ukupnog fosfora bio je u kategoriji vrlo dobrog i
dobrog ekološkog stanja. Koncentracija fosfora u izlaznom toku s UBP-a u ljeti i jesen nije
bila u potpunosti sukladna s vaţećim zakonskim propisima i vodopravnoj dozvoli
(maksimalna dopuštena koncentracija ukupnog fosfora u pročišćenim otpadnim vodama = 1
mg/L). U uzorcima vode iz Ţupanijskog kanala (Slika 2., lokacija uzorkovanja 9), najviše
vrijednosti fosfora izmjerene su u jesenskom terminu uzorkovanja (Slika 5.).
Slika 5. Koncentracija ortofosfata i ukupnog fosfora u pročišćenoj otpadnoj vodi i vodotocima
(Slika 2., Lokacije uzorkovanja 1-9)
0
1
2
3
4
5
6
7
1 2 3 4 5 6 7 8 9
mg
PO
43
- -P
/L
Ortofosfati
proljeće ljeto jesen
0
1
2
3
4
5
6
7
8
1 2 3 4 5 6 7 8 9
mgP
/L
Ukupni fosfor
proljeće ljeto jesen

41
4.1.5. Rezultati mjerenja teških metala
Prema rezultatima koje prikazuje Tablica 5, najveći sadrţaj ţeljeza od 23 675 µg/L izmjeren
je u šećeranskoj otpadnoj vodi u vrijeme kampanje prerade šećerne repe. Visoka vrijednost od
8 388 µg/L utvrĎena je u uzorku potoka OĎenica u proljeće. Naniţe koncentracije ţeljeza
izmjerene su u pročišćenoj otpadnoj vodi. Najviše koncentracije kadmija i bakra izmjerene su
u kanalu Manteč nakon ispuštanja pročišćenih otpadnih voda (Slika 2., lokacije uzorkovanja 5
i 6). Najviša vrijednost kroma izmjerena je u šećeranskoj otpadnoj vodi u proljeće, dok su
prosječne vrijednosti kroma za svaku mjernu postaju komparabilne. Najveći sadrţaj ţive i
olova izmjeren je u komunalnoj otpadnih vodi koja dolazi na UreĎaj, pročišćenoj otpadnoj
vodi i šećeranskoj otpadnoj vodi, a s povećanjem udaljenosti od ureĎaja koncentracija ţive se
smanjuje. U šećeranskim otpadnim vodama odreĎena je najviša koncentracija nikla.
Tablica 5. Koncentracija ţeljeza, kadmija, kroma, ţive, olova, nikla, cinka i bakra u uzorcima
voda (μg/L)
Proljeće Lokacija uzorkovanja
(prema Slici 2.) Fe Cd Cr Hg Pb Ni Zn Cu Ljeto
Jesen
1
8 388 0,06 6,08 <0,20 1,86 10,00 23,00 56,00
984 0,33 7,53 2,99 4,56 6,00 43,00 34,00
750 0,16 4,03 0,94 5,86 4,65 2,58 7,75
2
30 0,10 14,87 6,25 4,27 6,00 128,0 90,00
984 <0,20 6,84 3,25 0,58 3,00 70,00 35,00
558 0,28 1,10 0,31 1,31 1,96 28,59 13,30
3
9 197 0,24 17,56 4,23 3,17 76,00 46,00 43,00
- - - - - - - -
23 675 0,41 5,86 <0,20 6,84 <1,00 18,59 18,50
4
50 0,06 14,81 0,50 1,41 14,00 44,00 46,00
155 <0,20 2,27 5,79 0,44 4,00 26,00 60,00
32 0,14 1,12 <0,20 1,46 <1,00 <1,00 4,90
5
703 0,80 7,70 0,87 5,58 31,00 90,00 270,0
215 <0,20 <1,00 1,74 0,48 4,00 28,00 27,00
140 <0,10 <1,00 <0,20 1,97 2,94 7,52 5,13
6
419 1,62 12,77 1,21 5,83 38,00 101,00 274,00
390 <0,20 5,37 <0,20 0,58 4,00 26,00 21,00
370 0,14 1,54 <0,20 2,16 <1,00 5,50 6,10
7
207 0,03 7,84 <0,20 2,98 19,00 37,00 80,00
440 <0,20 7,10 <0,20 0,83 3,00 32,00 21,00
472 0,34 <1,00 <0,20 2,46 <1,00 <1 3,40
8
138 0,03 11,14 0,83 1,95 10,00 29,00 81,00
1 710 <0,20 5,15 1,62 0,33 4,00 27,00 21,00
764 0,30 <1,00 <0,20 2,38 <1,00 16,15 4,14
9
128 0,05 12,91 0,45 2,14 8,00 27,00 61,00
490 <0,20 1,30 <0,20 0,53 2,00 30,00 20,00
464 0,177 2,72 <0,20 2,38 <1,00 16,15 4,14

42
Vrijednosti teških metala u muljevima (Slika 6.) u svim su istraţivanim uzorcima bile niske,
osim na jesen. Koncentracija teških metala u svim je ispitivanim terminima bila značajno veća
u aktivnom mulju UBP-a nego u mulju na deponiji tvornice šećera. U aktivnom mulju su se,
prema izmjerenim vrijednostima, isticale visoke vrijednosti cinka, olova i bakra, posebice u
uzorcima uzetima na jesen.
Slika 6. Koncentracija teških metala u muljevima
4.1.6. Rezultati mjerenja farmaceutika
Prema Tablica 6., prosječni udio mjerenih antibiotika u ukupnoj koncentraciji odreĎivanih
farmaceutika u otpadnim vodama koje dolaze na UreĎaj bio je 13,62 %, a u pročišćenim
otpadnim vodama 27,45 %. Srednja vrijednost antibiotika u komunalnim otpadnim vodama
iznosila je 15 367,45 ng/L s udjelom sulfa lijekova od 0,049 % (76,05 ng/L), dok je u
pročišćenim otpadnim vodama srednja vrijednost antibiotika bila 7 715,39 ng/L a udio sulfa
lijekova 0,839 % (64,76 ng/L). U komunalnim otpadnim vodama koje dolaze na UreĎaj
proljeće ljeto jesen proljeće ljeto jesen
Aktivni mulj s ureĎaja Mulj s deponije
Cd 1,39 0,31 5,52 0,05 0,15 0,27
Cr 36,13 3,43 131,00 4,06 8,21 4,74
Hg 2,42 3,12 0,00 0,25 0,23 1,98
Pb 15,93 0,00 810,00 4,00 6,64 5,62
Ni 100,00 9,00 0,00 2,25 10,69 1,57
Zn 466,00 418,00 1685,00 32,25 36,93 77,17
Cu 336,00 149,00 590,00 18,22 17,63 13,80
0,00
200,00
400,00
600,00
800,00
1000,00
1200,00
1400,00
1600,00
1800,00
µg
/L
Cd
Cr
Hg
Pb
Ni
Zn
Cu

43
koncentracija antibiotika od jutra prema večeri povećala se za gotovo pet puta, dok je bila
konstantna u pročišćenim vodama. Koncentracija sulfa lijekova od jutra do večeri se
prepolovila, dok je bila stalna u pročišćenim otpadnim vodama. Prosječne vrijednosti
sulfametoksazola bile su komparabilne za svaku lokaciju uzorkovanja, osim večeri. Prosječna
vrijednost eritromicina u komunalnim otpadnim vodama bila je 19,57 ng/L, dok su vrijednosti
eritomicina u pročišćenim otpadnim vodama bile ispod granice detekcije. Koncentracija
ukupnih farmaceutika, antibiotika i sulfametoksazola u sedimentu se smanjivala s
udaljavanjem od UBP-a, dok eritromicin nije utvrĎen niti na jednoj lokaciji uzorkovanja
sedimenta.
Tablica 6. Koncentracija odreĎivanih farmaceutika u vodi (ng/L) i sedimentu (ng/g)
Voda
Ulaz komunalnih otpadnih voda Izlaz pročišćenih otpadnih voda
Jutro Podne Večer Jutro Pola dana Večer
Farmaceutici 112 333,45 107 688,88 118 557,90 21 085,90 35 383,55 27 830,73
Antibiotici 5 479,82 14 287,57 26 334,96 7 545,84 7 949,07 7 651,23
Sulfa lijekovi 90,88 87,88 49,39 65,71 60,12 68,44
Sulfametoksazol 57,74 67,81 26,55 51,68 51,02 53,14
Eritromicin 2,27 43,94 12,5 0 0 0
Sediment
Prijemnik pročišćenih otpadnih voda
Kanal Manteč Ţupanijski kanal
Farmaceutici 158 789,49 33 446,11 28 210,90
Antibiotici 144,54 17,51 18,58
Sulfametoksazol 19,6 5,07 0,99
Eritromicin 0 0 0

44
4.2. Ţivotinjske vrste i obrada tkiva
4.2.1. Ribe
4.2.1.1. Zdravstveni status riba
Općim kliničkim pregledom i patološko-anatomskom pretragom riba uhvaćenih tijekom
različitih godišnjih doba na tri reprezentativna mjesta uzorkovanja nisu utvrĎeni znakovi
bolesti. Na škrgama babuški iza prijemnika pročišćenih voda (kroz sva godišnja doba)
ustanovljene su hemoragične promjene uz opseţno izlučivanje sluzi i edem. Mikroskopskim
pregledom na nativnim preparatima škrga, koţe i peraja kod riba ulovljenih u potoku OĎenica
(Slika 2., Lokacija uzorkovanja 1) u proljeće je ustanovljena slaba invadiranost ektoparazitom
Dactylogyrus spp. Na ribama ulovljenima u Ţupanijskom kanalu na utoku kanala Manteč
(Slika 2. Lokacija uzorkovanja 8) u jesenskom razdoblju utvrĎeni su ektoparaziti
Dactylogyrus spp. (na jednoj ribi) i Gyrodactylus spp. (na dvije ribe). Najjača invadiranost
ektoparazitom Dactylogyrus spp. uz oštećenja na koţi (oštećenja ljuski, krvarenja na perajama
i repu), aneurizme na škrgama i telohaneloza po svim perajama utvrĎena je kod riba
ulovljenih u Ţupanijskom kanalu kod naselja Brezovica (Slika 2., lokacija uzorkovanja 9).
4.2.1.2. Histologija škrga, bubrega, jetre i slezene
Histopatološki nalaz riba iz efluenta otkriva strukturne i funkcionalne promjene na tkivu
škrga, ponajviše na sekundarnim lamelama Na tkivu škrga babuški ulovljenih u proljeće, ljeto
i jesen, nalaze se promjene u smislu sljepljivanja vršaka susjednih sekundarnih lamela u
potoku OĎenica, do sljepljivanja lamela i tvorbi „mosta“ koji premošćuje interlamelani
prostor (kanal Manteč i Ţupanijski kanal). Lamelarna fuzija primijećena je u proljeće. Ljeti su
se isticale atrofične i nekrotične lezije, dok je u jesen utvrĎena hipertrofija epitelnih i
interlamelarnih stanica. Ribe ulovljene u Ţupanijskom kanalu imale su izrazitu hiperplaziju i
hipertrofiju epitelnih i interlamelarnih stanica.

45
Slika 7. Škrge babuški ulovljenih u proljeće (a, b, c), ljeto (d, e, f) i jesen (g, h, i). Skala: a, b,
c: 0,5 mm; d, e, f: 0,2 mm; g, h, i: 0,1 mm. (a) Sljepljivanje vršaka nekoliko susjednih
sekundarnih lamela (potok OĎenica). (b) Sljepljivanje lamela; lateralni rubovi spojenih lamela
u nizu tvore “most” koji premošćuje interlamelani prostor (kanal Manteč). (c) Kovrčanje
lamela i meĎulamelarno premošćivanje (Ţupanijski kanal). (d) Lamelarni crteţ s pravilno
rasporeĎenim mukoznim stanicama u meĎulamelarnom prostoru (potok OĎenica). e)
Razdvajanje sekundarnih lamela praćeno hemoragičnim promjenama, oštećenjem kapilara i
hipersekrecijom mukoznih stanica (kanal Manteč); skraćenje sekundarnih lamela, atrofične i
nekrotične promjene stanica u interlamelarnom prostoru i potpornom hrskavičnom tkivu. (f)
Hipertrofija i hiperplazija stanica u interlamelarnom prostoru, skraćivanje/poduljivanje
sekundarnih lamela; hiperplazija mukoznih stanica (Ţupanijski kanal). (g) Pravilna histološka
slika škrga (potok OĎenica). (h) Hipertrofija epitelnih i interlamelarnih stanica, nekroza i jaka
infiltracija limfocitima; interlamelarni edem, odvajanje od primarne lamele (kanal Manteč).
(i) Hiperplazija i hipertrofija mukoznih stanica (Ţupanijski kanal).
Oštećenja na bubrezima odnose se na promjene nefrona i tubularnom sistemu. Različiti stadiji
atrofije i nekroze glomerula zabiljeţeni su tijekom sezona, a najveće promjene su bile

46
izraţene u ljeto i jesen (Slika 8.). U ljetnim mjesecima u kanalu Manteč, histološka slika
bubrega babuški pokazuje peritubularnu dilataciju i povećan broj malih intenzivno bazofilnih
nakupina stanica u blizini tubula, kao i povećanu infiltraciju makrofaga izmeĎu bubreţnih
kanalića. U Ţupanijskom kanalu tijekom ljetne sezone primijećeni su tipični multifokalni
granulomi, a vidljiva je tubularna degeneracija i znakovi smanjena lumena kanalića. U jesen
kod babuške iz Ţupanijskog kanala uočena je teška nekroza bubreţnog epitela, edem
bubreţnih tubula i prisutnost velikih eozinofilnih granula unutar stanica, te različiti stupnjevi
atrofije i nekroze glomerula.
Slika 8. Tkivo bubrega babuški ulovljenih u ljeto (a, b) i jesen (c, d) u kanalu Manteč (a,c) i
Ţupanijskom kanalu (b, d). Skala: 0,1 mm. (a) Peritubularna dilatacija (široka strelica),
bazofilne nakupine stanica (pune strelice), infiltracije makrofaga (srcolika strelica). (b)
Multifokalni granulomi (široke strelice), tubularna degeneracija (srcolika strelica), oštećenje
glomerula (puna strelica). (c) bazofilni nefroni (puna strelica), suţeni lumen nefrona (srcolika
strelica). (d) nekroza epitela (široke strelice), edem tubula (dugačke strelice), eozinofilne
granule (pune strelice), atrofija i nekroza glomerula (puna strelica).

47
Tkivo jetre babuški iz efluenta bilo je zahvaćeno patološkim promjenama tijekom ljeta i u
jesen, u obliku kariolize, binuklearnih hepatocita i distenzije sinusoida (Slika 9.). Promjene su
uočljive u riba iz kanala Manteč, kao i nizvodno, u riba iz Ţupanijskog kanala, pogotovo u
smislu nekroza i upalnih promjena.
Slika 9. Tkivo jetre babuški ulovljenih u ljeto (a, b) i jesen (c, d), u kanalu Manteč (a,c) i
Ţupanijskom kanalu (b, d). Skala: 0,1 mm. (a) Hepatociti gusto zbijeni, nejasnih obrisa;
gubitak staničnih membrana (srcolika strelica), karioliza (puna strelica), binuklearni hepatociti
(široka strelica). (b) Različiti stadiji apoptoze hepatocita; promjene na jezgrama u vidu
promjera i gustoće kromatina (srcolika strelica); ţarišna nekroza jetre (puna strelica). (c)
Vakuolarno proširenje sinusoida (d) Upalna leukocitna infiltracija (puna strelica); krvne
stanice izmeĎu hepatocita (srcolika strelica); anizokarioza i piknotične jezgre (široka strelica).
Histopatološke promjene na slezeni zapaţene su tijekom proljeća, u vidu intracelularnih
vakuola i granulomatoznih lezija kod riba iz efluenta i Ţupanijskog kanala (Slika 10.).

48
Slika 10. Tkivo slezene babuški ulovljenih u proljeće , u kanalu Manteč (a,b) i Ţupanijskom
kanalu (c). Skala: a, b: 0,05 mm; c: 0,1 mm. (a) Pravilna histološka struktura; ravnomjerna
raspodjela i dobra organiziranost melanomakrofagnih centara (MMC) (puna strelica) i
proširene sinusoide (široka strelica). (b) Intracelularne vakuole. (c) Granulomatozne lezije
(srcolika strelica) i smanjenje broja i veličina MMC (široka strelica).
4.2.1.3. Hematologija, biokemijski parametri u krvi riba
Vrijednosti GGT, GSH-Px i PON 1 u većini uzoraka nisu bile mjerljive, vjerojatno radi
hemolize, pa stoga nisu prikazane. Vrijednosti Hct (Tablica 7.) uglavnom nisu pokazivale
statistički značajne razlike izmeĎu skupina. Spearman Rank Order Correlation test pokazao
je kako varijable Hct za ribe iz Ţupanijskog kanala u ljeti i jesen imaju tendenciju pada, dok
ostale rastu (negativni koeficijent korelacije i p < 0,05).
Usporedba vrijednosti URE, CRE, TP, ALB, GLU, ALT, ALP, TRIG, CHOL i SOD babuški
kroz sezone prikazana je na Slici 11. One Way Analysis of Variance test pokazao je
signifikantnost (p < 0,05) za CHOL u plazmi riba iz Manteča u odnosu na ribe iz Ţupanijskog
kanala u jesen, te CHOL u plazmi riba iz OĎenice u ljeti u odnosu na ribe iz Ţupanijskog
kanala u jesen. Signifikantne razlike uočene su i za CRE i TRIG u plazmi riba iz OĎenice
(proljeće/jesen), za CRE u riba iz OĎenice (proljeće) u odnosu na Manteč (jesen). Isto tako,
TRIG je pokazivao signifikantnost u plazmi riba iz Manteča (proljeće/jesen). U proljeće je
zamijećen signifikantan rast vrijednosti URE, TP, ALB i TRIG u riba iz Manteča u usporedbi
sa svim ostalim babuškama, kao i signifikantan pad vrijednosti SOD u usporedbi sa
babuškama iz ostala dva kanala.
Spearman Rank Order Correlation test pokazao je kako parovi varijabli sa p vrijednostima
ispod 0,05 i pozitivnim koeficijentom korelacije, poput SOD u riba iz Ţupanijskog kanala

49
(proljeće/ljeto) imaju tendenciju da se zajedno povećavaju. Za parove varijabli sa negativnim
koeficijentom korelacije i sa p < 0,05, jedna varijabla ima tendenciju rasta, dok sve druge
padaju, poput: URE u riba iz Ţupanijskog kanala (proljeće/jesen), TRIG u riba iz Ţupanijskog
kanala (proljeće/jesen), ALT u riba iz Ţupanijskog kanala (proljeće)/OĎenice (jesen), SOD u
riba iz Ţupanijskog kanala (ljeto/jesen), Hct u riba iz Ţupanijskog kanala (proljeće/jesen).
Tablica 7. Vrijednosti hematokrita krvi babuške kroz sezone
Lokacija uzorkovanja
Proljeće
Ljeto Jesen
OĎenica (Lokacija uzorkovanja 1
prema Slici 2.) 33,60 ± 14,52 29,00 ± 13,21 36,06 ± 11,48
Manteč (Lokacija uzorkovanja 8 prema
Slici 2.) 42,00 ± 8,37 27,20 ± 18,36 27,86 ± 3,85(*)
Ţupanijski kanal nizvodno (Lokacija
uzorkovanja 9 prema Slici 2.) 47,14 ± 7,26 37,56 ± 10,08 42,44 ± 6,38(*)
Vrijednosti su prikazane kao srednja vrijednost ± standardna devijacija (SD); vrijednosti
označene sa (*) su signifikantne (p < 0,05).

