Embed Size (px)
Citation preview

DETECTION OF MALARIA ANTIBODIES USING PLASMODIUM KNOWLESI
ANTIGEN
Dissertation Submitted for the Degree of
Mdiittv of $l)ilo!B;opI)p
MICRDBIDLDGY ' ^ l ^ r' TO ' -̂ ^5te JO j II I ^ THE FACULTY OF MEDICINE
' ALIGARH MUSLIM UNIVERSITY ALIGARH
FARRAH MIRZA
Approved :-
Prof. SOHAIL AHMAD Supervisor
PARASITIC IMMUNOLOGY LABORATORY DEPARTMENT OF MICROBIOLOGY
JAWAHARLAL NEHRU MEDICAL COLLEGE ALIGARH MVSLIM UNIVERSITY
ALIGARH (INDIA)
1 9 9 2

2 7 JUN 1994
DS2329
•• ; ii^

DEDICATED
TO
MUMMY. DADDY AND GUDIYA: WITH YOU ALL HERE
'HEAVEN IS A PLACE ON EARTH'

PARASITIC IMMUNOLOGY LABORATORY DEPARTMENT OF MICROBIOLOGY
JAWAHARLAL NEHRU MEDICAL COLLEGE ALIGARH MUSLIM UNIVERSITY
ALIGARH
CERTIFICATE
This is to certify that the work presented in this
dissertation was carried out by Ms. Farrah Mirza in my
laboratory. It is suitable for the award of the degree of
Master of Philosophy fM.Phil) in Microbiology.
(Prof.Sohail Ahmad) M.S. (U.S.A.), Ph.D.

LIST OF CONTENTS
INDEX PAGE
LIST OF FIGURES i
LIST OF TABLES ii
ACKNOWLEDGEMENTS iii
ABSTRACT vi
INTRODUCTION 1
1. Historical Review 2 2. Classification 3 3. Malaria Life Cycle 4 4. Immunity to Malaria 6 5. Malaria vaccines 18 6. Plasmodium knowlesi - The simian malaria
parasite 24
AIMS AND OBJECTIVES 26
MATERIALS 28
METHODS
- Maintenance of Plasmodium knowlesi cell 30 1 ine
- Counting of Parasites 30 - Staining and Fixation of Blood Films 31 - Antigen Isolation 31 - Preparation of Water Soluble Whole Antigen 32 - Protein Estimation 32 - Measurement of pH 32 - Indirect Haemagglutination Test 32 - Treatment of Infected Animals 34
RESULTS 35
DISCUSSION AND CONCLUSION 47
BIBLIOGRAPHY 55

LIST OF FIGURES
FIGURE 1 : Seropositivity of Malaria Specific antibodies by the IHA tests in the Blood film positive and Blood film negative cases
PAGE
38
FIGURE 2 : Photograph of Indirect Haemaggluti- 41 nation test
FIGURE 3 : Photomicrograph of a blood smear 42 from an infected monkey showing schizout stage of the parasite
FIGURE 4 Distribution of P. vivax and 44 P. falciparum in Blood film positive malaria patients
FIGURE 5 : Frequency of IHA titres in malaria 46 cases and controls

11
LIST OF TABLES
PAGE
TABLE I Seropositivity of malaria antibodies in clinically diagnosed malaria patients using indirect haemaggluti-nation test (IHA)
37
TABLE II : Seropositivity of malaria antibodies in normal control patients using IHA test
39
TABLE III : Sex distribution of malaria patients
side positive 40
TABLE IV : Distribution of Plasmodium vivax and Plasmodium falciparum in s1ide positive malaria patients
43
TABLE V Distribution of patients
IHA titres in malaria 45

ill
ACKNOWLEDGEMENTS
With the passage of time things often take on a dream
like quality. Thus, today, nearly two years after first
Joining this department for my M.Phil, my memories of the
first few days are a bit hazy. All I seem to remember is the
nervousness which I think didn't have much to do with the
change of an entire continent (having done my M.Sc. from
Nigeria) but was more due to the apprehension of having to
meet my new Supervisor, Dr. Sohail Ahmad.
However, now I can sit back and remember things with a
smile, for my M.Phil stands completed and who can I thank
more than Dr. Sohail Ahmad who was always there to push me
forward; it was his words of encouragement and his confidence
in my abi1ities which saw me through the 1abyrinth of
research work which needs to be done before one reaches the
place where I am today - very near a M.Phil degree. I Just
can't find words to thank him for everything, all I can say
is that, to me this laboratory and department would somehow
fail to exist without this pillar of strength.
I must also thank Dr.Ashok Bal who now captains the
ship of the department of Microbiology and makes sure that it
overcomes all tides that pass its way.

IV
The mouthwatering doughnuts, fabulous biryanis, all the
other wonderful delicacies and above all the kindness and
encouraging smile make it extra hard for me to forget one
person - Mrs Sohail Ahmad. Ma'am, you have really made this
laboratory 'A home away from home'.
How can I forget Dr.Maris Manzoor Khan who always lent
a patient ear to all my problems and managed to find somewhat
ingenious solutions for them. His sense of humour was the
buoy that kept me from drowning despite the numerous tides
that I had to tackle.
Hemant Bhai, without you who would I have gone to for
all that useful information and those wonderful suggestions?
Not to forget - the markers, the samosas and numerous cups of
'Chai'.
Thanks - Raka di, Ghazzanfar bhai, Khalid bhai. Sabiha,
Azra, Huma, Ritu, Chandrama, Dr.MukerJee and Zakir for being
the 'Wind beneath my wings'.
Munna Lai and BriJ Mohan despite being most helpful
will welcome the new season with a sigh of relief because
with my M.Phil complete there will definitely be a decline in
their busy 'errand' schedule.
Special thanks are due to the people at the Animal
house because without them I might have been 'caged' - with
the monkeys!

Mr. Suhi: i Is meticulous typing lids worked wonders for the
few pages whicn have taken on a new shape - that of a
presentable thesis.
Zafar Eqbal was ever ready to help even though mostly
he ended up running around in circles. However, as they say
'Its the thought that counts! ' Thank you.
Arif, but for you, I'd still oe browsing through my
papers without going forward even a stop. You have
definitely proved that you are the silver lining in Al igarn ' s
c;-Or,'a, Oops ! I me An t c !oud.

VI
ABSTRACT
PIasmodi urn know!es 1 cell line was maintained and
allowed to multiply, in vivo, in Rhesus monkeys. Daily blood
films were made to observe the appearance of parasites in the
peripheral blood. The parasitaemia reached to about 80^ on
the 12th day of infection.
The parasites were isolated through the use of numerous
centrifugations, washings and lysis of the erythrocytes by
the use of saponin. The free parasites were disrupted by
ultrasonication and the suspension was centrifuged again to
obtain the antigen.
Blood films and sera specimens were taken from 50
patients with clinically diagnosed malaria from the J.N.M.C.
Hospital Out Patient Department and from those who were
admitted to the J.N. Medical College Hospital wards, Aligarh,
India.
The blood films were stained by Leishman stain and
observed under the microscope for the presence of malaria
parasites. Indirect haemagglutination tests were done on the
sera samples to determine malaria parasite specific antibody
titres against P. knowlesi antigen.

VI1
The parasitologic examination of slides yielded 21
(42%) blood films positive for malaria parasite and 29 (58%)
negative blood films. The sera of patients with positive
blood films gave a positive antibody titre (_>1:16) in &)%
cases, while the sera from patients with negative blood films
were about 73* IHA positive.
26 control patients were also included in the study
(that is those with no clinical evidence of malaria and
showing a negative blood film) from whom blood films and sera
specimens were taken. Only 1 (4%) serum sample gave a
positive titre in the above control groups.

INTRODUCTION

Malaria is a communicable disease caused by Sporozoon
parasites of the genus Plasmodium and transmitted to man by
certain species of infected female Anopheline mosquitoes.
The disease is characterised clinically by 3 stages.
(a) Cold stage: sudden onset of fever with rigor and
sensation of extreme cold accompanied by shivering.
(b) Hot stage: patient feels burning hot and casts off
all his clothes, there is intense headache.
(c) Sweating stage: fever comes down with profuse
sweating. There is enlargement of spleen and
secondary anaemia with a tendency to relapses (WHO,
1986) .
Malaria is a threat to almost half the world's
population. It is a major health problem in tropical
countries, where approximately three hundred million cases
occur annually. In Africa alone, almost one to two million
children die of the disease every year (Siddiqui, 1991). In
India an alarming increase in malaria cases has been
reported since 1965 and the maximum number has been
recorded in 1976 (6.5 x 10^) (Ghatak ^^ al-, 1987). The
number of confirmed cases recorded in 1988 were 1.78
million (WHO, 1990).

2 HISTORICAL REVIEW
Malaria is caused by a protozoan parasite of the
genus Plasmodium. The first accurate clinical descriptions
of malarial fevers were given by Hippocrates in 400 B.C.;
(Boyd, 1949) who mentioned the classic triad of chills,
fever and sweating. It seems that malarial fevers were
also known in ancient China and that Arab physicians during
the eight to the thirteenth century were commonly
acquainted with intermittent fevers (Boyd, 1949).
The first treatment of malaria came in the middle of
the seventeenth century with the introduction of the bark
of a Peruvian tree which was subsequently called cinchona.
Pelletier and Caventon, in 1820, succeeded in extracting
quinine from cinchona which was found to have an
antimalarial effect (Scott, 1939).
Meckel, in 1847, showed black granules, embedded in
protoplasmic masses, in the blood of patients suffering
from malaria. In 1880, Laveran, a French Army surgeon
discovered the malaria parasite in Algiers, North Africa.
In 1891, Romanowsky in Russia developed a new method of
staining blood films which made possible complete studies
of the malaria parasite. It was proved by Gerhardt (1884)
that malaria could be induced in healthy persons by

3
inoculation of blood of severely ill malarious patients.
Manson (1894) hypothesised that mosquitoes transmitted
malaria and Ronald Ross (1897) found malaria parasite of
man, growing as cysts (oocysts) on the stomach wall of an
Anopheline mosquito which had previously fed on a malaria
patient.
CLASSIFICATION
There is still a lot of discussion and uncertainty
about the classification of malaria parasites. The
classification of the phylum protozoa introduced by the
society of Protozoo1ogists (Honigberg gjfc. al- , 1964)
provides a generally accepted system based on morphological
and biological criteria, as well as on phylogenetic
considerations. According to this classification, the
malaria parasites belong to the subphylum Sporozoa, class
Telesporea and subclass Coccidia.
All members of the Subphylum Sporozoa are parasitic
of which some produce resistant spores or have a stage in
their life cycle which is derived from a resistant spore;
these forms contain one or more sporozoites. All
Telosporea possess sporozoites and reproduce sexually. The
common characteristic of subclass Coccidia is the
intracellular nature of the trophozoites.

MALARIA LIFE CYCLE
More than 100 plasmodial species have been described,
causing malaria in a wide range of vertebrates and
exhibiting narrowly defined host specificity. Four species
infect man, P. falciparum. £. vivax. P, malaniafi, and P.
ovale, the first being the most lethal. P. vivax has the
widest distribution throughout the world. In India, 65 to
69 per cent of the infections are reported to be due to P.
vivax. 25 to 30 per cent due to P. falciparum and 4 to 8
per cent due to mixed infections. P. malariae is said to be
responsible for less than one per cent of infections in
India (Park and Park, 1386).
The malaria parasite undergoes two cycles of
development, the human or asexual cycle and the mosquito or
the sexual cycle. Man is the intermediate host and mosquito
the definitive host.
Asexual Cycle: The asexual cycle begins when an infected
mosquito bites a person and injects sporozoites. Four
phases are described in the human cycle.
(a) Pre-erythrocytic phase: The sporozoites disappear with
half an hour from peripheral circulation and reach the
liver cells, where they undergo a cycle of development
which lasts for 6 to 8 days. At the termination of this

5
phase a shower of parasites (cryptozoites) are liberated
into the blood circulation (Garnham, 1966; Ayala, 1977).
(b) Erythrocytic Phase: The parasite passes through the
stages of trophozoite and schizont. The phase ends with
the liberation of merozoites which infect fresh red cells.
This release of merozoites into the blood stream causes
fever, the prime symptom of malaria.
(c) Gametogony: Some of the merozoites penetrate the red
cells but instead of developing into schizonts, they become
gametocytes. These are the sexual forms of the parasite
which are infective to mosquito (Aikawa gj;: ,aJ.. , 1978).
(d) Exoerythrocyti c Phase: In £. f_aJJLiP_aJliLDl the
intrahepatic schizonts rupture almost simultaneously and
there is no persistent tissue phase. The intrahepatic
schizonts of other Plasmodia do not burst all at the same
time and may remain dormant for an indeterminate period.
The cycle remains potentially active for 1 to 3 years for
£• vi vax and P. ovale. In P. malari ae recurrences are
possible 10, 2 0 and even 40 years after sporozoite
inoculation. The exoerythrocytic cycle is sustained
exclusively by blood schizogony or released from liver,
cells are not capable of reinfecting the liver (Park and
Park, 1986).

