Embed Size (px)
Citation preview

Investigative Ophthalmology & Visual Science, Vol. 31, No. 5, May 1990Copyright © Association for Research in Vision and Ophthalmology
Effects of Hematoporphyrin Derivative and Lighton Y79 Retinoblostomo Cells in Vitro
Yoshifaka Ohnishi, Mosakozu Murakami, and Hiroshi Wakeyama
Retinoblastoma Y79 cells exposed to a hematoporphyrin derivative and light were examined withregard to the production of intracellular lipid peroxide and morphologic changes, in the presence orabsence of oxygen. The intracellular lipid peroxide was related to the dose of hematoporphyrinderivative and the duration of photoradiation, under aerobic conditions. The formation of lipid perox-ide was not inhibited with superoxide dismutase and catalase, but it was inhibited with mannitol andl,4-diazabicyclo[2,2,2]octane, which were inhibitors of hydroxyl radicals and singlet oxygen, respec-tively. The mitochondria were apparently the target organelle in Y79 retinoblastoma cells. InvestOphthalmol Vis Sci 31:792-797,1990
Hematoporphyrin is a photodynamic substancewhich accumulates and is retained in tumor far morethan in normal cells.1 Photodynamic effects on reti-noblastoma cells have been investigated using hema-toporphyrin derivative (HpD) and light.23 Clinically,photodynamic therapy for malignant tumors was firstapplied by Dougherty et al in 1978.4
Since 1981, we have used photodynamic therapyfor 14 patients with retinoblastoma. A HpD andargon laser were used. This therapy has proven to beeffective for treating selected cases of retinoblas-toma.5 The mechanism of this therapy seems to relateto the phototoxicity of singlet oxygen produced byHpD exposed to light.6 As cellular sites of the actionof singlet oxygen and the relationship between singletoxygen and cell death are poorly understood,6"18 weinvestigated relationships between increases in intra-cellular lipid peroxide (LP) and cell death, underconditions of photoradiated HpD, with or withoutoxygen.
Materials and MethodsHpD was prepared following the procedure of Lip-
son et al.19 Retinoblastoma cell line Y79 cells werecultured in RPMI supplemented with 10% fetal calfserum. Y79 cells were incubated in culture mediumcontaining 10 ng HpD per 106 cells/ml for 4 hr, andthen were irradiated 5 cm distant from a 15-W fluo-
From the Department of Ophthalmology, Faculty of Medicine,Kyushu University, Fukuoka, Japan.
Supported by Fukuoka Anticancer Society grant 26-13.Submitted for publication: July 21, 1989; accepted September
11, 1989.Reprint requests: Dr. Yoshitaka Ohnishi, Department of Oph-
thalmology, Kyushu University 60, Faculty of Medicine, 3-1-1Maidashi, Fukuoka 812, Japan.
rescent bulb for 0-20 min in an incubator at 37°C.These cells were washed and homogenized ultrasoni-cally in 1.15% KC1, and the LP levels were measured(Fig. 1). Anaerobic conditions were obtained by re-placing the air in the experimental box with argon gasat 4°C. Cell viability was estimated by trypan bluestaining.
For the morphologic study, the Y79 cells werefixed, dehydrated, embedded, sectioned, and ob-served under a transmission electron microscope.
Results
First, the optimal HpD concentration and optimalincubation time for Y79 retinoblastoma cells takingup sufficient amounts of HpD were determined.Cell-associated HpD was measured fluorophotomet-rically, according to the method of Kessel andCheng.20 When the Y79 cells were incubated with0-200 n$ HpD per 106 cells/ml of culture medium for4 hr, the uptake of HpD increased with increase inthe HpD concentration in the culture medium. Theuptake was adequate at 10 ng HpD/106 cells (data notshown). The uptake then was measured for variousincubation times at 10 ng HpD/106 cells. Cell-asso-ciated HpD reached a maximum after the 4-hr incu-bation (data not shown). In the following experi-ments, the concentration was fixed at 10 ^g/106 cells,and the incubation time was 4 hr unless otherwiseindicated.
