Embed Size (px)
Citation preview

CHANTAT NYIRANSENGIYUMVA
EFFET DE DIFFÉRENTS ÉLÉMENTS MINÉRAUX SUR LA CROISSANCE ET LE DÉVELOPPEMENT DU CHAMPIGNON
HELMINTHOSPORIUM SOLANI, AGENT RESPONSABLE DE LA GALE ARGENTÉE DE LA POMME DE TERRE
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en Biologie Végétale pour l'obtention du grade de Maître es Sciences (M.Sc.)
DEPARTEMENT DE PHYTOLOGIE FACULTÉ DES SCIENCES DE L'AGRICULTURE ET DE L'ALIMENTATION
UNIVERSITÉ LAVAL QUÉBEC
2007
© Chantai Nyiransengiyumva, 2007

RESUME
Cette étude a évalué in vitro l'effet de différents éléments minéraux sur la croissance
et le développement de Helminthosporium solani, agent responsable de la gale argentée de
la pomme de terre. Les résultats obtenus ont démontré que la source et la concentration
d'éléments minéraux étudiés avaient une influence sur la croissance mycélienne et la
production de conidies chez ce champignon. L'augmentation des concentrations d'azote et
de calcium dans le milieu de culture a généralement diminué la croissance mycélienne mais
stimulé la production de conidies viables, tandis que l'augmentation des concentrations de
phosphore et de potassium a eu l'effet contraire. À la lumière des résultats obtenus, il
apparaît évident que ces travaux pourraient trouver des applications dans la mise au point
de milieux de culture favorables au développement de H. solani. Ils pourraient également
présenter un intérêt pour l'élaboration de substrats répressifs envers la gale argentée de la
pomme de terre.

m
AVANT-PROPOS
J'ai le plaisir d'exprimer ma plus profonde gratitude au Dr Russell Tweddell pour
avoir accepté la direction de ma recherche, pour la confiance qu'il m'a accordée, pour sa
rigueur, pour le temps qu'il a consacré à la lecture et à la correction du présent travail et
pour avoir fait preuve d'une patience toute particulière tout au long de ma maîtrise.
J'adresse mes sincères remerciements au Dr Tyler Avis, co-directeur de ma recherche
pour ses encouragements, ses corrections et suggestions. Les nombreuses discussions que
nous avons eues et les conseils qu'il m'a donnés ont mené à l'aboutissement de ce travail.
Je tiens à le remercier de l'aide précieuse et du temps qu'il a consacré à la réalisation du
présent mémoire. Son encadrement si précieux dont j'ai bénéficié, son esprit d'ouverture,
sa bonne humeur et l'atmosphère tranquille dans laquelle il m'a permis de travailler ont été
très appréciés.
Je souhaite remercier le Dr Pierre-Mathieu Charest, Directeur du programme de
Biologie végétale pour sa grande générosité, ses conseils et ses encouragements, qu'il
trouve ici l'expression de mon profond respect.
Je tiens à remercier énormément le Dr Carole Martinez pour son aide dont j 'ai
bénéficié au début de ma maîtrise.
Je tiens à souligner l'assistance technique réalisée par Mélanie Michaud, Sandra Le
Toumeux et Anaïs Le Goaziou, je les en remercie profondément.
J'adresse également tous mes remerciements au gouvernement rwandais et au
gouvernement du Québec pour le financement de mes études.
Une pensée va à l'endroit de tous mes collègues avec qui j 'ai partagé le laboratoire, le
bureau et le repas, à savoir Valérie, Jean-Pierre, Sophia et Joseph.
Je ne saurai comment remercier suffisamment mon conjoint Jean-Bosco Ntagungira
et ma fille Vistella-Gaju pour leur patience, leur compréhension, leur soutien moral et leur
amour.

IV
TABLE DES MATIERES
RÉSUMÉ.... : ii AVANT-PROPOS iii TABLE DES MATIÈRES iv LISTE DES FIGURES vi LISTE DES TABLEAUX viii
INTRODUCTION GÉNÉRALE I
CHAPITRE! 3 REVUE DE LITTÉRATURE 3
1.1 Généralités sur la culture de la pomme de terre 3 1.1.1 Origine et systématique 3 1.1.2 Importance de la pomme de terre 3 1.1.3 Principales maladies de la pomme déterre 3
1.2 Généralités sur la gale argentée de la pomme déterre 4 1.2.1 Agent pathogène responsable 4
1.2.1.1 Taxonomie 4 1.2.1.2 Description ; 4 1.2.1.3 Hôte 5
1.2.2 Symptômes de la gale argentée 5 1.2.3 Cycle et facteurs de développement de la gale argentée 7
1.2.3.1 Cycle de développement de la gale argentée.. 7 1.2.3.2 Facteurs de développement de la gale argentée 8
1.2.3.2.1 Semence et sol 8 1.2.3.2.2 Délai défanage-récolte 8 1.2.3.2.3 Conditions d'entreposage 8
1.2.4 Importance économique de la gale argentée 9 1.2.5 Moyens de lutte 9
1.2.5.1 Lutte chimique 9 1.2.5.2 Lutte génétique 10 1.2.5.3 Lutte biologique 10 1.2.5.4 Pratiques culturales 11
1.3 Éléments essentiels aux champignons 11 1.3.1 Carbone 11 1.3.2 Azote 12 1.3.3 Éléments inorganiques 12
1.3.3.1 Macro-éléments 13 1.3.3.2 Micro-éléments 13
1.3.4 Vitamines et facteurs de croissance 14

1.4 Effet de l'azote, du phosphore, du potassium, du calcium et du magnésium sur le développement de différentes maladies des cultures 14
1.4.1 Azote 14 1.4.2 Phosphore 17 1.4.3 Potassium 18 1.4.4 Calcium..... 18 1.4.5 Magnésium 19
1.5 Effet des éléments minéraux sur le développement de la gale argentée de la pomme déterre 20
1.6 Hypothèse et objectifs de recherche 20 1.6.1 Hypothèse de recherche 20 1.6.2 Objectifs de recherche 20
CHAPITRE 2 21 MATÉRIEL ET MÉTHODES 21
2.1 Helminthosporium solani 21
2.2 Effet des éléments minéraux sur la croissance et le développement de H. solani 21 2.2.1 Détermination de la croissance mycélienne 23 2.2.2 Détermination du nombre de conidies produites 23 2.2.3 Détermination du nombre de conidies viables 23
2.3 Dispositif expérimental et analyse statistique 24
CHAPITRE 3 '. 25 RÉSULTATS 25
3.1 Effet des éléments minéraux sur la croissance mycélierme de H. solani 25 3.1.1 Effet de l'azote sur la croissance mycélienne de//. solani 25 3.1.2 Effet du phosphore sur la croissance mycélienne de H. solani 28 3.1.3 Effet du potassium sur la croissance mycélienne de H. solani 30 3.1.4 Effet du calcium sur la croissance mycélienne de if. solani 32 3.1.5 Effet du magnésium sur la croissance mycélierme de H. solani 34
3.2 Effet des éléments minéraux sur le nombre de conidies produites 37
3.3 Effet des éléments minéraux sur le nombre de conidies viables produites 40
CHAPITRE 4 44 DISCUSSION 44
CONCLUSION GÉNÉRALE.... 51 RÉFÉRENCES BIBLIOGRAPHIQUES 52

VI
LISTE DES FIGURES
Figure 1. Helminthosporium solani et gale argentée de la pomme de terre. A. H. solani sur milieu de culture V8, B. H. solani sur gélose nutritive, C. Conidiophores et conidies de H. solani, D. Conidies, E. Symptômes de la gale argentée sur pomme de terre rouge, F. Symptômes de la gale argentée sur pomme de terre blanche 6
Figure 2. Cycle de la gale argentée 7 Figure 3. Effet de la concentration d'azote sous forme de NaNOs sur la croissance
mycélienne de Helminthosporium solani 25 Figure 4. Effet de la concentration d'azote sous forme de NH4CI sur la croissance
mycélienne de Helminthosporium solani 26 Figure 5. Effet de la concentration d'azote sous forme de NaNOi sur la croissance
mycélierme de Helminthosporium solani 26 Figure 6. Effet de la concentration d'azote sous forme de NH4NO3 sur la croissance
mycélienne de Helminthosporium solani 27 Figure 7. Effet de la concentration d'azote sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation... 28 Figure 8. Effet de la concentration de phosphore sous forme de Na2HP04-7H20 sur la
croissance mycélienne de Helminthosporium solani.... 29 Figure 9. Effet de la concentration de phosphore sous forme de Na3P04'12H20 sur la
croissance mycélienne de Helminthosporium solani ; 29 Figure 10. Effet de la concentration de phosphore sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurées après huit semaines d'incubation. 30 Figure 11. Effet de la concentration de potassium sous forme de KCl sur la croissance
mycélienne de Helminthosporium solani 31 Figure 12. Effet de la concentration de potassium sous forme de K2SO4 sur la croissance
mycélienne de Helminthosporium solani 31 Figure 13. Effet de la concentration de potassium sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation... 32 Figure 14. Effet de la concentration de calcium sous forme de CaCl2-2H20 sur la
croissance mycélienne de Helminthosporium solani 33 Figure 15. Effet de la concentration de calcium sous forme de CaS04-2H20 sur la
croissance mycélienne de Helminthosporium solani 33 Figure 16. Effet de la concentration de calcium sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation... 34 Figure 17. Effet de la concentration de magnésium sous forme de MgS04-7H20 sur la
croissance mycélierme de Helminthosporium solani 35 Figure 18. Effet de la concentration de magnésium sous forme de MgCl2-2H20 sur la
croissance mycélienne de Helminthosporium solani 35 Figure 19. Effet de la concentration de magnésium sous différentes formes sur la
croissance mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation 36
Figure 20. Effet de la concentration d'azote sous différentes formes sur le nombre de conidies produites par le champignon Helminthosporium solani 38
Figure 21. Effet de la concentration de calcium sous différentes formes sur le nombre de conidies produites par le champignon Helminthosporium solani 38

vu
Figure 22. Effet de la concentration de magnésium sous différentes formes sur le nombre de conidies produites par le champignon Helminthosporium solani 39
Figure 23. Effet de la concentration de phosphore sous différentes formes sur le nombre de conidies produites par le champignon Helminthosporium solani 39
Figure 24. Effet de la concentration de potassium sous différentes formes sur le nombre de conidies produites par le champignon Helminthosporium solani 40
Figure 25. Effet de la concentration d'azote sous différentes formes sur le nombre de conidies viables produites par le champignon Helminthosporium solani 41
Figure 26. Effet de la concentration de phosphore sous différentes formes sur le nombre de conidies viables produites par le champignon Helminthosporium solani 42
Figure 27. Effet de la concentration de potassium sous différentes formes sur le nombre de conidies viables produites par le champignon Helminthosporium solani 42
Figure 28. Effet de la concentration de calcium sous différentes formes sur le nombre de conidies viables produites par le champignon Helminthosporium solani 43
Figure 29. Effet de la concentration de magnésium sous différentes formes sur le nombre de conidies viables produites par le champignon Helminthosporium solani 43

via
LISTE DES TABLEAUX
Tableau 1. Effet de l'azote sur différentes maladies des cultures 14 Tableau 2. Concentrations d'azote, de phosphore, de potassium, de calcium et de
magnésium testées dans cette étude 22 Tableau 3. Sources d'éléments minéraux testées dans cette étude 22 Tableau 4. Effet de l'azote sur le champignon Helminthosporium solani 44 Tableau 5. Effet du phosphore sur le champignon Helminthosporium solani 47 Tableau 6. Effet du potassium sur le champignon Helminthosporium solani 48 Tableau 7. Effet du calcium sur le champignon Helminthosporium solani 49 Tableau 8. Effet du magnésium sur le champignon Helminthosporium solani 49

INTRODUCTION GENERALE
La pomme de terre {Solarium tuberosum L.) est l'une des cultures légumières les plus
importantes au Canada, occupant en 2005 une superficie de 163 135 ha, pour une valeur
totale à la ferme estimée à 920,3 millions de dollars (Statistique Canada, 2007). Au cours
de cette même année, la production canadienne de pommes de terre a atteint 4 386 584
tonnes dont 488 709 tonnes au Québec, soit 11,1 % de la production canadienne (Statistique
Canada, 2007).
La culture de la pomme de terre est limitée par de nombreuses maladies fongiques
parmi lesquelles, la gale argentée {Helminthosporium solani Dur. & Mont.) montre une
recrudescence depuis quelques années. Affectant uniquement le tubercule au niveau du
périderme, cette maladie se caractérise par des taches d'aspect argenté ou bronzé
(Errampalli et ai, 2001). Longtemps considérée comme une maladie d'importance
secondaire, la gale argentée est devenue une maladie économiquement importante suite au
développement de souches de H. solani résistantes au thiabendazole (Hide et al, 1988), à
l'absence de cultivars résistants et aux nouvelles exigences esthétiques du marché
(Errampalli et al, 2001). Actuellement, il n'existe aucune méthode permettant de lutter
efficacement contre cette maladie.
Dans ce contexte, une étude récente menée au Centre de recherche en horticulture
(CRH) de l'Université Laval a montré pour la première fois la présence au Québec de sols
suppressifs envers la gale argentée de la pomme de terre (Martinez et al, 2002). Cet effet
suppressif pourrait selon cette étude être attribuable aux propriétés chimiques du sol. À cet
effet, plusieurs études ont montré que la sévérité de certaines maladies des plantes cultivées
peut être affectée par certains éléments minéraux essentiels à la croissance de la plante
(Ruber et Watson, 1974 ; Huber, 1978 ; 1980) et que ces derniers peuvent être exploités
pour limiter le développement d'une grande variété de maladies (Engelhard, 1989). Des
exemples remarquables sont ceux des flétrissements fusariens causés par Fusarium
oxysporum avec des travaux qui datent des armées 1920 et qui décrivent l'effet bénéfique
de l'utilisation des amendements en chaux (Jones et al, 1989). Le développement d'un
grand nombre de maladies de la pomme de terre [gale commune {Streptomyces scabies),
rhizoctone brun (Rhizoctonia solani), gale poudreuse (Spongospora subterranea),

verticilliose (Verticillium dahliae et V. albo-atrum), altemariose {Alternaria solani et A.
alternata), brûlure tardive {Phytophthora infestans)] est affecté par les éléments minéraux
présents dans le sol (Lambert et al., 2005). L'effet de ces éléments sur le développement
des maladies peut résulter de changements structuraux ou physiologiques au niveau de la
plante, d'une augmentation de l'activité des antagonistes microbiens ou d'une toxicité
directe envers l'agent pathogène (Lambert et al., 2005). Ainsi, la manipulation ou la
modification des propriétés chimiques du sol apparaît comme une pratique culturale
intéressante pour le contrôle des maladies des plantes (Huber, 1989).
Dans le présent travail, une étude in vitro a été réalisée afin d'évaluer l'effet de
l'azote, du phosphore, du potassium, du calcium et du magnésium sur la croissance
mycélieime et le nombre de conidies total et viables produites par le champignon H. solani.
Les résultats obtenus pourront servir aux travaux fiaturs destinés à élaborer des plans
spécifiques de gestion des propriétés du sol dans la culture de la pomme de terre pour
mieux lutter contre le développement de la gale argentée.
Ce travail comporte quatre chapitres. Le premier chapitre expose la revue de
littérature concernant différents aspects de la gale argentée et le rôle des éléments minéraux
dans la lutte aux maladies des plantes. Le deuxième chapitre décrit le matériel et les
méthodes utilisés lors de ces travaux. Les chapitres 3 et 4 présentent les résultats obtenus et
la discussion, respectivement.

