Embed Size (px)
Citation preview

Human Cancer Biology
Detection and Characterization of a Novel Subset ofCD8þCD57þ T Cells in Metastatic Melanoma with anIncompletely Differentiated Phenotype
Richard C. Wu1,4,5, Shujuan Liu1, Jessica A. Chacon1,4, Sheng Wu1, Yufeng Li1,4, Pariya Sukhumalchandra3,James L. Murray2, Jeffrey J. Molldrem3, Patrick Hwu1,4, Hanspeter Pircher6, Gregory Liz�ee1,4, andLaszlo G. Radvanyi1,2,4
AbstractPurpose: Tumor-specific T cells are frequently induced naturally in melanoma patients and infiltrate
tumors. It is enigmatic why these patients fail to experience tumor regression. Given that CD8þ T cells
mediate antigen-specific killing of tumor cells, the focus of this study was to identify alterations in the
differentiation of CD8þ residing at the tumor site, with emphasis on a population expressing CD57, a
marker for terminal differentiation.
Experimental Design:We conducted flow cytometric analysis of CD8þ tumor-infiltrating lymphocytes
(TIL) isolated from 44 resected melanoma metastases with known T-cell differentiation markers. For
comparison, peripheral blood mononuclear cells were isolated from matched melanoma patients. We
sorted different CD8þ subsets found in TIL and determined their effector functions. In addition, we carried
out Vb clonotype expression analysis of T-cell receptors to determine lineage relationship between theCD8þ
TIL subsets.
Results: Themajority of CD8þ TIL was in the early-effectormemory stage of differentiation. A significant
population consisted of an oligoclonal subset of cells coexpressing CD27, CD28, CD57, and Granzyme B,
with little or no perforin. These cells could be induced to proliferate, produce a high level of IFN-g , anddifferentiate into CD27�CD57þ, perforinhigh mature CTL in vitro. Addition of TGF-b1 prevented further
differentiation.
Conclusions: Our studies identified a novel subset of incompletely differentiated CD8þ CTL coexpres-
sing early effector memory and late CTL markers. This population resembles that found in patients with
uncontrolled chronic viral infections. TGF-b1, frequently produced by melanoma tumors, may be a key
cytokine inhibiting further maturation of this subset. Clin Cancer Res; 18(9); 2465–77. �2012 AACR.
IntroductionCD8þCTLsare critical effector cells inantitumor responses
knowntomediate theantigen-specific cytolysisof tumorcellsthrough the action of granzymes, such as granzyme B (GB),
and the pore-forming protein perforin (Perf). Terminallydifferentiated effector (TTDE) CD8þCD57þCTL, which high-ly expresses both GB and Perf, have been identified as themost potent cells for combating chronical viral infection aswell as inducing effective cytotoxic killing of tumor cells (1–6). Most healthy adults contain a distinct population oflong-lived CMV-specific CD8þCD57þCD27�CD28�GBhigh
Perfhigh TTDE lymphocytes detectable in the peripheral bloodthat exhibit spontaneous antiviral CTL activity (5–8). Dif-ferentiation of CD57þ TTDE CTL fromCD27þ precursors hasbeen shown to be critical for maintaining a pool of matureCTL in the blood, which controls CMV and EBV infections inhumans (4, 8–11). However, differentiation toward suchTTDE phenotype can be altered in cases of chronic antigenstimulation, as has been reported for individuals infectedwithhuman immunodeficiencyvirus (HIV)andEpstein-Barrvirus (EBV) reactivation (7, 12), inwhich anaccumulation ofincompletely differentiated CD8þCD27þCD57þ CTL leadsto a loss of control of viral replication (8).
In cancer patients, the role of the CD8þCD57þ T cells inthe control of tumor growth has not been clearly defined
Authors' Affiliations: Departments of 1Melanoma Medical Oncology,2Breast Medical Oncology, and 3Stem Cell Transplantation, University ofTexas, MD Anderson Cancer Center; 4Graduate Program in Immunology,University of Texas Graduate School of Biomedical Sciences; 5M.D./Ph.DProgram, University of Texas Medical School at Houston, Houston, Texas;and 6Department of Immunology, Institute of Medical Microbiology andHygiene, University of Freiburg, Freiburg, Germany
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
R.C. Wu and S. Liu contributed equally to the work.
Corresponding Author: Laszlo G. Radvanyi, Department of MelanomaMedical Oncology, Box 904, 1515 Holcombe Blvd., Houston, TX 77030.Phone: 713-563-9489; Fax: 713-563-3424; E-mail:[email protected]
doi: 10.1158/1078-0432.CCR-11-2034
�2012 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org 2465
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

(13). It was previously reported that in malignant melano-ma, renal cell carcinoma, and gastric carcinoma, patientswho harbored higher percentages of CD8þCD57þ lympho-cytes in peripheral blood had shorter overall survival (14–16). However, in those studies, the exact substage of dif-ferentiation of the CD8þCD57þ T lymphocytes was notfurther analyzed. CD8þCD57þ T cells in the peripheralblood of advanced gastric carcinoma patients have beenshown to have lower Perf content and diminished IFN-gproduction as compared with healthy individuals (17). Onthe contrary, CD8þCD57þ cells with a TTDE phenotype(CD27�CD28�GBhighPerfhigh) accumulate in the peripher-al blood of normal aged individuals and are the main IFN-g–producing CD8þ T lymphocyte subset (18). They havealso been positively associated with potent antitumor effec-tor function and improved relapse-free survival in leukemiapatients after stem-cell transplant (1, 19).
Little is known about the frequency and function of theCD8þCD57þ TTDE CTL population in tumor-infiltratinglymphocytes (TIL) in solid tumors, such as melanoma orbreast cancer. Recently, a distinct population of CD8þ CTLin human peripheral blood with fully mature TTDE charac-teristics (CD27�CD28�GBhighPerfhigh) was found to expressa natural killer (NK) cell marker, CD56 (5, 6, 20).CD8þCD56þTTDECTL (CD3þ) thatdidnot coexpressCD57exhibited an even higher spontaneous cytolytic activity thanCD57þ CTL against tumor targets; they also showed higherPerf andGB expression and could reexpress CD45RA (20). Itis currently not known whether CD8þ TILs preferentiallydifferentiate to become CD56 or CD57-expressing CTL.
We undertook a detailed analysis of the phenotypicdifferentiation of CD8þCD57þ CTL in 44 melanoma and5 breast cancer metastases with multicolor flow cytometry.
Previous studies focused exclusively on T cells infiltrates inlymph mode (LN) metastases (21). In our studies, weincluded a large number of non-LN visceral metastases toexclude any possible contribution by nontumor specific,resident LN immune cells. This is important because mel-anoma patients usually die from visceral, non-LN metasta-ses. We did not find an appreciable population ofCD8þCD56þ CTL in freshly isolated melanoma TIL, butfrequently observed a significant population of CD57þ Tcells. Moreover, we found that the majority of the CD57þ
TIL in melanoma and breast cancer pleural effusions alsocoexpressedCD27andCD28,markers of effectormemory Tcells (TEM; refs. 12, 22, 23), with moderate GB expressionand low to absent Perf expression, similar to theCD8þCD27þCD57þ T-cell population that was recentlydescribed in HIV-infected patients (8). TheseCD8þCD27þCD57þ T cells could be induced to furtherdifferentiate into CD27�CD57þ, Perfhigh mature CTL aftershort-term T-cell receptor (TCR) stimulation and exhibitenhanced cytotoxic capacity. In addition, we show thatfurther differentiation of this lymphocyte subset could beinhibited by TGF-b1, an immunosuppressive cytokineknown to be produced by many types of tumor cells,including melanoma (24–27). These results suggest thatCD8þ TIL show aberrant differentiation along the CD57þ
CTL lineage, an effect that may result from suppressivefactors found within the tumor microenvironment.
Materials and MethodsReagents
Flow cytometry antibodies were purchased from BDBiosciences, eBioscience, or Beckman Coulter. Humanrecombinant interleukin (IL)-2 (ProLeukin) was a generousgift fromNovartis. KLRG1-Alexa 488monoclonal antibody(mAb) was a gift from Dr. H. Pircher (Freiburg, Germany).
Patient TIL and peripheral blood mononuclear cellsamples
TIL and blood samples for laboratory studies wereobtained from patients with stages IIIc to IV melanomawho were undergoing surgery at The University of TexasMD Anderson Cancer Center according to an InstitutionalReview Board (IRB)–approved protocol and patient con-sent. The method for fresh isolation of TILs from mela-noma metastases is described in detail online in Supple-mentary Methods. Typically 0.5 � 106 to 5 � 106 lym-phocytes were isolated initially from tumor fragments,depending on the size of tumors. The dividing TIL lineswere cultured in high-dose (HD) IL-2 as described previ-ously (28). Peripheral blood mononuclear cells (PBMC)were isolated by density gradient separation with Ficoll-Isopaque from normal donors from buffy coats obtainedfrom the Gulf Coast Regional Blood Center or peripheralblood from stage IV melanoma patients collected in hepar-izined tubes (BD Biosciences). The PBMCs were cryopre-served in 10% dimethyl sulfoxide, 90% human AB serumuntil analysis.
Translational RelevanceUncovering how CD8þ differentiation and effector
function can be altered in the tumor microenvironmentis critical to understand why most tumors continue toprogress, despite being infiltrated with CD8þ T cells.Although other investigators have examined CTL differ-entiation in lymph node (LN) metastases in melanoma,it is less clear whether the differentiation of the CD8þ
CTL is also defective in non-LN, visceral metastases. Inthis study, we elucidated the state of CD8þ CTL differ-entiation in LN and non-LN melanoma metastases andbreast cancer pleural effusions. We discovered a novelpopulation of incompletely differentiated CD8þ tumor-infiltrating lymphocytes. We found that TGF-b1, a keyimmunosuppressive cytokine in the tumor microenvi-ronment, could inhibit further differentiation of thesecells. These results suggest that CD8þdifferentiation intopotent CTL is altered in cancer. Thus, modifying thisaltered differentiation in the tumor microenvironmentshould also be the goal of current immunotherapeuticapproaches.
