Embed Size (px)
Citation preview

Hydrobiologia 520: 153–163, 2004.© 2004 Kluwer Academic Publishers. Printed in the Netherlands.
153
Density of red alder (Alnus rubra) in headwaters influences invertebrateand detritus subsidies to downstream fish habitats in Alaska
Mark S. Wipfli∗ & Jake MusslewhitePacific Northwest Research Station, USDA Forest Service, 2770 Sherwood Lane, Juneau, AK 99801-8545, U.S.A.∗Present address: Alaska Cooperative Fish and Wildlife Research Unit, USGS, Institute of Arctic Biology,209 Irving I Building, University of Alaska Fairbanks, Fairbanks, AK 99775-7020, U.S.A.E-mail: [email protected]
Received 2 April 2003; in revised form 4 December 2003; accepted 5 January 2004
Key words: alder, headwater stream, subsidy, invertebrate; detritus, salmonid, riparian forest management
Abstract
We investigated the influence of red alder (Alnus rubra) stand density in upland, riparian forests on invertebrateand detritus transport from fishless headwater streams to downstream, salmonid habitats in southeastern Alaska.Red alder commonly regenerates after soil disturbance (such as from natural landsliding or timber harvesting),and is common along streams in varying densities, but its effect on food delivery from headwater channels todownstream salmonid habitats is not clear. Fluvial transport of invertebrates and detritus was measured at 13 sitesin spring, summer and fall during two years (2000–2001). The 13 streams encompassed a riparian red alder densitygradient (1–82% canopy cover or 0–53% basal area) growing amongst young-growth conifer (45-yr-old stands thatregenerated after forest clearcutting). Sites with more riparian red alder exported significantly more invertebratesthan did sites with little alder (mean range across 1–82% alder gradient was about 1–4 invertebrates m−3 water,and 0.1–1 mg invertebrates m−3 water, respectively). Three-quarters of the invertebrates were of aquatic origin;the remainder was of terrestrial origin. Aquatic taxa were positively related to the alder density gradient, whileterrestrially-derived taxa were not. Streams with more riparian alder also exported significantly more detritus thanstreams with less alder (mean range across 1–82% alder gradient was 0.01–0.06 g detritus m−3 water). Based onthese data, we predict that headwater streams with more riparian alder will provide more invertebrates and supportmore downstream fish biomass than those basins with little or no riparian alder, provided these downstream foodwebs fully utilize this resource subsidy.
Introduction
Timber harvesting is widely viewed as having broadlynegative effects on stream ecosystems in the PacificNorthwest of North America and other places (NRC,1996). Clearcutting has historically been the mostcommon harvesting technique and under some cir-cumstances can lead to increased soil erosion andstream sedimentation (Waters, 1995; NRC, 1996), lossof large woody material and fish habitat (Gregory &Bisson, 1997), loss of allochthonous inputs (Bisson& Bilby, 1998), and increased solar radiation thatcan lead to dangerously warmer water temperaturesfor salmonids (Beschta et al., 1987; Bjornn & Reiser,
1991). Although many negative impacts can occur inforest ecosystems soon after some timber harvestingand may persist for decades in terrestrial ecosystems(Alaback, 1982), less is understood about the long-term consequences of post-harvest, forest successionon stream ecosystems. Regenerating forests shouldstabilize soils and provide woody material (albeitsmaller pieces in the short term than what the old-growth forests provided) potentially creating pools andcover for fish habitat, and provide leaf litter and shadeto streams (Naiman & Bilby, 1998). Although muchof the regenerating riparian canopy is red alder (Alnusrubra Bong.) in the Pacific Northwest, larger conifersare viewed as more desirable, providing stable and

154
longer term woody material for pool formation andfish habitat, sediment retention, and bank stabilization(Bilby & Bisson, 1998).
Red alder is a pioneer species that often colonizesrecently disturbed soil surfaces such as abandoned log-ging roads, log transfer sites, landslide areas, streambanks and other soil-disturbed or high light envir-onments in the Pacific Northwest (Hulten, 1968). Itquickly establishes following timber harvesting andcan persist for 40 to 80 years until being overtopped byconifers (Hibbs et al., 1994). Though not considered avaluable source of wood for fish habitat, red alder hasgreat potential to alter forest ecosystem productivityand biodiversity (Hibbs et al., 1994). Through nitrogenfixation it can increase soil nitrogen concentrations(Binkley et al., 1994) and understory biomass and spe-cies richness (Newton & Cole, 1994). And it is morepalatable and nutritious than conifers for terrestrialinvertebrates (Southwood, 1961; McComb, 1994).
If red alder affects terrestrial productivity and biod-iversity, it may have similar effects on streams andtheir riparian habitats. The nitrogen it provides shouldelevate stream, hyporheic and riparian soil nitrogenconcentrations (Binkley et al., 1994). More nitro-gen, in nitrogen-limited systems, should lead to elev-ated in-stream primary production and subsequentlyhigher secondary productivity (Allan, 1995; Borch-ardt, 1996). Higher quality (e.g., lack of secondaryplant compounds, high N content) allochthonous in-puts from alder (versus conifers) (McComb, 1994)may also provide better food resources that shouldsustain more detritivores in streams. In addition, ter-restrial invertebrates falling into streams may be moreabundant in the presence of alder. Wipfli (1997) andAllan et al. (2003) found red alder in young-growthforests along fish-bearing tributaries may providemore terrestrial invertebrates for drift-feeding streamfishes than systems with little alder.
Red alder in headwater forests could also havesubstantial effects on downstream fishes. Wipfli andGregovich (2002) showed that fishless headwaterstreams subsidize downstream fish-bearing food webswith invertebrates and detritus that originates in up-land forests and associated headwaters and that ter-restrial invertebrates that fall into headwater streamsfrom adjacent riparian habitats are part of this subsidy.Piccolo & Wipfli (2002) demonstrated that uplandforest management in southeastern Alaska alters thissubsidy, and that regenerating riparian forests that con-tain dense stands of red alder increased the amountof invertebrates transported to food webs lower in the
drainage. Streams with dense canopies of conifer andlacking alder supplied fish-bearing food webs with farfewer invertebrates (Piccolo & Wipfli, 2002). Addi-tionally, invertebrates of terrestrial origin form a largeportion of the food base for juvenile coho salmon, cut-throat trout, and Dolly Varden char in small rearingstreams in southeastern Alaska, and riparian vegeta-tion type may influence the amount of terrestrial preyentering stream food webs (Wipfli, 1997; Allan et al.,2003).