CHAPITRE 1
REVUE DE LITTÉRATURE
I.l Généralités sur la culture de la pomme de terre
1.1.1 Origine et systématique
La pomme de terre est une plante herbacée vivace qui appartient au règne Plantae,
division Magnoliophyta, classe Magnoliopsida, ordre Solanales, famille Solanaceae, genre
Solarium et espèce tuberosum. Elle est originaire de l'Amérique du sud et fut introduite en
Europe à la fin du 16 ""® siècle par deux portes d'entrée différentes, la première étant
l'Espagne vers 1570 et la seconde les îles Britanniques entre 1588 et 1593 (Spire et
Rousselle, 1996). Introduite de façon commerciale au Canada avant 1623, elle ftit cultivée
pour la première fois en Nouvelle-Ecosse, et au fil des ans, la culture s'est étendue vers
l'ouest du pays (Boiteau, 1996).
1.1.2 Importance de la pomme de terre
La pomme de terre est une culture légumière économiquement importante. Au
Canada, elle occupait en 2005 une superficie de 163 135 ha pour une production de 4 386
584 tormes. Le Québec a produit la même année 488 709 tonnes, soit 11,1 % de la
production canadienne (Statistique Canada, 2007). Actuellement, la pomme de terre est une
culture très importante dans l'alimentation humaine et elle occupe la quatrième position de
la production mondiale après la blé, le maïs et le riz (Randall, 1993). Le tubercule de
pomme de terre contient une quantité relativement importante de potassium, de fer, d'iode,
de vitamines (surtout la vitamine C), de fibres, de protéines et de glucides mais elle est
pauvre en calcium et en sodium (Burton, 1966).
1.1.3 Principales maladies de la pomme de terre
La pomme de terre est affectée par de nombreuses maladies fongiques, bactériennes
et virales qui affectent la plante en totalité ou en parties (racines, tiges, feuilles et
tubercules) pendant la phase de culture au champ et/ou pendant la phase de conservation
des tubercules (Gaucher, 1998). Les principales maladies fongiques sont la brûlure tardive

(P. infestans), le rhizoctone brun {R. solani), la gale argentée (H. solani), la dartrose
(Colletotrichum coccodes), la pourriture sèche (Fusarium sambucinum), l'altemariose {A.
solani, A. alternata), la pourriture à sclérotes {Sclerotinia sclerotium), la verticilliose
{Verticillium spp.), la pourriture aqueuse {Pythium ultimum), la pourriture rose {P.
erythroseptica) et la gale poudreuse (S. subterranea). Les principales maladies bactériennes
sont la jambe noire et la pourriture molle {Erwinia carotovora ssp. carotovora, E.
carotovora ssp. atroseptica), la gale commune (Streptomyces scabies) et la pourriture brune
(Ralstonia solanacearum). Les principales maladies virales sont les mosaïques bénignes
[virus A, S et X de la pomme de terre (VAP, VSP, VXP)], la mosaïque rugueuse [virus Y
de la pomme de terre (VYP)], l'enroulement de la pomme de terre [virus de l'enroulement
de la pomme de terre (VEP)] et la filosité (viroïde des filosités des tubercules de la pomme
de terre) (Gaucher, 1998).
1.2 Généralités sur la gale argentée de la pomme de terre
1.2.1 Agent pathogène responsable
1.2.1.1 Taxonomie
Harz a nommé pour la première fois en Autriche en 1871, le champignon pathogène
responsable de la gale argentée "Dematum atrovirens". Par la suite, le nom du champignon
a été modifié au gré des variations de la classification: Spondylocladium atrovirens (Harz,
1871) Harz ex Saccardo, 1886), puis Branchysporium solani, Cladosporium abietium,
Helminthosporium atrovirens (Ellis, 1961) pour arriver à sa dénomination actuelle H.
solani (Durieu et Montagne, 1849). H. solani appartient au règne Mycota, division
Eumycota, subdivision Deuteromycotina, classe Hyphomycetales, ordre Moniliales, famille
Dematiaceae, genre Helminthosporium et espèce solani (Alexopoulos et Mims, 1979).
1.2.1.2 Description
Sur milieu de culture artificiel, H. solani a une teinte hyaline à brune (Figures la et b)
avec des hyphes d'un diamètre variant de 1 à 8 |am au point d'origine des conidiophores.
Les conidiophores croissent seuls ou en bouquets et, dans la plupart des cas, en position
terminale et latéralement à l'hyphe (Figure le). Ils sont droits, simples, lisses et de couleur

brune (Ellis, 1961). Les conidies sont ellipsoïdales, lisses et de couleur variant d'hyaline à
brune (Figure Id). Elles ont une longueur de 24 ^m à 85 |im et possèdent 2 à 8
pseudosepta. Elles sont de forme effilée d'une largeur de 7 |im à 11 ^m à la base et de 2
\xm à leur sommet (Ellis, 1961).
1.2.1.3 Hôte
Parmi les plantes cultivées, H. solani est inféodé uniquement à la culture de la
pomme de terre {S. tuberosum) et le tubercule est le seul organe parasité par l'agent
pathogène (Kamara et Huguelet, 1972). Sa croissance et sa fructification sur le tubercule
sont étroitement liées aux conditions de température et d'humidité (Jouan et al, 1974). Sa
croissance in vitro dépend du pH et de la composition du milieu de culture (Singh, 1973).
1.2.2 Symptômes de la gale argentée
Les symptômes se manifestent à la surface du tubercule sous forme de taches
circulaires à contour irrégulier d'aspect bronzé chez les cultivars à peau rouge (Figure le)
ou argenté chez les cultivars à peau blanche (Figure If). L'aspect argenté ou bronzé
caractéristique serait attribuable au fait que les cellules du périderme se décollent et
renferment une mince couche d'air (Hunger et Mcintyre, 1979 ; Heiny et Mcintyre, 1983).
Au niveau de ces taches, on observe de minuscules ponctuations noires correspondant aux
fructifications du champignon (Gaucher, 1998). Si à la récolte, l'intensité de la maladie est
plutôt faible, c'est au cours de la période de conservation des tubercules que la maladie
s'aggrave, lorsque les conditions de température, d'humidité et d'aération sont favorables
au développement du champignon (Gaucher, 1998). Les taches s'accroissent en nombre et
en taille et confluent en de larges plages qui peuvent, dans des cas extrêmes, occuper toute
la surface du tubercule (Gaucher, 1998). La maladie ne cause pas de pertes de rendement à
la récolte, mais elle cause une perte de poids des tubercules en conservation suite à une
augmentation de la perte d'eau (Errampalli et al, 2001 ; Read et Hide, 1984). En culture,
lorsque la plante est fortement atteinte, la gale argentée peut être à l'origine d'accidents se
traduisant par un retard de végétation, voire une absence totale de levée, ce qui entraîne une
diminution de la production (Jouan et al., 1974 ; Errampalli et al, 2001).

Figure 1. Helminthosporium solani et gale argentée de la pomme de terre. A. H. solani sur
milieu de culture V8 (Tiré de Michaud, 2001), B. H. solani sur gélose nutritive, C.
Conidiophores et conidies de H. solani (Photo de Sophia Boivin), D. Conidies (Photo de
Sophia Boivin), E. Symptômes de la gale argentée sur pomme de terre rouge (Tiré de
Hervieux, 2000), F. Symptômes de la gale argentée sur pomme de terre blanche (Photo de
British Potato Council).

7
1.2.3 Cycle et facteurs de développement de la gale argentée
1.2.3.1 Cycle de développement de la gale argentée
Les symptômes de la gale argentée se développent rapidement après la plantation à
partir d'une semence infectée (Figure 2). La maladie se propage par les conidies présentes à
la surface des tubercules. Les conidies migrent généralement le long des stolons jusqu'aux
tubercules-fils et les infectent au niveau du talon (Jellis et Taylor, 1977). Les conidies
peuvent aussi survivre dans le sol et résister à l'hiver de façon saprophytique sur les débris
végétaux (Kamara et Huguelet, 1972 ; Mérida et Loria, 1994). Bien que dans certaines
conditions, 1'inoculum du sol puisse être impliqué dans la transmission de la maladie (Bisht
et Bains, 1995 ; Bains et al., 1996), le tubercule de semence constitue la principale source
d'inoculum (Burke, 1938; Santerre, 1967; Read et Hide, 1984). Les conditions qui
prévalent dans les entrepôts, particulièrement la forte humidité relative sont propices à la
croissance et la sporulation de H. solani et au développement de la gale argentée. À cet
effet, la gale argentée est souvent considérée comme une maladie d'entreposage (Rodriguez
et al., 1995). Une partie de l'infection est attribuée aux contacts physiques survenant entre
les tubercules sains et les tubercules infectés. Le déplacement et l'emballage des pommes
de terre sont d'autres modes possibles de dispersion des conidies (Secor, 2006).
Sol : transmission aux tubercules-fils
Semence infectée
Entrepôt : évolution et dissémination Déplacement et emballage dispersion
Inoculum du sol
Figure 2. Cycle de la gale argentée (Tiré de Michaud, 2001).

8
1.2.3.2 Facteurs de développement de la gale argentée
Le développement de la gale argentée dépend de plusieurs facteurs à savoir l'état
sanitaire des semences, le sol, le délai défanage-récolte et les conditions d'entreposage.
1.2.3.2.1 Semence et sol
Le tubercule de semence constitue la principale source d'inoculum (Burke, 1938 ;
Santerre, 1967 ; Read et Hide, 1984). Bains et al. (1996) ont montré que les tubercules
produits à partir de semences exemptes de gale argentée ont montré des symptômes de cette
maladie lorsque cultivés dans des champs où la culture de la pomme de terre n'a pas été
pratiquée pendant les quatre années précédentes. Ceci suggère que le champignon puisse
survivre dans le sol pendant plusieurs années. H. solani est capable de se développer et de
sporuler sur des feuilles sénescentes de différentes espèces végétales utilisées deins les
rotations avec la pomme de terre comme la luzerne (Medicago sativa), le maïs (Zea mays)
et le blé (Triticum spp.) (Mérida et Loria, 1994). Ces résultats indiquent que le parasite
puisse survivre dans le sol grâce à son activité saprophytique. Le champignon peut aussi se
développer sur les parties souterraines de la pomme de terre (tige et racines) en état de
sénescence (Jouan et al., 1974).
1.2.3.2.2 Délai défanage-récolte
La destruction du feuillage avant maturité favorise d'une façon nette l'installation du
champignon et le pourcentage de tubercules malades est d'autant plus élevé que la récolte
est effectuée tard après le défanage. Le taux d'infection par H. solani à la récolte peut varier
entre 4 et 94 % (Gaucher, 1998). Ainsi, la sévérité de la maladie est corrélée à la durée de la
période passée au champ par le tubercule (Jouan et ai, 1974).
1.2.3.2.3 Conditions d'entreposage
Au cours de l'entreposage des tubercules de pomme de terre, la gale argentée évolue
plus ou moins rapidement selon les conditions ambiantes de température et d'humidité.

L'étude sur l'action de la température sur la croissance du champignon effectuée par Jouan
et al. (1974) a montré que la température optimale de croissance de H. solani se situe entre
20 et 25°C. À 6°C, la croissance du champignon est très lente tandis que à 2°C, la
croissance est nulle. Cette température n'est toutefois pas létale (Jouan et al, 1974). La
germination des spores a lieu à une température comprise entre 2 et 35°C (Jouan et al,
1974). À une humidité relative de 100 %, le nombre de spores produites croît avec la
température dans l'intervalle de 2-20°C (Jouan et al., 1974). La fructification du
champignon est importante dans l'intervalle de 85-100 % d'humidité relative avec une
optimum situé aux alentours de 90 %. En dessous de 80 % d'humidité relative, le nombre
de spores produites baisse rapidement pour devenir nul en dessous de 55 % d'humidité
relative. Certains travaux ont montré qu'il existe une quantité importante de conidies libres
pouvant infecter les tubercules conservés dans les entrepôts de pomme de terre (Mérida et
Loria, 1994). Les conidies provenant des tubercules infectés peuvent être dispersées par le
système de ventilation et infecter par conséquent un grand nombre de tubercules
(Rodriguez er a/., 1996).
1.2.4 Importance économique de la gale argentée
La gale argentée sévit dans de nombreux pays producteurs de pommes de terre
(Elimane, 1996). Longtemps considérée comme une maladie d'importance secondaire, la
gale argentée est devenue au cours des dernières années une maladie économiquement
importante suite au développement de souches de H. solani résistantes au thiabendazole
(Hide et al, 1988), à l'absence de cultivars résistants et aux nouvelles exigences esthétiques
du marché (Errampalli et al, 2001).
1.2.5 Moyens de lutte
1.2.5.1 Lutte chimique
Le thiabendazole, seul fongicide homologué au Canada pour le traitement post
récolte des tubercules de pomme de terre, fut abondamment utilisé pendant de nombreuses
années. Depuis quelques années, des souches de H. solani résistantes à ce fongicide et au

10
thiophanate-méthyl (un autre fongicide de la famille des benzimidazoles) ont été recensées
en Angleterre (Hide et ai, 1988), aux États-Unis (Mérida et Loria, 1990 ; Rodriguez et al.,
1990 ; Secor et al., 1996), au Canada (Carnegie et al., 1994 ; Kawchuck et al., 1994 ;
Bernard et Bains, 1996) et au Québec (Platt, 1997). En conséquence, l'application post
récolte de thiabendazole ou de thiophanate-méthyl en traitement de semence ne permet plus
un contrôle efficace de la maladie.
Différentes études ont démontré que l'application en traitement de semence de
fenpiclonil (Gaucher, 1998), imazalil (Hide et al., 1994), mancozeb (Gaucher, 1998) ou
fludioxonil (Errampalli et al., 2001) permet une réduction intéressante de gale argentée. Le
mancozeb et le fludioxonil sont homologués au Canada pour le traitement des semences
(CRAAQ, 2004).
1.2.5.2 Lutte génétique
Des croisements interspécifiques avec les espèces sauvages de Solanum (S.
demissum, S. chacoense. S: aculae) ont été effectués pour améliorer la résistance aux
maladies de certains cultivars de S. tuberosum au Canada et les travaux de sélection et
d'évaluation continuent (Errampalli et al., 2001). Cependant, aucun cultivar de pomme de
terre résistant à la gale argentée n'a été identifié (Errampalli et al., 2001).
1.2.5.3 Lutte biologique
Pseudomonas corrugata (Chun et Shetty, 1994), Cephalosporium sp. (Secor et
Gudmestad, 1999), Bacilluspolymyxa (Lange et al., 1995), Pseudomonasputida (Elson et
al., 1997 ; Michaud et al., 2002 ), Nocardia globerula et Xanthomonas campestris (Elson et
al., 1997) ont montré la capacité d'inhiber le développement de H. solani sur le tubercule
de pomme de terre. De même, Alcaligenes piechaudii, Aquaspirillum autotrophicum,
Arthrobacter oxydans. Bacillus cereus, B. mycoïdes, Kocuria rosea, K. varians,
Paenibacillus pabuli, Pseudomonas chlororaphis, P. fluorescens, Rhodococcus erytropolis,
R. globulus et Streptomyces griseus (Michaud et al., 2002) ont diminué le développement

11
de la gale argentée sur le tubercule. À ce jour, aucun agent de lutte biologique n'est
homologué pour lutter contre cette maladie.
1.2.5.4 Pratiques cultiirales
Certaines pratiques culturales peuvent réduire l'incidence de la maladie, comme
l'utilisation de semences saines (Santerre, 1967 ; Secor et al., 1996), le séchage rapide des
tubercules après la récolte (Hide et Boorer, 1991 ; Firman et Allen, 1993) et la rotation des
cultures (Richard et Boivin, 1994 ; Stevick Haux, 1998). Une gestion appropriée des
conditions d'entreposage est un autre moyen de limiter le développement de la maladie
(Secor et Gudmestad, 1999).
1.3 Éléments essentiels aux champignons
Comme les autres organismes, les champignons ont besoin pour leiir croissance et
leur développement du carbone, de l'azote, des éléments inorganiques (macro- et micro
éléments), de vitaniines et de facteurs de croissance.
1.3.1 Carbone
Le carbone est un élément fondamental de structure et de fonctionnement de toutes
les cellules vivantes. Les champignons utilisent une large gamme de composés carbonés
allant de petites molécules comme les sucres (glucose, fructose, saccharose, mannose), les
acides organiques (acide acétique, acétate d'éthyl, acide oxalique, acide propionique, acide
fiimarique), les alcools (methanol, éthanol, propanol, butanol) jusqu'aux macromolécules
(protéines, lipides, polysaccharides, lignine) (Kirk, 1971 ; Perlman, 1965 ; Sestakova,
1976). Ces différents composés carbonés interviennent dans deux fonctions physiologiques
essentielles aux champignons : (1) ils fournissent le carbone nécessaire pour la synthèse des
constituants importants (hydrates de carbone, protéines, lipides, acides nucléiques), (2) et
leur oxydation fournit une source d'énergie nécessaire au fonctionnement adéquat des
processus vitaux de la cellule fongique (Garraway et Evans, 1991).

