Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Dried peel fraction of Citrus sinensis partially reversespathological changes in rat model of liver cirrhosis
Shakir Ali • Ram Prasad • Mohammed Naime • Hina Zafar • Amena Mahmood •
Indusmita Routray • Mehmet Yalniz • Ibrahim H. Bahcecioglu • Kazim Sahin
Received: 16 March 2010 / Accepted: 13 October 2010 / Published online: 3 December 2010
� Springer-Verlag 2010
Abstract Citrus sinensis is a seasonal fruit. Its zester is
rich in bioactive phytochemicals, such as limonene,
b-sitosterol, and ascorbic acid, which possess pharmaco-
logical action. In this study, we report the effect of fraction
prepared from dried peel of C. sinensis on biochemical and
histopathological changes in rat model of liver cirrhosis.
Liver cirrhosis was induced in rats by administering thi-
oacetamide at a concentration of 0.03% in drinking water
for 16 weeks. Thioacetamide was discontinued after
16 weeks and from the 18th week rats were given the
extract orally for 9 weeks. Following the completion of the
treatment, animals were killed and biochemical and histo-
pathological changes associated with liver cirrhosis were
evaluated. The treatment was found to reverse the elevated
levels of alkaline phosphatase, c-glutamyl transferase, and
other biochemical markers related to oxidative stress and
selected drug metabolizing enzymes. Histopathology of the
hepatic tissue confirmed the curative effect of the extract,
and corroborated with the biochemical findings. HPTLC
fingerprinting of the test fraction confirmed the presence of
limonene, b-sitosterol, and ascorbic acid, which may par-
tially explain the effect. The extract was also found to
possess the anti-proliferative activity, determined by mea-
suring the incorporation of radioactive thymidine by the
hepatic DNA. The study indicates the inhibitory action of
the test preparation on collagen accumulation in the
extracellular matrix, and hence suggests its use as a
potential therapeutic agent in liver fibrosis and cirrhosis.
Keywords Citrus sinensis � Liver cirrhosis �Animal model
Introduction
Citrus sinensis (sweet orange), family Rutaceae, is a sea-
sonal fruit widely grown in warm climates worldwide. Its
zester, the outer-most layer of the rind, contains the oil
glands and has a strong flavor similar to the fleshy inner part
of the fruit. Citrus species have been studied for medicinal
properties [1–3] and reported to contain numerous bioactive
compounds, such as flavanone glycosides (narirutin,
naringin, hesperidin, and neohesperidin) and natural anti-
oxidants [4–6]. Particularly, the phenolic acids have
attracted more attention for their antioxidant behavior and
beneficial health-promoting effects in chronic and degen-
erative diseases [7]. The fruit has been found to prevent the
oxidative stress by enhancing the total antioxidant capacity
and elevating liver antioxidant enzymes while modulating
cardiovascular risk factors [8]. Epidemiological studies
have suggested the protective role of the fruit in a variety of
human cancers [9], which might be attributed to limonoids
[10, 11]. D-Limonene, a key ingredient of orange peel
inhibits N-nitrosomorpholine-induced hepatocarcinogenesis
This work belongs to the PhD thesis of R. Prasad carried under the
supervision of S. Ali at Jamia Hamdard University.
S. Ali (&) � R. Prasad � M. Naime � H. Zafar � A. Mahmood �I. Routray
Department of Biochemistry, Faculty of Science,
Jamia Hamdard University, Hamdard Nagar,
New Delhi 110062, India
e-mail: [email protected]
M. Yalniz � I. H. Bahcecioglu
Department of Gastroenterology, Faculty of Medicine,
Firat University, 23119 Elazig, Turkey
K. Sahin
Department of Animal Nutrition, Faculty of Veterinary
Medicine, Firat University, 23119 Elazig, Turkey
123
Mediterr J Nutr Metab (2011) 4:57–67
DOI 10.1007/s12349-010-0033-8

[12]. Other bioactive compounds in the fruit, for example,
hesperidin and naringin, are powerful plasma lipid lower-
ing agents, increase the plasma antioxidant activity [13],
and have been reported to protect the liver against
CCL4-induced oxidative stress in rats [14]. Hesperidin also
attenuates lipopolysaccharide-induced hepatotoxicity, pos-
sibly by preventing the cytotoxic effects of NO and
oxygen-free radicals [15]. Considering the multiple phar-
macological role of bioactive compounds in C. sinensis, we
proposed to study its effect on liver cirrhosis in animal
model.
Materials and methods
Preparation of test fraction from dried orange peel
Citrus sinensis was purchased from commercial fruit
market in Delhi. The fruit was peeled and the peel was air
dried. A small amount of the peel was preserved under the
specimen voucher number BRL/A-3/03. Rest of the peel
was crushed in iron mortar, and subjected to extraction in
Soxhlet apparatus at 60–70 �C for 6 h continuously in 50%
ethanol. The extracted material was evaporated to dryness
under reduced pressure (40–50 �C), and suspended in dis-
tilled water in 1% gum acacia as suspending agent. The
suspended extract was stored in sterilized airtight vials in
refrigerator until further use.
Chemicals and other consumables for this study were
purchased from the standard commercial sources in India.
Animals and experimental design
Female rats (Wistar strain, inbred), weighing 160–180 g,
were used throughout the study. Animals were acclima-
tized for 1 week in polypropylene cages, and kept in an
environmentally controlled room (temperature: 21 ± 2 �C,
humidity: [40%) with a 12-h light–dark cycle. They were
provided pellet diet and water ad libitum. The study was
approved by the Animal Ethics Committee.
Animals were fasted overnight before the experiment and
divided into the following four groups: Group I (NC) con-
sisted of the control rats treated with normal saline for the
entire duration of the experiment; Group II (CS) consisted
of rats which were administered the orange peel extract
alone for 9 weeks to see the effect of the peel extract on rats;
Group III (LC) received thioacetamide for 16 weeks to
induce liver cirrhosis, this group was left untreated for next
10 weeks and served as positive control; Group IV (T)
consisted of the cirrhotic rats that received the test extract
for 9 weeks. On completion of the treatment, animals were
killed, and the tissue was subjected to histopathological and
biochemical examination. The experimental protocol is
shown in Fig. 1. The dose of the test extract was 800 mg/kg
body weight per day; equivalent dose in human weighing
60 kg, as calculated using the standard conversion formula
[16], comes out to be 2.3 g/day, t.d.s.