Sexual Cycle: The sexual cycle or sporogony is initiated
when an Anopheline mosquito capable of transmitting malaria
feeds on a malaria patient. In the lumen of the midgut,
the gametocytes escape from the host cells and soon undergo
changes preparatory to fertilization. The male gametocyte
(microgametocyte) through a process known as
"exf1agellation" form a number of sperm-like bodies.
Fertilization occurs when a microgamete- enters a
macrogamete by chemotaxis and a zygote is formed. The
motile zygote or ookinate penetrates the stomach wall of
the mosquito and after about 24 hours develops into an
oocyst on the outer surface of the stomach (Sinden, 1975;
Speer £i M-, 1975). The oocyst grows rapidly and develops
within its numerous sporozoites. The oocyst ruptures into
the body cavity after about 2 weeks and many of the
sporozoites eventually find their way into the salivary
glands of the mosquito which now become infective to man
(Joklik fii al., 1984).
IMMUNITY TO MALARIA
Innate resistance influences the pattern of acquired
malarial immunity which differs widely in distinct host-
parasite combinations and even with the same parasite in
closely related host species. The mechanisms of innate
resistance has been shown to influence the development of

7
a l l asexual blood forms but none of those res is tances seem
to act upon gametocytes ( M i l l e r and Car te r , 1976).
I n d i v i d u a l s t h a t lack the Duffy b lood-group ant igens
on t h e i r e r y th rocy tes are r e s i s t a n t t o P. v ivax i n f e c t i o n ,
because the receptor f o r the P. v ivax merozoi tes on the red
c e l l i s assoc ia ted w i t h ant igens of the Duffy blood group
(Butcher ^ a l - , 1973; M i l l e r e i a l . , 1973, 1978; Barnwell
e t a l . • 1 9 8 9 ) . T h i s e x p l a i n s t h e l o n g r e c o g n i s e d
res is tance t o P. v ivax i n f e c t i o n of West A f r i c a n blacks and
t h e i r descendants i n the USA. Because these popu la t ions
have a h igh f requency o f the D u f f y n e g a t i v e genotype
( M i l l e r e i a_L- , 1 9 7 6 ) . H e t e r o z y g o t e s f o r
haemoglobinopathies and other d i so rders of the red c e l l s
( t h a l a s s e m i a and g 1 u c o s e - 6 - p h o s p h a t e d e h y d r o g e n a s e
d e f i c i e n c y ) promote i n n a t e r e s i s t a n c e t o P.. f a l e i oarum
i n f e c t i o n (Wea the ra l l , 1987). Such p r o t e c t i o n i s probably
t r i gge red by m o d i f i c a t i o n s of p a r a s i t e development w i t h i n
the e r y t h r o c y t e s o f these i n d i v i d u a l s . However, i t has
been proposed t h a t these simple p h y s i o l o g i c a l e f f e c t s may
f o s t e r p r o c e s s e s t h a t l e a d t o an enhancement i n t h e
i n t e n s i t y a n d / o r s p e c i f i c i t y o f t h e a d a p t i v e immune
response, e n a b l i n g i n d i v i d u a l s c a r r y i n g t h e s e genes t o
acqu i r e c l i n i c a l immunity t o m a l a r i a f a s t e r t h a n o t h e r s
( A l l i s o n , 1984 , B a y o u m i , 1 9 8 7 ) . I n s u p p o r t o f t h i s

8
hypothesis Bayoumi sX. al (1990) found that lymphocytes of
individuals carrying the HbAS genotype showed higher
reactivity to malaria antigens in proliferative assay than
lymphocytes from HbAA controls living in the same area.
Protection to malaria, acquired by individuals living
in endemic areas is largely governed by the transmission
pattern. MacDonald (1957) identified two epidemiological
extremes in which the disease exists, one where instability
of transmission is the dominant feature and the disease is
characterized by substantial seasonal fluctuations,
producing cyclical epidemics in the entire population
regardless of age. In the other case, transmission and
morbidity remain stable over the seasons and years, with
the population exposed to frequent heavy inoculations. The
individuals appear to acquire protection against clinical
attacks after several years (Theander, 1992) and the
disease therefore affects mainly children.
McGregor (1960) proposed that individuals living in
areas with stable malaria pass through five stages before
immunity is acquired. In stage one (age group 0-2 months)
infants are protected against malaria and parasites are
only found in the peripheral blood of approximately 10*
(Gilles, 1961).

9
During stage two (2-6 months) parasites are found
more frequently in the blood (Gilles, 1961) and the
children often experience the first mild clinical attack of
malaria.
Stage three (7 months-2 years) is characterized by
repeated attacks of malaria which are associated with
severe illness. Parasites are almost always found in the
peripheral blood in high densities. Malaria disrupts the
normal pattern of growth in this age, producing severe
anaemia and high mortality rate (McGregor gi aJ_. , 1956).
Stage four (3-4 years) represents the acquisition of
the first phase of clinical immunity characterized by a
rapid decline in the severity and.frequency of clinical
episodes, despite the persistence of a relatively dense
parasitaemia.
In the fifth stage clinical malaria attacks are rare
and mild. Adults living in the area often harbour the
parasite without clinical symptoms. Reports by Colbourne
(1955) on susceptibility to malaria in students returning
to Africa after studies in Europe, and by Mastbaum and
Goeckel (as quoted by Bruce-Chawatt, 1963) on malaria
outbreaks suggest that immunity diminishes in individuals
living in areas free of transmission. Whether this is due

10
t o i n t r o d u c t i o n of new p a r a s i t e s t r a i n s when exposure was
r e s t o r e d or due t o lack o f i m m u n o l o g i c a l memory i s no t
known.
Thus i n d i v i d u a l s l i v i n g in areas of s t ab l e malar ia
t ransmiss ion develop p r o t e c t i o n to the devas ta t ing e f f e c t s
of the d isease over a number o f y e a r s . Such p r o t e c t e d
i n d i v i d u a l s are c h a r a c t e r i z e d by t h e r a r i t y o f c l i n i c a l
symptoms and t h e i r mi ldness i f an a t tack of ma la r ia occurs .
Acquired immunity i s on ly p a r t i a l , as pa ras i t es are found
i n low d e n s i t y i n t h e p e r i p h e r a l b l o o d o f p r o t e c t e d
i n d i v i d u a l s and immunity wanes once t h e exposu re t o t h e
pa ras i t e ceases.
Defence mechanisms o f immun i ty a re med ia ted and
regu la ted through an i n t e r p l a y between d i f f e r e n t pa r ts of
the system. The c l a s s i c a l d i v i s i o n i n t o the s p e c i f i c and
non -spec i f i c pa r ts of the system and between B and T - c e l l
m e d i a t e d i m m u n i t y becomes l e s s d i s t i n c t when i t i s
understood t h a t T - c e l l s f o r instance s t i m u l a t e non -spec i f i c
N a t u r a l K i l l e r (NK) c e l l s and m o n o c y t e s t h r o u g h t h e
s e c r e t i o n o f I n t e r l e u k i n -2 - f I L - 2 ) and i n t e r f eron- lJ
( IFN->^ ). T - c e l l s are on l y a c t i v a t e d when exposed t o
a n t i g e n by a n t i g e n p r e s e n t i n g c e l l s . The e f f e c t o f
an t ibod ies are o f t en t o opsonize micro-organisms so t ha t

11
they are easily phagocytized, the T-cells regulate the
production of antibodies by B-cells. (Theander, 1992).
The premise that macrophages, or their products are
important for protection is supported by experiments in
which mice were protected against sporozoite-induced
(Nussenzweig, 1967) or blood-stage induced malaria
infections CClark ei al., 1976, 1977) by non-specific
stimulation of the phagocytic defence systems through the
administration of BCG. Cells of the macrophage lineage and
neutrophils are also able to kill liver stages of the
parasites and have been shown to infiltrate liver tissue
and destroy rat hepatocytes infected with P. berqhei (Meis
gLl a,I. , 1987).
Monocytes or macrophages eliminate malaria parasites
both by phagocytosis and through secretion of molecules
toxic to the parasite. Peripheral blood monocytes from
malaria unsensitized donors are able to attach to and
ingest P. falciparjum merozoites. Phagocytosis is markedly
enhanced in the presence of immune serum (Krusmith and
Druilhe, 1983). Results obtained in a mouse model indicate
that non-activated macrophages primarily bind Plasmodia via
receptors for carbohydrate containing glycoproteins, while
activated macrophages bind them via Fc receptors (Brown and
Kreier, 1986). Phagocytosis seems to be regulated by the

12
T-cells as Plasmodium antinen-stimu1ated T-cel1s from
spleens of malaria infected mice produced a lymphokine
capable of activating macrophages for enhanced Fc-receptor
mediated phagocytosis (Ockenhouse and Shear, 1983). Malaria
parasites trigger macrophages to release parasite toxic
mediators and the activity is regulated by T-cells
(Ockenhouse and Shear. 1984).
Macrophages play an important role as antigen
presenting cells. T-cell receptors co-recognize complexes
of antigen and products of major histocompatibility complex
(MHO on the surface of other cells. Subpopulations of
antigen presenting cells handle antigens differently, and
the way monocytes process and present antigens play a role
in activation of subpopulations of T-cells, whereby they
influence the outcome of the activation process (Theander.
1992).
Besides their role in protection, the macrophages
appear to play a role in the pathogenesis of the disease.
Malaria antigen can stimulate tumour necrosis factory:
(TNF- oC ) and IL-1 production in macrophages (Jakobsen e_t
al-, 1990), although these can induce anti-parasitic
effects. They probably also contribute to the symptoms arid
complications of malaria (Clark ejt a±. . 1989). Both

13
monokines can induce fever and they seem to play a critical
role in the pathogenesis of cerebral malaria (Grau et al..
1987).
Neutrophils can inhibit the growth of P. falciparum
in vitro (Brown and Smalley, 1981; Nnalue and Friedman,
1988). They probably kill the parasite and have been shown
to phagocytize parasitized erythrocytes j_n vivo (Kharazmi
e.t al. , 1987 ) and in vitro (Celada et al- . 1983).
Little is known about the role of eosinophils in the
protection against malaria as the infection does not result
in eosinophi1ia. The in vitro growth of P. falciparum can
be inhibited by granule proteins secreted by purified
eosinophils. Both the eosinophil cationic protein and
other granule products seem to be toxic to parasites as
monoclonal antibody against the cationic protein only
partially blocked the inhibitory effect (Waters et a]..
1987) .
T-cells might participate in the protection against
malaria as (1) helper cells for antibody production, (2)
through the activation of non-specific effector cells like
macrophages or granulocytes, (3) through the production of
substances toxic to the parasites and (4) as cytotoxic
cells (Theander, 1992).