50
Slika 11. Biokemijski parametri i parametri oksidacijskog stresa; (log10 osnovne skale) u
plazmi babuški po sezonama prema mjestu ulova (Slika 2., lokacije uzorkovanja 1, 8, 9) URE
(urea), CRE (kreatinin), TP (ukupne bjelanjčevine), ALB (albumin), GLU (glukoza), ALT
(alanin aminotransferaza), ALP (alkalna fosfataza), TRIG (trigliceridi), CHOL (kolesterol),
SOD (aktivnost superoksid dismutaze).
-2,0
-1,0
0,0
1,0
2,0
3,0
4,0
5,0
6,0
URE CRE TP ALB GLU ALT ALP TRIG CHOL SOD
log
10 o
snovvn
e s
kale
Proljeće
1.
8.
9.
-2,0
-1,0
0,0
1,0
2,0
3,0
4,0
5,0
6,0
URE CRE TP ALB GLU ALT ALP TRIG CHOL SOD
log
10 o
snovn
e s
kale
Ljeto
1.
8.
9.
-2,0
-1,0
0,0
1,0
2,0
3,0
4,0
5,0
URE CRE TP ALB GLU ALT ALP TRIG CHOL SOD
log
10 o
snovn
e s
ka
le
Jesen
1.
8.
9.
c
a
b
b
d
c c
a
a
d d
a
b
a

51
Stupci pogreške predstavljaju standardnu devijaciju srednje vrijednosti. Razlike u
vrijednostima parametara označenih istim malim slovima (a, b, c, d) statistički su značajne (p
< 0,05).: a-prosječna vrijednost TRIG-a u proljeće bila je u krvi riba iz Ţupanijskog kanala
nizvodno 3,3 mmol/L, a u jesen 4,1 mmol/L; b-prosječna vrijednost SOD-a na istoj lokaciji
bila je u proljeće 502,5 U/L, u ljeto 505,3 U/L, a u jesen 299,8 U/L; c – prosječna vrijednost
CRE u potoku OĎenica bila je u proljeće 50 µmol/L, u jesen 30,3 µmol/L a u Ţupanijskom
kanalu nizvodno u jesen 34,4 µmol/L; d- prosječna vrijednost CHOL u potoku OĎenica u ljeto
iznosila je 3,5 mmol/L, u jesen 4,5 mmol/L, a u jesen u Ţupanijskom kanalu nizvodno 9,1
mmol/L.
4.2.1.4. Procjena genotoksiĉnosti (ENA i MN test)
Statistička analiza učestalosti pojave eritrocita sa nepravilnim jezgrama (ENA) pokazala je
najviše srednje vrijednosti kod riba iz Manteča u proljeće (13,19 ‰) i ljeto (12,77 ‰). Znatno
niţe vrijednosti zabiljeţene su u OĎenici (1,74-2,39 ‰) dok se prosječna učestalost ENA kod
babuški iz Ţupanijskog kanala kretala od 5,64 u proljeće do 7,57 ‰ u jesen. MeĎutim
statistički značajna razlika (p < 0,05) utvrĎena je samo u jesen izmeĎu riba iz Ţupanijskog
kanala i Manteča (Slika ). Učestalost pojave mikronukleusa (MN) bila je niska te su se srednje
vrijednosti kretale od 0,04 do 0,16 ‰.
Slika 12. Učestalost eritrocita sa nepravilnim jezgrama (‰) kod babuške; ab (p < 0,05)
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
Proljeće Ljeto Jesen
Ođenica
Manteč
Kanal
a
b

52
4.2.2. Gujavice
4.2.2.1. Gujavice: Test kontakta na filtar papiru
Prilikom izlaganja gujavica otpadnoj vodi pomoću filtar papir kontakt testa došlo je do
statistički značajne akumulacije fluorescentnog supstrata R123 u tkivima, što upućuje na
pristutnost MXR inhibitora u svim ispitivanim uzorcima. Razlika u jačini inhibicije ABC
transportera razlikovala se ovisno o sezoni uzorkovanja (Slika 14., Slika 15., Slika16.).
Najveća akumulacija R123 zabiljeţena je prilikom izlaganja gujavica otpadnoj vodi
uzorkovanoj u proljeće, posebice nakon izlaganja uzorku broj 1 (OĎenica) razrijeĎenom 100 x
(pribliţno 2,7 puta povećana u odnosu na kontrolne vrijednosti) te uzorku broj 10 (aktivni
mulj) razrijeĎenom 10 000 x (pribliţno 2,9 puta povećana u odnosu na kontrolne vrijednosti).
Vrijednosti akumulacije R123 u tkivu gujavica izlaganih otpadnoj vodi uzorkovanoj u ljeto
bile su nešto niţe, no i dalje statistički značajne. U ljetnim mjesecima najveći inhibitorni
potencijal uzrokovao je uzorak broj 1 razrijeĎen 100 x (akumulacija R123 pribliţno 2,2 puta
povećana u odnosu na kontrolne vrijednosti) te uzorak broj 4 razrijeĎen 100 x (pribliţno 1,7
puta povećana u odnosu na kontrolne vrijednosti). Jesensko uzorkovanje ukazalo je na najveći
inhibitorni potencijal uzoraka broj 1 i 9 razrijeĎenih 100 x (akumulacija R123 2,6 te 1,8 puta
povećana u odnosu na kontrolne vrijednosti, pojedinačno).

53
Slika 13. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h tretmana izlaganja otpadnoj vodi uzorkovanoj u proljeće 2014. god. Isprekidana
linija označava kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤
0,05.
Slika 14. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24h tretmana izlaganja otpadnoj vodi uzorkovanoj u ljeto 2014.god. Isprekidana linija
označava kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD,*p ≤ 0,05.

54
Slika 15. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h tretmana izlaganja otpadnoj vodi uzorkovanoj u jesen 2014. god. Isprekidana
linija označava negativnu kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ±
SD, *p ≤ 0,05.
4.2.2.2. Mjerenje lipidne peroksidacije u gujavicama nakon izlaganja otpadnim vodama
i otpadnom mulju
Izlaganje gujavica uzorcima otpadnih voda ukazalo je na statistički značajno povećanje
TBARS koncentracije u tkivima gujavica. Najveća TBARS koncentracija zabiljeţena je
nakon izlaganja gujavica 100 x razrijeĎenim uzorcima otpadne vode (Slika 16., Slika 17.,
Slika 18.). Navedeno razrijeĎenje uzrokovalo je najveće povećanje TBARS koncentracije
prilikom izlaganja gujavica uzorcima 2 i 3 prikupljenim u proljeće 2014 (pribliţno 1,6,
odnosno 1,5 puta u odnosu na kontrolne vrijednosti). TBARS vrijednosti u ostalim mjesecima
bile su nešto niţe, no još uvijek statistički značajne. Najveća TBARS koncentracija u
proljetnim uzorcima izmjerena je nakon izlaganja gujavica 100 x razrijeĎenom uzorku broj 1
(pribliţno 1,3 puta povećana) te uzorku broj 2 (pribliţno 1,5 puta povećana). U jesen, najveće
povećanje koncentracije iznosilo je pribliţno 1,4 puta u odnosu na kontrolu, te je zabiljeţeno
u uzorcima 2 i 4 (UBP-ulaz i UBP-izlaz).

55
Slika 16. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u proljeće 2014. g. Simboli:
' = 24-h izlaganje, '' = 48-h izlaganje, ''' = 7-d izlaganje. Isprekidana linija označava kontrolnu
vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤ 0,05.

56
Slika 17. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u ljeto 2014. g. Simboli: ' =
24-h izlaganje, '' = 48-h izlaganje, ''' = 7-d izlaganje. Isprekidana linija označava kontrolnu
vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤ 0,05.
Slika 183. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u jesen 2014. g. Simboli: ' =
24-h izlaganje, '' = 48-h izlaganje, ''' = 7-d izlaganje. Isprekidana linija označava kontrolnu
vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤ 0,05.
4.2.2.3. Razina inhibicije MXR proteina uslijed izlaganja otpadnom mulju
Porast akumulacije R123 zabiljeţen je prilikom izlaganja objema koncentracijama
dehidriranog aktivnog mulja (Slika 19., Slika 20., Slika 21.). Najveća koncentracija R123
izmjerena je u tkivima gujavica izlaganima 24 h većoj koncentraciji aktivnog mulja, čiji je
porast iznosio priblţno 2,2; 1,1; 1,8 u odnosu na kontrolne vrijednosti, ovisno o periodu
uzorkovanja (proljeće; ljeto; jesen). 24-satno izlaganje gujavica manjoj koncentraciji aktivnog
mulja (30 %) rezultiralo je takoĎer statistički značajnim porastom R123 akumulacije u
tkivima, i to pribliţno 0,9; 0,8; 1,0 puta većim u odnosu na kontrolne vrijednosti izmjerene u

57
proljeće, ljeto, jesen. Sedmi dan pokusa koncentracija fluorescentnog supstrata u tkivima
gujavica bila je slična kontrolnim vrijednostima, stoga statistički nije značajna.
Slika 19. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja uzorkovanog u
proljeće. Isprekidana linija označava negativnu kontrolnu vrijednost, dok puna linija označava
pozitivnu (MK571) kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤
0,05.
Slika 20. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja uzorkovanog u ljeto.

58
Isprekidana linija označava negativnu kontrolnu vrijednost, dok puna linija označava
pozitivnu (MK571) kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤
0,05.
Slika 21. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja uzorkovanog u
jesen. Isprekidana linija označava negativnu kontrolnu vrijednost, dok puna linija označava
pozitivnu (MK571) kontrolnu vrijednost. Podaci su izraţeni kao srednja vrijednost ± SD, *p ≤
0,05.
4.2.2.4. Razina oksidacijskog stresa uslijed izlaganja otpadnom mulju
Tijekom izlaganja gujavica E. fetida dvjema koncentracijama dehidriranog aktivnog mulja,
koncentracije u gujavicama izlaganima niţoj koncentraciji (30 %) bile su više nego li je to bio
slučaj prilikom izlaganja većoj koncentraciji (70 %). Nakon samo 24 h izlaganja niţoj
koncentraciji došlo je do statistički značajnog porasta TBARS-a u tkivu gujavice. Najveće
TBARS koncentracije zabiljeţene su nakon 48 h izlaganja manjoj koncentraciji aktivnog
mulja, te su pojedinačno prema redu uzorkovanja iznosile povećanje od pribliţno 1,9 ( Slika
22., Slika 23., Slika 24.) puta u odnosu na kontrolne vrijednosti. Trend smanjivanja TBARS
vrijednosti u gujavicama 7. dan trajanja pokusa zabiljeţen je u svim ispitanim uzorcima.
Zanimljivo, tijekom testiranja veće koncentracije (70 %) dehidriranog aktivnog mulja, niti u
jednom od uzoraka nije zabiljeţen statistički značajan porast TBARS-a.

59
Slika 22. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u proljeće. Isprekidana linija označava
negativnu kontrolnu vrijednost.
Slika 23. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u ljeto. Isprekidana linija označava
negativnu kontrolnu vrijednost.

60
Slika 24.. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u jesen. Isprekidana linija označava
negativnu kontrolnu vrijednost.
4.3. AMES test
Iz rezultata koje prikazuju Tablica 8, Tablica 9, Tablica 10 moţe se vidjeti da je laboratorijski
postupak ispravan: na pločama bez bakterija nije bilo spontanih revertanata koji bi ukazivali
na onečišćenje; broj spontanih revertanata bio je unutar očekivanih vrijednosti za TA 98 i TA
100 soj; izravni i neizravni mutageni spojevi uzrokovali su očekivano visoki broj revertanata.
Na temelju dobivenih rezultata utvrĎeno je da u svim testiranim uzorcima otpadnih voda ne
dolazi do pojave dvostruko većeg broja revertanata te da jednako tako rezultati ne ukazuju na
prisustvo premutagenih i/ili mutagenih tvari, prema meĎunarodno prihvaćenim kriterijima za
bakterijski Ames test.

61
Tablica 8. Rezultati analize mutagenog potencijala uzoraka otpadne vode uzorkovane u
proljeće Ames bakterijskim testom. Rezultati predstavljaju brojeve pozitivnih (His+)
revertanata po test ploči, uz pripadajuće srednje vrijednosti.
Uzorak
Salmonella typhimurium
TA 98 TA 100
br. His+
revertanata
Srednja
vrijednost
br. His+
revertanata
Srednja
vrijednost
1 25
27 26
205
177 191
2 23
21 22
209
225 217
3 20
34 27
225
230 227,5
4 29
30 29,5
220
195 207,5
5 19
25 22
233
202 217,5
6 25
29 27
221
240 230,5
7 25
22 23,5
252
210 231
8 19
18 18,5
196
186 191
9 28
23 25,5
209
271 240
10 20
28 24
194
226 210
Kontrole postupka
Kontrola bez bakterija 0
0 0
0
0 0
Kontrola-spontani
revertanti
29
33
22
28
178
180
200
186
Kontrola NOPD (20
µg/po ploči)
› 1300
› 1300
› 1 300
Kontrola 2-AF (20
µg/ploči)
› 1 800
› 1 800
› 1 800

62
Tablica 9. Rezultati analize mutagenog potencijala uzoraka otpadne vode uzorkovane u ljeto
Ames bakterijskim testom. Rezultati predstavljaju brojeve pozitivnih (His+) revertanata po
test ploči, uz pripadajuće srednje vrijednosti.
Uzorak
Salmonella typhimurium
TA 98 TA 100
br. His+
revertanata
Srednja
vrijednost
br. His+
revertanata
Srednja
vrijednost
1 15
13 14
125
133 129
2 19
24 21,5
120
129 124,5
3 21
23 22
135
129 132
4 19
24 21,5
139
132 135,5
5 25
23 24
117
125 121
6 27
24 25,5
132
139 135,5
7 20
17 12,3
135
129 132
8 22
26 24
118
125 121,5
9 25
29 27
132
126 129
10 16
19 17,5
127
135 131
Kontrole postupka
Kontrola bez bakterija 0
0 0
0
0 0
Kontrola-spontani
revertanti
16
17
21
18
122
129
127
126
Kontrola NOPD (20
µg/po ploči)
› 1 300
› 1 300
› 1300
Kontrola 2-AF (20
µg/ploči)
› 1 800
› 1 800
› 1 800

63
Tablica 10. Rezultati analize mutagenog potencijala uzoraka otpadne vode uzorkovane u jesen
Ames bakterijskim testom. Rezultati predstavljaju brojeve pozitivnih (His+) revertanata po
test ploči, uz pripadajuće srednje vrijednosti.
Uzorak
Salmonella typhimurium
TA 98 TA 100
br. His+
revertanata
Srednja
vrijednost
br. His+
revertanata
Srednja
vrijednost
1 13
15 14
119
138 128,5
2 9
12 10,5
132
141 136,5
3 9
17 13
129
165 147
4 18
15 16,5
177
155 166
5 19
10 14,5
137
138 137,5
6 7
20 13,5
137
143 140
7 12
19 15,5
139
127 133
8 15
19 17
172
145 158,5
9 20
20 20
165
157 161
10 10
13 11,5
197
164 180,5
Kontrole postupka
Kontrola bez bakterija 0
0 0
0
0 0
Kontrola-spontani
revertanti
14
14
16
14,6
135
121
143
133
Kontrola NOPD (20
µg/po ploči)
› 1 300
› 1 300
› 1300
Kontrola 2-AF (20
µg/ploči)
› 1 800
› 1 800
› 1 800

64
4.4. Mikrobiološke analize vode, mulja i tkiva riba
Slika 25. pokazuje log10 vrijednosti broja pojedinih skupina bakterija po mL vode ili g mulja
na svim lokacijama uzorkovanja za svaki vremenski period. Za mulj s deponije (Slika 2.,
lokacija uzorkovanja 11) prikazane su log10 vrijednosti broja pojedine skupine bakterija u 1
gramu uzorka.
Najveći broj ukupnih kolonija na 37 °C i 22 °C, ako se isključi aktivni mulj na UBP-u i kruti
otpadni mulj s deponije, zabiljeţen je u jesen u šećeranskim i pročišćenim otpadnim vodama.
U OĎenici je u svim ispitivanim terminima prisutnost ovih bakterija bila najmanja. U
istraţenim uzorcima voda u kanalu Manteč nakon Biljnog ureĎaja Lukač (Slika 2. lokacija
uzorkovanja 6) u proljeće i ljeto dolazi do povećanja broja kolonija na 37 °C i 22 °C. Za
razliku od jesenskog perioda, u Ţupanijskom kanalu nakon ulijevanja kanala Manteč, kao
prijemnika pročišćenih otpadnih voda, dolazi do povećanja broja kolonija na 37 °C i 22 °C.
Koliformnim bakterijama, fekalnim koliformima, bakterijom E. coli i enterokokima najviše su
bile zagaĎene komunalne otpadne vode a učinkovitost UPB-a u smanjenju njihovog broja bila
je preko 90 %. U Ţupanijskom kanalu u proljeće i ljeto broj koliformnih bakterija bio je
nizvodno povećan, a isti trend bio je sa E. coli u jesen. U proljetnom i jesenskom periodu broj
koliformnih bakterija, fekalnih koliforma, E. coli i enterokoka bio je veći u potoku OĎenica
nego u Ţupanijskom kanalu u koji se ulijeva prijemnik pročišćenih otpadnih voda, čime
postaje upitna teza da se radi o mikrobiološki nezagaĎenom potoku.