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2466
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

Pleural effusions were also collected from metastaticbreast cancer patients under an IRB-approved protocol.Lymphocytes in pleural effusions were isolated by centri-fugation over 70% and 100% Ficoll-Isopaque double layersto separate tumor cells from lymphocytes. The lymphocyteswere collected at the 100% Ficoll-Isopaque layer andwashed in TIL culture medium (TIL-CM; Gemini Bio Pro-ducts, Sacramento, CA) before flow cytometry staining.
Surface and intracellular staining by flow cytometryFreshly isolated TILs and TILs cultured and expanded
with IL-2, as well as thawed normal donor or patientPBMCs, were washed twice in Dulbecco’s PBS (D-PBS) andstained with LIVE/DEAD Fixable Aqua Dead Cell Stain Kit(Life Technologies) according to manufacturer’s instruc-tion. Cells were thenwashed twice in fluorescence-activatedcell sorting (FACS) wash buffer [D-PBSþ 1% bovine serumalbumin (BSA)] and stained for 30 minutes at 4�C withantibodies conjugatedwith fluorochromes against differentT-cell cell surface and intracellular markers. In the cases ofHLA-A2.1þ patients, the TIL suspensions were additionallystained with MART-1 peptide (ELAGIGILTV) HLA-A2.1tetramer, gp100peptide (IMDQVPFSV)HLA-A2.1 tetramer,or control HLA-A2.1 tetramer containing HIV gag peptide(SLYNTVATL; Beckman Coulter). The cells were finallyfixed in D-PBS, 1% paraformaldehyde and 0.25% ethanol.Intracellular staining for GB and Perf was done by accord-ing to manufacturer’s protocol (BD Biosciences). Flowcytometric analysis was conducted with an FACSCanto IIflow cytometer (Becton Dickinson). The positive and neg-ative regions of the staining of the indicated surface mar-kers were determined by comparing against the unstainedsamples (Supplementary Fig. S1B). Data were analyzed byFACSDiva (BD Biosciences) or FlowJo software (Tree StarInc.). Proliferation was assessed by intracellular stainingfor Ki-67 with an anti–Ki-67-APC antibody (BDBiosciences).
Sorting and 3H-thymidine incorporation assayIL-2–cultured TILs were stained with anti–CD8-Pacific
Blue, anti–CD27-APC-Cy7, and anti-CD57-FITC (FITC,fluorescein isothiocyanate) in PBS containing 1% BSAand 5% goat serum. The CD8þCD27þCD57� andCD8þCD27þCD57þ subsets were sorted by an InFlux cellsorter (BD Biosciences). A total of 5 � 104 viable cells perwell were plated into 96-well Costar 3361 High-bind plates(Sigma-Aldrich) precoated overnight with anti-CD3(OKT3; Ortho Biotech) or anti-CD3 and anti-CD28(eBioscience) agonistic antibodies. After 3 days, the cellswere pulsed with 1 mCi of [3H]-thymidine (methyl-T-thy-midine; PerkinElmer Inc.) for 18 hours. The incorporated[3H] thymidine was determined as counts perminute by a bliquid scintillation counter (Beckman Coulter).
Human Th1/Th2 multiplex cytokine analysisThe tissue culture supernatants collected in triplicate
from sorted TIL subsets under unstimulated and stimulatedconditions were collected and plated on the multiplex
ELISA plate configured to detect a panel of human Th1/Th2 cytokines (IFN-g , IL-2, IL-4, IL-5, IL-10, IL-12 p70, andIL-13), according to the manufacturer’s instructions (MesoScaleDiscovery). The signals were captured and analyzed bythe SECTOR Imager 2400 (Meso Scale Discovery). Theconcentration of each cytokine was calculated from its eachrespective standard curves.
Differentiation assay in vitroCD8þ TILs were sorted into subsets as described previ-
ously and stimulated with anti-CD3 or anti-CD3 and anti-CD28 antibodies precoated onNunc plates (Thermo FisherScientific). IL-2 (200 IU/mL) was added to each culture tomaintain cell viability. Human TGF-b1 (R&D Systems) wasused at 1 ng/mL for the TGF-b1–treated group. After 7 days,the cells were stained for CD8, CD27, CD28, CD56, andCD57. Intracellular staining for GB and Perf was done asdescribed above.
Redirected cytotoxic T-cell assayAnalysis of CTL activity on the sorted CD8þ subsets was
done according to a novel flow cytometric method mea-suring the cleavage of caspase-3 in anti-CD3 coated targetcells as described previously (29). Briefly, 5 � 106 murinemastocytoma target cells (P815)were labeledwith a fixable,far red fluorescent tracer, CellTrace Far Red DDAO-Succi-nimidyl Ester (DDAO-SE; Invitrogen) according to manu-facturer’s instructions, washed, resuspended at a density of2.5�106/mL, andpulsedwith 200mg/mLof anti-CD3mAbin a low serum–containing media (RPMI-1640 with 2%FBS) at room temperature for 30 minutes. Unpulsed targetcells served as controls. Labeled, pulsed P815 target cellswere added to the sorted TIL subsets at effector:target (E:T)ratios of 1:1 and 3:1, or 1:10 and 1:20 in a round bottom96-well plate and spun down for 5 minutes at low centri-fuge speed (300 rpm) to maintain optimal contactbetween target cells and effector T cells. The cells werecoincubated for 3 hours before harvesting. The 3-hour timepoint was chosen to prevent the apoptotic target cells frombecoming necrotic and losing the cleaved caspase-3 signal.The cells were stained intracellularly with an anti-cleavedcaspase-3-PE mAb (BD Biosciences). Target cells weredistinguished from effector T cells by the far red tracerDDAO-SE (which fluoresce in the APC channel), and theextent of the caspase-3 cleavage in the target cell popula-tion was analyzed by the FACSanto II flow cytometer (BDBiosciences).
Statistical analysisGraphPad Prism version 5.0 (GraphPad Software) was
used for graphing and statistical analysis. Either the 2-tailed,paired Student’s t test or the Wilcoxon rank-sum test wasused to analyze the statistical significance in differencesbetween 2 groups, and 2-tailed nonparametric Kruskal–Wallis test followed by Dunn multiple comparison testwere used for more than 2 sample groups. P � 0.05 wasdeemed to be statistically significant.
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2467
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

ResultsPatient tumor samples and experimental approach
In this study, tumors were surgically excised from stagesIII/IIIc and IV (M1a–M1c) melanoma patients as part of anongoing adoptive T-cell therapy clinical trial at MD Ander-son Cancer Center. Supplementary Table S1 shows thecharacteristics of the melanoma patients, including patientage, sex, tumor location, disease stage, and anatomical sites.The tumors were processed immediately after surgery formulticolor flow cytometry staining and expansion of TIL.The panels of fluorochrome-conjugated antibodies used inall experiments and their clone numbers are shown inSupplementary Table S2.
Presence of a CD8þCD57þ subset highly positive forCD27 and CD28 and absence of CD8þCD56þ T cells inTILs freshly isolated from melanoma tumors
As CD8þ T cells transition from TEM into fully differen-tiated (TTDE) CTL, they generally acquire GB and Perfexpression, downregulate CD27 and CD28, and expressCD57, or in some cases, CD56. This TTDE phenotype(CD27�CD28�CD56þ or CD27�CD28�CD57þ) is associ-ated with immediate cytolytic activity, with no requirementfor antigenic restimulation to induce CTL function
(3, 6, 20, 23). Todetermine the extent of the TTDE phenotypein tumors, we first gated on the live CD8þ lymphocytes, asdetermined by the Aqua Live/Dead exclusion dye (Invitro-gen), on TIL freshly isolated from tumors (SupplementaryFig. S1). We then analyzed them for CD4, CD8, CD27,CD28, CCR7, and CD45RA expressions with multicolorflow cytometry. A majority of the bulk as well as tumorantigen–specific CD8þ TIL from melanoma tumors dis-played a TEM phenotype (CD27þ, CD28þ, CCR7�,CD45RA�; Supplementary Figs. S3 and S4; refs. 22, 23).The specificity of the MART-1 and gp100 tetramer stainingwas verified against a negative control, HIV gag tetramer(data not shown) as well as comparing against HLA-A2�
patients (Supplementary Fig. S2). We then stained the TILsfor CD8, CD27, CD28, CD56, and CD57 and analyzed therelationship between end-stage CTL markers (CD57 andCD56) versus TEM markers (CD27 and CD28). We foundthat a significant population of CD57 single, positive cellswas seen without CD56 expression (see Fig. 1A for arepresentative example). Figure 1B shows this predomi-nance of CD8þCD57þ cells (mean percentage � SEM ¼16.2 � 3.5) over CD8þCD56þ cells (mean percentage �SEM¼ 1.0� 0.6; pairedWilcoxon rank-sum test; P < 0.005)in the CD8þ subset in freshly isolated TIL for 10 separate
A
CD56
34
CD8
039
061
TIL #2156
SS
C
CD
57
C TIL #2156
44
49
CD8
5
CD27
2
74
20
18
792
1
CD27
TIL #2276
Gated on
Live CD8+ TIL
% o
f C
D8
+ T
-cell s
ub
set 50
40
30
20
10
0
56+
57+
4
93
CD8
90
12
7
CD27
93
2
6
0
CD27
CD
57
CD
57
CD
28 C
D28 C
D28
CD
28
P < 0.005
B
Figure 1. Lack of CD56-expressing end-stage CTL and the appearance of a novel subset of CD8þCD57þ melanoma TILs coexpressing CD27 and CD28. A,fresh TIL isolates were stained with a panel of fluorochrome-conjugated antibodies for CD4, CD8, CD27, CD28, CD56, and CD57. Results were shown bygating on the live cells that were determined by the Live/Dead Fixable Aqua Dead Cell Stain Kit (Life Technologies). A minimum of 100,000 gated light scatterevents per sample were acquired on the flow cytometer. CD56 and CD57 expression in the CD8þ subset in a representative patient TIL sample is shown. B, asummary of the CD8þCD56þ and CD8þCD57þ populations in 10 freshly isolated TILs from melanoma tumors. Significant difference was calculated by a 2-tailed, paired Wilcoxon rank-sum test. C, the gated CD8þCD57þ and CD8þCD57� subpopulations were further analyzed separately for CD27 versus CD28expression. Examples of TILs from 2 melanoma patient tumor samples are shown.
