Embed Size (px)
Citation preview

THE JOURNAL 0 1986 by The American Society of Biological Chemists, Inc.
OF BIOLOGICAL CHEMISTRY Vol. 261, No. 3, Issue of January 25, pp. 1071-1076,1986 Printed in U.S.A.
Nonrandom Utilization of Acetylation Sites in Histones Isolated from Tetrahymena EVIDENCE FOR FUNCTIONALLY DISTINCT H4 ACETYLATION SITES*
(Received for publication, May 21, 1985)
Louis G. Chicoine, Ira G. Schulman, Ronald Richman, Richard G. Cook$, and C. David Allis From the Verna and Marrs McLean Department of Biochemistry and the $Howard Hughes Medical Institute and Department of Microbiology and Immunology, Baylor College of Medicine, Houston, Texas 77030
Macro- and micronuclei of the ciliated protozoan Thus, these data show that H3 is also acetylated in a Tetrahymena thermophila afford a unique opportunity nonrandom fashion in macronuclei. to study histone acetylation under conditions where Finally, we have determined which acetylation sites postsynthetic “transcription”-related acetylation and are utilized in macro- or micronuclear H4 when it is synthetic “deposition”-related acetylation are nonov- undergoing active synthesis and deposition. We have erlapping. Recent studies have demonstrated that at determined that two sites, lysines 4 and 11, are used least two general systems of acetylation operate in exclusively in newly synthesized diacetylated H4 Tetrahymena. One is postsynthetic, macronuclear spe- whether it is destined for macro- or micronuclei. Inter- cific, and may be related to gene expression in that estingly, one of these sites (1 1) is not utilized in steady- nucleus (Vavra, K. J., Allis, C. D., and Gorovsky, M. state populations of diacetylated H4 isolated from ma- A. (1982) J. Biol. Chem. 257,2591-2598). The other cronuclei of growing cells (which use 4 and 7 exclu- is synthetic, common to macro- and micronuclei, and sively). This observation suggests the interesting pos- is likely related to histone deposition during repli- sibility that functionally distinct acetylation sites may cation (Allis, C. D., Chicoine, L. G., Richman, R., and exist in histone H4. Schulman, I. G. (1985a) Proc. Natl. Acad. Sci. U. S. A.,
A unique feature of H3 and H4 in Tetrahymena is 82,8048-8052).
that neither are blocked at their amino termini. We Modification of histones by acetylation of specific internal have exploited this fact as well 8s the resolving CaPa- lysine residues is an active metabolic process whose func-
crosequencing techniques to examine whether llti1iza- cent evidence has been obtained (Vavra et al., 1982; Allis et Of different NHz-termind sites is ran- al., 1985a; Gorovsky, 1985) suggesting that at least two general dom or nonrandom during the progression toward high
acetylation states. Of the four acetylation sites which systems of histone acetylation operate in Tetrahymena. One have been identified in H4 (in Tetrahymena these are of these is presumably identical between macro- and micro- lysines at positions 4, 7, 11, and 15), we find that nuclei of growing, starved, or mating cells and is related to lysine 7 is the site of postsynthetic acetyla- histone deposition at the time of DNA replication. The other tion in populations of monoacetylated H4 isolated from is macronuclear specific and may play a role in the modulation macronuclei. This site is retained in populations of Of transcriptional activity in this nucleus. &acetylated H4, which are acetylated exclusively at Numerous groups have identified the sites of acetylation in lysines 4 and 7. Our data also suggest that there is non-H1 histones by amino acid sequence analysis (see Isen- some preference for using lysine 11 (as compared to berg, 19791, and in the case of certain histones, H3 and H4, 15) as the third site of acetylation in triacetylated these acetylation sites have been highly conserved throughout molecules. The data demonstrate that the postsynthetic evolution (Gorovsky, 1985). However, few reports have ad- acetylation-deacetylation process is surprisingly non- dressed the question whether specific acetylation sites are random for H4 in Tetrahymena macronuclei. preferred more than others (Candido and Dixon, 1971; Ruiz-
