Embed Size (px)
Citation preview

INFECTION AND IMMUNITY, Sept. 2010, p. 3650–3659 Vol. 78, No. 90019-9567/10/$12.00 doi:10.1128/IAI.00480-10Copyright © 2010, American Society for Microbiology. All Rights Reserved.
Development and Validation of an In Vivo Candida albicansBiofilm Denture Model�
Jeniel E. Nett,1,2 Karen Marchillo,1 Carol A. Spiegel,3 and David R. Andes1,2*Departments of Medicine,1 Medical Microbiology and Immunology,2 and Pathology and Laboratory Medicine,3
University of Wisconsin, Madison, Wisconsin
Received 7 May 2010/Returned for modification 16 June 2010/Accepted 25 June 2010
The most common form of oral candidiasis, denture-associated stomatitis, involves biofilm growth on an oralprosthetic surface. Cells in this unique environment are equipped to withstand host defenses and surviveantifungal therapy. Studies of the biofilm process on dentures have primarily been limited to in vitro models.We developed a rodent acrylic denture model and characterized the Candida albicans and mixed oral bacterialflora biofilm formation, architecture, and drug resistance in vivo, using time course quantitative cultureexperiments, confocal microscopy, scanning electron microscopy, and antifungal susceptibility assays. We alsoexamined the utility of the model for measurement of C. albicans gene expression and tested the impact of aspecific gene product (Bcr1p) on biofilm formation. Finally, we assessed the mucosal host response to thedenture biofilm and found the mucosal histopathology to be consistent with that of acute human denturestomatitis, demonstrating fungal invasion and neutrophil infiltration. This current oral denture model mimicshuman denture stomatitis and should be useful for testing the impact of gene disruption on biofilm formation,studying the impact of anti-infectives, examining the biology of mixed Candida-oral bacterial flora biofilminfections, and characterizing the host immunologic response to this disease process.
Candida spp. frequently establish biofilm lifestyles, prolifer-ating as multicellular communities attached to surfaces, suchas a medical device (21, 31). Adherent and encased in a poly-meric extracellular matrix, biofilm cells are phenotypically dis-tinct from free-floating planktonic cells. Not only are theseorganisms capable of proliferating in healthy hosts by survivingimmune factors, they demonstrate increased resistance to com-monly used antifungal drug therapies (1, 35, 39, 55). The ma-jority of Candida biofilm infections are associated with place-ment of medical devices, such as a dentures, venous catheters,or urinary catheters (29). Treatment of these infections re-mains difficult due to the extraordinary drug resistance exhib-ited during biofilm growth. Although often costly and incon-venient, device removal is typically required for eradication ofinfection (48).
Oral candidiasis commonly occurs in those who are colo-nized with Candida and have risk factors for development ofdisease, such as HIV infection, chemotherapy treatment, orantibiotic exposure (63, 67). Local breaks in mucosa caused bymucositis, radiation, and trauma are also predisposing factors.Oral candidiasis may occur as an acute or chronic illness andcan involve the tongue, buccal mucosa, or palate. The mostcommon form of oral Candida infection, denture stomatitis, isprevalent in the elderly population and affects up to 70% ofdenture wearers (58, 66, 70). The condition begins with Can-dida biofilm growth on the denture-mucosa interface. Biofilmgrowth progresses over the denture surface, leading to inflam-mation of the denture-exposed palatal mucosa (54, 67). Clin-ical manifestations range from no symptoms to severe pain and
difficulty swallowing (70). While Candida albicans is responsi-ble for the majority of oral candidiasis (70 to 80%), infectionby Candida glabrata, Candida tropicalis, and Candida parapsi-losis has been reported less frequently (20, 63).
In vitro biofilm models have the advantage of lower cost andare ideal for high-throughput screening assays and antifungaltesting (12, 56, 61, 71). These models have provided importantphenotypic information regarding drug resistance in Candidabiofilms. The models have also been valuable for characteriz-ing the role of surface adherence properties, such as topogra-phy and hydrophobicity. However, it is difficult for in vitromodels to account for all the factors which likely play a roleduring biofilm infection (38). For example, cells in the in vitrosystems are not exposed to salivary immune components andproteins which may precondition, or coat, the surface andpromote adherence. Although several of the host componentswhich play a key role in promoting Candida adherence havebeen defined, the interaction is likely a complex phenomenon(42). In the denture biofilm niche, additional components hy-pothesized to impact this growth state include companion oralbacterial flora, contact between the device biofilm and the hostmucosal surface, and salivary flow dynamics. Each of theseconditions would be challenging to mimic in vitro.
Several in vivo models have been developed for study of oralcandidiasis and denture stomatitis (58). The earliest studiesutilizing acrylic plates fitted to the palates of primates foundthat Candida infection of these prostheses mimicked the his-topathology of human denture stomatitis (8, 9). However, thecost and difficulty of maintaining primates prevented wide-spread use. An alternative model utilized rats with moldedacrylic plates (47). Advantages of the rat model include re-duced size and cost. The rat mucosal surface is also relativelyeasy to colonize with Candida, although maintenance of colo-nization often requires antibiotic administration (58). The rat
* Corresponding author. Mailing address: 600 Highland Ave., RoomH4.572, Madison, WI 53792. Phone: (608) 263-1545. Fax: (608) 263-4464. E-mail: [email protected].
� Published ahead of print on 6 July 2010.
3650

models have been integral for describing the basic pathologiesof oral candidiasis and denture stomatitis. The investigationshave primarily been directed toward the host response, andin-depth examination of the Candida biofilm phenotype hasnot been undertaken.
Here we adapt and test a rat model for Candida-associateddenture stomatitis, with a focus on Candida biofilm formation.We chose to use a common acrylic denture material to form anoral device directly in the animal. One major advantage of thecurrent device placement is the ability to form the denturewithout the costly and time-consuming ex vivo mold process.The model mimics denture stomatitis with respect to Candidabiofilm formation and histology. The experimental systemproved useful for testing of the impact of gene products andmeasurement of the response to treatment. It also demon-strated the value for study of mixed microbial biofilm interac-tions, comparisons with other models of biofilm niche infec-tion, such as a venous catheter model, and examination of thehost response to biofilm.
MATERIALS AND METHODS
Animals and denture maintenance. Specific-pathogen-free male Sprague-Dawley rats at weights of 350 g (Harlan Sprague-Dawley, Indianapolis, IN) wereused for all studies. Animals were maintained in accordance with the AmericanAssociation for Accreditation of Laboratory Care criteria, and all studies wereapproved by the Institutional animal Care Committee. Rats were housed indi-vidually in an environmentally controlled room in metabolic cages to avoid oralcontamination with bedding and animal waste. Animals were fed an Ensureliquid diet while dentures were in place to avoid contamination of biofilms withfood particles. The animals were examined for signs of distress at 6 h aftersurgery and then every 12 h throughout the study. The oral mucosal area wasexamined twice daily for signs of inflammation or purulence.
Prior to placement of denture appliances, rats were immunosuppressed with asingle dose of cortisone (200 mg/kg subcutaneously) on the day of infection tomimic a protocol commonly utilized for rodent oral candidiasis (49). For a subsetof experiments, either ampicillin sodium-sulbactam sodium at 100 mg intraperi-toneally (i.p.) twice a day (BID) or aztreonam at 50 mg/kg subcutaneously BIDwas administered 48 h prior to denture placement to reduce enteric bacterialcolonization common to rodents due to coprophagy, or the consumption of feces.Dosing regimens were chosen based upon those previously shown to be effectivein the treatment of rodent systemic bacterial infections (D. R. Andes, unpub-lished data).