12
1.3.2 Azote
L'azote est un élément indispensable pour la croissance et le développement des
champignons. L'azote est nécessaire pour la synthèse des constituants cellulaires
d'importance capitale (acides aminés, protéines, purines, pyrimidines, acides nucléiques,
glucosamine, chitine, différentes vitamines) (Garraway et Evans, 1991). La plupart des
champignons sont capables d'utiliser les sources d'azote inorganique simples (nitrates,
nitrites, ammonium) aussi bien que les sources organiques (urée, acides aminés) (Garraway
et Evans, 1991). À l'exception des protéines, la plupart des composés azotés sont
suffisament petits pour entrer directement dans la cellule sans dégradation extracellulaire
préalable. Ainsi, la membrane cellulaire constitue le premier point au niveau duquel les
champignons peuvent exercer un contrôle d'incorporation de l'azote. Étant donné que les
nitrates et les nitrites entrent apparemment dans les cellules fongiques par diffusion, les
principaux facteurs qui affectent leur utilisation seraient ceux qui impliquent le
métabolisme et non le transport. Ainsi, l'incapacité de certains champignons à croître sur
un milieu de culture contenant des nitrates est souvent due à l'absence de l'enzyme nitrate
reductase, tandis que l'incapacité à croître sur un milieu contenant des nitrites serait plutôt
attribuable à l'absence de l'enzyme nitrite reductase ou à l'action inhibitrice des nitrites
envers certains patrons métaboliques (Garraway et Evans, 1991). L'incorporation de
l'ammonium, de l'urée et des acides aminés est contrôlée au niveau de la surface cellulaire
et au niveau intracellulaire. Ainsi, les facteurs qui affectent la forme et la charge des
molécules de transport (température, pH), la réserve de l'adénosine triphosphate (ATP)
nécessaire pour le transport actif (niveau d'oxygène, pH, source de carbone) et les systèmes
de feed-back intracellulaires (concentration de sulfure et d'azote sous forme d'acides
aminés) affectent aussi l'utilisation de ces sources (Garraway et Evans, 1991).
1.3.3 Éléments inorganiques
Un élément inorganique est considéré comme nutriment essentiel pour les
champignons lorsque son absence dans un milieu de culture conduit à la réduction de leur
croissance et de leur reproduction (Garraway et Evans, 1991). Les champignons ont des
besoins relativement faibles en éléments inorganiques (100 mg/1 pour les phosphates et
moins de 1 mg/1 pour les composés contenant le cuivre) comparativement à leurs besoins en

13
carbone et en oxygène (g/1). Les quantités des différents éléments inorganiques essentiels
pour la croissance des champignons varient selon les espèces, la composition du milieu de
culture et l'environnement (Garraway et Evans, 1991). Les éléments inorganiques sont
groupés en deux catégories : les macro-éléments qui sont nécessaires en grandes quantités
(magnésium, phosphore, potassium, soufre, calcium) et les micro-éléments, aussi appelés
éléments en trace, requis en plus petites quantités (cuivre, fer, manganèse, zinc, molybdène)
(Garraway et Evans, 1991).
1.3.3.1 Macro-éléments
Dans la liste qui suit les rôles connus des différents macro-éléments sont
présentés :
- Magnésium : intervient dans l'activation d'enzyme et dans le métabolisme de l'ATP
(adenosine triphosphate);
- Phosphore : intervient dans la formation des acides nucléiques, dans le transfert d'énergie
et dans le métabolisme intermédiaire;
- Potassium : intervient dans l'activité enzymatique, dans le métabolisme des hydrates de
carbone et dans la balance ionique;
- Soufre : intervient dans la formation des acides aminés, des vitamines et des autres
composés sulfohydriles;
- Calcium : intervient dans l'activité enzymatique, dans la structure des membranes et il
n'est pas universellement requis par tous les champignons (Griffm, 1994).
1.3.3.2 Micro-éléments
Dans la liste qui suit les rôles connus des différents micro-éléments sont
présentés :
- Cuivre : intervient dans l'activité enzymatique et dans la formation des pigments;
- Fer : intervient dans la formation des cytochromes, du hème des apoenzymes et des
pigments;
- Manganèse : intervient dans l'activité enzymatique, dans le cycle de Krebs (acide
tricarboxylique : TCA) et dans la synthèse des acides nucléiques;

14
- Zinc : intervient dans l'activité enzymatique, dans les acides organiques et autres
métabolismes intermédiaires;
- Molybdène : intervient dans l'activité enzymatique, dans le métabolisme des nitrates et la
vitamine B12 (Griffm, 1994).
1.3.4 Vitamines et facteurs de croissance
Les vitamines et les facteurs de croissance sont des composés organiques requis en
quantités minimes pour la croissance des champignons (Griffin, 1994). Les vitamines sont
généralement requises en concentration micromolaire ou moins et jouent le rôle de
coenzymes. Les vitamines généralement essentielles aux champignons sont : la thiamine
(Bi), la biotine (B7), la pyridoxine (BÔ), la riboflavine (B2), la niacine (B3), l'acide p-
aminobenzoïque, l'acide pantothénique (B5), la cyanocobalamine (B12) et l'inositol (Griffin,
1994). Les facteurs de croissance généralement utilisés par les champignons sont: les
acides aminés, les sterols, les purines et les pyrimidines (Griffin, 1994).
1.4 Effet de l'azote, du phosphore, du potassium, du calcium et du magnésium sur le
développement de différentes maladies des cultures
1.4.1 Azote
L'azote a été intensivement étudié dans la relation "nutrition de l'hôte-sévérité de la
maladie" depuis plusieurs années (Huber et Watson, 1974). L'eizote présente différents
effets selon la culture et l'agent pathogène étudié (Tableau 1).
Tableau 1. Effet de l'azote sur différentes maladies des cultures* Maladie
Maladies de semis et fonte de semis
Pourritures racinaires
Hôte
Betterave à sucre
Citronnier
Citronnier
Haricot
Haricot
Pathogène
Rhizoctonia solani
Phytophthora citrophtora Fusarium spp.
R. solani
Fusarium solani f. sp phaseoli
Forme d'azote*"
Nitrate
Diminue
Diminue
ND'=
Diminue
Diminue
Ammonium
Augmente
Augmente
Augmente
Augmente
Augmente

15
Tableau 1. Effet de l'azote sur différentes maladies des cultures^ (suite) Maladie
Pourritures racinaires
Pourriture de la tige
Piétin-verse
Pourriture racinaire
Pourriture de la tige et de la racine
Piétin-échaudage
Rhizoctone ocellé
Racine noire
Taches
Rouille méridionale
Gale commune
Flétrissement
Hôte
Petit pois
Soya
Petit pois
Maïs
Pomme de terre
Maïs
Maïs
Maïs
Pin
Pin
Blé
Blé
Blé
Tabac cultivé (Nicotiana tabacum) Coton
Pomme de terre
Blé
Blé
Betterave à sucre
Gazon
Tomate
Pomme de terre
Oeillet
Coton
Tomate
Pomme de terre
Tomate
Pathogène
Aphanomyces euteiches
Pythium ultimum
A. euteiches
Pythium spp.
Fusarium spp.
Diplodia zeae
Porta weirii
Armillaria mellea
Fusarium spp.
Helminthosporium sativum Cercosporella herpotrichoides Thielaviopsis basicola
Phymatotrichum omnivorum R. solani
Gaeumannomyces graminis Rhizoctonia cerealis
Aphanomyces cochlioides G. graminis
Sclerotium rolfsii
Streptomyces scabies
Phialophora cinerescens F. oxysporum f. sp. vasinfectum F. oxysporum f. sp. lycopersici Verticillium spp.
Forme d'azote''
Nitrate
Diminue
Augmente
Diminue
Augmente
Diminue
Augmente
Diminue
Diminue
Diminue
Diminue
Diminue
Diminue
Augmente
Diminue
Augmente
Diminue
Diminue
ND
Diminue
Augmente
Augmente
Diminue
Diminue
Augmente
Ammonium
Augmente
Diminue
Augmente
Diminue
Augmente
Diminue
Augmente
Augmente
Augmente
ND
Augmente
Augmente
Diminue
Augmente
Diminue
Augmente
Augmente
Diminue
Augmente
Diminue
Diminue
Augmente
Augmente
Diminue

16
Tableau 1. Effet de l'azote sur différentes maladies des cultures^ (suite)
Maladie
Jaunissement
Flétrissement
Chancre
Maladie de Stewart
Pourriture annulaire
Blanc
Pourriture des fruits et racines (anthracnose)
Rouille striée
Rouille de la tige
Taches septentrionales
Piriculariose
Taches chocolatées
Nematode à kystes
Nematode des racines
Chancre
Pourriture de stockage
Virus
Hôte
Chou
Tabac cultivé
Tomate
Tomate
Maïs
Pomme de terre
Blé
Tomate
Blé
Blé
Maïs
Riz
Fève
Soya
Tabac cultivé
Pêcher
Prunier
Patate sucrée
Tabac cultivé
Pomme de terre
Tabac sauvage (Nicotiana glutinosd)
Tabac cultivé
Tabac sauvage
Pomme de terre
Pathogène
F. oxysporum f. sp. conglutinans Pseudomonas solanacearum
Corynebacterium michiganense Envinia stewartii
Corynebacterium sepedonicum Erysiphe graminis f. sp. tritici Colletotrichum phomoides
Puccinia striiformis f. sp. tritici Puccinia graminis f. sp. tritici Helm inthosporium turcicum Piricularia oryzae
Botrytis fabae
Heterodora glycines
Heterodora tabacum
Xanthomonas campestris p.v. pruni
Rhizopus stolonifer
Virus Y de la pomme de terre
Virus Y de la pomme de terre
Mosaïque de la tomate
Mosaïque de la tomate
Mosaïque du tabac
Virus X de la pomme de terre
Forme d'azote''
Nitrate
Diminue
Augmente
Augmente
Augmente
Augmente
Augmente
Augmente
Augmente
Augmente
Diminue
Diminue
Diminue
Augmente
Augmente
Diminue
Diminue
ND
ND
Augmente
Augmente
ND
ND
Ammonium
ND
Diminue
ND
ND
ND
ND
Diminue
Diminue
Diminue
Augmente
Augmente
Augmente
Diminue
Diminue
ND
Augmente
Augmente
Diminue
Diminue
Diminue
Diminue
Diminue
° Tableau adapté de Huber et Watson (1974)
'' L'addition de la forme d'azote (nitrate ou ammonium) diminue ou augmente la sévérité de la maladie.
' ND non déterminé

17
En outre, d'autres études effectuées sur l'azote ont montré qu'une concentration
élevée d'azote (450 ppm) n'a pas significativement augmenté la maladie de chancre
{Clavibacter mishiganense) chez la tomate comparativement à une concentration de 240
ppm d'azote (Berry et ai, 1988). Duffy et Défago (1999) ont montré que la maladie du
pourridié fusarien (Fusarium oxysporum f. sp. radicis-lycopersici) chez la tomate a
augmenté suite à l'augmentation de la concentration d'ammonium [NH4CI ; (NH4)6Mo7024 ;
(NH4)2S04]. Elle a par ailleurs été réduite par une application du nitrate de calcium et de
petites concentrations de nitrate d'ammonium (39 à 79 mg d'azote/1). Cependant, des
concentrations supérieures à 100 mg d'azote/1 ont augmenté la sévérité de cette maladie.
Hoffland et al. (1999) ont montré que l'augmentation de la concentration d'azote appliqué
diminue la sévérité de la moisissure grise causée par Botrytis cinerea chez la tomate. Par
contre, l'azote n'a pas eu d'effet sur le flétrissement fusarien causé par Fusarium
oxysporum f sp. lycopersici chez la tomate alors que l'augmentation de la concentration
d'azote a favorisé l'augmentation de la moucheture (Pseudomonas syringae pv. tomato) et
du blanc (Oïdium lycopersicum) chez la tomate (Hoffland et al., 2000). Nam et al. (2006)
ont montré que des concentrations élevées d'azote dans la solution nutritive ont augmenté
la sévérité de l'anthracnose (Colletotrichum gloeosporioides) chez une culture de fraisier en
système hydroponique fermé.
1.4.2 Phosphore
Woltz et Jones (1973 a, b, c ; 1981) ont montré qu'un niveau élevé de phosphore
augmentait la sévérité du flétrissement fusarien (F. oxysporum f. sp. lycopersici) chez la
tomate cultivée en pots et au champ. SagduUaev et Berezhnova (1974) ont montré que les
amendements en phosphore augmentaient la sévérité du flétrissement fusarien (F.
oxysporum f sp. melonis) chez le cantaloup. Dick et Tisdale (1938) ont montré que
l'augmentation de la dose de phosphore favorise l'augmentation de la sévérité du
flétrissement fusarien {F. oxysporum f. sp. vasinfectum) chez le coton. Brennan (1988) a
montré que les plants de blé cultivés sans phosphore ont été infectés par le piétin-échaudage
{Gaeumannomyces graminis var. tritici) tandis que l'augmentation de la concentration de
phosphore a permis la réduction de cette maladie. Duffy et Défago (1999) ont montré que le

18
pourridié fusarien (F. oxysporum f. sp. radicis-lycopersicî) a augmenté chez la tomate suite
à l'augmentation de la concentration de NaH2P04-H20. Sanogo et Yang (2001) ont montré
que la nutrition phosphatée a un effet sur le syndrome de la mort subite du soya [F. solani î.
sp. glycines {Fsg)\. En effet, le phosphate de calcium a augmenté la sévérité de la maladie
de 21 %, le phosphate de potassium de 32 % et le phosphate de sodium de 43 %. La
germination in vitro des conidies du Fsg n'a pas été affectée de façon significative par les
éléments nutritifs phosphatés ; par contre la croissance mycélienne a été favorisée sur
milieu de culture amendé de phosphate de potassium et de phosphate de sodium (Sanogo et
Yang, 2001).
1.4.3 Potassium
Sanogo et Yang (2001) ont montré que la nutrition potassique a un effet sur le
syndrome de la mort subite du soya \F. solani f. sp. glycines {Fsgy\. L'application du
chlorure de potassium a favorisé une réduction moyenne de 36 % de la sévérité de la
maladie comparativement au traitement témoin. Par contre, la sévérité de la maladie a
connu une augmentation moyenne de 32 % avec l'application du phosphate de potassium,
de 43 % avec le sulfate de potassium et de 45 % avec le nitrate de potassium. La
germination in vitro des conidies du Fsg n'a pas été affectée de façon significative par les
éléments nutritifs potassiques; par contre la croissance mycélienne a été favorisée sur
milieu de culture amendé de nitrate de potassium (Sanogo et Yang, 2001).
1.4.4 Calcium
Les sels de calcium ont montré un effet sur la croissance de Leucostoma persoonii,
agent responsable du chancre du pêcher. La plus grande réduction de la croissance in vitro a
été causée par le propionate de calcium (85 %), puis l'hydroxyde de calcium (76 %) et le
silicate de calcium (73 %). Le trempage des fruits pendant 15 minutes dans des solutions de
silicate de calcium et de propionate de calcium a diminué l'étendue des lésions de 70 %
(Biggs et al, 1995). Une autre étude réalisée par Biggs (1999) a montré que le chlorure de
calcium, le propionate de calcium et le silicate de calcium (1 000 \ig Ca/ml ) n'a eu aucun
effet sur la germination des conidies de deux espèces de Colletotrichum causant la

19
pourriture amère du pommier. Ces trois sels de calcium ont toutefois réduit la croissance de
Colletotrichum spp. en milieu de culture liquide et ont réduit la sévérité de la maladie sur
les fruits traités (Biggs, 1999). Biggs et al. (1997) ont montré que tous les sels de calcium à
l'exception du formate, du pantothenate et du phosphate dibasique de calcium ont réduit la
croissance sur le milieu de culture solide à base de dextrose de pomme de terre (PDA) de
Monilia fructicola, agent responsable de la pourriture brune du pêcher. Le chlorure de
calcium appliqué en post-récolte, a réduit significativement l'incidence et la sévérité de la
pourriture altemarienne (Alternaria spp.) chez la pomme (Biggs et al., 1993). Wisniewski
et al. (1995) ont montré que l'augmentation de la concentration du chlorure de calcium (25-
175 mM) entraîne une diminution de la germination et de la croissance du tube germinatif
in vitro du B. cinerea et du Pénicillium expansum, causant respectivement les moisissures
grise et bleue chez la pomme entreposée. Le chlorure de calcium a inhibé la croissance in
vitro du pathogène Rhizopus stolonifer, agent responsable de la moisissure chevelue des
fruits et des légumes en post-récolte. L'ajout de calcium dans le milieu de culture liquide à
base de dextrose de pomme de terre (PDB) a provoqué une réduction de la germination des
spores et un ralentissement de la croissance du tube germinatif de R. stolonifer (Tian et al.,
2002).
1.4.5 Magnésium
Selon Wisniewski et al. (1995), le chlorure de magnésium (25-175 mM) n'a pas eu
d'effet sur la germination et la croissance du tube germinatif in vitro de B. cinerea et de P.
expansum. La fertilisation avec du chlorure de magnésium a augmenté le flétrissement
fusarien causé par F. oxysporum f. sp. lycopersici chez la tomate (Jones e/ al., 1989) et par
F. oxysporum f. sp. apii chez le céleri (Schneider, 1985). Selon Duffy et Défago (1999), le
sulfate de magnésium n'a pas eu d'effet sur le pourridié fusarien {F. oxysporum f. sp.
radicis-lycopersici) chez la tomate. Taylor (1954) a montré que la susceptibilité du maïs à
la brûlure des feuilles (Helminthosporium maydis) diminue lorsque le niveau de magnésium
augmente dans les tissus foliaires.