Gross morphology, liver weight/body weight ratio,
and tissue histopathology
Gross morphological examination of the liver of each rat
was done and documented. The liver weight and body
weight of each animal was taken to calculate the liver
weight and body weight ratio and a small piece of the
hepatic tissue was cut and fixed in 10% buffered formalin.
Histopathological sections were stained using hematoxy-
lin–eosin (HE), and examined by a pathologist who was not
aware of the treatment groups.
Biochemical analysis
After completion of the treatment, blood was collected
from the abdominal aorta and the serum was prepared for
biochemical markers of liver cirrhosis. Hepatic tissue was
subjected to further biochemical analysis. The procedure
for the preparation of sample and subcellular fractionation
was similar to the one described in one of our earlier
publications [17]. Serum aminotransferases [18], alkaline
phosphatase [19], and hepatic c-glutamyl transferase [20],
Fig. 1 Schematic illustration of the treatment protocol design to
induce liver cirrhosis and study the effect of test fraction in rat.
Following the completion of 16 weeks treatment with thioacetamide,
rats were left untreated for 1 week to ensure that the thioacetamide or
its metabolites are eliminated and the constituents in the extract do not
interfere with the metabolites or the drug metabolizing system [17]
58 Mediterr J Nutr Metab (2011) 4:57–67
123

lipid peroxidation [21], reduced glutathione [22], glutathi-
one reductase [23], glutathione peroxidase [24], and cata-
lase [25] were measured in all groups. Selected Phase I and
Phase II drug metabolizing enzymes, xanthine oxidase [26]
and glutathione s-transferase [27] were also measured in
the post-mitochondrial supernatant.
HPTLC fingerprinting and TLC autography of the test
fraction
The chemical fingerprinting of the extract was performed
on HPTLC. Briefly, 2.0 mg of the dried peel extract was
suspended in 1 mL methanol and filtered through mem-
brane filter (pore size: 0.45 l). CAMAG HPTLC system
equipped with an automatic TLC sampler (Linomat 5), TLC
scanner 3, and integrated software winCATS version 3 were
used. Chromatography was performed on a pre-coated
silica gel HPTLC plate of 0.20-mm thickness. Sample was
applied onto the plate with an automatic TLC sampler
(Linomat 5) under the flow of N2 gas. Linear ascending
development was carried out in the CAMAG twin trough
chamber (10 cm 9 10 cm), which was pre-saturated with
the mobile phase for 30 min at room temperature. For
limonene, hexane:diethyl ether:glacial acetic acid (80:20:1)
was used as the mobile phase. For b-sitosterol and ascorbic
acid, respectively, the mobile phases were as follows,
hexane:ethyl acetate:acetic acid (75:25:1), and acetoni-
trile:acetone:water:acetic acid (80:5:15:2). Following the
completion of the run, the plate was dried in oven at 60 �C
for 5 min, and the post-chromatographic derivatization of
limonene and b-sitosterol was carried out in 5% anisalde-
hyde followed by heating at 110 �C for 2 min. For finger-
printing, the plate was scanned in absorption mode at
250–500 nm under computerized CAMAG TLC scanner 3.
Standards of limonene, b-sitosterol, and ascorbic acid were
analyzed along with the extract. Standards were prepared in
methanol.
TLC autographic assay was performed to demonstrate
the antioxidant activity of the extract according to the
method described by Galvez et al. [28]. Briefly, 5 lL
extract (in methanol) was spotted on silica gel plate
(10 9 10 cm) and run on a TLC plate using a solvent
system consisting of hexane, ethyl acetate, and glacial
acetic acid (75:25:05) for 20 min. TLC plate was dried and
sprayed with 0.2% 2,2-diphenyl-2-picrylhydrazyl (DPPH)
prepared in methanol, and examined. Yellow or light spots
against a purple background indicated the presence of
antioxidants in the extract. Radical scavenging activity of
the extract against stable DPPH was also determined
spectrophotometrically. In presence of the antioxidants,
the DPPH is reduced and change color from deep violet to
light or yellow, which can be measured at 517 nm in
spectrophotometer.
Free radical scavenging activity of the herbal extract
was determined as described by Yamaguchi et al. [29].
Briefly, 5 mg of dried 50% ethanolic extract of C. sinensis
was dissolved in 1 mL methanol. Final reaction mixture
consisted of 0.1 mL sample dissolved in methanol, 0.3 mL
methanol, and 0.4 mL of 0.3 mM DPPH (in methanol).
The mixture was vortexed for 15 s and left to stand at room
temperature for 10 min. The absorbance was read at
517 nm using UV spectrophotometer. Pyrogallol was used
as control. Scavenging effect of DPPH was calculated as
follows: Scavenging activity (%) = [(OD of blank - OD
of sample)/OD of blank] 9 100.
Treatment protocol for cell proliferation assay
The anti-proliferative activity of the test preparation was
studied on thioacetamide mediated increase in [3H]-thy-
midine incorporation into hepatic DNA synthesis. Briefly,
24 female rats were randomly divided into four groups,
each having six animals. Group I was administered nor-
mal saline. Group II and IV were orally administered the
test preparation (800 mg/kg body weight/day, orally) for
three consecutive days. On third day of experiment,
Groups III and IV were given a single intraperitoneal
injection of thioacetamide (400 mg/kg body weight)
freshly dissolved in 0.9% NaCl. Thioacetamide was
administered 45 min after the administration of the last
dose of the test preparation. After 16 h of thioacetamide
administration, all groups were injected [3H]-thymidine
(20 lCi/0.2 mL saline/100 g body wt, i.p.) and killed
exactly after 2 h by cervical dislocation. Liver was
excised quickly, washed in ice-cold saline (0.9% NaCl),
and processed for subcellular fractionation for the esti-
mation of hepatic DNA synthesis.