The formation of LP after light exposure was de-termined using various concentrations of HpD (Fig.2). After 10 min of photoradiation, the intracellularLP level was increased as the concentration of HpDin the medium was increased. However, no increaseof the LP level was observed in the absence of pho-
792
Downloaded from iovs.arvojournals.org on 03/06/2019
No. 5 EFFECTS OF HEMATOPORPHYRJN AND LIGHT ON Y79 CELLS / Ohnishi er ol 793
Cell homogenate in 1.15% KCI (100 mg protein/ml)
0.5ml
+3ml 0.5% Trichloroacetic acid
+ 1ml 0.6% Thiobarbituric acid
Heated in a 100 C water bath for 45min
cooled
+4 ml n-butanol
Shaken vigorously
Centrifuged at 2000g for 10 min
2.0
water phase butanol phase ( Malondialdehyde )
IAbsorbance at 535nm
Fig. 1. Method used to measure LP.
toradiation. In another control experiment, Y79 cellsincubated with 100 or 200 Mg HpD/106 cells werephotoradiated under anaerobic conditions. In thiscase, the formation of LP did not increase over thebaseline value.
The LP formation of cells incubated with 10 /ugHpD/106 cells was measured at various times of pho-toradiation (Fig. 3). The LP level increased with thetime of photoradiation. Photoradiation without HpDdid not yield any increase in LP formation. Underanaerobic conditions, LP formation by cells incu-bated with HpD did not increase for up to 20 min ofphotoradiation. These results show that Y79 cellstaking up HpD essentially need both light and molec-ular oxygen (O2) for LP formation, and suggest that
c'3
1ao>"3E
«a2 1.0a
°) aerobic condition
o anaerobic condition
HpD
-HpD
10Photoradiation time (minutes)
20
Fig. 3. Photoradiation time and LP production of cells incubatedwith 10 jig HpD per 106 cells/ml culture medium.
the active oxygens mediate the formation of LP. Wethen performed experiments using radical scavengers.
Y79 cells taking up HpD were exposed to light inthe existence of various radical scavengers (Fig. 4).Superoxide dismutase (SOD), the scavenger for su-
under aerobic condition + photoradiation
• under aerobic condition + no photoradiation
D under anaerobic condition + photoradiation
100 200
Dose of " cells)
Fig. 2. HpD uptake and LP production after 10 min of photora-diation.
»-• HpD + Lightfy o-a HpD+ SOD
/ ' / a-a HpD + Light +Catalase
* - * HpD • Light + Mannitol fOH)
O-o HpD + Light +(1,4-Diaiabicyclol2,2,2loctane)
10 20Photoradiation time (minutes)
Fig. 4. Influences of various radical scavengers on LP produc-tion.
Downloaded from iovs.arvojournals.org on 03/06/2019

794 INVESTIGATIVE OPHTHALMOLOGY b VISUAL SCIENCE / May 1990 Vol. 31
peroxide (O2 ), and catalase, the scavenger for hy-drogen peroxide (H2O2), did not inhibit the forma-tion of LP. Mannitol, the scavenger for the hydroxylradical (-OH), and l,4-diazabicyclo[2,2,2]octane,the scavenger for the singlet oxygen, partially inhib-ited the production of LP by the cells. Thus, eithersinglet oxygen or • OH is the mediator of LP forma-tion.
The relationship between LP formation and celldeath also was investigated (Fig. 5). At 5 min of pho-toradiation, LP formation occurred but cell deathratio was identical to the ratio in control experiment.When the cells were photoradiated for over 5 min, thecell death ratio increased abruptly and reached 83%at 20 min of photoradiation. The cell death ratio inthe absence of oxygen, light, or HpD was significantlylower than that in the presence of all three elements(Table 1). These results suggest that the oxygen-radi-cal-mediated reaction might lead to tumor cell deathin cases of photodynamic therapy with HpD.