Current federal timber-harvesting guidelines forupland forests in southeastern Alaska allow clearcut-ting in some headwaters that contain perennial andintermittent streams (USDA Forest Service, 1999).The small, high gradient streams that drain theseupland forested headwaters receive variable riparianbuffering during timber harvesting. These streamstypically drain into downstream salmonid habitats in-fluencing communities in higher order streams lowerin the drainage (Gomi et al., 2002), thus it is import-ant to understand how forest regeneration in uplandforests influences food delivery to these fish-bearingfood webs.
The objectives of this study were to investigatehow a range of red alder density in young, regen-erating riparian stands of even-aged conifer affectsthe delivery of invertebrates and detritus (via fluvialtransport) from forested headwaters to downstreamfish-bearing habitats. Specifically, we were interestedin understanding how red alder density affects theamount (number and mass) of invertebrates (terrestrialand aquatic) and mass of detritus transported. Thisstudy was one component of a broader study thatset out to identify possible ecosystem-level patternsin response to red alder abundance in regenerating,young-growth conifer forests (Wipfli et al., 2003). Itis a next logical step in headwater stream research insoutheastern Alaska, following the findings by Pic-colo & Wipfli (2002) that indicated both dense coniferand red alder stands can affect material transport fromheadwaters, to uncover headwater-downstream trophicrelationships across a broad range of red alder-coniferriparian stand densities. This information should helpguide upland forest management in areas containingheadwater streams in both old-growth and regenerat-ing young-growth forested ecosystems in southeasternAlaska and other parts of the world where Alnus spp.occur.

155
Materials and methods
Study sites
This study was conducted in 13 small (mean dis-charge 0.9–116 L s−1), perennial, headwater streamswithin the Maybeso (12 streams) and Harris (onestream) drainage basins on eastern Prince of WalesIsland, southeastern Alaska (Fig. 1). The Maybesoand Harris basins are 46 km2 and 108 km2, respect-ively. The study area was extensively clearcut duringthe 1950s and 1960s as part of the first large-scale,commercial timber harvest in southeastern Alaska. In2000–2001, we sampled the fluvial transport of in-vertebrates and coarse detritus (>250 µm) seasonally(spring, summer and fall) in our 13 streams. Thesestreams contained a range of red alder density (1–82% canopy coverage and 0–53% basal area) mixedin the naturally regenerating (40 years post-cut) con-ifer – Sitka spruce (Picea sitchensis (Bong.) Carriére),western hemlock (Tsuga heterophylla (Raf.) Sarg.),and Alaska yellow cedar (Chamaecyparis nootkaten-sis (D. Don) Spach). Red alder was found throughoutthe drainage but was more concentrated along streams(personal observation). Streams within a size range(mean active channel width 0.7–3.4 m; stream length<5 km), gradient (11–27◦), and red alder canopycover (1–82%) were selected for the study (Fig. 1,Table 1). Sites within these two drainage basins werechosen because they met the criteria of all the re-search components of the larger, multi-disciplinary(wildlife, forest vegetation, geomorphology, fish andfish habitat, and stream chemistry and ecology) study(Wipfli et al., 2003). A ‘transition zone’ ranging 1–50 m long on each stream was established to markthe upper range of salmonid occurrence (>10◦ gradi-ent). A ‘transition point’ marked the upper limit ofthis transition zone, dividing fish-bearing from fish-less reaches of the stream. The transition point wasoften an impassable barrier to upstream fish move-ment, a high-gradient and shallow reach, or othernotable feature that precluded salmonid occurrenceupstream, and marked the downstream end of thestudy reach. Macroinvertebrate and detritus transferacross the fishless/fish-bearing interface was collectedat this point to eliminate any possible suppression ofmacroinvertebrate populations from fish predation inthe upstream study reaches. Resident salmonid speciesin the fish-bearing reaches downstream were juvenilecoho salmon (Oncorhynchus kisutch Walbaum), cut-throat trout (O. clarki Richardson), and Dolly Varden
Figure 1. Stream study sites (13) within the Harris and Maybesodrainage basins on eastern Prince of Wales Island, southeasternAlaska. Both drainage basins empty into saltwater in TwelvemileArm.
char (Salvelinus malma Walbaum). Generally overtwo-thirds of the total stream length of the studystreams was fishless.
Stream channel gradient, active channel width,canopy coverage, and basal area of riparian trees weresurveyed along a 300 m reach of each stream dir-ectly upstream from the transition point. Gradient wasmeasured with a hand-held clinometer and canopycoverage was estimated every 50 m along the streamusing a circular viewing tube (10 cm inside-diameter× 30 cm long) (n = 7 for all streams that were 300 mor longer above sampling station). To estimate can-opy coverage and composition, the observer stood inthe center of the stream channel, looked through thevertically-held tube, and recorded the percentage ofalder, conifer, and open canopy. Basal area was de-
156
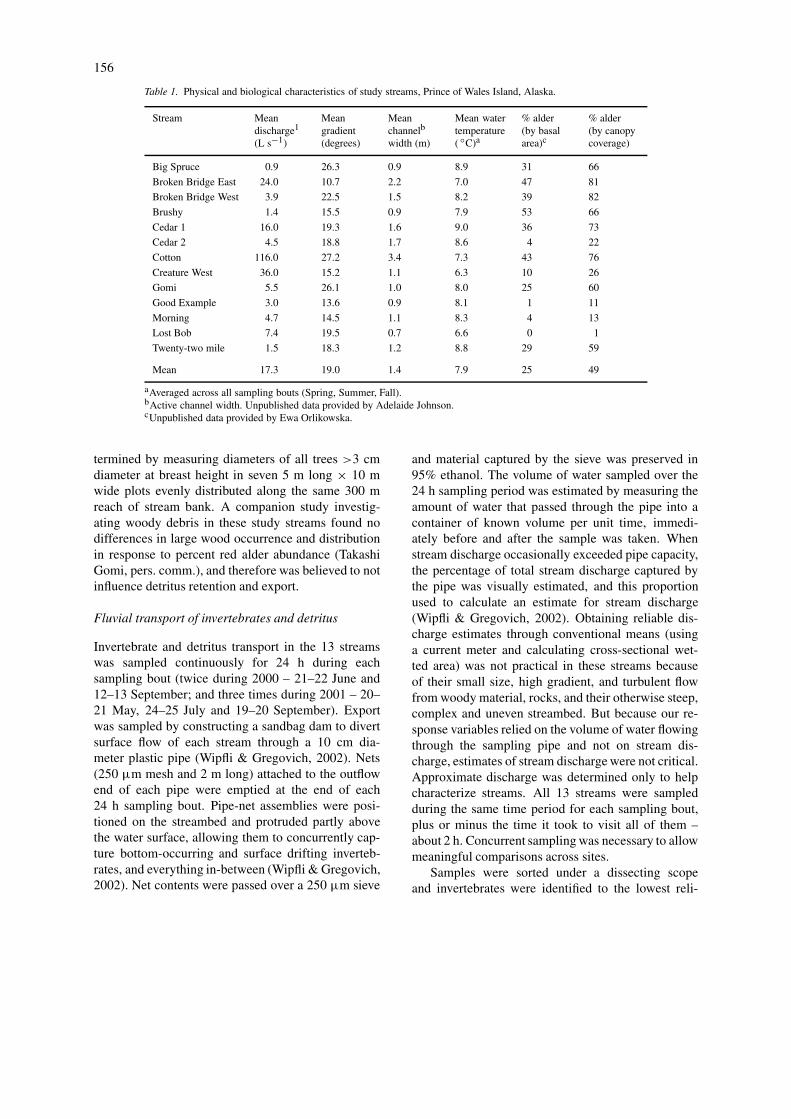
Table 1. Physical and biological characteristics of study streams, Prince of Wales Island, Alaska.