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2468
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

patient samples. Upon further analysis of the stainingprofiles from the freshly isolated TIL samples, we foundthat many CD8þCD57þ T cells coexpressed CD27 andCD28, which could be considered as a hybrid of late-stageCTLandearly TEMmarkers (Fig. 1C).We also found that PD-1, a marker for exhausted CD8þ T cells or TEM in humans(30, 31, Supplementary ref. 1), was more enriched in theCD27þCD57þ than the CD27þCD57� subset (Supplemen-tary Fig. S5A; Kruskal–Wallis test, P < 0.05), in the tumorsfrom 10 additional metastatic melanoma patients (Supple-mentary Fig. S5B), which suggests that the CD27þCD57þ
"hybrid" phenotype may represent a more end-stage effec-tor phenotype. We also stained PBMCs from patients and
normal donors to determine the extent of CD27þCD28þ
CD8þCD57þ T cells. Comparing TILs and PBMCs from thesame patient, we found that the CD8þ T cells in the PBMCshad only a few (<5%) of CD57þ T cells coexpressing CD27and CD28, while almost 60% of the CD8þCD57þ TILcoexpressed CD27 and CD28 (Fig. 2A). Similarly, only afew (<5%) of the CD8þCD57þ in PBMCs of normal donorscoexpressed CD27 and CD28 (Fig. 2A). In contrast, mela-noma TILs contained, on average, more than 20% of theCD8þCD57þ subset coexpressing CD27 and CD28 whenthe results of a larger number of patient samples wereanalyzed (Fig. 2B). An examination of the melanoma anti-gen-specific CD8þ subsets (MART-1 and gp100) in fresh TIL
BA TIL #2163
CD
57 16
CD
28
19 60
1110 50
60
50
60
CD8+, CD27+
60
CD8+, CD28+ CD8+, CD27+, CD28+CD8+, CD27+
P = 0.0007 50
CD8+, CD28+
P = 0.0006
CD8+, CD27+, CD28+
P = 0.0004
CD8 CD27
Pt. PBMC #2163
D28
41
D5
7
30
20
30
40
10
20
30
40
20
30
40
CD
CD2776
ND PBMC #1
CD8
CD
20
10
0
10 10
0 0TIL ND
PBMCPatient
PBMCTIL ND
PBMCPatient
PBMCTIL ND
PBMCPatient
PBMC
53
CD
57
0 3
1977
CD
28
C3
CD
27
92 8
00
TIL#2058
AR
T-1
tetr
am
er+
ND PBMC #2
CD8 CD27
35
D57
1 2
D28
00
0.7
CD8
0 t
etr
am
er+
CD
27
CD57
TIL#2276
84 14
MA
83CD8
CD
14
CD27
CD
CD8
gp
10
CD57
TIL#2276
2 0
% C
D57 s
ub
set
Figure 2. Relatively few fully differentiated CD8þCD27�CD28�CD57þ TILs were found in metastatic melanomas compared with peripheral blood of patientsand normal donors. TILs isolated from melanoma metastases and PBMCs from stage III/IV melanoma patients were stained in a panel of fluorochrome-conjugated antibodies against CD4, CD8, CD27, CD28,CD56, andCD57 (same as in Fig. 1). Aminimumof 100,000 gated light scatter events per samplewereacquired on the flow cytometer. PBMCs from 2 normal donors were similarly stained for comparison. Results were shown by gating on the CD8þ livelymphocytes thatwere determined by the Live/DeadFixable AquaDeadCell Stain Kit (Life Technologies) and analyzed for CD27andCD28expressions. A, theresults from 1 representative TIL isolate from tumor #2163, 1 representative patient PBMC sample, and 2 normal donors. B, analysis of the CD57þ subsetcoexpressingCD27andCD28 in theCD8þT-cell population in a larger group ofmelanomaTIL samples (n¼18),melanomapatient PBMC (n¼12), andnormaldonor PBMC (n¼ 6) samples. The percentage of gated CD8þCD57þ subset coexpressing CD27, CD28, or both, was plotted for each sample type. Each dotrepresents a single samplewith bars indicating the averages for each sample group.Results of theStudent t test between the TIL andpatient PBMCare shownfor each subset (P < 0.05 indicates statistical significance). C, melanoma metastases from HLA-A2.1þ patients were stained with antibodies for the samemarkers as in A together with tetramer containing either a human HLA-A2.1 MART-1 epitope or the human gp100 epitope. A minimum of 100,000 gated lightscatter per samplewere acquired on the flowcytometer. TheCD8þT-cell subset was gated and analyzed forMART-1 or gp100 tetramerþ cells. The tetramerþ
cells were then gated and analyzed for CD27 and CD57 expression. Shown are 2 examples of the analysis done with TILs from 2 different patient metastases(#2058 and #2276).
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2469
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

isolates by tetramer staining revealed these tetramerþ cellswere either CD27þCD57� or CD27þCD57þ, with a negli-gible fraction of cells having a CD27�CD57þ orCD57�CD27� phenotype (Fig. 2C).
CD8þCD57þCD27þ melanoma TILs are GBþ,but Perf�/low
We next determined the intracellular GB and Perf expres-sion, markers used to identify end-stage (TTDE) CTL, inCD8þCD27þCD57þ TIL in melanoma. We also obtainedpleural effusions from newly diagnosed metastatic breastcancer patients to determine whether CD8þCD57þ coex-pressing CD27 and CD28were also found in other forms ofcancer, and what their GB and Perf expression might be.First, inmelanoma TILs, we found that themajority of gatedCD8þCD27þCD57þ and CD8þCD27þCD57� cellsexpressed GB, but most of the cells in both subsets hadlow or negative Perf expression (Fig. 3A). Using the samegating approach, we analyzed fresh melanoma TIL samplesfrom 7 stage IVmelanoma patients, PBMC samples from 17stage IV melanoma patients, and PBMC from 6 normaldonors. Shown in Fig. 3B was an example of the analysis
that compared CD8þCD57þ T lymphocytes’ GB and Perfcontents in a patient’s TIL against the same patient’s PBMCand that of a normal donor. In Fig. 3C, 48.0% � 8.6%(mean� SEM) of CD8þCD27þCD57þ TILs from 7 patientswere found to express GB, whereas only 20.5% � 6.0%expressed Perf. In contrast, averages of 94% � 1.7% and80.9% � 4.2%, of the CD8þCD57þ lymphocytes in theperipheral blood of 17melanoma patients were positive forGB and Perf, respectively. The difference was statisticallysignificant for both GB and Perf (P < 0.001, respectively;Kruskal–Wallis test) When we analyzed lymphocytes iso-lated from breast cancer pleural effusions, we also foundthatmanyCD8þCD57þ cells coexpressed CD27 andCD28,and that these cells were predominantly GBþwith very littleor no Perf expression (Supplementary Fig. S6).
CD8þCD27þCD57þ TIL persist in culture with IL-2, canbe induced to enter cell cycle, and are potent IFN-gproducers after TCR stimulation
Several studies have shown thatCD8þCD57þ cells donotdivide appreciably, or at all, in response to TCR stimulation,indicating that they are senescent cells (9, 32). We therefore
B
ND PBMC
A
CD57
CD57
CD27
58
33
870
0 13
46
54
0
0
CD57
0
0 19
81
12 0
88 0
CD57
TIL #2163
0
0
CD57
26
74
CD5797
30
0
CD57
95
4
Pt. PBMC #2163
CD57
91
8
97
3
CD57
11
89
GB Perf
C
ns
P < 0.05
P < 0.001
P < 0.05
P < 0.001
ns
CD
57
GB
Per
f
GB
Per
f
GB
GB
GB
Per
fP
erf
Per
f
TIL Pt.PBMC
NDPBMC
TIL Pt.PBMC
NDPBMC
CD57
100
80
60
40
20
0% o
f C
D8+
T-c
ell s
ub
set
Figure 3. CD8þCD57þ subset in themelanomamicroenvironment has a uniqueGBþPerf�/low phenotype comparedwith the same subset in patient and normaldonor PBMCs. A, TIL isolates were stained for CD8, CD27, CD28, CD57, GB, and Perf expression, as described in Materials and Methods. A minimum of100,000 gated light scatter events per samplewere acquired on the flowcytometer. Resultswere shownby gating on the live cells thatwere determined by theLive/Dead Fixable Aqua Dead Cell Stain Kit. The gated CD8þCD57þ and CD8þCD57� subpopulations were further analyzed separately for GB and Perfexpression. One representativemelanomaTIL sample is shown. B, results comparingGB andPerf expression in theCD8þCD57þTIL andCD8þCD57þT cellsin PBMC from the same patient (#2163) and in a representative normal donor. C, summary of GB and Perf expression in CD8þCD57þ TIL subsets from anumber of melanomametastases (n¼ 7) compared with PBMCs of patients (n¼ 17) and PBMCs from a group of normal donors (n¼ 16). The percentages ofCD8þCD57þT cells expressingGBor Perf are shown. Each dot represents a single samplewith the bars indicating the averages andSEM for each TIL'sCD8þ
subset. Significant differences between the groups were calculated by the 2-tailed, nonparametric Kruskal–Wallis test followed by the Dunn multiplecomparison test. ns, not significant.
