Embed Size (px)
Citation preview

J. Insect Physiol. Vol. 39, No. 8, PP. 657-664. 1993 Printed in Great Britain
0022-1910/93 $6.00+0.00 Pergamon Press Ltd
Effects of Soybean Protease Inhibitors on the Growth and Development of Larval Helicoverpa armigera K. A. JOHNSTON,* J. A. GATEHOUSE,* J. H. ANSTEE*
Received 18 September 1992; revised 25 January 1993
Soybean Kunitz trypsin inhibitor (SBTI) and soybean BowmawBirk trypsin-cbymotrypsin inhibitor (SBBI) were incorporated into artificial diet and fed to newly moulted 3rd~instar larvae of Helicouerpa armigera, the corn earworm.
1. Neither SBTI nor SBBI affected larval survival for 14 days. After this time, many of the larvae fed diets with high levels of inhibitor suddenly died. This effect was most marked with SBTI diets.
2. When compared with the effect of the control diet (no inhibitor), inhibitors in the diet reduced total larval biomass and mean larval weight. This effect on larval growth was much greater with dietary SBTI than with SBBI. The relative effectiveness of the inhibitors in oioo is in agreement with previous data showing that SBTI was a more effective inhibitor of the H. armigera trypsin-like gut protease in vitro.
3. The presence of inhibitor in the diet reduced the mean larval weight at moult, suggesting a possible response of ecdysis to semi-starvation. This effect was greater with SBTI than with SBBI.
4. Faecal output was used as a measure of food intake. By day 10 of the feeding trials, the mean faecal output/larva was similar for larvae fed the control and SBBI diets, but it was significantly lower for larvae fed dietary SBTI. However, when faecal output was corrected to account for developmental states and weights of larvae, no differences were found between the effects of control and inhibitor diets.
5. Feeding larvae a diet containing 0.234 mM SBTI significantly reduced the trypsin-like enzyme activities found in their gut contents when compared with the levels found in larvae fed the control diet.
The possible mechanisms of action of SBTI and SBBI in aiuo are discussed, as are their relative potentials for protecting crop plants against damage by larval H. urmigera.
Helicoverpa armigera Growth Development Protease inhibitors
INTRODUCTION
Helicoverpa armigera, the corn earworm, is a major pest, particularly on cotton and legumes (Fitt, 1989). In common with many other species of insect pest, H. armigera tends to develop resistance to a range of pesticides. The development and use of crop cultivars that are resistant to pest damage is an increasingly important complement to the chemical control of such pests.
Incorporating or increasing levels of plant protease inhibitors in crop plants has potential for increasing crop resistance to pests (Ryan, 1990). Serine protease activity has been found in a wide variety of lepido- pteran pests, including H. armigera (Johnston et al.,
*Department of Biological Sciences, University of Durham, South Road, Durham DHl3LE, U.K.
1991). Many of these proteases are inhibited by plant serine protease inhibitors (Ahmad et al., 1980; Sasaki and Suzuki, 1982; Hamad and Attias, 1987; Broadway, 1989; Houseman et al., 1989; Johnston et al., 1991). Various serine protease inhibitors have also been shown to adversely affect the growth and development of lepidopteran larvae (Shukle and Murdock, 1983; Broadway and Duffey, 1986a; Hilder et al., 1987; Johnson et al., 1989). Furthermore, Hilder et al. (1987) showed that the introduction and expres- sion of the cowpea trypsin inhibitor gene into tobacco significantly reduced damage caused by Heliothis virescens.
A previous study (Johnston et al., 1991) reported the presence of serine protease activity in midgut of larvae of H. armigera and characterized the effects of various plant protease inhibitors on this activity. These results are summarized in Table 1.
657

658 K. A. JOHNSTON et al
TABLE I Art$cial larval diet Maximal level
Ki of inhibition Inhibitor (in M BApNA) (%)
Cowpea protease inhibitor 8.8 x 10-s 80 Lima bean trypsin inhibitor 1.9 x lo-’ 90 Soybean Bowman-Birk inhibitor 2.5 x IO-’ 90 Soybean trypsin inhibitor 9.6 x 1O-9 99
Larvae were fed an artificial diet of the composition shown in Table 2 (essentially that of Shell Research Ltd).