Stream Meandischarge1
(L s−1)
Meangradient(degrees)
Meanchannelb
width (m)
Mean watertemperature( ◦C)a
% alder(by basalarea)c
% alder(by canopycoverage)
Big Spruce 0.9 26.3 0.9 8.9 31 66
Broken Bridge East 24.0 10.7 2.2 7.0 47 81
Broken Bridge West 3.9 22.5 1.5 8.2 39 82
Brushy 1.4 15.5 0.9 7.9 53 66
Cedar 1 16.0 19.3 1.6 9.0 36 73
Cedar 2 4.5 18.8 1.7 8.6 4 22
Cotton 116.0 27.2 3.4 7.3 43 76
Creature West 36.0 15.2 1.1 6.3 10 26
Gomi 5.5 26.1 1.0 8.0 25 60
Good Example 3.0 13.6 0.9 8.1 1 11
Morning 4.7 14.5 1.1 8.3 4 13
Lost Bob 7.4 19.5 0.7 6.6 0 1
Twenty-two mile 1.5 18.3 1.2 8.8 29 59
Mean 17.3 19.0 1.4 7.9 25 49
aAveraged across all sampling bouts (Spring, Summer, Fall).bActive channel width. Unpublished data provided by Adelaide Johnson.cUnpublished data provided by Ewa Orlikowska.
termined by measuring diameters of all trees >3 cmdiameter at breast height in seven 5 m long × 10 mwide plots evenly distributed along the same 300 mreach of stream bank. A companion study investig-ating woody debris in these study streams found nodifferences in large wood occurrence and distributionin response to percent red alder abundance (TakashiGomi, pers. comm.), and therefore was believed to notinfluence detritus retention and export.
Fluvial transport of invertebrates and detritus
Invertebrate and detritus transport in the 13 streamswas sampled continuously for 24 h during eachsampling bout (twice during 2000 – 21–22 June and12–13 September; and three times during 2001 – 20–21 May, 24–25 July and 19–20 September). Exportwas sampled by constructing a sandbag dam to divertsurface flow of each stream through a 10 cm dia-meter plastic pipe (Wipfli & Gregovich, 2002). Nets(250 µm mesh and 2 m long) attached to the outflowend of each pipe were emptied at the end of each24 h sampling bout. Pipe-net assemblies were posi-tioned on the streambed and protruded partly abovethe water surface, allowing them to concurrently cap-ture bottom-occurring and surface drifting inverteb-rates, and everything in-between (Wipfli & Gregovich,2002). Net contents were passed over a 250 µm sieve
and material captured by the sieve was preserved in95% ethanol. The volume of water sampled over the24 h sampling period was estimated by measuring theamount of water that passed through the pipe into acontainer of known volume per unit time, immedi-ately before and after the sample was taken. Whenstream discharge occasionally exceeded pipe capacity,the percentage of total stream discharge captured bythe pipe was visually estimated, and this proportionused to calculate an estimate for stream discharge(Wipfli & Gregovich, 2002). Obtaining reliable dis-charge estimates through conventional means (usinga current meter and calculating cross-sectional wet-ted area) was not practical in these streams becauseof their small size, high gradient, and turbulent flowfrom woody material, rocks, and their otherwise steep,complex and uneven streambed. But because our re-sponse variables relied on the volume of water flowingthrough the sampling pipe and not on stream dis-charge, estimates of stream discharge were not critical.Approximate discharge was determined only to helpcharacterize streams. All 13 streams were sampledduring the same time period for each sampling bout,plus or minus the time it took to visit all of them –about 2 h. Concurrent sampling was necessary to allowmeaningful comparisons across sites.
Samples were sorted under a dissecting scopeand invertebrates were identified to the lowest reli-

157
Figure 2. Mean macroinvertebrate density (a) and biomass (b)across 1–82% red alder canopy coverage, exported from head-water streams on Prince of Wales Island, Alaska. Error barsrepresent one standard error (n = 5). r and P -values are forlogarithm-transformed data.
able taxon, measured to the nearest millimeter (totallength excluding antennae and cerci) and enumerated.Invertebrate biomass (mg dry mass) was estimatedfor individual invertebrates by using published taxa-specific length-weight regression equations (Rogerset al., 1977; Sample et al., 1993; Burgherr & Meyer,1997). Detritus was dried (at 60 ◦C for 48 h), weighed,burned to ash (at 500 ◦C for 5 h), and re-weighed todetermine ash-free dry mass (AFDM).