We have also investigated the same question with Carrillo et al., 1975), and as far as we are aware, no study has macronuclear H3 (which contains acetylation sites at addressed whether acetylation site utilization is random or lysines 9, 14, 18, and 23)- Our data demonstrate that nonrandom during the progression toward high acetylation unlike H4, lysines at position 9 or 14 are likely to be states. utilized as sites of acetylation within a population of In this study we have exploited the fact that neither H3 monoacetylated H3. Both of these acetylation sites are nor H4 in Tetrahymena is blocked at their amino termini to
9 is used initially as the site of monoacetylation, 14 is site utilization is random or nonrandom. In particular, we used secondarily (and vice versa). Our data show, moreover, that there is a preference for lysine have addressed whether different acetylation sites are utilized 18 85 the third acetylation site (in triacetylated H3). during “transcription”- and “deposition”-related H4 acetyla-
tion. Our results demonstrate that surprising regularity exists among the amino acid sites utilized during postsynthetic
Grant HD 16259 to C. D. A. The costs of publication of this article Of H4 and H3. we have were defrayed in part by the payment of page charges. This article determined which internal lysines in H4 are acetylated under must therefore be hereby marked “aduertisement” in accordance with conditions of active synthesis and deposition. Interestingly, 18 U.S.C. Section 1734 solely to indicate this fact. two of four possible acetylation sites are used exclusively
1071
bility of acid-urea gel electrophoresis and current mi- tion(s) is poorly understood and remains controversial. Re-
retained in diacetylated H3 which suggests that if site investigate the question of whether or not histone acetylation
*This research was supported by National Institutes of Health
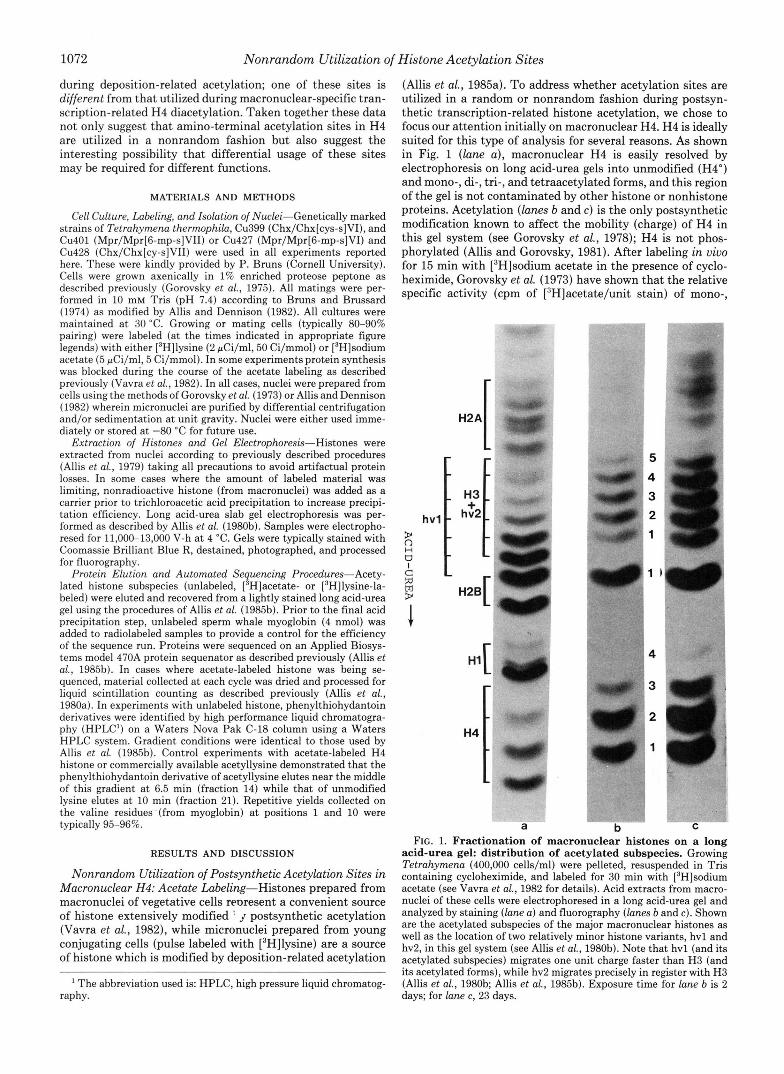
1072 Nonrandom Utilization of Histone Acetylation Sites
during deposition-related acetylation; one of these sites is different from that utilized during macronuclear-specific tran- scription-related H4 diacetylation. Taken together these data not only suggest that amino-terminal acetylation sites in H4 are utilized in a nonrandom fashion but also suggest the interesting possibility that differential usage of these sites may be required for different functions.
MATERIALS AND METHODS
Cell Culture, Labeling, and Isolation of Nuclei-Genetically marked strains of Tetrahymena thermophila, Cu399 (Chx/Chx[cys-SIVI), and Cu401 (Mpr/Mpr[G-mp-s]VII) or Cu427 (Mpr/Mpr[G-mp-s]VI) and Cu428 (Chx/Chx[cy-s]VII) were used in all experiments reported here. These were kindly provided by P. Bruns (Cornel1 University). Cells were grown axenically in 1% enriched proteose peptone as described previously (Gorovsky et al., 1975). All matings were per- formed in 10 mM Tris (pH 7.4) according to Bruns and Brussard (1974) as modified by Allis and Dennison (1982). All cultures were maintained at 30 "C. Growing or mating cells (typically 80-90% pairing) were labeled (at the times indicated in appropriate figure legends) with either [3H]lysine (2 pCi/ml, 50 Ci/mmol) or [3H]sodium acetate (5 pCi/ml, 5 Ci/mmol). In some experiments protein synthesis was blocked during the course of the acetate labeling as described previously (Vavra et al., 1982). In all cases, nuclei were prepared from cells using the methods of Gorovsky et al. (1973) or Allis and Dennison (1982) wherein micronuclei are purified by differential centrifugation and/or sedimentation at unit gravity. Nuclei were either used imme- diately or stored at -80 "C for future use.