Initial studies of the metabolic cages demonstrated that the rats continued topractice coprophagy. To address this, we modified a tube caging procedure thathas been utilized in nutritional absorption studies (15). Specifically, an additionalgroup of animals were housed in polyvinyl chloride (PVC) plastic tube caging inwhich the ability to turn and reach fecal material was eliminated. Animal mon-itoring during this additional housing restraint was increased to every 6 hthroughout the study.
Denture placement. A 32-gauge stainless steel Babcock orthodontic wire (Mil-tex) was threaded across the hard palate and secured between cheek teeth of ananesthetized rat to serve as an anchor for the denture material (Fig. 1). Cheekteeth were etched using Uni-Etch 32% semigel etchant, with benzalkoniumchloride (Bisco, Inc., Schaumburg, IL) for microretention. A metal spatula wasplaced over the hard palate to create an approximately 3- by 5-mm area acrossthe palate and less than 1 mm of space between the hard palate and the denturematerial. This space was used for Candida inoculation. Cold-cure acrylic tem-porary crown and bridge material (Maxitemp HP; Henry Schein, Melville, NY)was applied over the cheek teeth and wire to create a surface approximately 8 by10 mm in size and approximately 2-mm thick. Material was rapidly shaped witha small spatula to produce a rectangular denture, which was allowed to solidifyfor 5 min, after which the spatula was removed.
Organisms and inoculum. Candida albicans strains DAY185 and CJN702(bcr1�/bcr1�) were used for all studies (17, 43, 44). The strains were stored in15% (vol/vol) glycerol stock at �80°C and maintained on yeast extract-peptone-dextrose (YPD) medium plus uridine (1% yeast extract, 2% peptone, 2% dex-trose, and 80 �g/ml uridine) prior to experiments. Prior to denture inoculation,cells were grown at 30°C in YPD plus uridine liquid media with orbital shakingat 200 RPM overnight. To prepare the inoculum, cells were washed with phos-phate-buffered saline (PBS), enumerated by hemocytometer counting, and re-suspended in saline at 108 cells/ml. The final inoculum concentration was con-firmed by microbiologic enumeration.
Infection of denture. After removal of the spatula, the hard palate beneath theacrylic device was inoculated with 108 C. albicans cells/ml (0.1 ml). Animals weresacrificed after 6, 24, 48, or 72 h for histopathology or device removal and study.For treatment studies, antifungals were applied to mature biofilms after 48 h ofbiofilm growth.
Fungal and bacterial qualitative and quantitative cultures from dentures. Todetermine the viable burden of Candida and the accompanying oral flora, mi-crobiologic counting was performed on dentures. Devices were placed in 2 ml0.15 M NaCl, sonicated for 10 min, and vortexed. For Candida microbiology, 1:10dilutions were plated on Sabouraud dextrose agar (SAB) plus 2.5 �g/ml chlor-amphenicol. For determination of aerobic bacteria, dilutions were plated onMueller-Hinton agar plus 2.5 �g/ml amphotericin B. For anaerobic cultures,specimens were passed into an anaerobic chamber (80% N2, 10% H2, and 10%CO2; Coy Laboratory Products, Grass Lake, MI), where dilutions were made inprereduced anaerobically sterilized PY (peptone yeast extract; Anaerobe Sys-tems, Morgan Hill, CA) broth and placed on reduced plates of LKV (laked bloodwith kanamycin and vancomycin), PEA (phenylethyl alcohol agar), and EYA(egg yolk agar) that were incubated in the chamber at 35°C for 5 days (mediafrom Remel, Lenexa, KS). Dilutions were passed out of the chamber, plated, andincubated in 5 to 7% CO2 for 1 day. Organisms were identified by Gram stainmorphology and rapid testing, e.g., catalase, coagulase, PYR (pyrrolidonyl ami-nopeptidase), and streptococcal grouping (13). Experiments were performed ontwo or three separate occasions.
SEM. Acrylic devices were processed for scanning electron microscopy (SEM)as previously described for venous catheters (3). After removal of each denture,it was washed with phosphate-buffered saline (PBS) and placed in fixative (1%[vol/vol] glutaraldehyde and 4% [vol/vol] formaldehyde in PBS) overnight. Thesamples were rinsed with PBS, placed in 1% osmium tetroxide for 30 min, andrinsed with PBS. The samples were subsequently dehydrated in a series ofethanol washes (30% for 10 min, 50% for 10 min, 70% for 10 min, 95% for 10min, and 100% for 10 min). Final desiccation was accomplished by critical-point
FIG. 1. Placement of rat denture. Stainless steel wire was threaded between cheek teeth. Acrylic denture material was applied over palate andaround cheek teeth (8 by 10 mm). The spatula left a space (3 by 5 mm) for Candida inoculation.
VOL. 78, 2010 C. ALBICANS BIOFILM DENTURE MODEL 3651

drying (Tousimis, Rockville, MD). Specimens were mounted on aluminum stubsand sputter coated with gold. Dentures were imaged on a Jeol 6100 at 10 kV. Theimages were processed for display using Adobe Photoshop.
Confocal microscopy. For confocal microscopy, denture biofilms were re-moved, gently rinsed with PBS, and visualized using one of two stain sets. Forimaging of live fungal elements, biofilm architecture, and the extracellular ma-trix, dentures were stained with the FUN-1 (50 �M) component of the Live/Deadyeast viability kit (Molecular Probes, Eugene, OR) and concanavalin A (ConA)conjugate (Alexa Fluor 488 conjugate, 200 mM), according to the manufacturer’sinstructions. Briefly, the device was incubated with each dye in PBS with 2%glucose for 30 min at 30°C. Both live and dead yeast cells were stained with theFUN-1 dye and visualized with a diffusely distributed yellow-green fluorescence.Metabolically active cells process this dye, which results in a shift to red fluo-rescence within intravacuolar structures. ConA binds to glucose and mannoseresidues of cell wall polysaccharides and is visualized as green fluorescence.Colocalization of fluorescent dyes (FUN-1 and ConA) in metabolically activecells results in a yellow to orange fluorescence during multichannel image cap-ture. The biofilms were imaged on an Axiovert 200 with excitation and emissionfilters at 490/617 nm and 490/528 nm for FUN-1 and ConA, respectively, aspreviously described for venous catheters (3).
The second dye set was selected for simultaneous imaging of yeast, hyphae,and bacteria within the mixed-species denture biofilm. This set included SYTO9 dye (Molecular Probes, Eugene, OR) to visualize bacteria in the mixed bio-films, as previously described (23). Calcofluor white was selected for its ability tobind to Candida hyphal cell walls, and FUN-1 was chosen for imaging of yeast.Dentures were stained with calcofluor white (22.5 �g/ml), SYTO 9 (5 �M), andFUN-1 (50 �M) in PBS with 2% glucose for 30 min at 30°C and imaged onAxiovert 200 with excitation and emission filters at 360/457 nm, 490/528, and490/617 nm, respectively. Images were obtained at 63� or 100� magnificationand compiled using Adobe Photoshop.
Histopathology. To discern if the model mimicked denture stomatitis, weperformed mucosal biopsy and histopathology at the end of the study period (48h). After animals were sacrificed and the dentures were removed, the tonguesand palates were dissected. The tissues were fixed in 10% buffered formalin andembedded in paraffin (26). Sections were stained with either hematoxylin andeosin (H&E) or Gomori’s methenamine silver (GMS) for C. albicans. Imageswere obtained at 200� magnification.