Estimation of hepatic DNA synthesis
The isolation of hepatic DNA and incorporation of [3H]-
thymidine in hepatic DNA was performed according to
the method described by Smart et al. [30]. The liver
homogenate (20%, w/v) was prepared in ice-cold distilled
water. After homogenization, equal volume of ice cold
TCA (10%) was added and centrifuged at 5,000 rpm for
10 min. The supernatant was discarded. The precipitate
was dissolved in 5 mL ice-cold TCA (5%), and centri-
fuged at 5,000 rpm for 10 min. After the centrifugation,
supernatant was discarded and the pellet was dissolved in
5 mL of ice-cold perchloric acid (PCA) (10%) and kept
for 18 h at 4 �C. After 18 h, all tubes were centrifuged at
5,000 rpm for 10 min and the pellet was obtained. The
pellet was mixed with 5 mL of ice-cold PCA (5%)
and centrifuged at 5,000 rpm for 10 min at 4 �C. The
precipitate obtained after the final centrifugation was
Mediterr J Nutr Metab (2011) 4:57–67 59
123

incubated with 5 mL of warm PCA (10%) in boiling
water bath for 30 min. The sample was again centrifuged
and the supernatant filtered through a Whatman-50 filter
paper to get clear solution, which was used for DNA
estimation and counting radioactivity. For counting
radioactivity in the sample, 200 lL of the solution was
added to the scintillation vial containing 5 mL of scin-
tillation fluid and counted in the scintillation counter
(LKB-Wallace1410). The amount of DNA in the filtrate
was estimated by the diphenylamine method of Giles and
Myers [31]. The amount of [3H] thymidine incorporated
was expressed as d.p.m. (disintegration per minute)/lg
DNA.
Statistical analysis
The data were analyzed using one-way ANOVA. Mean ±
S.E.M. (n = 6) was calculated in each group. The signifi-
cance of difference between the positive control and the
treated group was determined; p \ 0.05 was considered
statistically significant.
Results
Effect of treatment on organ indices and serum marker
of liver cirrhosis (Tables 1, 2)
Percent liver weight body weight ratio, which doubled in
the liver cirrhosis group (6.3%), was found to decrease
significantly in treated rats (4%) (Table 1). A significant
increase in ALP, and also GGT was found in liver cir-
rhosis. Treatment with the orange peel faction could bring
down the increased values close to normal (Table 2).
Gross morphology and histopathology of the liver
Deposition of collagen, which is secreted by the activated
hepatic stellate cells (HSC), is a characteristic feature of
liver cirrhosis that results in gross morphological changes
and changes in the hepatic tissue architecture. The whole
organ and the sections from the treated and the control
group (Fig. 2) show characteristic nodules on the liver in
the LC group (Fig. 2c). The texture of the liver in the
control and test fraction treated rats was almost similar in
comparison to the LC group. Analysis of the representative
photomicrographs of sections from various groups provides
an insight into the hepatic tissue architecture. Control rats
(NC and CS) had normal lobular architecture with central
veins and radiating hepatic cords (Fig. 2a, b). On the other
hand, the LC group had proliferating bile ducts in the septal
area and a large nodule of regenerating hepatocytes; vac-
uolization of hepatocytes can be seen in the upper left
corner of sections from the cirrhotic liver, where the septa
are comparatively thick (Fig. 2c). Sections from the treated
group (Fig. 2d) showed hepatic parenchyma and regener-
ating hepatocytes and a more natural architecture (Fig. 2d).
Effect on the oxidative stress biomarkers
Elevated lipid peroxidation (LPO) is an important factor
for the development of liver fibrosis. It is also an oxidative
Table 1 Effect of the test fraction on liver weight body weight ratio
Group Liver weight Body weight Liver wt/Body
wt ratio (%)
NC 08.24 ± 0.47 266.67 ± 15.08 3.08
CS 08.10 ± 0.25 250.50 ± 6.50 3.23
LC 10.92 ± 0.79* 173.33 ± 5.42* 6.30
T 09.75 ± 0.15 220.10 ± 8.90 4.42
Data represent Mean ± S.E.M. (n = 6)
NC normal control, CS rats receiving the test fraction alone, LC liver
cirrhosis, T post-treatment group (the group of rats receiving the test
fraction following the induction of liver cirrhosis)
* p \ 0.05, when compared to the control group of rats. No signifi-
cant difference was found in NC and CS groups. Administration of
the test extract caused a decrease in the liver weight body weight ratio
Table 2 Effect of the test fraction on serum markers of hepatic injury
Groups ALT AST ALP GGT
NC 72.28 ± 0.71 (100) 130.77 ± 2.78 (100) 14.68 ± 0.86 (100) 302.00 ± 12.51 (100)
CS 71.08 ± 1.82 (98.33) 130.82 ± 1.18 (100.03) 15.31 ± 1.26 (104.29) 295.91 ± 16.05 (97.98)
LC 75.01 ± 1.16 (103.77) 139.87 ± 2.30 (106.95) 21.10 ± 0.93* (143.73) 359.93 ± 18.56* (119.18)
T 70.85 ± 1.14 (98.02) 132.25 ± 2.29 (101.13) 15.67 ± 0.69 (106.74) 305.33 ± 14.90 (101.10)
Data represent Mean ± S.E.M. (n = 6)
NC normal control, CS rats receiving the test fraction alone, LC liver cirrhosis, T post-treatment group (the group of rats receiving the test
fraction following the induction of liver cirrhosis), ALT alanine aminotransferase (units/mL), AST aspartate aminotransferase (units/mL), ALPalkaline phosphatase (equivalent units/mL), and GGT c-glutamyl transpeptidase (nmol of p-nitroanilide/mg protein)
* p \ 0.05, when compared to the control group of rats. Values in bracket show percent change with respect to the normal control. Post-
treatment with the peel extract normalized the activity of ALP, GGT, and AST to almost normal level
60 Mediterr J Nutr Metab (2011) 4:57–67
123

stress biomarker that can be measured in the whole tissue
lysate as malondialdehyde and expressed as nmol/mg
protein. LPO increased to 127% in the LC group over the
control group (598.9 ± 4.2 vs. 472.3 ± 9, *p \ 0.05).