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2470
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

asked whether the CD8þCD27þCD57þ cells found in mel-anoma TIL were capable of further proliferation and cyto-kine production in the presence of IL-2. Culturing of iso-lated melanoma TILs in culture medium with HD IL-2(3,000 IU/mL) is a standard method to expand TILs foradoptive cell therapy (28, 33). Routinely, we find that a 2-week culture period of isolated TILs with IL-2 expands theCD8þ T cells between 200- and 300-fold (data not shown).We stained 2-week IL-2–expanded TIL for CD8, CD27,CD57, andPerf. The percentage of theCD27þCD57þ subsetincreased from 4% to 20% of the gated CD8þ subset (seeSupplementary Fig. S7A). A small percentage of cells(�10%) in both CD27þCD57þ and CD27þCD57� subsetsexpressed another marker for senescence and terminaldifferentiation, KLRG-1 (Data not shown; refs. 34, 35). Tofurther examine the proliferative capacity of these cells, westained the cultured TIL with Ki-67 at the end of the 2-weekculture period and found that both CD8þCD27þCD57þ
and CD8þCD27þCD57� subsets contained a significantfrequency of Ki-67þ cells (Supplementary Fig. S7B).We then asked whether the CD8þCD27þCD57þ TILs
could enter cell cycle and produce cytokines after TCRstimulation. Two-week IL-2–cultured TILs were isolated,washed, and then recultured with low-dose (LD) IL-2(200 IU/mL) alone (to prevent apoptosis due to cytokinewithdrawal), or restimulatedwith LD IL-2 plus plate-boundanti-CD3 or anti-CD3 plus anti-CD28 mAb for 7 daysfollowed by Ki-67 staining (all CD8þCD27þCD57þ T cellswere CD28þ; see also earlier results in Fig. 1 and 2). Thegated CD8þCD27þCD57þ cells recultured with LD IL-2alone were negative for Ki-67 staining. However, a signif-icant proportion of these cells were Ki-67þ following stim-ulation with anti-CD3 or anti-CD3 and anti-CD28 (Fig.4A).The results above indicate that the CD8þCD27þCD57þ T
cells in isolated melanoma TILs have proliferative capacity,but it could also be that CD8þCD27þCD57� T cells pro-liferated and induced CD57 expression when cultured withIL-2. Thus, tomore directly examine the proliferative capac-ity of CD8þCD27þCD57þ TILs, we sorted these cells andexamined their response to TCR stimulation and IL-2. TheCD8þCD27þCD57þ and CD8þCD27þCD57� TIL subsetsfrom 4 different melanoma patients were each stimulatedwith anti-CD3 plus anti-CD28mAbs and labeledwith [3H]-thymidine after 3 days. Both sorted subsets showed amarked induction of [3H]-thymidine incorporation afterTCR stimulation, although the sorted CD8þCD27þCD57�
subset showed a higher proliferative capacity than thesorted CD8þCD27þCD57þ subset in response to CD3 andCD28 stimulation in a majority of patients (Fig. 4B).Analysis of the supernatants from a parallel set ofcultures of activated cells found that both the sortedCD8þCD27þCD57þ and CD8þCD27þCD57� TIL pro-duced Th1/Th2 cytokines (IFN-g , IL-5, and IL-13) uponTCR stimulation, with the CD27þCD57þ subset producingsignificantly more IFN-g than the CD27þCD57� subset inTIL in 3 out of the 4 patients studied (Fig. 4C; Supplemen-tary Fig. S8). Thus, the CD8þCD27þCD57þ T lymphocytes
found in melanoma TILs were capable of continued celldivision and were potent IFN-g producers. In addition, wecarried out Vb spectratyping and noted that the sortedCD8þCD27þCD57þ TILs from both the melanoma andbreast cancer samples had a striking oligoclonality in someVb families than the CD8þCD27þCD57� TIL (data notshown), similar to observations on CD8þCD57þ T cells inother pathologic settings (19, 36).
TCR stimulation drives CD8þCD27þCD57þ TILs todifferentiate into CD8þCD27�CD57þPerfhigh cells thatare inhibited by TGF-b1
Here, we asked whether CD8þCD27þCD57þ TILswere capable of further differentiation towardCD8þCD27�CD57þ CTL expressing increased Perf. Wefirst took bulk TIL in vitro and activated them in LD IL-2mediumwith anti-CD3 plus anti-CD28 and tracked the fateof the cells after 7 days by staining and analyzing for changesin CD27, CD57, and Perf expression in the CD8þ subset.This process induced further cell division (data not shown)associated with a decrease in the frequency of theCD27þCD57þ and CD27þCD57� CD8þ cells and anincrease in the percentage of the CD27�CD57þ andCD27�CD57� populations (Supplementary Fig. S9A).Both the percentage and level (MFI) of Perf expression wasalso increased in each cell subset (Supplementary Fig. S9B).To better delineate how TCR stimulation specificallyaffects the predominant CD8þCD27þCD57þ andCD8þCD27þCD57� subsets in TILs,we sorted these subsetsfrommelanomaTILs and recultured the sorted cellswith LDIL-2 or IL-2 and anti-CD3 plus anti-CD28. The postsortpurity of the 2 populationswas verified (Supplementary Fig.S10). The sorted cells were also verified to have CD3and CD28 expression, as previously shown (Fig. 1, anddata not shown). The 7-day TCR stimulation induced alarge decrease in the percentage of the sortedCD8þCD27þCD57þ cells from 72% to 37% (Fig. 5A) or66% to 6% (Fig. 5B), with a concomitant increase in thepercentage of CD8þCD27�CD57þ cells from 6% to 49%(Fig. 5A, bottom), or CD8þCD27�CD57� cells from 4% to54% in other patient (Fig. 5B, bottom).On the contrary, thesorted CD8þCD27þCD57� cells differentiated into aCD8þCD27�CD57� population with TCR stimulation(increase from 6% to 34% and 13% to 73%, respectively,in 2 different patients), although a minor subpopulationconverted into CD8þCD27þCD57þ cells in LD IL-2 culturealone (Fig. 5A and B, top).
We also tracked changes in GB and Perf expression andfound that both proteins increased in the sortedCD8þCD27þCD57þ and CD8þCD27þCD57� TIL popula-tions following TCR stimulation for 7 days, although theextent of the increase in Perf expression was greater in thesorted CD8þCD27þCD57þ subset in 3 different patients(Fig. 6A). The increase in Perf expression in both sortedsubsets led to an enhanced cytotoxic activity, in a redirectedlysis assay using anti-CD3 antibody–coated P815 targetcells (Fig. 6B). We chose the redirected lysis assay to assessthe cytotoxic activity between the TIL subsets to avoid
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2471
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034
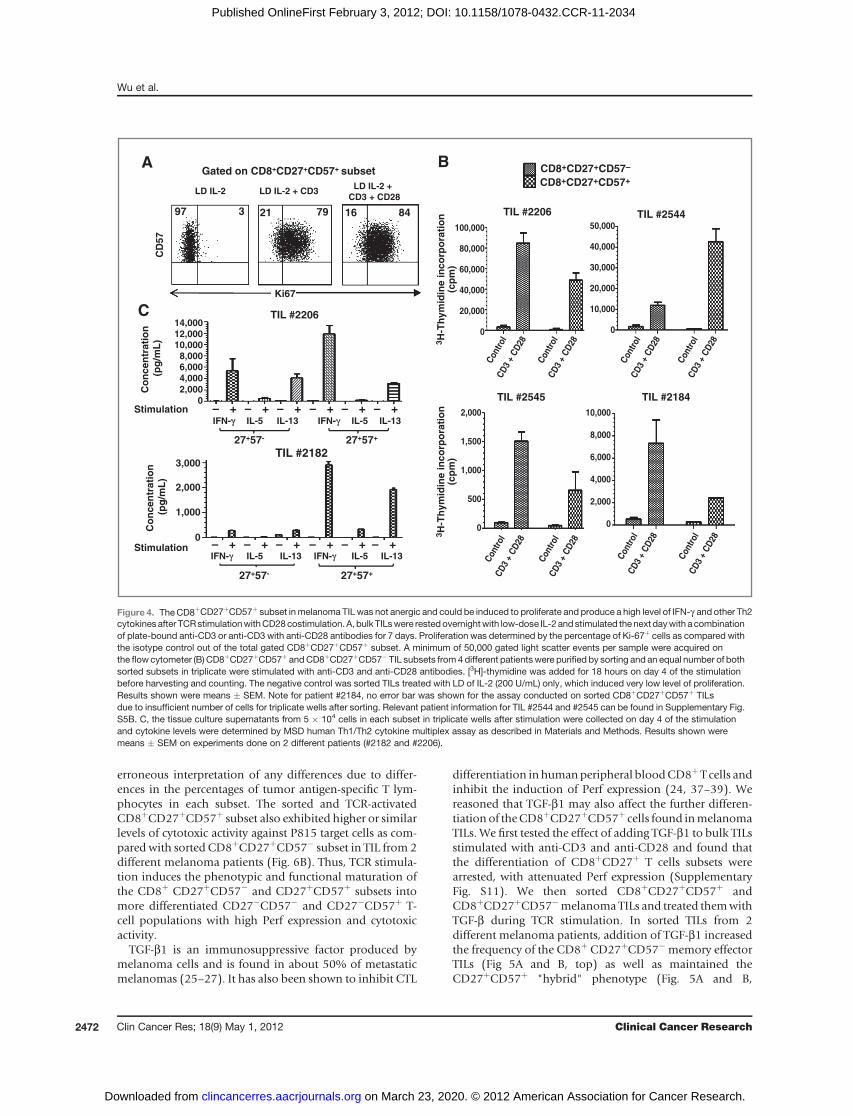
erroneous interpretation of any differences due to differ-ences in the percentages of tumor antigen-specific T lym-phocytes in each subset. The sorted and TCR-activatedCD8þCD27þCD57þ subset also exhibited higher or similarlevels of cytotoxic activity against P815 target cells as com-pared with sorted CD8þCD27þCD57� subset in TIL from 2different melanoma patients (Fig. 6B). Thus, TCR stimula-tion induces the phenotypic and functional maturation ofthe CD8þ CD27þCD57� and CD27þCD57þ subsets intomore differentiated CD27�CD57� and CD27�CD57þ T-cell populations with high Perf expression and cytotoxicactivity.