Drug resistance assays. Previous studies have demonstrated biofilm recalci-trance to antifungal therapy. We included both topical and systemic antifungaltreatment with a triazole, fluconazole, and an echinocandin, micafungin, todetermine the susceptibility of the denture biofilm to these therapies. For topicaltherapy, regimens included a 0.15 M NaCl control, topical administration of
micafungin at 5 mg/kg daily, or topical administration of fluconazole at 16 mg/kgtwice daily. Therapy was applied directly to mature (48-h-old) biofilms. Systemicdrug treatments included micafungin at 5 mg/kg intraperitoneally once daily orfluconazole at 16 mg/kg subcutaneously twice daily. The dosing regimens werechosen based on maximal efficacy in treatment of oral candidiasis in animalmodels (27, 51). All therapies were continued for 48 h, after which devices wereremoved and processed for viable burden determination as described above.
Ex vivo antifungal efficacy was also assessed on biofilm cells that were detachedfrom the biofilm and resuspended, as described previously for biofilm cells froma central venous catheter (3). Devices were removed at 48 h and placed in PBS.Organisms were dislodged by scraping the surface with a plastic loop, vortexing,and water bath sonicating as described above. Following biofilm removal fromthe biofilm, cells were resuspended for antifungal susceptibility testing to am-photericin B, micafungin, and fluconazole using NCCLS methods (37). Compar-isons were made to susceptibility testing with planktonic cells using the standardNCCLS protocol and identical inocula (103 CFU/ml).
RNA collection and quantitative reverse transcription-PCR (RT-PCR). Bio-film cells were dislodged from dentures by scraping, vortexing, and sonicating.Cells were flash frozen with liquid nitrogen in AE buffer (50 mM sodium acetateat pH 5.2, 10 mM EDTA), and RNA was collected using the hot method aspreviously described (3, 40). The RNA integrity was assessed using an AgilentBioanalyzer 2100 with an RNA Nano chip before use.
Quantitative real-time reverse transcription-PCR (RT-PCR) was used toquantify the mRNA abundances of a subset of transcripts (CDR1, CDR2,ERG25, ALS1, SAP5, GDB1, and BGL2). Transcripts for study were chosenbased on differential expression in the central venous catheter biofilm model andincluded those involved in drug resistance, biofilm adhesion, and glucan modi-fication (40, 45). The TaqMan probe and primer sets were designed using PrimerExpress (Applied Biosystems, Foster City, CA) (Table 1). The QuantiTect probeRT-PCR kit (Qiagen, Valencia, CA) was used in an ABI Prism 7700 version 1.7sequence detection system (Applied Biosystems) as previously described (3, 40).Reactions were performed in triplicate and analyzed using the threshold cycle(2���CT) method (34). The comparative expression method generated data astranscript fold changes normalized to a constitutive reference gene transcript(ACT1) and relative to log-phase planktonic cells grown in RPMI at 37°C on anorbital shaker set to 200 RPM.
RESULTS
Denture placement and animal well-being. Following oraldevice placement, the animals did not appear ill throughout
TABLE 1. C. albicans primer combinations used in quantitative real-time RT-PCR
Primer Sequence (5�–3�)a
ACT1 RT For .............................................................................................................AGCTTTGTTCAGACCAGCTGATTACT1 RT Rev.............................................................................................................GGAGTTGAAAGTGGTTTGGTCAAACT1 RT Probe .........................................................................................................FAM-CCAGCAGCTTCCAAACCT-TAMSpALS1 RT For..............................................................................................................TTAGCATCTGCAAGTGAAGAAGACAALS1 RT Rev .............................................................................................................ATGGAGCTTCTGTAGGACTGGTTGALS1 RT Probe..........................................................................................................FAM-AGCGGTTCTCATGAATCAGCATCCACAA-TAMSpBGL2 RT For .............................................................................................................CTCGCAACTGTTCTTACTTCAGTTGBGL2 RT Rev.............................................................................................................TGACGTCTTTACAAGTACCGTCATCBGL2 RT Probe .........................................................................................................FAM-TGAAAGCCAAATCACCCATGGCG-TAMSpCDR1 RT For.............................................................................................................CCATTAACCATCAGCACTTACDR1 RT Rev............................................................................................................CCGTTCTACCACCTTTTTGCACDR1 RT Probe.........................................................................................................FAM-AGTCTATCAAACTCAGCCA-TAMSpCDR2 RT For.............................................................................................................CCACATGTCCGACATACCTGGCDR2 RT Rev............................................................................................................GGAATCTGGGTCTAATTGTTCATGACDR2 RT Probe.........................................................................................................FAM-CCATTCAACGGCAACAT-TAMSpERG25 RT For...........................................................................................................TGGATTGGCAGCAGAATATGCERG25 RT Rev ..........................................................................................................GCAAGTTACCAGTGATAAGACACCAERG25 RT Probe.......................................................................................................FAM-CAATCGGAATACCAACCGTACCCAATCCTA-TAMSpGDB1 RT For.............................................................................................................TCTCACATCGGTGCAAATGGGDB1 RT Rev............................................................................................................CTCTACCCCAGCATCTCATATAATCAGDB1 RT Probe.........................................................................................................FAM-TGGCCGCTGGGTTACCTCATTTCA-TAMSpSAP5 RT For ..............................................................................................................CCAGATGATCCAACTGTTGAAGCSAP5 RT Rev .............................................................................................................TCCACATCACCATCTCTACCAGTCSAP5 RT Probe ..........................................................................................................FAM-AGTTTACTCCCTCAGAATTTCCCGTCGATG-TAMSp
a FAM, 6-carboxyfluorescein; TAMSp, 6-carboxytetramethylrhodamine.
3652 NETT ET AL. INFECT. IMMUN.

the course of the experiments (72 h) and continued normalintake of the food and water. The oral cavity did not appearvisibly inflamed, and no purulence was noted.
Time course experiment analysis. Time course experimentsshowing viable counts from the dentures demonstrated an in-creasing viable burden of both Candida spp. and bacteria from6 to 48 h after denture placement and inoculation (Fig. 2A). Atthe 6-h time point, �2 log10 CFU/device of viable Candida spp.and bacteria was adherent to the device surface. By the finaltime point (48 h), there were approximately 6 log10 CFU/device of viable Candida spp. and 8 log10 CFU/device of viablebacteria present on each denture, based on microbiologic platecounts. Classification of bacterial organisms suggested thepresence of a polymicrobial biofilm consisting of both aerobicand anaerobic organisms (Fig. 2B). The aerobic Gram-positivecoccus species included viridans group Streptococcus spp., En-terococcus spp., and Streptococcus agalactiae. In early studies,Escherichia coli was a prominent species identified. We felt thiswas likely related to the rodent practice of coprophagy. Weattempted to address enteric oral colonization in two ways.First, we administered a broad-spectrum antibiotic (ampicillin-
sulbactam) prior to denture placement. Antibiotic treatmentresulted in a 10-fold decrease in the number of biofilm-asso-ciated bacteria and a 100-fold increase in C. albicans bacteria(Fig. 2C). For the next experiment, we housed the animals sothat coprophagy was not possible and administered an antibac-terial agent with a narrow aerobic Gram-negative bacterialspectrum of activity, aztreonam. Both conditions were under-taken 2 days prior to denture placement, and the housingcontinued throughout the denture biofilm study. These condi-tions resulted in a �1,000-fold reduction in aerobic Gram-negative rods (3.5 log10 CFU/device compared to 7 log10 CFU/device) (data not shown).
Denture biofilm imaging. (i) SEM. We used SEM to assessbiofilm formation and architecture over the 48-h period fol-lowing denture placement and inoculation. The choice of theimaging time course experiment was based upon the identifi-cation of architecturally early and mature biofilms in previousstudies (3, 11, 24, 33, 55, 56). Imaging of the denture demon-strated adherent yeast cells after 6 h (Fig. 3). After 24 h, aconfluent layer of yeast cells had covered the denture surface,and an extracellular matrix became apparent. Over the courseof 48 h, a mature biofilm composed of two fungal morpholo-gies (yeast and hyphae) and host cells encased in extracellularmatrix was visualized. In addition, bacteria were embedded inthe biofilm. These bacilli and cocci were found growing ingroups associated with Candida biofilm cells and matrix mate-rial.