Feeding trials, where pure enzyme inhibitors are incor- porated in artificial diets and fed to insects, provide valuable information about which types of inhibitor have potential as resistance factors in genetic manipu- lation and plant breeding programmes. For instance, Larocque and Houseman (1990) demonstrated that serine protease inhibitors from various sources had very variable effects on growth of larvae of Ostrinia nubilalis. They also stress the importance of determining the biochemical interactions between plant inhibitors and insect proteases in order to understand the potential antimetabolic effects of these inhibitors.
The stock vitamin solution was Vanderzant’s Fortifi- cation Mixture (Singh, 1977). All the ingredients, except the agar and water, were mixed well with 34 ml of the distilled water. If inhibitor was being added, the inhibi- tor was first dissolved in a little of the water, prior to mixing with the above ingredients. The agar was dissolved, with heating, in 54ml of the distilled water and allowed to cool to 60°C. It was then quickly added to and mixed with the other dietary ingredients, detailed in Table 2, to produce a smooth even texture. The diet was then poured into Petri dishes and allowed to set.
Feeding trials
The present paper details results from feeding trials, which determined the effects of soybean trypsin inhibitor (SBTI) and soybean Bowman-Birk inhibitor (SBBI), on growth and development of larvae of H. armigera. SBTI, a Kunitz trypsin inhibitor, was chosen for the bioassay because it was 10-20 times more effective against the protease activity from midgut of larvae of H. armigera than the other inhibitors tested (see above). The trypsin- chymotrypsin Bowman-Birk inhibitor, SBBI, was less effective than SBTI at inhibiting the larval protease in vitro. Thus, the bioassays comparing the relative effectiveness of SBTI and SBBI in vivo help determine whether in vitro assays reflect the true potential of the inhibitors as resistance factors. Also, SBTI and SBBI are derived from the same crop, Glycine max, but they belong to two different major classes of serine protease inhibitor.
Studies on the effects of diet on growth and develop- ment were carried out using 3rd-instar larvae of known age. Insects of known age were collected as follows: newly moulted 2nd-instar larvae were isolated from the general population of larvae and were then examined at 9:00 and 18 : 00 h daily; any 3rd-instar larvae which appeared between sampling times were removed and placed in labelled cages. At the time of collection, 3rd~instar larvae were weighed and were divided into control and experimental groups. The same number of larvae from each chronological age were placed on each test diet, ensuring that variation between populations fed on each diet was minimized. Ten larvae were placed on each diet.
The larvae were initially placed in small, airtight plastic containers (3 cm dia x 4 cm high) with a cube of food of approx. 5 mm3. By the time larvae had moulted to the 4th instar, small airholes had been made in the lids and the amount of food increased to 10 mm3. Food was provided daily and in cases where little diet was eaten, the food was changed every 2 days to reduce microbial contamination.
MATERIALS AND METHODS
Soybean trypsin inhibitor (SBTI) (Catalogue No. T9003), soybean Bowman-Birk inhibitor (SBBI) (Catalogue No. T9777) and Ncr benzoyl-DL-arginine p - nitroanilide (BApNA) were obtained from the Sigma Chemical Co. Haricot beans and Brewers yeast were obtained locally. All other chemicals were obtained from British Drug Houses (BDH) Ltd, Poole, Dorset, U.K.
The control diet used was that outlined above with no inhibitor added. The levels of inhibitor used in the diet, expressed as percentage weight of inhibitor per wet weight of diet (w/w) (and corresponding mM con- centration), were as follows: SBTI; 0.1% (0.047 mM), 0.5% (0.234mM) and 2.0% (0.935 mM). SBBI;
Insect cultures
The insects used were larval H. armigera, and these were reared at 26°C 40% r.h., using Insect Rearing Method (SCP/3) of Shell Research Ltd, Sittingbourne, Kent, U.K. The original stock population was obtained from J. Radcliffe, Shell Research Ltd. The Ministry of Agriculture, Food and Fisheries (MAFF) licence number for obtaining and culturing H. armigera was PHF 346/166/(70).
