Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS 141, 389-~92 (1970)
Displacement of Histones from Deoxynucleoprotein by Protamine’
During spermatogenesis in some organisms (e.g., fish) the histones of sperm&ids gradually disappear as protamine appears (1, 2). Since there is no turnover of DNA in this process, protamines seem to displace the histones either directly or in- directly. Mirsky and Ris (3) demonstrated that protamines in vitro could displace histones directly from calf thymus nucleoprotein and it thus seemed possible that a direct displacement mechanism might be involved in spermatogenesis. In this early work, however, the displaced histone was not characterized in current terms and it was not clear whether the conditions and characteristics of the in vitro displacement approximated those of the in vivo system. The present study was under- taken to characterize the displaced histones and to make a comparison of the in vivo and in vitro systems.
Quite recently Marushige and Dixon (4) estab- lished that arginine-rich histones are replaced by protamine earlier in spermatogenesis than are the lysine-rich histones. Marushige, Ling, and Dixon (5) also obtained evidence that the displacement consists, at least in part, of an active process, namely, phosphorylation of the histones. The work about to be reported showed that the direct dis- placement of histones by protamines in tilro did not release the major histone classes in the same order found in tivo. Indeed, the characteristics of the in vitro system were so different from those of the in tivo system that it seems probable that the dependence of in viva displacement upon active processes is profound.
Protamine chloride, Grade V, obtained from Sigma Chemical Co. (St. Louis, MO.) was purified by chromatography on Amberlite IRC-50 (6). The column was developed successively with 250 ml of each of the following buffers; 0.1 M sodium phos- phate-8.5yo guanidinium chloride, pH 6.8, 0.1 M sodium phosphate-14Ya guanidinium chloride, pH 6.8, and 0.1 M sodium phosphatee4Oya guanidin-
1 Supported by USPHS Grants AM 12618 and AM 08845 and by Biomedical Sciences Support Grant FR-7006 from the General Research Support Branch, Division of Research Resources, Bureau of Health Professions Education and Manpower Training, National Institutes of Health. Support of this work by the Agricultural Experiment Sta- tion is also acknowledged.
ium chloride, pH 6.8. The last peak contained the protamine which was subsequently desalted by elution from Sephadex G-100 (Pharmacia) with 0.02 N HCI. The pooled fraction from Sephadex was lyophilized, dissolved in distilled water, and adjusted to pH 7 by addition of sodium phosphate. The solution was stored at - 19”.
Chicken erythrocyte deoxyribonucleoprotein was prepared from fresh blood by the method of Murray et al. (7). The resulting gelatinous pellet w&4 frozen until just prior to incubation when it was thawed, homogenized, and assayed with diphenylamine according to Dische (8).
Incubations of deoxyribonucleoprotein with protamine were all carried out at room tempera- ture. Deoxynucleoprotein was suspended at vari- ous concentrations equivalent to 0.5 to 5 mg/ml DNA, in 4.7 mM MgCl,, 0.016 M Tris, pH 7.2. Pro- tamine was added to give final concentrations varying from 0.2-0.8 mu. In kinetic studies the viscous mixture (total volume 12 ml) was shaken for a period of 3 hr during which l.O-ml aliquots were removed at intervals for analysis; in all other cases each incubate of 1.0 ml was analyzed directly after shaking gently for 1 hr. The reaction in each sample was stopped by filtration through a Milli- pore filter with a pore size of 0.45 p. Displaced pro- teins were analyzed by electrophoresis on poly- acrylamide gels according to Reisfeld et al. (9) at 10 mA/gel. After staining with amido black dye and destaining, the gels were scanned in a Gilford recording spectrophotometer.
It is widely held (e.g., (10) that the histones and their binding to chromatin do not differ much from one tissue to the next. For this reason and for the sake of convenience, it was decided that the de- oxynucleoprotein to be studied would be that from chicken erythrocytes rather than that from fish testes.2 The erythrocyte system allowed a good comparison of displacement by salt, since it has been described in detail (11). Moreover, chicken erythrocytes possess an extra histone, with well-
2 After completing this work we learned that Marushige and Dixon (personal communication) had observed in vitro that protamine displaced histone from the chromatin of trout testes in a manner similar to that described here, and that the displacement was insensitive to N&l concen- tration at least up to 0.25 M.