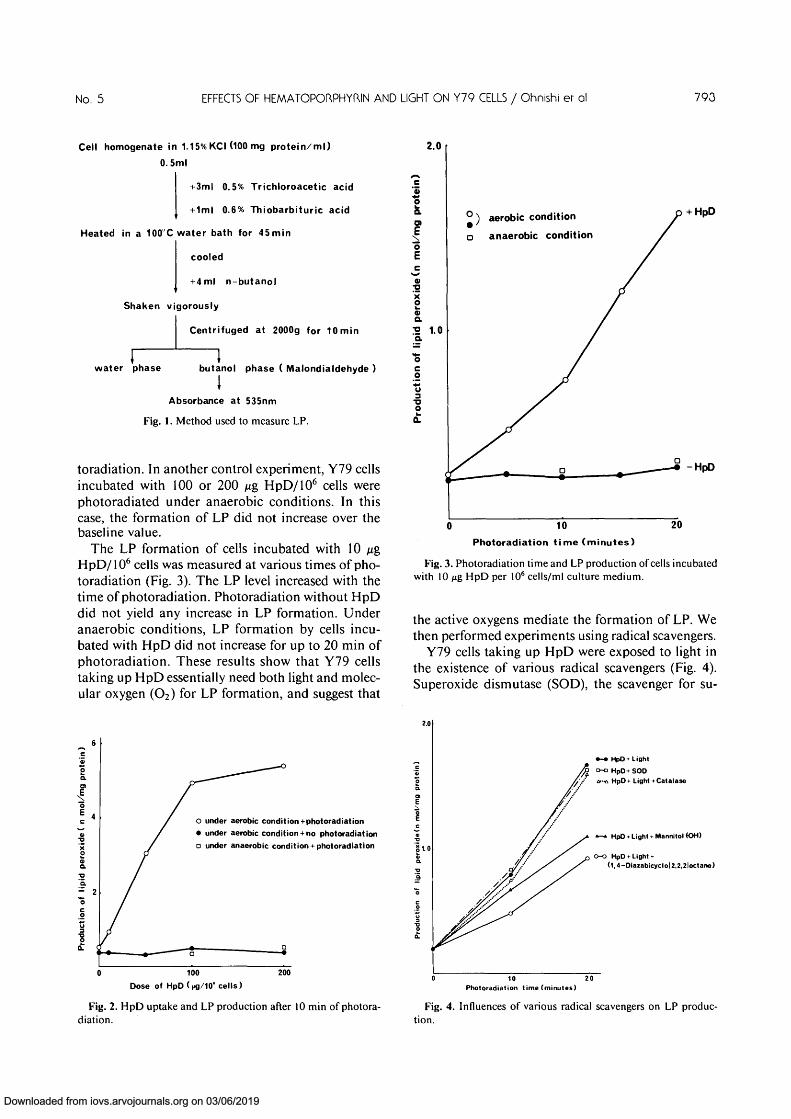
Transmission electron micrographs of Y79 cellsexposed to light for 20 min without HpD infiltrationrevealed a normal appearance. The Y79 cells exposedto HpD for 4 hr but not to light showed a normalappearance (Fig. 6). There were numerous mitoticfigures. The mitochondria in Y79 cells exposed tolight for 5 min in the case of infiltration of 10 ^gHpD/106 cells for 4 hr appeared contracted, and there
Table 1. Cell death ratio (%) in the absence orexistence of oxygen, light, and hematoporphyrinderivative (HpD)
1.0 =
Photoradiationtime (min)
1520
HpD +
95% air+ 5% Co2
81 ±892 ±7
light
Argon
26 ± 527 ± 7
Light H
95% air +
24 ±623 ±9
HpDH
5% Co2
25 ± 729 ± 10
10 20
Photoradiation time (minutes)
Fig. 5. Relationship between LP production and cell death.