20
1.5 Effet des éléments minéraux sur le développement de la gale argentée de la pomme de terre
Adams et al. (1970) ont étudié l'effet des propriétés du sol sur la gale argentée et ont
observé une corrélation négative entre la teneur du sol en nitrates et la sévérité de la gale
argentée chez les tubercules-fils. Martinez et al. (2002) ont pour leur part montré
l'existence d'une corrélation négative entre la sévérité de la gale argentée et les teneurs en
nitrates et fer des sols. Pour faire suite à ces travaux, le présent projet de recherche vise à
étudier in vitro l'effet direct de l'azote et de certains macro-éléments sur la croissance
mycélienne, le nombre de conidies total et le nombre de conidies viables produites par H.
solani, agent responsable de la gale argentée.
1.6 Hypothèse et objectifs de recherche
1.6.1 Hypothèse de recherche
L'azote, le phosphore, le potassium, le calcium et le magnésium ont un effet direct
sur la croissance mycélienne, le nombre total de conidies et le nombre de conidies viables
produites par l'agent pathogène H. solani.
1.6.2 Objectifs de recherche
- Évaluer in vitro l'effet de différentes concentrations et de différentes sources
d'azote, de phosphore, de potassium, de calcium et de magnésium sur la croissance
mycélienne du champignon H. solani.
- Évaluer in vitro l'effet de différentes concentrations et de différentes sources
d'azote, de phosphore, de potassium, de calcium et de magnésium sur le nombre total de
conidies produites par le champignon H. solani.
- Évaluer in vitro l'effet de différentes concentrations et de différentes sources
d'azote, de phosphore, de potassium, de calcium et de magnésium sur le nombre de
conidies viables produites par le champignon H. solani.

21
CHAPITRE 2
MATÉRIEL ET MÉTHODES
2.1 Helminthosporium solani
La souche ULaval-1 de Helminthosporium solani a été employée tout au long de ces
travaux. Elle est disponible sous le numéro DAOM 233452 (Department of Agriculture,
Ottawa, Mycology). Elle a été isolée à partir d'un tubercule de pomme de terre présentant
des symptômes de gale argentée au Laboratoire de mycologie du Centre de recherche en
horticulture de l'Université Laval. Cette souche a été cultivée à 24°C, sous obscurité, en
boîte de Pétri sur le milieu de culture V8 agar. Ce milieu est composé de 200 ml de jus de
V8 clarifié (Campbell's Soup Company Ltd, Toronto, ON), 3 g de CaCOa, 15 g de Bacto
Agar (Difco Laboratories, Becton Dickinson, Sparks, MD) et de 800 ml d'eau distillée. Le
pH a été ajusté à 7,2. La souche était conservée à 4°C (Michaud, 2001).
2.2 Effet des éléments minéraux sur la croissance et le développement de H. solani
Afin d'évaluer l'effet des éléments minéraux sur la croissance et le développemnt de
H. solani, le champignon a été cultivé en plat de Pétri sur gélose nutritive (milieu de base)
dont la composition en azote, phosphore, potassium, calcium et magnésium variait. Le
milieu développé par Singh (1968) légèrement modifié a été utilisé comme milieu de base
pour ces travaux. La composition du milieu était la suivante : 30 g/1 saccharose (EMD
Chemicals, Merck KgaA, Darmstadt, Germany), 15 g/1 agar (Difco Laboratories, Becton
Dickinson, Sparks, MD), 2 g/1 NaNOa (Laboratoire Mat, Québec, Québec), 1 g/1 K2HPO4
(Sigma-Aldrich, St. Louis, MO), 0,5 g/1 MgS04-7H20 (Sigma-Aldrich), 0,5 g/1 KCl (Sigma
Chemical Co., St. Louis, MO), 0,01 g/1 FeS04-7H20 (EM Science, EM Industries, Merck
KgaA, Darmstadt, Germany), 4,6 mg/1 d'acide citrique anhydre (BDH, Toronto, Ontario), 5
mg/1 ZnS04-7H20 (BDH), 0,391 mg/1 CuS04-5H20 (Sigma Chemical Co.), 0,05 mg/1
MnS04-H20 (Sigma Chemical Co.), 0,05 mg/1 H3BO4 (BDH), 0,05 mg/1 Na2Mo04-2H20
(Sigma-Aldrich) et 500 |j,g/l vitamine Bi (thiamine) (Gibco BRL, Life Technologies, Grand
Island, NY) ajoutée stérilement après autoclavage ; le pH était ajusté à 6,8 avec HCl (IN)
(Fisher Scientific, Nepean, Ontario) ou NaOH (IN) (EM Science), après ajout des

22
différentes concentrations de l'élément à l'étude. Les concentrations et les sources de
minéraux testées sont respectivement présentées aux Tableaux 2 et 3. Les différentes
géloses ont été inoculées avec une pastille (0,7 cm de diamètre) de V8 agar portant du
mycélium de H. solani en croissance active et incubées sous obscurité à 25°C pour une
période de huit semaines.
Tableau 2. Concentrations d'azote, de phosphore, de potassium, de calcium et de
magnésium testées dans cette étude
Éléments minéraux Concentrations (ppm)
Azote
Phosphore
Potassium
Calcium
Magnésium
0 0
0
0
0
3,3 1,778
7,1 0,272
0,5
33 17,78
71,1 2,72 5
330 177,8 711,18
27,21
50
3300 1778
7111,8 272,1
500
Tableau 3. Sources d'éléments minéraux testées dans cette étude
Éléments minéraux Sources
Azote
Phosphore
Potassium
Calcium
Magnésium
NaNOa (Laboratoire Mat) NH4CI (Laboratoire Mat) NH4NO3 (Laboratoire Mat) NaNOa (Sigma Chemical) Na2HP047H20 (Sigma Chemical Co.) Na3P04-12H20 (Sigma Chemical Co.) KCl (Sigma Chemical Co.) K2SO4 (Sigma Chemical Co.) CaS04-2H20 (Sigma Chemical Co.) CaCl2-2H20 (Sigma Chemical Co.) MgS04-7H20 (Sigma-Aldrich) MgCl22H20 (Sigma Chemical Co)

23
2.2.1 Détermination de la croissance mycélienne
La croissance mycélienne (CM) a été évaluée hebdomadairement pendant huit
semaines en mesurant la surface de gélose couverte par le mycélium. Elle a été calculée
selon la formule (1) et exprimée en cm . Cette formule destinée à évaluer la surface du
thalle mycélien a été retenue car elle tient compte de la forme elliptique du thalle.
(1 ) C M — omycélium" ^pastille
OÙ Smycéiium = Surfacc totale du mycélium = '/2 C x '/a L x 71
Spastiiie = Sxirface de la pastille = (r pastiiie x 71) = 0,38465 cm^
C : diamètre court du thalle mycélien
L : diamètre long du thalle mycélien
2.2.2 Détermination du nombre de conidies produites
Le dénombrement des conidies a été effectué après la période de culture de huit
semaines. Les conidies ont été récupérées avec 2 ml d'eau distillée stérile par grattage du
mycélium (Section 2.2.1) à l'aide d'une pipette pasteur stérile courbée sous forme de bâton
de hockey. La suspension conidienne a été récupérée dans un tube stérile. Après agitation
de la suspension au vortex, les conidies ont été dénombrées à l'aide d'un hémacytomètre
(Hausser Scientific Company, Horsham, PA). La production conidienne a été exprimée en
nombre de conidies par unité de surface mycélienne (conidies/cm^).
2.2.3 Détermination du nombre de conidies viables
La suspension conidienne obtenue (Section 2.2.2) a été étalée sur un milieu de culture
solide pour dénombrer le nombre d'unités formatrices de colonies (UFC). Ainsi, après
agitation au vortex, 100 |j,l de la suspension ont été prélevés et étalés sur le milieu potato
dextrose agar (PDA, Difco Laboratories). Le dénombrement des UFC a été effectué quatre
jours après l'étalement par comptage des colonies mycéliennes visibles à l'œil nu. Le

24
nombre de conidies viables a été exprimé en UFC par unité de surface mycélienne
(UFC/cm^).
2.3 Dispositif expérimental et analyse statistique
Le dispositif expérimental utilisé était un plan complètement aléatoire comprenant 15
répétitions. Une boîte de Pétri représentait l'unité expérimentale. L'analyse de variance
(ANOVA) a été effectuée et, lorsque significative, les moyennes des traitements étaient
comparées à l'aide du test Least Significant Difference (LSD) protégé (P = 0,05) au moyen
du logiciel JMP version 5.1 (SAS Institute, Cary, NC).
25
CHAPITRE 3
RÉSULTATS
3.1 Effet des éléments minéraux sur la croissance mycélienne de H. solani
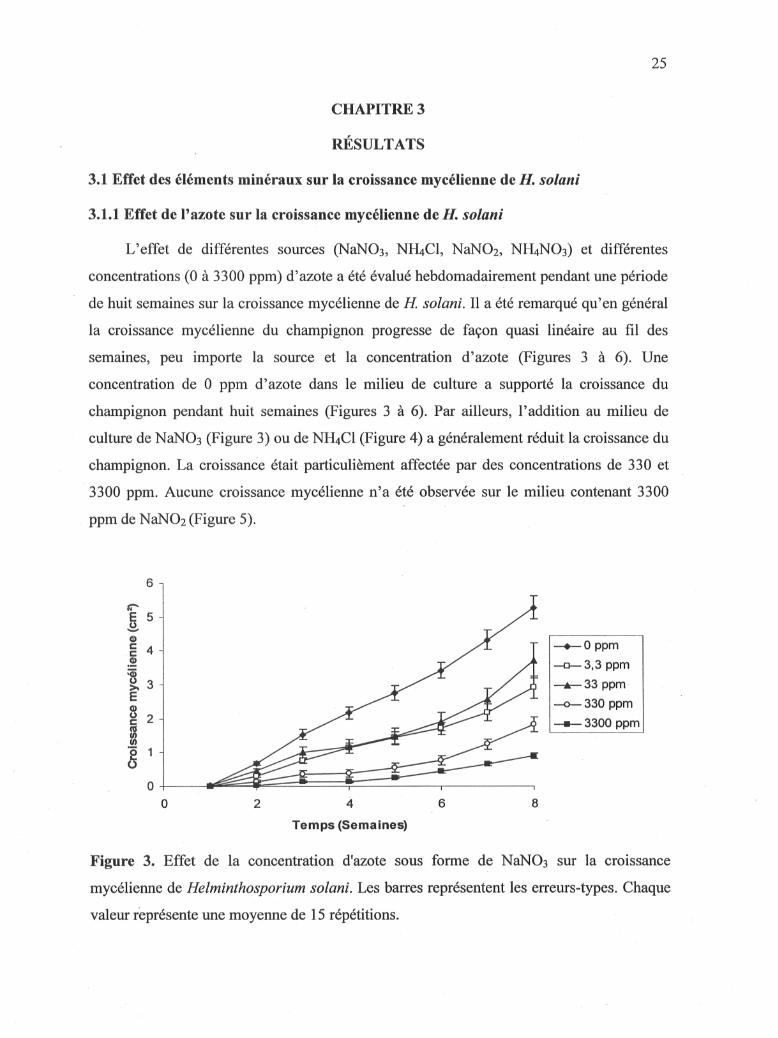
3.L1 Effet de l'azote sur la croissance mycélienne de H. solani
L'effet de différentes sources (NaNOs, NH4CI, NaN02, NH4NO3) et différentes
concentrations (0 à 3300 ppm) d'azote a été évalué hebdomadairement pendant une période
de huit semaines sur la croissance mycélienne de H. solani. Il a été remarqué qu'en général
la croissance mycélienne du champignon progresse de façon quasi linéaire au fil des
semaines, peu importe la source et la concentration d'azote (Figures 3 à 6). Une
concentration de 0 ppm d'azote dans le milieu de culture a supporté la croissance du
champignon pendant huit semaines (Figures 3 à 6). Par ailleurs, l'addition au milieu de
culture de NaNOa (Figure 3) ou de NH4CI (Figure 4) a généralement réduit la croissance du
champignon. La croissance était particuhèment affectée par des concentrations de 330 et
3300 ppm. Aucune croissance mycélienne n'a été observée sur le milieu contenant 3300
ppm de NaNOi (Figure 5).
• 0 ppm ■ 3,3 ppm ■ 33 ppm ■ 330 ppm ■ 3300 ppm
Temps (Semaines)
Figure 3. Effet de la concentration d'azote sous forme de NaNOs sur la croissance
mycélienne de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyenne de 15 répétitions.

26
iH -
"g 12 -u « 10 -c 0) •5 8 o >> E 6 0) u « 4 -^ 2 ? o ^
0 -
-♦—0 ppm
-o—3,3 ppm
-A— 33 ppm
-o— 330 ppm
- * - 3300 ppm
Temps (semaines)
Figure 4. Effet de la concentration d'azote sous forme de NH4CI sur la croissance
mycélienne de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyerme de 15 répétitions.
-♦—0 ppm
-D—3,3 ppm
-A— 33 ppm
-o— 330 ppm
-m— 3300 ppm
Temps (semaines)
Figure 5. Effet de la concentration d'azote sous forme de NaNOi sur la croissance
mycélierme de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyenne de 15 répétitions.

27
a -îT 8 -E A 7 ^ 0)
ë 6-0) :â> 5 -, u >« E 4 -o ë 3 ra 8 7-2 , o 1
0 -
-♦—0 ppm
-D—3,3 ppm
-A— 33 ppm
-o—330 ppm
-*—3300 ppm
Temps (semaines)
Figure 6. Effet de la concentration d'azote sous forme de NH4NO3 sur la croissance
mycélienne de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyenne de 15 répétitions.
Après huit semaines d'incubation, on observe que des concentrations élevées d'azote
réduisent la croissance de H. solani par rapport aux concentrations plus faibles et ce pour
toutes les sources d'azote à l'exception du NaNOa (Figure 7). Plus spécifiquement, des
concentrations supérieures à 33 ppm de NaNOa ont réduit de façon significative la
croissance de H. solani par rapport au traitement 0 ppm. Pour la forme NH4CI, il n'y a pas
eu de différence significative de croissance dans les milieux contenant 0, 3,3 et 33 ppm. Par
contre, dans les milieux contenant 330 et 3300 ppm de NH4CI la croissance était
significativement réduite. L'azote sous forme de NaNOi a également eu un effet significatif
sur la croissance mycélienne du champignon, mais cet effet n'a suivi aucune tendance
particulière en fonction de la concentration utilisée. Une concentration de 3300 ppm de
NaNOi a inhibé complètement la croissance de H. solani. Sous forme de NH4NO3, une
diminution significative de la croissance de H. solani a été observée lorsque la
concentration était de 330 et 3300 ppm.