u A l t h o u g h T and B - c e l l s can c o n v e y some i m m u n i t y ,
o p t i m a l p r o t e c t i o n i s o b t a i n e d by t h e t r a n s f e r o f T - c e l l s
o f t h e Ly-1 pheno type , i n c o m b i n a t i o n w i t h B - c e l l s .
A l t h o u g h T and B - c e l l s can c o n v e y some i m m u n i t y
o p t i m a l p r o t e c t i o n i s o b t a i n e d by t h e t r a n s f e r o f T - c e l l s
o f t h e L y - I pheno type , i n c o m b i n a t i o n w i t h B - c e l l s .
However, T - c e l l s a re a l s o ab le t o convey immuni ty i n
a n t i b o d y i n d e p e n d e n t s y s t e m s . B - c e l l d e f i c i e n t m i c e
t e r m i n a t e t h e i r i n f e c t i o n i n t h e same way as
i m m u n o l o g i c a l l y i n t a c t m i c e , t h e p r o t e c t i o n depends on the
presence o f an i n t a c t s p l e e n (Grun et al. , 1985) . T - c e l l s
m i g h t a l s o p l a y a r o l e i n p r e v e n t i n g t r a n s m i s s i o n o f
m a l a r i a as H a r t e e i a j_ . ( 1 9 8 5 ) w e r e a b l e t o r e d u c e
t r a n s m i s s i o n by 95% by p a s s i v e t r a n s f e r o f T - c e l l s f r o m
immune t o u n p r o t e c t e d a n i m a l s .
S e r o - e p i d e m i o l o g i c a l s t u d i e s o f t o t a l m a l a r i a
s p e c i f i c a n t i b o d i e s have i n d i c a t e d t h a t t h e r e i s an age
dependent i n c r e a s e i n a n t i b o d y l e v e l s i n i n d i v i d u a l s l i v i n g
i n m a l a r i a endemic a r e a s ; t h e l e v e l o f t hese a n t i b o d i e s i s
not an index o f f u n c t i o n m a l a r i a immun i t y , bu t an i n d i c a t o r
o f t h e d e g r e e o f m a l a r i a e x p o s u r e i n t h e p o p u l a t i o n
( V o l l e r , 1 9 7 1 ) . The a n t i b o d i e s a r e d i r e c t e d a g a i n s t
s p o r o z o i t e ( N a r d i n e t a l . , 1979) as w e l l as b lood s tage

15
(Cohen and Butcher. 1971) and gametocyte (Mendis et a1..
1987) antigens.
Complement does not seem to be able to kill parasites
directly, but could play a role as an opsonin for
neutrophils and macrophages (Salmon gt al., 1986).
The acquisition of immunity is a slow process during
which the individual is exposed to the parasite on numerous
occasions. The parasite could (1) prevent or divert the
immune recognition and (2) inhibit the immune responses to
antigens that have, or could be recognized by the immune
system. During blood stage infections, the parasite is
mostly intracellular and exposed to the extracellular
environment only for short periods of time before it
invades a new host cell. The parasite is partly hidden
from the immune system. The erythrocytes infected with
late trophozoites and schizonts bind to the endothelial
cells of the capillary system, thus avoiding the effector
cells of the spleen and other tissues.
More than twenty P. f alci parum proteins have been
shown to share amino acid sequences with human proteins
(McLaughlin gt al., 1987). The blood stage antigen Pf 11.1
shows homology with a thymic peptide (Dubois et al., 1988).
a gametic stage antigen contains segments homologous with

16
epidermal growth f a c t o r (Kaslow et_ a j . . , 1988) and the
cen t ra l repeat segment of the c i rcumsporozo i te p r o t e i n is
homologous w i t h a m i tochondr ia l p r o t e i n (McLaughlin e t a l . ,
1987). The e f f e c t of these homo log ies m i g h t be t h a t
o the rw ise immunogenic segments o f m a l a r i a p r o t e i n s are
recognized as ' S e l f and t o l e r a t e d by the immune system.
Paras i tes have been repor ted to cross the p lacenta and gain
access t o the f e t a l c i r c u l a t i o n b e f o r e and d u r i n g b i r t h
(Reinhardt e t al. , 1978). Although these c h i l d r e n seldom
develop c l i n i c a l ma lar ia (McGregor, 1984), to le rance might
be induced f o l l o w i n g the prenata l or p e r i n a t a l exposure to
malar ia ant igens (Pombo e t aj_-, 1988).
The pa ras i t es appear t o be able to undergo an t igen ic
v a r i a t i o n w i t h i n a s i n g l e c l o n e and exp ress a l t e r n a t i v e
forms of a molecule from a given genie r e p e r t o i r e (Hommel
e t al . . 1983, Fenton ejt a j_- , 1985 ) . Such d i v e r s i t y i n
m a l a r i a p a r a s i t e s i s l i k e l y t o c o n t r i b u t e t o the s low
development of immuni ty . Many m a l a r i a p r o t e i n s c o n t a i n
shor t amino ac id sequences t h a t are ex tens i ve l y repeated in
tandem a r r a y s . These r e p e a t e d sequences encode
immunodominant e p i t o p e s t o wh ich much o f t he a n t i b o d y
response a g a i n s t the p a r a s i t e i s d i r e c t e d (Kemp eX. a l . ,
1987). An t ibod ies aga ins t one such ep i tope o f t en c ross-
react w i t h o ther repeated ep i topes , e i t h e r w i t h i n the same

17
molecule or on other parasite proteins. Anders (1986)
proposed that these cross-reactions interfere with the
maturation of hiqn affinity antibody directed against the
parasi te.
Acute malaria infection causes marked perturbations
in the immune system of the host. On one hand, the
infection induces a powerful polyclonal activation of the
immune system resulting in hyper-gamma globulinaemia and
production of auto-antibodies, and on the other hand the
infection results in a depression of immune reactivity.
Symptoms and signs of respiratory and gastrointestinal
infections are common in children with acute malaria, but
this may be attributed directly to the parasite (Greenwood
ejt a.!., 1989). Clinical reports have indicated that
Salmonella infections are especially severe in malarious
children (Bennett and Hook, 1959). Although clinical
evidence for immuno suppression during malaria is not
overwhelming, results of these studies on P.. falciparum
malaria plus convincing evidence showing immunosuppression
during malaria in animals has led to a series of studies in
which immune reactivity during P. falciparum malaria was
characterized jm vi tro (Weidanz, ^962].

18 MALARIA VACCINES
Accordinq to WHO, about 100 million people are
clinically ill with malaria at any given time, and around 1
million - mostly very young children in Africa - die from
it each year. The effects are so widespread in the tropics
- ranging from lethargy to total incapacitation - that
malaria has been blamed for impeding the development of
entire nations and the problems with malaria will be
getting worse in the years ahead.
The main strategy for fighting malaria through the
1960s was to use pesticides to kill mosquitoes that carry
the parasite and to treat infected individuals with quinine
- related drugs, mainly chloroquine. However, the most
lethal strain of the parasite, P.. falciparum is now
genetically resistant to chloroquine. Health authorities
are switching to a new compound, mefloquine, but already
there are signs in Asia and Africa that the parasite is
resistant to this as well. It is only a matter of time
before the resistance spreads (Cherfas, 1990).
Malaria vaccines will be based on pure parasite
antigens which specifically stimulate protective immune
responses. Protective antigens are present in several of
the developmental forms of the parasite and future vaccines

19
may contain antigens from one or more of these, at present
most of the relevant research concerns P.. f a1 c i parum.
Vaccine trials currently envisaged are: (i) Sporozoites,
(ii) asexual erythrocytic stage, and (iii) gametes and
other forms developing in the mosquito midgut (WHO, 1986).
S-antigens of P. falci parum have also been successfully
employed for vaccination experiments (Ristic et al., 1985).
Vaccination of rodents, monkeys, birds and humans
with X-irradiated sporozoites induces protective immunity
to subsequent challenge with living sporozoites (Strickland
and Nussenzweig, 1982). Preliminary attempts at sporozoite
vaccination were made in 1971 - 1975 using X-irradiated
sporozoites of P. f al ci parum and P_. v i v a x which were
innoculated by mosquito bites into previously non-exposed
human volunteers. This bold trial brought to light the
information that the immunity to the Sporozoite challenge
became detectable 7-10 days after innoculation, immunity
was species and stage specific and appeared after 10 days
of immunization and persisted for 3-5 months and, no side
effects to the vaccine were observed (WHO, 1986). The most
promising achievement with regard to the development of a
sporozoite vaccine was gene cloning of P. knowlesi antinens
in Escherichi a coli. This was done by extracting the
messenger RNA encoding circumsporozoite protein from

20
infected mosquitoes through conversion, by enzyme reverse
transcriptase, into CDNA. The fragments of CDNA were
inserted into bacterial plasmids which were in turn
introduced into £. coli. The bacterial clones excreted
peptides binding to the monoclonal antibodies and against
circumsporozoite protein of P. knowlesi (Sharma and Godson,
1985) .
Sequencing analysis of circumsporozoite protein
showed that the immunodominant antigenic determinant
(epitope) contains a sequence of 12 amino acids and the
peptides share common antigens with the original
circumsporozoite protein (WHO, 1985).
A sporozoite vaccine, if fully effective, would
prevent the establishment of plasmodial development in the
host and thus induce sterile immunity. Parasitaemia,
asexual or sexual, would therefore not occur; clinical
illness would not supervene and the subject would remain
incapable of infecting mosquitoes. A vaccine of this type
should be able effectively to interrupt the natural
transmission of malaria; however, subjects effectively
immunized against sporozoites will remain susceptible to
challenge with asexual erythrocytic stages of the parasite
(WHO. 1986).

21
The merozoite or an asexual erythrocytic-stage
vaccine is expected to induce an immunity that operates by
restricting the replication of asexual blood stage
parasites without necessarily inducing sterile immunity.
Consequently its function will be to reduce the morbidity
and mortality due to malaria. However, persons immunized
by this type of vaccine will remain susceptible to
sporozoite infection and the parasites development in the
liver will occur unimpeded; low grade asexual parasitaemia
may occur and gametocytogenesis may evolve normally so that
immunized persons would remain capable of infecting
mosquitoes. It is unlikely that the asexual blood stage
vaccines if used alone will achieve the interruption of
malaria transmission in any endemic area; however they can
be used in highly susceptible groups in endemic areas to
induce a level of immunity which prevents serious illness
after infection. The merozoite vaccine acts as a curative
measure, and will overcome the problem of drug resistance.
Irradiated parasitized erythrocytes have been extensively
utilized as experimental immunogens. Some success was
achieved with rodent malaria (Wellde and Sadun, 1967) in
which irradiated erythrocytes infected with P. faJclEAriM
were used to vaccinate owl monkeys (Wellde et a_i. . 1979).
Two significant findings of these studies show that a
degree of protection could be obtained without using an

22
adjuvant; and that four immunizinq doses were required to
achieve significant degree of immunity.
The third type of vaccine will operate by inducing in
the human host serum antibodies that effectively block the
fertilization of female gamete by male gametes v*ithin the
mosquito gut, or inactivate the fertilized zygote or the
ookinete. Such transmission blocking vaccines will
apparently be capable of interrupting malaria transmission
at the mosquito level, but will neither protect the human
host against sporozoites, asexual blood stages, nor prevent
the development of gametocytaemia. However, they are
expected to reduce the overall rate of malaria transmission
in endemic areas and thereby play an important role in
control (WHO, 1986). The major prerequisite for producing
a gamete vaccine is the availability of large number of
infectious P. falci parum gametes and the continuous in
vitro cultivation of P. falciparum has given great impetus
for achieving this goal.
The S-antigens of P. falciparum are the subject of
great deal of research for vaccine production. Th.ey incucf
significant protective immunity in Squirrel monkeys against.
a virulent challenge by P. falciparum (Ristic e_t aj. • •
1985). Shamansky e_t a_l. (1985) have purified two S-

23
antigens (83 and 100 KDa) and have proposed that these may
be good candidates for potential host protective antigens
in malarial immunoprophylactic regimens.
Development of successful malaria vaccines will
depend on both the production of protective antigens by
chemical synthesis or recombinant technology as well as the
availability of safe and effective adjuvants. The best
protection has been achieved only when monkeys were
immunized in conjunction with Freund's complete adjuvant
(FCA).
Results of clinical trials with sporozoite and
merozoite vaccines with alum as an adjuvant have shown
limited immunogenicity and protective success. Since FCA
is unacceptable for clinical use, as it has toxic side
effects, and results of clinical trials with alum have
shown limited success, as it shows humoral response but is
deficient in inducing a ce11-mediated response; several
laboratories have sought to establish a safe and effective
adjuvant. Saponin has been found to be an effective
adjuvant with rodent malaria antigen but its efficacy with
purified or subunit human malaria antigens has yet to be
studied. Based on knowledge generated on the immunology of
malaria, it is evident that a malaria vaccine adjuvant must

24
be capable of inducing both humoral and cell mediated
responses (Siddiqui, 1991a).
The prospect for the development of effective malaria
vaccines is reasonably good, but the realisation of this
goal will require some time. Parasite antigenic diversity
and genetic restrictions of immunological responses to
these antigens in the human population pose great problems
to the effectiveness of subunit malaria vaccines now beinn
developed.
Plasmodium knowlesi - THE SIMIAN MALARIA PARASITE
The parasite is remarkable for its unique quotidian
periodicity in the blood and it is almost 10051̂ lethal in M.
malatta: however in M. fasciculari s it is not lethal as the
animal almost always survives the infection. The prepatant
period is about 6 days and the animal soon becomes very ill
as schizogony is rapid, resulting in rapid multiplication
of the organisms. The animal becomes very weak 48 hours
before death, for it refuses food, and has pallor of the
skin and conjunctiva. There is high fever which becomes
subnormal shortly before death (Garnham, 1980).