Weight/volume Ingredient
1.15g Agar (Oxoid No. 1)
20g Finely ground haricot bean seed 3.2 g Finely powdered Brewers yeast 0.1 g Sorbic acid 0.2 g Methyl-4-hydroxybenzoate 0.32 g Ascorbic acid
lg Aureomycin 0.26 ml 10% formaldehyde solution 0.67 ml Stock vitamin solution (see above) 0.8 ml Linseed oil
88 ml Distilled water
TABLE 2

EFFECT OF INGESTED SOYBEAN PROTEASE INHIBITORS 659
0.031% (O.O37mM), 0.152% (0.190mM) and 0.606% (0.750 mM).
Individual weight and developmental state were monitored at the same time each day, starting with the newly moulted 3rd-instar larvae. The faecal output was collected at the same time each day, freeze-dried and then weighed. Dried faecal weight was taken as an indication of food intake.
Estimation of serine protease activities in guts of larvae
Midgut serine protease activity was determined at various times (12 f 4.5 h) throughout the 3rd stadium: newly moulted 3rd-instar larvae were collected and divided into control-fed and inhibitor-fed (diet contain- ing 0.234 mM SBTI) groups. At known times, five larvae from each group were killed by decapitation and their gut contents dissected out. The gut contents from each larva were homogenized separately, in 500 ~1 distilled water. The proteins in these homogenates were then precipitated with 1 ml cold acetone and stored at -20°C. Five insects from each of the two groups were sampled in this way at known times throughout the 3rd stadium. When about 70-80% of remaining larvae had moulted to the 4th instar, the assay was terminated.
The acetone precipitated proteins were centrifuged down, for 15 min, at 8OOOg, and then redissolved in 100 ~1 distilled water. The amount of protein in the solutions was estimated by measuring absorbances at 260 and 280 nm and using the nomograph of Warburg and Christian (Dawson et al., 1986). Serine protease activity was assayed for at pH 9.8 [determined optimum pH for the enzyme activity (Johnston et al., 1991)] in borate buffer, with 1 mM BApNA in 50% dimethyl- formamide as substrate. The assays were carried out at 30°C in a Titretek plate. Incubation media consisted of 200~1 buffer, 20~1 substrate and the reaction was started by addition of 10 ~1 gut extract. Enzyme activity was determined by measuring the change in absorbance at 405 nm, over a 30min incubation time, using a Multiscan Titretek plate reader.
Statistical treatment of data
Analysis of variance consisted of the F-test, followed by the Student-Newman-Keuls multiple range test wherever the F-test showed the means to be signifi- cantly different. In all cases, 95% confidence limits (0.05-a P < 0.05) were used.
RESULTS
E#ect of inhibitors on larval survival
Figure 1 shows differential survival of larvae fed control and experimental diets. With both inhibitors, there was no identifiable effect on larval survival for approx. 14 days. After this time a rapid effect on survival was observed with 70% of the larvae fed 0.750 mM SBBI and 0.234 mM SBTI, and 100% of those fed 0.935 mM SBTI, dead by 22 days. Thus, SBTI was the more potent
80
60
80
60
40
20
0
(4
0 5 10 15 20 25
TIME (Days)
(W
I 1 I I I I 1 0 5 10 15 20 25
TIME (Days) FIGURE 1. The effects of dietary inhibitors on larval survival. (A) With SBTI at the levels of 0.047mM (a), 0.234mM (A) and 0.935mM (0). (B) With SBBI at the levels of 0.038mM (Cl), 0.190 mM (A) and 0.750 mM (0). The control diet contained no inhibitor (m). Ordinate: % survival. Abscissa: time in days from the
start of the 3rd instar.
agent in reducing larval survival. The LD,,, calculated after 18 days, was 0.46mM for dietary SBTI and 0.75 mM for dietary SBBI.