Statistical analyses
Response variables were invertebrate density (numberm−3) and biomass (mg m−3), and detritus mass (mgAFDM m−3). Invertebrate density data were logar-ithmically transformed to meet ANOVA assumptions.Raw data were not normally distributed. Data (aver-aged across seasons and years; n = 5) were regressedacross % red alder canopy cover for all tests (test forslope >0 on transformed data P = 0.05). Simple lin-
Figure 3. Mean macroinvertebrate biomass of aquatic and terrestrialinvertebrates across season (a) and across % red alder canopy cover-age (b) exported from headwater streams on Prince of Wales Island,Alaska. Percent occurrence based on biomass. Error bars representone standard error (n = 13 in Spring, 26 in Summer, and 26 in Fallfor (a); and n = 5 for each of 13 means in (b)). r and P -values arefor logarithm-transformed data.
ear regressions (SAS, 1998) were run on transformeddata, while original (untransformed) data were usedfor presentation purposes. Values for r and P werereported on transformed data.
Results
Headwater streams with more riparian red alder trans-ported significantly more invertebrates to downstreamfish habitats than did sites with less alder (P = 0.010;r = 0.47) (Fig. 2a). Mean transport for sites at thelow end of the alder density continuum (< 20% aldercanopy coverage) was about 1 invertebrate m−3 water,and sites with up to 82% alder canopy exported about 4invertebrates m−3 water. When converted to biomass,transport across this 1–82% alder gradient ranged 0.1-1 mg invertebrates m−3 water, again, a significantrelationship across the red alder gradient (P = 0.002;r = 0.60) (Fig. 2b).
158
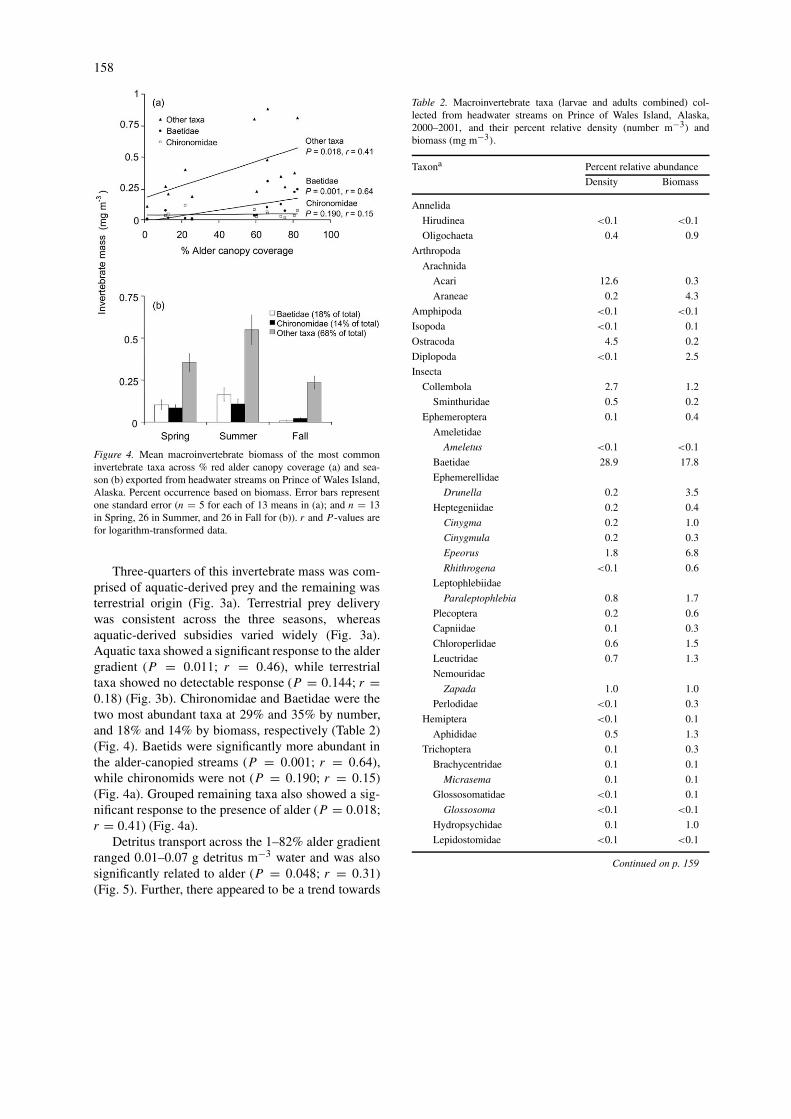
Figure 4. Mean macroinvertebrate biomass of the most commoninvertebrate taxa across % red alder canopy coverage (a) and sea-son (b) exported from headwater streams on Prince of Wales Island,Alaska. Percent occurrence based on biomass. Error bars representone standard error (n = 5 for each of 13 means in (a); and n = 13in Spring, 26 in Summer, and 26 in Fall for (b)). r and P -values arefor logarithm-transformed data.
Three-quarters of this invertebrate mass was com-prised of aquatic-derived prey and the remaining wasterrestrial origin (Fig. 3a). Terrestrial prey deliverywas consistent across the three seasons, whereasaquatic-derived subsidies varied widely (Fig. 3a).Aquatic taxa showed a significant response to the aldergradient (P = 0.011; r = 0.46), while terrestrialtaxa showed no detectable response (P = 0.144; r =0.18) (Fig. 3b). Chironomidae and Baetidae were thetwo most abundant taxa at 29% and 35% by number,and 18% and 14% by biomass, respectively (Table 2)(Fig. 4). Baetids were significantly more abundant inthe alder-canopied streams (P = 0.001; r = 0.64),while chironomids were not (P = 0.190; r = 0.15)(Fig. 4a). Grouped remaining taxa also showed a sig-nificant response to the presence of alder (P = 0.018;r = 0.41) (Fig. 4a).
Detritus transport across the 1–82% alder gradientranged 0.01–0.07 g detritus m−3 water and was alsosignificantly related to alder (P = 0.048; r = 0.31)(Fig. 5). Further, there appeared to be a trend towards
Table 2. Macroinvertebrate taxa (larvae and adults combined) col-lected from headwater streams on Prince of Wales Island, Alaska,2000–2001, and their percent relative density (number m−3) andbiomass (mg m−3).