Extraction of Histones and Gel Electrophoresis-Histones were extracted from nuclei according to previously described procedures (Allis et al., 1979) taking all precautions to avoid artifactual protein losses. In some cases where the amount of labeled material was limiting, nonradioactive histone (from macronuclei) was added as a carrier prior to trichloroacetic acid precipitation to increase precipi- tation efficiency. Long acid-urea slab gel electrophoresis was per- formed as described by Allis et al. (1980b). Samples were electropho- resed for 11,000-13,000 V-h at 4 "C. Gels were typically stained with Coomassie Brilliant Blue R, destained, photographed, and processed for fluorography.
Protein Elution and Automated Sequencing Procedures-Acety- lated histone subspecies (unlabeled, [3H]acetate- or [3H]lysine-la- beled) were eluted and recovered from a lightly stained long acid-urea gel using the procedures of Allis et al. (1985b). Prior to the final acid precipitation step, unlabeled sperm whale myoglobin (4 nmol) was added to radiolabeled samples to provide a control for the efficiency of the sequence run. Proteins were sequenced on an Applied Biosys- tems model 470A protein sequenator as described previously (Allis et al., 1985b). In cases where acetate-labeled histone was being se- quenced, material collected at each cycle was dried and processed for liquid scintillation counting as described previously (Allis et al., 1980a). In experiments with unlabeled histone, phenylthiohydantoin derivatives were identified by high performance liquid chromatogra- phy (HPLC') on a Waters Nova Pak C-18 column using a Waters HPLC system. Gradient conditions were identical to those used by Allis et al. (1985b). Control experiments with acetate-labeled H4 histone or commercially available acetyllysine demonstrated that the phenylthiohydantoin derivative of acetyllysine elutes near the middle of this gradient at 6.5 min (fraction 14) while that of unmodified lysine elutes at 10 min (fraction 21). Repetitive yields collected on the valine residues (from myoglobin) a t positions 1 and 10 were typically 95-96%.
RESULTS AND DISCUSSION
Nonrandom Utilization of Postsynthetic Acetylation Sites in Macronuclear H4: Acetate Labeling-Histones prepared from macronuclei of vegetative cells remesent a convenient source of histone extensively modified ' y postsynthetic acetylation (Vavra et al., 1982), while micronuclei prepared from young conjugating cells (pulse labeled with [3H]lysine) are a source of histone which is modified by deposition-related acetylation
The abbreviation used is: HPLC, high pressure liquid chromatog- raphy.
(Allis et al., 1985a). To address whether acetylation sites are utilized in a random or nonrandom fashion during postsyn- thetic transcription-related histone acetylation, we chose to focus our attention initially on macronuclear H4. H4 is ideally suited for this type of analysis for several reasons. As shown in Fig. 1 (lane a), macronuclear H4 is easily resolved by electrophoresis on long acid-urea gels into unmodified (H4") and mono-, di-, tri-, and tetraacetylated forms, and this region of the gel is not contaminated by other histone or nonhistone proteins. Acetylation (lanes b and c) is the only postsynthetic modification known to affect the mobility (charge) of H4 in this gel system (see Gorovsky et al., 1978); H4 is not phos- phorylated (Allis and Gorovsky, 1981). After labeling in vivo for 15 min with [3H]sodium acetate in the presence of cyclo- heximide, Gorovsky et al. (1973) have shown that the relative specific activity (cpm of [3H]acetate/unit stain) of mono-,
: F F
H41 I I
4
3
a b FIG. 1. Fractionation of macronuclear histones on a long
acid-urea gel: distribution of acetylated subspecies. Growing Tetrahymena (400,000 cells/ml) were pelleted, resuspended in Tris containing cycloheximide, and labeled for 30 min with [3H]sodium acetate (see Vavra et al., 1982 for details). Acid extracts from macro- nuclei of these cells were electrophoresed in a long acid-urea gel and analyzed by staining (lane a) and fluorography (lanes b and c). Shown are the acetylated subspecies of the major macronuclear histones as well as the location of two relatively minor histone variants, hvl and hv2, in this gel system (see Allis et al., 1980b). Note that hvl (and its acetylated subspecies) migrates one unit charge faster than H3 (and its acetylated forms), while hv2 migrates precisely in register with H3 (Allis et al., 1980b; Allis et al., 198513). Exposure time for lane b is 2 days; for lane c, 23 days.