TGF-b1 is an immunosuppressive factor produced bymelanoma cells and is found in about 50% of metastaticmelanomas (25–27). It has also been shown to inhibit CTL
differentiation inhumanperipheral bloodCD8þT cells andinhibit the induction of Perf expression (24, 37–39). Wereasoned that TGF-b1 may also affect the further differen-tiation of theCD8þCD27þCD57þ cells found inmelanomaTILs. We first tested the effect of adding TGF-b1 to bulk TILsstimulated with anti-CD3 and anti-CD28 and found thatthe differentiation of CD8þCD27þ T cells subsets werearrested, with attenuated Perf expression (SupplementaryFig. S11). We then sorted CD8þCD27þCD57þ andCD8þCD27þCD57�melanoma TILs and treated themwithTGF-b during TCR stimulation. In sorted TILs from 2different melanoma patients, addition of TGF-b1 increasedthe frequency of the CD8þ CD27þCD57� memory effectorTILs (Fig 5A and B, top) as well as maintained theCD27þCD57þ "hybrid" phenotype (Fig. 5A and B,
AC
D57
Ki67
97 3 21 79 16 84
LD IL-2 LD IL-2 + CD3LD IL-2 +
CD3 + CD28
Gated on CD8+CD27+CD57+ subsetB
TIL #2206
CD8+CD27+CD57–
CD8+CD27+CD57+
TIL #2545 TIL #2184
C
27+57- 27+57+
Stimulation
IFN-γγ IL-5 IL-13 IFN-γ IL-5 IL-13
IFN-γ IL-5 IL-13 IFN-γ IL-5 IL-13
Stimulation – + – + – + – + – + – +
– +– +– +– +– +– +
3,000
14,000
12,000
10,000
8,000
6,000
4,000
2,000
0
100,000
80,000
60,000
40,000
20,000
0
2,000
1,500
1,000
500
0
10,000
8,000
6,000
4,000
2,000
0
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
Con
trol
CD
3 +
CD
28
50,000
40,000
30,000
20,000
10,000
0
3H
-Th
ym
idin
e i
nc
orp
ora
tio
n(c
pm
)
3H
-Th
ym
idin
e i
nc
orp
ora
tio
n(c
pm
)
2,000
1,000
0
27+57- 27+57+
Co
nc
en
tra
tio
n
(pg
/mL
)C
on
ce
ntr
ati
on
(pg
/mL
)
TIL #2182
TIL #2206
TIL #2544
Figure 4. TheCD8þCD27þCD57þ subset inmelanoma TIL was not anergic and could be induced to proliferate and produce a high level of IFN-g and other Th2cytokinesafter TCRstimulationwithCD28costimulation. A, bulk TILswere restedovernightwith low-dose IL-2 and stimulated thenext daywith a combinationof plate-bound anti-CD3 or anti-CD3 with anti-CD28 antibodies for 7 days. Proliferation was determined by the percentage of Ki-67þ cells as compared withthe isotype control out of the total gated CD8þCD27þCD57þ subset. A minimum of 50,000 gated light scatter events per sample were acquired onthe flow cytometer (B) CD8þCD27þCD57þ andCD8þCD27þCD57� TIL subsets from 4 different patients were purified by sorting and an equal number of bothsorted subsets in triplicate were stimulated with anti-CD3 and anti-CD28 antibodies. [3H]-thymidine was added for 18 hours on day 4 of the stimulationbefore harvesting and counting. The negative control was sorted TILs treated with LD of IL-2 (200 U/mL) only, which induced very low level of proliferation.Results shown were means � SEM. Note for patient #2184, no error bar was shown for the assay conducted on sorted CD8þCD27þCD57þ TILsdue to insufficient number of cells for triplicate wells after sorting. Relevant patient information for TIL #2544 and #2545 can be found in Supplementary Fig.S5B. C, the tissue culture supernatants from 5 � 104 cells in each subset in triplicate wells after stimulation were collected on day 4 of the stimulationand cytokine levels were determined by MSD human Th1/Th2 cytokine multiplex assay as described in Materials and Methods. Results shown weremeans � SEM on experiments done on 2 different patients (#2182 and #2206).
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2472
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

bottom), which were both associated with low GB expres-sion and more attenuated Perf expression (Fig. 6A). TGF-b1prevented the loss of CD28 expression (data not shown).TGF-b1 also strongly diminished the cytotoxic activity ofCD8þ CD27þCD57þ and CD27þCD57� TIL subsets afterstimulationwith anti-CD3and anti-CD28 for 7 days, as seenin 2 different melanoma patients (Fig. 6B). Thus, the differ-entiation of the CD27þCD57þ and CD27þCD57� CD8þ
TILs into mature CD27� GBhighPerfhigh CTLs in vitro wasblocked by TGF-b1, which resembled the phenotype of thePerflow, CD8þ TILs freshly isolated frommelanoma tumors.
DiscussionWe found that the vast majority of CD8þ T cells infiltrat-
ingmetastaticmelanomas in a large number of patients hada CD27þCD28þGBþPerf�/low phenotype reminiscent ofearly effector memory cells (7, 22, 23). Importantly, a highpercentage of the tumors analyzed were from non-LNvisceral metastases. Very few highly differentiated CD8þ
T cells that were CD27�CD28� with high Perf and GBcoexpressionwere found.Unexpectedly,manyof these highCD27-expressing CD8þ T cells also coexpressed CD57(HNK-1), a marker usually attributed to highly differenti-ated end-stage CTL with high Perf and GB levels and potentcytolytic activity (5, 6, 23, 32). A similar subset ofCD8þ cells having this unusual early-effector memoryphenotype with CD57 expression was found in pleuraleffusions of breast cancer patients. Some of theseCD8þCD27þCD28þCD57þ TILs were also specific formelanoma tumor antigens, as found by costaining withMART-1 and gp100 peptide tetramers, suggesting that
they were not merely bystander CD8þ T cells that werenot tumor specific. However, many of the otherCD8þCD27þCD28þCD57þTILsmay alsohavebeen tumorspecific, recognizing other undefined antigens; this willneed to be addressed in future studies. The appearance ofthis subset in metastatic cancer patients has striking paral-lels to a population of CD8þCD27þCD28þCD57þ T cellsfound to accumulate in the peripheral blood of patientsunable to controlHIV, EBV, orCMV infection (4, 8, 10). Thecirculating gp100 tetramerþ CD8þ T cells containing aCD27þCD57þ subset has also been observed in the periph-eral blood of early-stage melanoma patients receivinggp100 peptide vaccination (Supplementary ref. 2).Although it is important to examine how the state of CD8þ
T-cell differentiation and the appearance of thisCD8þCD27þCD28þCD57þ may change with tumor stage,our study was limited by the lack of access to early-stagemelanoma tissue samples. We did not detect appreciableFoxp3 expression in the CD8þCD27þCD57þ subset (datanot shown), indicating that these are not "regulatory"CD8þ
T cells. In addition, a study by Anichini and colleaguesfound that a subpopulation of tumor-infiltratingCD8þFoxp3þ T cells were CD57� (21).
Overall, our results suggest that CD8þCTLdifferentiationinto potent Perfhigh killer cells in the melanoma tumormicroenvironmentmay be arrested as effectormemory cells(CD8þCD27þCD28þCD57�); some of these cells begin toexpress CD57 (an end-stage CTL marker) but then fail todownmodulate CD27 and CD28 and induce Perf. Alterna-tively, it is possible that CD8þCD57þ TIL with properties ofTTDE do arise, but these cells are too short lived to be
56 25
713
18
73
2
8
52
4
2
5
4
66
25
10
54 29
6 19 32
2626
Sorted CD8+CD27+CD57- subsetB
CD57
CD
27
Sorted CD8+CD27+CD57+ subset
A
7220
2 6
37
49
7
8
4235
1112
377
20 1
CD57
Sorted CD8+CD27+CD57+ subset
Sorted CD8+CD27+CD57- subset
TGF-ββ +
CD3 + CD28
CD3 + CD28TGF-β +
CD3 + CD28
CD
27
LD IL-2
LD IL-2
LD IL-2 CD3 + CD28
CD3 + CD28TGF-β +
CD3 + CD28
TGF-β +
CD3 + CD28
TIL #2384 TIL #2541
41
1342
34 11
CD3 + CD28
2666
6 2
LD IL-2
CD57CD57
Figure 5. Sorted CD8þCD27þCD57þ T cells in TIL could further differentiate into CD27�CD57þCTL upon TCR stimulation, whichwas inhibited by the additionof TGF-b1. TILs were sorted into CD8þCD27þCD57þ and CD8þCD27þCD57� subsets as before. Postsort purity of the sorted populations is shown inSupplementaryFig. S10. A, the sortedTIL subsets (frompatient #2384)were stimulatedwith anti-CD3andanti-CD28 antibodies for 7days. Thecellswere thenrestained for CD8,CD27, andCD57 expression andanalyzed by flowcytometry. To some cultures of sorted, restimulated cells, 1 ng/mLof TGF-b1was addedonday 0withCD3 andCD28. Cellswere harvested similarly on day 7of the stimulation and stained forCD8, CD27, andCD57. Aminimumof 50,000gated lightscatter events per sample were acquired on the flow cytometer. B, similar experiment carried out on TIL from a different patient (#2541), whose relevantinformation was described in Supplementary Fig. S5B.