Taxona Percent relative abundance
Density Biomass
Annelida
Hirudinea <0.1 <0.1
Oligochaeta 0.4 0.9
Arthropoda
Arachnida
Acari 12.6 0.3
Araneae 0.2 4.3
Amphipoda <0.1 <0.1
Isopoda <0.1 0.1
Ostracoda 4.5 0.2
Diplopoda <0.1 2.5
Insecta
Collembola 2.7 1.2
Sminthuridae 0.5 0.2
Ephemeroptera 0.1 0.4
Ameletidae
Ameletus <0.1 <0.1
Baetidae 28.9 17.8
Ephemerellidae
Drunella 0.2 3.5
Heptegeniidae 0.2 0.4
Cinygma 0.2 1.0
Cinygmula 0.2 0.3
Epeorus 1.8 6.8
Rhithrogena <0.1 0.6
Leptophlebiidae
Paraleptophlebia 0.8 1.7
Plecoptera 0.2 0.6
Capniidae 0.1 0.3
Chloroperlidae 0.6 1.5
Leuctridae 0.7 1.3
Nemouridae
Zapada 1.0 1.0
Perlodidae <0.1 0.3
Hemiptera <0.1 0.1
Aphididae 0.5 1.3
Trichoptera 0.1 0.3
Brachycentridae 0.1 0.1
Micrasema 0.1 0.1
Glossosomatidae <0.1 0.1
Glossosoma <0.1 <0.1
Hydropsychidae 0.1 1.0
Lepidostomidae <0.1 <0.1
Continued on p. 159

159
Table 2. Continued.
Taxona Percent relative abundance
Density Biomass
Limnephilidae 1.0 0.9
Cryptochia <0.1 <0.1
Ecclisomyia <0.1 <0.1
Psychoglypha <0.1 0.2
Philopotamidae 0.1 0.4
Polycentropodidae 0.1 0.1
Rhyacophilidae 0.9 5.3
Lepidoptera 0.1 0.6
Coleoptera 0.1 1.6
Misc. terrestrial 0.1 11.2
Amphizoidae <0.1 1.7
Curculionidae <0.1 <0.1
Dytiscidae 0.2 1.4
Elmidae <0.1 0.2
Hydrophilidae 0.2 2.7
Ametor <0.1 0.4
Staphylinidae 0.1 0.5
Diptera 1.5 2.5
Ceratopogonidae <0.1 <0.1
Chaoboridae 0.1 0.1
Chironomidae 34.6 13.5
Culicidae 0.1 0.2
Dixidae 2.1 1.0
Empididae 0.3 1.4
Ephydridae <0.1 <0.1
Psychodidae 0.1 0.1
Sciomyzidae 0.1 0.2
Simulidae 0.5 0.6
Tipulidae 0.2 1.5
Dicranota <0.1 <0.1
Tipula <0.1 0.2
Hymenoptera 0.2 0.3
Formicidae 0.1 0.3
Mollusca
Gastropoda 0.2 0.3
Nematomorpha <0.1 <0.1
aMacroinvertebrates were identified to the lowest reliable taxa (i.e.individuals that could not be positively identified to a certain taxo-nomic lever were assigned to the next higher category), so percentrelative abundance by count or biomass of a higher taxon (i.e. fam-ily) does not include those from the taxon or taxa below them (i.e.genus).
greater total invertebrate export during the spring andsummer than fall (Fig. 6a), but there was no discern-able pattern in organic matter transport across seasons(Fig. 6b).
Figure 5. Mean detritus mass, across 1–82% red alder canopy cov-erage, exported from headwater streams on Prince of Wales Island,Alaska. Error bars represent one standard error (n = 5 for each of13 means). r and P -values are for logarithm-transformed data.
Figure 6. Mean macroinvertebrate biomass (a) and detritus (b)exported seasonally from headwater streams on Prince of WalesIsland, Alaska. Error bars represent one standard error (n = 13).
Discussion
The fishless headwaters in this study provided down-stream fish-bearing food webs with invertebrates anddetritus, and riparian red alder along these streamsaugmented this subsidy – more alder translated intomore export. These results were consistent with previ-

160
ous findings where fishless, headwater streams drain-ing mountainous terrain throughout coastal southeast-ern Alaska were discovered to be food conduits fordownstream fish habitats (Wipfli & Gregovich, 2002),and that forest management, including dense coniferand red alder regeneration, influenced this subsidy(Piccolo & Wipfli, 2002). Allochthonous input fromriparian plants is an important food source for shred-ders in streams and may limit shredder communities(Cummins et al., 1989; Richardson, 1991). Broadleafspecies, particularly alder, are preferred by aquatic de-tritivores (Iversen, 1974; Friberg & Jacobsen, 1994).The higher quality leaf litter can increase invertebrategrowth and abundance (Iversen, 1974; Sedell et al.,1975), potentially leading to more export, as measuredin this study. This invertebrate (terrestrial and aquatic)and detrital subsidy should benefit the receiving foodwebs (Polis et al., 1997; Wipfli, 1997; Nakano et al.,1999; Kawaguchi & Nakano, 2001; Allan et al., 2003)as documented with marine-based subsidies for ju-venile salmonids in streams in the Pacific Northwest(Bilby et al., 1998) and Alaska (Wipfli et al., 2003).The freshwater ecosystems along the northern westcoast of North America are probably, at least in part,food limited (Chapman, 1966; Mundie, 1974). Foodabundance therefore likely plays a major role in reg-ulating salmonid production in these systems as hasbeen shown elsewhere (Hayes et al., 2000). Althoughhabitat likely sets the boundaries within which fishproduction occurs, prey abundance probably plays animportant role in governing fish production withinthese physical limits.
The increased prey and detritus export presum-ably resulting from riparian alder may be a reflectionof greater upstream ecosystem productivity (Alaback,1982). We caution however that actual cause-effectrelationships are difficult in retrospective studies likethis because there is no way to be absolutely certainthat the higher food transport was a result of alder orfrom a factor or factors that contributed to alder regen-eration in the first place. But if stream and riparianproductivity is higher with increasing alder densit-ies as these data suggest, whether it be a result ofhigher organic matter quality, more sunlight penetra-tion through the alder canopy, more nitrogen or othernutrients (Hibbs et al., 1994), or factors indirectly re-lated to alder (e.g., soil fertility), then the managementof riparian zones in headwater uplands where alder ispresent could have profound effects on the fish and fishhabitats they fuel. Although speculative, maintaininga certain level of alder in headwaters may help sustain
more productive food webs downstream. Headwaterstreams are abundant in southeastern Alaska (Wipfli& Gregovich, 2002) and comprise 70–90% of channelnetworks there and throughout the Pacific Northwest(Benda & Dunne, 1997; Gomi et al., 2002). Theytherefore have great potential to influence downstreamhabitats, affecting fish and other species in thosehabitats.