Nonrandom Utilization of Histone Acetylation Sites 1073
di-, and tri- (plus tetra-) acetylated H4 subspecies is 1.0, 2.1, and 3.8, respectively. This suggests that labeling under these conditions approaches steady state (the distribution of acetate label following a 30-min labeling is shown in Fig. 1, lane b). Therefore, by monitoring [3H]acetate radioactivity at each cycle of automated sequencing, it is possible to determine which residues are being utilized as acetylation sites in a particular acetylated subspecies. Our ability to carry out an analysis such as this is made considerably easier in Tetrahy- mena because H4 in Tetrahymena, unlike most other H4s (Isenberg, 1979), is not blocked at its amino terminus. Tet- rahymena H4 has been sequenced by several groups (Glover and Gorovsky, 1979; Hayashi et al., 1984b) and four potential sites of acetylation have been identified (lysines at positions 4, 7, 11, and 15).
Fig. 2 shows the results obtained when the [3H]acetate counts released at each cycle of an automated sequencing run are determined for populations of mono-, di-, tri-, and tetra- acetylated H4 subspecies. The results clearly demonstrate that the lysine at position 7 in the H4 sequence is the site of acetylation used exclusively in populations of monoacetylated H4 (panel A ) . This result is particularly striking since it was a formal possibility that each of the four potential acetylation sites could have been used in the total population of mono- acetylated H4 molecules (with each individual H4 polypeptide having one of the 4 sites utilized, random model). Equally striking is the fact that significant amounts of acetate are detected at cycles 4 and 7 in populations of diacetylated H4 (panel B ) . Therefore, under these conditions, lysines at posi- tions 4 and 7 are used exclusively as acetylation sites in diacetylated macronuclear H4. These results also suggest that the lysine at position 7 represents the first site of acetylation
H4 mono-
12
-7 H4 di-
d 10 20
CYCLE/RESIDUE FIG. 2. Sites of acetylation utilized in populations of mono-,
di-, tri-, and tetraacetylated H4. Acetylated subspecies of macro- nuclear H4 (from cells labeled as in Fig. 1) were fractionated on a long acid-urea gel (see Fig. l ) , excised, eluted, and subjected to automated sequencing procedures. Shown is the distribution of [3H] acetate label released at each cycle from populations of mono- ( A ) , di- ( B ) , tri- ( C ) , and tetraacetylated (D) H4. Asterisks along the abscissa of each plot denote the four potential acetylation sites in the H4 molecule (lysines at positions 4, 7,11, and 15; Glover and Gorov- sky, 1979; Hayashi et aL, 1984b). Also shown in panels C and D (dashed line) is the expected acetate radioactivity calculated using a repetitive yield of 95.5% and assuming the acetate radioactivity at lysine 4 is 100%.
utilized in H4, with that in position 4 being used secondarily. The results obtained with tri- and tetraacetylated H4 are
less clear. If a third acetylation site is utilized exclusively when diacetylated H4 is converted to triacetylated H4, one might have expected acetate radioactivity to be more or less evenly distributed between the two sites used for diacetylation (4 and 7, panel B ) and the third site with essentially no radioactivity detected in the fourth site. Similarly, one would expect all 4 of the sites to be utilized in tetraacetylated H4. However, factors such as repetitive yield and cross-contamination be- tween neighboring bands (see Fig. l) undoubtedly affect the data presented in Fig. 2. Plotted in Fig. 2 (panels C and D, dashed lines) is the acetate radioactivity expected for lysines at positions 4, 7, 11, and 15 calculated from a repetitive yield of 95% and assuming equal efficiency of acetate labeling at these sites (position 4 = 100%). Both the tri- and tetra- acetylated forms of H4 show peaks of radioactivity at posi- tions 11 and 15 which fall below the calculated repetitive yield curve; this is presumably due to artificially high radioactivity in positions 4 and 7 resulting from cross-contamination of the tri and tetra forms with the more prevalent mono- and dia- cetylated species. Thus, we are unable to conclude whether there is exclusive or random usage of sites 11 and/or 15 in the triacetylated form of H4.
The data presented in Fig. 2 are also complicated by the kinetics of acetylation-deacetylation reactions (see Covault and Chalkley, 1980 for a thorough discussion of these prob- lems). The conditions of acetate labeling which we have utilized (30 min in the presence of cycloheximide) are long in terms of half-lives of the fast and slow acetylation reactions characterized by Chalkley and his associates (Covault and Chalkley, 1980), and Gorovsky et al. (1973) have shown that a 15-min acetate labeling in Tetrahymena approaches steady state with macronuclear H4. Nonetheless, it is possible that the third and fourth sites of acetylation utilized to generate tri- and tetraacetylated H4 subspecies represent positions which are metabolized more slowly in the H4 molecule. To test whether increased acetate label could be driven into these sites by blocking deacetylation, we have repeated the experi- ment shown in Fig. 2 except that the acetate labeling and all subsequent nucleus purifications were carried out in the pres- ence of 15 mM sodium butyrate (these conditions have been shown to block deacetylation in Tetrahymena macronuclei, Vavra et al., 1982). Essentially identical results were obtained to what is shown in Fig. 2 (panel c ) when the position and extent of [3H]acetate label was monitored at each cycle of triacetylated H4 from butyrate-treated cells (data not shown).
