Embed Size (px)
Citation preview

8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 1/6
Brief Definitive Report
C T L A - 4 : A Ne g a t iv e R e g u l a t o r o f A u t o i m m u n e D i s e as e
By Nitin J. Karandikar,* Carol L.Vanderlugt,* Theresa L.Walunas,*Stephen D. Miller,*5 and Jeffrey A. Bluestone*g
From the *Departm ent of Microbiology-Immunology and Interdepartmental Immunobiology Center,
Northwestern University Medical School, Chicago, Illinois 6061 I; and *The Ben May Institute and
the Committee of Immunology, The University of Chicago, Chicago, Illinois 6063 7
Summary
CTLA-4, a CD28 homologue expressed on act ivated T cel ls , binds wi th high aff ini ty to the
CD 28 l igands, B7-1 (CD80) and B7-2 (CD86). Th is study was designed to examine the role of
CTLA-4 in regula t ing autoimmune disease . Murine re lapsing-remit t ing experimental autoim-
mu ne encephalomyel i t is (R-EAE ) is a demyel inat ing disease m ediated by PLP139-151-specif ic
CD 4 + T cel ls in sJL/J m ice . An t i -C TLA -4 m Abs (or thei r F(ab) fragments) enhanc ed in vi t ro
pro l ife rat ion and p ro- in f lammatory cy tok ine p roduc t ion by PLP139-151-pr imed lymph node
cells. Add ition of either reagent to in vitro activation cultures poten tiated the ability of T cells
to adoptiv ely transfer disease to naive recipients. In vivo administration of anti-C TL A- 4 mA bto recipients o f PLP13 9-151-s pecific T cells resulted in accelerated and e xacerba ted disease. Fi-
nally, ant i -CT LA -4 t rea tment of mice during disease remission resul ted in the exacerbat ion of
relapses. Collectively , these results suggest that CT LA -4 m ediates the dow nreg ulatio n o f on go -
ing imm une responses and plays a major role in regula t ing autoimm unity.
T cell activation requires th e d elivery of at least tw o sig-
nals to the T cell by the antigen-presenting cell
(APC). O ne signal is antigen-specific and is prov ided by the
l igat ion of the T cell receptor (TClq.) by an t igen -M HC
complexes, whereas the second "cost imulatory " signal may
be pro vided by the B7 family of molecules, B7-1 (CD80)or B7-2 (CD86), expressed on the APC. Interact ions be-
tween CD28 on the T cel l and the B7 molecules on the
APC are cri t ica l, both in vivo and in vi t ro, in preven t ing T
cell unresponsiveness (1-4) . The discovery of the CD 28
hom ologu e, cytotox ic T lymph ocyte associated molecule~4
(CTLA-4) , by sub t rac t ive mR N A c lon ing o f cy to tox ic T
cells (5) has increased the complexity of costimulatory inter-
actions. CTLA-4 binds with higher affinity to the B7-1 and
B7-2 molecules (6) . CTLA-4Ig, a soluble recombinant
fo rm of CTLA -4 , is a po ten t i nh ib i to r o f B7 /CD 28 in t e r -
actions and inhibits in vivo antigen-specific responses (2, 3,
7, 8). Ho wev er , the regula tory funct ion o f the nat ive
CTLA-4 molecule on the T cel l surface has only recent lybeen apprecia ted.
The cy top lasmic t ai l o f human and mur ine CTL A-4 is
highly conserved (9), suggesting that i t is involved in an
impor tant signaling function. Surface expression of CT LA -4
seems to be a t ightly regulated event with transient expres-
sion on activated T cells, peaking at 48-72 h after activa-
tion (10, 11). Initial studies suggested that anti-CTLA-4
Drs. Miller and Bluestoneare co-seniorauthors.
ant ibodies could synergize wi th ant i -CD3 and ant i -CD28
to enhance T cell proliferation (10, 12). Thus, i t was con-
cluded that CTL A- 4 was an addi t ional cost imulatory mo le-
cule that had funct ions cooperat ive wi th those o f CD 28.
However , recent studies have indicated that CTLA-4 may,
in fact, serve as a negative regu lator o f T cell activation. Wehave shown that soluble ant i -CTLA-4 mAb as wel l as i t s
F(ab) fragments increased T cell proliferation in murine
ML1L responses (11). A dditionally, intact cross-linked anti-
CTLA-4 mAb, but not i t s F(ab) fragments, could inhibi t
proliferation in T cells stimulated with anti-CD3 and anti-
CD28 (11, 13). Thus, i t is l ikely that CTLA-4 functions to
downregulate T cell responses such that blockade of CT LA -
4/B 7 interaction results in enhance d T cell activation. Oth er
studies also support a negative regulatory role for CTLA-4.
Cross- l inking of CT LA -4 af ter the l igation of CD3 and
CD28 st rongly inhibi ted prol i fera t ion and IL-2 product ion
(14). In vivo t rea tment of mice with ant i -C TLA -4 was
shown to enh ance c lonal expansion of ant igen-specif ic Tcel ls af ter immunizat ion (15) and enhance ant i - tumor im-
muni ty (16) . In fac t , CTLA-4 knockout mice express a
lymphoproliferative disorder resulting in massive infiltra-
tion o f several organs leading to early death (17, 18). T o-
gether, these studies support an essential role for CTLA-4
in the do wn regu lation of potentially autoaggressive T cells.