65
Slika 25 . Rezultati mikrobioloških ispitivanja vode i mulja (Slika 2., lokacija uzorkovanja 1-9
(voda), lokacija uzorkovanja 10-11 (mulj))
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10 c
fu/m
L
Broj kolonija na 37 °C
proljeće ljeto jesen
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10 c
fu/m
L
Broj kolonija na 22 °C
proljeće ljeto jesen
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10 c
fu/1
00
mL
Koliformne bakterije
proljeće ljeto jesen
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10 c
fu/1
00 m
L
Fekalni koliformi
proljeće ljeto
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10
cfu
/10
0 m
L
E.coli - uzorkovanje u jesen
0
1
2
3
4
5
6
7
8
9
10
1 2 3 4 5 6 7 8 9 10 11
log
10 c
fu/1
00
mL
Crijevni enterokoki
proljeće ljeto jesen

66
Rezultate ispitivanja bakterija Pseudomonas aeruginosa, Listeria monocitogenes i Salmonella
sp. u uzorcima vode i mulja prikazuje Tablica 11, a u Tablicama 12.-14, Slikama 27.-29.
prikazan je postotak bakterijskih izolata u uzorcima vode i mulja.
Tablica 11. Prisutnost Pseudomonas aeruginosa, Listeria monocytogenes i Salmonella sp. u
uzorcima vode i mulja
Proljeće Lokacija uzorkovanja (Prema Slici
2.)
Psudomonas
aeruginosa/100
mL
Listeria monocytogenes/100 mL Salmonella sp./100 mL
Ljeto
Jesen
1
neg(-) neg(-) -
neg(-) neg(-)
neg(-)
9,8x101 neg(-) neg(-)
2
2,0x103 neg(-)
poz(+)
3,0x103 poz(+) neg(-)
1,7x105 neg(-), L.inocua,L.grayi poz(+) poz(+)
3
neg(-) neg(-)
-
- - -
neg(-) neg(-) neg(-)
4
neg(-) neg(-)
neg(-)
neg(-) neg(-) neg(-)
2,0x102 neg(-) neg(-)
5
neg(-) neg(-)
-
neg(-) neg(-), L.ivanovii poz(+) neg(-)
1,1x103 neg(-), L.ivanovii poz(+) poz(+)
6
neg(-) neg(-)
-
1,5x101 neg(-), L.ivanovii poz(+) neg(-)
1,7x103 neg(-), L.welchimeri poz(+) neg(-)
7
neg(-) neg(-)
-
neg(-) neg(-) neg(-)
neg(-) neg(-) neg(-)
8
neg(-) neg(-) -
neg(-) neg(-), L.ivanovii poz(+) neg(-)
neg(-) neg(-), L.ivanovii poz(+) neg(-)
9
neg(-) neg(-)
-
neg(-) neg(-) -
2,0x101 neg(-) neg(-)
10
neg(-) neg(-) -
3,0x103 neg(-), L.inocua poz(+) neg(-)
>2,4x104 neg(-), L.grayi poz(+) poz(+)
11
neg(-) neg(-) -
- neg(-), L.inocua poz(+) neg(-)
1,0x102 neg(-), L.grayi poz(+) poz(+)

67
Najveći broj Pseudomonas aeruginosa prisutan je u komunalnim otpadnim vodama a iz
uzoraka šećeranskih otpadnih voda, kanala Manteč na mjestu ulijevanja u Ţupanijski kanal i
Ţupanijskog kanala nizvodno ova bakterija nije izolirana. U uzorcima iz potoka OĎenica
izoliran je veći broj Pseudomonas aeruginosa nego iz Ţupanijskog kanala. Na UBP-u se iz
otpadnih voda ukloni više od 99 % ovih bakterija.
Listeria monocytogenes pronaĎena je samo u komunalnim otpadnim vodama ljeti. Salmonella
sp. izolirana je u proljeće i ljeto u komunalnim otpadnim vodama i u jesen u uzorcima
muljeva.

68
Tablica 12. Postotak izolata u uzorcima vode (1-9) i mulja (10-11) u proljeće (API 20E i
MALDI-TOF MS)
MALDI-TOF MS 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
Aeromonas bestiarum - - - 2,44 - - - - - - -
Aeromonas caviae - - - - - - - - - - 2,44
Aeromonas hydrophila - 2,44 - - - - - - - - -
A. hydrophila ssp. - - - - 2,44 - - - - - -
Aeromonas media - - - 2,44 - - - - - - -
Aeromonas popoffii - 2,44 - - 2,44 - 4,88 - 2,44 - -
Aeromonas sorbia - - - - - - - 2,44 - - -
Aeromonas sp. - 2,44 - - - - - - - - -
Aeromonas sp. (2) - - - - 2,44 4,88 - 7,32 - - -
Aeromonas veronii - - - 2,44 - - - - - - -
Bacillus simplex - - - - - - 2,44 - - - -
Lysinibacillus fusirormis - - - - 2,44 2,44 - - - - -
Lysinibacillus sfaericus - - - - - - - - - - 2,44
Myroides odoratimimus 4,88 - - - - - - - - - -
Proteus vulgaris - - 7,32 - - - - - - - -
Providencia alcalifaciens - - 2,44 - - - - - - 2,44 -
Providencia retgeri 4,88 - - - - - - - - 7,32 -
Pseudomonas kilonensis - - - - - - - - - - 2,44
Pseudomonas thievervale - - - - - - - - - - 2,44
Serratia liquefaciens - 2,44 - - - - - - - - -
Shewanella baltica - - - - - - - - 2,44 - -
Shewanella profunda - - - - - 2,44 - - - - -
Shewanella putrefaciens - - - - - - - - 2,44 - -
API 20E 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
A.hydrophila group 1 - 3,85 - 11,54 11,54 3,85 3,85 11,54 7,69 - 7,69
A.hydrophila group 2 - 7,69 - - - 3,85 - 3,85 - - -
Ps.fluorescens/putida - - - - - - - - - 7,69 -
Serratia liquefaciens - 3,85 - - - - - - - - -
Shewanella putrefaciens - - - - - 3,85 3,85 - 3,85 - -
Slika 26. Postotak izolata MALDI-TOF MS/API 20E u uzorcima voda i mulja u proljeće
2,44
2,44
2,44
2,44
2,44
12,2
2,44
2,44
14,64
2,44 2,44 4,88 2,44
4,88
7,32
4,88
12,2
2,44
2,44
2,44
2,44
2,44
2,44
MALDI-TOF MS Aeromonas bestiarumAeromonas caviaeAeromonas hydrophilaA. hydrophila ssp.Aeromonas mediaAeromonas popoffiiAeromonas sorbiaAeromonas sp.Aeromonas sp. (2)Aeromonas veroniiBacillus simplexLysinibacillus fusirormisLysinibacillus sfaericusMyroides odoratimimusProteus vulgarisProvidencia alcalifaciensProvidencia retgeriPseudomonas kilonensisPseudomonas thievervaleSerratia liquefaciensShewanella balticaShewanella profunda
61,55 15,39
7,69
3,85 11,55
API 20E
A.hydrophilagroup 1
A.hydrophilagroup 2
Ps.fluorescens/putida
Serratialiquefaciens
Shewanellaputrefaciens

69
Tablica 13. Postotak izolata u vode (1-9) i mulja (10-11) u ljeto (API 20E i MALDI-TOF MS)
MALDI-TOF MS 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
Aeromonas caviae - - - - - 2,22 - - - - -
Aeromonas hydrophila - - 2,22 4,44 - 2,22 - 2,22 - 2,22 2,22
Aeromonas veronii - - 2,22 - 4,44 - 8,89 6,67 2,22 - -
Alcaligenes faecalis - 2,22 - - - - - - - - -
Bacillus cereus - - - - 4,44 2,22 - - - - 2,22
Bacillus megaterium - - - - - - - - 2,22 - 4,44
Enterobacter cowanii - - - - - - - - 4,44 - -
Escherichia coli - - 2,22 2,22 - - - - - - -
Klebsiella oxitoca - - - - - - - - - 2,22 -
Klebsiella pneumoniae - - - - - - - - - 4,44 -
Morganella morganii 2,22 - - - - - - - - - -
Protues mirabilis - 6,67 - - - - - - - - -
Providencia rettgeri - - 2,22 2,22 - - - - - - -
Pseudomonas oleovorans - - - - - 4,44 - - - - -
API 20E 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
A.hydrophila group 1 - 6,90 6,90 - 6,90 6,90 - - - - -
A.hydrophila group 2 10,3 3,45 - 6,90 - 3,45 13,8 3,45 - - 3,45
Bordetella/Alcaligenes/M
oraxella spp. - - - - - - 3,45 - - - -
Klebsiela ornithinolytica - - - 3,45 - - - - - - -
Klebsiella oxitoca - - - - - - - - 3,45 - -
Klebsiella terrigena - - - - - - - - 3,45 - -
Pantoea spp. 2 - - - - - - - 3,45 - - -
Rachnella aquatilis - - - - - - - 3,45 - - -
Serratia marcescens - - 3,45 - - - - - - - -
Vibrio cholerae - - - - - - - 3,45 - - -
Slika 27. Postotak izolata u uzorcima vode i mulja MALDI-TOF MS/API 20E - ljeto
2,22
15,54
24,44
2,22
8,88
6,66 4,44
4,44 2,22
4,44
2,22 6,67
4,44 4,44
MALDI-TOF MS Aeromonas caviae
Aeromonas hydrophila
Aeromonas veronii
Alcaligenes faecalis
Bacillus cereus
Bacillus megaterium
Enterobacter cowanii
Escherichia coli
Klebsiella oxitoca
Klebsiella pneumoniae
Morganella morganii
Protues mirabilis
Providencia rettgeri
Pseudomonasoleovorans
27,6
44,8
3,45
3,45
3,45
3,45
3,45 3,45
3,45 3,45
API 20E A.hydrophila group 1
A.hydrophila group 2
Bordetella/Alcaligenes/Moraxella spp.KlebsielaornithinolyticaKlebsiella oxitoca
Klebsiella terrigena
Pantoea spp. 2
Rachnella aquatilis
Serratia marcescens
Vibrio cholerae

70
Tablica 14. Postotak izolata u vode (1-9) i mulja (10-11) u jesen (API 20E i MALDI-TOF
MS)
MALDI-TOF MS 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
Actinobacter tandoii - - - - - - - - - 2,38 -
Aeromonas bestiarum 2,38 - - - - - - - - - -
Aeromonas caviae - - - - - - - 2,38 - - -
Aeromonas hydrophila - - - - 7,14 4,76 - 2,38 - - -
Aeromonas ichthiosmia - - - - 2,38 - - 2,38 - - -
Aeromonas media 2,38 - - - - - - - - - 2,38
Aeromonas veronii - - - - - 2,38 2,38 - 2,38 - -
Alcaligenes faecalis - - - - - - - - - 2,38 -
Bacillus megaterium - - - - - 2,38 - - - - -
Buttiauella gaviane 2,38 - - - - - - - - - -
Candida auilliermondii
(ana) (Pkichia
gulliermondii (telo)
- - - - - - - - 2,38 - -
Citrobacter braakii - - - - - - 2,38 - - - -
Comamonas testosteroni - - 4,76 - - - - - - - -
Delftia acidovorans - - 2,38 - - - - - - - -
Enterobacter asburiae - - - 2,38 - - - - - - -
Enterobacter cloacae - - - - - - - - - - 2,38
Enterobacter ludwigii - - - - - - 2,38 - - - -
Klebsiella pneumoniae - - - 2,38 - - - - - - -
Myroides odoratus 2,38 - - - - - - - - - -
Proteus vulgaris - - - - - - - 2,38 - - -
Providencia retgerii - - - 2,38 - - - - - - -
Providencia rustigianii - - 2,38 - - - - - - - -
Ps. chlororaphis - - - - - - - - 4,76 - -
Psudomonas putida - 2,38 - - - - 2,38 - - - -
Psudomonas vulgaris - 7,14 - - - - - - - - -
Roultella ornithinolytica - - - 2,38 - - - - - - -
Stenotrophomonas
maltophilia - - - - - - - - - - 2,38
Wauteriella falsenii - - - - - - - - - 2,38 -
API 20E 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11.
A.hydrophila group 1 11,5 - - - 7,69 3,85 - 3,85 - - 3,85
A.hydrophila group 2 - - - - 3,85 3,85 3,85 - 3,85 - 3,85
Citrobaccter braakii - - - - - - 7,69 - - - -
Enterobacter cloacae - - - - - - - - - - 3,85
Enterobacter sakazakii - - - - - - - - - - 3,85
Klebsiella oxitoca - - - 11,5 - - - - - - -
Klebsiella terrigena - - - 3,85 - - - - 7,69 - -
Ps. fluorescens/putida - - - - - - 3,85 - - - -
Serratia fonticola 3,85 - - - - - - - - - -
Serratia odorifera 1 - - - - 3,85 - - - - - -

71
Slika 28. Postotak izolata u uzorcima vode i mulja MALDI-TOF MS/API 20E - jesen
Rezultati bakteriološkog ispitivanja briseva s koţe i škrga riba prikazani su na Slici 29. Broj
aerobnih bakterija na +37 °C i +22 °C, koliforminih bakterija i enterokoka na svim ispitanim
2,38 2,38 2,38
14,28
4,76
4,76
7,14
2,38 2,38
2,38 2,38 2,38
4,76
2,38 2,38
2,38
2,38
2,38
2,38
2,38
2,38
2,38
4,76
4,76
7,14
2,38 2,38 2,38
MALDI-TOF MS
Actinobacter tandoii
Aeromonas bestiarum
Aeromonas caviae
Aeromonas hydrophila
Aeromonas ichthiosmia
Aeromonas media
Aeromonas veronii
Alcaligenes faecalis
Bacillus megaterium
Buttiauella gaviane
Candida auilliermondii/Pkichia gulliermondii
Citrobacter braakii
Comamonas testosteroni
Delftia acidovorans
Enterobacter asburiae
Enterobacter cloacae
Enterobacter ludwigii
Klebsiella pneumoniae
Myroides odoratus
Proteus vulgaris
Providencia retgerii
Providencia rustigianii
Ps. chlororaphis
Psudomonas putida
Psudomonas vulgaris
Roultella ornithinolytica
Stenotrophomonas maltophilia
Wauteriella falsenii
30,74
19,25
7,69 3,85
3,85
11,5
11,54
3,85
3,85
3,85
API 20E
A.hydrophila gr. 1
A.hydrophila gr.2
Citrobaccter braakii
Enterobacter cloacae
Enterobacter sakazakii
Klebsiella oxitoca
Klebsiella terrigena
Ps. fluorescens/putida
Serratia fonticola
Serratia odorifera 1

72
lokacijama značajnije je veći u ljeto a kvasaca i plijesni u jesen. E. coli nije utvrĎena a
Pseudomonas aeruginosa izoliran je na škrgama riba iz Ţupanijskog kanala nakon utoka
efluenta.
Slika 29. Prikaz mikrobiološke analize briseva s organa riba; K-koţa, Š-škrge
0
1
2
3
4
5
6
7
8
1. 8. 9. 1. 8. 9.
LJETO JESEN
log
10cfu
/1m
L
Broj kolonija na 37 °C i 22 °C
Broj kolonija u 1 ml na 37˚C K
Broj kolonija u 1 ml na 37˚C Š
Broj kolonija u 1 ml na 22˚C K
Broj kolonija u 1 ml na 22˚C Š
0
1
2
3
4
5
6
7
8
9
1. 8. 9. 1. 8. 9.
LJETO JESEN
log
10cfu
/100 m
L
Koliformne bakterije
Koliformne bakterije K
Koliformne bakterije Š
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1. 8. 9. 1. 8. 9.
LJETO JESEN
log
10cfu
/10
0 m
L
Crijevni enterokoki
Enterokoki K Enterokoki Š
0
0,5
1
1,5
2
2,5
3
1. 8. 9. 1. 8. 9.
LJETO JESEN
log
10 c
fu/1
mL
Kvasci i plijesni
Kvasci K Kvasci Š Plijesni K Plijesni Š

73
U Tablicama 15.-17. i Slikama 30.-32. prikazan je postotak izolata briseva s organa riba u
proljeće.
Tablica 15. Postotak izolata briseva s organa riba u proljeće (API 20E i MALDI-TOF MS) Š-
škrge, S-slezena, J-jetra, B-bubreg
MALDI-TOF MS 1. 8. 9.
Š J B Š J Š SL J
A.salmonicida spp. Salomicida 2,82
Acinetobacter generi 0,70
Acinetobacter guilouiae 0,70
Acinetobacter johnsonii 2,82
Acinetobacter pittii 2,11
Acinetobacter tondoii 0,70
Aeromonas bestiarum 14,1 2,82 0,70 1,56
Aeromonas encheleia 1,56
Aeromonas johnsonii 0,70
Aeromonas media 2,82
Aeromonas popoffii 0,70 1,56
Aeromonas sorbia 0,70 7,82 0,47
Aeromonas veronii 2,82 0,70 1,56 0,47
Jonthinobacterium lividum 0,70
Pseudomonas fragi 5,64
Pseudomonas graminis 2,82 2,82
Pseudomonas orientalis 0,70
Rheinheimera soli 14,1 2,11
Shewanella baltica 14,1 5,63 0,47
API 20E 1. 8. 9.
Š J B Š J Š SL J
A.hydrophila group 1 5,13 5,13 3,37
A.hydrophila group 2 2,56 12,8 12,8 4,72 12,8 12,8
Ochrobactrum anthropi 12,8 4,05
Ps.fluorescens/putida 5,13 5,13
Pseudomonas aeruginosa 0,67
Slika 30. Postotak izolata briseva s organa riba u proljeće (API 20E i MALDI-TOF MS)
2,8
0,7
0,7 2,8
2,1 0,7
19,2
1,6
0,7
2,8
2,3 9,0 5,6
0,7
5,6 5,6
0,7
16,2
20,2
MALDI-TOF MS A.salmonicida spp. SalomicidaAcinetobacter generiAcinetobacter guilouiaeAcinetobacter johnsoniiAcinetobacter pittiiAcinetobacter tondoiiAeromonas bestiarumAeromonas encheleiaAeromonas johnsoniiAeromonas mediaAeromonas popoffiiAeromonas sorbiaAeromonas veroniiJonthinobacterium lividumPseudomonas fragiPseudomonas graminisPseudomonas orientalisRheinheimera soliShewanella baltica
13,6
58,5
16,9
10,3
0,7 API 20 E
A.hydrophila group 1
A.hydrophila group 2
Ochrobactrum anthropi
Ps.fluorescens/putida
Pseudomonas aeruginosa

74
Tablica 16. Postotak izolata briseva s organa riba u ljeto (API 20E i MALDI-TOF MS)
Š-škrge, S-slezena, J-jetra, B-bubreg
MALDI-TOF MS OĎenica (1) Ţupanijski k. (8) Ţupanijski k. (9)
Š S J Š S J B Š S J B
Acitenobacter johnsonii 2,82 5,65 2,26 6,46
Aeromonas ichthiosmia 5,65
Aeromonas veronii 2,82 5,65 11,3 7,53 3,23 5,65 6,78 11,3
Enterobacter amnigenus 1,61
Enterobacter cloacae 3,76 4,52
Klebsiella oxitoca 5,65 2,26
Pseudomonas fulva 2,82
Shewanella putrefaciens
veronii 2,26
API 20E OĎenica (1) Ţupanijski k. (8) Ţupanijski k. (9)
Š S J Š S J B Š S J B
A.hydrophila group 1 3,13 1,88 1,34 4,70
A.hydrophila group 2 2,35 4,70 1,57 6,27 6,27 2,69 4,70 3,76 9,40
Enterobacter cloacae 1,57 1,88
Enterobacter sakazakii 3,13 1,88
Flavimonas oryzihabitans 1,88 2,68
Klebsiela oxytoca 4,70 1,88
Ochrobactrum anthropi 2,35
Pantoea ssp 1 9,40 3,13 1,34
Photobactrum damsela 2,35
Ps.fluorescens/putida 2,35
Serratia odorifera 1 1,57
Vibrio colerae 1,34 3,76
Slika 31. Postotak izolata s organa riba (MALDI-TOF MS/API 20 E)
17,19
5,65
54,26
1,61
8,28
7,91
2,82
2,26
MALDI-TOF MS Acitenobacter johnsonii
Aeromonas ichthiosmia
Aeromonas veronii
Enterobacter amnigenus
Enterobacter cloacae
Klebsiella oxitoca
Pseudomonas fulva
Shewanella putrefaciens
veronii
11,05
41,71
3,45 5,01
4,56
6,58
2,35
13,87
2,35
2,35 1,57
5,1
API 20E A.hydrophila group 1
A.hydrophila group 2
Enterobacter cloacae
Enterobacter
sakazakiiFlavimonas
oryzihabitansKlebsiela oxytoca
Ochrobactrum
anthropiPantoea ssp 1
Photobactrum
damselaPs.fluorescens/putida
Serratia odorifera 1
Vibrio colerae