Following the extract treatment, the value decreased to
506.2 ± 11.5, which in percent-activity was in the range
close to the normal control (Table 3). Reduced glutathione
(GSH), which decreased significantly (*p \ 0.05) in the
Fig. 2 Gross morphology of
the liver and respective
histopathological sections of the
following groups: a normal
control, b the group receiving
the test fraction, c rats with liver
cirrhosis, and d the group post-
treated with the test fraction. PTportal triad, CV central vein,
BDP bile duct proliferation
Mediterr J Nutr Metab (2011) 4:57–67 61
123

LC group, was almost normal in the treated group
(Table 3). Glutathione reductase (GR), the enzyme
required for the recycling of GSH (from its oxidized form,
GSSG) also decreased in the LC group (400.02 ± 7.97),
and attained the value similar to the control in treated rats
(518.97 ± 11.26 vs. 509.10 ± 13.21) (Table 3). The per-
oxide metabolizing enzymes, catalase, and GPx (Table 4)
decreased significantly (*p \ 0.05) in the LC group and
tend to increase in the treated rats.
Effect on selected phase I and II drug metabolizing
enzymes
Non-microsomal Phase I drug metabolizing enzyme xan-
thine oxidase (XO) was particularly studied for its ability to
generate reactive oxygen species (ROS) while catalyzing
the reaction. XO was found to increase (p [ 0.05) when
compared to the mean control value (Table 5), and
decreased in the group receiving the extract post-treatment.
This effect can be correlated to the activity levels of
peroxide metabolizing enzymes (Table 4), oxidative stress
markers (Table 3), serum markers of liver cirrhosis
(Table 2), and other signs and symptoms of hepatic injury
(Table 1; Fig. 2). Further, GST, which is a phase II drug
metabolizing enzyme, increased in the LC group and
decreased significantly following the treatment with the
test preparation (Table 5).
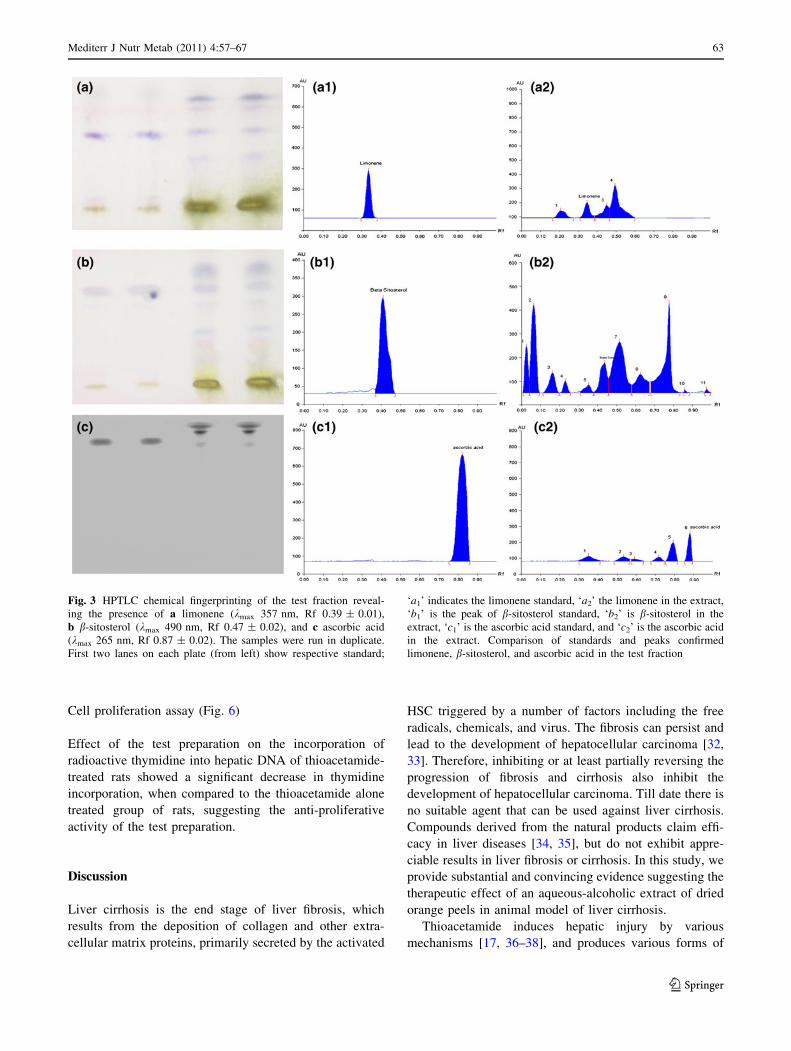
HPTLC finger printing and the TLC autography
of the test fraction
HPTLC chemical fingerprinting of the extract showed
several peaks at 250–500 nm. Photographic images of the
HPTLC plates in Fig. 3 demonstrate the presence of lim-
onene, b-sitosterol, and ascorbic acid in the extract; the
structures of limonene, b-sitosterol, and ascorbic acid are
shown in Fig. 4. TLC autographic assay was used to
demonstrate the presence of antioxidants in the extract. The
extract showed 94% free radical scavenging activity when
compared to pyrogallol (Fig. 5).