Th e present study was con ducted to di rectly examine the
role of CTLA-4 in regulating autoantigen-specific responses
in autoimmune disease . Relapsing-remit t ing experimental
78 3 j. Exp . Med. 9 The Rockefeller University Press 90022-1007/96/08/783/06 $2.00Volume 184 August 1996 783-788
Published August 1, 1996

8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 2/6
a u t o i m m u n e e n c e p h a l o m y e l i t i s ( R- E A E) i s a w e l l - e s t a b -
l i s h ed n m r i n e m o d e l f o r m u l t i p l e s c le r os i s (M S) c h a r a c t e r -
i z ed b y C D 4 + T c e l l - m e d i a t e d d e m y e l i n a t i o n i n t h e c e n -
t ra l n e r v o u s s y s t e m ( CN S) ( 19 ). R- EA E i s i n d u c e d i n SJ L / J
m i c e by i m m u n i z a t i o n w i th t h e i m m u n o d o m i n a n t e p i t o p e
o n p r o t e o l i p i d p r o t e i n ( PLP13 9 - 15 1 ) o r b y t h e a d o p t i v e
t ransfer of PLP 139-15 1-spec i f i c T ce l ls (20) . The c l in ica l
d i sease i s charac ter ized by an acute para ly t ic phase fo l lowed
b y r e c o v e r y a n d s u b s e q u e n t r e la p s e s a c c o m p a n i e d b y d e -v e l o p m e n t o f T c e l l r e s p o ns e s to e n d o g e n o u s e p i t o p e s o n
m y e l i n p r o t e i n s ( e. g ., PLP17 8 - 19 1 ) , a p h e n o m e n o n t e r m e d
e p i t o p e s p re a d in g ( 21 , 2 2) . C D 2 8 / B 7 - m e d i a t e d c o s t im u l a -
tory in te rac t ions p lay a major ro le in the es tabl i shment and
p r o g re s s io n o f P , - E A E . T r e a t m e n t o f m i c e w i t h C T L A - 4 I g
h a s b e e n s h o w n t o i n h i b i t d i se a s e i n d u c t i o n ( 8) . A n t i - B 7 - 1
o r a n t i - B7 - 2 t r e a t m e n t s h a v e d i f f er e n t ia l ef fe c ts o n a u t o i m -
m u n e d i s e as e s i n c l u d i n g R- E A E ( 2 3, 24 ) a n d a u t o i m m u n e
diabe tes (25) . Together , these s tudies emphas ize the com-
p l e x i t y o f c o s t i m u l a t o r y i n t e r a c ti o n s b e t w e e n B7 m o l e c u l e s
a n d CD 2 8 o r CT LA - 4 . I n th i s s t u d y , w e h a v e d i re c t ly a d -
d r es s ed t h e r o l e o f CT LA - 4 i n m o d u l a t i n g PLP 139 - 15 l - s p e -
c i f ic T ce l l r esponses and in regu la t ing d i sease us ing an a nt i -C T L A - 4 m A b , k n o w n t o p r e v e n t C T L A - 4 / B 7 i n t e r a c t i o n s .
Both in tac t Ig and F(ab) f ragments had s imi la r e f fec t s in a l l
the responses s tudied , sugges t ing tha t these reagents were
b l o c k i n g C T L A - 4 - m e d i a t e d e v e n t s . F i r s t , a n t i - C T L A - 4
m A b e n h a n c e d i n v i t r o p r o l i f e r a t i o n a n d p r o d u c t i o n o f
p r o - i n f l a m m a t o r y c y to k i n es b y P L P 1 3 9 - 1 5 1 - p r i m e d l y m p h
n o d e c ei ls . S e c o n d , a d d i t i o n o f a n t i - C T L A - 4 t o i n v it r o a c -
t iva t ion cul tures resul t ed in potent ia t ion o f the abi l i ty of these
ce ll s to adopt iv e ly transfer R-E AE to na ive rec ip ient s . Thi rd ,
i n v i v o b l o c k a d e o f CT LA - 4 i n r e c i p i e n t m i ce , a f t er a d o p -
t ive t ransfer of pr i me d ce l l s , a l so resul t ed in exa cerba t ion o f
di sease . Fina l ly , t r ea tm ent of mice dur in g rem iss ion , a f t e r
t h e a c u t e p h a s e o f R . - EA E r e s u l te d i n g r e a t e r i n c i d e n c e a n d
sever i ty of re lapses . These resul t s indica te tha t C TL A- 4 has
a n i m p o r t a n t r o l e in t h e d o w n r e g u l a t i o n o f T c e l l r e s p o ns e s
i n a u t o i m m u n e d i s e a s e a n d m a y b e a m a j o r r e g u l a t o r y
p l a y er in t h e c o n t r o l o f o n g o i n g i m m u n o p a t h o l o g y .
Materia ls and M ethod s
Mice. Female SJL/J mice, 6-7 wk old, were purchased from
Harlan Laboratories (Indianapolis, IN), housed in the North west-
ern animal care facility, and maintained on standard laboratory food
and water ad libitum. Paralyzed mice were afforded easier access
to food and water .
Peptides. PLP139-151 (HSLGK WLGH PDKF) was purchased
from Peptides Internat ional (Louisville, KY). A mino acid comp o-sition was verified by mass spectrometry and purity (>98%) was
assessed by HP LC .
In Vitro Proliferation and Cytokine A ssay s. Mice were immu-
nized subcutaneously with 100 ~1 of a Complete Freund's Adju-
vant (CFA) emulsion containing 200 ~,g of Mycobacterium tubercu-
losis H37Ra (Difco Labs. Inc. Detroit, MI) and 25 btg of
PLP13 9-151 distr ibuted over three spots on the f lank. On day
10, draining lymph node cells were harvested and cultured in 96-
well micro titer plates at a density of 5 • 105 cells/well in a total
volume of 200 ~xl Dulbecco's M odified Eagle Medium (DME)
containing 7% fetal bovine serum, 1 mM glutamine, 1% Pen-
Strep, 1 mM nonessential amin o acids and 5 X 10 -5 M 2 -me r-
captoethanol (complete DME-7; al l products from Sigma Chem.
Co. , St. Louis, MO ). Ce lls were allow ed to incubate at 37~ for
60 min w ith the indicated ant ibodies (10 ~g/m l) before the indi-
cated doses of antigen were added. Cultures were pulsed with 1 IxCi
of [3H]TdR after 72 h, harvested at 96 h and [3H]TdR uptake
determ ined by scintillation counts. Results are expressed as mean
of triplicate cultures -+ SEM (backgrou nd counts sub tracted). Su-
pernatants collected at 24 and 48 hours from replicate cultures
were assayed for IL-2, IL-4, and IFN -y levels using ELISA Mini-
Kits (Endogen, Cambridge, MA). Minimal detect ion l imits of the
assays were "~ pg/ ml for IL-2 and IL-4 and 30 pg/nfl for IFN- %
Flow Cytometry. FAC S analysis was perfo rmed using the
FAC Scan flow cytomete r as described previously (11, 24). PE-
conjugated ant i-CD3 and FITC-conjugated control Ig were pur-
chased from PharM ingen (La Jolla, CA).