75
Tablica 17. Postotak izolata briseva s organa riba u jesen (API 20E i MALDI-TOF MS)
Š-škrge, S-slezena, J-jetra, B-bubreg
MALDI-TOF MS OĎenica (1) Ţupanijski k. (8) Ţupanijski k. (9)
Š S J Š S J B Š S J B
Acitenobacter johnsonii 3,33 5,00 2,50 6,67
Aeromonas ichthiosmia 5,00
Aeromonas veronii 3,33 5,00 1,67 10 10 1,67 5,00 7,50 10,0
Enterobacter amnigenus 1,67
Enterobacter cloacae 3,33 2,50
Klebsiella oxitoca 5,00
Klebsiella oxitoca 5,00
Pseudomonas fulva 3,33
Shewanella putrefaciens veronii
2,50
API 20E OĎenica (1) Ţupanijski k. (8) Ţupanijski k. (9)
Š S J Š S J B Š S J B
A.hydrophila group 1 3,04 3,03 1,82 4,55
A.hydrophila group 2 2,27 4,55 1,52 6,06 6,06 3,03 4,55 4,55 9,09
Enterobacter cloacae 1,52 1,82
Enterobacter sakazakii 3,04 1,82
Flavimonas oryzihabitans 1,82 1,52
Klebsiela oxytoca 4,55 1,82
Ochrobactrum anthropi 2,77
Pantoea ssp 1 9,09 3,03
Photobactrum damsela 2,77
Ps.fluorescens/putida 2,77
Serratia odorifera 1 1,52
Vibrio colerae 1,52 4,55
Slika 32. Postotak izolata briseva s organa riba u jesen (API 20E i MALDI-TOF MS)
17,5
5
54,17
1,67
5,83 5
5
3,33 2,5
MALDI-TOF MS Acitenobacter
johnsonii
Aeromonas
ichthiosmia
Aeromonas veronii
Enterobacter
amnigenus
Enterobacter cloacae
Klebsiella oxitoca
Klebsiella oxitoca
Pseudomonas fulva
Shewanella
putrefaciens veronii
12,44
41,68
3,34 4,86
3,34
6,37
2,77
12,12
2,77
2,77 1,52
6,07
A.hydrophila gr.1
A.hydrophila gr.2
Enterobacter
cloacaeEnterobacter
sakazakiiFlavimonas
oryzihabitansKlebsiela oxytoca
Ochrobactrum
anthropiPantoea ssp 1
Photobactrum
damselaPs.fluorescens/putid
aSerratia odorifera 1

76
4.5. Osjetljivost izdvojenih sojeva prema antimikrobnim lijekovima
Osjetljivost prema antimikrobnim lijekovima bakterija izoliranih iz vode i mulja na
lokacijama uzorkovanja prema Slici 2. za sva tri ispitivana perioda (proljeće, ljeto, jesen)
prikazana je na Slici 33. Izvršeno je ispitivanje osjetljivosti na oksitetraciklin (OTC30),
amoksicilin (AMC30), oksolinsku kiselinu (OA2), eritromicin (E15), sulfametoksazol (SX50),
florfenikol (FFC30), norfloksacin (NOR10) i flumekvin (UB30). Osim prema amoksicilinu,
većina je sojeva, čak i iz referentnog potoka OĎenica (1), pokazala neosjetljivost prema
sulfametoksazolu i eritromicinu, što ukazuje na onečišćenje potoka OĎenica antimikrobnim
lijekovima. Neosjetljivost na pretraţivane antimikrobne lijekove bila je najizraţenija u
izolatima iz aktivnog mulja.
Proljeće Ljeto Jesen
Slika 33. Osjetljivost prema antimikrobnim lijekovima u vodi i mulju
Oznake: neosjetljivost R (=1); umjerena osjetljivost I (=2); najveća osjetljivost S (=3)
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1.
0
2
OTC
SM
X
AM
C
FFC
E
OA
NO
R
UB
1.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
2.
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
2.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
2.
0
2
4OTC
SMX
AMC
FFC
E
OA
NOR
UB
3.
0123OTC
SMX
AMC
FFC
E
OA
NOR
UB
3.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
3.

77
Slika 33. nastavak
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
4.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
4.
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
4.
0123OTC
SMX
AMC
FFC
E
OA
NOR
UB
5.
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
5.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
5.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
6.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
6.
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
6.
0
2
4OTC
SMX
AMC
FFC
E
OA
NOR
UB
7.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
7.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
7.
-1
1
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9.
0
2
4OTC
SMX
AM
C
FFC
E
OA
NOR
UB
9.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9.

78
Slika 33. Nastavak
0
1
2
3OTC
SM
X
AM
C
FFC
E
OA
NO
R
UB
10.
0123OTC
SMX
AMC
FFC
E
OA
NOR
UB
10.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
10.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
11.
0
2
4OTC
SMX
AM
C
FFC
E
OA
NOR
UB
11.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
11.

79
Najveću osjetljivost, bakterijski izolati iz tkiva riba lovljenih u potoku OĎenica, pokazali su
prema flumekvinu, oksolinskoj kiselini i norfoksacinu, u području od najveće do umjerene
osjetljivosti bili su oksitetraciklin, florfenikol i eritromicin, a u području izmeĎu umjerene
osjetljivost i neosjetljivosti bili su sulfametaksazol i amoksicilin (Slika 34).
Proljeće Ljeto Jesen
Slika 34. Osjetljivost prema antimikrobnim lijekovima bakterija poraslih iz briseva organa
riba u potoku OĎenica (Slika 2., lokacija uzorkovanja 1) prema sezonama i vrsti tkiva.
Oznake: neosjetljivost R (=1); umjerena osjetljivost I (=2); najveća osjetljivost S (=3); Š-
škrge, B-bubreg, J-jetra, SL-slezena
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/B
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/B
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
1/SL
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
1/SL

80
Najveću osjetljivost, bakterijski izolati iz tkiva riba lovljenih neposredno nakon izlijevanja
kanala Manteč u Ţupanijski kanal, u sve tri sezone, pokazali su na norfoksacin, zatim
oksolinsku kiselinu i flumekvin, u području izmeĎu najveće i umjerene osjetljivosti bili su
oksitetraciklin i florfenikol, umjerena osjetljivost pokazala se prema eritromicinu, a u
području izmeĎu umjerene osjetljivosti i neosjetljivosti bili su amoksicilin i sulfametaksazol
(Slika 35).
Proljeće Ljeto Jesen
Slika 35. Osjetljivost prema antimikrobnim lijekovima sojeva bakterija poraslih iz briseva
organa riba na lokaciji uzorkovanja 8 (Ţupanijski kanal) prema slici 2.. Oznake: neosjetljivost
R (=1); umjerena osjetljivost I (=2); najveća osjetljivost S (=3); Š-škrge, B-bubreg, J-jetra,
SL-slezena
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
8/Š
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
8/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8/B
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
8/B
0
1
2
3OTC
SMX
AM
C
FFC
E
OA
NOR
UB
8/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8/SL
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
8/SL

81
Najveću osjetljivost, bakterijski izolati iz tkiva riba lovljenih u Ţupanijskom kanalu nizvodno,
pokazali su prema flumekvinu, oksitetraciklinu, oksolinskoj kiselini i florfenikolu, u
području izmeĎu najveće i umjerene osjetljivosti bili su prema eritromicinu i
sulfametaksazolu, a u području od umjerene osjetljivosti i neosjetljivosti bili su prema
amoksicilinu (Slika 36).
Proljeće Ljeto Jesen
Slika 36. Osjetljivost prema antimikrobnim lijekovima sojeva bakterija poraslih iz briseva
organa riba na lokaciji uzorkovanja 9. (Ţupanijski kanal nizvodno) prema Slici 2.. Oznake:
neosjetljivost R (=1); umjerena osjetljivost I (=2); najveća osjetljivost S (=3); Š-škrge, B-
bubreg, J-jetra, SL-slezena
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/Š.
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/Š
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/B
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/B
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/B
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/SL
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/J
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/SL
0
1
2
3OTC
SMX
AMC
FFC
E
OA
NOR
UB
9/SL

82
Bakterije izolirane iz organa svih ulovljenih riba na sve tri lokacije bile su neosjetljive do
umjereno osjetljive na antimikrobne lijekove amoksicilin i sulfametaksazol. Umjereno
osjetljive do najviše osjetljive bile su na slijedeće antimikrobne lijekove: eritormicin,
florfenikol, oksitetraciklin, oksolinska kiselina, norfloksacin i flumekvin. Od svih bakterija iz
roda Aeromonas, čak je 50 % bilo rezistentno prema sulfametoksazolu, koji se pokazao kao
antimikrobni lijek koji je polučio najveću neosjetljivost u svim izolatima iz riba (osim
amoksicilina, za koji je neosjetljivost očekivana).

83
5. RASPRAVA
Ovo multidisciplinirano istraţivanje obuhvatilo je fizikalno-kemijsku, mikrobiološku i
ekotoksikološku karakterizaciju vode, mulja i sedimenta pod utjecajem UBP-a, kao i analize
vezane za indikatorske organizme, posebno ribe i gujavice, pokazujući specifične i
nespecifične promjene koje efluent izaziva u biološkim sustavima u ispitivanim medijima.
Ovakav multidisciplinarni pristup izabran je zbog mogućih pogrešnih intepretacija
pojedinačnih analiza i stoga podrazumijeva širi ekološki pristup te u konačnici daje
pretpostavke o mogućim posljedicama na ljudsko zdravlje.
Iako većina ekoloških studija uzima u obzir samo ograničenu količinu bioloških odgovora,
parametri koji pojedinačno ne pokazuju jednoznačan odgovor na onečišćujuće tvari, u
kombinaciji sa ostalim parametrima mogu biti korisni indikatori odgoĎenog utjecaja na okoliš
(Bernet i sur. 2000). Taj se utjecaj moţe pokazati ili kao izravan štetni učinak na stanice i/ili
tkiva, modifikacija fizioloških ili biokemijskih reakcija, povećanje infekcija uzrokovanih
fakultativnim patogenima ili kao smanjena otpornost organizma na infekcije (Schmidt-
Posthaus i sur. 2001). Stoga njihova determinacija na razini cijelog organizma moţe otkriti
subletalne i kronične učinke.
Rezultati ovog istraţivanja potvrĎuju hipotezu da učinci antibiotika, patogenih bakterija i
promjene bioloških odgovora vezanih za učinkovitost rada ureĎaja za pročišćavanje otpadnih
voda seţu daleko nizvodno od UBP-a.
Količina otpadne vode u procesu prerade šećerne repe moţe biti značajna, sa velikim
potencijalom zagaĎenja, posebno u odnosu na količinu zemlje i otpadaka koji dolaze sa
mehanizacijom dok se repa vadi za vrijeme kiše, a koja dospjeva u tvornicu sa korijenom, kao
i veliku količinu mulja koja ulazi u sustav putem pranja repe (Cheesman 2004). Posljedično, u
efluentu se značajno povećava suspendirana tvar, KPKCr , BPK-5 i amonijak. Prerada sirovog
šećera iz šećerne trske (proljeće) nije imala isti utjecaj na efluent kao prerada šećerne repe,
najviše zbog manje količine vode tijekom prerade, jer se ne koristi kao u preradi šećerne repe
za predpranje. MeĎutim, u procesu prerade šećerne trske ekstremne vrijednosti nitrata, dušika,
fosfata i fosfora dospijevaju efluentom u vodotoke (El-Shafai i sur. 2004). Tako povišeni
dušik i fosfor mogu rezultirati obraštajem algama, smanjenjem otopljenog kisika u vodi, što
djeluje štetno ili dovodi do uginuća riba i drugih vodenih organizama. Ingaramo i suradnici
(2009) predlaţu tri indeksa za učinak na okoliš proizvodnje šećerne trske: indeks potrošnje
vode koji uzima u račun efikasnost korištenja vode; indeks promjena u okolišu koji evaluira

84
povećanje KPK iznad dopuštene vrijednosti po lokalnom zakonodavstvu, indeks promjena
okolišu koji izračunava specifični porast KPK u efluentu s obzirom na potrošnju svjeţe vode.
U radu pogona za proizvodnju šećera potrebno je voditi računa o navedenenim postavkama.
Otpadne vode nakon prerade šećerne repe (Slika 2., Lokacija uzorkovanja 3) nadmašuju
gradske otpadne vode koje dolaze na UBP (Slika 2., Lokacija uzorkovanja 2) po vrijednosti
mnogih parametara, kao što su suspendirana tvar, KPKCr, KPKMn ,BPK-5, nitrati, nitriti, dok
se podudaraju u koncentraciji amonijačnog i ukupnog dušika. Do manje kontaminacije od
strane šećeranskih otpadnih voda moglo bi se doći uvoĎenjem naknadnog korištenja voda u
proizvodnom procesu, recikliranjem i regeneracijom već korištene vode od pranja šećerne
repe i iz tehnološkog procesa proizvodnje šećera.
Daljnji utjecaj na okoliš moţe se procijeniti mjerenjem indeksa utjecaja na okoliš kojim se
odreĎuju vrijednosti KPK u efluentu iznad dopuštenih vrijednosti (Ingaramo i sur. 2009). U
ovom radu KPK opada ispod dopuštenih vrjednosti u efluentu (Slika 2., lokacija uzorkovanja
4) i nizvodno (Slika 2., lokacije uzorkovanja 5-9).
UsporeĎujući koncentracije nitrita, ukupnog dušika i ukupnog fosfora u nezagaĎenim vodama
(Slika 2., lokacija uzorkovanja 1), ulaz netretirane gradske otpadne vode na ureĎaj za
pročišćavanje (Slika 2., lokacija uzorkovanja 2) i ulaz otpadne vode iz šećerane na ureĎaj za
pročišćavanje (Slika 2., lokacija uzorkovanja 3) sa literaturnim podacima (El-Shafai i sur.
2004), naše vrijednosti bile su značajno veće, uglavnom zbog visokih vrijednosti ukupnog
dušika na svim mjestima. Iako se u naš referentni vodotok, (Slika 2., lokacija uzorkovanja 1)
ne ulijevaju pročišćene otpadne vode, rezultati istraţivanja ukazuju na onečišćenje usljed
drugih antropoloških djelovanja (poljoprivredna proizvodnja, otpad animalnog podrijetla, i
slično).
Veliki broj ispitivanja obavljen je na ribi babuška koja je na svim mjestima izlova ulovljena u
najvećem broju u odnosu na druge vrste. Babuška naseljava nizinske rijeke, često u zajednici
sa podvodnom vegetacijom, prisutna je u poplavnim područjima, vrlo je tolerantna na niske
koncentracije kisika i visoke koncentracije različitih zagaĎivala (USFWS 2012). Riba
ulovljena na mjestima uzorkovanja nije značajno varirala s obzirom na biometrijske
vrijednosti i nalaze zdravstvenog pregleda, iako su babuške iz udaljenijih lokacija od ureĎaja
bile više infestirane nametnicima nego ribe izlovljene u kanalu u koji se ulijeva efluent, što se
moţe pripisati njihovoj mogućnosti migracije preko vodotoka pri traţenju boljih lokacija sa
hranom.
Od svih pronaĎenih nametnika na ribama u ovom radu prevalentni su jednorodni metilji
(Monogenea), škrţni paraziti koji svojom pojavnošću nisu preferirali samo specifične dijelove

85
istraţivanih vodotoka. Do istih rezultata došli su Billard i Khan (2003), koji su ustanovili da
su škrţni paraziti prisutni na ribama bez obzira na to jesu li one izloţene utjecaju komunalnih
ili industrijskih otpadnih voda. Osim toga, Kelly i Janz (2008) su ustanovili da je stupanj
infestacije, mjeren kao abundancija i biomasa intestinalnih nametnika i brojnost jednorodnih
metilja na škrgama štuka (Esox lucius), bio najveći u kontrolnoj skupini štuka, srednji u slabo
izloţenim štukama, dok ribe drţane pri visokoj izloţenosti nisu nosile nikakve parazite, što
korespondira s našim nalazima na ribama iz kanala koji prima efluent. Temperatura vode nije
bila značajno povezana sa brojnošću ovih ektoparazita, iako su male do srednje infestacije
zabiljeţene u proljeće i ljeto. MeĎutim, postojala je i negativna korelacija izmeĎu prevalencije
jednorodnih metilja i koncentracije olova u vodi, posebno u u proljeće (u efluentu otpadnih
voda) i u ljeto (nezagaĎena voda), kao što su ranije utvrdili Bayoumy i sur. (2008). Isti autori
su takoĎer zapazili i značajnu pozitivnu korelaciju infestacije jednorodnim metiljima i
koncentracija nikla u vodi, čemu idu u prilog i ova istraţivanja. Onečišćujuće tvari kao što su
teški metali mogu smanjiti imunološke sposobnosti riba koje obitavaju u tim vodama, čineći
ih tako osjetljivijima na nametnike (Overstreet, 1993), pa infestacija babuške jednorodnim
metiljima stoga sluţi kao pomoćni bioindikatorski model.
Strukturne i funkcionalne promjene naĎene u tkivu škrga babuški, posebno u kanalu u koji se
ulijeva pročišćena otpadna voda (Slika 2., lokacija uzorkovanja 5), odnose se u velikoj mjeri
na sekundarne lamele (škrţne listiće), što ukazuje na onečišćenje okoliša, kao što je i
pretpostavljeno u hipotezi ovog rada. Kompaktnost sekundarnih lamela i formiranje
interlamelarnih mostova smanjuje ukupnu respiratornu površinu škrga, što rezultira
smanjenjem kapaciteta difuzije, a time i izmjene plinova dišnog područja. Munshi i Singh
(1971) drţe da se nakon duţe izloţenosti onečišćivačima u riba stvara adaptivni fiziološki
odgovor koji priječi prolazak iritansa u krvotok, a utvrdili su da pod utjecajem ksenobiotika
dolazi do odvajanja epitelnih stanica, nakon čega slijedi reasocijacija i reagregacija u vidu
interlamelarnih mostova, kao i u ovom istraţivanju. Flores-Lopes i Thomaz (2011) takoĎer su
uočili nekoliko vrsta promjena tkiva škrga uzrokovanih različitim razinama degradacije
okoliša, kao što su odizanje epitela, nekroza, fuzija, hipertrofija, hiperplazija i proliferacija
škrţnih lamela. Ribe u ovom istraţivanju, izlovljene iz kanala koji prima efluent (Slika 2,
lokacija uzorkovanja 5), bile su izloţene visokim koncentracijama kroma, nikla i cinka
(Tablica 5). U ranijim radovima utvrĎeno je smanjenje dišnog području šarana koji su bili
izloţeni teškim metalima (Mustafa i sur. 2012; Schjolden i sur. 2007; Srivastava i Punia 2011;
Ţikić i sur. 1997). Histopatološke promjene na tkivu škrga mogu biti u vezi sa ksenobioticima