E#ect of inhibitors on larval biomass and growth
The inclusion of the serine proteinase inhibitors, SBTI and SBBI, in diets, resulted in a reduction in total larval biomass produced when compared with that of control- fed insects (Fig. 2). This effect was most marked with SBTI diets, with 0.047 mM SBTI effecting a reduction of 64.1% in total larval biomass by day 14. Much higher levels of SBBI were needed to achieve a similar reduction

660 K. A. JOHNSTON et al.
(4 2000
z
z 1500
I ._ m 1000 3 ir 1
2000
E
3 1500
I ii 1000
3 k -I Z 500
z I-
O
l-
0 0 5 10 15 20
TIME (Days)
(B)
0 5 10 15 20
TIME (Days)
FIGURE 2. The effect of dietary inhibitors on total larval biomass produced. (A) With SBTI at the levels of 0.047 mM (O), 0.234 mM (A) and 0.935 mM (0). (B) With SBBI at the levels of 0.038 mM (IJ), 0.19OmM (A) and 0.750mM (0). The control diet contained no inhibitor (m). Ordinate: total larval biomass, in mg. Abscissa: time in
days from the start of the 3rd instar.
of total larval biomass in this time; 0.750 mM SBBI reducing total larval biomass by 50%. Despite these effects on total larval biomass, the presence of inhibitors in the diet had no effect on larval mortality by day 14 (Fig. 1). Thus, the substantial differences in total larval biomass over this period were due to the effects of the inhibitors on larval growth.
After day 14, the presence of high levels of inhibitors in the diets increased larval mortality as well as affecting larval growth, so that in the case of larvae fed diets containing 0.234 mM and 0.935 mM SBTI, there was a decline in total larval biomass (Fig. 2). With high levels
of SBBI, the larvae which died were the smallest in the population, and so the increased mortality after day 14 had less effect on total larval biomass than it had with SBTI.
Figure 3 shows the effect of SBTI in the diets on larval growth with time. It can be seen that larval growth, as indicated by an irreversible increase in weight, was greatly reduced by the the inclusion of this inhibitor in the diet. Analysis of variance indicates that 0.047 mM SBTI diets significantly (P < 0.05) reduced the mean larval weight when compared with that observed with the control diet. Higher concentrations of SBTI in the
(4 200 -
200 1 (B)
5 10 15 20
TIME (Days)
0 5 10 15 20
TIME (Days)
FIGURE 3. The effect of dietary inhibitor on larval growth, expressed as average larval weight. (A) With SBTI at the levels of 0.047 mM (n), 0.234mM (A) and 0.935mM (0). (B) With SBBI at the levels of 0.038 mM (a), 0.190 mM (A) and 0.750 mM (0). The control diet contained no inhibitor (a). Ordinate: mean larval weight, in mg. Abscissa: time in days from the start of the 3rd instar. Error bars:
&standard error of the mean.

EFFECT OF INGESTED SOYBEAN PROTEASE INHIBITORS
(A) 3 200 -
(61 T
3rd to 4th moult 4th to 5th moult 5th to 6th mouk
MOULTS
W
3rd to 4th moult 4th to 5th mouk 5th to 6th moult
MOULTS
661
FIGURE 4 The effects of dietary inhibitor on average larval weight at moult. (A) With SBTI at the levels of 0.041 mM ( 0.234mM (m) and 0.935 mM (B). (B) With SBBI at the levels of 0.038 mM ( ), 0.190mM (m) and 0.750mM (m). The control diet contained no inhibitor (0). Ordinate: average weight of larvae, on moulting (in mg). Abscissa: moults. Error bars: *standard error of the mean. Above each column, the number of larvae which had moulted to the next instar, is given
in parentheses.
diet tended to further reduce the rate of increase in mean larval weight, although these further reductions were not statistically significant (P 2 0.05). Dietary SBBI also reduced mean larval weight compared with controls, but this reduction was not statistically significant (P 2 0.05), and whereas a progressive decrease in larval weight with SBTI concentration was observed, with SBBI the highest concentration of inhibitor (0.750 mM) gave a reduction in mean larval weight intermediate between those given by 0.035 mM SBBI (least reduction) and 0.190 mM SBBI (greatest reduction).
The presence of protease inhibitor in the diet reduced the mean weight at which larvae moulted (Fig. 4). At the 4th larval moult, this response was significant (P < 0.05)
in larvae fed high levels of SBTI, and by the 5th moult, all larvae fed on diets containing SBTI, even at the lowest concentration, were significantly (P < 0.05) smaller than those fed the control diet. The number of moults successfully completed was also reduced as the inhibitor concentrations in the diets increased (Fig. 4); no larvae fed the highest level of SBTI moulted to the 6th instar. Until the 5th moult, dietary SBBI had no significant effect (P > 0.05) on the mean weight of larvae at the moult. At the 5th moult, the larvae fed 0.035 mM SBBI and 0.190 mM SBBI were significantly (P < 0.05) lighter than those fed the control diet. However, larvae fed the highest concentration of SBBI, 0.750 mM, were not significantly (P > 0.05) different in weight from

662 K. A. JOHNSTON et al.
those fed the control diet. This was probably due to selective survival of the largest individuals from groups which had been fed on diets containing SBBI, as noted above; the degree of selectivity increased as concen- tration of SBBI in the diet was increased.