389
390 COMMUNICATIONS
3
2
I 4
A a
0 -0
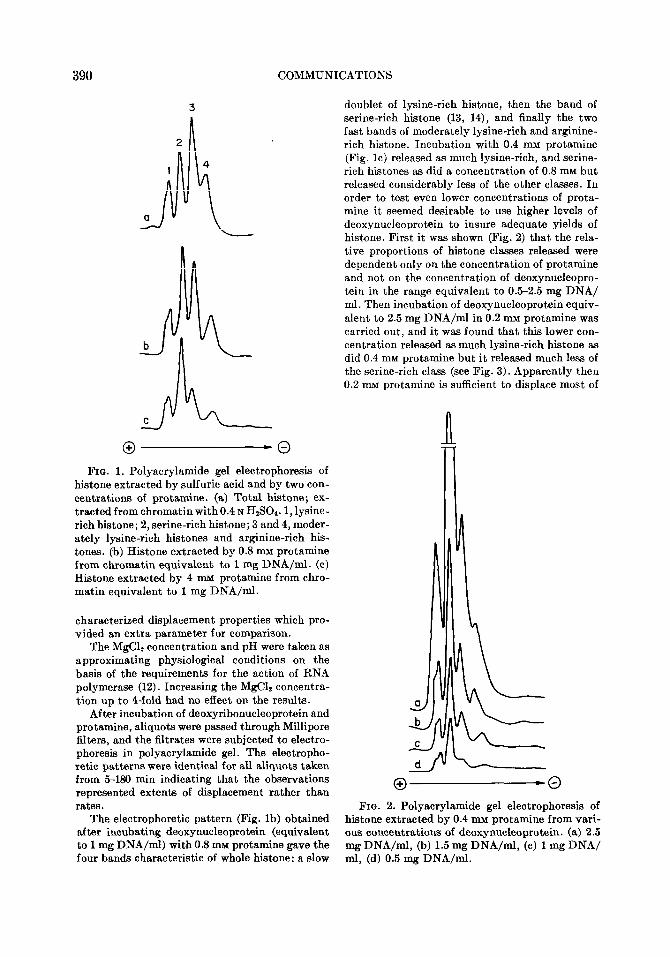
FIG. 1. Polyscrylamide gel electrophoresis of histone extracted by sulfuric acid and by two con- centrations of protamine. (a) Total histone; ex- tracted from chromatin with0.4 N H801.1, lysine- rich histone; 2, serine-rich histone; 3 and 4, moder- ately lysine-rich histones and arginine-rich his- tones. (b) Histone extracted by 0.8 mu protamine from chromatin equivalent to 1 mg DNA/ml. (c) Histone extracted by 4 mu protamine from chro- matin equivalent to 1 mg DNA/ml.
characterized displacement properties which pro- vided an extra parameter for comparison.
The MgClt concentration and pH were taken as approximating physiological conditions on the basis of the requirements for the action of RNA polymerase (12). Increasing the MgClz concentra- tion up to 4-fold had no effect on the results.
After incubation of deoxyribonucleoprotein and protamine, aliquots were passed through Millipore filters, and the filtrates were subjected to electro- phoresis in polyacrylamide gel. The electropho- retie patterns were identical for all aliquots taken from 5-130 min indicating that the observations represented extents of displacement rather than rates.
The electrophoretic pattern (Fig. lb) obtained after incubating deoxynucleoprotein (equivalent to 1 mg DNA/ml) with 0.8 mu protamine gave the four bands characteristic of whole histone: a slow
doublet of lysine-rich histone, then the band of serine-rich histone (13, 14), and finally the two fast bands of moderately lysine-rich and arginine- rich histone. Incubation with 0.4 nnu protamine (Fig. lc) released as much lysine-rich, and serine- rich histones as did a concentration of 0.8 mu but released considerably less of the other classes. In order to test even lower concentrations of prota- mine it seemed desirable to use higher levels of deoxynucleoprotein to insure adequate yields of histone. First it was shown (Fig. 2) that the rela- tive proportions of histone classes released were dependent only on the concentration of protamine and not on the concentration of deoxynucleopro- tein in the range equivalent to 0.5-2.5 mg DNA/ ml. Then incubation of deoxynucleoprotein equiv- alent to 2.5 mg DNA/ml in 0.2 mu protamine was carried out, and it was found that this lower con- centration released as much lysine-rich histone as did 0.4 mM protamine but it released much less of the serine-rich class (see Fig. 3). Apparently then 0.2 mu protamine is sufficient to displace most of
0 -0 FIG. 2. Polyacrylamide gel electrophoresis of
histone extracted by 0.4 mu protamine from vari- ous concentrations of deoxynucleoprotein. (a) 2.5 mg DNA/ml, (b) 1.5 mg DNA/ml, (c) 1 mg DNA/ ml, (d) 0.5 mg DNA/ml.