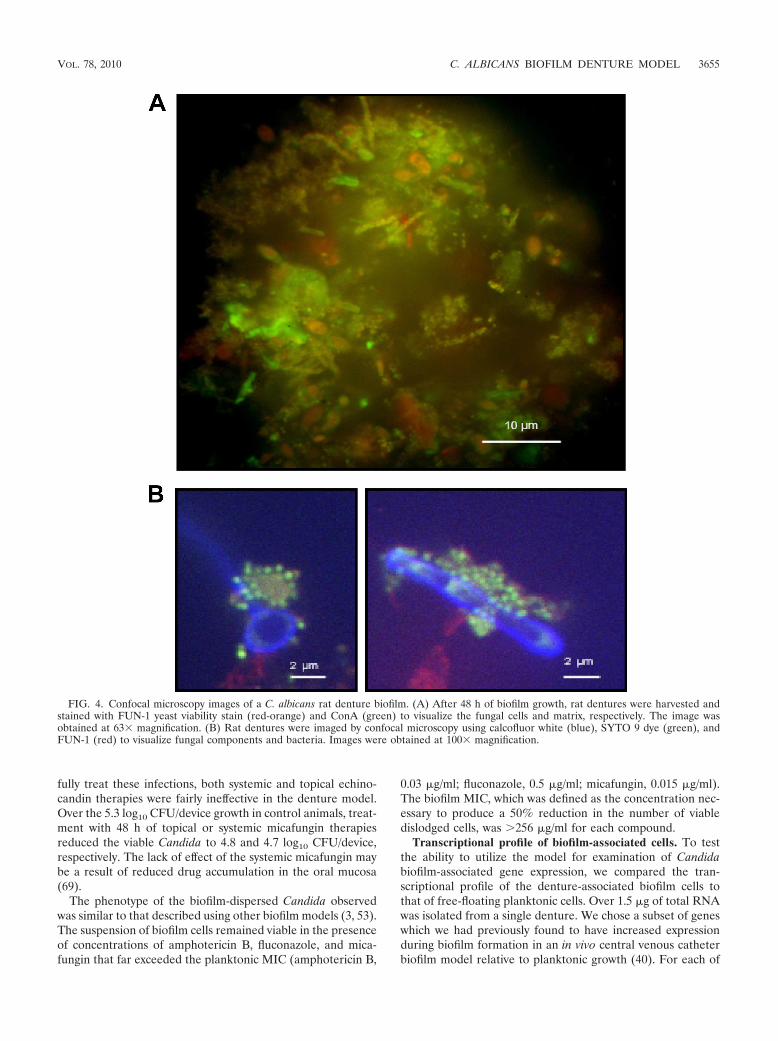
(ii) Confocal microscopy. Two sets of fluorescent dyes wereused to examine biofilm formation on the denture surfaces.First, imaging with the Live/Dead FUN-1 dye (red) confirmedthe presence of viable yeast in a mature biofilm attached to thedenture surface after 48 h (Fig. 4A). The extracellular matrixsurrounding the cells was visualized using ConA carbohydratestaining and appears green in the image capture.
The second set of dyes attempted to image both the Candidaspp. and associated bacteria simultaneously. We utilizedSYTO 9 staining to image the bacteria attached to the dentureand embedded in the biofilm. The utility of the combination ofSYTO 9 and calcofluor white in the imaging of mixed-speciesbiofilms has previously been demonstrated for Candida andStaphylococcus aureus biofilms (23). In addition to these dyes,we included FUN-1 to visualize live yeast cells. In the mul-tichannel image, bacteria appear yellow, and the predominantform was coccus (Fig. 4B). The clusters were found attached tocalcofluor white-stained hyphae and yeast cells, which appearblue.
Comparison of the reference strain and the biofilm-defectivemutant. We next sought to test the ability of the model todetect the phenotype of a Candida strain previously shown toproduce poor biofilms. We chose the bcr1�/bcr1� strain, whichlacks a zinc finger transcription factor and produces minimalbiofilms in vitro and in a central venous catheter in vivo biofilmmodel (43, 44). We hypothesized that the mutant would alsoexhibit a biofilm defect in the rat denture model. Compared toan otherwise isogenic reference strain, the bcr1�/bcr1� infec-tion of the rat denture resulted in 4-fold-fewer adherent cells(bcr1�/bcr1�, 2 � 105 Candida CFU/device; reference strain,8.7 � 105 Candida CFU/device). Interestingly, a 50%-higherburden of bacteria was found within the bcr1�/bcr1� biofilm
FIG. 2. Microbiology of rat dentures infected with C. albicans bio-film infection. (A) Rat dentures were harvested after 6, 24, or 48 h ofgrowth, and microbiological counting was used to determine the num-ber of organisms present in the biofilms. (B) Rat dentures were har-vested after 48 h of biofilm growth, and adherent bacteria were enu-merated and categorized based on Gram staining, morphology, andaerobic growth. GN, Gram negative; GP, Gram positive. (C) Rats withdentures in place received no C. albicans inoculum (control), C. albi-cans inoculum only (infection), or treatment with ampicillin-sulbactamprior to C. albicans inoculum (infection � Amp-Sulb). Microbiologicalcounting was used to determine the number of organisms present inthe biofilms after 48 h of growth. All experiments were performed ontwo occasions, and microbiologic counting was performed in duplicate.
VOL. 78, 2010 C. ALBICANS BIOFILM DENTURE MODEL 3653

(1.5 � 103) compared to that associated with the referencestrain biofilm (1 � 103).
Antifungal drug susceptibility. Similar to reports from invitro biofilm studies and the central venous catheter in vivobiofilm model, the triazole, fluconazole, did not effectivelyreduce the burden of viable Candida in this denture model (3,
6, 11, 28, 30, 32, 35, 53). The devices of control animals con-tained approximately 5.3 log10 CFU/device, while fluconazoletreatment decreased the burden by less than 0.5 log10 CFU/device (to 4.5 and 4.8 log10 CFU/ml for topical and systemictherapies, respectively). Although previous in vitro and in vivocatheter biofilm studies have shown echinocandins to success-
FIG. 3. Scanning electron microscopy (SEM) images of a C. albicans rat denture biofilm. (A) Rat dentures were harvested after 6, 24, or 48 hof growth, processed for SEM, and imaged. Scale bars for images at 50� and 1,000� magnification represent 600 �M and 30 �M, respectively.(B) Hyphal elements and bacteria are visualized in a denture biofilm imaged at 2,000� magnification.
3654 NETT ET AL. INFECT. IMMUN.
fully treat these infections, both systemic and topical echino-candin therapies were fairly ineffective in the denture model.Over the 5.3 log10 CFU/device growth in control animals, treat-ment with 48 h of topical or systemic micafungin therapiesreduced the viable Candida to 4.8 and 4.7 log10 CFU/device,respectively. The lack of effect of the systemic micafungin maybe a result of reduced drug accumulation in the oral mucosa(69).
The phenotype of the biofilm-dispersed Candida observedwas similar to that described using other biofilm models (3, 53).The suspension of biofilm cells remained viable in the presenceof concentrations of amphotericin B, fluconazole, and mica-fungin that far exceeded the planktonic MIC (amphotericin B,
0.03 �g/ml; fluconazole, 0.5 �g/ml; micafungin, 0.015 �g/ml).The biofilm MIC, which was defined as the concentration nec-essary to produce a 50% reduction in the number of viabledislodged cells, was �256 �g/ml for each compound.