Effect of inhibitors on food intake
Faecal output was used as a measure of food intake. The mean faecal output per larva per 24 h was estimated over the initial lo-day period of the feeding trial. The data for day 1, which demonstrate the immediate effects of dietary inhibitor on larval food intake, showed no significant (P > 0.05) difference between treatments in amounts of faeces produced (0.8-2.0 mg faeces per larva per day) at this time. By day 10, the faecal output of larvae fed on control diets was not significantly different from those fed diets containing SBBI (7-14 mg faeces per larva per day). In contrast, larvae fed diets containing SBTI showed significantly (P < 0.05) lower levels of faecal output. The mean faecal output of larvae fed 0.935 mM SBTI increased only approx. twofold between days 1 and 10, whereas that of the control-fed insects had increased by about sevenfold over this period. However, no significant differences were found between the treat- ments when faecal output was expressed per unit weight of larva, at the start of the 3rd stadium and at the midpoints of the 3rd and 4th stadia. This suggests that neither inhibitor had a significant effect on food intake when weight and developmental state of the larvae were taken into consideration.
The accumulative effect of dietary inhibitors on total faecal output was also measured. By day 10, low levels of SBTI (0.047 mM) in the diet had reduced total faecal output by 60%. Although increases in SBTI level caused a progressive reduction in faecal output, the highest level of dietary SBTI tested (0.935 mM) only reduced total output by a further 10%. Reduction in the total faecal output was much less for SBBI; levels of 0.038 mM in the diet reduced total faecal output by approx. 13%, while the higher levels of SBBI (20.19 mM) reduced faecal output by approx. 25%.
Eflect of dietary SBTI on larval serine protease activity
The presence of SBTI in the diet, at levels of 0.5% of the wet weight (0.234 mM), significantly (P < 0.05) reduced serine proteinase activity, as measured by BApNAase activity, in the guts of larvae fed this diet, compared with that of larvae fed the control diet (Fig. 5). Thus, since the serine protease activity detected by BApNA represents at least 80% of total proteolysis in H. armigera (Johnston et al., 1991), SBTI has been shown to inhibit proteolysis strongly in vivo. After moulting to the 3rd stadium, the serine protease activity tended to increase in larvae fed the control diet. This response was suppressed in larvae fed SBTI. The protein levels in the gut contents of larvae fed diets containing SBTI were only slightly lower than those of larvae fed the control diet, apart from those measured at 30 h after the 3rd moult (data not presented). At 30 h, the protein
-
3rd M,
7 0 15 30 45 60 75 9a
Y TIME (hrsl after 2nd MOULT
FIGURE 5. The protease activity in the gut contents of 3rd~instar larvae fed diets with (n---n) and without (W--W) SBTI at 0.234 mM. Ordinate: the mean protease activity, measured by hydroly- sis of BApNA, at pH 9.8, 30°C expressed as AOD,,,, x 10’ per min per larval gut. Absicca: time in hours after the 2nd mouh. Error bars:
*standard error of the mean.
levels in the control animals was much higher than those of the inhibitor-fed group.