Induction of R-E AE by Active Immunization with PLP13 9-
151. Mice were immunized with 80 txg PLP139 -151 in CFA
as previously described (20).
Adoptive Transfer qf R- EA E. D o n o r mice were immunized
with 25 ~g of PLP139 -151 in CF A. D raining lymph node cel ls
were harvested on day t0 and cultured in ei ther T25 ( total vol-
ume : 15 ml) or T75 (30 ml) flasks at 8 • 1()~' cells/ml with 20#,g/ml of PLP139-15 1. In ant ibody-containing cultures, cells
were incubated at 37~ for 60 rain with the indicated mAb s (11)
L~g/ml) before th e add ition of antigen. O n day 4, cells were
washed in PBS, counted, and transferred i.p. into naive recipients
in a total volume o f 0.5 ml PBS. R ecipient m ice received a sub-
optimal dose of 6-7 • 106 live cells each.
Antibody Treatments. The fol lowing mAbs were employed:
hamster control lg (Cappel Research Products, Cochranvil le, PA)
and the ant i -CTL A-4 mAb (UC l l -4F 10) (11). Ant ibodies and
F(ab) fragments were produced and purified as previously de-
scribed (11, 25). Mice were treated, as indicated, either starting
on the day of adoptive transfer or after recovery from the initial
paralytic episode of R-EA E (~25 d post- immunizat ion). 50 txg
of antibody was administered i.p. every other day for five treat-
ments (250 txg total) and m ice w ere m onito red fo r clinical signsof disease for an additional 20-40 d.
Clinical Evaluation. Clinical severity was assessed on a 0 to 5
scale as described (24). The data are plotted as the daily mean
clinical score for all animals in a particular treatment group. A re -
lapse was defined as a sustained increase of at least one full grade
in clinical score after the animal had previously improved at least
a full clinical score and had stabilized. Mean maximal severity is
the mean of the highest cl inical score reached by mice within an
experim ental group whi ch displayed clinical signs.
Statistical Ana lyses . Comparison of the percentage of animals
showing clinical disease and/or relapses between any two groups
of mice were analyzed by X2 using Fisher's exact probability.
Comparisons of the mean day of onset of disease/relapse, and
mean peak disease severi ty between any two groups of mice w ere
analyzed by the Student's t test. P values <0.05 were considered
significant.
Resul ts and Discuss ion
An ti -C TL A- 4 mAb Enhances PLP139-151-Specific In Vitro
Proliferative and Cyto kine Responses. R e c e n t i n v i t r o s t u d -
i es ha v e e m p l o y e d a l l o g e n e ic M L R o r a n t i - C D 3 s t i m ul a -
t i o n t o a ss ess t h e e x p r e s s i o n a n d f u n c t i o n o f CT LA - 4 o n T
784 Anti-C TLA -4 mAb Treatment Exacerbates Murine EAE
Published August 1, 1996
8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 3/6
HAM. CT. ~ oCTL/b4 ~ oCTLA-4 Fab
zO
wglIL-I
Owa.
A
0 10 20 30 40 50 00 70 80
ACPM (X 104)
z0F.O,-iaOr=.
,. I
0 50 100 150 200 250
pglml
B
3 0 0
z Io
O-sel
OwQ..>
zig
0
I C
500 1000 1500 2000
pg/ml
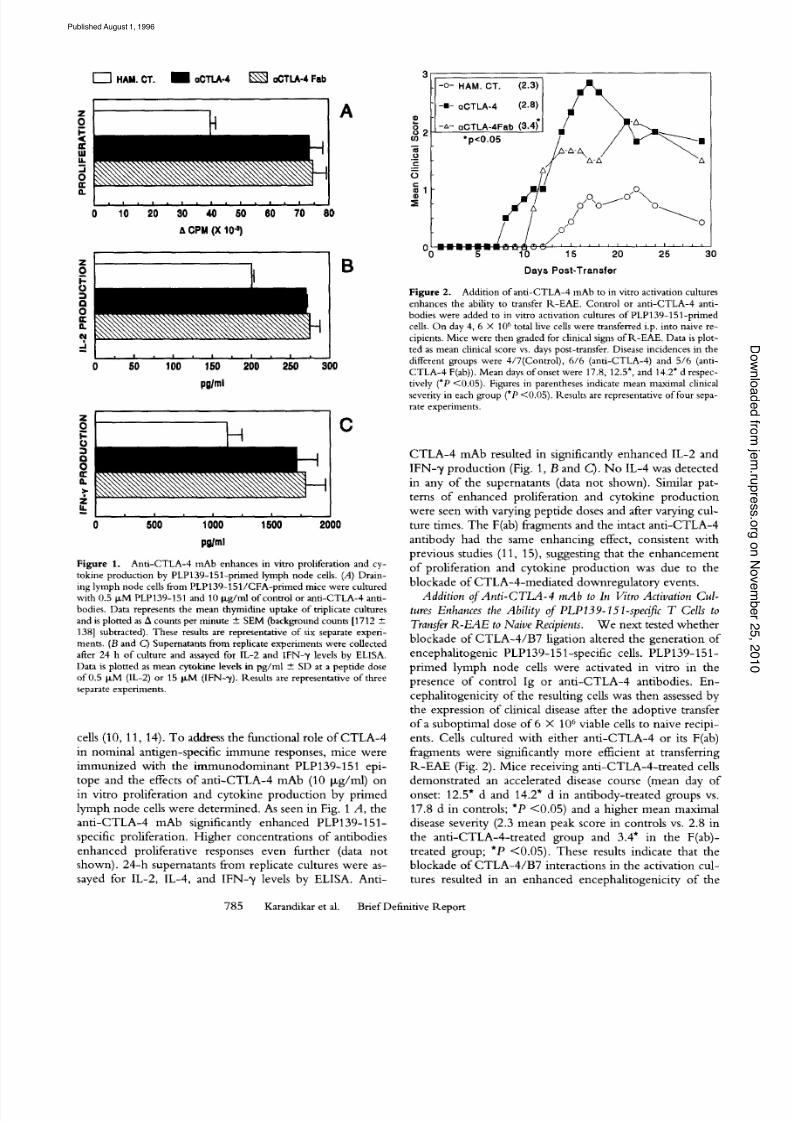
Anti-CTLA-4 mAb enhances in vi tro proliferation and cy-i g u r e 1 .tokine production by PLP139-151-primed ymph node cells. (A) Drain-ing lymph node cells from PLP139-151/CFA-primedmice were culturedwith 0.5 btM PLP139-151 and 10 b~g/mlof control or anti-CTLA-4 anti-bodies. Data represents the mean thymidine uptake of triplicate culturesand is plotted as A counts per minute + SEM (backgroundcounts [1712 +138] subtracted). These results are representative of six separate experi-ments. (B and C) Supernatants rom replicate experiments were collectedafter 24 h of culture and assayed for 1L-2 and IFN-7 levels by ELISA.Data is plotted as mean cytokine levels n pg/ml + SD at a peptide doseof 0.5 I-tM (IL-2) or 15 la,M (IFN-~/). Results are representativeof threeseparate experiments.