Table 3 Effect of the test fraction on oxidative stress markers
Groups LPO GSH GR
NC 472.34 ± 09.01 (100) 1,392.79 ± 22.37 (100) 518.97 ± 11.26 (100)
CS 470.75 ± 15.38 (99.66) 1,395.44 ± 11.49 (100.19) 522.06 ± 25.34 (100.59)
LC 598.86 ± 4.24* (126.78) 1,305.14 ± 23.69* (93.70) 400.02 ± 7.97* (77.07)
T 506.18 ± 11.53 (107.16) 1,379.62 ± 12.86 (99.05) 509.10 ± 13.21 (98.09)
Data represent Mean ± S.E.M. (n = 6)
NC normal control, CS rats receiving the test fraction alone, LC liver cirrhosis, T post-treatment group (the group of rats receiving the test
fraction following the induction of liver cirrhosis), LPO lipid peroxidation (nmol malondialdehyde/mg protein), GSH reduced glutathione (lmol/
g tissue), and GR glutathione reductase (nmol NADPH oxidized/min/mg protein)
* p \ 0.05, when compared to the control group of rats. Values in bracket show percent change with respect to the normal control. While the
level of GSH and GR reduced significantly (*p \ 0.05), LPO increased (* p \ 0.05). Post-treatment with the extract almost normalized the
values
Table 4 Effect of the test fraction on peroxide metabolizing enzymes
Groups GPx Catalase
NC 198.16 ± 10.01 (100) 2,289.57 ± 54.32 (100)
CS 174.55 ± 8.70 (88.08) 1,853.67 ± 69.85 (80.96)
LC 139.97 ± 3.53* (70.63) 1,451.27 ± 43.18* (63.38)
T 162.59 ± 6.35 (82.04) 1,680.59 ± 43.88 (73.40)
Data represent Mean ± S.E.M. (n = 6)
NC normal control, CS rats receiving the test fraction alone, LC liver
cirrhosis, T post-treatment group (the group of rats receiving the test
fraction following the induction of liver cirrhosis), GPx glutathione
peroxidase (nmol NADPH oxidized/min/mg protein)
*p \ 0.05, when compared to the control group of rats. Values in
bracket show percent change with respect to the normal control. Activity
of both GPx and catalase decreased in the LC group (*p \0.05), and
was found to increase in the post-treated rats; the activity of catalase was
expressed as the nmole H2O2 consumed/min/mg protein
Table 5 Effect of the test fraction on selected Phase I and Phase II
drug metabolizing enzymes
Groups XO GST
NC 174.25 ± 5.06 (100) 653.31 ± 22.49 (100)
CS 159.54 ± 6.91 (91.55) 639.12 ± 11.42 (97.82)
LC 193.34 ± 4.46* (110.95) 707.83 ± 8.65* (108.34)
T 163.16 ± 10.58 (93.63) 611.72 ± 22.74 (93.63)
Data represent Mean ± S.E.M. (n = 6). *p \ 0.05, when compared
to the control group of rats. Values in bracket show percent change
with respect to the normal control. Both XO and GST increased in the
LC group and decreased in the treated rats
NC normal control, CS rats receiving the test fraction alone, LC liver
cirrhosis, T post-treatment group (the group of rats receiving the test
fraction following the induction of liver cirrhosis), XO xanthine
oxidase (lmol of uric acid/mg protein), GST glutathione s-transferase
(nmol of CDNB conjugate/mg protein)
62 Mediterr J Nutr Metab (2011) 4:57–67
123
Cell proliferation assay (Fig. 6)
Effect of the test preparation on the incorporation of
radioactive thymidine into hepatic DNA of thioacetamide-
treated rats showed a significant decrease in thymidine
incorporation, when compared to the thioacetamide alone
treated group of rats, suggesting the anti-proliferative
activity of the test preparation.
Discussion
Liver cirrhosis is the end stage of liver fibrosis, which
results from the deposition of collagen and other extra-
cellular matrix proteins, primarily secreted by the activated
HSC triggered by a number of factors including the free
radicals, chemicals, and virus. The fibrosis can persist and
lead to the development of hepatocellular carcinoma [32,
33]. Therefore, inhibiting or at least partially reversing the
progression of fibrosis and cirrhosis also inhibit the
development of hepatocellular carcinoma. Till date there is
no suitable agent that can be used against liver cirrhosis.
Compounds derived from the natural products claim effi-
cacy in liver diseases [34, 35], but do not exhibit appre-
ciable results in liver fibrosis or cirrhosis. In this study, we
provide substantial and convincing evidence suggesting the
therapeutic effect of an aqueous-alcoholic extract of dried
orange peels in animal model of liver cirrhosis.
Thioacetamide induces hepatic injury by various
mechanisms [17, 36–38], and produces various forms of
Fig. 3 HPTLC chemical fingerprinting of the test fraction reveal-
ing the presence of a limonene (kmax 357 nm, Rf 0.39 ± 0.01),
b b-sitosterol (kmax 490 nm, Rf 0.47 ± 0.02), and c ascorbic acid
(kmax 265 nm, Rf 0.87 ± 0.02). The samples were run in duplicate.
First two lanes on each plate (from left) show respective standard;
‘a1’ indicates the limonene standard, ‘a2’ the limonene in the extract,
‘b1’ is the peak of b-sitosterol standard, ‘b2’ is b-sitosterol in the
extract, ‘c1’ is the ascorbic acid standard, and ‘c2’ is the ascorbic acid
in the extract. Comparison of standards and peaks confirmed
limonene, b-sitosterol, and ascorbic acid in the test fraction
Mediterr J Nutr Metab (2011) 4:57–67 63
123

liver diseases depending on the dosage and duration of
administration: higher dose produce necrosis [39], ful-
minant hepatic failure, and early death [40], whereas lower
doses administered over a prolonged period of time cul-
minates in liver fibrosis/cirrhosis [41] and hepatocellular
carcinoma [42]. In this study, thioacetamide was used to
induce liver cirrhosis in rat model [43]. This model has
many of the alterations owing to the liver cirrhosis in
human [44–46]. In the experimental animals, liver cirrhosis
was confirmed by studying the gross organ morphology,
histology, and biochemical changes. Liver weight body
weight ratio of control and the extract-treated rats were
compared and found that the dried peel extract could
decrease the ratio significantly (Table 1). This effect could
be attributed to the changes in nutrient absorption and
metabolic utilization and efficiency that have been descri-
bed in earlier studies on thioacetamide-induced liver cir-
rhosis [47]. Further, portal hypertension and impaired bile
acid metabolism have been reported in liver cirrhosis [48],
both influencing the nutrient utilization. Biochemical
analysis of serum markers of hepatobiliary diseases (liver
cirrhosis) shows increased alkaline phosphatase and GGT
[49, 50], which has also been reported in this study
(Table 2). Extract treatment could cause a significant
decrease in the elevated levels of these enzymes. Serum
Fig. 4 Structures of a limonene, b b-sitosterol, and c ascorbic acid
Fig. 5 a TLC autographic
image demonstrating the
antioxidant activity of
C. sinensis (C.S) and pyrogallol
(P). b Comparison of the free
radical scavenging activity of
pyrogallol and C. sinensis
Fig. 6 Effect of the test preparation on [3H]-thymidine uptake by the
hepatic DNA. Data represent Mean ± SEM (n = 6). *p \ 0.05,
when compared to group III, and #p \ 0.05, when compared to Group
I. Group I: normal control, Group II: control rats receiving the test
fraction alone, Group III: treated with thioacetamide, and Group IV:
the group treated with the test fraction followed by thioacetamide
64 Mediterr J Nutr Metab (2011) 4:57–67
123

biochemical markers of liver cirrhosis corroborated with
the gross organ morphology (Fig. 2) and the tissue histo-
pathology. The extract was found to not only protect the
liver against hepatocyte damage and decrease collagen
deposition, but also ameliorated oxidative stress as indi-
cated by decreased LPO and increased GSH level in the
extract-treated rats (Table 3).