The phenomenon of invertebrate drift is whatmakes the subsidy of invertebrates from headwatersto salmonid habitats possible. And although drift dis-tance (Brittain & Eikeland, 1988; Allan, 1995) willhelp determine whether an invertebrate enters fish hab-itat downstream, from a consumer (fish) perspectiveit does not matter where a given food item actuallyoriginates (e.g., 1 km vs. 0.1 m away). The importantresult of this phenomenon is that a portion of up-stream (in-stream or riparian) production from fishlesshabitats enters food webs inhabited by fish or otherconsumers. Drift distance will certainly dictate howmuch total and species-specific biomass is exportedfrom the donor habitat to the receiving habitat, butfor any given food transported across the boundarybetween these two habitats, distance traveled (i.e., driftdistance) is irrelevant.
To be most meaningful, estimates of prey from anygiven source (in this case from headwaters) need tobe put into the context of other prey sources. Prey de-livery to fish habitats from headwaters in southeasternAlaska is about 1.2 mg dry mass m−2 d−1 (yearly aver-age) in streams with riparian zones not influenced bytimber harvesting (Piccolo & Wipfli, 2002; Wipfli &Gregovich, 2002). Aquatic invertebrate production ap-pears to be about 2.8 mg dry mass m−2 d−1 for south-eastern Alaska salmonid rearing tributaries (Duncanet al., 1989). Allochthonous prey subsidies to thesestreams in southeastern Alaska can be high during cer-tain times of the season, ranging 10 to 75 mg dry massm−2 d−1 (Wipfli, 1997; Allan et al., 2003), dwarfingother sources. Considering these other sources, head-waters may provide a relatively small portion of preyto fish habitats on an annual basis, but there may besome important seasonal and spatial considerations tothis prey resource. Certain fishes and other speciesmay take advantage of a localized and reliable sourceof prey due to limited food supplies elsewhere (Groot& Margolis, 1991; Bramblett et al., 2002). In thisstudy terrestrial invertebrate subsidies were consist-ent across seasons, whereas aquatic-derived prey werehighly variable. Wipfli & Gregovich (2002) found thatheadwaters are a year-round source of invertebrates

161
and detritus for downstream habitats in southeasternAlaska. In contrast, marine subsidies are also im-portant for some freshwater systems, but usually onlyduring brief periods (Bilby et al., 1998; Wipfli et al.,1998).
Drift density will also influence the amount of in-vertebrates (and detritus) delivered from headwaters.Invertebrate drift densities in headwater streams in thisstudy were low (<1–4 individuals m−3 water) relat-ive to streams in other parts of the world (<1–116individuals m−3 water) (O’Hop & Wallace, 1983; Al-lan, 1995; Giller & Malmqvist, 1998). Organic mattertransport was also much lower in these southeasternAlaska headwater streams (0.01–0.07 g m−3 water)than those reported elsewhere (0.5–4.0 g m−3 water)(Wallace et al., 1982; Cuffney & Wallace, 1988; Wal-lace et al., 1991). This suggests that headwater streamswithin watersheds in areas where invertebrate driftdensities and detritus export are higher may have evenmore potential to subsidize downstream food webs,provided such streams are abundant.
Conclusion
These results have important implications for resourcemanagement in southeastern Alaska, and through-out the world. We detected significant relationshipsbetween the amount of invertebrates and detritus ex-ported from headwater streams and the density ofred alder along those streams. Similar patterns withforest vegetation and wildlife abundance were also de-tected in the broader, ecosystem-level study where amore abundant forest understory and greater abund-ance of songbirds were detected (Wipfli et al., 2003).Red alder is widely distributed throughout the westcoast of North America (Hibbs et al., 1994), andother species of alder (with similar biological proper-ties and presumably similar ecological effects) occurthroughout the world (Murai, 1964). Developing ormaintaining an alder component in forested ecosys-tems affects plant and animal species, populationsand communities (Harrington et al., 1994; McComb,1994). Regenerating alder following disturbances suchas clearcutting may help sustain food web productivityin, along, and away from streams, at least until thealder are topped by later successional species (Hibbset al., 1994; Piccolo & Wipfli, 2002). Further, un-derstanding how resource management practices andnatural disturbances influence alder regeneration andtrophic pathways in these food webs may aid man-
agers in making decisions when confronted with man-agement options that include alder. Food resources aswell as habitat (i.e., shelter) are clearly both importantfor sustaining fish and wildlife resources.
This study illustrated the trophic linkages betweenupland forests, headwater streams and their riparianzones, and downstream fish habitats lower in the drain-age. It also demonstrated how upland forest regenera-tion (i.e., red alder stand density) appears to affect thedelivery of invertebrates and detritus to downstreamfood webs. Further, it raised questions regarding for-ested headwater ecosystems, including the influenceof upland and riparian forests on headwater streamproductivity (i.e., Does more invertebrate and de-tritus transport actually mean greater aquatic/riparianproductivity in headwaters, or is this elevated ex-port a symptom of something else?), forest-streamlinkages (i.e., Where are the terrestrial invertebratesthat fall into streams coming from: canopy, under-story, forest floor litter, or soil-born, or even dispersalor migrants from distant habitats?), hyporheic-streamlinkages (i.e., How much of the transported aquaticbiomass is actually coming from the streambed v. thehyporheos, and are hyporheic habitats important refu-gia for invertebrates during dry periods of little or nosurface flow in these headwater habitats?), headwater-downstream linkages (i.e., How far downstream doinvertebrates and detritus travel?), fish effects (i.e.,How much food delivered from fishless headwaters tofish-bearing stream reaches is actually available to andassimilated by consumers, and is this subsidy essen-tial for supporting downstream production?), temporaland spatial effects (i.e., How do these subsidies fromheadwaters vary seasonally, and across drainages andecoregions?), and forest management effects (Howdoes timber harvesting and subsequent forest succes-sional pathways, both short and long-term, influencetrophic linkages?). Finally, how important to foodwebs is this source of invertebrates and detritus relat-ive to other sources (e.g., other allochthonous inputs)?With greater attention being placed on headwatersand the effects of associated resource management onthese habitats (Meyer & Wallace, 2001; Gomi et al.,2002), answering these and related questions is vital toimproving our understanding of headwater ecosystemfunction and their interactions with downstream andadjoining habitats, and when and how these forestedheadwater ecosystems should be managed in Alaska,and throughout the world.

