Embed Size (px)
Citation preview

ORIGINAL PAPER
Does the influence of earthworms on water infiltration,nitrogen leaching and soil respiration depend on the initialsoil bulk density? A mesocosm experiment with the endogeicspecies Metaphire posthuma
Pascal Jouquet & Gaëlle Huchet & Nicolas Bottinelli &Thuy Doan Thu & Toan Tran Duc
Received: 23 September 2011 /Revised: 1 December 2011 /Accepted: 18 December 2011 /Published online: 4 January 2012# Springer-Verlag 2011
Abstract Soil compaction has a negative impact on bothearthworm abundance and diversity. Recent studies, however,suggest that earthworm cast properties are not influenced bythe initial soil bulk density. With time, earthworms couldtherefore transform soils with different bulk densities into asoil with the same physical state and thus with a similarecological functioning. This study aimed to test this hypoth-esis in two laboratory incubation experiments. First, we mea-sured the influence of soil bulk density (1.1 or 1.4 gcm−3) onthe production of cast by the endogeic earthworm speciesMetaphire posthuma. In a second experiment, we investigatedthe effect of M. posthuma on water infiltration, NH4
+, andNO3
− leaching and soil respiration at the same two soil bulkdensities. Although initially higher, earthworm casting activ-ity in soil at 1.4 gcm−3 decreased until it reached the samelevel of activity as earthworms in soil at 1.1 gcm−3. Thisbehavioral plasticity led to a transformation of compactedand loose soils, with their own functioning, to a third andsimilar state with similar hydraulic conductivity, nitrogen
leaching, and soil respiration. The consequences for soilorganization and soil functioning are discussed.
Keywords Earthworms .Metaphire posthuma . Soil bulkdensity . Soil organic matter . Nitrogen leaching . Soilorganization
Introduction
In the past decades, agricultural intensification and the socialand environmental demands to develop sustainable practices,have led to increased knowledge about the influence of culti-vation systems on soil biodiversity and its effect on ecosystemfunctioning (Altieri 1999; Barrios 2007). Where present,earthworms usually play an important role in the turnover ofsoil organic matter and nutrients in natural and agriculturalecosystems and they have been described as soil engineersbecause they create biogenic structures with different biolog-ical, physical, and chemical properties to the surrounding soilaggregates (Lavelle et al. 1997; Jouquet et al. 2006). Bycreating burrows and egesting casts, they also modify soilporosity and several studies have reported their impact onwater infiltration and solute transport in the field (e.g.,Lamandé et al. 2003; Shipitalo et al. 2004; Jouquet et al.2007, 2008; Capowiez et al. 2009; Zehe et al. 2010).
An extensive body of literature describes the influence ofsoil cultivation practices on earthworm populations. Theirsensitivity to chemical and physical disturbances has madethem a good indicator of soil quality and their presence in thefield is often considered as an indication of good agriculturalpractice (Paoletti 1999). For instance, it is generally acceptedthat soil compaction due to intense machinery traffic and
P. Jouquet (*) :G. HuchetIRD, UMR 211 BIOEMCO, Centre IRD Bondy,32 Avenue H. Varagnat,93143 Bondy Cedex, Francee-mail: [email protected]
P. Jouquet :G. Huchet : T. D. Thu : T. T. DucSFRI, Dong Ngac,Tu Liem,Hanoï, Viet Nam
N. BottinelliINRA, UR1138 INRA, Biogéochimie des Ecosystèmes Forestiers,Centre INRA de Nancy,54280 Champenoux, France
Biol Fertil Soils (2012) 48:561–567DOI 10.1007/s00374-011-0652-3

tillage leads to a decrease in earthworm number and biomass(Chan 2001; Kladivko 2001). The inverse relationship be-tween soil bulk density and earthworm abundance resultsfrom either decreased earthworm activity in compacted soilsdue to the high soil bulk density slowing down their foraging,feeding, and mating activities (Söchtig and Larink 1992; Binetet al. 1997) or the indirect effect of soil compaction on soilproperties (McKenzie et al. 2009). However, although thenegative impact of soil compaction on earthworm populationshas been observed, few studies have examined the directinfluence of soil bulk density on the properties of the biogenicstructures themselves (Buck et al. 2000).