COMMUNICATIONS 391
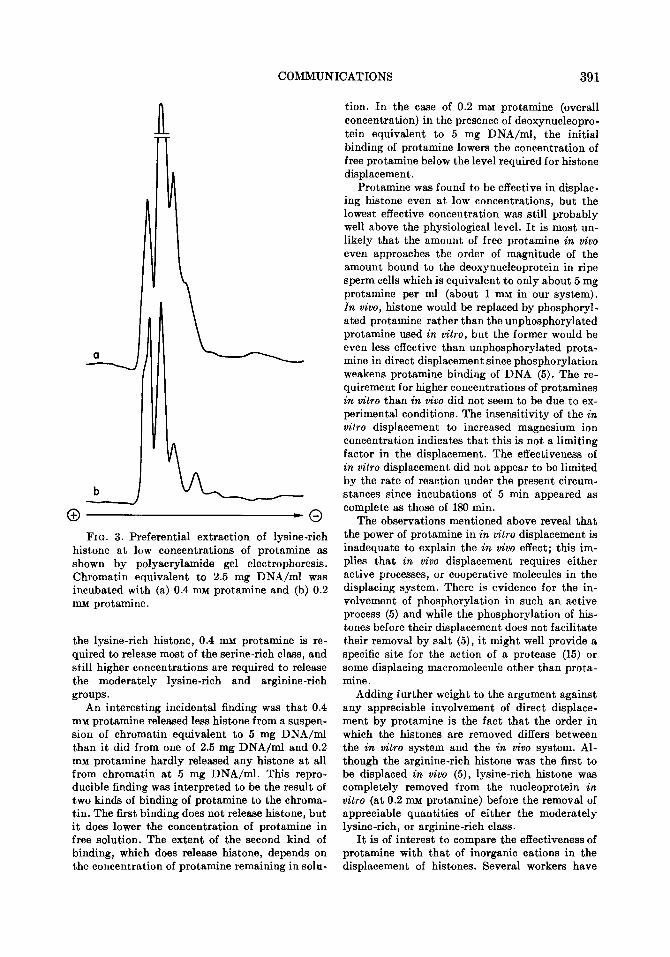
O-0 FIG. 3. Preferential extraction of lysine-rich
histone at low concentrations of protamine as shown by polyacrylamide gel electrophoresis. Chromatin equivalent to 2.5 mg DNA/ml was incubated with (a) 0.4 mu protamine and (b) 0.2 mM protamine.
the lysine-rich histone, 0.4 mM protamine is re- quired to release most of the serine-rich class, and still higher concentrations are required to release the moderately lysine-rich and arginine-rich groups.
An interesting incidental finding was that 0.4 mu protamine released less histone from a suspen- sion of chromatin equivalent to 5 mg DNA/ml than it did from one of 2.5 mg DNA/ml and 0.2 mu protamine hardly released any histone at all from chromatin at 5 mg DNA/ml. This repro- ducible finding was interpreted to be the result of two kinds of binding of protamine to the chroma- tin. The first binding does not release histone, but it does lower the concentration of protamine in free solution. The extent of the second kind of binding, which does release histone, depends on the concentration of protamine remaining in soln-
tion. In the case of 0.2 mu protamine (overall concentration) in the presence of deoxynucleopro- tein equivalent to 5 mg DNA/ml, the initial binding of protamine lowers the concentration of free protamine below the level required for histone displacement.
Protamine was found to be effective in displac- ing histone even at low concentrations, but the lowest effective concentration was still probably well above the physiological level. It is most un- likely that the amount of free protamine in z&o even approaches the order of magnitude of the amount bound to the deoxynucleoprotein in ripe sperm cells which is equivalent to only about 5 mg protamine per ml (about 1 mu in our system). In wivo, histone would be replaced by phosphoryl- ated protamine rather than the unphosphorylated protamine used in vitro, but the former would be even less effective than unphosphorylated prota- mine in direct displacement since phosphorylation weakens protamine binding of DNA (5). The re- quirement for higher concentrations of protamines in vitro than in tivo did not seem to be due to ex- perimental conditions. The insensitivity of the in vitro displacement to increased magnesium ion concentration indicates that this is not a limiting factor in the displacement. The effectiveness of in vitro displacement did not appear to be limited by the rate of reaction under the present circum- stances since incubations of 5 min appeared as complete as those of 130 min.