Production of ROS cause the generation of lipid per-
oxides, which have deleterious effects on cell and trigger/
sustain the metabolic and signaling pathways leading to
hepatic injury [51]. LPO is a degradative process in the
tissue arising from the production and propagation of free
radical reactions primarily involving the membrane poly-
unsaturated fatty acids and production of end products,
such as MDA and 4-hydroxynonenal [52]. MDA, one of
the major products of LPO, has been extensively used as a
sensitive oxidative stress marker [21]. Increased LPO often
depletes the level of GSH, an endogenous antioxidant that
counteracts the effect of oxidative stress due to LPO and
other causes, and also affects GR. In this study, both GSH
and GR decreased and the LPO increased, which is con-
sidered a factor for the development of liver cirrhosis.
Further, the peroxide metabolizing enzymes and the non-
microsomal phase I drug metabolizing enzyme xanthine
oxidase and GST also increased in the LC group, which
suggests the role of ROS in the development of liver
fibrosis/cirrhosis. XO generates ROS while catalyzing the
reaction. Increased production of ROS has been demon-
strated to trigger hepatic injury.
Xanthine oxidase is a Mo–Fe–S flavin hydroxylase that
initiates the production of hydroperoxides and stimulates
the tissue antioxidant armory including the catalase to
counteract the damaging effect of peroxides. As shown in
Table 4, activity of catalase decreased in the LC group, but
improved in the treated rats. Induction of XO observed in
the LC group (Table 5) indicates excess generation of
superoxide anion radicals and other ROS. As discussed
earlier, ROS is reported to activate the HSC proliferation,
which is a key event in liver cirrhosis [53]. The peel extract
could attenuate the elevated activity level of XO, inhibiting
the generation of ROS, thereby, inhibiting the activation of
HSC and, therefore, the progression of fibrosis/cirrhosis. In
this study, we suggest XO as a possible cause that might
trigger/sustain liver cirrhosis. In the group of rats receiving
the extract (Group IV), activity level of XO decreased
(Table 5), which may be attributed to the presence of
caffeic acid in oranges [54] that has been reported to inhibit
xanthine oxidase [55]. The role of XO is further substan-
tiated by measuring the activity levels of peroxide metab-
olizing enzymes (Table 4), oxidative stress biomarkers
(Table 3), serum markers of liver cirrhosis (Table 2), and
other signs and symptoms of hepatic injury (Table 1;
Fig. 2). Further, GST, which is a phase II drug
metabolizing enzyme, increased in the LC group and
decreased significantly following the post-treatment
(Table 5). GST is essential for eliminating the toxic
chemical moieties by conjugating them with GSH. This
study also demonstrates the anti-proliferative activity of the
test preparation (Fig. 6).
An overview of the findings reported in this study pro-
vides an insight into the mechanism of action of the peel
extract. While emphasizing the ability of test preparation in
attenuating the production of ROS by XO, this study sug-
gests the role of other bioactive molecules, such as the
limonene, b-sitosterol, and ascorbic acid (Figs. 3, 4) found
in the extract. Limonoids and the antioxidants have a
variety of pharmacological actions. Limonene, a major
constituent of several citrus oils (orange, lemon, mandarin,
lime, and grapefruit) inhibits rat mammary and tumor
development [56, 57], hepatic preneoplastic lesions, cell
proliferation [12], and GST [58]. GST, besides acting as a
phase II enzyme, is also involved in prostaglandin (PG)
metabolism, which has been reported to contribute to
hepatic injury.
Several studies have shown the possible benefits of
antioxidants from plant sources in altering, reversing, or
forestalling the negative effects of oxidative stress. The
antioxidants, such as polyphenols, carotenoids, and vita-
mins have an important role in cellular defense against
oxidative insult, which is pivotal to liver pathology [59,
60]. The antioxidant and the free radical activity of the
extract have been demonstrated in vivo using TLC autog-
raphy and DPPH assay (Fig. 5); free radical scavenging
action is considered to be one of the mechanisms for
antioxidation [61]. The antioxidants found in the extract
might be responsible for its effect on HSC proliferation
and collagen synthesis. The exact mechanism, however,
remains elusive, and would be the subject of future
research particularly with regard to its effects on
NF-KappaB and JNK pathways.
Conclusion
Taken together, the results provide evidence suggesting the
role of the dried peel extract of C. sinensis in liver fibrosis
and cirrhosis in animal model. The extract contains mul-
tiple constituents that appear to act synergistically. An
overview of the results led us to propose that the test
fraction acts by multiple mechanisms, which can be
attributed to the presence of several constituents in it,
instead of a single molecule.
Acknowledgments Authors acknowledge UGC for providing the
infrastucture development grant to the Department of Biochemistry,
Jamia Hamdard.
Mediterr J Nutr Metab (2011) 4:57–67 65
123

Conflict of interest None.