Interestingly, when Barré et al. (2009) compared the me-chanical state of earthworm casts sampled in compacted andloose soil, they concluded that the porosity of earthworm castsis not dependent on the initial soil density and that earthwormsmay provide an efficient mechanical resilience to soils. Thissimilar physical state may be explained by soil aggregatesbeing mixed and then reorganized when earthworms ingestthe soil, resulting in a more simple organization of cast aggre-gates (e.g., more homogeneous distribution and less variablequality of organic matter) compared to the surrounding soil(Jouquet et al. 2009, 2011; Nguyen Hong et al. 2011). It couldtherefore be hypothesized that, over time, earthworms trans-form both compacted and loose soils to a similar physical statewith a similar ecological functioning. However, this assump-tion remains unverified in the field since the effect of earth-worms on soil organization and functioning is masked by thelarge set of abiotic (i.e., climate, pedology, topography) andbiotic factors (human, plants, and soil organisms from verte-brates to microorganisms) operating at the different scales,which regulate soil functioning (Dokuchaev 1899 fromBockheim et al. 2005).
In this study, we used mesocosm experiments to test theability of earthworms to modify the functioning of two soils,only differing in their bulk density. Incubation experimentswere carried out with the endogeic earthworm speciesMetaphire posthuma. Recent studies showed that this earth-worm species produces a large amount of casts (∼fivefold itsown weight per day) which are less stable that the surround-ing soil aggregates (Bottinelli et al. 2010a). As a conse-quence, rainfall events and water infiltration dissolve thesecast aggregates more easily than the surrounding soil aggre-gates leading to a compaction of the bulk soil (Bottinelli etal. 2010a). This study model was therefore useful to test twohypotheses: (1) earthworm development is reduced in themore compacted soil, but (2) the production of earthwormcasts with similar properties regardless of the initial soil bulkdensity leads to a similar organization and functioning of thewhole bulk soil. To test these hypotheses, we first comparedthe activity ofM. posthuma in loose (density 1.1 gcm−3) andcompacted soil (density 1.4 gcm−3). In a second experiment,we examined whether the influence of earthworms on soil
functioning was dependent on the initial bulk soil density.Three key ecological functions were assessed as indicatorsof soil functioning: water infiltration, N leaching, and soilrespiration.
Material and methods
Study site and model
The study was carried out from May to June 2009 at the Soiland Fertilizer Research Institute (Hanoi, Vietnam, 20° 57′N,105° 29′E). The soil is a loamy soil (clay, almost exclusivelykaolinite, 13%; silt, 50%; sand, 37%) with low organicmatter and mineral nutrients (C, 0.47 gkg−1; total N,0.29 gkg−1; total mineral N, 6.82 mg kg−1), low pH(5.61), and a bulk density of 1.1 gcm−3. This soil is almostexclusively colonized by the earthworm M. posthuma,which is a medium-sized endogeic geophagous earthworm(∼10 cm in length and 5 mm diameter on average at theadult stage) usually observed in the upper 10 cm of the soil.In northern Vietnam, M. posthuma produces approximatelyfivefold its own weight in casts per day (Bottinelli et al.2010a). Its casts range from 2 to 3 mm in diameter and aremainly found belowground (Bottinelli et al. 2010a).
Experiment 1: influence of soil bulk density on earthwormactivity
Cast production was assessed following the procedure de-scribed by Bottinelli et al. (2010a). Approximately 50 kg ofsoil was collected from the upper 10 cm of the soil, air-dried, ground-down, and sieved at 1 mm. Aggregates wereput in polyvinyl chloride cylinders (20 cm in diameter and10 cm high) and manually packed down until densities of1.1 and 1.4 gcm−3 were reached. One individual earthwormwas added per cylinder and no additional food was providedduring the experiment. Before inoculation, juvenile earth-worms were set in moist tissue paper for 24 h to void theirguts. They were then weighed. Individual earthworms wereabout 3 cm long and weighed 0.5 g on average. A plastic lidcontaining small holes covered the top of the cylinders toallow air circulation. Boxes were incubated at 30°C afterremoistening (75% of the field capacity). Surface- and be-lowground casts were sampled after 1, 5, 9, and 15 days ofincubation (total number of box012 with n03 for eachdate). Casts were easily distinguished from the bulk soilby their rounded shape and size (see Fig. 1). They werecarefully hand sorted with a spatula, air-dried and weighed.After each incubation time, earthworms were removed andtheir fresh weights determined as above. Earthworm devel-opment was assessed by comparing initial earthwormweight with earthworm weight at each sampling date.