Nonrandom Utilization of Acetylation Sites in H4: Mass Ratio of Acetyllysine to Lysine-To circumvent the problems associated with acetate labeling, we have repeated the se- quencing with unlabeled material and examined the molar ratio of acetyllysine to lysine at each of the four potential H4 acetylation sites (4, 7, 11, and 15) in populations of mono-, di-, triacetylated H4 (fractionated on a long acid-urea gel). In the case of mono- and diacetylated H4, the acetyllysine to lysine ratio is approximately 2-3 times higher in those sites which have already been identified as the exclusive sites of acetylation in our acetate-labeling studies (7 in monoacety- lated H4; 4 and 7 in diacetylated H4; see Fig. 2). In the case of triacetylated H4, this ratio is again highest in residues 4 and 7; however, in this case the acetyllysine to lysine ratio determined for residue 11 is twice that determined for residue 15 (data not shown). This 2-fold apparent preference for site 11 over 15 as the third site of acetylation in triacetylated H4 is probably a minimal estimate since the amount of acetylly- sine observed at residue 15 is likely to be inflated due to some

1074 Nonrandom Utilization of Histone Acetylation Sites
contamination from the previous residue during sequencing. An alanine exists in the H4 sequence at position 14 (Glover and Gorovsky, 1979; Hayashi et al., 1984b), and alanine and acetyllysine co-migrate under our HPLC column conditions.2 Nonetheless, these data again suggest that there is some preference for using the lysine at position 11 as the third site of acetylation in H4.
Postsynthetic Acetylation Site Preference in H3"The ob- servation that acetylation site utilization is nonrandom for macronuclear H4 has prompted us to ask whether this is the case for any other macronuclear histones. While each of the other macronuclear core histones (H2A, H2B, and H3) are also modified postsynthetically by acetylation (see Fig. 1 and Vavra et al., 1982 and Gorovsky, 1985), we have chosen not to analyze acetylation site utilization in some of these histones for the following reasons. H2B exists in only a monoacetylated form (Gorovsky et al., 1978), and its apparently unique site of acetylation has already been determined (lysine 3; Glover, 1979; Nomoto et al., 1982). H2A consists of two major electro- phoretic forms, S and F (Gorovsky, 1985), both of which are modified by acetylation and phosphorylation (Allis and Go- rovsky, 1981; Vavra et al., 1982; Fusauchi and Iwai, 1984). Thus, it is not possible to use mobility on acid-urea gels alone as an indicator of acetylation states (mono- versus diacetyla- tion, for example). H2A is also blocked a t its amino terminus (Fusauchi and Iwai, 1983,1984).
Macronuclear H3, on the other hand, is another histone well suited for analyses similar to what have been described with H4. It is extensively modified by acetylation (see Fig. 1) and is not known to be phosphorylated to a significant degree (Allis and Gorovsky, 1981). Like H4, H3 has an unblocked amino terminus (Glover, 1979). However, such analyses are made more complicated because the H3 region of a long-acid- urea gel is contaminated by histone variants hvl and hv2 (see Allis et al., 1980b and Fig. 1). Acetylation sites in hvl have been determined (lysines a t positions 4, 7, 10, 13, 16, and 21; Allis et al., 1985b) and fortunately, none of these overlap with those in H3 (9, 14, 18, and 23; Glover, 1979; Hayashi et al., 1984a). In contrast, the putative sites of acetylation in hv2 have been identified (Allis et al., 1985b), and these are essen- tially identical to those found in H3 (hv2 is an H3 variant; Allis et al., 1980b). Thus, while it is possible to distinguish hvl and H3 acetylation sites in the same sequencing run (because there is no overlap), we are not able to positively distinguish hv2 and H3 sites in samples recovered from an acid-urea gel.
With these reservations in mind, Fig. 3 presents the data we obtained by monitoring [3H]acetate radioactivity (again from cells labeled as in Fig. 1) a t each cycle during the sequencing of populations of mono- ( A ) , di- (B), and tria- cetylated (C) H3. When the monoacetylated H3 region of the gel is analyzed in this fashion (Fig. 3, panel A ) , we observe significant acetate counts released at cycles 9 and 14, two known H3 (or hv2) acetylation sites (Glover, 1979; Hayashi et al., 1984a; Allis et al., 1985b). This result could be explained in two ways. First, two distinct populations of monoacetylated H3 could exist, one acetylated exclusively at lysine 9 and one acetylated exclusively at lysine 14. Alternatively, one peak could result from monoacetylated H3 and the other from monoacetylated hv2 with each being acetylated at different positions (9 and 14). Both of these acetylation sites are retained in the histone migrating at the position of diacety- lated H3 (panel B). If two distinct populations of monoacet- ylated H3 exist, each population must utilize the alternative
R. G. Cook and C . D. Allis, unpublished observations.
site (9 or 14) not utilized in the monoacetylated form. Similar arguments can be made if distinct populations of monoacet- ylated H3 and hv2 exist. Finally, our data demonstrate that there is preference for using lysine 18 as the third site of acetylation in triacetylated H3 and/or hv2 (panel C). Even though we are unable to measure the extent with which hv2 contamination affects the data presented in Fig. 3, it is clear that acetylation sites in H3 are also utilized in a nonrandom fashion.