Transcriptional profile of biofilm-associated cells. To testthe ability to utilize the model for examination of Candidabiofilm-associated gene expression, we compared the tran-scriptional profile of the denture-associated biofilm cells tothat of free-floating planktonic cells. Over 1.5 �g of total RNAwas isolated from a single denture. We chose a subset of geneswhich we had previously found to have increased expressionduring biofilm formation in an in vivo central venous catheterbiofilm model relative to planktonic growth (40). For each of
FIG. 4. Confocal microscopy images of a C. albicans rat denture biofilm. (A) After 48 h of biofilm growth, rat dentures were harvested andstained with FUN-1 yeast viability stain (red-orange) and ConA (green) to visualize the fungal cells and matrix, respectively. The image wasobtained at 63� magnification. (B) Rat dentures were imaged by confocal microscopy using calcofluor white (blue), SYTO 9 dye (green), andFUN-1 (red) to visualize fungal components and bacteria. Images were obtained at 100� magnification.
VOL. 78, 2010 C. ALBICANS BIOFILM DENTURE MODEL 3655

the selected genes, transcripts were more abundant for den-ture-associated biofilm cells than for planktonic cells (Table 2).While the venous catheter and denture expression studies werenot undertaken at the same time, it is interesting that the foldchange in expression of this subset of genes during biofilmgrowth varied by as much as 10-fold between the two in vivobiofilm models.
Histopathology. H&E staining of the rat oral mucosal sur-face demonstrated inflammation, marked by infiltration ofpolymorphonuclear cells in the keratinized and deeper layersof the epithelium (Fig. 5). GMS staining confirmed tissue in-vasion by Candida hyphae, consistent with the findings ofpseudomembranous candidiasis. Hyphae were also observed inthe superficial keratinized layer of the palate by GMS staining.The constellation of fungal invasion and neutrophilic inflam-mation is consistent with acute denture stomatitis (47, 58).Unlike models with prolonged infection, epithelial atrophy wasnot found (47).
DISCUSSION
Candida, one of the most common human fungal pathogens,adopts a biofilm lifestyle, often growing attached to a prosthe-sis or medical device (19, 22, 29). Many devices have beendemonstrated to serve as substrates for biofilm growth andinfection, including oral prostheses, catheters (venous or uri-nary), vascular stents, pacemakers, and joint implants (29).Advancing age and a number of pathological conditions of thehead and neck often result in a decline in oral health and theneed for prosthetic devices (70). Candida spp. and oral bacte-rial flora can grow as a biofilm on the surface of an oral device,resulting in denture stomatitis and often device malfunction.The device biofilm and associated mucosal inflammation neg-atively impact a patient’s ability to eat and speak. In addition,these mixed-species biofilms harbor pathogenic bacterial or-ganisms which commonly cause dental caries and less fre-quently cause more severe disease, such as pneumonia or en-docarditis (14, 60). Biofilm infections in general, includingthose associated with dentures, are notoriously difficult toeradicate due to associated resistance to both host defensesand anti-infective therapy (12, 25, 55).
Here, we adapted a rat denture model to mimic humanCandida denture stomatitis, the most common form of oralcandidiasis (67). The model was designed to study Candidabiofilm formation on the surface of a prosthetic oral device.The model mimics human denture stomatitis in terms of Can-dida biofilm formation, host factors, and anatomic location.The ecologic niche for denture stomatitis, the oral cavity, issimilar between rats and humans in terms of salivary flow, hostimmune response, and host salivary proteins. The role of salivain Candida adhesion is complex (18, 41, 42). Saliva has a
TABLE 2. Transcription profiles of in vivo denture-associatedbiofilm cells compared to those of planktonic cells
Transcript DescriptionRT fold
change in cells(biofilm/planktonic)
CDR1 Multidrug transport 2.5CDR2 Multidrug transport 19ERG25 Ergosterol biosynthesis 1.2ALS1 Adhesin 136SAP5 Secreted aspartyl proteinase 256GDB1 Glucanotransferase 3.3BGL2 Glucosyltransferase 1.6
FIG. 5. Histopathology of rat oral mucosa in the denture stomatitis model. Rat dentures were infected with C. albicans. After 48 h, animals weresacrificed, and dissected samples were fixed. Sections were stained with either hematoxylin and eosin (H&E) or Gomori’s methenamine silver(GMS) for C. albicans. Images were obtained at 200� magnification.
3656 NETT ET AL. INFECT. IMMUN.

physical cleansing effect and innate immune defense compo-nents, including lysozyme, histatin, lactoferrin, and IgA, whichcan act to decrease colonization and adherence (18, 50). How-ever, saliva proteins, such as mucins, may also facilitate adher-ence by absorbing to Candida or coating the oral device. Fullincorporation of the impact of the numerous and complexcomponent makeup of saliva on biofilm formation is difficult toaccomplish without use of an in vivo model.
Compared to other commonly used laboratory animals, wefound several advantages to employing a rat for the denturestomatitis model. Unlike smaller animals, such as mice, ratshave a larger oral cavity, easing denture placement. However,rats are much less expensive and easier to maintain than largeranimals, such as primates (58). The current model has theadvantage of not requiring a mold for manufacture of a pre-formed device (47, 58). With this technique, the device can beconstructed directly in the oral cavity and has the benefit of awell-fitted apparatus. One limitation of this method of place-ment is a small amount of variability (less than 10%) in sizeamong the devices.
Using this model, we found histopathologic changes to beconsistent with candidiasis and acute denture stomatitis. Sim-ilar to previously described mucosal models, we observed hy-phal invasion of the epithelial layer and inflammation markedby polymorphonuclear cells (47, 58). Unlike models with pro-longed mucosal or device infection, epithelial atrophy was notidentified, perhaps due to the relatively short duration of thecurrent studies (47). Future investigation, including a pro-longed time course experiment, may be of interest for exami-nation of the histopathology of chronic denture stomatitis.Given the animal tolerance to the denture model, longer stud-ies should be feasible. In addition, this model should be usefulfor examination of the host response to biofilm infectionthrough histopathological comparisons of animals with mixed-species biofilm infections and those with single-species biofilminfections or by study of the response to genetically manipu-lated strains.
Microscopic examination of the denture surface demon-strated a C. albicans biofilm. The biofilm consisted of adherentyeast and hyphae embedded in an extracellular matrix. In ad-dition, host cells and bacteria were identified in the biofilm.This biofilm architecture was similar to that described for a ratvenous catheter biofilm (3). The main difference was the pres-ence of a mixed microbial biofilm infection of the denturemodel. Confocal microscopy confirmed the presence of adher-ent bacteria on the surfaces of hyphae. These biofilm cellswere resistant to several antifungal drug classes, a charac-teristic typical of C. albicans biofilm cells, described in therat venous catheter model, and mimicking the therapeuticrecalcitrance of denture stomatitis observed in patients (1,3, 7, 16, 35, 39, 46, 55).
The current denture model provides an additional tool forexamining the impact of specific gene products on biofilmformation, architecture, and drug resistance in vivo. Using thismodel, we were able to examine the role of Bcr1p, a transcrip-tion factor implicated in biofilm formation, and corroboratedits importance in this niche as well (43, 44). A difference in therequirement of an adhesin, Als3p, for biofilm formation in vitroand in vivo has previously been demonstrated using a venouscatheter model (43). Similarly, we anticipate that biofilm for-
mation in the oral cavity, subjected to saliva and specific oralhost defenses, may require a unique set of gene products.Potential features may include the requirement of gene prod-ucts involved in quorum sensing, bacterial interactions, or ad-herence to epithelial cells. The current studies indicate theutility of the denture model for investigation of these ques-tions.