562 Biol Fertil Soils (2012) 48:561–567

Experiment 2: soil incubation procedure
In a second soil incubation experiment, 3.5 cm of soil materialsmaller than 1 mm (same soil as above) were put into polyvi-nyl chloride boxes (9×12 and 10 cm high). A 1-cm layer ofsand and gravel covered the bottom of the boxes and a plasticpipe closed by a waterproof silicone cap was inserted into thebottom corner. As for experiment 1, soil aggregates werepacked down to reach densities of either 1.10 or 1.40 gcm−3.The boxes were inoculated, or not (“control”), with one juve-nile earthworm individual (∼0.47 g in average) and incubatedat 30°C for 2 months after remoistening (75% of the fieldcapacity). Before inoculation, the earthworms’ guts werevoided as explained above. All boxes were opened twice aweek to renew the atmosphere and measure CO2 emissions.Every 2 weeks, five boxes per treatment were randomlysampled to analyze hydraulic conductivity (see next section)and measure earthworm biomass, as described above.Nitrogen leaching was also measured through the randomselection of five other boxes. In total, 25 boxes were producedper treatment (two bulk densities and presence vs. absence ofearthworms) with n05 replications, four dates (after 15, 30,45, and 60 days) and the utilization of 10 boxes at d08 weeks(five for hydraulic conductivity and five for nitrogen leachingassessments).
Water infiltration and nitrogen leaching
Every 2 weeks, five boxes per treatment were selectedrandomly to measure hydraulic conductivity (K, meter persecond) using a Decagon minidisk infiltrometer with a 40-mm diameter base. The infiltrometer disk was placed on thesoil surface covered with a thin layer of fine well-sortedsand to provide a flat surface, as described by Lamandé et al.(2003). Infiltration rates were measured at three water
potentials (−0.6, −0.2, and −0.05 kPa) in ascending orderat the same position. The multipotential technique ofReynolds and Elrick (1991) was used to derive the relation-ship between K and the water potential from infiltration meas-urements assuming Wooding’s solution for steady-stateaxisymmetrical infiltration (Wooding 1968). Hypotheses forapplying Wooding’s equation and calculating K at each po-tential are described by Coutadeur et al. (2002). Determiningnear-saturated K at different water potential allows estimatingthe relative importance of different pores to the movement ofwater into and through the soil. According to capillary theory,infiltration at pressure heads of −0.05, −0.2, and −0.6 kPa willexclude pores with diameters equal to or larger than 6, 1.5, and0.5 mm, respectively.
Every 2 weeks, 200-ml deionized water was slowlyadded to all the remaining boxes to mimic rainfall effects(intensity ∼50 mm h−1). Excess water was collected at theoutlet of the boxes through the plastic pipe and the amountof NH4
+ and NO3− were measured for five randomly chosen
boxes with the colorimetric HACH methods and reagents.
Microbial activity
Soil respiration was assessed by measuring C–CO2 emis-sions twice a week from five randomly selected boxes forevery sampling date. Carbon dioxide emissions were mea-sured through a vial containing 5 ml aqueous NaOH solu-tion (0.5 M) to trap the CO2 emitted and a vial containing5 ml of deionized water to maintain humidity in the boxes.The CO2 produced was determined by back titration (HCl0.2 M; pH 8.6) of the NaOH trap with excess BaCl2 (1.5 M)using a DL15-potentiometric titrator.
Statistical analyses
Before analyses, data were tested for homogeneity of vari-ance using the Levene’s test and log-transformed (log10)when required. Data were afterwards analyzed by analysisof variance (ANOVA) with treatments (soil density, pres-ence of earthworm, and sampling time) as the independentvariables. Comparisons between means were made with aLSD test. All statistical calculations were carried out usingR (R Development Core Team 2008) and the package “agri-colae” for post hoc analysis. Differences were consideredsignificant only when p values were lower than 0.05.