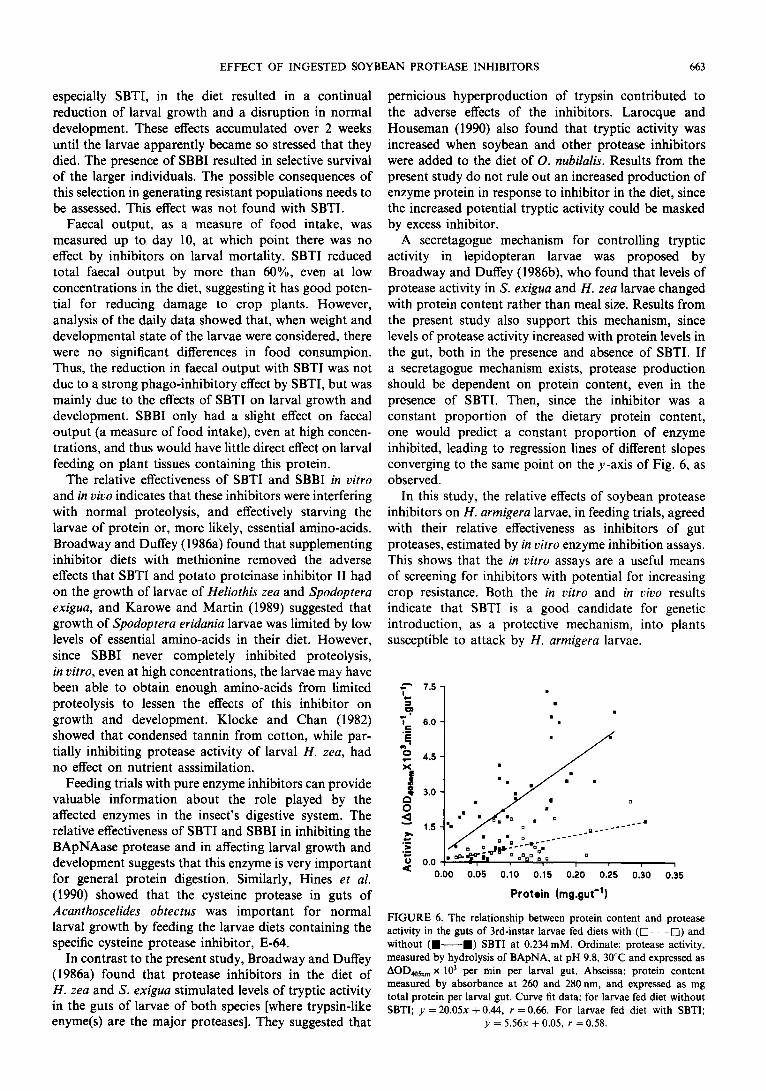
For the control and experimental groups, the total activity of protease in the gut was plotted against total protein levels in the gut contents of each larva (Fig. 6). There was a direct relationship between protease activity and protein levels in the larval gut in each treatment. The slopes of the regression lines calculated for each treat- ment were significantly different, with a decrease of approx. fourfold in the increase in amount of protease per amount of protein in larvae fed on diet containing SBTI, compared with control animals.
DISCUSSION
In agreement with earlier results that SBTI was a more effective inhibitor of H. armigera protease activity in vitro than SBBI (Johnston et al., 1991), the results of this study show that SBTI was also much more effective than SBBI in reducing larval survival and growth, and in affecting larval development. For instance, after 14 days, the biomass of larvae fed 0.047 mM SBTI in their diet was more than 50% lower than that of control larvae. A concentration of 0.750mM SBBI was needed in the diet to achieve the same effect. The presence of inhibitors in the diet significantly reduced the average weight of larvae at moult, with SBTI being effective at an earlier stage and at lower concentrations in the diet than SBBI. Thus, high levels of SBTI in the diet pro- duced Sth-instar larvae which were too small to pupate successfully. Moulting at a lower body weight than normal may be a mechanism to overcome the effects of an inadequate diet.
Increasing the levels of SBTI and SBBI in the diet increased mortality of larvae, but this effect only occurred after 14-15 days. High levels of inhibitor,
EFFECT OF INGESTED SOYBEAN PROTEASE INHIBITORS 663
especially SBTI, in the diet resulted in a continual reduction of larval growth and a disruption in normal development. These effects accumulated over 2 weeks until the larvae apparently became so stressed that they died. The presence of SBBI resulted in selective survival of the larger individuals. The possible consequences of this selection in generating resistant populations needs to be assessed. This effect was not found with SBTI.
Faecal output, as a measure of food intake, was measured up to day 10, at which point there was no effect by inhibitors on larval mortality. SBTI reduced total faecal output by more than 60%, even at low concentrations in the diet, suggesting it has good poten- tial for reducing damage to crop plants. However, analysis of the daily data showed that, when weight and developmental state of the larvae were considered, there were no significant differences in food consumpion. Thus, the reduction in faecal output with SBTI was not due to a strong phago-inhibitory effect by SBTI, but was mainly due to the effects of SBTI on larval growth and development. SBBI only had a slight effect on faecal output (a measure of food intake), even at high concen- trations, and thus would have little direct effect on larval feeding on plant tissues containing this protein.