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2473
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034
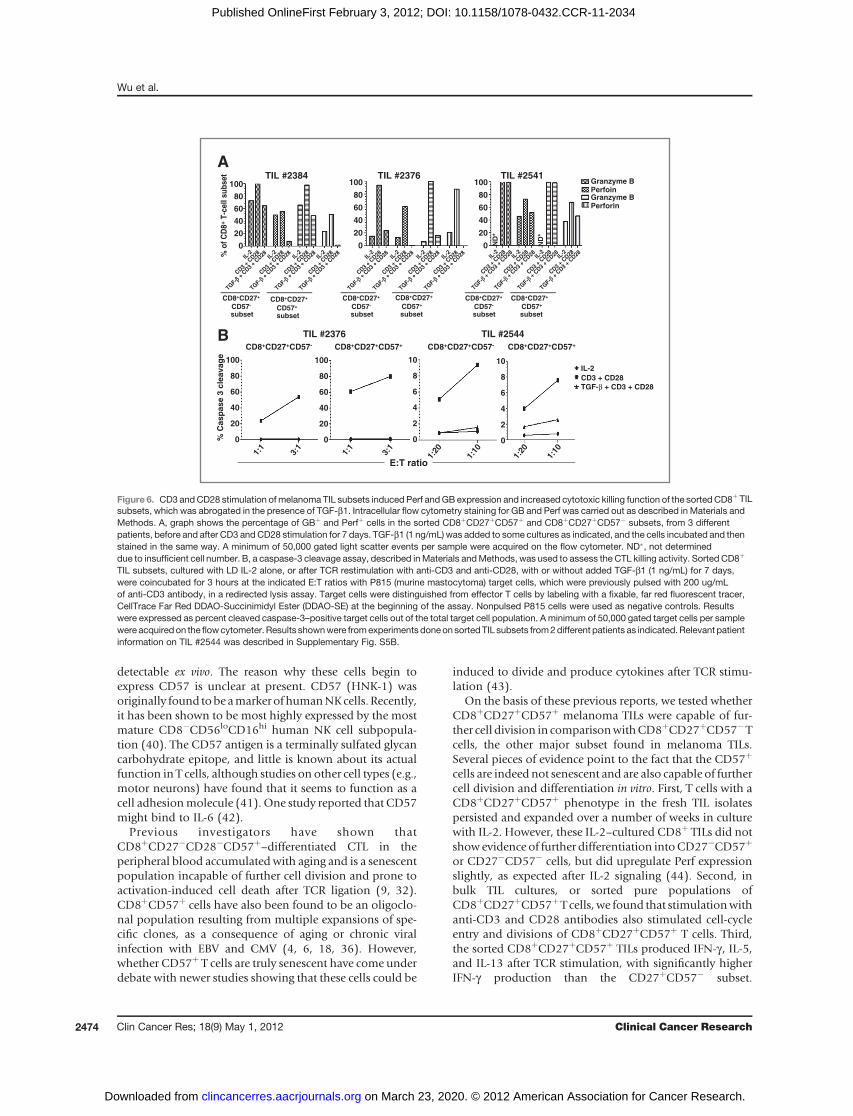
detectable ex vivo. The reason why these cells begin toexpress CD57 is unclear at present. CD57 (HNK-1) wasoriginally found tobe amarker of humanNKcells. Recently,it has been shown to be most highly expressed by the mostmature CD8�CD56loCD16hi human NK cell subpopula-tion (40). The CD57 antigen is a terminally sulfated glycancarbohydrate epitope, and little is known about its actualfunction in T cells, although studies on other cell types (e.g.,motor neurons) have found that it seems to function as acell adhesionmolecule (41). One study reported that CD57might bind to IL-6 (42).
Previous investigators have shown thatCD8þCD27�CD28�CD57þ–differentiated CTL in theperipheral blood accumulated with aging and is a senescentpopulation incapable of further cell division and prone toactivation-induced cell death after TCR ligation (9, 32).CD8þCD57þ cells have also been found to be an oligoclo-nal population resulting from multiple expansions of spe-cific clones, as a consequence of aging or chronic viralinfection with EBV and CMV (4, 6, 18, 36). However,whether CD57þ T cells are truly senescent have come underdebate with newer studies showing that these cells could be
induced to divide and produce cytokines after TCR stimu-lation (43).
On the basis of these previous reports, we tested whetherCD8þCD27þCD57þ melanoma TILs were capable of fur-ther cell division in comparisonwithCD8þCD27þCD57�Tcells, the other major subset found in melanoma TILs.Several pieces of evidence point to the fact that the CD57þ
cells are indeed not senescent and are also capable of furthercell division and differentiation in vitro. First, T cells with aCD8þCD27þCD57þ phenotype in the fresh TIL isolatespersisted and expanded over a number of weeks in culturewith IL-2. However, these IL-2–cultured CD8þ TILs did notshowevidence of further differentiation intoCD27�CD57þ
or CD27�CD57� cells, but did upregulate Perf expressionslightly, as expected after IL-2 signaling (44). Second, inbulk TIL cultures, or sorted pure populations ofCD8þCD27þCD57þT cells, we found that stimulationwithanti-CD3 and CD28 antibodies also stimulated cell-cycleentry and divisions of CD8þCD27þCD57þ T cells. Third,the sorted CD8þCD27þCD57þ TILs produced IFN-g , IL-5,and IL-13 after TCR stimulation, with significantly higherIFN-g production than the CD27þCD57� subset.
A
TIL #2376B TIL #2544
CD8+CD27+
CD57-
subset
CD8
100
80
60
40
20
0
100
80
60
40
20
0
1:1
IL-2
CD3
+ CD28
TGF-ββ
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28 IL
-2
CD3
+ CD28
TGF-β
+ CD3
+ CD28
1:1
3:1
1:20
1:10
1:20
1:103:1
100
80
60
40
20
0
10
8
6
4
2
0
10
8
6
4
2
0
100
80
60
40
20
0
100
80
60
40
20
0
Granzyme B
IL-2
CD3 + CD28
TGF-β + CD3 + CD28
PerfoinGranzyme BPerforin
+CD27+
CD57+
subset
TIL #2376
E:T ratio
% o
f C
D8
+ T
-cell s
ub
set
CD8+CD27+CD57- CD8+CD27+CD57+ CD8+CD27+CD57- CD8+CD27+CD57+
ND
*
ND
*
CD8+CD27+
CD57+
subset
CD8+CD27+
CD57-
subset
TIL #2541TIL #2384
CD8+CD27+
CD57-
subset
CD8+CD27+
CD57+
subset
% C
as
pa
se
3 c
lea
va
ge
Figure 6. CD3 and CD28 stimulation ofmelanoma TIL subsets induced Perf andGB expression and increased cytotoxic killing function of the sorted CD8þ TILsubsets, which was abrogated in the presence of TGF-b1. Intracellular flow cytometry staining for GB and Perf was carried out as described in Materials andMethods. A, graph shows the percentage of GBþ and Perfþ cells in the sorted CD8þCD27þCD57þ and CD8þCD27þCD57� subsets, from 3 differentpatients, before and after CD3 and CD28 stimulation for 7 days. TGF-b1 (1 ng/mL) was added to some cultures as indicated, and the cells incubated and thenstained in the same way. A minimum of 50,000 gated light scatter events per sample were acquired on the flow cytometer. ND�, not determineddue to insufficient cell number. B, a caspase-3 cleavage assay, described inMaterials andMethods, was used to assess the CTL killing activity. Sorted CD8þ
TIL subsets, cultured with LD IL-2 alone, or after TCR restimulation with anti-CD3 and anti-CD28, with or without added TGF-b1 (1 ng/mL) for 7 days,were coincubated for 3 hours at the indicated E:T ratios with P815 (murine mastocytoma) target cells, which were previously pulsed with 200 ug/mLof anti-CD3 antibody, in a redirected lysis assay. Target cells were distinguished from effector T cells by labeling with a fixable, far red fluorescent tracer,CellTrace Far Red DDAO-Succinimidyl Ester (DDAO-SE) at the beginning of the assay. Nonpulsed P815 cells were used as negative controls. Resultswere expressed as percent cleaved caspase-3–positive target cells out of the total target cell population. A minimum of 50,000 gated target cells per samplewere acquired on the flowcytometer. Results shownwere fromexperimentsdoneon sortedTIL subsets from2different patients as indicated.Relevant patientinformation on TIL #2544 was described in Supplementary Fig. S5B.