Sites of Acetylation in Newly Synthesized Micronuclear and Macronuclear H4"Data presented earlier (Fig. 2) show clearly that during macronuclear-specific acetylation of H4, lysines at positions 4 and 7 are used exclusively as acetylation sites in the population of diacetylated H4. The recent finding that newly synthesized micronuclear H4 is deposited in a diacetylated form as well (Allis et al., 1985a) tempted us to ask whether the same two acetylation sites are also used in deposition-related acetylation. To investigate this question and circumvent problems associated with macronuclear con- tamination in the micronuclear preparation, we have ex- ploited the high specificity of [3H]lysine labeling for micro- nuclear H4 in young conjugating cells (Allis et al., 1985a). Newly synthesized micronuclear histone (labeled for 30 min with [3H]lysine from 5.0 to 5.5 h of conjugation) was fraction- ated on a long acid-urea gel after which the diacetylated H4 subspecies was excised, eluted, recovered, and subjected to automated sequencing procedures. Material recovered from the sequencing run corresponding to cycles (residue) 4, 7, 11, and 15 (and as a control 1 and 9) was then subjected to HPLC using conditions where the phenylthiohydantoin derivatives of acetyllysine and unmodified lysine are well separated. Gradient fractions from each column run (residues 4, 7, 11, and 15) were then counted to determine if the [3H]lysine label representing newly synthesized micronuclear H4 elutes in the position of acetyllysine or unmodified lysine at these four positions. The results (Fig. 4A) demonstrate that the vast majority of the [3H]lysine counts at positions 4 and 11 elute from the HPLC column in the position of acetyllysine (large arrow) while that from residues 7 and 15 elutes as unmodified lysine. Essentially no counts above background were observed in any fraction of the run performed with residues 1 and 9 (this is expected since residues 1 and 9 are alanine and methionine, respectively, in macronuclear H4 (Glover and Gorovsky, 1979 and Hayashi et al., 1984b)). Furthermore, no counts above background were observed in any position of the gradients not shown in Fig. 4A. From these results we con- clude that the lysines a t positions 4 and 11 are the exclusive sites of acetylation in newly synthesized micronuclear H4.
The finding (Fig. 4A) that deposition-related micronuclear H4 acetylation affects lysines at positions 4 and 11, while 4 and 7 are the exclusive sites of postsynthetic acetylation in steady-state populations of diacetylated H4 isolated from macronuclei caused us to ask whether diacetylation of H4 at positions 4 and 11 correlates better with histone deposition per se (as opposed to what may be transcription-reluted di- acetylation at 4 and 7) or is a phenomenon unique to micro- nuclear H4 as opposed to macronuclear H4. To test these relationships further, we repeated the experiment shown in Fig. 4A except that diacetylated H4 was isolated from macro- nuclei of pulse-labeled ([3H]lysine for 5 min) growing cells. The results obtained (Fig. 4B) are essentially identical to those shown in Fig. 4A. The lysine label at positions 4 and 11 elutes at the position of acetyllysine, whiIe that at 7 and 15 elutes as unmodified lysine. Thus, we conclude that diacety- lation of H4 at lysines 4 and 11 is not a micronuclear-specific phenomenon but is instead a deposition-related phenomenon.

Nonrandom Utilization of Histone Acetylation Sites I I I I I I
C H3 tri hvl tr i hvl tetra
- - 4.5-
10 20 10 20 10 20 CYCLE/RESIDUE
FIG. 3. Sites of acetylation utilized in acetylated subspecies of H3 (and hv l and hv2). Acetylated subspecies of macronuclear H3 were prepared and analyzed as described in the legend to Fig. 2. Shown is the distribution of [3H]acetate released a t each cycle from populations of mono- (A) , di- ( B ) , and triacetylated ( C ) H3. Asterisks along the abscissa of each plot denote the four potential acetylation sites which have been identified in H3 (lysines a t positions 9, 14, 18, and 23; Glover, 1979; Hayashi et al., 1984a). Downward-pointing arrows denote acetate counts which are likely to arise from sequencing hvl which co-migrates with H3 almost precisely in this gel system (see Fig. 1). The acetylation sites in hvl have recently been identified (lysines at positions 4, 7, 10, 13, 16, and 21; Allis et al., 1985b).