Results
Experiment 1: earthworm casting activity
Earthworm biomass is shown in Fig. 2. The soil bulk densityand sampling date influenced earthworm biomass (ANOVA,
Fig. 1 Subsurface casts produced by M. posthuma at 10-cm depth inthe field. Casts range from 2 to 3 mm diameter (photo, P. Jouquet)
Biol Fertil Soils (2012) 48:561–567 563
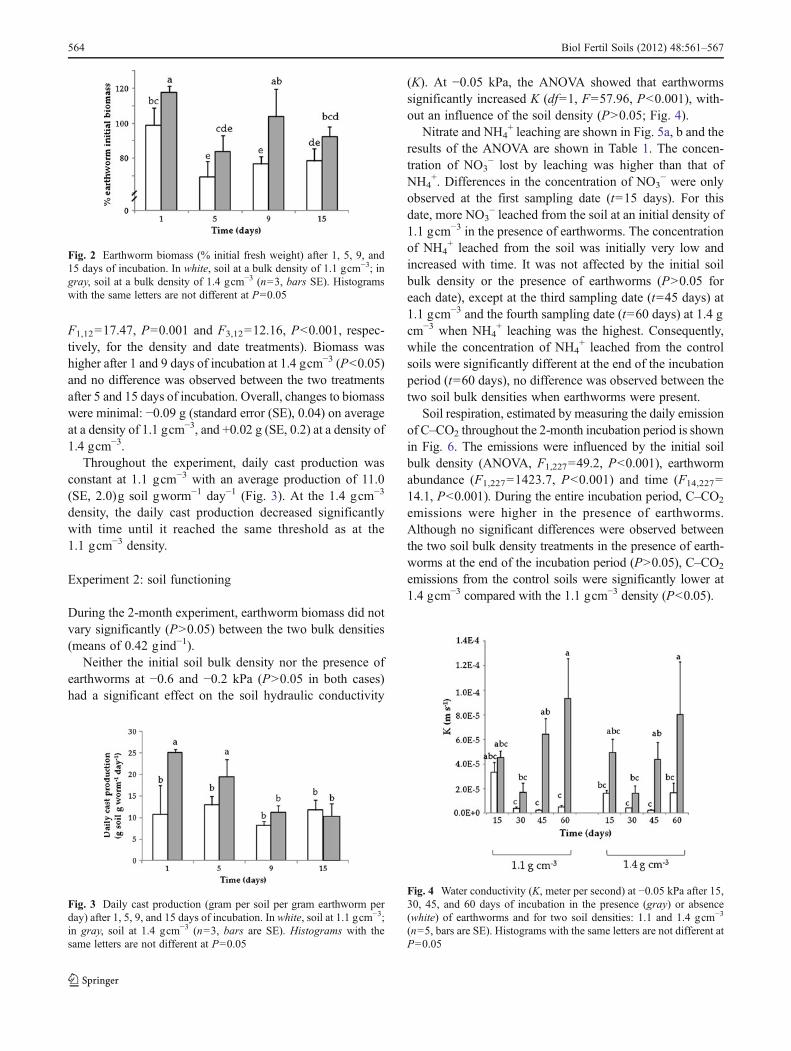
F1,12017.47, P00.001 and F3,12012.16, P<0.001, respec-tively, for the density and date treatments). Biomass washigher after 1 and 9 days of incubation at 1.4 gcm−3 (P<0.05)and no difference was observed between the two treatmentsafter 5 and 15 days of incubation. Overall, changes to biomasswere minimal: −0.09 g (standard error (SE), 0.04) on averageat a density of 1.1 gcm−3, and +0.02 g (SE, 0.2) at a density of1.4 gcm−3.
Throughout the experiment, daily cast production wasconstant at 1.1 gcm−3 with an average production of 11.0(SE, 2.0)g soil gworm−1 day−1 (Fig. 3). At the 1.4 gcm−3
density, the daily cast production decreased significantlywith time until it reached the same threshold as at the1.1 gcm−3 density.
Experiment 2: soil functioning
During the 2-month experiment, earthworm biomass did notvary significantly (P>0.05) between the two bulk densities(means of 0.42 gind−1).
Neither the initial soil bulk density nor the presence ofearthworms at −0.6 and −0.2 kPa (P>0.05 in both cases)had a significant effect on the soil hydraulic conductivity
(K). At −0.05 kPa, the ANOVA showed that earthwormssignificantly increased K (df01, F057.96, P<0.001), with-out an influence of the soil density (P>0.05; Fig. 4).