162
Acknowledgements
Thanks to the USDA Forest Service Craig RangerDistrict for technical support, Tim Max for statisticaladvice, Maria Lang for field and laboratory assist-ance, Takashi Gomi for providing an earlier versionof Figure 1 and Adelaide Johnson for providing somephysical characteristics data for Table 1. We thankEwa Orlikowska for canopy coverage and basal areaestimates. Thanks to J. David Allan, Jack Piccolo,Carol Volk and Richard Woodsmith for their insight-ful reviews of earlier drafts. This research was fundedby the Wood Compatibility Initiative (WCI), USDAPacific Northwest Research Station, Portland, OR.
References
Alaback, P. B., 1982. Dynamics of understory biomass in Sitkaspruce – Western hemlock forests of Southeast Alaska. Ecology63: 1932–1948.
Allan, J. D., 1995. Stream Ecology: Structure and Function ofRunning Waters. Chapman & Hall, London.
Allan, J. D., M. S. Wipfli, J. P. Caouette, A. Prussian & J. Rodgers,2003. Influence of streamside vegetation on terrestrial inver-tebrate subsidies to salmonid food webs. Canadian Journal ofFisheries and Aquatic Sciences 60: 309–320.
Benda, L. & T. Dunne, 1997. Stochastic forcing of sediment supplyto the channel network from landsliding and debris flow. WaterResources Research 33: 2849–2863.
Beschta, R. L., R. E. Bilby, G. W. Brown, L. B. Holtby &T. D. Hofstra, 1987. Stream temperature and aquatic habitat:fisheries and forestry implications. In Salo, E. O. & T. W.Cundy (eds), Streamside Management: Forestry and Fishery In-teractions. Contribution 57. University of Washington, Seattle:191–232.
Bilby, R. E. & P. A. Bisson, 1998. Function and distribution oflarge woody debris. In Naiman, R. J. & R. E. Bilby (eds),River Ecology and Management. Springer Publishers, New York:324–346.
Bilby, R. E., B. R. Fransen, P. A. Bisson & J. K. Walter, 1998.Response of juvenile coho salmon (Oncorhynchus kisutch) andsteelhead (Oncorhynchus mykiss) to the addition of salmon car-casses to two streams in southwestern Washington, U.S.A. Cana-dian Journal of Fisheries and Aquatic Sciences 55: 1909–1918.
Binkley, D., K. Cromack & D. D. Baker, 1994. Nitrogen fixationby red alder: biology, rates, and controls. In Hibbs, D. E., D. S.DeBell & R. F. Tarrant (eds), The Biology and Management ofRed Alder. Oregon State University Press, Corvallis: 57–72.
Bisson, P. A. & R. E. Bilby, 1998. Organic matter and trophic dy-namics. In Naiman, R. J. & R. E. Bilby (eds), River Ecology andManagement. Springer Publishers, New York: 373–398.
Bjornn, T. C. & D. W. Reiser, 1991. Habitat requirements ofsalmonids in streams. American Fisheries Society Special Pub-lication 19: 83–138.
Borchardt, M. A., 1996. Nutrients. In Stevenson, R. J., M. L. Both-well & R. L. Lowe (eds), Algal Ecology. Academic Press, SanDiego: 183–227.
Bramblett, R. G., M. D. Bryant & B. E. Wright, 2002. Seasonalmovements and habitat use by juvenile steelhead in Southeast
Alaska. Transactions of the American Fisheries Society 131:498–506.
Brittain, J. E. & T. J. Eikeland, 1988. Invertebrate drift – A review.Hydrobiologia 166: 77–93.
Burgherr, P. & E. I. Meyer, 1997. Regression analysis of linear bodydimensions vs. dry mass in stream macroinvertebrates. Archivfür Hydrobiologie 139: 101–112.
Chapman, D. W. 1966. Food and space as regulators of salmonidpopulations in streams. American Naturalist 100: 345–357.
Cuffney, T. F. & J. B. Wallace, 1988. Particulate organic matterexport from three headwater streams: discrete versus continu-ous measurements. Canadian Journal of Fisheries and AquaticSciences 45: 2010–2016.
Cummins, K. W., M. A. Wilzbach, D. M. Gates, J. B. Perry &W. B. Taliaferro, 1989. Shredders and riparian vegetation: leaflitter that falls into streams influences communities of streaminvertebrates. BioScience 39: 24–30.
Duncan, W. F., M. A. Brusven & T. C. Bjornn, 1989. Energy flowresponse models for evaluation of altered riparian vegetation inthree Southeast Alaskan streams. Water Research 23: 965–974.
Friberg, N. & D. Jacobsen, 1994. Feeding plasticity of twodetritivore-shredders. Freshwater Biology 32: 133–142.
Giller, P. S. & B. Malmqvist, 1998. The Biology of Streams andRivers. Oxford University Press, New York.
Gomi, T., R. C. Sidle & J. S. Richardson, 2002. Understand-ing processes and downstream linkages of headwater systems.BioScience 52: 905–916.
Gregory, S. V. & P. A. Bisson, 1997. Degradation and lossof anadromous salmonid habitat in the Pacific Northwest. InStouder, D. J., P. A. Bisson, & R. J. Naiman (eds), Pacific Salmonand their Ecosystems: Status and Future Options. Chapman andHall, New York: 277–314.
Groot, C. & L. Margolis, 1991. Pacific Salmon Life Histories.University of British Columbia Press, Vancouver.
Harrington, C. A., J. C. Zasada & E. A. Allen, 1994. Biology ofred alder (Alnus rubra Bong.). In Hibbs, D. E., D. S. DeBell &R. F. Tarrant (eds), The Biology and Management of Red Alder.Oregon State University Press, Corvallis, OR: 116–123.
Hayes, J. W., J. D. Stark & K. A. Shearer, 2000. Development andtest of a whole-lifetime foraging and bioenergetics growth modelfor drift-feeding brown trout. Transactions of the AmericanFisheries Society 129: 315–332.
Hibbs, D. E., D. S. DeBell & R. F. Tarrant, 1994. The Biologyand Management of Red Alder. Oregon State University Press,Corvallis, OR.
Hulten, E., 1968. Flora of Alaska and Neighboring Territories.Stanford University Press, Stanford, CA.
Iversen, T.M., 1974. Ingestion and growth in Sericostoma person-atum (Trichoptera) in relation to the nitrogen content of ingestedleaves. Oikos 25: 278–282.