^5
I n t r a v a s c u l a r h e m o l y s i s i s o f t e n p r e s e n t a long w i t h
h a e m o g l o b i n u r i a . t h e p h e n o m e n o n o f e x c e s s i v e b l o o d
d e s t r u c t i o n has been a t t r i b u t e d t o a u t o - i m m u n i z a t i o n
(Zuckerman. 1966) .

AIMS AND OBJECTIVES ̂

26
The present study aims to gheck the efficacy of P.
knowlesi antigens for their use in serodiagnostic testa for
human malaria. The role of serodiagnosis in malarial
infections is indeed limited. At best it could be used as
an adjunct to microscopic detection of parasites. Yet it
would make things simpler if P. knowles i antigen, which
can be easily obtained compared to antigens from human
malarial parasites, could be used for identification of
malaria specific human antibodies. Antigen pools from P.
knowlesi could then be used for conducting large scale
sero-epidemiological surveys in malaria endemic regions.
Present investigations were, therefore, aimed to test
the usefulness and the efficacy of P. knowlesi antigens in
the identification of human malaria antibodies by means of
indirect haemagglutination tests. If this antigen proves
useful in IHA test then, hopefully, the antigen from monkey
malaria can be usefully employed in several other
diagnostic tests such as ELISA, RIA and several others.
Slide smears with P. knowlesi infected monkey RBCs can also
be effectively used for the detection of malaria specific
human antibodies in IFA tests. Infact some preliminary
tests along these lines have already been done in one or
two other laboratories.

11
The Dresent investigations were therefore designed to
achieve the above objective of using P. knowTesi antigen in
IHA tests. The details of the experimental orotocols which
were followed are as under:
(1) P. knowlesi cell line was maintained in vivo using
Rhesus monkeys as an animal model.
(2) P. knowlesi antigen was isolated from the parasites
harvested from the blood of infected Rhesus monkeys.
(3) Serum samples were collected from clinically
diagnosed malaria patients irrespective of their
having a negative or a positive blood film.
(4) Serum samples were collected from patients with no
clinical evidence of malaria for the past five years
and negative malaria blood films to serve as
controls.
( 5 ) Indirect haemagg1utination tests were performed on
the serum samples using P. knowles i antigen for
serodiagnosis of malaria.

MATERIALS AND METHODS

28
ANIMALS
Two young and healthy four month old Rhesus monkeys
(Macaca mulatta) . a male and a female, each weighing about
3-5 kgs were purchased from a local animal dealer.
CELL-LINE
PI asmod i urn K n o w l e s i c e l l l i n e was o b t a i n e d f r o m t h e
M i c r o b i o l o g y D i v i s i o n o f t h e C e n t r a l D r u g R e s e a r c h
I n s t i t u t e . L u c k n o w , I n d i a .
ANTICOAGULANT
A c i d c i t r a t e d e . x t r o s e was o b t a i n e d f r o m t h e B l o o d
B a n k , J . N . M e d i c a l C o l l e g e , A l i g a r h .
STAIN
Leishman stain was purchased from BDH. England.
CHEMICALS
Sodium corbonate. sodium potassium tartarate.
barbital, sodium barbitone, copper sulphate, disodium
hydrogen phosphate, dihydrogen sodium phosphate and sodium
chloride were obtained from BDH (India). Tannic acid was
purchased from W.J. Bush and Co., England and Methanol from
Merck (India). Bovine serum albumin and saponin were
obtained from Sigma chemical Co., USA and chloroquine

29
injectible from IPCA Pvt. Ltd., Bombay Sheep red blood
cells were obtained from healthy sheeps in the Central
Animal House of the J.N. Medical College, Aligarh.
Sodium azide, used as a preservative, was obtained
from Rield, Germany.
GLASSWARE
A l l t h e g l asswa re used were o b t a i n e d f r o m Pyrex and
Co .and C o r n i n g ( I n d i a ) L t d . The m i c r o t i t r e " U ' b o t t o m
p l a t e s were f r om Cooke E n g i n e e r i n g C o . , A l e x a n d r i a .
G l a s s d i s t i l l e d w a t e r was u s e d t h r o u g h o u t t h e
e x p e r i m e n t and a s e p t i c c o n d i t i o n s m a i n t a i n e d w h e r e v e r
n e c e s s a r y .
SPECIMENS
Five ml of blood was taken from 50 patients with
clinically diagnosed malaria, who were attending the Out
Patients Department, or from those patients who were
admitted to the J.N. Medical College, Hospital Aligarh,
India.
The blood was drawn by a sterile syringe and needle
(21G) and allowed to clot in a sterile universal bottle, or
screw capped tube, which was later stored at 4*̂ C overnight.
The serum was obtained by centrifuging at 1,000 rpm for 30

30
minutes and stored in a sterile screw capped tuPe at -20°C
for further use.
Five ml of blood was also taken from 25 patients of
the same age group with no clinical evidence of malaria for
making malaria negative blood films to serve as controls.
MAINTENANCE OF Plasmodium knowlesi CELL LINE
The P. knowlesi cell line was maintained and allowed
to multiply In v i vo in Rhesus monkeys which were
inoculated, intravenously, with approximately 5.3 x 10
parasitized monkey erythrocytes. After the appearance of
parasites in the blood the parasitaemia reached B0% on the
5th day of their appearance; the animals were bled at this
stage and the blood collected in Acid Citrate Dextrose.
Five ml of the infected blood sample was inoculated into a
fresh animal, for subsequently maintaining the cell-line,
while the rest was pooled for antigen isolation. Normal,
uninfected monkeys were used as controls.
COUNTING OF PARASITES
Blood films were made every day by a finger prick to
observe the appearance of the parasite after which they
were used for counting the parasites.

31
STAINING AND FIXATION OF BLOOD FILMS
Fixation of the dried blood films was done by
using methanol for 4 minutes and the films dried again.
The staining was carried by mixing equal volumes of
Leishman stain and phosphate buffer (0.01 M, pH 7.2) and
then layering this solution on the fixed and dried blood
film for 30 minutes. The slides were then washed with
phosphate buffer, air dried and examined under the oil
immersion of a microscope (Carl Zeiss, Jena).
ANTIGEN ISOLATION
Parasites were harvested according to the procedure
by Spira and Zuckerman (1962). At 80S$ parasitaemia blood
was collected from the infected monkey in ACD by cardiac
puncture under anaesthesia (Pentabarbitone sodium). The
pooled cells were washed by centrifugation in chilled
normal saline. The buffy coat containing leukocytes,
plasma and platelets was discarded and the sediment with
normal and parasitized erythrocytes pooled. The pooled
erythrocytes were washed 4 times by centrifuging at 1500
rpm for 10 minutes and incubated with an equal volume of
saponin (0.05X in normal saline) with continuous stirring
in an ice bath for 10 minutes. The lysate was centrifuged
at 1500 rpm for 10 minutes at 4*^0, the supernatant

32
discarded and the pellet washed again 3-4 times with
chilled normal saline to remove erythrocytes. After the
final washing the pellet consisting of malaria parasite was
suspended in chilled normal saline.
PREPARATION OF WATER SOLUBLE WHOLE ANTIGEN
The parasites were disrupted by ultrasonication of
the parasite suspension in 3 stretches for 6 minutes
altogether, in a 50 watt, 9 Kc magnetostriction oscillator.
The suspension was centrifuged again at 1500 rpm for 20
minutes and the supernatant collected was stored as soluble
antigen,
PROTEIN ESTIMATION
Protein estimations were done according to Lowry ejt
al . . (1951) using a Bausch and Lomb Spectronic 21
Spectrophotometer at room temperature.
MEASUREMENT OF pH
The pH was measured by an Elico LI-120 pH meter using
sodium tetraborate (0.1 M, pH 8.18) and Potassium hydrogen
pthalate (0.05 M, pH 4.0) as standard buffers.
INDIRECT HAEMAGGLUTINATION TEST (IHA)
The IHA test was done according to the method
described by Mathews §:t al-, (1975).

33
Sheep erythrocytes (SRBC) were washed 4 times with
phosphate buffered saline (pH 7.2) after which a 0. 3S6 (v/v)
suspension of the SRBC was prepared using PBS. The
suspension was mixed with an equal volume of 1/20,000
tannic acid and incubated at 4*̂ 0 for 30 minutes with
intermittent shaking. Tanned SRBC were washed thrice with
PBS-BSA for the removal of free tannic acid and resuspended
at 3%. A 0.1 ml tanned SRBC suspension was mixed with an
equal volume of the diluted antigen and incubated at 37°C
for 30 minutes with intermittent shaking. The antigen
coated SRBC were washed 3 times with PBS-BSA and was used
for making a ^ .5% suspension.
An 0.025 ml of serially diluted serum sample was
added to an equal volume of antigen coated SRBC suspension
in a 'U' bottom microtitre plate. The control wells
containing tanned RBC, normal sera and antigen were
included in the test. The plates were incubated at room
temperature in a humid chamber for 1 hour and then at 4°C
overnight. The highest dilution of the test serum giving a
positive carpet like pattern was recorded as the end point.
Both positive and negative controls were used along
with the test samples.

34
TREATMENT OF INFECTED ANIMALS
5 mg/kg body weight of Chloroquine was injected to
all experimentally infected monkeys before the animals
reached the stage of maximum parasitaemia.