Nitrate and NH4+ leaching are shown in Fig. 5a, b and the
results of the ANOVA are shown in Table 1. The concen-tration of NO3
− lost by leaching was higher than that ofNH4
+. Differences in the concentration of NO3− were only
observed at the first sampling date (t015 days). For thisdate, more NO3
− leached from the soil at an initial density of1.1 gcm−3 in the presence of earthworms. The concentrationof NH4
+ leached from the soil was initially very low andincreased with time. It was not affected by the initial soilbulk density or the presence of earthworms (P>0.05 foreach date), except at the third sampling date (t045 days) at1.1 gcm−3 and the fourth sampling date (t060 days) at 1.4 gcm−3 when NH4
+ leaching was the highest. Consequently,while the concentration of NH4
+ leached from the controlsoils were significantly different at the end of the incubationperiod (t060 days), no difference was observed between thetwo soil bulk densities when earthworms were present.
Soil respiration, estimated by measuring the daily emissionof C–CO2 throughout the 2-month incubation period is shownin Fig. 6. The emissions were influenced by the initial soilbulk density (ANOVA, F1,227049.2, P<0.001), earthwormabundance (F1,22701423.7, P<0.001) and time (F14,2270
14.1, P<0.001). During the entire incubation period, C–CO2
emissions were higher in the presence of earthworms.Although no significant differences were observed betweenthe two soil bulk density treatments in the presence of earth-worms at the end of the incubation period (P>0.05), C–CO2
emissions from the control soils were significantly lower at1.4 gcm−3 compared with the 1.1 gcm−3 density (P<0.05).
Fig. 2 Earthworm biomass (% initial fresh weight) after 1, 5, 9, and15 days of incubation. In white, soil at a bulk density of 1.1 gcm−3; ingray, soil at a bulk density of 1.4 gcm−3 (n03, bars SE). Histogramswith the same letters are not different at P00.05
Fig. 3 Daily cast production (gram per soil per gram earthworm perday) after 1, 5, 9, and 15 days of incubation. In white, soil at 1.1 gcm−3;in gray, soil at 1.4 gcm−3 (n03, bars are SE). Histograms with thesame letters are not different at P00.05
Fig. 4 Water conductivity (K, meter per second) at −0.05 kPa after 15,30, 45, and 60 days of incubation in the presence (gray) or absence(white) of earthworms and for two soil densities: 1.1 and 1.4 gcm−3
(n05, bars are SE). Histograms with the same letters are not different atP00.05
564 Biol Fertil Soils (2012) 48:561–567

Discussion
Influence of soil bulk density on earthworm activity
The detrimental effect of soil compaction on earthwormactivity has been described several times (e.g., Binet et al.
1997; Stovold et al. 2004; Chan and Barchia 2007). In ourstudy, M. posthuma activity was constant throughout theexperiment in soil at a density of 1.1 gcm−3, which is alsothe soil density in the field where the earthworms weresampled. However, at a density of 1.4 gcm−3, M. posthumacasting production was high at the beginning of the incuba-tion period and then decreased until it reached the samelevel of production as that observed for earthworms in the1.1 gcm−3 soil. Therefore, the first hypothesis of this studycan be rejected and our findings highlight that in compactedsoil, M. posthuma is capable of a certain degree of behav-ioral plasticity which allows it to modify its environment toone more suitable for its activities. This observation is inagreement with that of Buck et al. (2000) who reported thatLumbricus terrestris is forced to ingest an increased amountof soil to construct its burrows in compacted soil. It alsoexplains why a significantly higher earthworm biomass wasobserved at density 1.4 gcm−3 than at density 1.1 gcm−3.This result is also consistent with the concepts of extendingphenotype engineering (Jouquet et al. 2006) and self-organization applied to the soil system (Lavelle et al.2006). Through the creation of elemental biogenic structures(casts and galleries), some earthworm species are able tomodify the soil until a level of organization is obtained thatis better suited to their ecological requirements.