One method of determining the gene products required forgrowth and proliferation in an environmental niche is throughgene expression profiling. This model provides a means toexamine the transcriptome of the C. albicans denture-associ-ated biofilm cells. In current experiments, transcript abun-dances of a subset of biofilm-associated genes were compareddirectly between in vivo denture biofilm conditions and plank-tonic conditions (Table 2) (35, 40, 53, 59, 68). Biofilm cellsfrom both the denture and catheter in vivo models exhibited adirection of expression similar to that of planktonic growthprofiles with transcriptional abundance for each of the chosengenes. However, the magnitude of expression varied betweenthe two models. For example, the transcript abundance ofSAP5, a member of the extracellular aspartyl proteinase genefamily known to function in pathogenicity by degrading tissuebarriers, destroying host defense molecules, or digesting pro-teins for nutrient supply, was higher for the denture biofilmmodel (�250-fold increase versus 30-fold increase) (36, 57,64). This finding is consistent with prior studies implicating thegene product in mucosal candidiasis pathogenicity (36, 57, 64).Variability in gene expression between the sites likely reflectsthe differences in environmental factors, including tempera-ture, nutrient availability, oxygen level, and surface adherenceproperties. Further exploration for potential biofilm niche sim-ilarities and differences in global gene expression may provideimportant pathogenesis and therapeutic insights into these dif-ficult-to-manage infections.
Of the biofilm-associated phenotypes, the profound drugresistance characteristic of biofilm cells is of specific interestdue to the difficulty in eradication of these organisms (1, 35, 39,55). Using the current model, we were able to test the suscep-tibility of denture-associated C. albicans cells both in vivo andex vivo. Similar to previous investigations, the biofilms cellswere extraordinarily resistant to the triazole, fluconazole, whentested on disrupted biofilm cells or following in vivo systemic ortopical treatment (2, 35, 53, 62). Unlike previous investiga-tions, micafungin, an echinocandin, did not demonstrate sig-nificant anti-Candida biofilm activity using these methods (4,30). The reason for this apparent discrepancy is unclear butmay reflect C. albicans growth in a mixed-species biofilm at thisdistinct site. The model may prove to be beneficial as an ad-ditional environmental condition for testing of potential anti-fungal or anti-Candida biofilm drugs.
One unique feature of the Candida denture biofilm relativeto most systemic biofilms is the mixed-species nature of theprocess in the former. Although Candida spp., including C.albicans, C. glabrata, and C. tropicalis, are predominant organ-isms, many types of bacterial organisms may be present. Themost common bacterial organisms include Streptococcus spp.,Staphylococcus spp., and anaerobes, but molecular analysis ofdenture biofilms obtained from patients with stomatitis iden-tified over 50 bacterial species involved in the process (10, 52).Interestingly, the bacterial species found on the dentures of
VOL. 78, 2010 C. ALBICANS BIOFILM DENTURE MODEL 3657

patients with stomatitis appear distinct from those adherent tothe dentures of healthy patients. The impact of bacterial mi-croflora during development of denture stomatitis has notbeen fully explored, but investigations have begun to examinethe interaction between Candida spp. and bacteria in mixedbiofilms. Candida has been reported to form mixed biofilmswith both Staphylococcus and Streptococcus during denture andcatheter infection (5, 65). These bacterial species have beenshown to interact with Candida and, in some instances, impactadhesion and antimicrobial susceptibility (1, 23). The currentdenture model provides ideal tools for examination of mixedbiofilms.
The intricacies of the bacterial-fungal interaction highlightthe importance of the oral microflora during denture place-ment. One potentially important difference in the oral cavitiesof rodents and humans is the higher burden of enteric florapresent in the mouths of rodents due to coprophagy. We foundthe rat oral cavity to harbor many Gram-negative organisms,presumably linked to this behavior. To inhibit growth of theseorganisms and more closely mimic human denture stomatitis,we treated the animals with a broad-spectrum antibiotic priorto denture placement and housed animals in metabolic cageswith wire cage floors. In a subset of experiments, animals weretreated with an antibiotic specific to Gram-negative organismsand were housed in a tube which did not allow the animal toturn in place (15). These conditions greatly decreased co-prophagy and the burden of Gram-negative organisms. Weconsidered the possibility of using gnotobiotic, germ-free ani-mals and colonizing a subset of animals with pathogens typicalof human denture stomatitis. However, we currently do nothave a gnotobiotic facility, and at this time, the cost would beprohibitive.
In conclusion, these studies indicate the utility of the den-ture model for testing the impact of gene products, studyingbiofilm architecture, and examining biofilm drug resistance.Furthermore, the model is potentially valuable for the inves-tigation of mixed-species biofilms, bacterial-fungal interac-tions, and the impact of individual host factors, such as salivaryproteins and immune components. Future investigations mayinclude gnotobiotic animals with defined microflora to exam-ine the impact of specific bacteria on C. albicans biofilm for-mation and drug resistance. To better understand the role ofthe host response in this unique process, studies may includeanimals with genetically defined immune deficiencies or withreduced salivation. Comparisons to other biofilm infectionmodels promise to identify fungal and host elements distinctfor the oral biofilm environment.
ACKNOWLEDGMENTS
We thank William R. Gengler for helpful suggestions on the place-ment of oral rodent devices and Thomas Warner for assistance withhistopathology. We thank Aaron Mitchell and Clarissa Nobile forproviding strain CJN702.
This work was supported by the National Institutes of Health (grantRO1 AI073289-01).
REFERENCES
1. Al-Fattani, M. A., and L. J. Douglas. 2006. Biofilm matrix of Candida albi-cans and Candida tropicalis: chemical composition and role in drug resis-tance. J. Med. Microbiol. 55:999–1008.
2. Al-Fattani, M. A., and L. J. Douglas. 2004. Penetration of Candida biofilmsby antifungal agents. Antimicrob. Agents Chemother. 48:3291–3297.
3. Andes, D., J. Nett, P. Oschel, R. Albrecht, K. Marchillo, and A. Pitula. 2004.Development and characterization of an in vivo central venous catheterCandida albicans biofilm model. Infect. Immun. 72:6023–6031.
4. Bachmann, S. P., K. VandeWalle, G. Ramage, T. F. Patterson, B. L. Wickes,J. R. Graybill, and J. L. Lopez-Ribot. 2002. In vitro activity of caspofunginagainst Candida albicans biofilms. Antimicrob. Agents Chemother. 46:3591–3596.
5. Baena-Monroy, T., V. Moreno-Maldonado, F. Franco-Martinez, B. Aldape-Barrios, G. Quindos, and L. O. Sanchez-Vargas. 2005. Candida albicans,Staphylococcus aureus and Streptococcus mutans colonization in patientswearing dental prosthesis. Med. Oral Patol. Oral Cir. Bucal 10(Suppl. 1):E27–E39. (In Spanish.)
6. Baillie, G. S., and L. J. Douglas. 2000. Matrix polymers of Candida biofilmsand their possible role in biofilm resistance to antifungal agents. J. Antimi-crob. Chemother. 46:397–403.
7. Bissell, V., D. H. Felix, and D. Wray. 1993. Comparative trial of fluconazoleand amphotericin in the treatment of denture stomatitis. Oral Surg. OralMed. Oral Pathol. 76:35–39.
8. Budtz-Jorgensen, E. 1973. Cellular immunity in acquired candidiasis of thepalate. Scand. J. Dent. Res. 81:372–382.
9. Budtz-Jorgensen, E. 1971. Denture stomatitis. IV. An experimental model inmonkeys. Acta Odontol. Scand. 29:513–516.
10. Campos, M. S., L. Marchini, L. A. Bernardes, L. C. Paulino, and F. G.Nobrega. 2008. Biofilm microbial communities of denture stomatitis. OralMicrobiol. Immunol. 23:419–424.