28
NaN03 NH4C1 NaN02 NH4N03
n o ppm I I 3,3 ppm m33 ppm ■ 330 ppm
■ 3300 ppm
Figure 7. Effet de la concentration d'azote sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation. Pour
chacune des sources, les concentrations testées sont exprimées en ppm d'azote. Pour chaque
source, les valeurs surmontées d'une même lettre ne sont pas significativement différentes
selon un test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15
répétitions.
3.1.2 Effet du phosphore sur la croissance mycélienne de H. solani
L'effet de deux sources (Na2HP04-7H20, Na3P04-12H20) et de différentes
concentrations (0 à 1778 ppm) de phosphore a été évalué hebdomadairement pendant une
période de huit semaines sur la croissance mycélienne de H. solani. Il a été remarqué que la
croissance mycélienne du champignon H. solani progresse rapidement après la quatrième
semaine d'incubation (Figures 8 et 9) ; on note par la suite un début de croissance
logarithmique.

29
-♦—0 ppm -o—1,778 ppm -A—17,78 ppm -o—177,8 ppm -•—1778 ppm
0 2 4 6 8
Temps (semaines)
Figure 8. Effet de la concentration de phosphore sous forme de Na2HP04-7H20 sur la
croissance mycélienne de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyenne de 15 répétitions.
-0 ppm -1,778 ppm -17,78 ppm -177,8 ppm -1778 ppm
0 2 4 6 8
Temps (semaines)
Figure 9. Effet de la concentration de phosphore sous forme de Na3P04-12H20 sur la
croissance mycélienne de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyenne de 15 répétitions.

30
Après huit semaines d'incubation, on note que les concentrations de Na2HP04-7H20
n'ont eu aucun effet significatif sur la croissance de H. solani (Figure 10). Sous forme de
Na3P04-12H20, l'addition des concentrations de 177,8 et 1778 ppm a stimulé la croissance
(Figure 10).
D O ppm □ 1,778 ppm H 17,78 ppm ■ 177,8 ppm ■ 1778 ppm
Na2HP04.7H20 Na3P04.12H20
Figure 10. Effet de la concentration de phosphore sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurées après huit semaines d'incubation. Pour
chacune des sources, les concentrations testées sont exprimées en ppm de phosphore. Pour
chaque source, les valeurs surmontées d'une même lettre ne sont pas significativement
différentes selon un test LSD protégé {P = 0,05). Chaque valeur représente une moyenne de
15 répétitions.
3.1.3 Effet du potassium sur la croissance mycélienne de H. solani
L'effet de deux sources (KCl, K2SO4) et de différentes concentrations (0 à 7111,8
ppm) de potassium a été évalué hebdomadairement pendant une période de huit semaines
sur la croissance mycélienne de H. solani. Les résultats obtenus ont montré que les fortes
concentrations de KCl et de K2SO4 stimulaient la croissance du champignon (Figures 11 et
12).

31
0 -
s- 2,5 E o "• « c 2 -c .S
.■fl)
^ 1.5 E S s 1 M M S O 0,5
0 ■—
/ 1 / -
/%///-'
jT T v ^ * * ^ y " ^ v ^
— ■ - - a*^^ T 1 1
-♦—0 ppm -a—7,1 ppm -^—71,1 ppm -o—711,18 ppm -•—7111,8 ppm
Temps (semaines)
Figure 11. Effet de la concentration de potassium sous forme de KCl sur la croissance
mycélienne de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyenne de 15 répétitions.
-♦—0 ppm -o—7,1 ppm -A—71,1 ppm -o—711,18 ppm -•—7111,8 ppm
Temps (semaines)
Figure 12. Effet de la concentration de potassium sous forme de K2SO4 sur la croissance
mycélienne de Helminthosporium solani. Les barres représentent les erreurs-types. Chaque
valeur représente une moyenne de 15 répétitions.

32
Après huit semaines d'incubation, il n'y a pas eu de différence significative de
croissance entre 0, 7,1 et 71,1 ppm de KCl. Par contre, une concentration de 7111,8 ppm de
KCl a significativement augmenté la croissance mycélienne en comparaison avec 0, 7,1 et
71,1 ppm (Figure 13). La concentration de K2SO4 ajoutée au milieu a également influencé
significativement la croissance du champignon (Figure 13). Une concentration de 7111,8
ppm a permis d'obtenir la croissance mycélienne la plus importante.
nO ppm H 7,1 ppm i l 71,1 ppm ■ 711,18 ppm ■ 7111,8 ppm
KCl K2S04
Figure 13. Effet de la concentration de potassium sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation. Pour
chacune des sources, les concentrations testées sont exprimées en ppm de potassium. Pour
chaque source, les valeurs surmontées d'une même lettre ne sont pas significativement
différentes selon un test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de
15 répétitions.
3.1.4 Effet du calcium sur la croissance mycélienne de Jï. solani
L'effet de deux sources (CaCl2-2H20, CaS04-2H20) et de différentes concentrations
(0 à 272,1 ppm) de calcium a été évalué hebdomadairement pendant une période de huit
semaines sur la croissance mycélienne de H. solani. La croissance mycélienne la plus
importante a été obtenue lorsque le milieu était amendé avec 27,21 ppm de CaCl2-2H20
(Figure 14) et 0,272 ppm de CaS04-2H20 (Figure 15). La concentration de 272,1 ppm
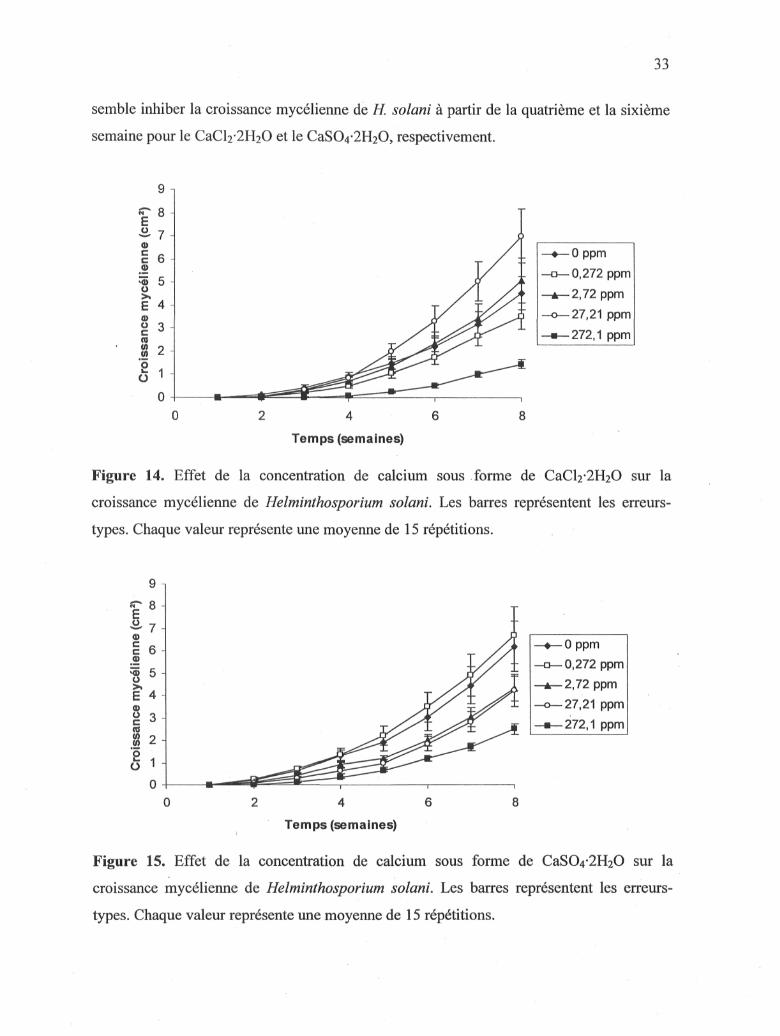
33
semble inhiber la croissance mycélienne de H. solani à partir de la quatrième et la sixième
semaine pour le CaC^^HaO et le CaS04-2H20, respectivement.
-♦—0 ppm
-C3—0,272 ppm
-à—2,72 ppm
-o—27,21 ppm
-•—272,1 ppm
Temps (semaines)
Figure 14. Effet de la concentration de calcium sous forme de CaCl2'2H20 sur la
croissance mycélienne de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyenne de 15 répétitions.
-♦—0 ppm -0—0,272 ppm -A—2,72 ppm -o—27,21 ppm -•—272,1 ppm
2 4
Temps (semaines)
Figure 15. Effet de la concentration de calcium sous forme de CaS04-2H20 sur la
croissance mycélienne de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyenne de 15 répétitions.
34
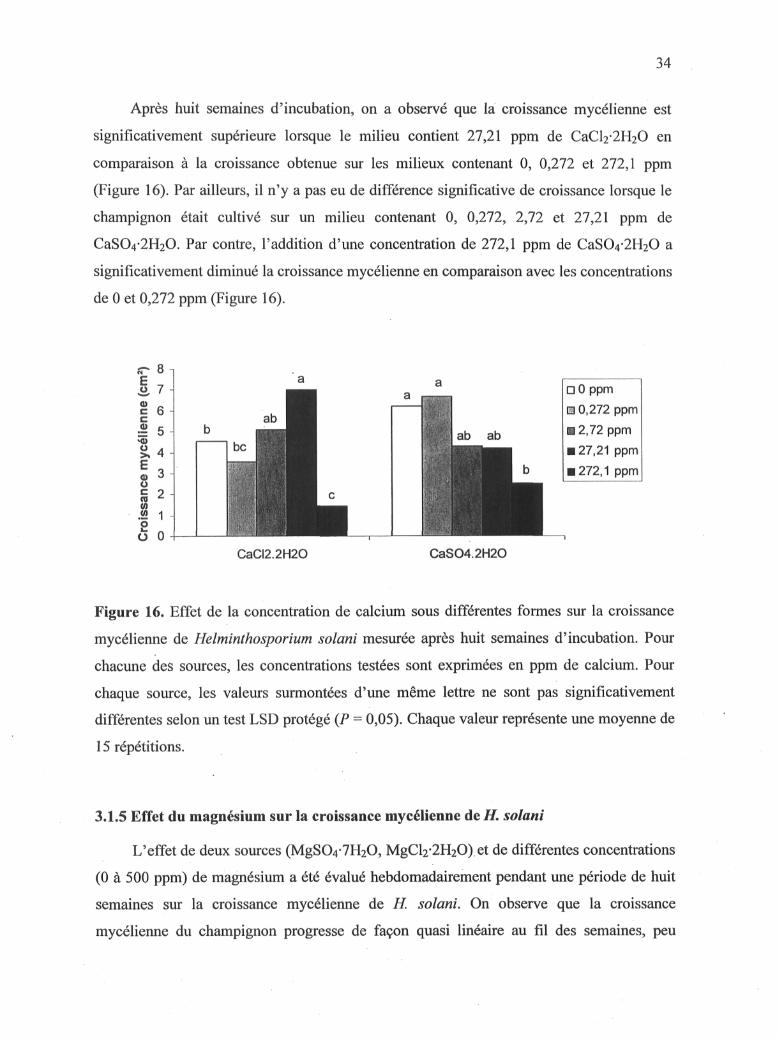
Après huit semaines d'incubation, on a observé que la croissance mycélienne est
significativement supérieure lorsque le milieu contient 27,21 ppm de CaCl2-2H20 en
comparaison à la croissance obtenue sur les milieux contenant 0, 0,272 et 272,1 ppm
(Figure 16). Par ailleurs, il n'y a pas eu de différence significative de croissance lorsque le
champignon était cultivé sur un milieu contenant 0, 0,272, 2,72 et 27,21 ppm de
CaS04-2H20. Par contre, l'addition d'une concentration de 272,1 ppm de CaS04-2H20 a
significativement diminué la croissance mycélienne en comparaison avec les concentrations
de 0 et 0,272 ppm (Figure 16).
D O ppm m 0,272 ppm
2,72 ppm 27,21 ppm 272,1 ppm
CaCI2.2H20 CaS04.2H20
Figure 16. Effet de la concentration de calcium sous différentes formes sur la croissance
mycélienne de Helminthosporium solani mesurée après huit semaines d'incubation. Pour
chacune des sources, les concentrations testées sont exprimées en ppm de calcium. Pour
chaque source, les valeurs surmontées d'une même lettre ne sont pas significativement
différentes selon un test LSD protégé {P = 0,05). Chaque valeur représente une moyenne de
15 répétitions.
3.1.5 Effet du magnésium sur la croissance mycélienne de H. solani
L'effet de deux sources (MgS04-7H20, MgCl2-2H20) et de différentes concentrations
(0 à 500 ppm) de magnésium a été évalué hebdomadairement pendant une période de huit
semaines sur la croissance mycélienne de H. solani. On observe que la croissance
mycélienne du champignon progresse de façon quasi linéaire au fil des semaines, peu

35
importe la source et la concentration du magnésium (Figures 17 et 18). La concentration de
500 ppm de magnésium dans le milieu de culture a permis la meilleure croissance
mycélienne pour le MgS04-7H20 (Figure 17) et les concentrations de 0,5, 5 et 50 favorisent
la meilleure croissance mycélienne pour le MgCl2-2H20 (Figure 18).
—♦—0 ppm
—D—0,5 ppm
—*—5 ppm
—o— 50 ppm
—■— 500 ppm
Temps (semaines)
Figure 17. Effet de la concentration de magnésium sous forme de MgS04-7H20 sur la
croissance mycélienne de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyerme de 15 répétitions.
-0 ppm -0,5 ppm -5 ppm - 50 ppm - 500 ppm
0 2 4 6 8
Temps (semaines)
Figure 18. Effet de la concentration de magnésium sous forme de MgCl2-2H20 sur la
croissance mycélierme de Helminthosporium solani. Les barres représentent les erreurs-
types. Chaque valeur représente une moyerme de 15 répétitions.

36
Après huit semaines d'incubation, on observe que la concentration de MgS04-7H20
et de MgCl2-2H20 n'a pas eu d'effet significatif sur la croissance mycélienne de H. solani
(Figure 19).
8
7
6
5
4
3 ^
2
1
0
-
~
^ -
a
a
Wm
a
HpÇa
» \ 'f
a n 0 ppm H 0,5 ppm la 5 ppm ■ 50 ppm ■ 500 ppm
MgS04.7H20 MgCI2.2H20
Figure 19. Effet de la concentration de magnésium sous différentes formes sur la
croissance mycélienne de Helminthosporium solani mesurée après huit semaines
d'incubation. Pour chacune des sources, les concentrations testées sont exprimées en ppm
de magnésium. Pour chaque source, les valeurs surmontées d'une même lettre ne sont pas
significativement différentes selon un test LSD protégé {P = 0,05) Chaque valeur
représente une moyenne de 15 répétitions.

37
3.2 Effet des éléments minéraux sur le nombre de conidies produites
Les résultats obtenus ont montré qu'à l'exception du MgS04-7H20, la concentration
des éléments minéraux incorporés au milieu de culture a influencé significativement le
nombre de conidies produites par H. solani. Dans le cas des sources d'azote, les travaux ont
montré que l'incorporation de 330 ppm de NaNOs et de NaN02 et de 3300 ppm de NH4CI
et de NH4NO3 a augmenté significativement le nombre de conidies produites (Figure 20).
Tout comme dans le cas de l'azote, l'incorporation de concentrations élevées de
CaS04-2H20, CaCl2-2H20 et de MgCl2-2H20 a favorisé le nombre de conidies produites.
En effet, l'incorporation de 272,1 ppm de CaS04-2H20 (2875,15 conidies/cm^), de 272,1
ppm de CaCl2-2H20 (6997,04 conidies/cm^) (Figure 21) et de 500 ppm MgCl2-2H20
(2228,86 comdies/cm ) (Figure 22) a augmenté significativement le nombre de conidies
produites. À la lecture des résultats concernant l'effet des concentrations de phosphore, il
apparaît évident que l'incorporation de fortes concentrations a réduit le nombre de conidies
produites (Figure 23). Les milieux contenant 177,8 et 1778 ppm de Na2HP04-7H20 ou de
Na3P04l2H20 ont causé une réduction significative du nombre de conidies produites
comparativement aux milieux contenant 0 ppm et 1,778 ppm. Dans le cas des sources de
potassium, les thalles mycéliens cultivés dans un milieu supplémenté avec 7111,8 ppm de
KCl ont produit significativement moins de conidies (875,78 conidies/cm^) que les thalles
mycéliens cultivés sur un milieu amendé avec 71,1 (5526,46 conidies/cm^) ou 711,18 ppm
(4850,07 conidies/cm^) (Figure 24). Enfin, les thalles mycéliens cultivés sur un milieu
supplémenté avec 7111,8 ppm de K2SO4 ont produit significativement moins de conidies
(1184,91 conidies/cm^) que les thalles mycéliens cultivés sur un milieu amendé avec 7,1
ppm de K2SO4 (4255,92 conidies/cm^).