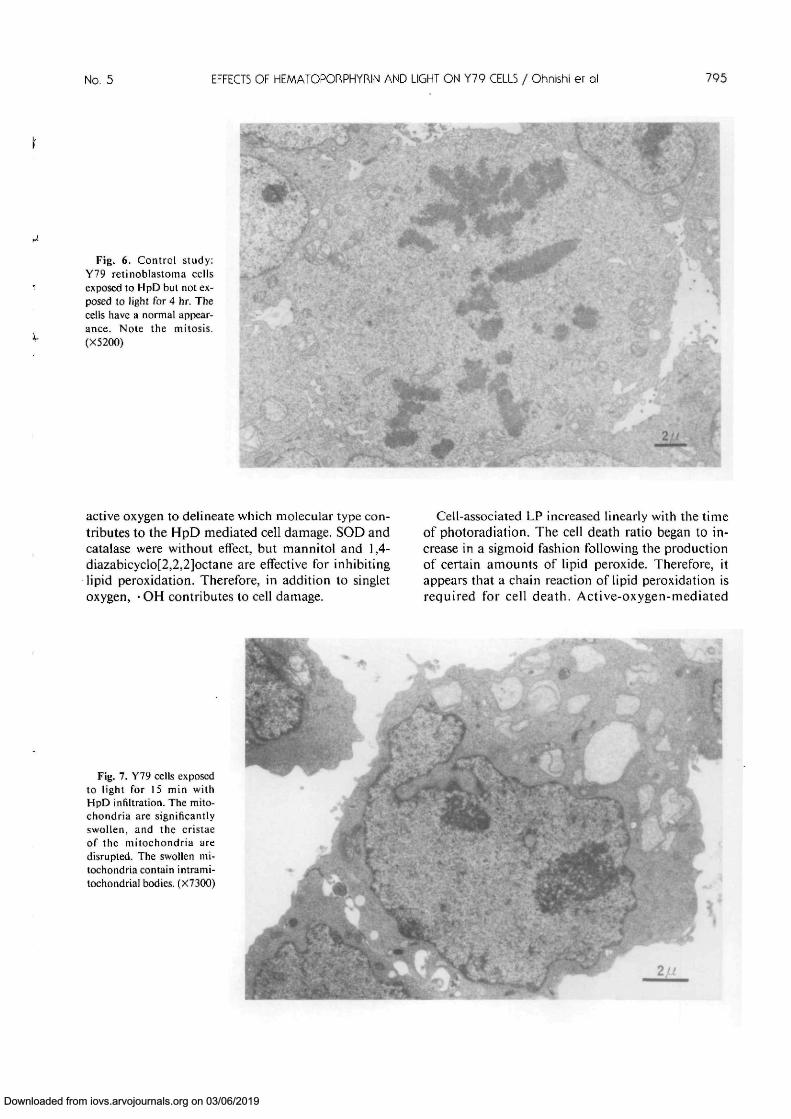
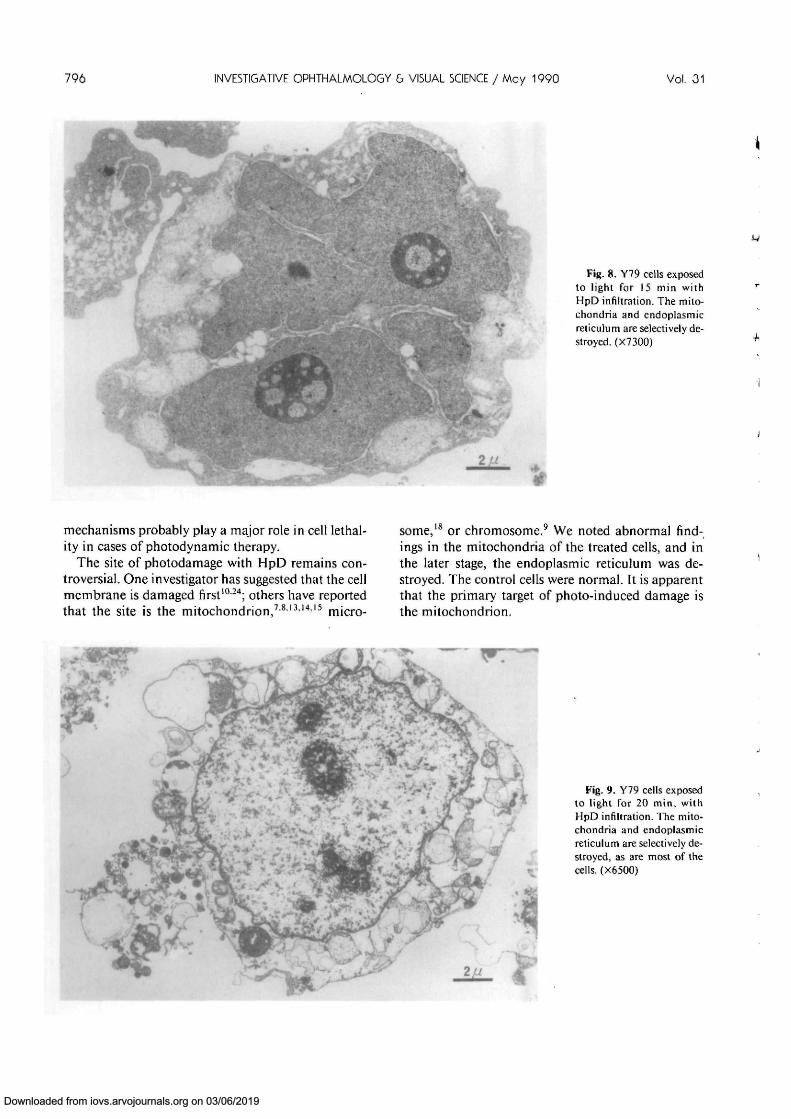
was an increase in density of the matrix. Mitochon-dria in the Y79 cells exposed to light for 10 min withHpD infiltration were slightly swollen. The mito-chondria in the cells exposed to light for 15 min withHpD infiltration were significantly swollen, and thecristae disrupted. These swollen mitochondria con-tained intramitochondrial bodies (Fig. 7). The mito-chondria and endoplasmic reticulum in the cells ex-posed to light for 15 min with HpD infiltration wereselectively destroyed, as were most of the cells (Fig.8). The plasma membranes in the cells exposed tolight for 20 min with HpD infiltration were disrupted,and the nuclei were pyknotic (Fig. 9). These morpho-logic studies revealed that the first changes occur inthe mitochondria.
Discussion
Porphyrin and hematoporphyrin are well-studiedphotosensitizers.21 Two types of mechanisms havebeen proposed for the photoinactivation of biomole-cules. In the so-called type II reaction, singlet oxygenmay mediate the photoinactivation. It remains to bedetermined whether singlet oxygen contributes totumor cell death in cases of photodynamic therapywith HpD, and the requirement for oxygen is not wellrepresented. We performed experiments using strictanaerobic conditions and obtained definite evidencefor the requirement of active