86
u efluentu, povećanom koncentracijom teških metala, mikrorganizmima na škrgama i
narušenom kvalitetom vode te mogu posluţiti kao vaţno ishodište u razlikovanju različitih
mjesta zagaĎenja (Omar i sur. 2013). Poznato je da se topljivi i suspendirani teški metali,
naročito bakar, lako apsorbiraju putem škrga riba i mogu uzrokovati štetne promjene strukture
tkiva (Mustafa i sur. 2012). U riba iz efluenta u ljeto i jesen oštećenja strukture škrga
uključuju skraćenja sekundarnih lamela, atrofiju i upalne procese sa nekrozom. Slični rezultati
dobiveni su u studijama praćenja stanja okoliša i potvrĎena su laboratorijskim ispitivanjima
(Georgieva i sur. 2010; Barišić i sur. 2015). Škrge riba iz efluenta bile su izloţene
progresivnijim promjenama koje uključuju epitelnu hiperplaziju s lamelarnom fuzijom,
hipertrofiju epitela i proliferaciju mukoznih stanica. Takve promjene na škrgama zabiljeţili su
i drugi autori (Marioara i sur. 2009; Parvathi i Sarsu 2011; Nikalje i sur. 2012) koji ih opisuju
kao tipične lezije na škrgama nakon produljene izloţenosti različitim teškim metalima.
Oštećenja tkiva jetre variraju u odnosu na godišnja doba, lokacije i vrstu zagaĎivala i
pokazuju bioakumulacijski karakter (Kessabi i sur. 2014), što dovodi do postupnog
propadanja stanica. Dilatacija sinusoida, pronaĎena u jetri riba na lokaciji s najvećim
utjecajem efluenta u jesen, vjerojatno je posljedica dugotrajnog zagušenja krvi i začepljenja
sinusoida koje su se proširile kako bi omogućile lakši protok metabolita i antioksidanata
(Nikalje i sur. 2012). Uočeni upalni klasteri izmeĎu jetrenih lobula mogu ukazivati na reakciju
na bakterijske podraţaje, kao što je Aeromonas spp. izoliran iz vode i tkiva jetre (Topić
Popović i sur. 2015). Uslijed toksičnog utjecaja teških metala dolazi do teškog oštećenja tkiva
bubrega i disfunkcije bubrega u raznih vrsta riba (Omar i sur. 2013). U ovom radu uočeno je
da do najvećih promjena dolazi na ribama iz efluenta ljeti, kad je u bubreţnom tkivu
ustanovljen povećan broj bazofilnih bakterija, dok je u jesen prevladavala nefroza. Prema
Cormieru i sur. (1995) razvoj nefroze i bazofilnog klastera moţe posluţiti kao opći
pokazatelj ekološkog stanja nefrotoksičnosti. TakoĎer, vakuolarna degeneracija slezene
utvrĎena je kod riba pod najvećim utjecajem efluenta i pokazatelj je onečišćenja teškim
metalima (Georgieva i sur. 2010).
Hematologija riba moţe biti vrijedan alat za klinička ispitivanja. Pod uvjetom da se
primjenjuju standardizirane tehnike, moguće je usporeĎivati parametre iz krvi riba izmeĎu
nezagaĎenih kontrolnih voda i populacija izloţenih onečišćivačima, s tim da je za odreĎene
vrste poznat i dostupan skup referentnih vrijednosti (Clauss i sur. 2008).
Osnovne vrijednosti hematokrita šarana (Cyprinus carpio) su od 28-32%, a pod uvjetima
stresa i povišenih temperatura vode vrijednost hematokrita se povećava (Hrubec i Smith 2000;

87
Stoskopf 1993). Iako u ovom istraţivanju nisu naĎene značajne razlike izmeĎu grupa babuški,
najviše ukupne vrijednosti zabiljeţene su u proljeće u Ţupanijskom kanalu nizvodno. Ribe iz
efluenta imale su najviše vrijednosti Hct-a u proljeće, ali ove vode su takoĎer imale najviše
vrijednosti koncentracije bakra (270 µg/L), slično nalazima Mustafa i sur. (2012), gdje su Hct
vrijednosti u šarana bile značajno povećane uslijed nedostatka kisika i uz visoke vrijednosti
Cu kojima su bili izloţeni. Visoke vrijednosti suspendirane tvari, KPKMn, BPK-5 i dušika u
kanalu u koji se ulijeva efluent u proljeće (kada je tvornica šećera preraĎivala sirovi šećer iz
šećerne trske), uz niske vrijednosti otopljenog kisika, zasigurno doprinose oslabljenoj
osmoregulaciji i oštećenju stanica škrga. Onečišćena voda tako uzrokuje dodatni stres sa
mogućom adrenergičnom stimulacijom krvotvornih organa uzrokujući oslobaĎanje
pohranjenih eritrocita u krvotok (Mustafa i sur., 2012). Iako imaju visoku toleranciju na teške
metale i otpornost na gubitak iona (Schjolden i sur. 2007), povišene koncentracije teških
metala moguće su pridonijele povećanju metabolita u plazmi babuški u našem istraţivanju
(TRIG ,CHOL, ALB, URE, TP u ribama iz efluenta u proljeće).
CHOL u plazmi značajno je varirao u jesen (ribe iz kanala u koji se ulijeva efluent u odnosu
na ribe iz Ţupanijskog kanala), kada su pojedinačne koncentracije bile najniţe i najviše kroz
sva sezonska doba i lokacije. Iako sezonska variranja vrijednosti CHOL u šarana ranije nisu
ustanovljena (Svetina i sur. 2002), koncentracija CHOL u krvi riba umjereno osjetljivih na
prisutnost zagaĎivala najvjerojatnije ovise o koncentraciji onečišćujućih tvari. Gluth i Hanke
(1985) izloţili su šarane subletalnim koncentracijama različitim organskim pesticidima i
zapazili smanjenje kolesterola u svim slučajevima. Do sličnog smanjenje CHOL dolazi i
nakon dugotrajnog izlaganja šarana ţivi (Heath 1995). Trigliceridi u plazmi takoĎer pokazuju
značajne razlike u riba iz kanala u koji se ulijeva efluent u proljeće u odnosu na ulovljenu ribu
u jesen, gdje su jesenske vrijednosti bile najniţe na svim mjestima uzorkovanja i u svim
sezonskim dobima. Takav drastičan pad moţe biti povezan i sa nedostatkom hrane u kanalu u
koji se ulijeva efluent, tim više što su GLU i CHOL takoĎer imali svoje minimalne vrijednosti
u tom kanalu u jesen. Ovi parametri predstavljaju glavne energetske metabolite koji se često
koriste u procjeni zdravlja riba i procjeni stresora u prirodi (Čoţ-Rakovac i sur. 2005).
TakoĎer, takve su rezerve energije ribama potrebne da ublaţavaju učinke stresa i sluţe kao
energetski puferi (Topić Popović i sur. 2012). Stoga, s obzirom na pad tih vrijednosti, vrlo je
vjerojatna imunosupresija i slabljenje imunološkog sustava šarana u kanalu u koji se ulijeva
efluent u jesen (Javed i Usmani 2015). MeĎutim, budući da ALT i ALP aktivnosti nisu bile
povišene u kanalu u koji se ulijeva efluent, štetni učinci, zbog polupropusnosti membrane koje
sprječavaju gubitak enzima iz oštećenog tkiva (jetre), mogu biti prolaznog karaktera (Topić

88
Popović i sur. 2006.). Visoku toleranciju šaranskih vrsta na teške metale moguće je objasniti
njihovom sposobnošću da izbjegnu hipoksiju i gubitak iona iz plazme (Schjolden i sur.
2007). Utjecaj teških metala doprinosi strukturnom i funkcionalnom oštećenju tkiva škrga,
kao i povećanju vrijednosti nekih metabolita plazme. Značajan porast vrijednosti URE, TP,
ALB i TRIG zapaţen je u riba iz kanala koji prihvaća efluent (Slika 2., lokacija uzorkovanja
5).
Povećanje koncentracije URE vjerojatno je posljedica inhibicije izlučivanja dušikovih
otpadnih produkata i nakupljanja URE u krvi (Bernet i sur. 2000), kao i histopatoloških lezija
na škrgama. Premda se povezuje s onečišćenjem, povišen TP u vezi je i s nutritivnim statusom
riba, kao što i povećanje vrijednosti organske tvari u pročišćenim otpadnim vodama moţe
dovesti do povećanje biomase beskraljeţnjaka (Bernet i sur. 2000), zbog čega dolazi do
povećanja TP i ALB. Uočena je jasna razlika izmeĎu riba koje su bile pod utjecajem efluenta i
riba koje nisu povezane s otpadnim vodama, a stres je na jetru imao štetne učinke. Slični
rezultati vezani za oštećenja tkiva uzrokovana stresom potvrĎeni su u radu Li i sur. (2011).
Oštećenja bubrega pod utjecajem stresa doseţu do riba lovljenih nizvodno od UBP-a na
posljednjoj mjernoj postaji (Slika 2., lokacija uzorkovanja 9), što pokazuju i vrijednosti CRE i
URE.
Potencijalno toksični spojevi u okolišnim uzorcima mogu inducirati oksidacijski stres kod
mikroorganizama, biljaka te mnogih kraljeţnjaka i beskraljeţnjaka. Oksidacijski stres nastaje
uslijed pomaka ravnoteţe u oksidacijsko-redukcijskim procesima, što rezultira stvaranjem
slobodnih radikala. Slobodni radikali najčešće obuhvaćaju reaktivne vrste kisika (ROS) i
molekulu . Nagli porast koncentracije slobodnih radikala moţe rezultirati oštećenjem
svih vrsta bioloških molekula (proteina, membranskih lipida, ugljikohidrata, nukleinskih
kiselina), što se u konačnici očituje u promjenama strukture i funkcije stanice, tkiva, organa
(Sohal i sur. 2002; Del Rio i sur. 2005). Budući da je SOD jedan od glavnih antioksidacijskih
enzima koji sluţe za detoksikaciju ROS-a, smanjenje njegove aktivnosti mijenja redoks status
stanica što se moţe pripisati inhibiciji radikala superoksida (Ural, 2013) implicirajući
smanjenu sposobnost da zaštiti stanice. Komunalne otpadne vode pokazuju smanjenjenu
SOD aktivnost, odraţavajući oštećenost SOD proteina zbog hiperprodukcije ROS-a (Karadag
i sur. 2014). U proljeće je zabiljeţeno značajno smanjenje SOD-a u riba koje su ulovljene u
kanalu prijemniku efluenta. U ovom radu SOD aktivnost je bila sezonskog karaktera, a SOD
vrijednosti varirale su najviše u Ţupanijskom kanalu s tendencijom povećanja u proljeće i u
ljeto, u ljeto su imale tendenciju pada, dok su se u jesen povećavale. Najveće ukupne
vrijednosti SOD-a izmjerene su u proljeće i ljeto u riba iz Ţupanijskog kanala nizvodno. U

89
pokusima s vodnim organizmima obično dolazi do povećanja enzimske aktivnosti
antioksidanasa, a visoka vrijednost SOD-a ukazuje na sposobnost riba za zaštitu od toksičnog
učinka radikala superoksidnog aniona (Huang i sur. 2007). Dokazano je to nakon izlaganja
zebrice (Danio rerio) fenolima, gdje je trend SOD aktivnosti u početku biopovišen, a s
vremenom je došlo do aklimatizacije (Zhang i sur. 2014.).
Hematološki i biokemijski parametri u riba mogu se značajno promijeniti pod utjecajem
različitih stresora. Premda promjene prikazane u ovom radu ukazuju da je kvaliteta vode
jedan od najvaţnijih čimbenika koji utječu na ukupnu fiziologiju istraţivanih riba, njihova
oštećenja su nespecifična za širok raspon tvari, što nameće potrebu evaluacije i s drugim
indikatorima i biomarkerima. U tom smislu se za procjenu onečišćenja voda često koristi MN
i ENA (Klobučar i dr, 2003;. Talapatra i Banerjee, 2007; Pavlica i sur, 2008;. Güner i
Muranh, 2011; Omar i sur., 2012; Carrola i sur., 2014). Tvorba mikronukleusa posljedica je
kromosomskih oštećenja ili disfunkcije diobenog aparata, a moţe nastati od acentričnih
dijelova kromosoma ili cijelih kromosoma koji su isključeni iz glavne jezgre tijekom stanične
diobe (Udroiu, 2006). Iako mehanizam formiranja nije u potpunosti objašnjen, ENA se
smatra genotoksičnim analogom mikronukleusa. U ovom istraţivanju prisutnost MN-a
zabiljeţena je svega kod nekoliko riba na sva tri mjesta uzorkovanja s vrlo niskim
frekvencijama. Razina ENA bila je veća (iako ne značajno) kod riba ulovljenih u kanalu koji
prihvaća vodu s prijemnika pročišćenih otpadnih voda što korespondira sa višim
koncentracijama teških metala. Diskrepanciju izmeĎu MN i ENA opisali su i drugi autori
(Günner i Muranh 2011; Monteiro i sur. 2011), koji su utvrdili značajnu indukciju nuklearnih
abnormalnosti, ali ne i mikronukleusa kod riba koje su bile izloţene bakru, kadmiju i olovu.
Gujavice E. fetida učinkovita su vrsta za program biomonitoringa koji ima za cilj prevenciju
degradacije ekosustava uslijed antropogenih aktivnosti (Tondoh i sur. 2007). One mogu
preţivjeti u vrlo zagaĎenom okolišu zbog sposobnosti staničnog izbacivanja ksenobiotika
posredstvom transportnih proteina i detoksikacijskim enzimima. Iako se uglavnom koriste
kao indikatori otpadnih voda koje su kontaminirane organskim antropogenim onečišćenjem
u tlu (Kinney i sur. 2010), utvrĎeno je da akumuliraju toksikante na razinama iznad onih
objavljenih za otpadne vode (Markman i sur. 2007). Mehanizam obrambene funkcije MXR i
akumulacija fluorescentnih supstrata u tkivu gujavica moţe biti smanjen specifičnim
inhibitorima (Bošnjak i sur. 2014). Zaštitna funkcija mehanizma MXR moţe biti inhibirana
sa više onečišćivača prisutnih u okolišu (Kurelec 1997; Markman i sur. 2007). Prisutnost u
okolišu kemosenzitivnih tvari ili inhibitora obrambenog sustava MXR u različitim

90
organizmima (vodeni, kopneni) moţe uzrokovati povećanje intracelularne akumulacije i
toksičnih učinaka drugih ksenobiotika normalno odstranjenih pomoću MXR transportnih
proteina [P-glikoproteina (P-GPS), MRPs]. Ekotoksikološki značaj MXR inhibicije potvrĎen
je in vivo studijama koje su pokazale povećanu proizvodnju mutagenih metabolita u
školjkama i povećan broj apoptotičnih stanica u embrijima morskih jeţinaca nakon izlaganja
modelu MXR inhibitora. TakoĎer je potvrĎeno da su MXR inhibitori prisutni kod
konvencionalnih i novih umjetnih zagaĎivala, pa tako neki pesticidi i sintetički mošusni mirisi
pokazuju izrazito snaţan inhibitorni potencijal MXR-a na ekološki relevantnim
koncentracijama (Smital i sur. 2004). Biotest je pokazao prisutnost MXR inhibitora u svim
ispitivanim uzorcima, posebno u razrijeĎenim uzorcima. Time smo potvrdili koncept da se
mjerenje inhibicije MXR-a moţe koristiti kao vrijedan biomarker onečišćenja okoliša (Epel i
sur. 2008, Hackenberger i sur. 2012).
U vodama nizvodno od UBP-a lokalno stanovništvo lovi ribu za rekreaciju i prehranu.
Njihovom konzumacijom otvara se mogućnost štetnog utjecaja na zdravlje ljudi, premda
korištenje pročišćene otpadne vode nije nepoznato u uzgoju riba (El-Shafai i sur. 2004;
Harnisz i Tucholski 2010). Mikrobiološka kvaliteta voda u kojima ribe obitavaju odraţava se
na mikrobiološku floru riba. Ako ukupan broj heterotrofnih aeroba prelazi 5x104/mL vode, to
predstavlja rizik ulaska humanih patogena u jestiva tkiva riba (Mara i Cairncross 1989).
Ukupan broj bakterija i intestinalnih enterokoka utvrĎenih u ovom radu bio je značajno veći u
vodi iz kanala koji prima otpadnu vodu u usporedbi s druga dva mjesta uzorkovanja riba
(potok OĎenica i Ţupanijski kanal, Slika 2., lokacije uzorkovanja 1, 9). Da bi se riba mogla
konzumirati, broj fekalnih koliformnih bakterija u vodi ne bi smio prijeći 103/100 mL (WHO
2006). Fekalni koliformi su na sve tri lokacije izlova riba bili ispod navedne vrijednosti, a u
najvećem broju bili su pronaĎeni u potoku OĎenica (3,2 x 102/100 mL). Relativno niska
razina fekalnih koliforma moţe se akumulirati u ribljem probavnom sustavu, uzrokujći rizik
transmisije do konzumenata preko kontaminiranih tkiva u slučajevima neodgovarajućih
higijenskih postupaka u pripremi riba (El-Shafai i sur. 2004; Harnisz i Tucholski 2010).
Rezultati identifikacije izolata iz vode i riba uzorkovanih na tri lokacije po API 20E i
MALDI-TOF MS testovima umnogome korespondiraju. Većina očitanja ukazuje na
Aeromonas spp. Poznato je da izolati roda Aeromonas iz okoliša imaju širu fenotipsku
raznolikost nego izolati dobiveni iz kliničkih uzoraka (Donohue i sur. 2007). Iako API 20E
pokazuje ograničenu osjetljivost za razlikovanje vrsta, grupirajući sve Aeromonas vrste u A.
hydrophila grupa 1 ili grupa 2, i uz manu sporih fenotipskih reakcija, to je vrijedan prvi korak
za preliminarnu identifikaciju bakterija. MALDI-TOF MS spektar pokazuje mnoge

91
interspecijske sličnosti, svaku Aeromonas vrstu definira pripadajućom jedinstvenom
distribucijom mase, koja korespondira sa spektrom “otiska prstiju” ili sa njihovim m/z
potpisom. MALDI-TOF MS se pokazala kao visoko reproducibilna metoda za brzo
razlikovanje Aeromonas spp. i sličnih vrsta, kao i diferencijaciju usko povezanih vrsta roda
Aaeromonas, gdje spektri različitih izolata iste vrste dolaze grupirani zajedno. Klaster analiza
m/z potpisa grafički opisuje kako su vrste slične jedna drugoj kao otisci prstiju, ukazuje na
visok početni stupanj povezanosti, nakon čega slijede vrste sa slijedećim najvećim stupnjem
povezanosti (Donohue i sur. 2006; Eddabra i sur. 2012; Topić Popović i sur. 2015a).
U vodenom okolišu su najzastupljenije mezofilne bakterije roda Aeromonas, uključujući
kloriranu vodu za piće, fekalnu otpadnu vodu, podzemnu vodu, zagaĎene i nezagaĎene
vodotoke (Huddleston i sur. 2006). Imajući u vidu da je većina bakterijskih izolata sa tkiva
riba, kao i iz uzoraka vode i mulja u ovom radu iz roda Aeromonas, potrebno je naglasiti da
ove bakterije mogu biti uzročnici zoonoza. Tako su npr. A. salmonicida ssp. salmonicida, A.
hydrophyila, A. sobria i A. caviae riblji patogeni (Austin i Austin, 1999) koji mogu uzrokovati
bolest kod riba preko koţnih infekcija i sistemskih infekcija, dok A. hydrophyila unesen
hranom u ljudi moţe urokovati gastroenteritis, proljev i sistemske infekcije (Austin i Austin
1999; Cabello i sur. 2013; Goni-Urriza i sur. 2000; Huddleston i sur. 2006; Lehane i Rawlin
2000). Sa popularnošću rekreativnog ribolova u vodama nizvodno od UBP-a, moguće je
očekivati više infekcija vezanih za vrste iz roda Aeromonas, dok njihova dijagnostika i
liječenje mogu biti sloţeni, posebno uzimajući u obzir rezistentnost ribljih patogena na
antbiotke. Onečišćenje okoliša antimikrobnim i ostalim antibiotskim tvarima značajno
doprinosi širenju otpornosti na antibiotike (Huddleston i sur. 2006), posebno kroz širenje
rezistentnih plazmida izmeĎu nepovezanih bakterija (Kostich i Lazorchak, 2008).
Od svih bakterijskih izolata sa tkiva riba u ovom radu, najveću rezistetnciju pokazale su
bakterije roda Aeromonas na AMC i SMX. AMC je vrlo sličan ampicilinu, na koji su te
bakterije prirodno neosjetljive (Harnisz i Tucholski, 2010), stoga je ovaj rezultat donekle bio
očekivan. Jedan od mogućih razloga za visoku rezistenciju na SMX je taj što on sam nije
aktivan protiv Aeromonas spp., ali u kombinaciji sa trimetoprimom je uglavnom učinkovit
(Goni-Urriza i sur. 2000). Učestalost rezistencije Aeromonas spp. iz okoliša na SMX u
ovom radu je bila značajno veća nego u sličnim studijama (Huddleston i sur. 2006; Imzlin i
sur. 1996; Kampfer i sur. 1999), najvjerojatnije zato jer su naši izolati prethodno bili izloţeni
antibioticima i kemoterapeuticima zbog ispuštanja bolničkih otpadnih voda i njihovog
utjecaja na sastav efluenta. Moguće je da se iz istog razloga pojavljuju izolati iz roda
Aeromonas rezistentni na oksitetraciklin i eritromicin. Harnisz i sur. (2011) su utvrdili da su