RESULTS

35
Blood films made from peripheral blood of P. knowlesi
infected Rhesus monkeys showed the presence of parasites on
the 7th day following intravenous inoculation of
parasitized monkey erythrocytes. The parasitaemia reached
to a maximum count of about 80% on the 12th day. In the
maximum diseased stage the animals became critically ill
showing hyperthermia; these animals became weak, pale and
refused all kinds of food. The animals were treated before
they had maximum parasitaemia and the parasites were not
seen in the peripheral blood after 2 days of treatment.
Out of the 50 clinically diagnosed malaria patients
21 (^2%) had a positive blood film and 29 (58%) had a
negative blood film. When the IHA test was performed on
the 50 clinically diagnosed patients, 17 (81%) out of 21
blood film positive patients gave a positive IHA test and
21 (73%) out of the 29 blood film negative patients gave a
positive IHA test (Tableil). The 25 normal control
patients, which were all blood film negative, showed a
positive IHA test in only 1 (4%) patient (Table:II).
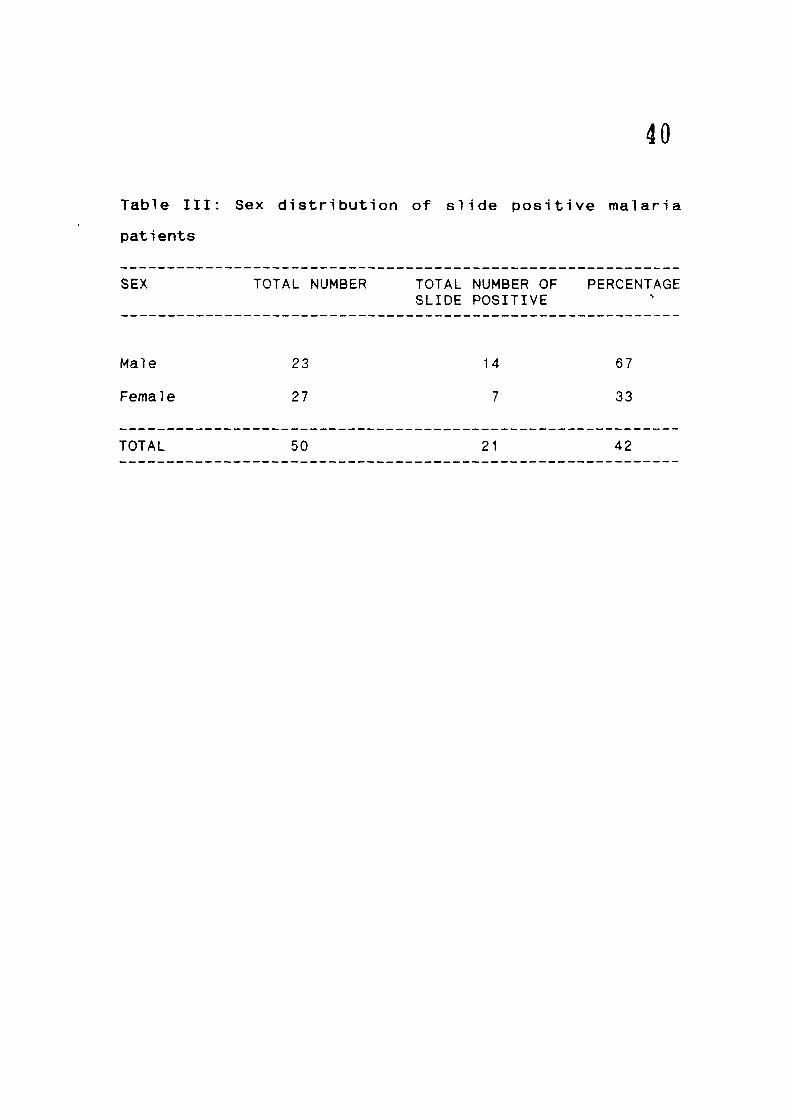
The 50 malaria patients consisted of 23 males and 27
females: the total number of blood smear positive were 14

36
(67X) and 7 (33%) f o r ma les and f e m a l e s r e s p e c t i v e l y
f T a b l e : I I I ) .
Of t h e 21 s l i d e p o s i t i v e p a t i e n t s 18 (865K) we re
i n f e c t e d w i t h P. v i v a x and 3 (14%) were i n f e c t e d w i t h
P. f a l c i p a r u m ( T a b l e : I V ) .

37
Table I: Seropositivity of malaria antibodies in clinically
diagnosed malaria patients using indirect haemagglutination
test (IHA)
GROUPS TOTAL NUMBER TOTAL NUMBER PERCENTAGE POSITIVE BY IHA
Blood film positive
Blood film negative
21
29
17
21
81
73
Total 50 38 76

FIGURE 1: SEROPOSITIVITY OF MALARIA SPECIFIC ANTIBODIES BY IHA TESTS IN THE BLOOD FILM POSITIVE & NEGATIVE CASES
Total Number
38
(H
* *
Total number of blood film positive cases.
Total number of IHA oositlve cases in the blood film positive group.
Total number of blood film negative cases.
Total number of IHA positive cases in the blood film negative group.

39
Table II: Seropositivity of malaria antibodies in normal
control patients using IHA test
GROUP TOTAL NUMBER TOTAL NUMBER PERCENTAGE POSITIVE BY IHA
Normal 25 Control
40
Table III: Sex distribution of s l i d e p o s i t i v e m a l a r i a
patients
SEX TOTAL NUMBER TOTAL NUMBER OF PERCENTAGE SLIDE POSITIVE
Male 23 14 6 7
Female 27 7 33
TOTAL 50 21 42

FIGURE 2 : Photograph of Indirect Haemagglutination test

41
t » • • • • ^̂ r -2 . ^ %'%%%: .^
'̂9 • # # '̂ • -̂ '• ' ' • # # # # # - ^ •
r### « . . . . ^ • • • # # • • •
? • # # • •
^ # • • • • • • •
c • • * 4 • • • • ^ • • • • • • • «
- ^
^>
- >
, ^
, •
» 1
• )

42

43
Table IV: Distribution of Plasmodium vivax and Plasmodium
falciparum in slide positive malaria patients
TOTAL NUMBER NUMBER OF NUMBER OF POSITIVE P. vivax P. falciparum
21 18 3
PERCENTAGE 86 14

44
FIGURE 4: DISTRIBUTION OF P. VIVAX AND P. FALCIPARUM IN BLOOD FILM POSITIVE
MALARIA PATIENTS
p. vivax 86%
P. falciparum 14%
...J

45
Table V: Distribution of IHA titres in malaria patients
GROUPS TOTAL TOTAL INDIRECT HAEMAGGLUTINATION TITRES NUMBER NUMBER TESTED POSITIVE < 16 16-64 128-512 > 1024
(> 16)
Blood Smear 21 17 4 6 10 1 Positive
Blood Smear 29 21 8 13 8 0 Negative
Control 25 1 24 1 0 0

FIGURE 6: FREQUENCY OF IHA TITRES IN MALARIA CASES AND CONTROL
) 1024
C. IHA t i t res in blood film negative cases.
B. IHA t i t res in blood film positive cases.
A. THA t i t res in Controls.

DISCUSSION AND CONCLUSION

47
Until some time back the only source of malaria
parasites was the infected host; however, the development
of continuous j_n vitro culture of P. falciparum has lead to
a large-scale production of infected blood stages in vitro.
Malaria parasites have various degrees of host
specificity and pathogenicity which affect the level of
parasitaemia obtained in any species. The degree of
infection with a given parasite may be affected by factors
such as age, sex and genetic strain of the host (Zuckerman
and Yoeli, 1954; Eling et al. , 1977; Miller 1976). Blood
collected from infected animals or humans which is to be
used for harvesting Plasmodia, may be subjected to certain
preliminary treatments to remove unwanted blood components
and to increase the proportion of parasitized erythrocytes.
Plasma is removed by centrifugation and washing, the buffy
coat is removed from the packed red cell mass to reduce the
number of platelets and leukocytes (Zuckerman et, al . .
1967). Leukocytes may be removed by suspending the washed
blood cells in several volumes of dextran solution or
obtaining them as supernatant on sedimentation (Zuckerman
et aj.. , 1967, Langer et aj.. , 1967). Sedimentation velocity
of washed blood cells on dextran (Levy and Chow, 1973) or

on sucrose density gradient (Williamson and Cover, 1966) or
on Ficol1-Hypaque solution which usually removes about 75^
leukocytes (Wallach and Conley, 1977). The slow speed
centrifugation, which we used in our experiment, removes
about 85* leukocytes. Homewood and Neame M976) reported
that the most efficient procedure for removing leukocytes
is the passage of diluted blood through a column of filter-
paper powder.
P. knowlesi infection in the Rhesus monkey (Macaca
mulatta) follows a characteristically rapid and fatal
course resulting in the death of host by day 10 post-
inoculation (Dutta et al., 1981). While running a typical
course of infection, it was revealed in differential
parasite counts that the infection in Rhesus monkey is
rather synchronous at a particular point and at this time
the parasites mostly appear in the same stage of
development (Khanna, 1984).
Resurgence of malaria has currently made us realise
the importance of developing some reliable serological
tests. Serology is an important epidemiological tool along
with blood slide examination, as malaria cannot always be
detected by direct microscopic examination of blood film.
Detection of antibody to malarial parasites can be useful
in the diagnosis of clinical cases as well as screening for

49
disease c a r r i e r s , mainly in n o n - e n d e m i c a r e a s , and in
epidemiological studies (Agarwal et aj.. , 1982; Biswas et.
a1 •, 1990). The microscopic examination of blood slides is
not the only parasitologic l a b o r a t o r y p r o c e d u r e used in
malaria surveillance work to determine the occurrence of
the disease. This method can only indicate the presence or
absence of patent parasitaemia at the time of examination;
it does not indicate the individual's malaria experience or
the state of his immunity CKagan, 1972).
Serologic tests are needed to detect infections in
those cases in which the parasites in the peripheral blood
are in such low numbers that they cannot be detected
readily by parasitological examinations. We need serology
to determine whether eradication measures have been
successful, that is, whether transmission of malaria in
endemic areas has been interrupted. Moreover, a proper
surveillance needs to be maintained in those regions in
which malaria has been successfully eradicated in order to
prevent reintroduction of the parasite, which could bring
about devastating epidemics in populations rendered
susceptible by the absence of infection (Sadun, 1972). A
practical serologic method for use in malaria programs must
meet the following criteria, (a) the test has to be simple
to perform; (D) the interpretation of results must be free

50
of subjectivity; (c) the test must be rapid; (d) the cost
must be minimal; (e) the tests results have to be
sufficiently sensitive and specific, and (f) the test must
be capable of producing repeatedly reliable results in
various laboratories. The indirect haemagglutination test
meets these criteria better than other available serologic
methods like indirect fluorescent antibody test and the
complement fixation test (Kagan, 1972).
The IHA test may have a wide range of applications in
epidemiology. For example it may be used (1) to measure the
level of malaria endemicity (Kagan et al. , 1969a); (2) to
verify the absence or presence of transmission from a
relatively small number of specimens (Mathews et. al . ,
1970a); (3) to delineate malarious areas (Mathews et al . .
1970b); (4) to detect seasonal changes of malaria
transmission (Kagan et aj.. , 1969b); (5) to investigate
suspected reintroductions of malaria into consolidation or
maintenance phase areas; and (6) to detect small foci of
malaria transmission (Mathews ei al., 1970b). The test is
especially useful in areas where the regular malaria
surveillance work is limited, and it can be used
effectively as a screening mechanism to detect potential
carriers of malaria.