References
1. Kris EPM, Hecker KD, Bonanome A et al (2002) Bioactive
compounds in foods: their role in the prevention of cardiovas-
cular disease and cancer. Am J Med 113:71–88
2. Silalahi J (2002) Anticancer and health protective properties of
citrus fruit components. Asia Pac J Clin Nutr 11:79–84
3. Izquierdo AG, Gil MI, Ferreres F (2002) Effect of processing
techniques at industrial scale on orange juice antioxidant and
beneficial health compounds. J Agr Food Chem 50:5107–5114
4. Rouseff RL, Martin SF, Youtsey CO (1987) Quantitative survey
of narirutin, naringin, hesperidin, and neohesperidin in citrus.
J Agr Food Chem 35:1027–1030
5. Bocco A, Cuvelier ME, Richard H, Berset C (1998) Antioxidant
activity and phenolic composition of citrus peel and seed extracts.
J Agr Food Chem 46:2113–2129
6. Manthey JA, Grohmann K (2001) Phenols in citrus peel
byproducts. Concentrations of hydroxycinnamates and poly-
methoxylated flavones in citrus peel molasses. J Agr Food Chem
49:3268–3273
7. Lodovici M, Guglielmi F, Meoni M, Dolara P (2001) Effect of
natural phenolic acids on DNA oxidation in vitro. Food Chem
Toxicol 39:1205–1210
8. Deyhim F, Lopez E, Gonzalez J et al (2006) Citrus juice mod-
ulates antioxidant enzymes and lipid profiles in orchidectomized
rats. J Med Food 9:422–426
9. Miller EG, Sanders APG, Couvillon AM (1994) Citrus limonoids
as inhibitors of oral carcinogenesis. Food Technol 48:110–114
10. Baker RA (1994) Potential dietary benefits of citrus pectin and
fibre. Food Technol 11:133–139
11. Lam LKT, Zang J, Hasegawa S (1994) Citrus limonoid reduction
of chemically induced tumorigenesis. Food Technol 48:104–108
12. Kaji I, Tatsuta M, Iishi H et al (2001) Inhibition by d-limonene of
experimental hepatocarcinogenesis in Sprague-Dawley rats does
not involve p21(ras) plasma membrane association. Int J Cancer
93:441–444
13. Gorinstein S, Leontowicz H, Leontowicz M et al (2007) Effect of
hesperidin and naringin on the plasma lipid profile and plasma
antioxidant activity in rats fed a cholesterol-containing diet. J Sci
Food Agri 87:1257–1262
14. Tirkey N, Pilkhwal S, Kuhad A, Chopra K (2005) Hesperidin, a
citrus bioflavonoid, decreases the oxidative stress produced by
carbon tetrachloride in rat liver and kidney. BMC Pharmacology
5:2–10
15. Kaur G, Tirkey N, Chopra K (2006) Beneficial effect of hes-
peridin on lipopolysaccharide-induced hepatotoxicity. Toxicol-
ogy 226:152–160
16. Shaw SR, Nihal M, Ahmad N (2007) Dose translation from
animal to human studies revisited. FASEB J 22:261–659
17. Ali S, Pawa S, Naime M et al (2008) Role of mammalian cyto-
solic molybdenum Fe–S flavin hydroxylases in hepatic injury.
Life Sci 82:780–788
18. Reitman S, Frankel S (1957) A colorimetric method for the
determination of serum glutamate oxaloacetate transaminase. Am
J Clin Pathol 28:53–56
19. Kind PRN, King EJJ (1954) Estimation of plasma phosphatase by
determination of hydrolyzed phenol with aminopyrines. J Clin
Pathol 7:322–326
20. Orlowski M, Meister A (1973) c-Glutamyl cyclotransferase dis-
tribution, isozymic forms and specificity. J Biol Chem
248:2836–2844
21. Bernheim F, Bernheim MLC, Wilburn KM (1948) The reaction
between thiobarbituric acid and the oxidation products of certain
lipids. J Biol Chem 174:257–264
22. Jollow DJ, Mitchell JR, Zampagtrone N, Gillete JR (1974) Bro-
mobenzene induced liver necrosis. Protective role of glutathione
and evidence for 3, 4-bromobenzene oxide as the hepatotoxic
metabolite. Pharmacology 11:151–169
23. Carlberg I, Mannervick B (1975) Purification and characteriza-
tion of the flavoenzymes glutathione reductase from rat liver.
J Biol Chem 250:5475–5480
24. Mohandas J, Marshall JJ, Duggin GG et al (1984) Differential
distribution of glutathione and glutathione related enzymes in
rabbit kidney. Possible implications in analgesic nephropathy.
Biochem Pharmacol 33:1801–1807
25. Claiborne A (1984) Catalase activity. Handbook of methods in
oxygen radical research. CRC Press, Boca Raton, pp 283–284
26. Stripe F, Corte DE (1969) The regulations of rat liver xanthine
oxidase. Conversion in vitro of the enzyme activity from dehy-
drogenase (type D) to oxidase (type O). J Biol Chem 244:
3855–3863
27. Habig WH, Pabst MJ, Fleischner G et al (1974) The identity of
glutathione-s-transferase B with ligandin, a major binding protein
of liver. PNAS 71:3879–3882
28. Galvez M, Cordero CM, Houghton PJ, Ayuso MJ (2005) Anti-
oxidant activity of methanol extract obtained from Plantago
species. J Agr Food Chem 53:1927–1933
29. Yamaguchi T, Takamura H, Matoba T, Terao J (1998) HPLC
method for evaluation of the free radical-scavenging activity of
foods by using 1, 1-diphenyl-2-picrylhydrazyl. Biosci Biotech
Bioch 6:1201–1204
30. Smart RC, Huang MT, Conney AH (1986) Sn1, 2-diacylglycerols
mimic the effects of TPA in vivo by inducing biochemical
changes associated with tumor promotion in mouse epidermis.