SUMMARY AND CONCLUSIONS
Exploitation of the biology of Tetrahymena (macro- uersus micronuclei) has allowed us to study sites of histone acetyla- tion under conditions where postsynthetic transcription-re- lated and synthetic deposition-related acetylation are nono- verlapping. The theme of two distinct functions for histone acetylation (deposition and transcription) is strengthened by our finding of distinct combinations of acetylation sites being used during these two processes. To analyze acetylation site utilization under various physiological conditions, we have exploited the fact that H4 and H3 are not blocked at their amino termini in Tetrahymena as is the case in many other organisms. With regard to macronuclear-specific postsyn- thetic acetylation, we find that utilization of H4 and H3 acetylation sites is surprisingly nonrandom. In certain in- stances, exclusive (or highly preferred) sites of acetylation have been identified a t each acetylated level (mono-, di-, etc.), suggesting that there may be an ordered sequence to the sites utilized during acetylation. As far as we are aware, this is the first report demonstrating such regularity in this aspect of histone metabolism.
While our data support a nonrandom process in the utili- zation of NH2-terminal acetylation sites, the molecular basis for this regularity is far from clear. Close inspection of the protein sequence surrounding these sites (see Gorovsky, 1985) has revealed few clues as to why certain sites are more preferred than others. (We note that more glycine residues surround sites 4 and 7 than 11 and 15 in the H4 sequence.) In a very general sense it seems that sites lying closer to amino termini are utilized before (and more often than) those lying closer to carboxyl temini. I t is, of course, possible that acetylation at the first position causes an allosteric change in the conformation of the protein rendering second, third, etc., sites more accessible. A more important issue is whether histones carrying different acetylation states have different functions. Since it is generally true that a higher percentage of histone is acetylated than the fraction of the genome transcribed into heterogeneous nuclear RNA (Covault and Chalkley, 1980), it is possible that only the highest acetylation states are functional in producing transcriptionally competent chromatin.
Of the four potential sites of acetylation that have been identified in the NH,-terminal arm of macronuclear H4 (ly-
A Ac-Lys
I
15 20
B
‘1. A c - L ~ s
7
15 20
1075
FRACTION NO
FIG. 4. Sites of acetylation on newly synthesized micro- and macronuclear deacetylated H4. Acid extracts from micro- ( A ) and macronuclei ( B ) were electrophoresed in a long acid-urea gel after which protein migrating at the position of diacetylated H4 was excised, eluted, and processed for automated protein sequencing (see “Materials and Methods”). Micronuclei were isolated from 5-h mating cells labeled with [3H]ly~ine while macronuclei were prepared from cells pulsed for only 5 min with [3H]lysine to avoid transcription- related acetylation reactions (see text for details). Following protein sequencing, material corresponding to residues 4, 7, 11, and 15 (and 1 and 9 as negative control) was subjected to HPLC fractionation under conditions where acetyllysine is well separated from unmodified lysine (see “Materials and Methods” for details). Column fractions were dried and subjected to liquid scintillation counting. Acetyllysine elutes near fraction 14; unmodified lysine elutes near fraction 21. Only this region of the gradient is shown; no counts above background (20-30 cpm) were observed elsewhere.

1076 Nonrandom Utilization of Histone Acetylation Sites
sines at positions 4, 7, 11, and 15; see Glover and Gorovsky, 1979; Hayashi et al., 1984a), we have determined that two of these sites (4 and 11) are used exclusively in newly synthesized diacetylated H4 destined for either micronuclei of mating cells or macronuclei of pulse-labeled growing cells. In contrast, lysines at positions 4 and 7 were found to be the exclusive sites of acetylation in poulations of diacetylated molecules isolated from macronuclei of growing cells. Thus, it seems that positions 7 and 11 represent preferred sites of acetylation for transcription- and deposition-related function(s), respec- tively. (We point out that a small proportion of total macro- nuclear H4 exists as tri- and tetraacetylated H4, and, there- fore, acetylation at sites 11 and 15 may serve as a transcrip- tion-related function as well as 4 and 7.) As far as we are aware, this is the first suggestion of functionally distinct acetylation sites in histone H4. While it seems likely that the specificity for deposition- and transcription-related acetyla- tion H4 sites may reside in the activities of at least two distinct H4 acetylases used in Tetrahymena, this question will be resolved only when appropriate enzymes have been purified and characterized.
Elegant genetic studies carried out in yeast by Grunstein and his associates (Wallis et al., 1983) have shown that H2B can function in uiuo even with large deletions at its amino terminus (including deletions which remove H2B acetylation sites). However, several points suggest that acetylation of H4 (and possibly H3) is functionally significant. First, a high degree of conservation exists between the sites of acetylation in H3 and H4 between Tetrahymena and several vertebrates (see Isenberg, 1979 and Gorovsky, 1985). This finding argues strongly that acetylation of these arginine-rich histones serves an important function(s) that has been rigidly maintained throughout evolution. Second, we have demonstrated in this study that specific (and different) acetylation sites are utilized for what is likely to be transcription- and deposition-related H4 diacetylation. Finally, we point out that mutagenesis studies have yet to be carried out (to our knowledge) with H3 and H4. Since these histones are known to play a fundamental role in nucleosome structure and assembly (Camerini-Otero et al., 1976; Bina-Stein and Simpson, 1977) and are specifi- cally the histones which display deposition-related acetyla- tion, it will be important to carry our functional tests with site-specific and deletion mutants of these histones.