92
bakterije rezistentne na tetraciklin pouzdan indikator antimikrobne rezistencije i
mikrobiološke kvalitete površinskih voda pod antropološkim utjecajem. U ovom radu
prikazana je antimikrobna osjetljivost na svim mjestima uzorkovanja za sve izolate i sva
ispitivana godišnja doba. Najveća prosječna neosjetljivost pokazala se prema AMC, zatim
prema SMX, E, FFC, OTC, OA, NOR i UB. Rezistentnost bakterija na antimkrobne tvari
moţe biti i veća, budući da analize uglavnom prikazuju rezistenciju uzgojivih bakterija koje
čine samo mali udio ukupnih bakterija prisutnih u vodenom okolišu (Cabello i sur. 2013).
TakoĎer, otpornost na antibiotike presudna je za nastajanje zaraznih bolesti, dok putevi
površinskim vodama mogu predstavljati ključnu rutu širenja gena otpornih na antibiotke
(Pruden i sur. 2012). Ovim smo potvrdili hipotezu da je u vodi, mulju i ribama pod utjecajem
komunalnih i industrijskih otpadnih voda moguća prisutnost bakterija rezistentnih na
antibiotike, a koje su patogene i za ljude. Sukladno hipotezi, prisutnost visokih koncentracija
SMX-a u vodi, takoĎer naĎenog i u sedimentu, doprinosi značajno većoj rezistenciji bakterija
roda Aeromanas u okolišu prema ovom antibiotiku. Njegova srednja vrijednost od 50,70 ng/L
korespondira sa srednjom vrijednosti od 48,40 ng/L izmjerenom u vodama zagaĎenim
urbanim i ruralnim aktivnostima u Kini (Xu i sur. 2014). Isti autor navodi srednju vrijednost
SMX-a od 16,10 ng/g za odgovarajući sediment, dok vrijednosti u sedimentu iz ovog rada
(Slika 2, lokacije uzorkovanja 4, 5, 9) opadaju sa udaljenošću od UBP-a (19,60>5,07>0,99
ng/g). U područjima onečišćenim antibioticima (Xue i sur. 2013), razina SMX-a u vodama
bila je do 191 ng/L, no ipak nije pronaĎen u sedimentu. Isti autor navodi 43,5 ng/L
eritromincina (E) u vodi, što je dvostruko više nego u otpadnoj vodi koja dolazi na UBP u
Virovitici, što dokazuje da SMX i E predstavljaju relativno visoki ekološki rizik za osjetljive
vodne organizme. Često sulfonamidi, napose SMX, pokazuju najveću učestalost i
koncentraciju u odnosu na druge antimikrobne tvari testirane u vodama (Chen i Zhou 2014).
U sedimentu dugoročno dolazi do akumulacije antimikrobnih tvari kojima su izloţeni bentički
i ostali vodni organizmi. U sedimentima se dugoročno akumuliraju lijekovi, postajući izvorom
kontaminacije za bentičke i ostale vodne organizme Gibs i sur. (2013) opisuju vjerojatni
uzrok padajućeg trenda pojavljivanja eritromicina i ostalih antibiotika u vodama zbog učinka
razrjeĎenja podzemnim vodama i sorpcije na dnu sedimenta. U sedimentu ukupna
koncentracija antibiotika istraţenih u ovom radu opada od 144,55 do 18,58 ng/g sa
udaljenošču od ureĎaja za pročišćavanje otpadnih voda, što je komparabilno s radom Chen i
Zhou (2014) (101,0 do 12,4 ng/g). Iako se pretpostavlja da su bolnice glavni izvor
farmaceutika u gradskim otpadnim vodama, farmaceutici dospijevaju u vode i iz drugih izvora
(kućanstava i dr.), npr. u Velikoj Britaniji 70 % i u SAD-u 75 %. Gotovo 70 % ukupnih

93
antibiotika se ispuštaju nepromijenjeni kao aktivni spojevi (Kümmerer 2009). Često su
njihovi metaboliti, zbog svoje biotransformacije, bolje topljivi u vodi nego njihovi lipofilni
osnovni spojevi. To moţe objasniti veće udjele sulfa lijekova i SMX u jutarnjim satima kad se
usporede sa sredinom dana i večeri. Antimikrobne supstance, ukoliko dospiju u okoliš,
nepovoljno utječu na vodne i kopnene organizme, a samim time i na ljudsku populaciju.

94
6. ZAKLJUĈAK
Rezultati ovog istraţivanja ukazuju da učinci antibiotika, patogenih bakterija i promjene
bioloških odgovora vezanih za ureĎaj seţu daleko nizvodno od UBP-a. Pročišćene otpadne
vode, u površinskim vodama kao njihovom prijemniku, mijenjaju okolišne parametre i
doprinose opterećenju ksenobioticima, što ima vaţnu ulogu u slici antimikrobne otpornosti.
Histološka analiza tkiva riba u vodama koje su pod utjecajem pročišćenih otpadnih voda,
vrijednosti hematoloških pokazatelja i biokemijskih pokazatelja plazme krvi riba, pokazuju
značajne promjene uvjetovane kompleksnošću okolišnih stresora, što doprinosi njihovom
cjelokupnom fiziološkom stanju. Promijenjeni fiziološki parametri najviše se očituju
oštećenjima škrga, stanica jetre i povećanjem SOD aktivnosti u krvi, dok istraţivanja
metabolita i enzima plazme, posebice CHOL i TRIG pokazuju jaku povezanost sa godišnjim
dobima i radom postrojenja tvornice šećera (prerada šećerne repe). Prisutnost teških metala u
okolišu doprinosi promjenama morfologije tkiva u vidu gubitka stanične i strukture tkiva, kao
i strukturne heterogenosti različitih ribljih organa. Navedene promjene su u korelaciji s
razinom oksidacijskog stresa potvrĎenim ovim istraţivanjem. Metaboličke promjene u razini
jetrenih enzima potvrĎuju uočene patohistološke lezije tkiva jetre. Zdruţeno interpretirani
rezultati patohistoloških i biokemijskih analiza mogu posluţiti kao vrijedan biomarker u
utvrĎivanju zdravstvenog statusa riba i utjecaja onečišćenja slatkovodnih ekosustava. U ovom
istraţivanju utvrĎena je visoka tolerancija babuške na vode smanjene kakvoće zbog utjecaja
pročišćenih komunalnih, bolničkih i industrijskih otpadnih voda. Dokazano je da su biološki
odgovori na onečišćenje i štetni učinci na parametre krvi i tkiva riba prisutni i u udaljenim
dijelu vodotoka nizvodno od UBP-a. PotvrĎeno je da je stanični mehanizam efluksa
posredstvom ABC transportnih proteina u gujavici izvrstan biomarker onečišćenja okoliša i
predlaţe se njeno korištenje u studijama biomonitoringa. Rezultati mikrobioloških analiza
ukazuju na dominaciju bakterija roda Aeromonas koje su pokazale visoki stupanj
neosjetljivosti na antimikrobne lijekove.
Iako je u ovom istraţivanju dokazana vrijednost multidisciplinarnog pristupa problemima
utjecaja pročišćenih otpadnih voda na biotu, potrebno je i nadalje razvijati biomarkere
osjetljivosti vrsta, identifikaciju najranjivijih razvojnih stadija i populacija, sa ciljem praćenja
zdravstvenog statusa istraţivanih organizama, patogenih i toksičnih onečišćivača u svrhu
integralnog pristupa upravljanju vodenim sustavima.

95
7. POPIS SIMBOLA I KRATICA
– amonijev molibdat tetrahidrat
- srebro(II)-sulfat
- natrij diklorizocijanurat (Izosan G)
– sulfanilna kiselina
- kalij-hidrogen-ftalat
– mravlja kiselina
– metilni alkohol
– amonijev formiat
– vodikov peroksid
- sulfatna kiselina
– nitratna kiselina
kalij bikromat
– kalij-permanganat
– kemijska potrošnja kisika po bikromatu
– kemijska potrošnja kisika po permanganatu
ALB – albumin
ALP – alkalna fosfataza
ALT – alanin amonotransferaza
AMC30 – amoksicilin
BU – Biljni ureĎaj Lukač
CHOL – kolesterol
CRE – kreatinin
E15 – eritromicin
ENA – eritrociti sa nepravilnim jezgrama (test), (engl. erythrocyte nuclear abnormalities)
FFC30 – florfenikol
GGT – gamma-glutamil transferaza
GLU – glukoza
GSH-Px – glutation peroksidaza
– kloridna kiselina
– ţiva(II)-sulfat

96
MALDI-TOF MS (engl. Matrix-assisted laser induced desorption ionization-time of flight-
mass spectrometry) - masena spektrometrija - matricom potpomognuta laserska
desorpcija/ionizacija i analizator vremena leta
MN – mikronukleus (test)
– natrijev klorid
– natrijev hidroksid
NOR10 – norfloksacin
o.v. – otpadna voda
OA2 – oksolinska kiselina
OTC30 – oksitetraciklin
SOD – superoksid dizmutaza
SX50 – sulfametaksazol
TBARS - reaktivna supstanca tiobarbiturne kiseline (engl. thiobarbituric acid reactive
substances)
TP – ukupne bjelančevine
TRIG – trigliceridi
TŠ – Tvornica šećera Viro
UB30 – flumekvin
UBP – UreĎaj za biološko pročišćavanje otpadnih voda grada Virovitice
UHPLC-QTOF-MS - kvadrupolna time of flight masena spektrofotometrija
URE – urea

97
8. LITERATURA
1. Affonso EG, Polez VLP, Correa CF, Mazon AF, Araujo MRR, Moraes G, Rantin FT
(2002) Blood parameters and metabolites in the teleost fish Colossoma macropomum
exposed to sulfide or hypoxia. Comparative Biochemistry and Physiology Part C:
Toxicology & Pharmacology 133, 3, 357-382.
2. Allen DA, Austin B. i Colwell RR (1983) Numerical taxonomy of bacterial isolates
associated with a freshwater fishery. Journal of General Microbiology 129, 2049-
2062.
3. Ašperger D i sur. (2013) Analitika okoliša, HINUS & Fakultet kemijskog inţenjerstva
i tehnologije, Zagreb
4. Austin B, Austin D (1987) Bacterial fish pathogens: Disease in farmed and wild fish,
364 pp., Ellis Horwood Limited, Chichester, England
5. Austin B, Austin D, (1999). Bacterial fish pathogens: disease in farmed and wild fish.
3rd ed. Springer-Praxix series in aquaculture and fisheries 455 pp.
6. Bard, SM. (2000) Multixenobiotic resistance as a cellular defense mechanism in
aquatic organisms, Aquat Toxicol 48, In: Laird, L(ed), 357–389
7. Barišić J, Dragun Z, Ramani S, Filipović Marijić V, Krasnići N, Čoţ-Rakovac R,
Kostov V, Rebok K, Jordanova M (2015) Evaluation of histopathological alterations
in the gills of Vardar chub (Squalius vardarensis Karaman) as an indicator of river
pollution. Ecotox Environ Safe 118: 158-166.
8. Bayoumy EM, Osman HAM, Laya F, El-Bana, Hassanain MA (2008) Monogenean
parasites as bioindicators for heavy metals status in some Egyptian Red Sea fishes,
Global Veterinaria 2: 3, 117-122.
9. Bernet D, Schmidt-Posthaus D, Wahii H, Burkhardt-Holm P (2000) Effect of
wastewater on fish health: an integrated approach to biomarker responses in brown
trout (Salmo trutta L.). Aquat Ecosyst Stress Recover 8, 143-151.
10. Bernet SM, Schmidt H, Wahli T, Burkhardt-Holm P (2001). Effluent from a sewage
treatment works causes changes in serum chemistry of brownt trout (Salmo truta L.).
Ecotox Environ Safe 48: 140-147.
11. Bhatia M, Goyal D (2013) Analyzing remediation potential of wastewater through
wetland plants. Environ Prog Stress Recover 8, 143-151.

98
12. Billard SM, Khan RA (2003). Chronic stress in cunner, Tautogolabrus adspersus,
exposed to minicipal and industrial effluents. Ecotox Environ Safe 55: 1, 9-18.
13. Biswas S, Rolain JM (2013) Use of MALDI_TOF mass spectrometry for
identification of bacteria that are difficult to culture. Journal of Microbiological
Methods 92, 1, 14-24.
14. Bošnjak I, Bielen A, Babić S, Šveer L, Topić Popović N, Strunjak-Perović I, Čoţ-
Rakovac R, Sauerborn Klobučar R (2014) First evidence of the P-gycoprotein gene
expression and multixenobitic resistance modulation in earthworm. Arhives of
Industrial Hygiene and Toxicology 65, 67-75.
15. Braga PAC, Tata S, Goncalves dos Santos V, Barreiro JR, Vilczaki Schwab N, Veiga
dos Santos M, Nogueira Eberlina M, Ramires Ferreira C (2012) Bacterial
identification: from the agar plate to the mass spectrometer. RSC Advances 4, 994-
1008.
16. Cabello FC, Godfrey HP, Tomova A, Ivanova L, Dolz H, Millnao A, Buschmann AH
(2013) Antimicrobial use in aquaticulture re-examined: its relevance in antimicrobial
resistane and to animal and human health. Microbiol 15 (7), 1917-1942.
17. Cabral JPS (2010) Water microbiology. Bacterial pathogens and water. International
Journal of Environmental Research and Public Health 7, 3657-3703.
18. Carrasco KR, Tilbury KL, Myers MS (1990) Assessment of the piscine micronucleus
test as an in sistu biological indicator of chemical contminant effects. Can J Fish
Aquat Sci 47, 2123-2136.
19. Carrola J, Santos N, Rocha MJ, Fontainhas-Fernandes A, Pardal MA, Monteiro RA,
Rocha E (2014) Frequency of micronuclei and of other nuclear abnormalities in
erythrocytes of the grey mullet from the Mondego, Douro and Ave estuaries-Portugal.
Environ Sci Pollut Res Int
20. Charlton-Menys V, Liu Y, Durrington PN (2006) Semiautomated method for
determination of serum paraoxonase actxvity using paraoxon as supstrate. ClinChem
52, 453-457.
21. Cheesman OD (2004). Environmental impacts of sugar production: the cultivation and
processign of sugarcane and sugar beet. Surrey, UK: CABI Bioscientce UK Centre.
22. Chen K, Zhou JL (2014) Occurrence and bahavior of antibiotics in water and
sediments trom the Huangpu River, Shangai, Chine. Chemosphere 95, 604-612.
23. Christensen PJ (1977) The history, biology and taxonomy of the Cytophaga group.
Canadian Journal of Microbiology 23, 1589-1653.

99
24. Clauss TM, Dove AD, Arnold JE (2008) Hematologic disorders of fish. Vet Clin
North Am Exot Anim Pract
25. Cooper GM, Hausman RE (2010) Stanica: Molekularni pristup. Medicinska naklada,
Zagreb
26. Cormier SM, Neiheisel TW, Wernsing P, Racine RN, Reimschuessel R (1995) New
nephron development in fish from polluted waters: a possible biomarker.
Ecotoxicology 4, 3: 157-68.
27. Couillard CD, Counteney CC, Maodaonaid RW (2008) Chemical-environment
interactions affecting the risk of imapcts on aquatic organisms; a review with a
Canadian perspective-interacitons affecting vulnerability. Environ Rev 16, 19-44.
28. Čolak S (2011) Biokemijski profil hemolimfe triju vrsta dekapodnih rakova.
Disertacija, pp. 130, Sveučilište u Zagrebu, Agronomski fakultet, Zagreb.
29. Čoţ-Rakovac R, Strunjak-Perović I, Hacmanjek M, Topić Popović N, Lipej Z,
Šoštarić B (2005) Blood chemistry and histological properties of wild and cultured sea
bass (dicentrarchus labrax) in the North Adriatic Sea. Vet. Res Commun 29: 677-687.
30. Čoţ-Rakovac R, Šmuc T, Topić Popović N, Strunjak-Perović I, Hacmenjak M, Jadan
M. (2008). Novel methods for assessing fish blood biochemical data. Journal of
Applied Ichthyology 24, 1, 77-80.
31. da Silva Souza T, Fontanetti CS (2006) Micronucleus test and observation of nuclear
alterations in erythrocytes of Nile tapia exposed to waters affected by refinery effluent.
Mutat Res 605: 87-93.
32. Dallinger R, Prosi F, Segner H, Back H (1987) Contaminated food and uptake of
heavy metals by fish: a review and proposla for further research. Öecologia 73: 91-98.
33. Darwano H, Duy SV, Sauv S (2014) A new protocol for the analysis of farmaceutical
pesticides and hormones in sedimentand suspended particulate matter from rivers and
municipal wastewaters. Arch Environ Contam Toxicol 66(4), 582-593.
34. de Campos Ventura B, de Fransceschi de Angelis D, Marin-Morales MA (2007)
Mutagenic and genotoxic effects of the Atrazine herbicide in Oreochromis niloticus
(Perciformes, Cichllidae) detected by the micronuclei test and the comet assay. Pestic
Biochem Phys 90: 42-51.
35. Del Rio D, Stewart AJ, Pellegrini N (2005) A review of fecent studeis on
malondialdehyde as tocic molecule and biological marker of oxidative stress. Nutr
Metab Cardiovasc Dis 15(4): 316-328.