The relative effectiveness of SBTI and SBBI in vitro and in viuo indicates that these inhibitors were interfering with normal proteolysis, and effectively starving the larvae of protein or, more likely, essential amino-acids. Broadway and Duffey (1986a) found that supplementing inhibitor diets with methionine removed the adverse effects that SBTI and potato proteinase inhibitor II had on the growth of larvae of Heliothis zea and Spodoptera exigua, and Karowe and Martin (1989) suggested that growth of Spodoptera eridania larvae was limited by low levels of essential amino-acids in their diet. However, since SBBI never completely inhibited proteolysis, in vitro, even at high concentrations, the larvae may have been able to obtain enough amino-acids from limited proteolysis to lessen the effects of this inhibitor on growth and development. Klocke and Chan (1982) showed that condensed tannin from cotton, while par- tially inhibiting protease activity of larval H. zea, had no effect on nutrient asssimilation.
Feeding trials with pure enzyme inhibitors can provide valuable information about the role played by the affected enzymes in the insect’s digestive system. The relative effectiveness of SBTI and SBBI in inhibiting the BApNAase protease and in affecting larval growth and development suggests that this enzyme is very important for general protein digestion. Similarly, Hines et al. (1990) showed that the cysteine protease in guts of Acanthoscelides obtectus was important for normal larval growth by feeding the larvae diets containing the specific cysteine protease inhibitor, E-64.
In contrast to the present study, Broadway and Duffey (1986a) found that protease inhibitors in the diet of H. zea and S. exigua stimulated levels of tryptic activity in the guts of larvae of both species [where trypsin-like enyme(s) are the major proteases]. They suggested that
pernicious hyperproduction of trypsin contributed to the adverse effects of the inhibitors. Larocque and Houseman (1990) also found that tryptic activity was increased when soybean and other protease inhibitors were added to the diet of 0. nubilalis. Results from the present study do not rule out an increased production of enzyme protein in response to inhibitor in the diet, since the increased potential tryptic activity could be masked by excess inhibitor.
A secretagogue mechanism for controlling tryptic activity in lepidopteran larvae was proposed by Broadway and Duffey (1986b), who found that levels of protease activity in S. exigua and H. zea larvae changed with protein content rather than meal size. Results from the present study also support this mechanism, since levels of protease activity increased with protein levels in the gut, both in the presence and absence of SBTI. If a secretagogue mechanism exists, protease production should be dependent on protein content, even in the presence of SBTI. Then, since the inhibitor was a constant proportion of the dietary protein content, one would predict a constant proportion of enzyme inhibited, leading to regression lines of different slopes converging to the same point on the y-axis of Fig. 6, as observed.
In this study, the relative effects of soybean protease inhibitors on H. armigera larvae, in feeding trials, agreed with their relative effectiveness as inhibitors of gut proteases, estimated by in vitro enzyme inhibition assays. This shows that the in vitro assays are a useful means of screening for inhibitors with potential for increasing crop resistance. Both the in vitro and in uivo results indicate that SBTI is a good candidate for genetic introduction, as a protective mechanism, into plants susceptible to attack by H. armigera larvae.
-7 7.5 1 . 4 . 4, .
7 6.0 - . . .c
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
Protein (mg.gut”)
FIGURE 6. The relationship between protein content and protease activity in the guts of 3rd~instar larvae fed diets with (0 ---0) and without (+---¤) SBTI at 0.234mM. Ordinate: protease activity, measured by hydrolysis of BApNA, at pH 9.8, 30°C and expressed as
AGD,,,, x 10’ per min per larval gut. Abscissa: protein content measured by absorbance at 260 and 280nm. and expressed as mg total protein per larval gut. Curve fit data: for larvae fed diet without SBTI; y = 20.05x + 0.44, r = 0.66. For larvae fed diet with SBTI;
y = 5.56.x + 0.05, r = 0.58.