oxygen to kill retino-blastoma cells. In ophthalmic research, HpD reten-tion in heterotransplanted tumor tissue has beennoted,122 but we found no report on the mechanismof cell death in photodynamic therapy. We measuredLP production to search for the effect of HpD oncells, because the method is simple and appropriatefor elucidating the contribution of active oxygen. Weused argon gas replacement for achieving the anaero-bic condition. This condition is adequate to block theNADPH oxidase of neutrophils, to produce the su-peroxide anion.23 NADPH oxidase has an unusuallyhigh affinity for molecular oxygen. These experi-ments clearly showed that HpD, light, and oxygen areessential factors for cell damage to be induced byphotodynamic therapy. We also used scavengers of
Downloaded from iovs.arvojournals.org on 03/06/2019
No. 5 EFFECTS OF HEMATOPORPHYRIN AND LIGHT ON Y79 CELLS / Ohnishi er QI 795
m-Fig. 6. Control study:
Y79 retinoblastoma cellsexposed to HpD but not ex-posed to light for 4 hr. Thecells have a normal appear-ance. Note the mitosis.(X5200)
A *
active oxygen to delineate which molecular type con-tributes to the HpD mediated cell damage. SOD andcatalase were without effect, but mannitol and 1,4-diazabicyclo[2,2,2]octane are effective for inhibitinglipid peroxidation. Therefore, in addition to singletoxygen, • OH contributes to cell damage.