100
36. Donohue MJ, Smallwoods AW, Pfaller S, Rodgers M, Shoemake JA (2006) The
development of a matrix-assisted laser desorption/ionization-mass spectrometry-based
method for the protein ffingerprinting and identification of Aeromonas species using
whole cells. J Microbiol Methods 65, 380-389.
37. Eddabra R, Prevost G, Scheftel JM (2012) Rapid discrimination of environmental
Vibrio by matrix-assisted laser desorption ionization time-of-fliht mass spectrometry.
Microbiol Res 167, 226-230.
38. Edwards CA i Bohlen PJ (1996) Biology and ecology of earthworms. 3rd ed.
Chapman & Hall, London, UK.
39. El-Bouri K, Johnston S, Rees E, Thomas I, Bme-Mannathoko N, Jones C, Reid M,
Ben-Ismaeil B, Davies AR, Harris LG, Mack D (2012) Comparison of bacterila
identification by MALDI-TOF mass spectrometry and coventional diagnostic
microbiology methods. British Journal of Biomedical Science. 69, 2, 47-55.
40. El-Shafai SA, Gijzen HJ, Nard FA, El-Gohary FA (2004) Microbial quality of tilapia
reared in fecal-contaminated ponds. Environ Res 95, 231-238.
41. Epel D, Luckenbach T, Stevenson CN, Macmanus-Spencer LA, Hamdoun A, Smital T
(2008) Efflux transporters: newly appreciated roles in protection against pollutants.
Environ Sci Technol 42, 3914–3920.
42. Esteban S, Gorga M, Petrović M, Gonzalez-Alonzo, Barcelo D, Vacarcel Y (2014)
Analysis and accurrence of endocrine-disrupting compaunds and estrogenic activity in
the surface waters of Central Spain. Sci Total Environ 466-467, 939-951.
43. Fenech M (2000) The in vitro micronucleus technique. Mutat Res 455:81-95.
44. Fenech M (2003) HUMN project: detailed description of the scoring criteria for the
cytokinesis-bolck micronucleus assay using isolated human lymphocyte cultures.
Mutation Res Genetic Tox Environ Mutagen 534: 65-75.
45. Ferri J, Topić Popović N, Čoţ-Rakovac R, Beer-Ljubić B, Strunjak-Perović I, Škeljo
F, Jadan M, Petrić M, Barišić J, Šimpraga M, Stanić R (2011) The effect of artificial
feed of blood biochemistry profile and liver histology of wild saddled bream, Oblada
melanura (Sparidae). Marine Environmental Reasearch 71, 3, 218-224.
46. Flores-Lopes F, Thomaz AT (2011) Histopathologic alterations observed in fish gills
as a tool in environmental monitoring. Braz J Biol 71 (1), 179-188.
47. Frerichs GN, Roberts RJ (1989) The bacteriology of teleosts. In: Roberts, RJ (ed.),
Fish Pathology 289-319. London: Bailliere Tindall.

101
48. Garrido L, Sanchez O, Ferrera I, Tomas N, Mas J (2014) Dynamics of microbi8al
diversity profiles in waters of different qualities. Approximation to an ecological
quality indicator. Sci Tot Environ 468-469, 1154-1161.
49. Georgieva E, Arnaudov A, Velcheva I (2010) Clinical, hematological and
morphological studies on ex situ induced copper intoxication in Crucian carp
(Carassius gibelio). J Cent Eur Agr 11(2):165-172.
50. Gibs J, Heckathorn HA, Meyer MT, Klapinski FR (2013) Occurence and partioninig
of antibiotic compounds found in the water column and bottom sediments from a
stream receving two wastewater treatment plant effluents in Northern New Yersey. Sci
Total Environ 458-460, 107-116.
51. Glancer-Šoljan M, Landeka Dragičević T, Zanoški M, Šoljan V (2005) Biotehnologija
i okoliš, Interna skripta. Sveučilište u Zagrebu, Prehrambeno-biotehnološki fakultet,
Zagreb.
52. Gluth G, Hanke W (1985) A comparison of physiological changes in carp, Cyprinus
carpio, induced by several pollutants at sublethal concentrations. I. The dependency on
exposure time. Ecotox Environ Safe 2, 179-188.
53. Goni-Uriza M, Pineau L, Capdepury M, Roques C, Caumette P, Quentin C (2000)
antimicrobial resistance of mesophilic Aeromonas spp. strains isolated from two
European rievers. J Antimicrob Chemother 46, 297-301.
54. Günner U, Muranh FDG (2011) Micronucleus test, nuclear abnormalities and
accumulation of Cu and Cd on Gambusia affinis (Baird & Girard, 1853). Turkish
Journal of Fisheries and Aquatic Sciences 11, 615-622.
55. Hackenberger BK, Velki M, Stepić S, Hackenberger DK (2012) First evidence for the
presence of efflux pump in the earthworm Eisenia andrei. Ecotoxicology and
Environmental Safety 75, 1, 40–45.
56. Harnisz M, Tucholski S (2010) Microbial quality of common carp and pikeperch
fingerlingculturated in a pond fed with treated wastewater. Ecol Eng 36, 466-470.
57. Heath AG (1995) Water Pollution and Fish Physiology. Boca Raton, Florida: CRC
Press Inc.
58. Hrubec TC, Smith SA (2000) Hematology of fish. In Schalm's Veterinary
Hematology, 5th ed. (Feldman BF, Zinkl JQ and Jain NC, eds) pp. 1120-1125.
Philadelphia: Lippincott Williams and Wilkins.

102
59. Huang DJ, Zhang YM, Song G, Long J, Liu JH, Ji WH (2007) Contaminants-induced
oxidative damage on the carp Cyprinus carpio collected from the Upper Yellow River,
China Environ Monit Assess 128: 483-488.
60. Huddleston JR, Zak JC, Jeter RM (2006) Antimicrobial suspectibilities of Aeromonas
spp. isolated from environmental sources. Appl Environ Microbiol 72 (11), 7036-
7042.
61. Imziln B, Lafdal YMO, Jana M (1996) Effect of wastewater stabilization ponds on
antimicrobial susceptibility and haemolysin occurrence among mitile Aeromonas
strains. World J Microbiol Biotechnol 12, 385-390.
62. Ingaramo A, Heluane H, Colombo M, Cesca M (2009) Water and wastewater eco-
efficiency indicators for the sugar cane industry. J Clean Prod 17, 487-495.
63. ISO (International Organization for Standardization) 16240, 2005. Water quality -
Determination of the genotoxicity of water and waste water – Salmonella/microsome
test (Ames test). Geneva, Switzerland. (n.d.).
64. Javed M, Usmani N (2015) Stress response of biomolecules (carbohyddrate, protein
and lipid profiles) in fish Channa punctatus inhabiting river polluted by thermal power
plant effluent. Saudi J Biol Sci 22: 237-242.
65. Jiraungkoorskul W, Sahaphong S, Kosai P, Kim MH (2007) Micronucleus test: the
effect of ascorbic acid on cadmium exposure in fish (Puntius altus). research J Environ
Toxicol 1: 27-36.
66. Kampfer P, Christmann C, Swings J, Huys G (1999) In vitro susceptibilities of
Aeromonas genomic spacies to 69 antimicrobial agensts. Sys Appl Microbiol 22, 662-
669.
67. Karadag H, Firat O, Firat O (2014) Use of oxidative stress biomarkers in Cyprinus
carpio L. for the evaluation of water pollution in ataturk Dam Lake (Adiyaman,
Turkey). B Environ Contam Tox 92: 289-293.
68. Karlson P (1988). Biokemija, pp. 396, Školska knjiga, Zagreb
69. Kaur R, Dua A (2014) Adverse effeccts on histology of liver and kidney in fish
Chamma punctatus exposed to wastewater from Tung Dhab drain in Amritsar, India J
Evnivron Biol 35, 265-293.
70. Kelly JM, Janz DM (2008). Altered energetics and parasitism in juvenile notrhern pice
(Esox lucius) ingabiting metal-mining contaminated lake. Ecotox Environ Safe 70: 3,
357-369.
71. Kepec S (2015). Virkom d.o.o. Virovitica, Izvještaj 2013./2014.g, Laboratorij.

103
72. Kessabi K, Hwas Z, Sassi A, Said K, Messaoudi I (2014) Heavy metal accumulation
and histomorphological alterations in Aphanius fasciatus (Pisces, Cyprinodontidae)
from the Gulf of Gabes (Tunisia). Environ Sci Pollut R 21, 24: 14099-14109.
73. Kinney CA, Furlong ET, Kolpin DW, Zaugg SD, Burkhardt MR, Bossio JP, Werner
SL (2010) Eartworm: Diagnostic indicators of wastewater-derived anthropogenic
organic contaminants in terrestrial environments. In: Contaminants of Emerging
Concern in the Envvironment. Ecological and Human Health Conciderations (ed.
Halden, R). ACS Sympsium Series pp 297-317.
74. Klobučar GI, Stambuk A, Pavlica M, Sertić Perić M, Kutuzović Hackenberger B,
Hylland K (2010) Genotoxicity monitoring of feshwater environments using caged
carp (Cyprinus carpio). Ecotoxicology 19(1): 77-84. doi: 10.1007/s 1007/s 10646-009-
0390-6. Epub Jul 21.
75. Klobučar GIV, Pavlica M, Erben R, Papeš D (2003) Application of the micronucleus
and comet assays to mussel Dreissena polymorpha haemocytes for genotoxicity
monitoring of freshwater environments. Aquatic Toxicology 64, 15-23.
76. Koeman JH, Köhler-Günther, Kurelec A, Riviere JL, Verstee G, Walker CH (1993)
Applications and objectives of biomarker research. In. Peakall DB Shugart LR (eds)
Biomarkers. Springer. Berlin Hidelberg New-York, pp 1-13.
77. Kolkwitz R, Marsson M (1902) Grundsätze für die biologische Beurteilung des
Wassers nach seiner Flora und Fauna. Mitteilungen aus der Königlichen
Prüfungsanstadt für Wasserversorgung und Abwässerbeseitigung 1; 33-72.
78. Kolkwitz R, Marsson M (1908). Ökologie der pflantzlichen Saprobienüä.
79. Kolkwitz R, Marsson M (1909). Ȍkologie der tierichen Saprobien. International
Revue der gesamten Hydrobiologie und Hydrographie 2; 126-152.
80. Kostich MS, Lazorchak JM (2008) Risks in aquatic organisms posed by human
pharmaceutical use. Sci Total Environ 389 (2-3), 329-339.
81. Kümmerer K (2009) Antibiotics in the aquatic environment - a review part 1.
Chemosphere 75, 417-434.
82. Kurelec B (1997) A new type of hazardous chemical: The chemosensitizers of
multixenobiotic resistance. Environmental Health Perspectives 105, 4, 855-860.
83. Kurelec B (1992) The multixenobiotic resistance mechanism in aquatic organisms.
Crit Rev Toxicol 22(1):23–43.
84. Lehane L, Rawlin GT (2000) Topically acquired bacterial zoonoses form fish: a
review. Med J Aust 173(5), 256-259.

104
85. Li Z-H, Velisek J, Grabic R, Li P, Kolarova J, Randak T (2011) Usae of hematological
and plasma biochemical parameters to assess the chronic effects of a fungicide
propiconazole on a freshwater teleost. Chemosphere 83, 572-578.
86. Liebmann H (1962). Biologie des Trinkwasseer, Badewasser, Fischwasser, Vorfluters
und Abwasser U. Handbuch der Frischwasser- und Abwasserbiologie, knjiga 1.
München: Oldenbourg.
87. Litman T, Druley TE, Stein WD, Bates SE (2001) From MDR to MXR: new
understanding of multidrug resistance systems, their properties and clinical
significance. Cell Mol Life Sci 58(7), 931-59.
88. Lucas FS, Therial C, Conçalves A, Servais P, Rocher V, Mouchel JM (2014)
Vaariation of raw wastewater microbiological yuality in dry and wet weater
conditions. Environ Sci Pollut Res 21 (8), 5318-5328.
89. Mara D, Cainccross S (1989) Guidelines for the Safe use of Wastewater and Exccreta
in Agriculture and Aquaculture: Measures for Public Health Protection. p.187, World
Health Organization, Geneva, Switzerland.
90. Marioara N, Dumitrescu G, Petculescu-Ciochină L, Bănăţean-Dunea I., Moţ M,
Tăpălagă I, Lunca M, Boca L (2009) Pathological tissue lesions induced by chronic
cadmium intoxication in silver Crucian carp Carassius auratus gibelio. SPASB 42, 2:
84-90 .
91. Markman S, Guschina IA, Barnsley S, Buchanan KL, Pascoe D, Muller CT (2007)
Endocrine disrupting chemicals accumulate in earthworms exposed to sewage
effluent. Chemosphere 70, 119-125.
92. Martin-Carnahan A, Joseph SW (2005) Order XII. Aeromonadales ord. nov, 2nd
edition In: Brenner, DJ, Krieg NR, Staley JT, Garrity GM (Eds.), Bergey's Manual of
Systematic Bacteriology, Vol.2. New York: Springer.
93. Martone-Rocha S, Piveli RP, Matte GR, Boria MC, Dropa M, Monita M, Peternella
FA, Matte MH (2010) Dynamics of Aeromonas species isolated from wastewater
treatment system. J Water Health 08.4, 703-711.
94. Monteiro V, Cavalcante DGSM, Vilela MBFA, Sofia SH, Martinez CBR (2011) In
vivo and in vitro eyposures for the evaluation of the genotoxic effedt sf lead on the
neotropical freshwater fish Prochilodus lineatus. Aquat Toxicol 104, 291–298.
95. Mortelmans K, Zeiger E (2000) The Ames Salmonella/microsome mutagenicity assay.
Mutation Research 455, 29-60.

105
96. Munshi JSD, Singh RK (1971) Investigation of the effect of insecticides and other
chemical substances on the respiratory epithelium of predatory and weed fishes .
Indian J Zool 22, 127-134.
97. Mustafa SA, Davies SJ, Jha AN (2012) Determination of hypoxia and dietary copper
mediated sub-lethal toxicity in carp. Cyprinus carpio, at different levels of biological
organization. Chemosphere 87, 413-422.
98. Naidoo S, Olaniran A (2014) Treated Wastewater Effluent as a Source of Microbial
Pollution of Surface Water Resources. Int J Environ Res Public Health 11(1), 249-270.
99. Nikalje SB, Muley DV, Angadi, SM (2012) Histopathological changes in liver of
freshwater major carp, Labeo rohita after acute and chronic exposure to textile mill
effluent. The Bioscan 7, 2: 215–220.
100. Novotny L, Dvorska L, Lorencova A, Beran V, Pavlik I (2004) Fish: a potential
source of bacterila pathogens for human beings. Veterinarni Medicina 49, 9, 343-358.
101. OECD (Organization for Economic Cooperation and Development. (1984). OECD
Guidelines for Testing of Chemicals. Section 2. Effects on Biotic Systems. Test No.
207: Earthworms, Acute Toxicity Tests (Filter Paper Test and Artificial Soil Test).
Paris: OECD.
102. Omar WA, Zaggkizk KH, Abdel-Khalek AA, Abo-Hegab S (2013) Risk Assessment
and Toxic Effects of Metal Pollution in Two Cultured and Wild Fish Species from
Highly Degraded Aquatic Habitats. Arch Environ Contam Toxicol 65(4): 753-764.
103. Omar WA, Zaghloul KH, Abdel-Khalek A, Abo-Hegab S (2012) Genotoxic effects of
metal pollution in two fish species, Oreochromis niloticus and Mugil cephalus, form
highly degraded aqutatic habitats. Mutat Res Genet Toxicol Environ Mutagen 746: 7-
14.
104. Overstreet RM (1993) Parasitic deseases of fishes and their relationship with toxicants
and other environmental factors. In: Couch JA, Fournie JW (eds), Pathobiology of
Marine and Estuarine Organisms, CRC, Press, pp 111-156.
105. Pantle E, Back H (1955) Die biologische überwachung der Gewäser und die
Darstellung der Ergebnise. Gas- und Wasserschaft 96: 1-604.
106. Parvathi KS, Sarsu C (2011) Effect of chromium on histological alteration of gill, liver
and kidney of freshwater teleost, Cyprinus carpio. Int J Fish Aquac 6(1):1-5.
107. Pavlica M, Podrug M, Štambuk A, Cjetko P, Klobučar GIV (2008) Seasonal
variability in micronuclei induction in haemocytes of mussels along the eastern
Adriatic coast. Polish Journal of Environmental Studies 17, 5, 765-771.

106
108. Pavlica M (2012) Genetika. Poglavlje 14. Genske mutacije. Web udţbenik. I izdanje,
Udţbenici zagrebačkog sveučilišta, elektronička izdanja. Zagreb.
109. Pearse, AGE (1968) Histochemistry, theoretical and applied. 3rd ed.J.and Churchill
Ltd. London.
110. Pejić S (2014) Prisutnost i uklanjanje mikroorganizama u vodama različitog porijekla -
završni rad, p.21. Sveučilište Josipa Jurja Strossmyera u Osijeku, Prehrambeno-
tehnološki fakultet, Osijek.
111. Polak-Juszczak L (2012) Bioaccumulation of mercury in the trophic chain of flatfish
from the Baltic Sea. Chemosphere 89: 585-591.
112. Polard T, Jean S, Merlina G, Laplanche C, Pinelli E, Gauthier L (2011) Giemsa versus
acridine orange staining in the fish micronucleus assay and validation for use in water
quality monitoring. Ecotoxicology and Environmental Safety 74, 1, 144-149.
113. Poli G, Parola M (1997) Oxidative damage and fibrogenesis. Freee Rad Biol Med, pp.
22:287-305.
114. Pruden A, Arabi M, Stortenboom HN (2012) Corelation between upstream human
activities and riverine antibiotic resistane genes. Environ Sci Technol 46, 11541-
11549.
115. Republika Hrvatska, Drţavni ured za reviziju, Područni ured Virovitica (2013.)
Izvješće o obavljenoj financijskoj reviziji Opće bolnice Virovitica za 2011.
116. Ribera I, Dolédec S, Downie IS, Foster GN (2001) Effecct of land disturbance and
stress on species trait of ground beetle assemblages. Ecology, A publication of the
ecological society of America.
117. Ringo E, Strom E (1994) Microflora of Arctic charr, Salvelinus alpinus (L.):
gastrointestinal microflora of free-living fish and effect of diet and salinity on
intestinal microflora. Aquaculture and Fisheries Management 25, 623-629.
118. Sauerborn Klobučar R, Ţaja R, Franjević D, Brozović A, Smital T (2010) Presence of
ecotoxicologically relevant Pgp and MRP transcripts and proteins in cyprinid fish. Arh
Hig Rada Toksikol 61:175-182.
119. Savigny JC (1826) Analyse d'un Mémoire sur les Lombricus par Cuvier . Mém Acad
Sci Inst France 5: 176-184.
120. Schjolden J, Sørensen J, Nilsson GE, Poléo ABS (2007) The toxicity of copper to
crucian carp (Carassius carassius) in soft water. Sci Total Environ 384: 1-3, 239-251.
121. Schmidt-Posthaus H, Bernet D, Wahli T, Burkhardt-Holm P (2001) Morphological
organ alterations and infectious diseases in brown trout Salmo trutta and rainbow trout

107
Oncorhynchus mykiss exposed to polluted river water. Diseases of Aquatic Organisms
44: 161-170.
122. Servais P, Garcia-Armisen T, George I, Billen G (2007) Fecal bacteria in the rivers of
the Seine drainage network; souce, fate and modeling. Sci Total Environ 375, 153-
167.
123. Sies H (1985) Oxidative stress. Orlando: Academic press.
124. Sinal CJ, Webb CD, Bend JR (1999) Differential in vivo effects of alpha-
naphthoflavone and beta-naphthoflavone on CYP1A1 and CYP2E1 in rat liver, lung,
heart, and kidney. J Biochem Mol Toxicol 13(1): 29-40.
125. Sladeček V (1973) System of water quality from the biological point of view. Arhiv
für Hydrobiologie, Beiheft Ergebnisse der Limnologie 7: 1-218.
126. Smital T, Luckenbach T, Sauerborn Klobučar R, Hamboun MA, Vega LR, Epel D
(2004) Emerging contaminats - pesticides, PPCPs, microbial degradation products and
natural substances as inhibitors of multixenobiotic defence in aquatic organisms.
Mutation Research 552, 101-117.
127. Sohal RS (2002) Role of oxidative stress and protein oxidation in the aging process.
Free Radic Biol Med 37.
128. Southgate P (1990) Disease in aquaculture. In: Brown, L. (ed.), Aquaculture for
Veterinarians, Fish Husbandry and Medicine.91-128. Oxford: Pergamon Press.
129. Southgate P (1993) In Aquaculuture for Veterinarians, p 92. New York: Brown, L. Ed;
Pergamon Press.
130. Srivastava R, Punia P (2011) Effect of heavy metal on biochemical and hematologic
parameters in Cyprinus carpio and its use as a bioindicator of pollution stress. J
Ecophysiol Occup Health 11(1-2), 21-28.
131. Stepić S (2010) Promjene toksičnih učinaka smjesa pesticida na molekularne markere
izloţenosti kompostne gujavice (Eisenia andrei), disertacija. Sveučilište Josipa Jurja
Strossmayera u Osijeku i Institu RuĎer Bošković Zagreb.
132. Stoskopf M (1993) Clinical pathology of carp, goldfish, and koi. In Fish Medicine
(Stoskopf, M., ed), pp. 450-453. Philadelphia: W. B. Saunders.
133. Stoskopf MK (1993) Fich medicine. pp 882, W.B. Saunders Co, Philadelphia.
134. Strunjak-Perović I, Čoţ-Rakovac R, Topić Popović N, Jadan M (2009) Seasonality of
nuclear abnormalities in gilthead sea bream Sparus aurata (L.) erythrocytes. Fish
Physiology and Biochemistry 35, 287-291.