51
P. falciparum antigen obtained from infected Actus
monkeys (Voller et al-, 1976; Mahajan, et al-, 1981), or
from i_n vi tro cultures (Spencer e_t_ aj_.,1981) has been
successfully used for diagnosis of malaria by the IHA test
(Farshy and Kagan, 1972; Wilson ejt aj.. , 1975). Due to
limitations in the availability of adequate amount of
P. faici parum antigen in a sufficiently purified form,
efforts are being made in several laboratories to evaluate
the possibility of using P. knowlesi antigen for diagnosis
of human malaria (Chandanani et aJ.-. 1981; Agarwal et al . .
1981). This Plasmodium species was selected because Rhesus
monkeys can be readily infected and can provide a large
volume of parasitized red blood cells for obtaining antigen
to be used for detection of P_. f al ci parum or P.. v i v a x
infections (Kagan, 1972; Agarwal et al-, 1983).
Our results show that 81% of the slide positive
malaria patients obtained a significant antibody titre of
2 1:16. This finding compares well with the of Agarwal et,
al (1981, 1982) and Marwah et. aJ.. (1989); who also showed
75%, 84.5% and 78% of significant antibody titres in slide
positive malaria patients respectively. The slide negative
patients showed a significant antibody titre in 73%, this
seems to be in contrast to Marwah et, aj_ (1989) and Agarwal
et al (1982) who showed 32% and 7.14% positive antibody

52
titres. The reason for this difference could be because
most of the patients attending J.N. Medical College
Hospital come from places in and around Aligarh which are
at some distance from Aligarh and as such they do not come
back again for slide parasitology; drugs are given to them
on the basis of clinical symptoms of malaria irrespective
of a negative laboratory diagnosis. The slides are
generally made at a time when the parasites in the
peripheral blood are in low numbers and as such the
parasites are not easily detected. Differences in the
method for preparations of P_. knowl es i antigen and the
storage conditions may explain the low titres reported by
Agarwal et al (1982) and Marwah et al (1989).
The sera specimens from the 25 healthy control group
were also subjected to IHA tests. Only 1 (4^) sample gave
an antibody titre of >_ 1:16, this is in accordance with the
results of Agarwal et aj. (1981; 1982) who showed positive
IHA values in 4% and ^ . 5% respectively. A positive IHA
result in 15.5* of normal healthy subjects has also been
reported by Srivastava et aJ. (1983).
In this study the sera were collected from 23 males
and 27 females and out of these 61% males and 335̂ females
were found to have a positive blood film. Out of the 50
slides made from the blood of clinically diagnosed malaria

53
patients only 21 slides showed malaria parasites giving a
slide positivity rate of 42%: this is a little lower than
that reoorted by Chowdhury ejt a_i (1983) who reported a
slide positivity rate of 67.5%. This could be because the
samples were collected from February to September and the
highest incidence of malaria being from September to
November only, and seemingly as such, the tip of the
iceberg was reached (Chowdhury et al., 1983).
Of the 21 slide positives 18 were of P. vivax and 3
of P. f alci parum; this can be compared to the work of
Marwah et al (1989) who also showed a higher incidence of
P. vivax (94X). But these findings are in contrast with a
study done in Senegal where P. falciparum was found to be
the predominant species (Vercruysse et aj.. , 1983).
Our results show that the IHA antibody titres of
slide positive malaria patients ranged from l 1:16
to 2 1:1024 with the highest number of positives having
titres between 1:128 to 1:512. In the slide negative
malaria patients most of the subjects having a positive IHA
antibody titre ranged from 1:16 to 1:64 titre. The 4 of
the slide positive sera gave a titre <_ 16, such a low titre
could be attributed to the fact that IHA test is rather
insensitive at detecting antibodies in the early phases of

54
primary infection. No slide negative patient had a
titre >. 1024 (Marwah et al-. 1989).
On the basis of this investigation and in the light
of other similar studies it can be concluded that antigen
prepared from £. knowlesi parasites can be successfully
employed in the IHA test for detecting malaria specific
human antibodies. An IHA positivity of 81% has been seen
in the present study and this is in agreement with the
results reported by other workers using heterologous
ant i gen (Lobel e_t al . , 1973; Agarwal ejtaj.., 1981).
Apparently, a high degree of specificity resides in the
test as was evident from the very low positivity (4%)
obtained among normal healthy controls. An IHA positivity
of 73% in patients with slide negative malaria suggest that
a small number of such persons may indeed be suffering from
malaria. If serology is to be employed, as an adjunct to
microscopic detection of parasites, the, the diagnosis of
clinical cases of malaria on the basis of IHA antibody
titres can be conveniently carried out on more or less a
routine basis.

BIBLIOGRAPHY

55 BIBLIOGRAPHY
A g a r w a l , S . S . , N a t h , A . , Sha rma , P . , S r i v a s t a v a , I . K . ,
D w i v e d i , S .R. and D u t t a . G .P . ( 1 9 8 3 ) . " C o m p a r a t i v e
e v a l u a t i o n o f P l a s m o d i um k n o w ! e s i and PI asmod i um
c v n o m o 1 q i a n t i g e n s i n t h e I n d i r e c t f l u o r e s c e n t
a n t i b o d y t e s t f o r human m a l a r i a . " I n d . J . Med. Res.
7 7 : 6 1 6 .
Agarwa l , S . S . , N a t h , A . , Sha rma , P . , S r i v a s t a v a , I . K . ,
D w i v e d i , S . R . , Yadava, R.L. and D u t t a , G.P. ( 1 9 8 2 ) .
" S e r o e p i d e m i o l o g y o f human m a l a r i a : I n d i r e c t
h a e m a g g l u t i n a t i o n u s i n g Plasmodium k n o w l e s i a n t i g e n . "
I n d . J . M a l a r i o l . 1 9 : 2 1 .
A g a r w a l , S . S . , Sharma, P . , N a t h , A . , S r i v a s t a v a , I . K . , R a i ,
J . and D u t t a , G.P. ( 1 9 8 1 ) . " I n d i r e c t h a e m a g g l u t i n a t i o n
and I n d i r e c t f l u o r e s c e n t a n t i b o d y t e s t s f o r human
m a l a r i a u s i n g PIasmodi um know les i a n t i g e n . " I n d . J .
M a l a r i o l . 1 8 : 6 7 .
A i k a w a , M . , M i l l e r , L . H . , J o h n s o n , J . and R a b b e g e , J .
( 1 9 7 8 ) . " E r y t h r o c y t e e n t r y by m a l a r i a l p a r a s i t e s : A
moving j u n c t i o n between e r y t h r o c y t e and p a r a s i t e . " J .
C e l l . B i o l . 7 7 : 7 2 .
A l l i s o n , A . C . ( 1 9 8 4 ) . " C e l l u l a r i m m u n i t y t o m a l a r i a and
babes ia p a r a s i t e s : A p e r s o n a l v i e w p o i n t . " Contemp.
Top. Immunob io l . 12: 463.

56
Anders, R.F. (1986). "Multiple cross-reactivities amongst
antigens of PI asmod i urn fa 1c i parum impair the
development of protective immunity against malaria."
Para. Immunol. 8: 529.
Ayala, S.C. (1977). "In Parasitic Protozoa, Kreir, J.P.
(ed.) vol.3, Academic Press, New York: 267.
Barnwell, J.W., Nichols, M.E. and Rubenstein, P. (1989).
"In vitro evaluation of the rate of duffy blood group
in erythrocyte invasion by Plasmodium vivax." J. Exp.
Med. 169: 1795.
Bayoumi, R.A.L. (1987). "The sickle-cell trait modifies
the intensity and specificity of the immune response
against PI asmodi um falci parum malaria and leads to
acquired protective immunity." Med. Hypotheses. 22:
287.
Bayoumi, R.A.L., Abu Zaid, Y.A., Abdulhadi, N.H., Theander.
T.G., Hviid, L., Saeed, B.O.,Ghalib, H.W., Nugud.
A.H.D., Jepsen, J.B.and Jepsen, S. (1990). "Cell
mediated immune responses to PI asmod i um falci parum
purified soluble antigens in Sickle-cell trait
subjects." Immunol. Letter. 25: 243.
Bennett, I.L. and Hook, E.W. (1959). "Infectious diseases
(Some aspects of Salmonellosis)." Ann. Rev. Med. 16:
1 .

57
Biswas , S . , Saxena, O.B. and Roy. A. ( 1 9 9 0 ) . "The n a t u r a l
o c c u r r e n c e o f c i r c u l a t i n g a n t i b o d i e s i n p o p u l a t i o n s o f
endemic m a l a r i o u s a r e a s . " I n d . J . M a l a r i o l . 27: 139.
Boyd, M . F . ( e d . ) f 1 9 4 9 ) . I n M a l a r i o l o g y , S a u n d e r s ,
P h i l a d e l p h i a , P e n n s y l v a n i a .
Brown, J . and S m a l l e y , M.E. ( 1 9 8 1 ) . " I n h i b i t i o n o f t h e i n
v i t r o g r o w t h o f P1asmod i urn f a l c i p a r u m by human
p o l y m o r p h o n u c l e a r n e u t r o p h i l l e u k o c y t e s . " C l i n . Exp.
Immunol . '46 : 106 .
B r o w n , K . M . a n d K r e i e r , J . P . ( 1 9 8 6 ) . " E f f e c t o f
m a c r o p h a g e - a c t i v a t i o n on P h a g o c y t e - P l a s m o d i u r n
i n t e r a c t i o n . " I n f e c t . Immum. 5 1 : 744.
B r u c e - C h w a t t , L . J . ( 1 9 6 3 ) . "A L o n g i t u d i n a l s u r v e y o f
n a t u r a l m a l a r i a i n f e c t i o n i n a g roup o f West A f r i c a n
a d u l t s . " W. A f r . Med. J . 12: 141-173 ; 199-217.
B u t c h e r , G . A . , M i t c h e l , G . H . and C o h e n , S. ( 1 9 7 3 ) .
" M e c h a n i s m s o f h o s t s p e c i f i c i t y i n m a l a r i a l
i n f e c t i o n . " N a t u r e , 2 4 4 : 4 0 .
C e l a d a . A . , C r u c h a n d , A. and P e r r i n , L . H . ( 1 9 8 3 ) .
" P h a g o c y t o s i s o f P lasmod ium f a l c i p a r u m p a r a s i t i z e d
e r y t h r o c y t e s by human p o l y m o r p h o n u c l e a r l e u k o c y t e s . "
J . P a r a s i t o l . 69 : 49 .
Chandanan i , R . E . . Maha.jan, R .C . . P rasad , R.N. and Gangu ly ,
N.K. ( 1 9 8 1 ) . " E v a l u a t i o n o f P I a s m o d i urn k n o w l e s i

58
antigen for serodiagnosis of human malaria infection."
Ind. J. Med. Res. 73:41.
Cherfas, J. (1990). "Malaria research - What Next?" Science
247:399.
Choudhury. D.S., Mai hot ra, M.S... Shukla, R.P., Ghosh, S.K.
and Sharma, V.P. (1983). "Resurgence of malaria in
Gadarpur PHC, District Nainital, Uttar Pradesh." Ind.
J. Malariol. 20: 49.
Clark, I.A., Allison, A.G. and Cox, F.E.G. (1976).
"Protection of mice against Babesia and Plasmodium
with BCG". Nature (London) 259:309.
Clark, I.A., Chaudhari, G. and Cowden, W.B. (1989). "Roles
of tumour necrosis factor in the illness and pathology
of malaria." Trans. Roy. Trop. Med. Hyg. 83:436.
Clark, I.A., Cox, F.E.G. and Allison, A.C. (1977).
"Protection of mice against Babesia spp. and
Plasmodium Spp. with killed Corynebacterium parvum."
Parasitology 74:9.
Cohen, S. and Butcher, G.A. (1971). "Serum antibody in
acquired malarial immunity." Trans. Roy Soc. Trop.
Med. Hyg. 65: 125.
Colbourne. M.J. (1955). "Malaria in Gold Coast students on
their return from the United Kingdom". Trans. Roy.
Soc. Trop. Med. Hyg. 49: 483.

59
D u b o i s , P . . D a r d e n n e , M . , F a n d e u r . T . , M e r c e r e a u -
P u i j a l o n , 0 . , M a t t e l , D. , M u H e r - H i l K B . , B l i s n i c k ,
T. and P e r e i r a de S i l v a , L. ( 1 9 8 8 ) . " S t r u c t u r e and
f u n c t i o n o f a t h y m i c p e p t i d e i s mimicked by Plasmodium
f a 1 c i p a r u m p e p t i d e s . " Ann . I n s t . P a s t e u r / I m m u n o l .
139: 557.
D u t t a , G . P . , S i n g h , P.P. and Sa ibaba , P. ( 1 9 8 1 ) . " P r e s b v t i s
e n t e l 1 us as a new h o s t f o r e x p e r i m e n t a l P I a s m o d i urn
k n o w l e s i i n f e c t i o n . " I n d . J . Med. Res. 73: 63 .
E l i n g , W. , Van Z o u , A. and J e r u s a l e m , C. ( 1 9 7 7 ) . "The
c o u r s e o f a P I a s m o d i urn b e r q h e i i n f e c t i o n i n s i x
d i f f e r e n t mouse s t r a i n s . " 2 . P a r a s i t e n k d . 54: 24 .
F a r s h y , D.C. and Kagan , I . G . ( 1 9 7 2 ) . "Use o f s t a b l e
s e n s i t i z e d c e l l s i n i n d i r e c t m i c r o h a e m a g g l u t i n a t i o n
t e s t f o r m a l a r i a . " Am. J . T rop . Med. Hyg. 2 1 : 868.
Fen ton , B . , Wa l ke r , A. and W a l l i k e r , D. ( 1 9 8 5 ) . " P r o t e i n
v a r i a t i o n i n c l o n e s o f Plasmodium f a l c i p a r u m d e t e c t e d
by two d i m e n s i o n a l e l e c t r o p h o r e s i s . " Mol . Biochem.
P a r a s i t o l . 16: 173.
Garnham, P.C.C. (1966). " M a l a r i a p a r a s i t e s and o t h e r
h a e m o s p o r i d i a . B l a c k w e l l , O x f o r d c i t e d i n Wa l t e r H.
W e r n s d o r f e r ( 1 9 8 0 ) . I m p o r t a n c e o f M a l a r i a i n t h e
w o r l d . " I n M a l a r i a v o l . 1 , K r e i e r , J . P . ( e d . ) .
Academic P r e s s , New Y o r k . London, 7 1 .