Earthworms influence soil functioning
Compaction tends to severely limit movement, aeration, andwater and nutrient flows unless porosity is created by pow-erful physical or biological processes (Lavelle et al. 2006).Earthworms influence soil porosity at different scales. Theyincrease the quantity of large interconnected macroporespaces by creating galleries, which then constitute preferen-tial flow paths for water (i.e., Capowiez et al. 2009; Ernst etal. 2009; Zehe et al. 2010). The production of casts in thesoil is also related to a modification in the proportion of the
Fig. 5 The concentration of a NO3− and b NH4
+ (milligram per liter)in the solution leached from the soil after 15, 30, 45, and 60 days ofincubation in the presence (gray) or absence (white) of earthworms andfor the two soil densities: 1.1 and 1.4 gcm−3 (n05, bars are SE).Histograms with the same letters are not different at P00.05
Table 1 Results of the ANOVA examining the effect of the initial soildensity (1.1 vs. 1.4 gcm−3), earthworm activity (presence vs. absence)and the sampling date (t015, 30, 45, and 60 days after the beginning ofthe experiment) on leaching of NO3
− and NH4+
NO3− NH4
+
df F F
Density (1) 1 3.57 0.01
Earthworms (2) 1 46.37*** 13.18***
Date (3) 3 0.59 104.59***
(1)×(2) 1 4.04 * 0.16
(1)×(3) 3 2.59 3.67*
(2)×(3) 3 48.95*** 5.00*
(1)×(2)×(3) 3 5.27** 4.62*
*p<0.05, **p<0.01, ***p<0.001
Fig. 6 Daily C–CO2 emissions (μC-CO2 g soil−1) in the presence (full
line) or absence (dashed line) of earthworms and for two soil densities:1.1 (triangle) and 1.4 gcm−3 (square; n05, bars SE). Values measuredat the end of the incubation period with the same letters are notdifferent at P00.05
Biol Fertil Soils (2012) 48:561–567 565

macro, meso, and microporosity (Lamandé et al. 2003;Bottinelli et al. 2010a, b). However, as outlined by Chan(2001), the macroporosity created by galleries is themost important factor controlling water movement insoil, preventing flooding and soil erosion. In our study,earthworms had a significant effect on K at −0.05 kPabut not at −0.2 and −0.6 kPa, suggesting that earth-worms affected water conductivity macropores rangingfrom 1.5 to 6 mm diameter through the creation ofmacropores (i.e., galleries and porosity between casts).Although higher hydraulic conductivity as a result ofearthworm activity is in agreement with the generalassumptions developed above, we did not expect thisresult. Using the same soil material, Bottinelli et al.(2010a) showed that casts produced by M. posthumaare less water stable than the surrounding soil aggre-gates and collapse rapidly in the presence of water. As aconsequence, their findings suggested that this earth-worm species is likely to decrease soil porosity andincrease soil compaction during rainfall events. Ourstudy therefore shows that, even if occurring, the com-pacting effect of earthworm casts (negative effect onwater infiltration) was counterbalanced by the decom-pacting effect of the galleries and porosity between casts(positive effect on water infiltration).
An abundant literature stresses the influence of earth-worms on soil organic matter turnover and nutrientcycling (see Lavelle and Spain 2006 for a review).Earthworms usually influence soil organic matter dy-namics at different temporal and spatial scales. At smallscales, earthworms are assumed to increase organic mat-ter mineralization with a positive effect on soil aerationand water infiltration. It is also generally assumed thatearthworm bioturbation increases nutrient leaching,sometimes to considerable soil depths (Dominguez etal. 2004). As expected, we observed higher C–CO2
emissions which reflect increasing C mineralization inthe presence of M. posthuma. Our study also confirmedincreased leaching of NO3
−. However, this effect wasonly observed at the beginning of the incubation exper-iment, after the first application of water, and a lowleaching of NO3
− was afterwards observed, probablybecause of the low pH of the soil (<6) which hasimpeded the nitrification process during incubation.Conversely, NH4
+ leaching was reduced in the presenceof earthworms; it was stable over time but higher in thecontrol soils after the third and fourth applications ofwater, respectively, at a density of 1.1 and 1.4 gcm−3.This time lag between the two soil bulk densities islikely to be explained by the reduced microbial activityin more compacted soils (Beylich et al. 2010).Moreover, microbes and earthworms may compete forN and/or the reorganization of the soil structure may
lead to less NH4+ being lost from the system in the
presence of earthworms.
Earthworm activity leads to homogenization of the soilfunctioning
In agreement with our second hypothesis, the main conclu-sion of this study is that M. posthuma activity transformedcompacted and loose soils, with their own functioning, to athird and similar state with the same hydraulic conductivity,nitrogen leaching and soil respiration. Consequently, ourstudy strongly supports the idea developed in the study ofBarré et al. (2009) at the soil aggregate level, which can nowbe extrapolated to the bulk soil, that, without any otherforcing actors (i.e., plant roots, other earthworm species…)and in soils only differing in their bulk densities, endogeicearthworms are likely to favor the homogenization of soilfunctioning.