11. Chandra, J., D. M. Kuhn, P. K. Mukherjee, L. L. Hoyer, T. McCormick, andM. A. Ghannoum. 2001. Biofilm formation by the fungal pathogen Candidaalbicans: development, architecture, and drug resistance. J. Bacteriol. 183:5385–5394.
12. Chandra, J., P. K. Mukherjee, S. D. Leidich, F. F. Faddoul, L. L. Hoyer, L. J.Douglas, and M. A. Ghannoum. 2001. Antifungal resistance of candidalbiofilms formed on denture acrylic in vitro. J. Dent. Res. 80:903–908.
13. Clinical and Laboratory Standards Institute. 2008. Abbreviated identifica-tion of bacteria and yeast, 2nd ed., vol. 28. Approved guideline. Clinical andLaboratory Standards Institute, Wayne, PA.
14. Coulthwaite, L., and J. Verran. 2007. Potential pathogenic aspects of dentureplaque. Br. J. Biomed. Sci. 64:180–189.
15. Cree, T. C., D. M. Wadley, and J. A. Marlett. 1986. Effect of preventingcoprophagy in the rat on neutral detergent fiber digestibility and apparentcalcium absorption. J. Nutr. 116:1204–1208.
16. Cross, L. J., J. Bagg, D. Wray, and T. Aitchison. 1998. A comparison offluconazole and itraconazole in the management of denture stomatitis: apilot study. J. Dent. 26:657–664.
17. Davis, D., R. B. Wilson, and A. P. Mitchell. 2000. RIM101-dependent and-independent pathways govern pH responses in Candida albicans. Mol. Cell.Biol. 20:971–978.
18. Dodds, M. W., D. A. Johnson, and C. K. Yeh. 2005. Health benefits of saliva:a review. J. Dent. 33:223–233.
19. Donlan, R. M. 2001. Biofilm formation: a clinically relevant microbiologicalprocess. Clin. Infect. Dis. 33:1387–1392.
20. Dorocka-Bobkowska, B., and K. Konopka. 2007. Susceptibility of Candidaisolates from denture-related stomatitis to antifungal agents in vitro. Int. J.Prosthodont. 20:504–506.
21. Douglas, L. J. 2003. Candida biofilms and their role in infection. TrendsMicrobiol. 11:30–36.
22. Douglas, L. J. 2002. Medical importance of biofilms in Candida infections.Rev. Iberoam. Micol. 19:139–143.
23. Harriott, M. M., and M. C. Noverr. 2009. Candida albicans and Staphylo-coccus aureus form polymicrobial biofilms: effects on antimicrobial resis-tance. Antimicrob. Agents Chemother. 53:3914–3922.
24. Hawser, S. P., and L. J. Douglas. 1994. Biofilm formation by Candida specieson the surface of catheter materials in vitro. Infect. Immun. 62:915–921.
25. Hawser, S. P., and L. J. Douglas. 1995. Resistance of Candida albicansbiofilms to antifungal agents in vitro. Antimicrob. Agents Chemother. 39:2128–2131.
26. Jensen, J., T. Warner, C. Johnson, and E. Balish. 1996. Oral immunizationof mice against candidiasis. J. Infect. Dis. 174:133–140.
27. Kamai, Y., M. Kubota, T. Hosokawa, T. Fukuoka, and S. G. Filler. 2001. Newmodel of oropharyngeal candidiasis in mice. Antimicrob. Agents Chemother.45:3195–3197.
28. Khot, P. D., P. A. Suci, R. L. Miller, R. D. Nelson, and B. J. Tyler. 2006. Asmall subpopulation of blastospores in Candida albicans biofilms exhibitresistance to amphotericin B associated with differential regulation of ergos-terol and beta-1,6-glucan pathway genes. Antimicrob. Agents Chemother.50:3708–3716.
29. Kojic, E. M., and R. O. Darouiche. 2004. Candida infections of medicaldevices. Clin. Microbiol. Rev. 17:255–267.
30. Kuhn, D. M., T. George, J. Chandra, P. K. Mukherjee, and M. A. Ghan-noum. 2002. Antifungal susceptibility of Candida biofilms: unique efficacy ofamphotericin B lipid formulations and echinocandins. Antimicrob. AgentsChemother. 46:1773–1780.
3658 NETT ET AL. INFECT. IMMUN.

31. Kumamoto, C. A., and M. D. Vinces. 2005. Alternative Candida albicanslifestyles: growth on surfaces. Annu. Rev. Microbiol. 59:113–133.
32. LaFleur, M. D., C. A. Kumamoto, and K. Lewis. 2006. Candida albicansbiofilms produce antifungal-tolerant persister cells. Antimicrob. Agents Che-mother. 50:3839–3846.
33. Lewis, R. E., D. P. Kontoyiannis, R. O. Darouiche, I. I. Raad, and R. A.Prince. 2002. Antifungal activity of amphotericin B, fluconazole, and vori-conazole in an in vitro model of Candida catheter-related bloodstream in-fection. Antimicrob. Agents Chemother. 46:3499–3505.
34. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-delta delta C(T)) method.Methods 25:402–408.
35. Mukherjee, P. K., J. Chandra, D. M. Kuhn, and M. A. Ghannoum. 2003.Mechanism of fluconazole resistance in Candida albicans biofilms: phase-specific role of efflux pumps and membrane sterols. Infect. Immun. 71:4333–4340.
36. Naglik, J. R., D. Moyes, J. Makwana, P. Kanzaria, E. Tsichlaki, G. Weindl,A. R. Tappuni, C. A. Rodgers, A. J. Woodman, S. J. Challacombe, M.Schaller, and B. Hube. 2008. Quantitative expression of the Candida albicanssecreted aspartyl proteinase gene family in human oral and vaginal candidi-asis. Microbiology 154:3266–3280.
37. NCCLS. 2002. Clinical Laboratory Standards Institute. Reference methodfor broth dilution antifungal susceptibility testing, 2nd ed. Document M27-A2. National Committee for Clinical Laboratory Standards, Wayne, PA.
38. Nett, J., and D. Andes. 2006. Candida albicans biofilm development, mod-eling a host-pathogen interaction. Curr. Opin. Microbiol. 9:340–345.
39. Nett, J., L. Lincoln, K. Marchillo, R. Massey, K. Holoyda, B. Hoff, M.VanHandel, and D. Andes. 2007. Putative role of beta-1,3 glucans in Candidaalbicans biofilm resistance. Antimicrob. Agents Chemother. 51:510–520.
40. Nett, J. E., A. J. Lepak, K. Marchillo, and D. R. Andes. 2009. Time courseglobal gene expression analysis of an in vivo Candida biofilm. J. Infect. Dis.200:307–313.
41. Nikawa, H., H. Nishimura, T. Hamada, H. Kumagai, and L. P. Samaranay-ake. 1997. Effects of dietary sugars and, saliva and serum on Candida bioflimformation on acrylic surfaces. Mycopathologia 139:87–91.
42. Nikawa, H., H. Nishimura, T. Hamada, H. Yamashiro, and L. P. Samaranay-ake. 1999. Effects of modified pellicles on Candida biofilm formation onacrylic surfaces. Mycoses 42:37–40.
43. Nobile, C. J., D. R. Andes, J. E. Nett, F. J. Smith, F. Yue, Q. T. Phan, J. E.Edwards, S. G. Filler, and A. P. Mitchell. 2006. Critical role of Bcr1-depen-dent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoSPathog. 2:e63.