Carcinogenesis 7:1865–1870
31. Giles KW, Myers A (1965) An improved diphenylamine method
for the estimation of deoxyribonucleic acid. Nature 206:93
32. Nissen NN, Martin P (2002) Hepatocellular carcinoma: the high-
risk patient. J Clin Gastroenterol 35:79–85
33. Okita K, Sakaida I, Hino K (2002) Current strategies for che-
moprevention of hepatocellular carcinoma. Oncology 62:24–28
34. Baek SH, Park M, Suh JH, Choi HS (2008) Protective effects of an
extract of young radish (Raphanus sativus L) cultivated with sulfur
(sulfur-radish extract) and of sulforaphane on carbon tetrachloride-
induced hepatotoxicity. Biosci Biotech Bioch 72:1176–1182
35. Ali S, Ansari KA, Jafri MA et al (2000) Nardostachys jatamansiprotects against liver damage induced by thioacetamide in rats.
J Ethanopharmacol 71:359–363
36. Hunter AL, Holscher MA, Neal RA (1997) Thioacetamide-
induced hepatic necrosis. I. Involvement of the mixed-function
oxidase enzyme system. J Pharmacol Exp Ther 200:439–448
37. Bruck R, Shirin H, Aeed H et al (2001) Prevention of hepatic
cirrhosis in rats by hydroxyl radical scavengers. J Hepatol
35:457–464
38. Bruck R, Schey R, Aeed H et al (2004) Protective effect of
pyrrolidine dithiocarbamate in a rat model of liver cirrhosis. Liver
Int 24:169–176
39. Ali S, Diwakar G, Pawa S et al (2001) Xanthine oxidase derived
reactive oxygen metabolites contribute to liver necrosis: protec-
tion by 4-hydroxypyrazolo[3,4-d] pyrimidine. BBA-Mol Basis
Dis 1536:21–30
40. Bruck R, Aeed H, Schey R (2002) Pyrrolidine dithiocarbamate
protects against thioacetamide-induced fulminant hepatic failure
in rats. J Hepatol 36:370–377
41. Pines M, Knopov V, Genina O et al (1997) Halofuginone, a
specific inhibitor of collagen type I synthesis, prevents dime-
thylnitrosamine-induced liver cirrhosis. J Hepatol 27:391–398
66 Mediterr J Nutr Metab (2011) 4:57–67
123

42. Dasgupta A, Chatterjee R, Chowdhury JR (1981) Thioacetamide
induced hepatocarcinoma in rat. Oncology 38:249–253
43. Oren R, Dotan I, Papa M et al (1996) Inhibition of experimentally
induced cirrhosis in rats by hypothyroidism. Hepatology 24:
419–423
44. Strnad P, Tao GZ, Zhou Q et al (2008) Keratin mutation pre-
disposes to mouse liver fibrosis and unmasks differential effects
of the carbon tetrachloride and thioacetamide models. Gastro-
enterology 134:1169–1179
45. Zimmermann T, Muller A, Machnik G et al (1987) Biochemical
and morphological studies on production and regression of
experimental liver cirrhosis induced by thioacetamide in Uje:
WIST rats. Z Versuchstierkd 30:165–180
46. Fontana L, Moreira E, Torres MI et al (1996) Serum amino acid
changes in rats with thioacetamide induced liver cirrhosis. Tox-
icology 106:197–206
47. Ortega MA, Torres MI, Fernandez MI et al (1997) Hepatotoxic
agent thioacetamide induces biochemical and histological alter-
ations in rat small intestine. Digest Dis Sci 42:1715–1723
48. Aris MI, Boyer JL, Fausto N et al (1994) The liver: biology and
pathology. Raven Press, New York
49. Galisteo M, Suarez A, Montilla MP et al (2006) Protective effects
of Rosmarinus tomentosus ethanol extract on thioacetamide-
induced liver cirrhosis in rats. Phytomedicine 13:101–108
50. Xiangnong L, Irving SB, Barry A (2002) Reproducible produc-
tion of thioacetamide-induced macronodular cirrhosis in the rat
with no mortality. J Hepatol 36:488–493
51. Kubow S (1995) Routes of formation of toxic consequences of
lipid oxidation products in foods. Free Radic Bio Med 12:63–81
52. Poli G (2000) Pathogenesis of liver fibrosis: role of oxidative
stress. Mol Aspects Med 21:49–98
53. Galli A, Svegliati BG, Ceni E et al (2005) Oxidative stress
stimulates proliferation and invasiveness of hepatic stellate cells
via a MMP2-mediated mechanism. Hepatology 41:1074–1084
54. Stohr H, Herrmann K (1975) On the occurrence of derivatives of
hydroxycinnamic acids, hydroxybenzoic acids, and hydrox-
ycoumarins in citrus fruits. Zeitschrift fur Lebensmittel-Unter-
suchung und-Forschung 159:305–306
55. Sarawek S, Feistel B, Pischel I, Butterweck V (2008) Flavonoids
of Cynara scolymus possess potent xanthine oxidase inhibitory
activity in vitro but are devoid of hypouricemic effects in rats
after oral application. Planta Med 74:221–227
56. Maltzman TH, Hurt LM, Elson CE et al (1989) The prevention of
nitrosomethylurea-induced mammary tumors by D-limonene and
orange oil. Carcinogenesis 10:381–781
57. Crowell PL, Gould MN (1994) Chemoprevention and therapy of
cancer by D-limonene. Crit Rev Oncol 5:1–22
58. Reicks MM, Crankshaw D (1993) Effects of D-limonene on
hepatic microsomal monooxygenase activity and paracetamol
induced glutathione depletion in mouse. Xenobiotica 23:809–819
59. Tedesco I, Russo GL, Nazzaro F et al (2001) Antioxidant effect
of red wine anthocyanins in normal and catalase inactive human
erythrocytes. J Nutr Biochem 12:505–511
60. Shi HL, Zhao BL, Xin WJ (1995) Scavenging effects of baicalin
on free radicals and its protection on erythrocyte membrane from
free radicals injury. Biochem Mol Biol Interact 35:981–994
61. Sini H, Devi KS (2004) Antioxidant activities of the chloroform
extract of Solanum trilobatum. Pharm Biol 42:462–466
Mediterr J Nutr Metab (2011) 4:57–67 67
123