cells (10, 11, 14). To address the fu nction al role of CT LA -4
in n ominal antigen-specific imm une responses, mice wereimmunized with the im muno domi nan t PLP139-151 epi-
tope an d the effects of an ti- CTL A- 4 mAb (10 Ixg/ml) on
in vitro proliferation and cytokine production by primed
lymph n ode cells were deter mined. As seen in Fig. 1 A, the
anti -CTL A-4 mAb significantly enhanced PLP139-151-
specific proliferation. Higher concentrations of antibodies
enhanced proliferative responses even further (data not
shown). 24-h supematants from replicate cultures were as-
sayed for IL-2, IL-4, and IFN-"v levels by ELISA. Anti-
3 t -~ HAM 'CT ' (2 '3 )
- I - eCTLA-4 (2 .8 [ B / ~
| ~ I t - " -oCTLA -4 F-b (3 .4 ) ] ,./ /=<~"-..
00 = = = ' = E I = ~ " = ' I ' o e C ' ' 1 5 . . . . 2 0 . . . . 2 5 . . . . 30
Days Post-Transfer
F i g u r e 2. Additionofanti-CTLA-4 ]nAb to in vitro activationculturesenhances the ability to transfer R-EAE. Control or anti-CTLA-4 anti-bodies were added to in vitro activation cultures of PLP139-151-primedcells. On day 4, 6 X 106 total live cellswere transferred i.p. into naive re-cipients. Mice were then graded for clinicalsigns of R-EAE. Data is plot-ted as mean clinicalscore vs. days post-transfer. Disease incidences n thedifferent groups were 4/7(Control), 6/6 (anti-CTLA-4) and 5/6 (anti-
CTLA-4 F(ab)). Mean days of onset were 17.8, 12.5", and 14.2" d respec-tively (*P <0.05). Figures n parentheses indicate mean maximal clinicalseverity in each group (*P <0.05). Results are representativeof four sepa-rate experiments.
CTLA -4 mAb resulted in significantly enhance d IL-2 and
IFN-y production (Fig. 1, B and C). No IL-4 was detected
in any of the supernatants (data not shown). Similar pat-
terns of enhanced proliferation and cytokine production
were seen with var ying peptide doses and after varying cul-
ture times. The F(ab) fragments and the intact anti-CTLA-4
antibody had the same enhancing effect, consistent with
previous studies (11, 15), suggesting that the enhancement
of proliferation and cytokine product ion was due to the
blockade of CTLA-4-m edia ted downregulatory events.
Addition of An ti -C TL A- 4 mAb to In Vitro Activation Cul-
tures Enhances the Abilit y of PLP139-15 1-spec ific T Cells to
Transfer R -E A E to Naive Recipients. We next tested whether
blockade of CTLA -4/ B7 ligation altered the generation of
encephalitogenic PLP139-151-specific cells. PLP139-151-
primed lymph node cells were activated in vitro in the
presence of control lg or anti-CTLA-4 antibodies. En-
cephalitogenicity of the resulting cells was then assessed by
the expression of clinical disease after the adoptive transfer
of a subo ptim al dose of 6 • 106 viable cells to naiv e recipi-
ents. Cells cultured with either anti-CTLA-4 or its F(ab)
fragments were significantly more efficient at transferringR-E AE (Fig. 2). Mice receiving anti-C TLA-4- treat ed cells
demonstrated an accelerated disease course (mean day of
onset: 12.5" d and 14.2" d in ant ibody- treat ed groups vs.
17.8 d in controls; *P <0.05) and a higher mean maximal
disease severity (2.3 mean peak score in controls vs. 2.8 in
the anti-CT LA-4- treat ed group and 3.4" in the F(ab)-
treated group; *P <0.05). These results indicate that the
blockade of CTLA -4/ B7 interactions in the activation cul-
tures resulted in an en hanced encephalitogenicity of the
785 Karandikar et al. Brief Definitive Repo rt
Published August 1, 1996
8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 4/6
c e l l s t r a n s f e r r e d t o n a i v e r e c i p i e n t s . S i n c e a n t i - CTLA - 4
m A b t r e a t m e n t a l s o r e s u l t e d i n e l e v a t e d p r o l i f e r a t i o n a n d
cytokin e produc t ion by PLP13 9-151-spec i f i c T ce ll s (Fig . 1),
t h e e n h a n c e d d i se a s e c o u r s e m a y b e d u e t o t h e t r a n s fe r o f a
h i g h e r n u m b e r o f e f fe c t o r T c e ll s a n d /o r o f e f f ec t o r ce ll s
c a p ab l e o f p r o d u c i n g e n h a n c e d l e ve ls o f p r o - i n f l a m m a t o r y
c y t o k i n e s .