38 25000
E •^ 20000
o 15000 u
Si 10000
NaN03 NH4CI
b a a a H a
NaN02
I 1 1 1 J NH4N03
nO ppm El3,3 ppm ■ 33 ppm ■ 330 ppm ■ 3300 ppm
Figure 20. Effet de la concentration d'azote sous différentes formes sur le nombre de
conidies produites par le champignon Helminthosporium solani. Pour chacune des sources,
les concentrations testées sont exprimées en ppm d'azote. Pour chaque source, les valeurs
surmontées d'une même lettre ne sont pas significativement différentes selon un test LSD
protégé (P = 0,05). Chaque valeur représente une moyenne de 15 répétitions.
8000
CaS04.2H20 CaCI2.2H20
nO ppm H 0,272 ppm a 2,72 ppm ■ 27,21 ppm ■ 272,1 ppm
Figure 21. Effet de la concentration de calcium sous différentes formes sur le nombre de
conidies produites par le champignon Helminthosporium solani. Pour chacune des sources,
les concentrations testées sont exprimées en ppm de calcium. Pour chaque source, les
valeurs surmontées d'une même lettre ne sont pas signifïcativement différentes selon un
test LSD protégé {P = 0,05). Chaque valeur représente une moyenne de 15 répétitions.

39
zouu -
o 2000 (0
o 1500 -
m
■§ 1000 -
m m
1 500 -o O
0 -
a
a
S Afffii
.f '
a
S
a
a
nO ppm
110,5 ppm
@5 ppm
■ 50 ppm
■ 500 ppm
MgS04.7H20 MgCI2.2H20
Figure 22. Effet de la concentration de magnésium sous différentes formes sur le nombre
de conidies produites par le champignon Helminthosporium solani. Pour chacune des
sources, les concentrations testées sont exprimées en ppm de magnésium. Pour chaque
source, les valeurs surmontées d'une même lettre ne sont pas significativement différentes
selon un test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15
répétitions.
_ 14000
aO ppm 1,778 ppm 17,78 ppm 177,8 ppm 1778 ppm
Na2HP04.7H20 Na3P04.12H20
Figure 23. Effet de la concentration de phosphore sous différentes formes sur le nombre de
conidies produites par le champignon Helminthosporium solani. Pour chacune des sources,
les concentrations testées sont exprimées en ppm de phosphore. Pour chaque source, les
valeurs surmontées d'une même lettre ne sont pas significativement différentes selon un
test LSD protégé {P = 0,05). Chaque valeur représente vme moyenne de 15 répétitions.

40
6000
KCl K2S04
nO ppm ■ 7,1 ppm
71,1 ppm 711,18 ppm 7111,8 ppm
Figure 24. Effet de la concentration de potassium sous différentes formes sur le nombre de
conidies produites par le champignon Helminthosporium solani. Pour chacune des sources,
les concentrations testées sont exprimées en ppm de potassium. Pour chaque source, les
valeurs surmontées d'une même lettre ne sont pas significativement différentes selon un
test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15 répétitions.
3.3 Effet des éléments minéraux sur le nombre de conidies viables produites
Dans le cas de l'évaluation de l'effet des éléments minéraux sur le nombre de
conidies viables produites par le champignon, les résultats ont montré qu'à l'exception du
NH4NO3, la concentration d'éléments minéraux incorporée au milieu de culture a eu une
influence significative. Concernant les sources d'azote, l'incorporation de 300 ppm de
NaN03 ou de NaN02 a augmenté significativement le nombre de conidies viables produites
comparativement au traitement 0 ppm pour le NaNOs et aux traitements 0 ppm et 3,3 ppm
pour le NaN02. Pour le NH4CI, il n'y a eu aucune différence significative entre les
concentrations 0, 3,3, 330 et 3300 ppm. Le milieu contenant 33 ppm a réduit
significativement le nombre de conidies viables comparativement au traitement 0 ppm
(Figure 25). Dans le cas des sources de phosphore, la concentration de 1,778 ppm a
augmenté significativement le nombre de conidies viables produites pour les deux sources
testées (Figure 26). L'incorporation de concentrations élevées de KCl ou de K2SO4 a réduit
significativement le nombre de conidies viables produites (Figure 27), La concentration de

41
7111,8 ppm a diminué significativement le nombre de conidies viables produites. Quant à
l'effet des concentrations de calcium, l'incorporation de 272,1 ppm de CaS04-2H20 ou de
CaCl2-2H20 a augmenté significativement le nombre de conidies viables produites (Figure
28). Dans le cas des sources de magnésium, la concentration de 5 ppm de MgS04-7H20 a
augmenté significativement le nombre de conidies viables produites (Figure 29). Quant au
MgCl2-2H20, la concentration de 50 ppm a réduit significativement le nombre de conidies
viables produites comparativement au traitement 0 ppm (Figure 29).
250
NaN03 NH4CI NaN02 NH4N03
nO ppm I l 3,3 ppm B 33 ppm ■ 330 ppm ■ 3300 ppm
Figure 25. Effet de la concentration d'azote sous différentes formes sur le nombre de
conidies viables produites par le champignon Helminthosporium solani. Pour chacune des
sources, les concentrations testées sont exprimées en ppm d'azote. Pour chaque source, les
valeurs surmontées d'une même lettre ne sont pas significativement différentes selon le test
LSD protégé {P = 0,05). Chaque valeur représente une moyenne de 15 répétitions.

42 1600
nO ppm B 1,778 ppm ■ 17,78 ppm ■ 177,8 ppm ■ 1778 ppm
Na2HP04.7H20 Na3P04.12H20
Figure 26. Effet de la concentration de phosphore sous différentes formes sur le nombre de
conidies viables produites par le champignon Helminthosporium solani. Pour chacune des
sources, les concentrations testées sont exprimées en ppm de phosphore. Pour chaque
source, les valeurs surmontées d'une même lettre ne sont pas significativement différentes
selon le test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15
répétitions.
1800 n 1600 -
1 - , ^
N E 1400 -£ 1200 -O « 1000 -n .5 800 -> o 600 -■o
g 400-o 200 -
n -
a nO ppm H 7,1 ppm 8 71,1 ppm ■ 711,18 ppm ■ 7111,8 ppm
KCl K2S04
Figure 27. Effet de la concentration de potassium sous différentes formes sur le nombre de
conidies viables produites par le champignon Helminthosporium solani. Pour chacune des
sources, les concentrations testées sont exprimées en ppm de potassium. Pour chaque
source, les valeurs surmontées d'une même lettre ne sont pas significativement différentes
selon le test LSD protégé {P = 0,05). Chaque valeur représente une moyenne de 15
répétitions.

43
E o
O,
(0
1000
800
600
5 400 (0 .2 1 200 c o O
CaS04.2H20 CaCI2.2H20
nO ppm Q 0,272 ppm a 2,72 ppm ■ 27,21 ppm ■ 272,1 ppm
Figure 28. Effet de la concentration de calcium sous différentes formes sur le nombre de
conidies viables produites par le champignon Helminthosporium solani. Pour chacune des
sources, les concentrations testées sont exprimées en ppm de calcium. Pour chaque source,
les valeurs surmontées d'une même lettre ne sont pas significativement différentes selon le
test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15 répétitions.
1200
MgS04.7H20
nO ppm H 0,5 ppm ■ 5 ppm ■ 50 ppm ■ 500 ppm
MgCI2.2H20
Figure 29. Effet de la concentration de magnésium sous différentes formes sur le nombre
de conidies viables produites par le champignon Helminthosporium solani. Pour chacune
des sources, les concentrations testées sont exprimées en ppm de magnésium. Pour chaque
source, les valeurs surmontées d'une même lettre ne sont pas significativement différentes
selon le test LSD protégé (P = 0,05). Chaque valeur représente une moyenne de 15
répétitions.

44
CHAPITRE 4
DISCUSSION
Les éléments minéraux sont essentiels à la croissance et au développement des
champignons. Les résultats obtenus ont démontré que la source et la concentration
d'éléments minéraux étudiés avaient une influence sur la croissance mycélienne, la
production totale de conidies et la production de conidies viables chez le champignon H.
solani.
Les résultats relatifs à l'effet de l'azote sont résumés au Tableau 4. Ils montrent qu'il
y a en général, une diminution de la croissance mycélienne et une augmentation du nombre
de conidies produites en présence de concentrations plus élevées. Le nombre de conidies
viables diminue avec l'augmentation de la concentration de NaNOa et de NaN02. Cette
diminution de la croissance mycélienne combinée à la diminution du nombre de conidies
viables produites avec des concentrations élevées de NaNOs pourrait revêtir un intérêt
particulier dans le contexte de la lutte contre le champignon H. solani. Ces résultats vont
dans le même sens que les travaux de Martinez et al. (2002) qui ont montré l'existence
d'une corrélation négative entre le contenu en nitrates du sol et la sévérité de la gale
argentée.
Tableau 4. Effet de l'azote sur le champignon Helminthosporium solani
Paramètres
Croissance mycélienne
Conidies produites
Conidies viables produites
NaNOj
Diminue avec l'augmentation de la concentration Augmentent légèrement avec l'augmentation de la concentration Diminuent avec l'augmentation de la concentration
Sources d'azote
NH4CI
Diminue avec l'augmentation de la concentration Augmentent avec l'augmentation de la concentration
Augmentent avec l'augmentation de la concentration
NaNOî
Non concluant
Augmentent légèrement avec l'augmentation de la concentration Diminuent avec l'augmentation de la concentration
NH4NO3
Diminue avec l'augmentation de la concentration Augmentent avec l'augmentation de la concentration
Aucun effet

45
La plupart des champignons peuvent utiliser les nitrates comme source d'azote. La
seule voie du métabolisme d'assimilation de l'azote sous forme des nitrates est leur
réduction en nitrites en présence de l'enzyme nitrate reductase, suivie d'une réduction en
ammonium (Pateman et Kinghorn, 1976 ; Deacon, 2006). Le nitrate reductase est un
enzyme qui catalyse le transfert d'électron du NADPH aux nitrates et il requiert le
molybdène et le fer comme cofacteurs. Certains groupes de champignons comme les
basidiomycètes, les saprolegniaceae et les blastocladiales sont incapables d'utiliser les
nitrates comme source d'azote puisqu'ils sont incapables de synthétiser le nitrate reductase
(Whitaker, 1976). Ceci ne semble pas être le cas de H. solani qui croît relativement bien sur
un milieu contenant des nitrates.
L'ammonium peut généralement être utilisé comme source d'azote par la majorité
des champignons. On attribue à l'ion ammonium un rôle de régulateur de croissance. Par
exemple, la présence de l'ammonium en milieu de culture est très importante pour la
formation de périthèces fertiles; la formation des périthèces infertiles étant attribuable à la
présence des nitrates ou des nitrites chez Calonectria camelliae (Shipton, 1977). Par contre,
la sporulation chez Saccharomyces cerevisiae est inhibée par l'ion ammonium. Cette
inhibition est accompagnée d'une perte de viabilité. Par exemple, 2 mM de (NH4)2S04
inhibent complètement la sporulation et réduisent la viabilité des spores à 50 % (Pinon,
1977). Le principal site d'action de l'ion ammonium chez cette espèce est l'ADN. L'ion
ammonium inhibe la méiose et la phase pré-méiotique de la synthèse d'ADN. La présence
de l'ion ammonium cause une dégradation massive de l'ADN au cours d'une incubation
prolongée du champignon (Pinon, 1977). De plus, l'ammonium inhibe la synthèse et la
dégradation des protéines chez S. cerevisiae au cours de la formation des asques (Croes et
al., 1978). Par ailleurs, Morton et MacMillan (1954) ont montré que l'utilisation de
l'ammonium par la plupart des champignons est accompagnée d'une chute de pH du milieu
de culture. Le pH du milieu de culture du champignon Scopulariopsis brevicaulis décroît
jusqu'à 2,7 en présence de l'ammonium, il en résulte une faible assimilation de l'azote et
une mauvaise croissance du champignon (Morton et MacMillan, 1954).
Bien que les nitrites soient utilisés par quelques champignons comme source d'azote,
ils sont toxiques pour plusieurs espèces. La croissance du champignon Neurospora crassa
est inhibée par des fortes concentrations des nitrites (Nicholas, 1965), comme c'est le cas

46
pour le champignon H. solani avec l'inhibition totale de la croissance à une forte
concentration de NaN02. Cette toxicité des nitrites peut être attribuable à leur capacité de
désamination des acides aminés et à une interférence avec le métabolisme des sulfures
(Pateman et Kinghom, 1976). La mauvaise croissance des champignons en présence des
nitrites est souvent associée à l'effet du pH. Il a été démontré que la croissance de
VAspergillus niger sur les nitrites est accélérée par l'ajustement du pH du milieu de culture
de 5,6 à 7,5 (Agnihotri, 1968). Les nitrites sont convertis en ammonium sous l'action de
l'enzyme nitrite reductase, enzyme catalysant le transfert des électrons du NADPH vers les
nitrites (Pateman et Kinghom, 1976). D'autres études ont montré que Sderotium rolfsii
croît en présence de niveaux extrêmement élevés de NH4NO3, (NH4)2S04 et NaNOs
(Avizohar-Hershenzon et Shacked, 1969 ; Chaudhuri et Maiti, 1978 ; Johnson, 1953).
De façon surprenante, il est à noter qu'une concentration de 0 ppm d'azote dans le
milieu de culture a supporté la croissance radiale du champignon pendant huit semaines et
ce pour les quatre sources d'azote. À cet effet, des travaux complémentaires devraient être
menés afin de comprendre ce phénomène. Il est possible que le champignon croisse en
longs filaments minces puisqu'il est en déficit d'azote. Ceci pourrait expliquer que la
croissance radiale devient importante puisqu'il est à la recherche de l'azote dans son
environnement immédiat. Pour mieux comprendre ce phénomène, il est souhaitable
d'étudier l'effet des éléments étudiés sur le poids sec du champignon H. solani en milieu
liquide afin d'avoir une autre mesure de la croissance mycélienne. Des tests préliminaires
ont cependant montré que le milieu de culture employé lors de ce travail mais sous forme
liquide ne supporte pas la croissance de H. solani. Un autre milieu de culture synthétique
devra donc être développé pour étudier la croissance mycélierme en culture liquide.
Alternativement, des essais in vitro sur une membrane perméable (cellophane) recouvrant
le milieu gélose pourraient être réalisés. Le champignon pourrait être cultivé sur cette
membrane et le mycélium pourrait être facilement récupéré afin de déterminer le poids sec.
Une autre possibilité pouvant expliquer le fait que la concentration de 0 ppm d'azote dans
le milieu de culture ait supporté la croissance radiale serait qu'une composante du milieu de
culture (inoculum, agar, sucres, éléments minéraux) contienne de l'azote. Les deux sources
les plus susceptibles seraient l'agar et la pastille d'inoculation. Une expérience préliminaire
a montré que l'agar seul (15 g/1 ; sans additif) supporte la croissance radiale de H. solani.