Acknowledgments-The original idea to look for nonrandom usage of acetylation sites in macronuclear histones evolved from discussions
when C. D. A. was a postdoctoral fellow in the laboratory of Dr. Martin Gorovsky (University of Rochester, Rochester, NY). We thank Drs. M. Gorovsky and C. Glover for their comments on the manuscript.
REFERENCES Allis, C. D., and Dennison, D. K. (1982) Deu. Bwl. 9 3 , 519-533 Allis, C. D., and Gorovsky, M. A. (1981) Biochemistry 20,3828-3833 Allis, C. D., Glover, C. V. C., and Gorovsky, M. A. (1979) Proc. Natl.
Allis, C. D., Bowen, J. K., Abraham, G. N., Glover, C. V. C., and Acad. Sci. U. S. A. 76,4857-4861
Gorovskv. M. A. (1980a) Cell 20.55-64 Allis, C. D.; Glover, C. V. C., Bowen, J. K., and Gorovsky, M. A.
Allis. C. D.. Chicoine. L. G.. Richman. R.. and Schulman. I. G. (1985a) (1980b) Cell 20,609-617
Proc. Natl. Acad. Sci. U.'S. A. 82,'8048-8052 Allis, C. D., Richman, R., Gorovsky, M. A., Ziegler, Y. S., Touchstone,
B., Bradley, W. A., and Cook, R. G. (1985b) J. Biol. Chem., in press Bina-Stein, M., and Simpson, R. T . (1977) Cell 11, 609-618 Bruns, P. J., and Brussard, T. B. (1974) J. Exp. 2001. 188,337-344 Camerini-Otero, R. D., Sollner-Webb, B., and Felsenfeld, G. (1976)
Candido, E. P. M., and Dixon, G. H. (1971) J. Biol. Chem. 246,3182-
Covault, J., and Chalkley, R. (1980) J. Biol. Chem. 255,9110-9116 Fusauchi, Y., and Iwai, K. (1983) J. Biochem. (Tokyo) 93,1487-1497 Fusauchi, Y., and Iwai, K. (1984) J. Biochem. (Tokyo) 95,147-154 Glover, C. V. C. (1979) Ph.D. thesis, University of Rochester Glover, C. V. C., and Gorovsky, M. A. (1979) Proc. Natl. Acad. Sci.
Gorovsky, M. A. (1985) in Molecular Biology of Ciliated Protozoans
Cell 8,333-347
3188
U. S. A. 76 , 585-589
(Gall. J. G.. ed) Academic Press. New York. in Dress Gorovsky, M: A.; Pleger, G. L., Keevert, 3. B. , aAd Johmann, C. A.
(1973) J. Cell Biol. 57, 773-781 Gorovsky, M. A., Yao, M.-C., Keevert, J. B., and Pleger, G. L. (1975)
in Methods Cell Biol. 9 , 311-327 Gorovsky, M. A., Glover, C., Johmann, C. A., Keevert, J. B., Mathis,
D. J., and Samuelson, M. (1978) Cold Spring Harbor Symp. Quunt. Biol. 42,493-503
Hayashi, T., Hayashi, H., Fusauchi, Y., and Iwai, K. (1984a) J. Biochem. (Tokyo) 95,1741-1749
Hayashi, H., Nomoto, M., and Iwai, K. (1984b) J. Biochem. (Tokyo)
Hunkapiller, M., Lujan, E., Ostrander, F., and Hood, L. E. (1983a)
Hunkapiller, M. W., Hewick, R. M., Dreyer, W. J., and Hood, L. E.
Isenberg. I. (1979) Annu. Reu. Biochem. 48. 159-191
96,1449-1456
Methods Enzymol. 91.227-236
(1983b) Methods Enzymol. 9 1 , 399-413
Nomoto, M., Hayashi, H., and Iwai, K. (1982) J. Biockem. (Tokyo)
Ruiz-Carrilo, A., Wangh, L. J., and Allfrey, V. G. (1975) Science 190,
_. . .
9 1,897-904
117-128 Vavra, K. J., Allis, C. D., and Gorovsky, M. A. (1982) J. Biol. Chem.
Wallis, J. W., Rykowski, M., and Grunstein, M. (1983) Cell 35 , 711-
~~ ~~~
257,2591-2598
719





