In Vivo Treatment with An ti -C TL A- 4 mAb Exacerbates
Adoptive R-EAE. W e n e x t e x a m i n e d w h e t h e r C T L A - 4 -m e d i a t e d e v e n t s d o w n r e g u l a t e d PLP13 9 - 15 1 - s p e c i f i c T
ce ll s in v ivo . Act iva ted PLP139-15 1-spec i f i c ce l ls were ado p-
t i v e l y t r a n s f e r r e d t o n a i v e S j L / j r e c i p i e n t s . Re c i p i e n t m i c e
w e r e t h e n t r e a t e d w i t h c o n t r o l I g , i n t a c t a n t i - CTLA - 4 , o r
a n t i - C T L A - 4 F ( a b ) f r a g m e n t s . A n t i - C T L A - 4 m A b t r e a t -
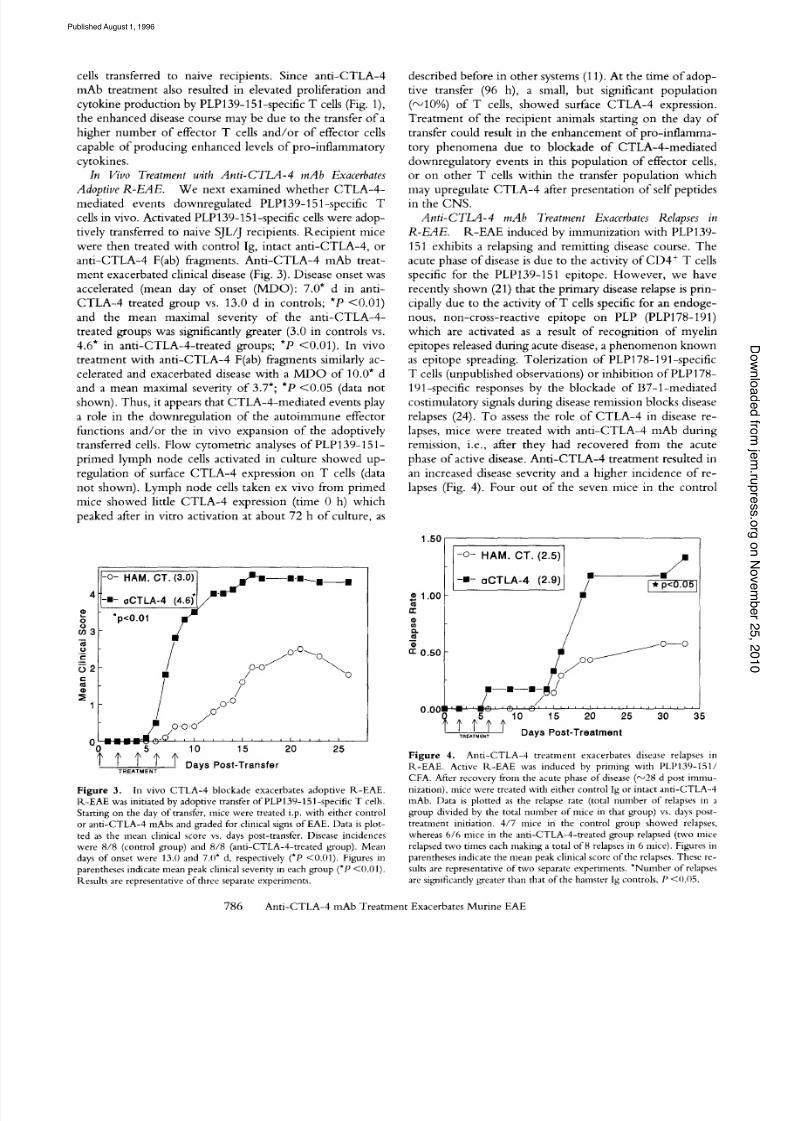
me nt exac erba te d c l in ica l d i sease (Fig . 3) . Disease onse t was
a c c e l e r a t e d ( m e a n d a y o f o n s e t ( M D O ) : 7 .0 * d i n a n t i -
CTLA - 4 t r e a t e d g r o u p v s . 13 .0 d i n c o n t r o l s ; *P < 0 .0 1 )
a n d t h e m e a n m a x i m a l s e ve r it y o f th e a n t i - C T L A - 4 -
t rea ted grou ps was s ign i f icant ly grea ter (3 .0 in c ont ro l s vs .
4 .6 * i n a n t i - CT LA - 4 - t r e a t e d g r o u p s ; *P < 0 .0 1 ) . I n v i v o
t r e a t m e n t w i t h a n t i - C TL A - 4 F ( ab ) f r ag m e n t s s im i l a rl y a c -
c e l e r a t e d a n d e x a c e r b a t e d d i s ea s e w i t h a M D O o f 10 .0 " da n d a m e a n m a x i m a l s e v e r i t y o f 3 .7 * ; *P < 0 .0 5 ( d a ta n o t
s h o w n ) . Th u s , i t a pp e a r s t h a t CTLA - 4 - m e d i a t e d e v e n t s p la y
a r o l e i n t h e d o w n r e g u l a t i o n o f t h e a u t o i m m u n e e f f e c to r
f u n c t i o n s a n d /o r t h e i n v i v o e x p a n s i o n o f t h e a d o p t i v e l y
t r a n sf e r re d c e l ls . F l o w c y t o m e t r i c a n a ly s es o f PLP 13 9 - 15 1 -
p r i m e d l y m p h n o d e c e ll s a c t i v a t e d i n c u l t u r e s h o w e d u p -
r e g u l a t i o n o f su r fa c e CTL A - 4 e x p r e s s i o n o n T c e ll s (d a ta
n o t s h o w n ) . Ly m p h n o d e c e ll s t a k e n e x v i v o f r o m p r i m e d
m i c e s h o w e d l i t tl e CT LA - 4 e x p r e s s i o n ( ti m e 0 h ) w h i c h
pea ked af te r in v i t ro ac t iva t io n a t abou t 72 h of cul ture , as
d e s c r i b e d b e f o r e i n o t h e r s y s t em s ( 11) . A t t h e t i m e o f a d o p -
t ive t ransfer (96 h) , a small , but s igni f icant popul a t io n
( " ~ o f T c e ll s, s h o w e d s u r fa c e CT LA - 4 e x p r e s si o n .