The observations mentioned above reveal that the power of protamine in in vitro displacement is inadequate to explain the in vivo effect; this im- plies that in tivo displacement requires either active processes, or cooperative molecules in the displacing system. There is evidence for the in- volvement of phosphorylation in such an active process (5) and while the phosphorylation of his- tones before their displacement does not facilitate their removal by salt (5), it might well provide a specific site for the action of a protease (15) or some displacing macromolecule other than prota- mine.
Adding further weight to the argument against any appreciable involvement of direct displace- ment by protamine is the fact that the order in which the histones are removed differs between the in vitro system and the in vivo system. Al- though the arginine-rich histone was the first to be displaced in wivo (5), lysine-rich histone was completely removed from the nucleoprotein in vitro (at 0.2 IMI protamine) before the removal of appreciable quantities of either the moderately lysine-rich, or arginine-rich class.
It is of interest to compare the effectiveness of protamine with that of inorganic cations in the displacement of histones. Several workers have

392 COMMUNICATION
exploited the early findings of Daly and Mirsky (16) that histones are released from nucleoprotein progressively by a series of increasing concentra- tions of NaCl or acids. In particular it has been shown (11) that lysine-rich histones are released first at 0.4-0.5 M NaCl, the serine-rich second at 0.6-0.7 M, and finally the other two classes between 1 and 2 M NaCI. The concentration of protamine which we found to remove just the lysine-rich and serine-rich histones was from 0.2-0.4 mu so that the protamine seems to be about lOOC-2000 times more effective than Na+ on a molar basis and about 3066 times more effective per positive charge.
Although the protamine is much more powerful than Na+, the order in which it displaces histones is like that of displacement by salt, and its in- creased effectiveness is plausible for the coopera- tive action expected of a polycation. In conclusion, then, it seems that the direct displacement of histones by protamines resembles displacement by salts and acids rather than the in a-ivo process seen in spermatogenesis.
REFERENCES
1. ALFERT, M., J. Biophys. Biochem. Cytol. 2, 109 (1956).
2. ANDO, T., AND HASHIMOTO, C., J. Biochem. Tokyo 46, 529 (1958).
3. MIRSKY, A. E., .~ND RIS, H., J. Gen. Physiol. 34, 475 (1961).
4. MARUSHIGE, K., AND DIXON, G. H., Develop. Biology 19, 397 (1969).
5. MARUSHIGE, K., LING, V., AND DIXON, G. H., J. Biol. Chem. 244, 5953 (1969).
6. RISMUSSEN, P. S., MURRAY, K., AND LUCK,
J. M., Biochemistry 1, 79 (1962). 7. MURRAY, K., VIDALI, G., AND NEELIN, J. M.,
Biochem. J. 107, 207 (1968). 8. DISCHE, Z., Mikrochemie 8, 4 (1930). 9. REISFELD, R. A., LEWIS, U. J., .~ND WILLIAMS,
D. E., Nature London 196, 281 (1962). 10. HNILIC~, L. S., Progr. Nucl. Acid Res. Mol.
Biol. 7, 25 (1967). 11. WILHELM, x., AND CHAMPAGNE, M., Eur. J.
Biochem. 10, 102 (1969). 12. CHAMBERLIN, M., .~ND BERG, P., Proc. Nat.
Acad. Sci. U.S.A. 48, 81 (1962). 13. NEELIN, J. M., in “The Nucleohistones” (J.
Bonner and P.O.P. Ts’o, eds.), p. 66. Hol- den-Day, San Francisco (1964).
14. HNILICA, L. S., Ezperientia 20, 13 (1964). 15. FURLAN, M., JERICIJO, M., Biochim. Biophys.
Acta 147, 135 (1967). 16. DALY, M. M., AND MIRSKY, A. E., J. Gen.
Physiol. 88, 405 (1955). KATHERINE EVANS
PAULA KONIGSBERG~
R. D.~VID COLE
Department of Biochemistry University of California Berkeley, California 94720
Received July 13, 1970 -
3 Present address: Department of Biology, State University of New York, Stony Brook, New York 11790.





