108
135. Svetina A, Matašin Z, Tofant A, Vučemilo M, Fijan N (2002) Haematology and some
blood chemical parameters of young carp till the age of three years. Acta Vet Hun 50:
4, 459-467.
136. Šimunić I, Senta A, Tomić F (2006) Potreba i mogućnost navodnjavanja
poljoprivrednih kultura u sjevernom dijelu Republike Hrvatske. Agronomski glasnik .
137. Šmuc T (2009) Kiselo-bazna svojstva Mn(III) meso-tetrakis((N-butil)piridin-2-il)
porfirina u vodenom mediju. Sveučilište u Zagrebu, Farmaceutsko-biokemijski
fakultet, Zagreb.
138. Štraus B (1988) Medicinska biokemija. p. 1058. Jugoslavenska medicinska naklada,
JUMENA, Zagreb.
139. Švob M (1974) Histološke i histokemijske metode. Svjetlost, Sarajevo.
140. Talapatra SN, Banerjee SK (2007) Detection of micronucleus and abnormal nucleus in
erytrocyte from the gill and kidney of Labeo bata cultivated in sewage-fed fish farms.
Food and Chemical Toxicology 45, 210-215.
141. Tondoh JE, Monin LM, Tiho S, Czuzdi C (2007) Can earthworms be used as bio-
indicators of land-use perturbations in semi-deciduous forest?. Biology and Fertility of
Soils 43, 585-592.
142. Topić Popović N (1997) Bakterije riba iz creskoga jezera Vrana. Magistarski rad, pp
108. Sveučilište u Zagrebu, Zagreb.
143. Topić Popović N, Čoţ-Rakovac R, Strunjak-Perović I (2007) Commercial
phoenotypic tests (API 20E) in diagnosis of fish bacteria. Vet Med 52 (2), 49-53.
144. Topić Popović N, Kazačinski L, Strunjak-Perović I, Čoţ-Rakovac R, Jadan M,
Cvrtila-Fleck Ţ, Barišić J (2012) Fatty acid and proximate composition of bluefin tuna
(Thunnus thynnus) muscle with regard to plasma lipids. Aquatic Res 43: 722-729.
145. Topić Popović N, Srebočan E, Čoţ-Rakovac R, Hacmenjek M, Strunjak-Perović I,
Jadan M (2008) Blood biochemistry of captive Atlantic bluefin tuna Thunnus thynnus
farmed in Adriatic Sea. Journal of Applied Ichthyology. 24, 614-616.
146. Topić Popović N, Strunjak-Perović I, Čoţ-Rakovac R, Hacmenjak M (2006) Plasma
metabolites and enzymes of bluefin tuna Thunnus thynnus and liver histology. Period
Biolog 108: 2, 127-131.
147. Topić Popović N, Strunjak-Perović I, Sauerborn Klobučar R, Barišić J, Babić S, Jadan
M, Kepec S, PKazazić S, Matijatko V, Ber Ljubić B, Car I, Repec S, Stipaničev D,
Klobučar GIV, Čoţ-Rakovac R (2015). Inpact of treated wastewater on organismic
biosensors at various levels of biological organization. Sci Total Environ 23-37.

109
148. Truhaut R (1977) Ecotoxicology: objectives, principles and perspectives. Ecotoxicol
Environ Saf 1(2): 151-73.
149. Udroiu I (2006) The micronucleus test in piscine erythrocytes. Aquatic Toxicology 79,
201-204.
150. Ural MS (2013) Chlorpyrifos-induced changes in oxidant/antioxidant status and
haematological parameters of Cyprinus carpio carpio: Ameliorative effect off
lycopene. Chemosphere 90: 2059-2064.
151. USFWS, U.S.Fish & Wildlife Service (2012) Federal and state endangered and
threatented species expenditures.
152. Van Gastel CA, Van Brummelen TC (1994) Incorporation of the biomarker concept in
ecotoxicology cells for a redefinition of terms. Ecotoxicology vol 5, pp 217-225.
153. Van Ngan P, Gomes V, Passos MJACR, Ussami KA, Campos DYF, da Silva Rocha
AJ, Pereira BA (2006) Biomonitoring of the genotoxic potential (micronucleus and
erythrocyte nuclear abnormalities assy) of the Admiralty Bay water surrounding the
Brazilian Antarctic Research Station "Comandante Rerraz", King George Island. Polar
Biol 30: 209-217.
154. van Straalen N (2003) Ecotoxicology becomes stress ecology. Environmental Science
& Technology 37: 324A-330A.
155. Varela AR, Manaia CM (2013) Human health implications of clinically relevant
bacteria in wastewater habitats. Enivironmental Science and Pollution Research, 20, 6,
3550-3569.
156. Wedemeyer GA, Yastuake WT (1977) Clinical methods for the assessment of the
effects of environmental stress on fish health. Technical Papers of the U.S. Fish and
Wildlife Service, Washington, DCop.1-18.
157. WHO (1993) Environmental health criteria 155: Biomarkers and risk assessment:
concepts and principles. World Health Organisation, Geneva.
158. WHO (2006) Wastewater and Excreta Use in Aquaculture (160 pp.). World Health
Organisation, Geneva.
159. Wiesmann U, Su Choi I, Dombrowki EM (2007) Fundamentals of Biological
Wastewater Treatment. Weinheim: Wiley-VCH Verlag GmbH&Co. KGaA.
160. Xue B, Zhang R, Wang Y, Liu X, Li J, Zhang G (2013) Antibiotic contamination in a
typical developing city in south Cihna: occurence and ecological risks in the Yonjang
River impacted by tributary discarge and anthropogenic activiteis. Ecotoxicol Environ
Saf 92, 229-236.

110
161. Zelikoff JT (1993) Metal pollution-induced immunomodulation in fish. Annu rev Fish
Dis 3:305-325.
162. Zhang Q, Wu J, Chen S, Feng Y (2014) Influences of organic pollutants in water on
antioxidant enzyme in zebrafish. J. Chem. Pharm. Res.6: 4, 1014-1021.
163. Ţarković N (2000) Mechanismus der Tumorentstehung. Pharmazeutische Zeitung,
145: 239-286.
164. Ţarković N, Lončarić I, Čipak A, Jurić G, Wonisch W, Borović S, Weag G, Vuković
T, Ţarković K (2001) Patofiziološke značajke sekundarnih glasnika slobodnih radikala
i oksidativni stres. U: Oksidativni stres i djelotvornost antioksidansa. Medicinska
naklada, Zagreb.
165. Ţikic V, Štajn AS, Ognjanović BJ, Palovic SZ, Saičić ZS (1997) Activities of
superoxide dismutasae and catalase in erythrocytes and transaminases in the plasma of
carps (Cyprinus carpio) exposed to cadmium. Physiol Res 46 (5), 391-394.

111
9. ŢIVOTOPIS
Slavko Kepec je roĎen 01. listopada 1958. godine u Virovitici. Osnovnu školu završio je u
Suhopolju, a gimnaziju u Virovitici. Diplomirao je 1982. godine na Prirodoslovno-
matematičkom fakultetu Sveučilišta u Zagrebu, smjer biologija i stekao zvanje diplomirani
inţenjer biologije.
Nakon završetka fakulteta zaposlio se kao profesor biologije i kemije u Centru za odgoj i
usmjereno obrazovanje u Virovitici. Na Filozofskom fakultetu Sveučilišta u Zagrebu poloţio
je predmete iz pedagoško-psihološke skupine predmeta za rad u školi.
1985. godine zapošljava se u Gradskom društvu crvenog kriţa u Virovitici na poslovima
provoĎenja akcija dobrovoljnog darivanja krvi i tečajeva iz prve pomoći za vozače.
Od 1991. godine do danas zaposlen je u poduzeću Virkom d.o.o. Virovitica na radnom mjestu
voditelj laboratorija. Poslovi koje obavlja odnose se na pripremu i kontrolu kakvoće vode za
piće. Radi na poslovima praćenja rada ureĎaja za biološko pročišćavanje otpadnih voda i
provoĎenju analiza otpadnih voda.
Popis radova:
1. Kepec S (1997) Prikaz rezultata ispitivanja vode za piće, otpadnih voda i ureĎaja za
biološko pročišćavanje otpadnih voda sustava vodoopskrbe i odvodnje Virovitice,
Hrvatska vodoprivreda, Zagreb
2. Kepec S (1998) Praćenje učinkovitosti uklanjanja ţeljeza i amonijaka, Hrvatska
vodoprivreda, Zagreb
3. Kepec S (2000) Osvrt na kakvoću vode za piće na području bivše općine Virovitica,
Hrvatska vodoprivreda, Zagreb
4. Kepec S (2002) Rezultati ispitivanja kakvoće vode za pi8će u javnom vodoopskrbnom
sustavu Virovitice, Zbornik radova znanstveno-stručnog skupa „Voda i javna
vodoopskrba“, Mlini
5. Kepec S (2002) Prikaz tehnološke linije gradskog dijela ureĎaja za biološko
pročišćavanje otpadnih voda s osvrtom na strukturu troškova njegovog rada, Zbornik
radova VIII. Stručno poslovnog skupa Hrvatske grupacije vodovoda i kanalizacije,
Šibenik

112
6. Kepec S, Markić M, Beslić M, Majnarić B, Siposz L (2006) Analiza rada samoispirnih
filtara u Hrvatskoj. Zbornik radova stručno-poslovnog skupa Suvremene tehnologije i
ureĎaji za pročišćavanje otpadnih voda, Poreč
7. Topić Popović N*, Strunjak-Perović I, Sauerborn Klobučar R, Barišić J, Babić S, Jadan
M, Kepec S, P.Kazazić S, Matijatko V, Beer Ljubić B, Car I, Repec S, Stipaničev D,
Klobučar GIV, Čoţ-Rakovac R (2015) Impact of treated wastewater on orgnismic
biosensors at various levels of biological organization, Science of the Total Environment,
538: 23-37
8. Topić Popović N, Benussi Skukan A, Dizidara P, Strunjak-Perović I, Kepec S, Barišić J*,
Čoţ-Rakovac R (2015) Prediction of Listeria monocitogenes growth as function of
environmental factors, Acta Alimentaria, Vol.44 (3), pp. 443-453
9. Topić Popović N*, P.Kazazić S, Strunjak-Perović I, Barišić J, Sauerborn-Klobučar R,
Kepec S, Čoţ-Rakovac R (2015) Detection and diversity of aeromonads from treated
wastewater and fish inhabiting effluent and downstream waters, Ecotoxicology and
Environmental Safety 120, 235-242

113
10. POPIS SLIKA I TABLICA
10.1. Popis slika
Slika 1. Projektirani i izgraĎeni UreĎaj za biološko pročišćavanje otpadnih voda grada
Virovitice (UBP)
19
Slika 2. Prikaz lokacija uzorkovanja 21
Slika 3. Babuška (Carassius gibelio), ulovljena 08.07.2014. u Ţupanijskom kanalu
nizvodno
Slika 4. Koncentracija amonijaka, nitrita, nitrata i ukupnog dušika u pročišćenoj otpadnoj
vodi i vodotocima (Slika 2., Lokacije uzorkovanja 1-9)
39
Slika 5. Koncentracija ortofosfata i ukupnog fosfora u pročišćenoj otpadnoj vodi i
vodotocima (Slika 2., Lokacije uzorkovanja 1-9)
40
Slika 6. Koncentracija teških metala u muljevima 42
Slika 7. Škrge babuški ulovljenih u proljeće (a, b, c), ljeto (d, e, f) i jesen (g, h, i) 45
Slika 8. Tkivo bubrega babuški ulovljenih u ljeto (a, b) i jesen (c, d) u kanalu Manteč (a,c)
i Ţupanijskom kanalu (b, d)
46
Slika 9. Tkivo jetre babuški ulovljenih u ljeto (a, b) i jesen (c, d), u kanalu Manteč (a,c) i
Ţupanijskom kanalu (b, d)
47
Slika 10. Tkivo slezene babuški ulovljenih u proljeće , u kanalu Manteč (a,b) i Ţupanijskom
kanalu (c)
48
Slika 11. Biokemijski parametri i parametri oksidacijskog stresa; (log10 osnovne skale) u
plazmi babuški po sezonama prema mjestu ulova (Slika 2., lokacije uzorkovanja
1, 8, 9)
50
Slika 12. Učestalost eritrocita sa nepravilnim jezgrama (‰) kod babuške 51
Slika 13. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h tretmana izlaganja otpadnoj vodi uzorkovanoj u proljeće 2014. god.
52
Slika 14. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24h tretmana izlaganja otpadnoj vodi uzorkovanoj u ljeto 2014.god.
53
Slika 15. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h tretmana izlaganja otpadnoj vodi uzorkovanoj u zimu 2014. god
53
Slika 16. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u proljeće 2014.
g.
54
Slika 17. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u ljeto 2014. g
55
Slika 18. TBARS koncentracija u tkivu gujavice E. fetida nakon izlaganja uzorcima otpadne
vode (nerazrijeĎenim, razrijeĎenim 100 x i 10.000 x) uzorkovane u jesen 2014. g.
56
Slika 19. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja
uzorkovanog u proljeće
57
Slika 20. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja
uzorkovanog u ljeto
57

114
Slika 21. Akumulacija fluorescentnog supstrata Rodamina 123 u tkivu gujavice E. fetida
nakon 24 h, 48 h i 7 d izlaganja dvjema koncentracijama otpadnog mulja
uzorkovanog u jesen
58
Slika 22. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u proljeće
59
Slika 23. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u ljeto
59
Slika 24. Koncentracija TBARS-a u tkivu gujavice E. fetida nakon 24 h, 48 h i 7 d izlaganja
dvjema koncentracijama otpadnog mulja uzorkovanog u jesen
60
Slika 25. Rezultati mikrobioloških ispitivanja vode i mulja 65
Slika 26. Postotak izolata MALDI-TOF MS/API 20E u uzorcima voda i mulja - proljeće 68
Slika 27. Postotak izolata u uzorcima vode i mulja MALDI-TOF SE/API 20E - ljeto 69
Slika 28. Postotak izolata u uzorcima vode i mulja MALDI-TOF SE/API 20E - jesen 71
Slika 29. Prikaz mikrobiološke analize briseva s organa riba; K-koţa, Š-škrge 72
Slika 30. Postotak izolata briseva s organa riba u proljeće (API 20E i MALDI-TOF MS) 73
Slika 31. Postotak izolata s organa riba (MALDI-TOF MS/API 20 E 74
Slika 32. Postotak izolata briseva s organa riba u jesen (API 20E i MALDI-TOF MS) 75
Slika 33. Osjetljivost prema antimikrobnim lijekovima u vodi i mulju 78
Slika 34. Osjetljivost prema antimikrobnim lijekovima bakterija poraslih iz briseva organa
riba u potoku OĎenica (Slika 2., lokacija uzorkovanja 1) prema sezonama i vrsti
tkiva
79
Slika 35. Osjetljivost prema antimikrobnim lijekovima sojeva bakterija poraslih iz briseva
organa riba na lokaciji uzorkovanja 8 (Ţupanijski kanal) prema Slici 2.
80
Slika 36. Osjetljivost prema antimikrobnim lijekovima sojeva bakterija poraslih iz briseva
organa riba na lokaciji uzorkovanja 9. (Ţupanijski kanal nizvodno) prema Slici 2.
81

115
10.2. Popis tablica
Tablica 1. Prikaz uzoraka prema lokacijama uzorkovanja 20
Tablica 2. Biometrijski podaci riba prema terminu i mjestu ulova 27
Tablica 3. Temperatura vode, suspendirana tvar, pH i elektroprovodljivost 36
Tablica 4. Otopljeni kisik, kemijska potrošnja kisika (KPKCr i KPKMn) i biološka
potrošnja kisika (BPK-5)
38
Tablica 5. Koncentracija ţeljeza, kadmija, kroma, ţive, olova, nikla, cinka i bakra u
uzorcima voda (μg/L)
41
Tablica 6. Koncentracija odreĎivanih farmaceutika u vodi (ng/L) i sedimentu (ng/g) 43
Tablica 7. Vrijednosti hematokrita krvi babuške kroz sezone 49
Tablica 8. Rezultati analize mutagenog potencijala uzoraka otpadne vode
uzorkovane u proljeće Ames bakterijskim testom
61
Tablica 9. Rezultati analize mutagenog potencijala uzoraka otpadne vode
uzorkovane u ljeto Ames bakterijskim testom
62
Tablica 10. Rezultati analize mutagenog potencijala uzoraka otpadne vode
uzorkovane u jesen Ames bakterijskim testom
63
Tablica 11. Prisutnost Pseudomonas aeruginosa, Listeria monocytogenes i
Salmonella sp. u uzorcima vode i mulja
66
Tablica 12. Postotak izolata u uzorcima vode (1-9) i mulja (10-11) u proljeće (API
20E i MALDI-TOF MS)
68
Tablica 13. Postotak izolata u vode (1-9) i mulja (10-11) u ljeto (API 20E i MALDI-
TOF MS)
69
Tablica 14. Postotak izolata u vode (1-9) i mulja (10-11) u jesen (API 20E i MALDI-
TOF MS
70
Tablica 15. Postotak izolata briseva s organa riba u proljeće (API 20E i MALDI-TOF
MS)
73
Tablica 16. Postotak izolata briseva s organa riba u ljeto (API 20E i MALDI-TOF
MS)
74
Tablica 17. Postotak izolata briseva s organa riba u jesen (API 20E i MALDI-TOF
MS)
75





