60
Garnham, P.C.C. (1980). "Malaria in its various vertebrate
hosts". In "Malaria." vol.1, Kreier, J.P. (ed.).
Academic Press, New York, 95.
Gerhardt, G. (1884). "Uber Intermittensimpfungen", Z. Klin.
Med. 7: 372.
Ghatak, S.N.. Bhattarchar.i i , S. and Dhar, M.M. (1987).
"CDRI Researches on Parasitic Diseases." Malaria:
157.
Gilles. H.M. (1961). "The natural history of stable
"falciparum malaria in the Preschool Child." W. Afr.
J. Med. J. 10:293.
Grau, Q.E., Fajardo, L.F., Piquet, P.F., Allet, B.,
Lambert, P.H. and Vassali, P. (1987). "Tumour necrosis
factor (cachectin) as an essential mediator in murine
cerebral malaria." Science 237: 1210.
Greenwood. B.M., Byass, P., Greenwood, A.M., Hayes, R.J.,
Menon, A., Shenton, F.C., Stephens, J. and Snow, R.W.
f1989). "Lack of association between acute
gastroenteritis, acute respiratory infections and
malaria in young Gambian children." Trans. Roy. Soc.
Trop. Med. Hyg. 83:595.
Grun, J.L., Long, C.A. and Weidanz, W.P. (1985). "Effects
of splenectomy on antibody independent immunity to
Plasmodium chabaudi malaria." Infect. Immun. 48:853.

61
Harte, P.G.. Rogers, N.C. and Targett. G.A.T. (1985).
"Role of T-cells in preventing transmission of rodent
malaria." Immunol. 56: 1.
Homewood. C.A. and Neame, K.D. (1976). "A comparison of
methods used for the removal of white cells from
malaria infected blood." Ann. Trop. Med. Parasitol.
70:249.
Homme 1, M., David. P.H. and Oligino, L.D. (1983). "Surface
alterations of erythrocytes in Plasmodium falciparum
malaria. Antigenic variation, antigenic diversity and
the role of spleen." J. Exp. Med. 157: 1137.
Honigberg, B.M., Balamuth, W. , Bovee, E.C., Corliss. J.O.,
Go.idics, M.. Hall, R.P., Kudo, R.R.. Levine, N.D.,
Loeblich. A.R. Jr., Weiser, J. and Wenrich, D.H.
(1964). "A revised classification of the Phylum
Protozoa." J. Protozool. 11: 7.
Jacobsen, P.H., Hviid, L., Theander, T.G., Riley, E.M.,
Grellier, P., Bruun, L., Dalsgaard, K., Schrevel , J.
and Jepsen, S. (1990). "Isolation and character
ization of a soluble complex of Plasmodium falciparum
with pyrogenic properties." APMIS. 99: 21.
Joklik, K.W., Willett, P.H. and Amos. B.D. (eds.) (1984).
"Malaria" In Zinsser Microbiology, (18th ed.).
Appleton-Century-Crofts/Norwalk , Connecticut, USA.
1223.

62
Kagan. I.G. (1972). "Evaluation of the indirect
haemagqlutination test as an epidemiologic technique
for malaria." Am. J. Trop. Med. Hyg. 21(5): 683.
Kagan, I.G.. Mathews, H.M., Rogers, W.A. Jr. and Fried.
J.A. (1969a). "Seroepidemiologic studies by indirect
haemagglutination tests for malaria. Military recruit
collections from Argentina, Brazil, Columbia and the
United States of America." Bull. W.H.O. 41:825.
Kagan, I.G., Mathews, H.M. and Sulzer, A.J. (1969b). "The
serology of malaria; Recent applications." Bull. N.Y.
Acad. Med. 45: 1027.
Kaslow, D.C., Quakyi, I.A., Syin, C , Raum, M.G., Keister,
D.B., Coligan, J.E., McCutchan, T.F. and Miller. L.H.
(1988). "A vaccine candidate from sexual stages of
human malaria that contains EGF-like domains." Nature
333: 74.
Kemp, D.J., Coppel, R.L. and Anders, R.F. (1987).
"Repetitive proteins and genes of malaria." Ann. Rev.
Microbiol. 41: 181.
Khanna, R. (1984). "Plasmodium knowlesi infection of Rhesus
monkeys - Comparison of certain parasitological and
haematological parameters in pre and post-infected
animals." M.Phil Thesis. Aligarh Muslim University.
Aliqarh. 70.

63
K h a r a z m i . A . , H o i b y , N. and T h e a n d e r , T . G . ( 1 9 8 7 ) .
"Pseudomonas a e r u g i n o s a s e p t i c a e m i a i n a p a t i e n t w i t h
s e v e r e PI asmodi urn f a 1 c i pa rum m a l a r i a . " T r a n s . Roy .
Soc. T r o p . Med. Hyg. 8 1 : 49 .
K r u s m i t h . S. and D r u i l h e , P. ( 1 9 8 3 ) . " A n t i b o d y dependent
i n g e s t i o n o f PIasmodium f a l c i p a r u m m e r o z o i t e s by human
b l o o d m o n o c y t e s . " P a r a s i t e Immunol . 5: 357.
Langer , B.W., P h i s p h u m v i d h i , P. and F r i e d l a n d e r , Y. ( 1 9 6 7 ) .
"MaTar ia p a r a s i t e m e t a b o l i s m : The pen tose c y c l e i n
Plasmodium b e r g h e i . " Exp. P a r a s i t o l . 20 : 68 .
Levy. M.R. and Chow, S . C . ( 1 9 7 3 ) . " A c t i v i t y and some
p r o p e r t i e s o f an a c i d p r o t e i n a s e f r o m n o r m a l and
Plasmodium be rghe i i n f e c t e d red c e l l s . " J . P a r a s i t o l
59: 1064.
L o b e l . H . O . , M a t h e w s , H . M . and K a g a n . I . G . ( 1 9 7 3 ) .
" I n t e r p r e t a t i o n o f IHA t i t r e s f o r t h e s t u d y o f m a l a r i a
e p i d e m i o l o g y . " B u l l . W.H.O. 4 9 : 4 8 5 .
Lowry . D . H . , R o s e n b r o u g h , N . J . , F a r r , A . L . and R a n d a l l .
R . J . ( 1 9 5 1 ) . " P r o t e i n measurement w i t h f o l i n phenol
r e a g e n t . " J . B i o l . Chem. 193: 265.
MacDona ld , G. ( 1 9 5 7 ) . "The e p i d e m i o l o g y and c o n t r o l o f
m a l a r i a . " O x f o r d U n i v . P r e s s , London, 53 .
M a h a i a n . R . C . . G a n g u l y , N . K . , T h a d a n i . M . , G i l l N. and
C h a n d a n a n i , R.E. ( 1 9 8 1 ) . "Comparison of ELISA w i t h

6i
IHA a n d I F A t e s t s i n m a l a r i a . " I n d . J . M e d . R e s .
7 3 : 8 7 4 .
M a r w a h , G . S . . A r o r a . S . a n d P r a b h a k a r , H . ( 1 9 8 9 ) .
" H a e m a g g l u t i n a t i o n a n t i b o d y t i t r e s and i m m u n o g l o b u l i n
l e v e l s i n c a s e s o f P l a s m o d i u m i n f e c t i o n . " I n d . J . Med.
R e s . 8 9 : 3 2 6 .
M a t h e w s , H . M . , F i s h e r , G . U . a n d K a g a n . I . G . ( 1 9 7 0 a ) .
" P e r s i s t e n c e o f m a l a r i a a n t i b o d y i n T o b a g o , W e s t
I n d i e s , f o l l o w i n g e r a d i c a t i o n , as m e a s u r e d by t h e
I n d i r e c t h a e m a g g l u t i n a t i o n t e s t . " Am. J . T r o p . Med.
H y g . 1 9 : 5 8 1 - 5 8 5 .
M a t h e w s , H . M . , F r i e d , J . A. and K a g a n , I . G . ( 1 9 7 0 b ) . "A
s e r o e p i d e m i o l o g i c s t u d y o f m a l a r i a i n t h e R e p u b l i c o f
P h i l i p p i n e s by t h e I n d i r e c t h a e m a g g l u t i n a t i o n t e s t . "
Am. J . E p i d e m i o l . 9 2 : 3 7 6 - 3 8 1 .
M a t h w s , H . M . , F r i e d , J . A . a n d K a g a n , I . G . ( 1 9 7 5 ) . " T h e
i n d i r e c t h a e m a g g 1 u t i n a t i o n t e s t f o r m a l a r i a .
E v a l u a t i o n o f a n t i g e n s p r e p a r e d f r o m P 1 a s m o d i urn
f a l c i p a r u m and P l a s m o d i u m v i v a x . " Am. J . T r o p . Med.
H y g . 2 4 : 4 1 7 .
M c G r e g o r . I . A . ( 1 9 6 0 ) . " D e m o g r a p h i c e f f e c t s o f m a l a r i a
w i t h s p e c i a l r e f e r e n c e t o t h e s t a b l e M a l a r i a o f
A f r i c a . " West A f r . Med. J . 9 : 2 6 0 .
M c G r e g o r . I . A . ( 1 9 8 4 ) . " E p i d e m i o l o g y , m a l a r i a a n d
p r e g n a n c y . : Am. J . T r o p . Med. H y g . 3 3 : 5 1 7 .

65
McGregor, I.A., GilTes, H.M.. Walters, J.H., Davies, A.H.
and Pearson. F.A. (1956). "Effects of heavy and
repeated malarial infections on Gambian infants and
children. Effects of erythrocytic parasitization."
Br. Med. J. 2:686.
McLaughlin, G.L., BenediK, M.J. and Campbell, G.H. (1987).
"Repeated immunogenic aminoacid sequences of
Plasmodium species share sequence homologies with
protein from human and human viruses." Am. J. Trop.
Med. Hyg. 37: 258.
Meckel, H. (1847). "Uber Schwarzer Pigment in der Milz und
in Blute einer Geisteskrauken." 2. Psychiatr. 4:198.
Meis, J.F.G.M., Jap, P.H.K., Hollingdale, M.R. and Verhave,
J.P. (1987). "Cellular responses against
exoerythrocytic forms of Plasmodium berqhei in rats."
Am. J. Trop. Med. Hyg. 37:506.
Mendis. K., Munesinghe, Y.D., Silva, N.Y., Keragalla, I.
and Carter, R. (1987). "Malaria transmission-blocking
immunity induced by natural infection of Plasmodium
vivax in humans." Infect. Immun. 55:369.
Miller, L.H. (1976). "Innate resistance in malaria." Exp.
Parasitol. 40: 132.
Miller. L.H. and Carter, R. (1976). "Innate resistance to
malaria". Expl. Parasitol. 40:132.