Tr e a t m e n t o f th e r e c i p i e n t a n i m a l s s t a r ti n g o n t h e d a y o f
t r an s f er c o u l d r e su l t i n t h e e n h a n c e m e n t o f p r o - i n f l a m m a -
t o ry p h e n o m e n a d u e t o b l o c ka d e o f C T L A - 4 - m e d i a t e d
d o w n r e g u l a t o r y e v e n t s i n t h is p o p u l a t i o n o f ef f e c t or c e ll s,
o r o n o t h e r T c e ll s w i t h i n t h e t r a n sf e r p o p u l a t i o n w h i c h
m a y u p r e g u l a t e C TL A - 4 a f t er p r e s e n t a t i o n o f s e lf p e p t i d e si n t h e CN S.
An ti -C TL A- 4 mAb Treatment Exacerbates Relapses in
R - E A E . R - E A E i n d u c e d b y i m m u n i z a t i o n w i t h P L P 1 3 9 -
151 exhibi t s a re laps ing and rem i t t ing d i sease course . Th e
acute phase o f d i sease i s due to the a c t iv i ty of C D 4 + T ce l ls
s p e c if i c f o r t h e PLP13 9 - 15 1 e p i t o p e . H o w e v e r , w e h a v e
recent ly shown (21) tha t the pr imary d i sease re lapse i s pr in-
c ipa l ly due to the ac t iv i ty of T ce l l s spec i f i c for an e ndo ge-
n o u s , n o n - c r o s s - r e a c t i v e e p i t o p e o n PLP ( PLP17 8 - 19 1 )
w h i c h a r e a c t iv a t e d a s a r e su l t o f r e c o g n i t i o n o f m y e l i n
epi topes re leased dur ing acute d i sease , a phenomenon known
a s e p i t o p e s p r e a d in g . To l e r i z a t i o n o f PLP17 8 - 19 1 - s p e c i f i c
T c e ll s ( u n p u b l i s h e d o b s e r v a t i o n s) o r i n h i b i t i o n o f PLP17 8 -19 1 - s p ec i f ic r e sp o n s e s b y t h e b l o c k a d e o f BT- l - m e d i a t e d
cos t im ula tor y s igna l s dur in g d i sease remiss ion b lo cks d i sease
re lapses (24). T o assess the ro le of CT LA -4 in d i sease re -
l a p s e s , m i c e w e r e t r e a t e d w i t h a n t i - CTLA - 4 m A b d u r i n g
r e m i s s i o n , i . e . , a f t e r t h e y h a d r e c o v e r e d f r o m t h e a c u t e
p h a s e o f a c t i v e d i se a se . A n t i - C TL A - 4 t r e a t m e n t r e s u l te d i n
an increased d i sease sever i ty and a h igher in c iden ce o f re -
lapses (Fig . 4) . Four o ut o f the seven m ice in the c ont r o l
4
o
0
8a
o
oa
Oof
- o - , , M o , . 01] . , d ' - . . . - . 9
_,,_ aCTLA-4 (4 .y n ' -
*p<O.01/
_0-0~. 0
//I o_o.o/~176176
5 10 15 20 25
f f f ~ Da ys Post-TransferTREATMENT
Figure 3. In vivo CTLA-4 blockade exacerbates adoptive R-EAE.1<-EAE was initiated by adoptive transfer of PLP 139-151 -specific T cells.Starting on the day o f transfer, mice were treated i.p. w ith either controlor anti-CTLA-4 mAbs and graded for clinical signs of EAE. Data is plot-ted as the me an clinical score vs. d ays post-transfer. Disease incidenceswere 8/8 (control grou p) and 8/8 (anti-CTLA-4-treated group). Meandays of onset were 13.0 and 7.0* d, respectively (*P <0.01). Figures inparentheses indicate mean peak clinical severity in each group (*P <0.01).Results are representative of three separate experiments.
1.50
1.00
r
mQ.
rr 0.50
0.00~
oC r LA - 4 (2.9) /"
j - - O - - O
/,-,-@/, ~ , , ~ l , ~ l , ~ , , i , , , , i i i i i i i i i i i i J i i
Days Post-Treatment
F i gur e 4 . Anti-C TLA -4 treatment exacerbates disease relapses inR-EAE. Active R-EAE was induced by priming with PLP139-151/CFA. After recovery from the acute phase of disease ('-~ d post immu-nization), mice were treated with either control I g or intact anti-CTLA -4mAb. Data is plotted as the relapse rate (total nmnber of relapses in agroup divided by the total number of mice in that group) vs. days post-treatment initiation. 4/7 mice in the control group showed relapses,whereas 6/6 mice in the anti-CTLA-4-treated group relapsed (two micerelapsed two times each making a total of 8 relapses n 6 mice). Figures inparentheses indicate the mean peak clinical score of the relapses. The se re-suits are representative of two separate experiments. *Number of relapsesare significantlygreater than that of the hamster Ig controls, P <0.05.
786 Anti-CT LA-4 mAb Treatment Exacerbates Murine EAE
Published August 1, 1996

8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 5/6
group show ed r e l apses whereas s ix ou t o f s ix mice i n t he
a n t i - C T L A - 4 t r ea t e d g r o u p r el ap s ed , w i t h t w o m i c e s h o w -
ing two relapses each (P <0.05) .