If confirmed in the field, this impact of earthworms onsoil organization is likely to be important in the regulation ofsoil functioning in heterogeneous environments, especiallyagro-ecosystems with areas of compacted and looser soils.As mentioned above, the soil organization regulates vitalecosystem functions, such as soil organic matter decompo-sition, water flow, and nutrient cycling. Therefore, furtherfield studies would be of interest to confirm our findings inthe field, with other species, and functional groups, anddetermine to what extent earthworms can decrease entropyin ecosystems.
Acknowledgments We would like to thank Laurence Goury forproviding us with an abundant source of literature on the influence ofsoil compaction on earthworms and Sandrine Caquineau for the anal-ysis of soil mineralogy. This project was supported financially byCNRS/INSU (VERAGREGAT project under the framework of theEC2CO program), IRD (unit research UMR-211-BIOEMCO) andCNRS (unit research UMR-7618-BIOEMCO) French institutes andthe Management of Soil Erosion Consortium (MSEC3) from IRD.
References
Altieri MA (1999) The ecological role of biodiversity in agroecosys-tems. Agr Ecosyst Environ 74:19–31
Barré P, McKenzie BM, Hallett PD (2009) Earthworms bring com-pacted and loose soil to a similar mechanical state. Soil BiolBiochem 41:656–658
Barrios E (2007) Soil biota, ecosystem services and land productivity.Ecol Econ 64:269–285
Beylich A, Oberholzer HR, Schrader S, Höper H, Wilke BM (2010)Evaluation of soil compaction effects on soil biota and soil bio-logical processes in soils. Soil Till Res 109:133–143
Binet F, Hallaire V, Curmi P (1997) Agricultural practices and thespatial distribution of earthworms in maize fields. Relationshipsbetween earthworm abundance, maize plants and soil compaction.Soil Biol Biochem 29:577–583
566 Biol Fertil Soils (2012) 48:561–567

Bockheim JG, Gennadiyev AN, Hammer RD, Tandarich JP (2005)Historical development of key concept in pedology. Geoderma124:23–36
Bottinelli N, Henry-des-Tureaux T, Hallaire V, Benard Y, Mathieu J,Duc Tran T, Jouquet P (2010a) How earthworms accelerate soilporosity under watering events. Geoderma 156:43–47
Bottinelli N, Hallaire V, Menasseri-Aubry S, Le Guillou C, Cluzeau D(2010b) Abundance and stability of belowground earthworm castsinfluenced by tillage intensity and depth. Soil Till Res 106:263–267
Buck C, Langmaack M, Schrader S (2000) Influence of mulch and soilcompaction on earthworm cast properties. Appl Soil Ecol 14:223–229
Capowiez Y, Cadoux S, Bouchant P, Ruy S, Roger-Estrade J, RichardG, Boizard H (2009) The effect of tillage type and croppingsystem on earthworm communities, macroporosity and waterinfiltration. Soil Till Res 105:209–216
Chan KY (2001) An overview of some tillage impacts on earthwormpopulation abundance and diversity: implications for functioningin soils. Soil Till Res 57:179–191
Chan KY, Barchia I (2007) Soil compaction controls the abundance,biomass and distribution of earthworms in a single dairy farm insouth-eastern Australia. Soil Till Res 94:75–82
Coutadeur C, Coquet Y, Roger-Estrade J (2002) Variation of hydraulicconductivity in a tilled soil. Eur J Soil Sci 53:619–628
Dokuchaev VV (1899) A contribution to the theory of natural zones:horizontal and vertical soil zones (in Russian). Mayor’s OfficePress, St. Petersburg, Russia, 62 pp
Dominguez J, Bohlen PJ, Parmelee RW (2004) Earthworms increasenitrogen leaching to greater soil depths in row crop agroecosys-tems. Ecosystems 7:672–685
Ernst G, Felten D, Vohland M, Emmerling C (2009) Impact of ecolog-ically different earthworm species on soil water characteristics.Eur J Soil Biol 45:207–213
Jouquet P, Dauber J, Lagerlof J, Lavelle P, Lepage M (2006) Soilinvertebrates as ecosystem engineers: intended and accidentaleffects on soil and feedback loops. Appl Soil Ecol 32:153–164
Jouquet P, Bernard-Reversat F, Bottinelli N, Orange D, Rouland C,Tran Duc T, Podwojewski P (2007) Influence of land use changeand earthworm activity on carbon and nitrogen dynamics in asteepland ecosystem in Northern Vietnam. Biol Fert Soils 44:69–77
Jouquet P, Podwojewski P, Bottinelli N, Mathieu J, Ricoy M, OrangeD, Tran Duc T, Valentin C (2008) Above-ground earthworm castsaffect water runoff and soil erosion in Northern Vietnam. Catena74:13–21
Jouquet P, Zangerlé A, Rumpel C, Brunet D, Bottinelli N, Tran Duc T(2009) Relevance of the biogenic and physicogenic classification.