66
M i l l e r , L. H. .. Duorak. J . A . . S h i r o i s h i . T. and Durocher . J .
( 1 9 7 3 ) . " I n f l u e n c e o f e r y t h r o c y t e membrane components
on m a l a r i a m e r o z o i t e i n v a s i o n . " J . E x p . Med .
138 :1597 .
M i l l e r , L . H . , Mason, S . J . , C l y d e , D.F. and M c G u i n i s s , M.H.
( 1 9 7 6 ) . "The r e s i s t a n c e f a c t o r t o Plasmodium v i v a x m
b l a c k s , t h e d u f f y b l ood group g e n o t y p e , Fy Fy. "New
E n g l . J . Med. 2 9 5 : 3 0 2 .
M i l l e r , L . H . , M c G u i n i s s , M .H . , Hoe land , P.V. and Sigmon, P.
( 1 9 7 8 ) . "The d u f f y b l ood group phenotype i n Amer ican
b l a c k s i n f e c t e d w i t h P I a s m o d i urn v i vax i n V i e t n a m . "
Am. J . T r o p . Med. Hyg. 27 : 1069.
N a r d i n , E . H . , Nussenzweig , R . S . , McGregor, I . A . and Bayan,
J . H . ( 1 9 7 9 ) . " A n t i b o d i e s t o S p o r o z o i t e s : t h e i r
f r e q u e n t o c c u r r e n c e i n i n d i v i d u a l s l i v i n g i n an area
o f hyperendemic m a l a r i a . " S c i e n c e , 206: 597.
N n a l u e , N.A. and F r i e d m a n , M . J . ( 1 9 8 8 ) . " E v i d e n c e f o r a
n e u t r o p h i l - med ia ted p r o t e c t i v e response i n m a l a r i a . "
Para . Immunol . 10 :47 .
N u s s e n z w e i g , R . S . ( 1 9 6 7 ) . " I n c r e a s e d n o n - s p e c i f i c
r e s i s t a n c e t o m a l a r i a p roduced by a d m i n i s t r a t i o n o f
k i l l e d C o r y n e b a c t e r i u m p a r v u m . " E x p . Paras i tol .
2 1 : 2 2 4 .

67
Ockenhouse. C.F . and Shear . H.L . ( 1 9 8 3 ) . " M a l a r i a induced
Lymphok ines: S t i m u l a t i o n o f macrophages f o r enhanced
P h a g o c y t o s i s . " I n f e c t . Immun. 4 2 : 7 3 3 .
Ockenhouse . C . F . and S h e a r , H . L . ( 1 9 8 4 ) . " O x i d a t i v e
k i l l i n g o f t h e i n t r a e r y t h r o c y t i c m a l a r i a p a r a s i t e
P Iasmod i urn y o e l i i by a c t i v a t e d m a c r o p h a g e s . " J .
Immunol . 132 :424 .
Park , J . E . and Pa rk . E. ( 1 9 8 6 ) . " M a l a r i a " . I n Textbook o f
P r e v e n t i v e and S o c i a l M e d i c i n e . P u b ! . B a n a r s i d a s
Bhanot , 1268 Nap ie r Town, J a b a l p u r , I n d i a , 355.
Pombo, D . , M a l a y , W . L . , B e r z o f s k y , J . A . and Good , M .F .
( 1 9 8 8 ) . "Neona ta l exposure t o immunogenic p e p t i d e s .
D i f f e r e n t i a l s u s c e p t i b i l i t y t o t o l e r a n c e i n d u c t i o n o f
h e l p e r T and B - C e l l s r e a c t i v e t o m a l a r i a l
c i r c u m s p o r o z o i t e p e p t i d e e p i t o p e s . " J . Immunol . 140:
3594.
R e i n h a r d t . M.C. . Ambro ise-Thomas, P . . C a v a l 1 o - S e r r a . R.,
M e y l a n , C. and G a u t i e r . R. ( 1 9 7 8 ) . " M a l a r i a a t
d e l i v e r y i n A b i d j a n . H e l v . Paed. A c t a . 33: 65.
R i s t i c . M. . James, M.A. , Shamansky, L . , Kakoma, I . , L i u ,
H . Y . and N i c h o l s , J . ( 1 9 8 5 ) . " C u l t u r e - d e r i v e d
P l ^Smod i um f a l c i p a r u m e x o a n t i g e n s : I s o l a t i o n and
c h a r a c t e r i z a t i o n o f p r o t e c t i v e p o l y p e p t i d e s . " I n Proc.
A s i a and P a c i f i c C o n f . on M a l a r i a . S i d d i q u i , W.A.
( e d . ) , H o n o l u l u , H a w a i i . 194.

00
Sadun, E .H . ( 1 9 7 2 ) . "The r e s e a r c h and d e v e l o p m e n t o f
s e r o l o g i c t e s t s f o r m a l a r i a . " Am. J . T r o p . Med. Hyn.
21 { 5 ) : 677.
Sa lmon . D . . V i l d e . J . L . . A n d r i e u , B . , S i m o n o v i c , R. and
L e b r a s . J . ( 1 9 8 6 ) . " R o l e o f immune s e r u m and
complement i n s t i m u l a t i o n o f t h e m e t a b o l i c b u r s t o f
human n e u t r o p h i l s by P1 asmod i urn f a l c i p a r u m .
I n f e c t . I m m u n . 5 1 : 8 0 1 .
S c o t t , H.H. ( 1 9 3 9 ) . "A H i s t o r y o f t r o p i c a l m e d i c i n e . "
A r n o l d , L o n d o n . C i t e d i n W a l t h e r H. W e r n s d o r f e r
( 1 9 8 0 ) . " I m p o r t a n c e o f m a l a r i a i n W o r l d . " M a l a r i a ,
volume 1 , e d . K r e i e r , J . P . . Academic P r e s s , New York .
London.
Shamansky, L . M . . L i u . H . Y . . H a g e r , L . P . and R i s t i c , M.
( 1 9 8 5 ) . " P u r i f i c a t i o n and C h a r a c t e r i s a t i o n o f c u l t u r e
d e r i v e d e x o a n t i g e n s o f Plasmodium f a l c i p a r u m " . Mol .
B i o l . P a r a s i t o 1 . 17 :299 .
Sharma, S. and Godson, G.N. ( 1 9 8 5 ) . " E x p r e s s i o n o f the
m a j o r s u r f a c e a n t i g e n o f P1asmod i um k n o w1e s i
s p o r o z o i t e s i n Y e a s t . " Sc ience 228.
S i d d i q u i . W.A. (1991o). " R o l e o f A d j u v a n t s i n M a l a r i a
V a c c i n e s . I n I m m u n o t h e r a p e u t i c P r o s p e c t s o f
I n f e c t i o u s D i s e a s e s . M a s i h i , K . N . and L a n g e , W.
( e d s . ) . S p r i n g e r - V e r l a n B e r l i n H e i d e l b e r g . 325.

69
Siddiqui. W.A. (1991). "Where are we in the Quest for
vaccines for malaria." Drugs 41(1): 1.
Sinden, R.E. (1975). "The sporogenic cycle of Plasmodium
yoeli i ni geriensi s : A scanning electron microscope
study." Protislogica. 11:31.
Speer. C.A., Rosa!es-Ronqui11o, M.C and Silverman, P.H.
(1975). "Motility of Plasmodium berghei ookinetes jn
vitro." J. Invertrbr. Pathol. 25.
Spencer, H.C., Collins, W.E., Chin, W. and Skinner, J.C.
(1981). "The enzyme linked immunosorbent assay for
malaria III. Antibody response in documented
Plasmodium falciparum infections." Am. J. Trop. Med.
Hyg. 30:747.
Spira. D. and Zuckerman, A. (1962). "Antigenic structure of
Plasmodium vinckei." Science 137: 536.
Srivastava, I.K., Sharma, P., Nath, A., Agarwal, S.S. and
Dutta, G.P. (1983). "Enzyme linked immunosorbent
assay test with P1asmodi um know!es i antigen in
diagnosis of malaria." Ind. J. Med. Res. 77: 431.
Strickland, G.T. and Nussenzweig, R.S. (1982). In
"Immunoparasitology Principles and Methods in Malaria
and Schisto somiasis Research." Strickland and Hunter.
K.W. Jr. (eds.), Prager, 39.

70
T h e a n d e r . T . G . ( 1 9 9 2 ) . " D e f e n c e m e c h a n i s m s and immune
e v a s i o n i n t h e i n t e r p l a y b e t w e e n t h e humane immune
sys tem and Plasmodium f a l c i p a r u m . " Dan ish Med. B u l l .
39M ) : 49 .
V e r c r u y s s e . J . , J a n c l o e s , M. and V e l d e r , V . ( 1 9 8 3 ) .
" E p i d e m i o l o g y o f s e a s o n a l f a l c i p a r u m m a l a r i a i n an
urban area o f S e n e g a l . " B u l l . W ld . H l t h . Orgn . 6 1 ( 5 ) :
821 .
V o l l e r , A. ( 1 9 7 1 ) . "The d e t e c t i o n and m e a s u r e m e n t o f
m a l a r i a l a n t i b o d i e s . " T r a n s . Roy . S o c . T r o p . Med.
Hyg. 65: 111 .
V o l l e r , A . . B a r t l e t t , A. and B i d w e l l , D.E. ( 1 9 7 6 ) . "Enzyme
immunoassays f o r p a r a s i t i c d i s e a s e s . " T r a n s . Roy .
Soc. T rop . Med. Hyg. 7 0 : 9 8 .
W a l l a c h , D .F .H . and Con ley , M. ( 1 9 7 7 ) . " A l t e r e d membrane
p r o t e i n s o f monkey e r y t h r o c y t e s i n f e c t e d w i t h s i m i a n
m a l a r i a . " J . M o l . Med. 2 : 119.
Wate rs , L . S . , Tave rne , J . , T a i . P-C, Sp ry . C . J . F . , T a r g e t t ,
G . A . T . and P l a y f l a i r . J . H . ( 1 9 8 7 ) . " K i l l i n g o f
P I a s m o d i um f a l c i p a r u m by e o s i n o p h i l s e c r e t o r y
p r o d u c t . " I n f e c t . Immun. 5 5 : 8 7 7 .
W e a t h e r a l l , D . J . ( 1 9 8 7 ) . "Common g e n e t i c d i s o r d e r s o f t h e
red c e l l and t h e m a l a r i a h y p o t h e s i s . " Ann. T rop . Med.
P a r a s i t o l . 8 1 : 539.

71
Weidanz, W.P. (1982). "Malaria and alterations of immune
reactivity." Brit. Med. J. 38: 167.
Wellde, B.T., Diggs, C.L. and Anderson, S. (1979).
"Immunization of Aotus trivi rqatus against Plasmodium
falciparum with irradiated blood forms." Bull.of WHO
57(1 ) : 153.
Wellde, B.T. and Sadun, E.H. (1967). "Resistance produced
in rats and mice by exposure to irradiated Plasmodi urn
berqhei." Exp. Parastiol. 21:310.
W.H.O. (1985). "Report on the joint meeting on the advanced
development of malaria vaccines." Geneva, Feb. 1985.
W.H.O. (1986). "Principles of malaria vaccine trials:
Memorandum from WHO meeting." Bull, W.H.O. 64(2):
185.
W.H.O. (1990). "WHO News and Activities". Bull. W.H.O.
68(5): 667.
Williamson. J. and Cover, B. (1966). "Separation of blood
cell free trypanosomes and malaria parasites on a
sucrose gradient." Trans. Roy. Soc. Trop. Med. Hyg.
60:425.
Wilson. M.. Fife. E.H., Mathews, H.M. and Sulzer, A.J.
(1975). "Comparison of the complement fixation,
indirect haemagglutination and indirect immuno-

72
fluorescence tests for malaria." Am. J. Trop. Med.
Hyg. 24: 755.
Zuckerman. A. (1966). "Recent studies on factors involved
in malaria anaemia." Mil. Med. 131: 1201-1216.
Zuckerman. A., Spira. D. and Hamburger, J. (1967). "A
procedure for the harvesting of mammalian Plasmodia."
Bui 1. W.H.O. 37: 431 .
Zuckerman, A. and Yoeli, M. (1954). "Age and sex factors
influencing Plasmodium berghei infections in intact
and splenectomized rats." J. Infec. Dis. 94:255.





