Col lect ively, the data presented in this paper suppor t the
hypot hes i s t ha t CTL A-4 i s a ma j or nega t i ve r egu l a t o r o f
au t o immune T ce l l f unc t i on bo t h i n v i t r o and i n v ivo and
emphas i zes t he com pl i ca t ed na t u r e o f cos t imula t o ry i n t e r -
a c ti o ns . W e h a v e s h o w n t h a t th e b l o c k a d e o f C T L A - 4 i n -
teract ions wi th i t s l igands resul ted in the enhancement ofevery effector funct ion s tudied, including prol i ferat ion,
p r o - i n f l a m m a t o r y c y t o k i n e p r o d u c t i o n , e n c e p h a l i t o g e n i c -
i t y , and t he ab i l i t y t o ma in t a in ongoing au t o immune d i s -
ease. These resul t s are consis tent wi th a mode l of cost im u-
latory interact ions , w her e ini tia l act ivat ion o f T cel ls af ter
an t i gen exposure is media t ed by CD 28 / B 7 in t e rac t i ons and
leads to the prol i ferat ion and d i f ferent ia t ion o f ef fector T
cel l s . Act ivated T cel l s express CTLA-4 on thei r surface,
which can b ind t he same B7- fami ly l i gands wi t h h igher a f -
f i n it y and r e sul t i n t he dow nregu l a t i on o f t he T ce ll r e -
sponse. Th e net T cel l response wo uld then b e a result of a
b a la n c e b e t w e e n C D 2 8 a n d C T L A - 4 - m e d i a t e d e v e n ts .
Ou r r ecen t re su lt s show t hat , dur ing t he cour se o f R-E A E ,
t he re i s p r e fe ren t i al u pregu l a t i on o f B7-1 , a s com pared t o
B 7 - 2 , b o t h i n t h e C N S a n d i n t h e s p l e n i c l y m p h o i d c o m -
par tmen t (24). Su ch dyna mic changes in cost imulatory l igand
express ion dur ing a chron i c i n f l ammat ory au t o immune r e -
sponse may r e f l ec t an a t t empt a t downregu l a t i ng a des t ruc -
t ive response. Based o n the paralle l la te upregulat ion of B7-1
and CTL A-4 , i t has been sugges ted t ha t B7-1 m ay be t he
pre fe r r ed l i gand fo r CTL A-4 (26) . Our p r e l imina ry da t asuggest that there are dramat ic di f ferences in B7 l igand ex-
pres s ion depe nding on t he l ym pho id t is sue examined . I t is
t empt in g t o specu la t e t ha t ce rt a in compar t me nt s o f t he im-
m u n e s y s t e m m a y b e i n v o l v e d i n p r o v i d i n g a n e n v i r o n -
m e n t w h i c h p r o m o t e s p r o - i n f l a m m a t o r y e v e n t s , w h e r e a s
o t he r compar t ment s may be p r imar i l y r e spons ib l e fo r t he
u l t ima t e do wnreg u l a t i on o f T ce l l r e sponses . O ur cur r en t
s tudies are di rected towards address ing these i ssues by care-
fu ll y fo l l owing t he expres s ion and func t i ons o f B7 m ole -
cules and thei r T cel l l igands in animals wi th an apparent
se l f r egu l a t ing a u t o im mu ne d isease such as obse rved dur ing
re l aps ing- remi t t i ng E A E .
This wor k was supported in part by National Multiple Sclerosis Society Grant RG2 775- A3, USPH S Na -
tional Institutes of Health Research Grants NS30871, N S26543, AI35294, CA 40216, A I35225, an d a grant
from P, epligen C orporation. T.L. Walunas is the recipient o f NIH Training Grant H LO 7381-16A1.
Address correspondence to Dr. Stephen D. Miller, Departmen t of Microb iology -Imm unolo gy, Northw est-
ern University Medical School, 303 Eas t Chica go Avenue, Chica go, IL 60611 or Dr. Jeffrey A. Bluestone,
Com mittee O n Im mun ology, Box MC 1089, 5841 S. Maryland Ave., Chicago, IL 60637.
Reference s
1. Mueller, D.L., M.K. Jenkins, and R .H . Schwartz. 1989.Clonal expansion versus functional clonal inactivation: a co-
stimulator' /signalling pathway determines the outcom e o f T
cell antigen receptor occupancy. Annu. Rev. Immunol. 7: 445-
480.
2. Lensch ow, D.J., Y. Z eng , J.R.. Thistlethwaite, A. Mon tag,
W. Brady, M.G . Gibson, P.S. Linsley, and J.A. Bluestone.
1992. Lon g-te rm survival ofxen og ene ic pancreatic islet grafts
induced by CTLA41g. Science (Wash. DC ). 257:789-792.
3. Linsley, P.S., P.M. Wallace, J. John son , M.G . Gibson, J.L.
Greene, J.A. Ledbetter, C. Singh, and M.A. Tepper . 1992.
Immunosuppression in vivo by a soluble form o f the C TLA -4
T cell activation molecule. Science (Wash. DC ). 257:792-795.
4. Miller, S.D., and W.J. Karpus. 1994. The immunopathogen-
esis and regulation of T-cell m ediated demye linating diseases.lmmunol. Today. 15:356-361.
5. Brunet, J.F., F.Denizot, M.F. Luciani, M. Roux -Do sseto , M.
Suzan, M.G. Mattei, and P. Golstein. 1987. A new m emb er
of the immunoglobulin superfamily--CTL A-4. Nature (Lond.).
328:267-270.
6. Linsley, P.S., W. Bra dy, M. Urn es, L.S. Grosmaire, N.K .
Damle, an dJ.A . Ledbetter. 1991. CT LA -4 is a secon d recep-
tor for the B cell activation antigen B7. J. Exp. Med. 174:
561-569.
7. Lenschow, D.J., Y. Zeng, K.S. Hathcock, L.A. Zuckerman,
G. Freem an, J.P,. Thistlethwaite, G.S. Gray, R,.J. Hod es, andJ.A. Bluestone. 1995. Inhibition of transplant rejection fol-
lowing treatment with anti-B7-2 and anti-B7-1 antibody.
Transplantation. 60:1171-1178.
8. Perrin, P.J., D. Scott, L. Quigley, P.S. Albert, O. Feder, G.S.
Gray, 1<. Abe, C .H. June, and M.K. P, acke. 1995. Ro le o f
B7 :CD 28/ CT LA -4 in the induction of chronic relapsing ex-
perimental allergic encephalomyelitis. J, Immunol. 154:1481-
1490.