Kawaguchi, Y. & S. Nakano, 2001. Contribution of terrestrial inver-tebrates to the annual resource budget for salmonids in forest andgrassland reaches of a headwater stream. Freshwater Biology 46:303–316.
McComb, W. C., 1994. Red alder: interactions with wildlife. InHibbs, D. E., D. S. DeBell & R. F. Tarrant (eds), The Biologyand Management of Red Alder. Oregon State University Press,Corvallis, OR: 131–138.
Meyer, J. L. & J. B. Wallace, 2001. Lost linkages and lotic ecology:rediscovering small streams. In Press, M. C., N. J. Huntly & S.Levin (eds), Ecology: Achievement and Challenge. BlackwellScientific Publications, Oxford: 295–317.

163
Mundie, J. H., 1974. Optimization of the salmonid nursery stream.Journal of the Fisheries Research Board of Canada 31: 1827–1837.
Murai, S., 1964. Phytotaxomomical and geobotanical studies ongenus Alnus in Japan. III. Taxonomy of whole world species anddistribution of each sect. Government Forest Experiment StationBulletin, Japan 171: 1–107.
Naiman, R. J. & R. E. Bilby, 1998. River Ecology and Management.Springer Publishers, New York, New York.
Nakano, S., H. Miyasaka & N. Kuhara, 1999. Terrestrial–aquaticlinkages: riparian arthropod inputs alter trophic cascades in astream food web. Ecology 80: 2435–2441.
Newton, M. & E. C. Cole, 1994. Stand development and succes-sional implications: pure and mixed stands. In Hibbs, D. E., D.S. DeBell & R. F. Tarrant (eds), The Biology and Managementof Red Alder. Oregon State University Press, Corvallis, Oregon:106–115.
NRC (National Research Council), 1996. Upstream: Salmon andSociety in the Pacific Northwest. National Academy Press,Washington, DC.
O’Hop, J. & J. B. Wallace, 1983. Invertebrate drift, discharge, andsediment relations in a southern Appalachian headwater stream.Hydrobiologia 98: 71–84.
Piccolo, J. J. & M. S. Wipfli, 2002. Does red alder (Alnus rubra)along headwater streams increase the export of invertebrates anddetritus from headwaters to fish-bearing habitats in southeasternAlaska? Canadian Journal of Fisheries and Aquatic Sciences 59:503–513.
Polis, G. A., W. B. Anderson & R. D. Holt, 1997. Toward an in-tegration of landscape and food web ecology: the dynamics ofspatially subsidized food webs. Annual Review of Ecology andSystematics 28: 289–316.
Polis, G. A. & S. D. Hurd, 1996. Linking marine and terrestrial foodwebs: allochthonous input from the ocean supports high second-ary productivity on small islands and coastal land communities.American Naturalist 147: 396–423.
Richardson, J.S., 1991. Seasonal food limitation of detritivores in amontane stream: an experimental test. Ecology 72: 873–887.
Rogers, L. E., R. L. Buschbom & C. R. Watson, 1977. Length–weight relationships of shrub-steppe invertebrates. Annals of theEntomological Society of America 70: 51–53.
Sample, B. E., R. J. Cooper, R. D. Greer & R. C. Whitmore, 1993.Estimation of insect biomass by length and width. AmericanMidland Naturalist 129: 234–240.
SAS Institute, 1998. SAS/STAT User’s Guide, Version 6, 4th edn.SAS Institute Inc., Cary, NC.
Sedell, J. R., F. J. Triska & N. S. Triska, 1975. The processing ofconifer and hardwood leaves in two coniferous forest streams: I.Weight loss and associated invertebrates. Verhandlungen interna-tionale Vereinigung Limnologie 19: 1617–1627.
Southwood, T. R. E., 1961. The number of species of insect as-sociated with various trees. Journal of Animal Ecology 30:1–8.
USDA, 1999. Record of decision: Tongass National Forest land andresource management plan. United States Forest Service, PacificNorthwest Research Station, Alaska Region, FS–639.
Wallace, J., B., T. F. Cuffney, J. L. Webster, G. J. Lugthart, K.Chung & B. S. Goldowitz, 1991. Export of fine organic particlesfrom headwater streams: effects of seston, extreme discharges,and invertebrate manipulation. Limnology and Oceanography36: 670–682.
Wallace, J. B., H. R. Ross & J. L. Meyer, 1982. Seston anddissolved organic carbon dynamics in a southern Appalachianstream. Ecology 63: 824–838.
Waters, T. F., 1995. Sediment in Streams: Sources, Biological Ef-fects and Control. American Fisheries Society Monograph 7.Bethesda, Maryland.
Wipfli, M. S., 1997. Terrestrial invertebrates as salmonid prey andnitrogen sources in streams: contrasting old-growth and young-growth riparian forests in southeastern Alaska, U.S.A. CanadianJournal of Fisheries and Aquatic Sciences 54: 1259–1269.
Wipfli, M. S. & D. P. Gregovich, 2002. Export of invertebrates anddetritus from fishless headwater streams in southeastern Alaska:implications for downstream salmonid production. FreshwaterBiology 47: 957–969.
Wipfli, M. S., J. P. Hudson & J. P. Caouette, 1998. Influence ofsalmon carcasses on stream productivity: response of biofilm andbenthic macroinvertebrates in southeastern Alaska, USA. Cana-dian Journal of Fisheries and Aquatic Sciences 55: 1503–1511.
Wipfli, M. S., R. L. Deal, P. E. Hennon, A. C. Johnson, R. T. Ed-wards, T. L. De Santo, T. Gomi, E. H. Orlikowska, M. D. Bryant,M. E. Schultz, C. LeSage, R. Kimbirauskus & D. V. D’Amore,2003. Compatible management of red alder-conifer ecosystemsin southeastern Alaska. In Monserud, R. A., R. Haynes & A.Johnson (eds), Compatible Forest Management. Kluwer Aca-demic Publishers, Dordrecht, The Netherlands: 55–81.
Wipfli, M. S., J. P. Hudson, J. P. Caouette & D. T. Chaloner,2003. Marine subsidies in freshwater ecosystems: salmon car-casses increase the growth rates of stream-resident salmonids.Transactions of the American Fisheries Society 132: 371–381.





