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2474
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

Interestingly, TILs in breast and kidney carcinoma havealso been observed to produce IL-13, which was hypothe-sized to negatively regulate indoleamine 2,3-dioxygenase(IDO) expression in tumors (45). Last, our sorting experi-ments revealed that TCR stimulation induced sortedCD8þCD27þCD57þ T cells to differentiate intoCD8þCD27�CD57þGBhighPerfhigh T cells, reminiscent ofTTDE CD57þ CTL in the peripheral blood; this observationwas not due to CD8þCD27þCD57� TIL contaminantsbecause sorted CD8þCD27þCD57� T cells did not differ-entiate into CD27�CD57þ T cells, but rather, differenti-ated directly into CD27�CD57� T cells, which furthersupports that CD8þCD57� and CD8þCD57þ TILs belongto distinct lineages or subsets. We do not know whetherthese CD8þCD27þCD57þ TILs persist as short-lived effec-tor cells at the tumor site or recirculate in the periphery.Addressing this question requires a murine model, but thisCD8þCD57þ T-cell subset does not exist in mice (11).On the basis of our initial observation that incompletely
differentiated Perflow CD8þ TILs accumulate in the tumormicroenvironment, we reasoned that some immunosup-pressive mechanism may account for the lack of furtherdifferentiation. TGF-b1 is known to be secreted by manytumors, including melanoma cells and certain types oftumor-associated macrophages (24–27). Previous studieshave also established that TGF-b1 inhibited the differenti-ation of na€�ve CD8þ T cells into effector cells (37, 39),impaired the production of inflammatory cytokines such asIFN-g and TNF-a by effector CD8þ T cells (24), and atten-uated Perf expression in lymphokine-activated killer cellsand murine CD8þ T cells (38, 46). The negative effect ofTGF-b1 on CTL function is evident in studies showingenhanced tumor eradication in mice with T cells lackingTGF-b signaling (47). Indeed, we found that TGF-b1 pre-vented the differentiation of CD8þCD27þ subsets intomoremature CD8þCD27�CTLs. This cytokine also potent-ly inhibited the expression of Perf and GB to a lesser extentin CD8þ TILs. In addition to TGF-b1, other factors in thetumor microenvironment, such as a lack of adequate cyto-kine signaling (e.g, IL-2 and IL-15), or lack of propercostimulatory signals (e.g., through 4-1BB), or the presenceof inhibitory costimulation through PD-1, BTLA, or TIM-3(30, 48) following TCR activation could also contribute tothis arrest in CTL differentiation. Alternatively, the presenceof other immunosuppressive cytokines, such as IDO, pros-taglandin E2, or IL-10 could also play a role (27, 49).In summary, we have identified a unique stage in which
CD8þCTL differentiation seems to be arrested in the tumormicroenvironment in metastatic melanoma and in breastcancer. This incompletely differentiated state of melanomaCD8þ TILs was also associated with the appearance of anunusual CD8þ effector memory subset coexpressing a lateterminally differentiated CTL marker, CD57. TheseCD8þCD27þCD28þCD57þ TILs, although expressing GB,have little or no Perf expression and therefore seem to be aunique effector memory subset in cancer. Immunosuppres-sive factors, such as TGF-b1, may lead to incomplete CTLdifferentiation and the appearance of this unique CD57þ
subset. It is possible that these unique CD57þ cellsmay alsorepresent in part CD8þCD27þCD57� TILs that haveattempted to differentiate into CD57þ cells, but wereblocked from further differentiation into CD27�Perfhigh
cells in the tumor microenvironment. On the contrary, theCD8þ CD27þCD57� effector memory TILs, which were infact the dominant CD8þ TIL subpopulation, may alsorepresent a putative CTL precursor subset, which, like thehybrid CD27þCD57þ subset, was also functionally sup-pressed by the immunosuppressive factors in the tumormicroenvironment. Finally, tumor antigen-specific CD8þ
populations could be found in both CD27þCD57� andCD27þCD57þ subsets. Thus, future studies will be neededto address whether the more differentiated CD8þCD27�
TILs exhibit better control of tumor growth in vivo than theirCD27þ precursors. In addition, it will be of interest tofurther examine whether CD8þCD27þCD57þ TILs candifferentiate intomore long-livedCD8þCD27�CD57þ cellsin vivo, which are reminiscent of the long-lived CD57þ Tcells in the blood that effectively control CMV and EBVinfection in humans (4, 10).
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsPhenotypic analysis of the TIL by flow cytometry: R.C. Wu, S.J. LiuFunctional studies of the TIL subsets: R.C. WuDesigned experiments: L.G. RadvanyiSupervised the work: L.G. RadvanyiProcured the melanoma tissue: L.G. RadvanyiWriting and editing the manuscript: L.G. RadvanyiProvided the breast cancer pleural effusions patient samples: J.L.MurrayProvided anti-human KLRG-1 mAb: H. PircherConducted the Vb spectratyping experiment: P. Sukhumalchandra, J.J.Murray
AcknowledgmentsThe authors thank the cooperation and efforts of the melanoma surgical
staff at MD Anderson Cancer Center who have generously provided all themelanoma specimens for this study, including Drs. Jeffrey Gershenwald,Jeffrey Lee, Merrick Ross, Amy Heimberger, Paul Mansfield, Janice Cormier,and Anthony Lucci. The authors also thank Renjith Ramachandran, SethWardell, Audrey Gonzalez, Christopher Toth, OJ Fulbright, and RahmatuMansaray, for processing tumor specimens; Dr. James L. Murray from theBreast Medical Oncology Department for providing the breast cancer pleuraleffusion samples; Dr. Victor Prieto and Katie McNeil, MD Anderson’sMelanoma SPORE Tissue Bank personnel, for help in obtaining the mela-noma samples used in this study; Drs. Kevin Kim, Natalia Martin-Orozco,Chantale Bernatchez, and Willem Overwijk for critical reading of the man-uscript; and Dr. Luis Vence, Himabindu Pappu, Leslie Wiltz of the MDAnderson Cancer Center Immunomonitoring Core Lab (IMCL) for helpingwith flow cytometry and cytokine analysis.
Grant SupportThisworkwas supportedbyNationalCancer Institute grantR01-CA111999
to P. Hwu and L. Radvanyi, by The University of Texas MD Anderson CancerCenter support grant (P30-CA16672) to the Immune Monitoring Core Laband to the Flow Cytometry Core Facility, and Award No. TL1RR024147 fromtheNationalCenter for ResearchResources toR.C.Wu. Thisworkalso receivedsupport from a Melanoma SPORE Developmental grant (P50-CA093459-05-DRP21) and a Team Science Award from the Melanoma Research Alliance(MRA) to L. Radvanyi andDr. Miriam and Sheldon AdelsonMedical ResearchFoundation (AMRF) and the Mulva Foundation.
The costs of publicationof this articlewere defrayed inpart by thepaymentof page charges. This articlemust therefore be herebymarked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received August 5, 2011; revised January 8, 2012; accepted January 23,2012; published OnlineFirst February 3, 2012.
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2475
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

References1. Dolstra H, Preijers F, Van de Wiel-van Kemenade E, Schattenberg A,
Galama J, de Witte T. Expansion of CD8þCD57þ T cells after allo-geneic BMT is related with a low incidence of relapse and withcytomegalovirus infection. Br J Haematol 1995;90:300–7.
2. Kagi D, Ledermann B, Burki K, Seiler P, Odermattt B, Olsen KJ, et al.Cytotoxicity mediated by T cells and natural killer cells is greatlyimpaired in perforin-deficient mice. Nature 1994;369:31–7.
3. Lieberman J, Trimble LA, Friedman RS, Lisziewicz J, Lori F, Shankar P,et al. Expansion of CD57 and CD62L-CD45RAþ CD8 T lymphocytescorrelates with reduced viral plasma RNA after primary HIV infection.AIDS 1999;13:891–9.
4. Lynne JE, Schmid I, Matud JL, Hirji K, Buessow S, Shlian DM, et al.Major expansions of select CD8þ subsets in acute Epstein-Barr virusinfection: comparison with chronic human immunodeficiency virusdisease. J Infect Dis 1998;177:1083–7.
5. OhkawaT, Seki S,DobashiH,KoikeY,HabuY, AmiK, et al. Systematiccharacterization of human CD8þ T cells with natural killer cell markersin comparison with natural killer cells and normal CD8þ T cells.Immunology 2001;103:281–90.
6. TakayamaE,KoikeY,OhkawaT,MajimaT, FukasawaM,ShinomiyaN,et al. Functional and Vbeta repertoire characterization of human CD8þ
T-cell subsets with natural killer cell markers, CD56þ CD57� T cells,CD56þ CD57þ T cells and CD56� CD57þ T cells. Immunology2003;108:211–9.
7. AppayV,DunbarPR,CallanM,KlenermanP,GillespieGM,Papagno L,et al.MemoryCD8þTcells vary in differentiation phenotype indifferentpersistent virus infections. Nat Med 2002;8:379–85.
8. Hoji A, Connolly NC, Buchanan WG, Rinaldo CR Jr. CD27 and CD57expression reveals atypical differentiation of human immunodeficiencyvirus type 1-specific memory CD8þ T cells. Clin Vaccine Immunol2007;14:74–80.
9. Brenchley JM, Karandikar NJ, Betts MR, Ambrozak DR, Hill BJ, CrottyLE, et al. Expression of CD57 defines replicative senescence andantigen-induced apoptotic death of CD8þ T cells. Blood 2003;101:2711–20.
10. van Baarle D, Kostense S, Hovenkamp E, Ogg G, Nanlohy N, CallanMF, et al. Lack of Epstein-Barr virus- and HIV-specific CD27� CD8þ Tcells is associated with progression to viral disease in HIV-infection.AIDS 2002;16:2001–11.
11. Okada T, Liai T, Kawachi Y, Moroda T, Takii Y, Hatakeyama K, Abo T.Origin of CD57þ T cells which increase at tumour sites in patients withcolorectal cancer. Clin Exp Immunol 1995;102:159–66.
12. KaechSM,Wherry EJ.Heterogeneity andcell-fate decisions in effectorandmemoryCD8þT cell differentiation during viral infection. Immunity2007;27:393–405.
13. Casado JG, Soto R, DelaRosa O, Peralbo E, del Carmen-Mu~noz-Villanueva M, Rioja L, et al. CD8 T cells expressing NK associatedreceptors are increased in melanoma patients and display an effectorphenotype. Cancer Immunol Immunother 2005;54:1162–71.