9. Dariavach, P., M.G. Mattei, P. Golstein, and M.P. Lefranc.
1988. Hum an Ig superfamily CTL A-4 gene: chromoso mal
localization and identity of protein sequ ence betwe en m urine
and human CTLA-4 cytoplasmic domains. Eur. J. Immunol.
18:1901-1905.
10. Linsley, P.S., J.L. Greene, P. Tan, J. Bradshaw, J.A. Ledbet-ter, C. Anasetti, and N.K. DaMe. 1992. Coexpression and
functional cooperation of CTLA- 4 and CD 28 on activated T
lymphocytes.J. Exp. Med. 176:1595-1604.
11. Walunas, T.L., D.J. Le nsch ow, C. Y. Bakker, P.S. Linsley,
G.J. Freeman, J.M. Green, C.B. Tho mpso n, and J.A. Blue-
stone. 1994. CTLA-4 can function as a negative regulator of
T cell activation. Immunity. 1:405-413.
12. Damle, N.K., K. Klussman, G. Leytze, S. Myrdal, A. Aruffo,
J.A. Ledbetter, and P.S. Linsley. 1994. Costimulatio n of T
lymp hocy tes with integrin ligands intercellular adhesion mo l-
78 7 Karandikar et al. Brief Definitive Re po rt
Published August 1, 1996

8/4/2019 CTLA-4- A Negative Regulator of Autoimmune Disease
http://slidepdf.com/reader/full/ctla-4-a-negative-regulator-of-autoimmune-disease 6/6
ecule-1 or vascular cell adhesion molecule-1 induces func-
t ional expression ofCT LA -4, a second receptor for B7.J. Im-
munol. 152:2686-2697.
13. Walunas, T .L., C .Y. Bakker, and J.A. B luestone. 1996.
CTLA-4 l igat ion blocks CD28-dependent T cel l act ivat ion.
J. Exp. Med. 183:2541-2550.
14. Krummel, M.F., andJ.P. All ison. 1995. CD28 and CTLA-4
have op posing effects on the response of T cells to stimu la-
t ion .J . Exp. Med. 182:459-465.
15. Keame y, E.R., T.L. Walunas, R.W . Karr , P.A. Morton,
D.Y. Loh, J .A. Bluestone, and M.K. Jenkins. 1995. An tigen-
dependent clonal expansion of a t race populat ion o f ant igen-
specific CD 4 + T cells in vivo is dependent o n CD2 8 costimu-
lat ion and inhibited by CTL A-4.J . Immunol. 155:1032-1036.
16. Leach, D.R ., M.F. Krum mel, a nd J.P. Allison. 1996. En -
hancement of ant itumor immuni ty by C TLA -4 b lockade .
Science (Wash. DC). 271 : 1734-1736 .
17. Tivol , E .A., F. Borriel lo, A.N. Schweitzer, W.P. Lynch, J .A.
Bluestone, and A.H. Sharpe. 1995. CT LA -4 deficient mice
exhibi t massive lymphoproliferat ion and mult i-organ lym-
phatic infiltration: a critical negative immunoregulatory role
o f CTLA - 4 . Immunity. 3:541-547.
18. Waterhouse, P.W.,J.M. Penninger, E. Timms, A. Wakeham,
A. Shahinian, K.P. Lee, C.B. Thompson, H. Griesser, andT.W . Mak. 1995. Lymphoproliferat ive disorders with early
lethali ty in mice deficient in CTL A-4. Science (Wash. DC ).
270:985-988.
19. Weker le, H. 1991. Im muno pathog enesis o f multiple sclerosis.
Acta Neurologica. 13:197-204.
20. McRae, B.L., M.K. Kennedy, L.J. Tan, M.C. Dal Canto,
and S.D. Miller. 1992. In ductio n of active and adop tive
chronic-relapsing experimental autoimmun e encephalo my-
elitis (EAE) using an encephalitogenic epitope of proteolipid
prote in .J . Neuroimmunol. 38:229-240.
21. McRae, B.L., C.L. Vanderlugt , M.C. Dal Canto, and S.D.
Miller. 1995. Functional evidence for epitope spreading in
the relapsing pathology of EAE in the SjL/J mouse. J. Exp.
Med. 182:75-85.
22. Lehmann, P.V., E.E. Sercarz, T. Forsthuber, C.M. Dayan,
and G. Gam mon. 1993. Determinan t spreading and the dy-
namics of the autoimm une T-cel l repertoire. Immunol. Today.
14:203-208.
23. Kuchroo, V.K., M.P. Das, J .A. Brown, A.M. Ranger, S.S.
Zamvil , R.A. Sobel , H.L. Weiner, N. Nabavi , and L.H.
Glimch er. 1995. B7-1 and B7-2 co stimulatory molecules dif-
ferential ly act ivate the T hl /T h2 developmental pathways:
application to autoimm une disease therapy. Cell. 80:707-718.
24. Miller, S.D., C.L. Vande rlugt, D.J. Lenschow , J.G. Pope,
N.J. Karandikar , M.C. Dal Canto, andJ.A. Bluestone. 1995.
Blockade of CD2 8/B7-1 interact ion prevents epi tope spread-
ing and clinical relapses ofm urin e EAE. Immunity. 3:739-745.
25. Lenschow, D.J., S.C. Ho, H. Sattar, L. Rhee, G. Gray, N.
Nabav i, K.C. Her old, and J.A. Bluestone. 1995. Differentialeffects of ant i-B7-1 and ant i-B7-2 mA b treatment on the de-
velopmen t of diabetes in the NO D mouse. J. Exp. Med. 181 :
1145-1155.
26. Lenschow, D.J., andJ.A. Bluestone. 1993. T cell co-stimula-
tion and in vivo tolerance. Curr. Opin. Immunol. 5:747-752.
788 Anti-CT LA-4 mAb Treatment Exacerbates Murine EAE
Published August 1, 1996





