47
Lorsqu'un gélifiant purifié (le Phytagel) a été employé, la croissance du champignon était
nulle. Dans ce contexte, des essais complémentaires devront être réalisés en remplaçant
l'agar par le Phytagel.
Les résultats relatifs à l'effet du phosphore sont résumés dans le Tableau 5.
Tableau 5. Effet du phosphore sur le champignon Helmînthosporium solani
Paramètres Sources de phosphore Na2HP04-7H20 Na3P04l2H20
Croissance mycélienne Aucun effet
Conidies produites Diminuent avec l'augmentation de la concentration
Conidies viables produites Diminuent à haute concentration
Augmente avec l'augmentation de la concentration Diminuent avec l'augmentation de la concentration
Diminuent à haute concentration
Les résultats obtenus par Sanogo et Yang (2001) sur l'effet des éléments nutritifs
phosphatés sur la croissance mycélienne et sur la germination in vitro de Fusarium solani
(Mart) Sacc. f sp. glycines (Fsg), agent responsable du syndrome de la mort subite du soya,
ont montré que la croissance mycélierme a été améliorée sur un milieu de culture amendé
de K2HPO4 et de Na2HP04 comme H. solani en présence de Na3P04-12H20. Le même
phénomène a été observé chez Candida sp., où la croissance a significativement augmenté
en présence d'une variété de sources organiques et inorganiques de phosphore (Shamis et
ai, 1968). La même croissance a toutefois été obtenue chez Alternaria tenuis, lorsqu'on
ajoutait le phosphate de calcium, le phosphate de sodium, le phosphate de potassium et le
phosphate d'ammonium dans un milieu de culture sans phosphore (Singh et Tandon,
1967a,b). Contrairement à H. solani, l'augmentation de la concentration de 125 à 150
Hg/ml du KH2PO4 a entraîné une diminution de 50 % de la sporulation chez Claviceps
purpurea (Kybal et al., 1968). Les niveaux très élevés de phosphore ont entraîné une
inhibition des enzymes importants dans le processus de sporulation chez ce champignon.
Ceci montre que les concentrations de phosphore qui sont optimales pour certains processus
sont inhibitrices pour certains autres (Garraway et Evans, 1984). Cela est évident pour le

48
champignon H. solani avec le Na3P04-12H20 comme source de phosphore, puisque les
concentrations élevées de phosphore qui favorisent la croissance mycélienne diminuent la
production totale de conidies et la production de conidies viables.
Les résultats relatifs à l'effet du potassium sont résumés dans le Tableau 6. Le
potassium sous forme de KCl et de K2SO4 augmente la croissance mycélienne et diminue la
production de conidies totale et la production de conidies viables à des concentrations
élevées.
Tableau 6. Effet du potassium sur le champignon Helminthosporium solani
Paramètres Sources de potassium KCl K2SO4
Croissance mycélienne
Conidies produites
Augmente avec l'augmentation de la concentration Diminuent à haute concentration
Augmente avec l'augmentation de la concentration
Diminuent à haute concentration
Conidies viables produites Diminuent à concentration Diminuent à haute concentration élevée
Contrairement à H. solani, la croissance mycélienne de Fsg n'a pas été favorisée par
le KCl et le K2SO4 (Sanogo et Yang, 2001). Ces mêmes auteurs ont également montré que
la germination in vitro n'a pas été affectée de façon significative par des éléments nutritifs
potassiques.
Les résultats relatifs à l'effet du calcium et du magnésium sont résumés dans les
Tableaux 7 et 8 respectivement. La croissance mycélienne du champignon H. solani
diminue avec l'augmentation de la concentration de calcium. La production totale de
conidies et la production de conidies viables ont, quant à elles, augmenté avec une
augmentation de la concentration peu importe la source de calcium. Le magnésium sous
forme de MgS04-7H20 a diminué le nombre de conidies viables à des concentrations
supérieures à 5 ppm sans toutefois affecter la croissance mycélienne et la production totale
de conidies.
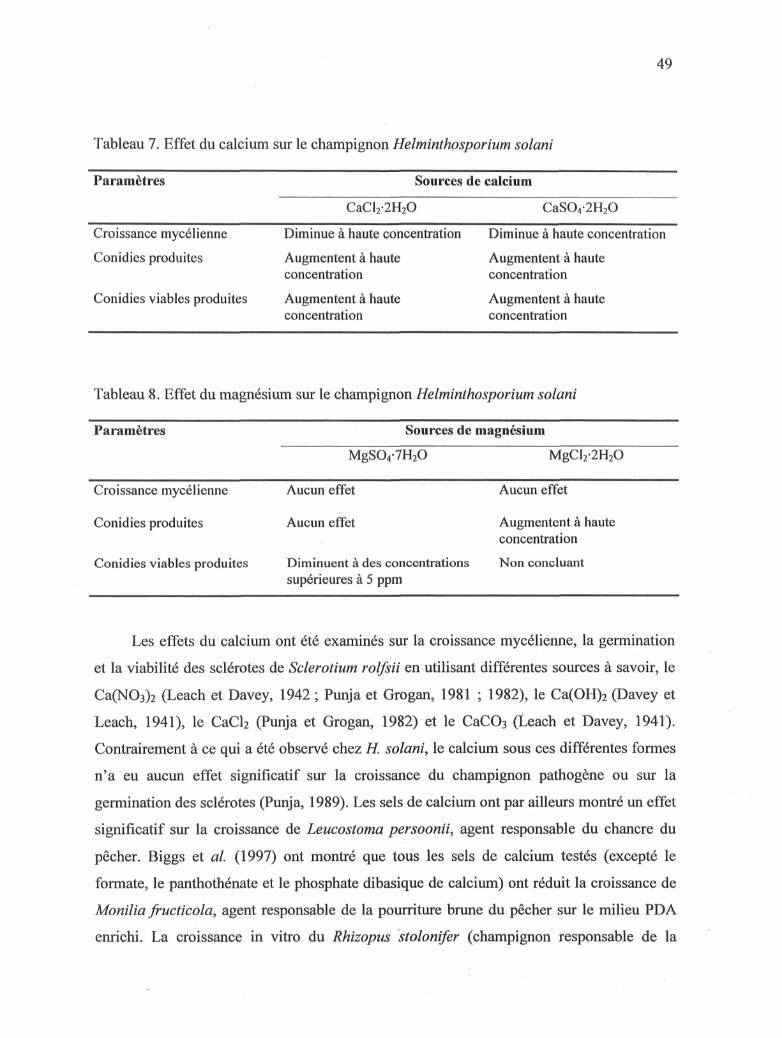
49
Tableau 7. Effet du calcium sur le champignon Helminthosporium solani
Paramètres Sources de calcium
CaCl2-2H20 CaS04-2H20
Croissance mycélienne
Conidies produites
Conidies viables produites
Diminue à haute concentration Diminue à haute concentration Augmentent à haute concentration
Augmentent à haute concentration
Augmentent à haute concentration
Augmentent à haute concentration
Tableau 8. Effet du magnésium sur le champignon Helminthosporium solani
Paramètres Sources de magnésium
MgS04-7H20 MgCl2-2H20
Croissance mycélienne
Conidies produites
Conidies viables produites
Aucun effet
Aucun effet
Diminuent à des concentrations supérieures à 5 ppm
Aucun effet
Augmentent à haute concentration Non concluant
Les effets du calcium ont été examinés sur la croissance mycélienne, la germination
et la viabilité des sclérotes de Sclerotium rolfsii en utilisant différentes sources à savoir, le
Ca(N03)2 (Leach et Davey, 1942 ; Punja et Grogan, 1981 ; 1982), le Ca(0H)2 (Davey et
Leach, 1941), le CaCb (Punja et Grogan, 1982) et le CaCOa (Leach et Davey, 1941).
Contrairement à ce qui a été observé chez H. solani, le calcium sous ces différentes formes
n'a eu aucun effet significatif sur la croissance du champignon pathogène ou sur la
germination des sclérotes (Punja, 1989). Les sels de calcium ont par ailleurs montré un effet
significatif sur la croissance de Leucostoma persoonii, agent responsable du chancre du
pêcher. Biggs et al. (1997) ont montré que tous les sels de calcium testés (excepté le
formate, le panthothénate et le phosphate dibasique de calcium) ont réduit la croissance de
Monilia fructicola, agent responsable de la pourriture brune du pêcher sur le milieu PDA
enrichi. La croissance in vitro du Rhizopus stolonifer (champignon responsable de la

50
pourriture des fruits et légumes en post-récolte) a été inhibée par le chlorure de calcium
(Tian et al, 2002) comme c'est le cas pour H. solani. Comme on peut le constater et selon
les différentes études réalisées, l'effet du calcium sur la croissance fongique dépend de la
source de calcium utilisée et du pathogène à l'étude.
Tout comme les travaux de Wisniewski et al. (1995) montrant que le chlorure de
magnésium (25-175 mM) n'a pas eu d'effet significatif sur la croissance du tube germinatif
de B. cinerea et de P. expansum, les résultats obtenus ont montré que ce sel n'a pas eu
d'effet sur la croissance mycélienne de H. solani.

51
CONCLUSION GÉNÉRALE
Ce projet qui s'inscrit dans un programme de recherche destiné à élaborer une
méthode respectueuse de l'environnement pour lutter contre la gale argentée {H. solanï) de
la pomme de terre avait pour objectif d'évaluer in vitro l'effet de l'azote, du phosphore, du
potassium, du calcium et du magnésium sur la croissance mycélienne et la production de
conidies chez le champignon H. solani. Les résultats obtenus ont permis de démontrer
l'influence de ces éléments minéraux sur le développement du champignon.
L'augmentation des concentrations d'azote et de calcium a généralement diminué la
croissance mycélienne, mais stimulé la production de conidies viables, tandis que
l'augmentation des concentrations de phosphore et de potassium a eu l'effet contraire.
À la lumière des résultats obtenus, il apparaît évident que ces travaux pourraient
trouver des applications dans la mise au point de milieux de culture favorables au
développement de ce champignon. Ils pourraient également présenter un intérêt pour
l'élaboration de substrats répressifs envers la gale argentée de la pomme de terre. À cet
effet, d'une part l'azote (NaNOs, NH4CI, NaN02 et NH4NO3) avec son fort effet inhibiteur
sur la croissance mycélienne et d'autre part le potassium (KCl et K2SO4) avec son effet
inhibiteur sur la production de conidies viables, semblent particulièrment intéressemts.
Dans les travaux fiiturs, il serait intéressant de faire une étude plus approfondie en
utilisant des concentrations sélectionnées selon une échelle plus restreinte (par exemple par
100 ppm) et de tester d'autres sources des éléments étudiés. Il serait également intéressant
d'évaluer l'effet de ces éléments sur le développement de la gale argentée et le
développement de H. solani in vivo, dans un premier temps sur un tubercule de pomme de
terre détaché et dans un deuxième temps en système de culture (en serre ou au champ).
Ainsi, les informations qui en découleront et celles obtenues in vitro pourront
éventuellement servir pour élaborer des plans stratégiques de fertilisation ou de
modification des propriétés telluriques afin de mieux contrôler la gale argentée.

52
REFERENCES BIBLIOGRAPHIQUES
Adams, A., Sander, N. et Nelson, D. C, 1970. Some propreties of soils affecting russet scab and silver scurf of potatoes. Am. Potato J. 47 : 49-57.
Agnihotri, V. P., 1968. Effects of nitrogenous compounds on Sclerotium formation in Aspergillus niger. Can. J. Microbiol. 14 : 1253-1258.
Alexopoulos, C. J. et Mims, C. W., 1979. Introductory Mycology : 3 "" édition. John Wiley & Sons, New York, NY, pages 334-566.
Avizohar-Hershenzon, A. et Shacked, P., 1969. Studies on the mode of action of inorganic nitrogenous amendments on Sclerotium rolfsii in soil. Phytopathology 59 : 288-292.
Bains, P. S., Bisht, V. S. et Bernard, D. A., 1996. Soil survival and thiabendazole sensitivity of Helminthosporium solani isolates from Alberta, Canada. Potato Res. 39 : 23-30.
Bernard, D. A. et Bains, P. S., 1996. Enzyme production by Helminthosporium solani, the causal agent of silver scurf disease on potato and its relationship to thiabendazole sensitivity. Can. J. Plant Pathol. 18 : 488.
Berry, S. Z., Madumadu, G. G. et Uddin, M. R., 1988. Effect of calcium and nitrogen nutrition on bacterial canker disease of tomato. Plant Soil 112 : 113-120.
Biggs, A. R., Ingle, M. et Solihati, W. D., 1993. Control of Alternaria infection of fruit of apple cultivar Nittany with calcium chloride and fungicides. Plant Dis. 77 : 976-980.
Biggs, A. R., El-Kholi, M. M. et El-Neshawy, S. M., 1995. Effect of calcium salts on growth, pectic enzyme activity and colonization of peach twigs by Leucostoma persoonii. Plant Dis. 79 : 886-890.
Biggs, A. R., El-Kholi, M. M., El-Neshawy, S. et Nickerson, R., 1997. Effect of calcium salts on growth, polygalacturonase activity and infection of peach fruit by Monilinia fructicola. Plant Dis. 81 : 399-403.
Biggs, A. R., 1999. Effects of calcium salts on apple bitter rot caused by two Colletotrichum spp. Plant Dis. 83 : 1001-1005.
Bisht, V. S. et Bains, P. S., 1995. Silver scurf of potato : disease dynamics over generation and fiingicidal control in the field. Can. J. Plant Pathol. 17 : 289.
Boiteau, G., 1996. Origine socio-historique de la pomme de terre. Dans La pomme de terre : production, amélioration, ennemis, maladies et utilisations. Rousselle, P., Robert., Y. et Crosnier, J. C, réds. I.N.R.A, Paris, France, pages 541-552.
Brennan, R. F., 1988. Effet of phosphorus deficiency in wheat on the infection of roots by Graeumannomyces graminis var. tritici. Aust. J. Agric. Res. 39 : 541-546 (Abstr.).
Burke, O. D., 1938. The silver scurf disease of potatoes. Cornell University Agricultural Experiment State Bulletin 692, 30 pages.

53
Burton, W. G., 1966. The potato. 2 ème édition. H. Veemnan & Zonen, Wageningen, The Netherlands, 382 pages.
Carnegie, S. F., Cameron, A. M., Hide, G. A. et Hall, S. M., 1994. The occurrence of thiabendazole resistant isolates of Polycytalum pustulans and Helminthosporium solani on seed potato tubers in relation to fungicide treatment and storage. Plant Pathol. 43 : 961-971.
Chaudhuri, S. et Maiti, S., 1978. Inhibitory activity of inorganic nitrogen sources to sclerotia of Sclerotium rolfsii. J. Plant Dis. Prot. 85 : 10-14.
Chun, W. W. C. et Shetty, K. K., 1994. Control of scurf disease of potatoes caused by Helminthosporium solani (Dur. & Mont.) with Pseudomonas corrugate. Phytopathology 84 : 1090.
CRRAQ. 2004. Répertoire 2004-2005 des traitements de protection des cultures. Centre de référence en agriculture et agroalimentaire du Québec (CRAAQ), Sainte-Foy, Canada, 296 pages.
Croes, A. F., Steijns, J. M. J. M., De Vries, G. J. M. L. et Van der Putte, T. M. J. A., 1978. Inhibition of meiosis in Saccharomyces cerevisiae by ammonium ions : Interference of ammonia with protein metabolism. Planta 141 : 205-209.
Davey, A. E. et Leach, L. D., 1941. Experiments with fungicides for use against Sclerotium rolfsii in soils. Hilgardia 13 : 523-547.
Deacon, J. W., 2006. Fungal biology. 4" edition. Institute of Cell and Molecular Biology, University of Edinburgh, Blackwell Publishing, U. K., 371 pages.
Dick, J. B. et Tisdale, H. B., 1938. Fertilizers in relation to incidence of wilt as affecting a resistant and a susceptible variety. Phytopathology 28 : 666-667 (Abstr.).
Duffy, B. K. et Défago, G., 1999. Macro- and microelements fertilizers influence the severity of fusarium crown and root rot of tomato in a soilless production system. Hort.Sci. 34:287-291.
Durieu, M.C. et Montagne, C, 1849. Flore d'Algérie, Cryptogamie, page 356. Ellis, M. B., 1961. Dematiaceous hyphomycetes. Mycol. Pap. 82 : 3-5. Elimane, R., 1996. La gale argentée de la pomme de terre. Dans La pomme de terre :
production, amélioration, ennemis, maladies et utilisations. Rousselle, P., Robert., Y. et Crosnier, J. C , réds. I.N.R.A, Paris, France, pages 296-299
Elson, M. K., Schisler, D. A. et Bothast, R. J., 1997. Selection of microorganisms for biological control of silver scurf {Helminthosporium solani) of potato tubers. Plant Dis. 81 : 647-652.
Engelhard, A. W., 1989. Historical highlights and prospects for the future. Dans soilbome plant pathogens : Management of diseases with macro- and microelements. Engelheird, A. W., red. APS Press, The American Phj^opathological Society, St. Paul, MN, pages 9-17