664 K. A. JOHNSTON er al.
REFERENCES
Ahmad Z., Saleemuddin M. and Siddi M. (1980) Purification and characterisation of three alkaline proteases from the gut of the larva of army worm, Spodoptera litura. Insect Biochem. 10, 667-673.
Broadway R. M. (1989) Characterization and ecological implications of midgut proteolytic activity in larval Pieris rapae and Trichoplusia
ni. J. them. Ecol. 15, 2101-2113. Broadway R. M. and Duffey (1986a) Plant proteinase inhibitors:
mechanisms of action and effect on the growth and physiology of larval Heliothis zea and Spodoptera exigua. J. Insect Physiol. 32,
827834. Broadway R. M. and Duffey S. S. (1986b) The effect of dietary protein
on the growth and digestive physiology of larval Heliothis zea and Spodoptera exigua. J. Insect Physiol. 32, 673-680.
Dawson M. C., Elliott D. C., Elliott W. H. and Jones K. M. (1986) Dam for Biochemical Research. Clarendon Press, Oxford.
Fitt G. P. (1989) The ecology of Heliothis species in relation to agroecosystems. A. Rev. Ent. 34, 17-52.
Hamad M. B. B. and Attias J. (1987) Isolation and partial character- isation of two alkaline proteases of the greater wax moth Gaheria
melloneIla L. Insect Biochem. 17, 6533658. Hilder V. A., Gatehouse A. M. R., Sheerman S. E., Barker, R. F. and
Boulter D. (1987) A novel mechanism of insect resistance engineered into tobacco. Nature 334, 160-163.
Hines M. E., Nielsen S. S., Shade R. E. and Pomeroy M. N. (1990) The effect of two proteinase inhibitors, E-64, and the Bowman-Birk inhibitor, on the developmental time and mortality of Acanthoscelides obtectus. Entomologia exp. appl. 57, 201-207.
Houseman J. G., Downe A. E. R. and Philogene B. J. R. (1989) Partial characterization of proteinase activity in the larval midgut of the European corn borer, Ostrinia nubilalis Hubner (Lepidoptera: Pyralidae). Can. J. 2001. 67, 864-868.
Johnson R., Narvaez J., An G. and Ryan C. A. (1989) Expression of proteinase inhibitors I and II in transgenic tobacco plants: effects on
natural defense against Manduca sexta larvae. Proc. natn. Acad. Sci.,
U.S.A. 86, 9871-9875. Johnston K. A., Lee M. J., Gatehouse J. A. and Anstee J. H. (1991)
The partial purification and characterisation of serine protease activity in midgut of larval Helicoverpa armigera. Insect Biochem. 21, 389-397.
Karowe D. N. and Martin M. M. (1989) The effects of quantity and quality of diet nitrogen on the growth, efficiency of food utilization nitrogen budget, and metabolic rate of fifth-instar Spodoptera eridania larvae (Lepidoptera: Noctuidae). J. Insect Physiol. 35,
699-708. Klocke J. A. and Chan B. G. (1982) Effects of cotton condensed tannin
on feeding and digestion in the cotton pest, Heliothis zea. J. Insect
Physiol. 28, 911-915.
Larocque A. M. and Houseman J. G. (1990) Effect of ingested soybean, ovomucoid and corn protease inhibitors on digestive processes of the European corn borer, Ostrinia nubilalis (Lepi- doptera: Pyralidae). J. Insect Physiol. 36, 691697.
Ryan C. A. (1990) Protease inhibitors in plants: genes for improving defenses against insects and pathogens. A. Rev. Phytopath. 28,
425-449. Sasaki T. and Suzuki Y. (1982) Alkaline proteases in digestive juice of
the silkworm, Bombyx mori. Biochim. biophys. Acta 703, l-10. Shukle R. H. and Murdock L. L. (1983) Lipoxygenase, trypsin
inhibitor and lectin from soybeans: effects on larval growth of Manduca sexta (Lepidotera: Sphingidae). Environ. Ent. 12,
787-791.
Singh P. (1977) Arttficial Diets for Insects, Mites and Spiders. IF1 Plenum, New York.
Acknowledgements-This work was funded by the Agricultural and Food Research Council of Great Britain. We thank Mr P. Loftus who maintained the insect cultures.





