Cell-associated LP increased linearly with the timeof photoradiation. The cell death ratio began to in-crease in a sigmoid fashion following the productionof certain amounts of lipid peroxide. Therefore, itappears that a chain reaction of lipid peroxidation isrequired for cell death. Active-oxygen-mediated
Fig. 7. Y79 cells exposedto light for 15 min withHpD infiltration. The mito-chondria are significantlyswollen, and the cristaeof the mitochondria aredisrupted. The swollen mi-tochondria contain intrami-tochondrial bodies. (X73OO) I
Downloaded from iovs.arvojournals.org on 03/06/2019
796 INVESTIGATIVE OPHTHALMOLOGY b VISUAL SCIENCE / May 1990 Vol. 31
Fig. 8. Y79 cells exposedto light for 15 min withHpD infiltration. The mito-chondria and endoplasmicreticulum are selectively de-stroyed. (X7300)
• _ • *
f
mechanisms probably play a major role in cell lethal-ity in cases of photodynamic therapy.
The site of photodamage with HpD remains con-troversial. One investigator has suggested that the cellmembrane is damaged first1024; others have reportedthat the site is the mitochondrion,78131415 micro-
some,18 or chromosome.9 We noted abnormal find-ings in the mitochondria of the treated cells, and inthe later stage, the endoplasmic reticulum was de-stroyed. The control cells were normal. It is apparentthat the primary target of photo-induced damage isthe mitochondrion.
Fig. 9. Y79 cells exposedto light for 20 min, withHpD infiltration. The mito-chondria and endoplasmicreticulum are selectively de-stroyed, as are most of thecells. (X6500)
Downloaded from iovs.arvojournals.org on 03/06/2019

No. 5 EFFECTS OF HEMATOPORPHYRIN AND LIGHT ON Y79 CELLS / Ohnishi er ol 797
Key words: hematoporphyrin derivative, lipid peroxide,Y79 retinoblastoma cell, mitochondria, photodynamictherapy
AcknowledgmentThe authors thank M. Ohara for comments on the man-
uscript.
References1. Gomer CJ, Rucker N, Mark C, Benedict WF, and Murphree
AL: Tissue distribution of 3H-hematoporphyrin derivative inathymic "nude" mice heterotransplanted with human retino-blastoma. Invest Ophthalmol Vis Sci 22:118, 1982.
2. Sery TW: Photodynamic killing of retinoblastoma cells withhematoporphyrin and light. Cancer Res 39:96, 1979.
3. Benedict WF, Lingua RW, Doiron DR, Dawson JA, andMurphree AL: Tumor regression of human retinoblastoma inthe nude mouse following photoradiation therapy: A prelimi-nary report. Med Pediatr Oncol 8:397, 1980.
4. Dougherty TJ, Kaufman JE, Goldfarb A, Weishaupt KR,Boyle D, and Mittleman A: Photoradiation therapy for thetreatment of malignant tumors. Cancer Res 38:2628, 1978.
5. Ohnishi Y, Yamana Y, and Minei M: Photoradiation therapyusing argon laser and a hematoporphyrin derivative for retino-blastoma: A preliminary report. Jpn J Ophthalmol 30:409,1986.
6. Weishaupt KR, Gomer CJ, and Dougherty TJ: Identificationof singlet oxygen as the cytotoxic agent in photo-inactivationof a murine tumor. Cancer Res 36:2326, 1976.
7. Coppola A, Viggiani E, Salzarulo L, and Rasile G: Ultrastruc-tural changes in lymphoma cells treated with hematopor-phyrin and light. Am J Pathol 99:175, 1980.