54
Errampalli, D., Saunders, J. M. et Holley, J. D., 2001. Emergence of silver scurf {Helminthosporium solani) as an economically important disease of potato. Plant Pathol. 50: 141-153.
Firman, D. M. et Allen, E. J., 1993. Effects of windrowing, irrigation and defoliation of potatoes on silver scmï {Helminthosporium solani) disease. J. Agric. Sci. 121 : 47-53.
Garraway, M, O. et Evans, R. C , 1984. Fungal nutrition and physiology. Wiley-Interscience Publication. John Wiley & Sons, New York, NY, 401 pages.
Garraway, M. O. et Evans, R. C, 1991. Fungal nutrition and physiology. Krieger Publishing Company, Malabar, FL, pages 71-211.
Gaucher, D., 1998. Maladies de la pomme de terre. Institut technique des céréales et des fourrages. Paris, 48 pages.
Griffin, D. H., 1994. Fungal physiology. Wiley-Liss, John Wiley & Sons, New York, NY, pages 130-157.
Harz, I., 1871. Spondylocladium atrovirehs. Bulletin of Society Imperial, Moscow 44 : 42. Heiny, D. K. et Mclntyre, G. A., 1983. Helminthosporium solani (Dur. & Mont.)
development on potato periderm. Am. Potato J. 60 : 773-789. Hervieux, V., 2000. Stratégies de lutte contre la tache argentée de la pomme de terre:
traitements fongicides de la semence et application post-&colte de sels organiques et inorganiques. Mémoire de Maîtrise, Faculté des Sciences de l'Agriculture et de l'Alimentation, Université Laval, 125 pages.
Hide, G. A. et Boorer, K. J., 1991. Effects of drying potatoes (Solanum tuberosum L.) after harvest on the incidence of disease after storage. Potato Res. 34 : 133-137.
Hide, G. A., Boorer, K. J., et Hall, S. M. 1994. Controlling potato tuber blemish diseases on cv. Estima with chemical and non-chemical methods. Ann. Appl. Biol. 124 : 253-265.
Hide, G. A., Hall, S. M. et Boorer, K. J., 1988. Resistance to thiabendazole in isolates of Helminthosporium solani, the cause of silver scurf disease of potatoes. Plant Pathol. 37 : 377-380.
Hoffland, E., Van Beusichem, M. L. et Jeger, M. J., 1999. Nitrogen availability and susceptibility of tomato leaves to Botrytis cinerea. Plant Soil 210 : 263-272.
Hoffland, E., Jeger, M. J. et Van Beusichem, M. L., 2000. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 218 : 239-247.
Huber, D. M. et Watson, R. D., 1974. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 12 : 139-165.
Huber, D. M., 1978. Disturbed mineral nutrition. Dans Plant Pathology, An Advanced Treatise. Vol. III. Horsfall, J. G. et Cowling, E. B., reds. Academic Press, New York, NY, pages 163-181.
Huber, D. M., 1980. Role of nutrients in defense. Dans Plant Pathology, An Advanced Treatise. Vol. V. Horsfall, J. G. et Cowling, E. B., reds. Academic Press, New York, NY, pages 381-406.

55
Huber, D. M., 1989. Introduction. Dans Soilbome plant pathogens : Management of diseases with Macro- and Microelements. Engelhard, A. W., red. APS Press, The American Phytopathological Society, St. Paul, MN, pages 1-8.
Hunger, R. M. et Mclntyre, G. A., 1979. Occurrence, development and losses associated with silver scurf and black dot on Colorado potatoes. Am. Potato J. 56 : 289-306.
Jellis, G. et Taylor, G. S., 1977. The development of silver scurf (Helminthospohum solani) disease of potato. Ann. Appl. Biol. 86 : 19-28.
Johnson, S. P., 1953. Some factors in the control of the southern blight organism, Sclerotium rolfsii. Phytopathology 43 : 363-368.
Jones, J. P., Engelhard, A. W. et Woltz, S. S., 1989. Management of Fusarium wilt of vegetables and ornamentals by macro- and microelement nutrition. Dans Soilbome Plant Pathogens : Management of Diseases with Macro- and Microelements. Engelhard, A. W., red. APS Press, The American Phytopathological Society, St. Paul, MN, pages 18-32.
Jouan, B., Lemaire, J. M., Perennec, P. et Sailly, M., 1974. Études sur la gale argentée de la pomme de terre {Helminthosporium solani Dur. & Mont.). Ann. Phytopathol. 6 : 407-423.
Kamara, A. M. et Huguelet, J. E., 1972. Host range and overwintering of Helminthosporium solani. Am. Potato J. 49 : 365.
Kawchuck, L. M., Holley, J. D., Lynch, D. R. et Clear, R. M., 1994. Resistance to thiabendazole and thiophanate-methyl in Canadian isolates of Fusarium sambucinum and Helminthosporium solani. Am. Potato J. 71 : 185-192.
Kirk, T. K., 1971. Effects of microorganisms on lignin. Ann. Rev. Phytopathol. 9 : 185-210.
Kybal, J., Majer, J., Komersova, I. et Wani, W. D., 1968. Phosphorus content during development of Clavicepspurpurea. Phytopathology 58 : 647-650.
Lambert, D. H., Powelson, M. L. et Stevenson, W. R., 2005. Nutritional interactions influencing diseases of potato. Am. J. Potato Res. 82 : 309-319.
Lange, R. M., Bisht, V. S., Chen, Z., Bains, P. S., Kawchuck, L. M. et Lynch, D. R., 1995. Antibiosis of Bacillus polymyxa and its metabolites against various potato pathogens. Can. J. Plant Pathol. 17 : 358.
Leach, L. D. et Davey, A. E., 1942. Reducing southern sclerotium rot of sugar beets with nitrogenous fertilizers. J. Agric. Res. 64 : 1-18.
Martinez, C, Michaud, M., Bélanger, R. R. et Tweddell, R. J., 2002. Identification of soils suppressive against Helminthosporium solani, the causal agent of potato silver scurf. Soil Biol. Biochem. 34 : 1861-1868.
Mérida, C. A. et Loria, R., 1990. First report of resistance of Helminthosporium solani to thiabendazole in the United States. Plant Dis. 74 : 614.
Mérida, C. A. et Loria, R., 1994. Comparison of thiabendazole sensitive and resistant Helminthosporium solani isolates from New York. Plant Dis. 78 : 187-192.

56
Michaud, M., 2001. Étude des sols suppressifs envers Helminthosporium solani, agent responsable de la tache argentée de la pomme de terre. Mémoire de Maîtrise, Faculté des Sciences de l'Agriculture et de l'Alimentation, Université Laval, 119 pages.
Michaud, M., Martinez, C, Simao-Beaunoir, A. M., Bélanger, R. R. et Tweddell, R. J., 2002. Selection of antagonist microorganisms against Helminthosporium solani, causal agent of poatato silver scurf Plant Dis. 86 : 717-720.
Morton, A. G. et MacMillan, A., 1954. The assimilation of nitrogen from ammonium salts and nitrate by fiingi. J . Exp. Bot. 5 ; 232-252.
Nam, M. H., Jeong, S. K., Lee, Y. S., Choi, J. M. et Kim, H. G., 2006. Effects of nitrogen, phosphorus, potassium and calcium nutrition on Straw^berry anthracnose. Plant Pathol. 55 : 246-249.
Nicholas, D. J. D., 1965. Utilization of inorganic nitrogen compounds and amino acids by fungi. Dans The Fungi, Vol 1., Ainsworth, G. C. et Sussman, A.S., reds.. Academic Press, New York, NY, pages 349-376.
Pateman, J. A. et Kinghorn, J. R., 1976. Nitrogen metabolism. Dans The Filamentous Fungi, Vol. 2., Smith, J. E. et Berry, D. R., reds., Wiley, New York, NY, pages 159-237.
Perlman, D., 1965. The chemical environment for fungal grov^h. 2 : Carbon sources. Dans The Fungi, Vol. 1, Ainsworth, G. C. et Sussman, A. S., reds.. Academic Press, New York, NY, pages 479-490.
Pinon, R., 1977. Effects of ammonium ions on sporulation of Saccharomyces cerevisiae. Exp. Cell. Res. 105 : 367-378.
Piatt, H. W., 1997. Resistance to thiabendazole in Fusarium species and Helminthosporium solani in potato tubers treated commercially in eastern Canada. Phytoprotection 78 : 1-10.
Punja, Z. K., 1989. Influence of nitrogen and calcium compounds on development of disease due to Sclerotium rolfsii. Dan^ Soilbome Plant Pathogens : Management of Diseases with Macro- and Microelements. Engelhard, A. W., red. APS Press, The American Phytopathological Society, St. Paul, MN, pages 75-89.
Punja, Z. K. et Grogan, R.G., 1981. Mycelial growth and infection withouth a food base by eruptively germinating sclerotia of Sclerotium rolfsii. Phytopathology 71 : 1099-1103.
Punja, Z. K. et Grogan, R. G., 1982. Effects of inorganic salts, carbonate-bicarbonate anions, and ammonia, and the modifying influence of pH on sclerotial germination of Sclerotium rolfsii. Phytopathology 72 : 635-639.
Randall, C. R., 1993. Potato Health Management : A Holistic Approach. Dans Potato Health Management. Randall, C. R., red. APS Press. The American Phytopathological Society, St. Paul, MN, pages 3-18.
Read, P. J. et Hide, G. A., 1984. Effects of silver scmf {Helminthosporium solani) on seed potatoes. Potato Res. 27 : 145-154.

57
Richard, C. et Boivin, G., 1994. Maladies et ravageurs de cultures légumières au Canada. La société canadienne de Phytopathologie et la Société d'Entomologie du Canada, Ottawa, Ontario, 590 pages.
Rodriguez, D. A., Secor, G. A. et Nolte, P., 1990. Resistance of Helminthosporium solani isolates to benzimidazole fungicides. Am. Potato J. 67 : 574-575.
Rodriguez, D. A., Secor, G. A., Gudmestad, N. C. et Grafton, K., 1995. Screening tuberbearing solatium species for resistance to Helminthosporium solani. Am. Potato J. 72 : 669-679.
Rodriguez, D. A., Secor, G. A., Gudmestad, N. C. et Franci, L. J., 1996. Sporulation of Helminthosporium solani and infection of potato tubers in seed and commercial storages. Plant Dis. 80 : 1063-1070.
Saccardo, P.A., 1886. Sylloge Fungorum, Vol. IV, page 483. Sagdullaev, M. M. et Berezhnova, W., 1974. The effects of phosphorus fertilizers on the
physiological properties, yield and resistance of melons to fusarium wilt. Agrokhimia 4 : 36-40.
Sanogo, S. et Yang, X. B., 2001. Relation of sand content, pH and potassium and phosphorus nutrition to the development of sudden death syndrome in soybean. Can. J. Plant Pathol. 23 : 174-180.
Santerre, J., 1967. Evaluation of the relative importance of soil and sowing as a source of inoculum in silver scurf repression tests. Can. J. Plant Sci. 47 : 695-702.
Sestakova, M., 1976. Assimilation spectrum of the yeast Candida utilis 49 used for producing fodder yeast from synthetic ethanol. Folia Microbiol. 21 : 384-390.
Schneider, R. W., 1985. Suppression of Fusarium yellows of celery with potassium, chloride and nitrate. Phytopathology 75 : 40-48.
Secor, G. A., 2006. Department of Plant Pathology. North Dakota State University, http : www.ndsu.nodak.edu/instruct/gudmesta/lateblight/basic-frame.htm (Consulté en septembre 2006).
Secor, G. A. et Gudmestad, N. C, 1999. Managing fungal diseases of potato. Plant Pathol. 21 : 213-221.
Secor, G. A., Gudmestad, N. C. et Rodriguez, D. A., 1996. North Dakota State University making progress on management of silver scurf. Valley Potato Grower, juillet, pages 26-27.
Shamis, D. L., Gelikova, N. S. et Sarsenova, L. N., 1968. Effect of different sources of phosphorus nutrition on biomass of fodder yeasts and the nitrogen and phosphorus content of them. Trans. Inst. Microbiol. Virisol. Akad. Nauk. Kay. USSR 11 : 18-24.
Shipton, W. A., 1977. Some nutritional factors regulating formation of fertile perithecia of Calonectria camelliae. Trans. Brit. Mycol. Soc. 69 : 59-62.
Singh, A., 1968. Studies on Helminthosporium solani, the causal organism of silver scurf of potato. Ph. D. Thesis, North Dakota State University, ND, 60 pages.

58
Singh, A., 1973. Effet of vitamins and carbon and nitrogen sources on the growth of Helminthosporium solani. Ind. Phytopathol. 26 : 510-516.
Singh, B. P. et Tandon, R. N., 1967a. Sulfur and phosphorus requirements of Alternaria tenuis Auct. isolated from papaya (Carica papaya L.) leaf Proc. Natl. Acad. Sci. India, Sect. B 37 : 199-203.
Singh, B. P. et Tandon, R. N., 1967b. Phosphorus requirements of certain isolates of Alternaria tenuis Auct. Proc. Natl. Acad. Sci. India, Sect. B 37 : 131-134.
Spire, D. et Rousselle, P., 1996. Origine socio-historique de la pomme de terre. Dans La pomme de terre : production, amélioration, ennemis, maladies et utilisations. Rousselle, P., Robert., Y. et Crosnier, J. C, réds., I.N.R.A, Paris, France, pages 1-48.
Statistique Canada, 2007. Division de l'Agriculture. Bulletin de la production canadienne de la pomme de terre. Catalogue 22-008 http://www.statcan.ca. (Consulté le 24 janvier 2007).
Stevick Haux, C. S., 1998. Integrated management practices control silver scurf Valley Potato Grower, May, pages 12-30.
Taylor, G. A., 1954. The effects of three levels of magnesium on the nutrient element composition of two inbred lines of com and on their susceptibility to Helminthosporium maydis. Plant Physiol. 29 : 87-91.
Tian, S. P., Fan, Q., Xu, Y. et Jiang, A. L., 2002. Effects of calcium on biocontrol activity of yeast antagonists against the postharvest fungal pathogen Rhizopus stolonifer. Plant Pathol. 51 : 352-358.
Wisniewski, M., Droby, S., Chalutz, E. et Eilam, Y., 1995. Effects of Ca ^ and Mg^^ on Botrytis cinerea and Pénicillium expansum in vitro and on the biocontrol activity of Candida oleophila. Plant Pathol. 44 : 1016-1024.
Whitaker, A., 1976. Amino acid transport in fungi : An essay. Trans. Brit. Mycol. Soc. 67 : 365-376.
Woltz, S. S. et Jones, J. P., 1973a. Interactions in source of nitrogen fertilizer and liming procedure in the control of Fusarium wilt of tomato. HortScience 8 : 137-138.
Woltz, S. S. et Jones, J. P., 1973b. Tomato Fusarium wilt control by adjustments in soil fertility. Proc. Fla. State Hort. Soc. 86 : 157-159.
Woltz, S. S. et Jones, J. P., 1973c. Tomato Fusarium wilt control by adjustments in soil fertility : A systematic approach to pathogen starvation. Agric. Res. Ed. Center Bradenton Res. Rept. GC 1973-1977.
Woltz, S. S. et Jones, J. P., 1981. Nutritional requirements oî Fusarium oxysporum : Basis for a disease control system. Dans Fusarium : Diseases, Biology and Taxonomy. Nelson, P. E., Toussoun, T. A. et Cook, R.J., réds., Perm. State University Press, University Park, pages 340-349.
























![minéraux cristallographie[1]](https://img.dokumen.tips/doc/110x75/5571fac1497959916993061e/mineraux-cristallographie1.jpg)




![Fossiles - La Combelle] : Accueil€¦ · minéraux et insectes. Insectes. Différents papillons. ... ainsi qu'une expo ion.ðe papil ons ,d'insectes . Exposition e rmneraux , ossi](https://img.dokumen.tips/doc/110x75/5f0e42887e708231d43e6117/fossiles-la-combelle-minraux-et-insectes-insectes-diffrents-papillons.jpg)