14. Akagi J, Baba H. Prognostic value of CD57(þ) T lymphocytes in theperipheral blood of patients with advanced gastric cancer. Int J ClinOncol 2008;13:528–35.
15. Characiejus D, Pasukoniene V, Jonusauskaite R, Azlauskaite N,Aleknavicius E, Mauricas M, et al. Peripheral blood CD8highCD57þ
lymphocyte levels may predict outcome in melanoma patients trea-ted with adjuvant interferon-alpha. Anticancer Res 2008;28:1139–42.
16. Characiejus D, Pasukoniene V, Kazlauskaite N, Valuckas KP, PetraitisT, Mauicas M, et al. Predictive value of CD8highCD57þ lymphocytesubset in interferon therapy of patients with renal cell carcinoma.Anticancer Res 2002;22:3679–83.
17. Chochi K, Ichikura T, Majima T, Kawabata T, Matsumoto A, SugasawaH, et al. The increase of CD57þ T cells in the peripheral blood and theirimpaired immune functions in patients with advanced gastric cancer.Oncol Rep 2003;10:1443–8.
18. Bandres E, Merino J, Vazquez B, Inog�es S, Moreno C, Subir�a ML. Theincrease of IFN-gamma production through aging correlates with theexpanded CD8(þhigh) CD28(-)CD57(þ) subpopulation. Clin Immunol2000;96:230–5.
19. Gorochov G, Debre P, Leblond V, Sadat-Sowti B, Sigaux F, Autran B.Oligoclonal expansion of CD8þ CD57þ T cells with restricted T-cellreceptor beta chain variability after bone marrow transplantation.Blood 1994;83:587–95.
20. PittetMJ, Speiser DE, Valmori D, Cerottini JC, RomeroP. Cutting edge:cytolytic effector function inhumancirculatingCD8þTcells closely cor-relates with CD56 surface expression. J Immunol 2000;164:1148–52.
21. Anichini A, Molla A, Vegetti C, Bersani I, Zappasodi R, Arienti F, et al.Tumor-reactive CD8þ early effector T cells identified at tumor site inprimary and metastatic melanoma. Cancer Res 2010. 70:8378–87.
22. Sallusto F, Geginat J, Lanzavecchia A. Central memory and effectormemory T cell subsets: function, generation, and maintenance. AnnuRev Immunol 2004;22:745–63.
23. Takata H, Takiguchi M. Three memory subsets of human CD8þ T cellsdifferently expressing three cytolytic effector molecules. J Immunol2006;177:4330–40.
24. AhmadzadehM,RosenbergSA. TGF-beta 1 attenuates the acquisitionand expression of effector function by tumor antigen-specific humanmemory CD8 T cells. J Immunol 2005;174:5215–23.
25. Bennicelli JL, Guerry Dt. Production of multiple cytokines by culturedhuman melanomas. Exp Dermatol 1993;2:186–90.
26. Derynck R, Jarrett JA, Chen EY, Eaton DH, Bell JR, Assoian RK, et al.Human transforming growth factor-beta complementary DNAsequence and expression in normal and transformed cells. Nature1985;316:701–5.
27. Polak ME, Borthwick NJ, Gabriel FG, Johnson P, Higgins B, Hurren J,et al. Mechanisms of local immunosuppression in cutaneous mela-noma. Br J Cancer 2007;96:1879–87.
28. Li Y, Liu S, Hernandez J, Vence L, Hwu P, Radvanyi L. MART-1–specific melanoma tumor-infiltrating lymphocytes maintaining CD28expression have improved survival and expansion capability followingantigenic restimulation in vitro. J Immunol 2010;184:452–65.
29. He L, Hakimi J, Salha D, Miron I, Dunn P, Radvanyi L. A sensitive flowcytometry-based cytotoxic T-lymphocyte assay through detection ofcleavedcaspase3 in target cells. J ImmunolMethods2005;304:43–59.
30. Sakuishi K, Apetoh L, Sullivan JM, Blazar BR, Kuchroo VK, AndersonAC. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustionand restore anti-tumor immunity. J Exp Med 2010. 207:2187–94.
31. Quigley M, Pereyra F, Nilsson B, Porichis F, Fonseca C, Eichbaum Q,et al. Transcriptional analysis of HIV-specific CD8þ T cells showsthat PD-1 inhibits T cell function by upregulating BATF. Nat Med2010;16:1147–51.
32. Le Priol Y, Puthier D, Lecureuil C, Combadi�ere C, Debr�e P, Nguyen C,et al. High cytotoxic and specific migratory potencies of senescentCD8þ CD57þ cells in HIV-infected and uninfected individuals. JImmunol 2006;177:5145–54.
33. Dudley ME, Wunderlich JR, Robbins PF, Yang JC, Hwu P, Schwart-zentruber DJ, et al. Cancer regression and autoimmunity in patientsafter clonal repopulation with antitumor lymphocytes. Science2002;298:850–4.
34. Bannard O, Kraman M, Fearon DT. Cutting edge: virus-specific CD8þT cell clones and the maintenance of replicative function during apersistent viral infection. J Immunol 2010;185:7141–5.
35. Ibegbu CC, Xu YX, Harris W, Maggio D, Miller JD, Kourtis AP. Expres-sion of killer cell lectin-like receptor G1 on antigen-specific humanCD8þ T lymphocytes during active, latent, and resolved infection andits relation with CD57. J Immunol 2005;174:6088–94.
36. Morley JK, Batliwalla FM, Hingorani R, Gregersen PK. OligoclonalCD8þ T cells are preferentially expanded in the CD57þ subset. JImmunol 1995;154:6182–90.
37. Kim YJ, Stringfield TM, Chen Y, Broxmeyer HE. Modulation of cordblood CD8þ T-cell effector differentiation by TGF-beta1 and 4-1BBcostimulation. Blood 2005;105:274–81.
38. Smyth MJ, Strobl SL, Young HA, Ortaldo JR, Ochoa AC. Regulation oflymphokine-activated killer activity and pore-forming protein geneexpression in human peripheral blood CD8þ T lymphocytes. Inhibitionby transforming growth factor-beta. J Immunol 1991;146:3289–97.
39. Willinger T, Freeman T, HerbertM, HasegawaH,McMichael AJ, CallanMF. Human naive CD8 T cells down-regulate expression of the WNT
Wu et al.
Clin Cancer Res; 18(9) May 1, 2012 Clinical Cancer Research2476
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

pathway transcription factors lymphoid enhancer binding factor 1 andtranscription factor 7 (T cell factor-1) following antigen encounter invitro and in vivo. J Immunol 2006;176:1439–46.
40. Lopez-Verges S, Milush JM, Pandey S, York VA, Arakawa-Hoyt J,Pircher H, et al. CD57 defines a functionally distinct population ofmature NK cells in the human CD56dimCD16þ NK-cell subset. Blood2010;116:3865–74.
41. Jungalwala FB. Expression and biological functions of sulfoglucuronylglycolipids (SGGLs) in the nervous system–a review. Neurochem Res1994;19:945–57.
42. Cebo C, Dambrouck T, Maes E, Laden C, Strecker G, Michalski JC,et al. Recombinant human interleukins IL-1alpha, IL-1beta, IL-4, IL-6,and IL-7 show different and specific calcium-independent carbohy-drate-binding properties. J Biol Chem 2001;276:5685–91.
43. Chong LK, Aicheler RJ, Llewellyn-Lacey S, Tomasec P., Brennan P,Wang EC. Proliferation and interleukin 5 production by CD8hiCD57þ Tcells. Eur J Immunol 2008;38:995–1000.
44. Zhang J, Scordi I, Smyth MJ, Lichtenheld MG. Interleukin 2 receptorsignaling regulates the perforin gene through signal transducer and
activator of transcription (Stat)5 activation of two enhancers. J ExpMed 1999;190:1297–308.
45. Godin-Ethier J, Pelletier S, Hanafi LA, Gannon PO, Forget MA,Routy JP, et al. Human activated T lymphocytes modulate IDOexpression in tumors through Th1/Th2 balance. J Immunol 2009;183:7752–60.
46. Thomas DA, Massague J. TGF-beta directly targets cytotoxic T cellfunctions during tumor evasion of immune surveillance. Cancer Cell2005;8:369–80.
47. Gorelik L, Flavell RA. Immune-mediated eradication of tumors throughthe blockade of transforming growth factor-beta signaling in T cells.Nat Med 2001;7:1118–22.
48. Derr�e L, Rivals JP, Jandus C, Pastor S, Rimoldi D, Romero P, et al.BTLA mediates inhibition of human tumor-specific CD8þ T cells thatcan be partially reversed by vaccination. J Clin Invest 2010. 120:157–67.
49. Haas AR, Sun J, Vachani A, Wallace AF, Silverberg M, Kapoor V, et al.Cycloxygenase-2 inhibition augments the efficacy of a cancer vaccine.Clin Cancer Res 2006;12:214–22.
Novel Nonsenescent CD8þCD57þ Subset in Human Cancer
www.aacrjournals.org Clin Cancer Res; 18(9) May 1, 2012 2477
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034

2012;18:2465-2477. Published OnlineFirst February 3, 2012.Clin Cancer Res Richard C. Wu, Shujuan Liu, Jessica A. Chacon, et al. PhenotypeCells in Metastatic Melanoma with an Incompletely Differentiated
T+CD57+Detection and Characterization of a Novel Subset of CD8
Updated version
10.1158/1078-0432.CCR-11-2034doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2012/02/03/1078-0432.CCR-11-2034.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/18/9/2465.full#ref-list-1
This article cites 48 articles, 23 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/18/9/2465.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/18/9/2465To request permission to re-use all or part of this article, use this link
on March 23, 2020. © 2012 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 3, 2012; DOI: 10.1158/1078-0432.CCR-11-2034





