8. Volden G, Christensen T, and Moan J: Photodynamic mem-brane damage of hematoporphyrin derivative-treated NHIK3025 cells in vitro. Photobiochem Photobiophys 3:105, 1981.
9. Moan J, Johannessen JV, Christensen T, Espevik T, andMcGhie JB: Porphyrin-sensitized photoinactivation of humancells in vitro. Am J Pathol 109:184, 1982.
10. Berns MW, Dahlman A, Johnson FM, Burns R, Sperling D,Guiltinan M, Siemens A, Walter R, Wright W, Hammer-Wil-son M, and Wile A: In vitro effects of hematoporphyrin deriva-tive. Cancer Res 42:2325, 1982.
11. Gibson SL and Hilf R: Photosensitization of mitochondrialcytochrome C oxidase by hematoporphyrin derivative and re-
lated porphyrins in vitro and in vivo. Cancer Res 43:4191,1983.
12. Moan J: The photochemical yield of singlet oxygen from por-phyrins in different states of aggregation. Photochem Photo-biol 39:445, 1984.
13. Lee See K, Forbes IJ, and Betis WH: Oxygen dependency ofphotocytotoxicity with hematoporphyrin derivative. Photo-chem Photobiol 39:631, 1984.
14. Miyoshi N, Hisazumi H, Ueki O, and Nakajima K: Cellularbinding of hematoporphyrin derivative in human bladdercancer cell line KK-47. Photochem Photobiol 39:359, 1984.
15. Das M, Dixit R, Mukhtar H, and Bickers DR: Role of activeoxygen species in the photodestruction of microsomal cy-tochrome P-450 and associated monooxygenases by hemato-porphyrin derivative in rats. Cancer Res 45:608, 1985.
16. Van Steveninck J, Tijssen K, Boegheim JPJ, Van der Zeer J,and Dubbelman TMAR: Photodynamic generation of hy-droxyl radicals by hematoporphyrin derivative and light. Pho-tochem Photobiol 44:711, 1986.
17. Hilf R, Murant RS, Narayanan U, and Gibson SL: Relation-ship of mitochondria and cellular adenosine triphosphatelevels to hematoporphyrin derivative-induced photosensitiza-tion in R3230AC mammary tumors. Cancer Res 46:211, 1986.
18. Kessel D: Site of photosensitization by derivatives of hemato-porphyrin. Photochem Photobiol 44:489, 1986.
19. Lipson R, Baldes E, and Olsen AM: The use of derivative ofhematoporphyrin in tumor detection. J Natl Cancer Inst 26:1,1961.
20. Kessel D and Cheng M-L: Biological and biophysical proper-ties of tumor-localizing component of hematoporphyrin deriv-ative. Cancer Res 45:3053, 1985.
21. Pottier R and Truscott TG: The photochemistry of hemato-porphyrin and related systems. Int J Radiat Biol 50:421, 1986.
22. Gomer CJ, Jester JV, Razum NJ, Szirth BC, and MurphreeAL: Photodynamic therapy of intraocular tumors examinationof hematoporphyrin derivative distribution and long-termdamage in rabbit ocular tissue. Cancer Res 45:3718, 1985.
23. Murakami M, Nakamura M, and Minakami S: NADPH oxi-dase of guinea-pig macrophages catalyses the reduction of ubi-quinone-1 under anaerobic conditions. Biochem J 237:541,1986.
24. Gibson SL, Murant RS, and Hilf R: Photosensitizing effects ofhematoporphyrin derivative and Photofrin II on the plasmamembrane enzymes 5'-nucleotidase, Na+K+-ATPase, andMg2+-ATPase in R3230AC mammary adenocarcinomas.Cancer Res 48:3360, 1988.
Downloaded from iovs.arvojournals.org on 03/06/2019





















![HPD 2 cifrado[1]](https://img.dokumen.tips/doc/110x75/5571fe4149795991699afd46/hpd-2-cifrado1.jpg)