44. Nobile, C. J., and A. P. Mitchell. 2005. Regulation of cell-surface genes andbiofilm formation by the C. albicans transcription factor Bcr1p. Curr. Biol.15:1150–1155.
45. Nobile, C. J., H. A. Schneider, J. E. Nett, D. C. Sheppard, S. G. Filler, D. R.Andes, and A. P. Mitchell. 2008. Complementary adhesin function in C.albicans biofilm formation. Curr. Biol. 18:1017–1024.
46. Olsen, I. 1975. Denture stomatitis. The clinical effects of chlorhexidine andamphotericin B. Acta Odontol. Scand. 33:47–52.
47. Olsen, I., and O. Bondevik. 1978. Experimental Candida-induced denturestomatitis in the Wistar rat. Scand. J. Dent. Res. 86:392–398.
48. Pappas, P. G., C. A. Kauffman, D. Andes, D. K. Benjamin, Jr., T. F. Ca-landra, J. E. Edwards, Jr., S. G. Filler, J. F. Fisher, B. J. Kullberg, L.Ostrosky-Zeichner, A. C. Reboli, J. H. Rex, T. J. Walsh, and J. D. Sobel.2009. Clinical practice guidelines for the management of candidiasis: 2009update by the Infectious Diseases Society of America. Clin. Infect. Dis.48:503–535.
49. Park, H., C. L. Myers, D. C. Sheppard, Q. T. Phan, A. A. Sanchez, E. E. J.,and S. G. Filler. 2005. Role of the fungal Ras-protein kinase A pathway ingoverning epithelial cell interactions during oropharyngeal candidiasis. Cell.Microbiol. 7:499–510.
50. Pereira-Cenci, T., A. A. Del Bel Cury, W. Crielaard, and J. M. Ten Cate.2008. Development of Candida-associated denture stomatitis: new insights.J. Appl. Oral Sci. 16:86–94.
51. Petraitis, V., R. Petraitiene, A. H. Groll, T. Sein, R. L. Schaufele, C. A.Lyman, A. Francesconi, J. Bacher, S. C. Piscitelli, and T. J. Walsh. 2001.
Dosage-dependent antifungal efficacy of V-echinocandin (LY303366)against experimental fluconazole-resistant oropharyngeal and esophagealcandidiasis. Antimicrob. Agents Chemother. 45:471–479.
52. Radford, D. R., S. J. Challacombe, and J. D. Walter. 1999. Denture plaqueand adherence of Candida albicans to denture-base materials in vivo and invitro. Crit. Rev. Oral Biol. Med. 10:99–116.
53. Ramage, G., S. Bachmann, T. F. Patterson, B. L. Wickes, and J. L. Lopez-Ribot. 2002. Investigation of multidrug efflux pumps in relation to flucon-azole resistance in Candida albicans biofilms. J. Antimicrob. Chemother.49:973–980.
54. Ramage, G., K. Tomsett, B. L. Wickes, J. L. Lopez-Ribot, and S. W. Redding.2004. Denture stomatitis: a role for Candida biofilms. Oral Surg. Oral Med.Oral Pathol. Oral Radiol. Endod. 98:53–59.
55. Ramage, G., K. Vandewalle, B. L. Wickes, and J. L. Lopez-Ribot. 2001.Characteristics of biofilm formation by Candida albicans. Rev. Iberoam.Micol. 18:163–170.
56. Redding, S., B. Bhatt, H. R. Rawls, G. Siegel, K. Scott, and J. Lopez-Ribot.2009. Inhibition of Candida albicans biofilm formation on denture material.Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 107:669–672.
57. Ripeau, J. S., M. Fiorillo, F. Aumont, P. Belhumeur, and L. de Repentigny.2002. Evidence for differential expression of Candida albicans virulencegenes during oral infection in intact and human immunodeficiency virus type1-transgenic mice. J. Infect. Dis. 185:1094–1102.
58. Samaranayake, Y. H., and L. P. Samaranayake. 2001. Experimental oralcandidiasis in animal models. Clin. Microbiol. Rev. 14:398–429.
59. Sanglard, D., K. Kuchler, F. Ischer, J. L. Pagani, M. Monod, and J. Bille.1995. Mechanisms of resistance to azole antifungal agents in Candida albi-cans isolates from AIDS patients involve specific multidrug transporters.Antimicrob. Agents Chemother. 39:2378–2386.
60. Senpuku, H., A. Sogame, E. Inoshita, Y. Tsuha, H. Miyazaki, and N. Ha-nada. 2003. Systemic diseases in association with microbial species in oralbiofilm from elderly requiring care. Gerontology 49:301–309.
61. Teughels, W., N. Van Assche, I. Sliepen, and M. Quirynen. 2006. Effect ofmaterial characteristics and/or surface topography on biofilm development.Clin. Oral Implants Res. 17(Suppl. 2):68–81.
62. Uppuluri, P., J. Nett, J. Heitman, and D. Andes. 2008. Synergistic effect ofcalcineurin inhibitors and fluconazole against Candida albicans biofilms.Antimicrob. Agents Chemother. 52:1127–1132.
63. Vazquez, J. A., and J. D. Sobel. 2002. Mucosal candidiasis. Infect. Dis. Clin.North Am. 16:793–820.
64. Villar, C. C., H. Kashleva, C. J. Nobile, A. P. Mitchell, and A. Dongari-Bagtzoglou. 2007. Mucosal tissue invasion by Candida albicans is associatedwith E-cadherin degradation, mediated by transcription factor Rim101p andprotease Sap5p. Infect. Immun. 75:2126–2135.
65. Wargo, M. J., and D. A. Hogan. 2006. Fungal-bacterial interactions: a mixedbag of mingling microbes. Curr. Opin. Microbiol. 9:359–364.
66. Webb, B. C., C. J. Thomas, M. D. Willcox, D. W. Harty, and K. W. Knox.1998. Candida-associated denture stomatitis. Aetiology and management: areview. Part 1. Factors influencing distribution of Candida species in the oralcavity. Aust. Dent. J. 43:45–50.
67. Webb, B. C., C. J. Thomas, M. D. Willcox, D. W. Harty, and K. W. Knox.1998. Candida-associated denture stomatitis. Aetiology and management: areview. Part 2. Oral diseases caused by Candida species. Aust. Dent. J.43:160–166.
68. White, T. C., S. Holleman, F. Dy, L. F. Mirels, and D. A. Stevens. 2002.Resistance mechanisms in clinical isolates of Candida albicans. Antimicrob.Agents Chemother. 46:1704–1713.
69. Wiederhold, N. P., and J. S. Lewis II. 2007. The echinocandin micafungin: areview of the pharmacology, spectrum of activity, clinical efficacy and safety.Expert Opin. Pharmacother. 8:1155–1166.
70. Wilson, J. 1998. The aetiology, diagnosis and management of denture sto-matitis. Br. Dent. J. 185:380–384.
71. Yoshinari, M., T. Kato, K. Matsuzaka, T. Hayakawa, T. Inoue, Y. Oda, K.Okuda, and M. Shimono. 2006. Adsorption behavior of antimicrobialpeptide histatin 5 on PMMA. J. Biomed. Mater. Res. B Appl. Biomater.77:47–54.
Editor: G. S. Deepe, Jr.
VOL. 78, 2010 C. ALBICANS BIOFILM DENTURE MODEL 3659



![· [thrush], and angular cheilitis as well as denture stomatitis." Like the bacte- rial biofilms, Candida also is capable of adhering to the surfaces of indwelling medical devices](https://img.dokumen.tips/doc/110x75/5d4e1faf88c993551d8b6a09/-thrush-and-angular-cheilitis-as-well-as-denture-stomatitis-like-the-bacte-.jpg)

