A comparison of approaches to discriminate the origin of soilaggregates. Eur J Soil Sci 60:1117–1125
Jouquet P, Ngo Thi P, Henry-des-Tureaux T, Chevallier T, Trand Duc T(2011) Is earthworm cast fragmentation a linear process? Conse-quences for SOM mineralization and nutrient leaching. Appl SoilEcol 47:24–30
Kladivko EJ (2001) Tillage systems and soil ecology. Soil Till Res61:61–76
Lamandé M, Hallaire V, Curmi P, Pérès G, Cluzeau D (2003) Changesof pore morphology, infiltration and earthworm community in aloamy soil under different agricultural managements. Catena54:637–649
Lavelle P, Spain A (2006) Soil ecology. Kluwer, Amsterdam, TheNetherlands, 651 pp
Lavelle P, Bignell D, Lepage M (1997) Soil function in a changingworld: the role of invertebrate ecosystem engineers. Eur J SoilBiol 33:159–193
Lavelle P, Decaëns T, Aubert M, Barot S, Blouin M, Bureau F,Margerie P, Mora P, Rossi JP (2006) Soil invertebrates and eco-system services. Eur J Soil Biol 42:3–15
McKenzie BM, Kuhner S, MacKenzie K, Peth S, Horn R (2009) Soilcompaction by uniaxial loading and the survival of the earthwormAporrectodea caliginosa. Soil Till Res 104:320–323
Nguyen Hong H, Rumpel C, Henry-des-Tureaux T, Bardoux G, BillouD, Tran Duc T, Jouquet P (2011) How do earthworms influenceorganic matter quality in tropical soil? A spatial dependent con-clusion. Soil Biol Biochem 43:223–230
Paoletti MG (1999) The role of earthworms for assessment of sustain-ability and as bioindicators. Agr Ecosyst Env 74:137–155
R Development Core Team (2008) R: a language and environment forstatistical computing. R Foundation for Statistical Computing,Vienna, Austria
Reynolds WD, Elrick DE (1991) Determination of hydraulic conduc-tivity using a tension infiltrometer. Soil Sci Soc Am J 55:633–639
Shipitalo MJ, Nuutinen V, Butt KR (2004) Interaction of earthwormburrows and cracks in a clayey subsurface-drained soil. Appl SoilEcol 26:209–217
Söchtig W, Larink O (1992) Effect of soil compaction on activity andbiomass of endogeic lumbricids in arable soils. Soil Biol Biochem24:1595–1599
Stovold RJ, Whalley WR, Harris PJ, White RP (2004) Spatial variationin soil compaction, and the burrowing activity of the earthwormAporrectodea caliginosa. Biol Fert Soils 39:360–365
Wooding RA (1968) Steady infiltration from a shallow circular pond.Water Resour Res 4:1259–1273
Zehe E, Blume T, Blöschl G (2010) The principle of maximum energydissipation: a novel thermodynamic perspective on rapid waterflow in connected soil structures. Phil Trans Royal Soc B Biol Sci365:1377–1386
Biol Fertil Soils (2012) 48:561–567 567







![Chemical Engineering Journal · [23] Mesocosm Meat processing wastewater Glyceria maxima TN: 46–49 New Zealand [24] Mesocosm Nutrient solution Canna sp., Calamus sp. TN: 76.94;](https://img.dokumen.tips/doc/110x75/60948f4f85c3c96d7a4daeca/chemical-engineering-journal-23-mesocosm-meat-processing-wastewater-glyceria-maxima.jpg)





















