Embed Size (px)
Citation preview

UNIVERSITA’ “G. d’ANNUNZIO” -CHIETI-PESCARA
DIPARTIMENTO DI SCIENZE BIOMEDICHE Sezione di Terapia Medica
CORSO DI PERFEZIONAMENTO IN
MEDICINA BIOINTEGRATA
www.centrostellamaris.it

Psico-Neuro-Endocrino- Immunologia-Genetica (PNEIG) in Medicina Biointegrata

Programma AA.2010-2011 • Definizione di PNEI. Il Sistema nervoso: le cellule del SN e loro
connessioni; il tronco encefalo, diencefalo, SNV. Sistema endocrino: gli ormoni, loro funzioni e comunicazioni. Controllo cerebrale sul Sistema endocrino. Assi neuroendocrini: ipotalamo-ipofisi, ipotalamo-ipofisi-surreni, ipotalamo-ipofisi-tiroide, ipotalamo-ipofisi-gonadi. Sistema immunitario: cenni di immunologia (teoria della selezione clonale, immunità naturale ed acquisita, sistema HLA, sistema immunitario come organo di senso, produzione di ormoni da parte del sistema immunitario che fungono da mediatori tra cervello e sistema endocrino). Cervello e psiche. La grande connessione: Sistema nervoso ed immunità; sistema dello stress; sistema endocrino e cervello. Endocrinosenescenza e P.N.E.I.. DCA prototipo di PNEIG.

Assi neuroendocrini

(ng) (µg)
Cascate ormonali • Stimoli
iniziali neuro-sensoriali
• Anelli di retroazione negativa (feedback)
Natura e unità di misura

Ipotalamo-ipofisi
strutture importanti del Sistema Nervoso, responsabili dei meccanismi di elaborazione, verifica e trasmissione di emozioni

• L’ipotalamo è organizzato in circa una dozzina di "nuclei" di cellule che si coordinano fra loro, la sua costituzione anatomica, è sostanzialmente simile in tutti i vertebrati; questo particolare neuroanatomico sembrerebbe non essersi evoluto... è come se fosse "nato" perfetto!
• La differente efficienza funzionale, nelle diverse specie viventi, sarebbe data dagli apprendimenti acquisiti, più che dall’assetto cellulare genetico. La base predisponente, è uguale per tutti; cambiano solo le informazioni depositate in memoria (conoscenze fatte).

• ha un ruolo di coordinamento e sincronizzazione nella gestione e nella trasmissione delle emozioni
• Gestisce l’apparato endocrino, tramite la collaborazione dell’Ipofisi
• Regola, coordina e sincronizza le attività cosiddette "autonome", con il coinvolgimento del Sistema Nervoso Vegetativo (SNV-SNA) (Temperatura corporea, fame e sete, ritmo sonno - veglia, funzioni sessuali, secrezione acida dello stomaco, etc.).
• Per svolgere tutte queste funzioni, l’Ipotalamo prende informazioni sia dall’interno che dall’esterno dell’organismo:

• dall’interno: dal Sistema limbico (“in dialogo" con Sistema Neuroendocrino e Neurovegetativo), da Amigdala e Ippocampo (attraverso il fornice) e dalla Corteccia tramite il Talamo (relé di informazioni). Verso il basso, è connesso con midollo allungato e midollo spinale: dialoga col SNA.
• dall’esterno: tramite il Nucleo soprachiasmatico, l’Ipotalamo riceve il segnale della luce e del buio che gli consente di regolare gli orologi biologici interni.

• La sua Eminenza mediana (fibre nervose e vasi sanguigni), attraversa la barriera ematoencefalica e va a costituire, all’esterno dell’encefalo nella fossa ipofisaria del corpo dell’osso sfenoide ("sella turcica“), la neuroipofisi. Lo stretta unione fra Ipotalamo e Neuroipofisi realizza una connessione forte fra Sistema Nervoso, Sistema Endocrino e Sistema Immunitario, garantendo la regolazione dei processi vitali fondamentali.

• Alla Neuroipofisi si collega, anteriormente, l’Adenoipofisi composta da tessuto ghiandolare e "governata" dall’Ipotalamo tramite una serie di ormoni ("releasing factors" - Fattori di rilascio) che giungono attraverso il circolo sanguigno ipotalamo - ipofisario e stimolano l’Ipofisi a secernere ormoni che attiveranno ghiandole endocrine (tiroide, surreni, pancreas, ovaie, etc.) e organi non strettamente endocrini (ad esempio, i reni) a svolgere le proprie mansioni.

G. Edelman in "Sulla materia della mente", descrive due grandi vie di connessione:
a) la via tronco-encefalica-limbica-autonoma legata a comportamenti affettivi, sessuali,
consumatori, di difesa e alla regolazione di alcuni fenomeni vitali di tipo vegetativo (frequenza cardiaca, respiratoria, digestione, sonno ecc) e "dialoga" con il sistema endocrino attraverso la produzione di ormoni che impiegano minuti o ore per raggiungere il bersaglio.

b) la via talamo - corticale che invece, rappresenta un sistema sviluppato per ricevere segnali dai recettori sensoriali e inviarli ai muscoli volontari. Collegamenti più veloci rispetto alla via precedente perché mediati da neurotrasmettitori. I due sistemi hanno il loro collegamento nel Talamo, stazione fondamentale per la circolazione delle informazioni che vengono dal basso e dall’alto, dalla periferia e dalla corteccia.

• Secondo J. W. Papez, gli impulsi nervosi partirebbero dalla corteccia cerebrale e, passando attraverso il giro del Cingolo e dell’Ippocampo, raggiungerebbero l’Ipotalamo. Dall’Ipotalamo i messaggi, passando per il Talamo, potrebbero ritornare alla Corteccia.
• Si realizzerebbe così uno scambio di informazioni che verrebbero via via sempre più verificate e riconosciute prima di essere trasmesse.

Ormoni ipofisari e ipotalamici
Ipotalamo
Ipofisi posteriore (neuroipofisi)
Ipofisi anteriore (adenoipofisi)
Ormoni di rilascio Ormoni di rilascio
Altre ghiandole endocrine Metabolismo
5 assi endocrini
Ormoni di rilascio
Bilancio idrico Produzione latte
Vasopressina (ADH) Ossitocina (OXT)
Ormoni di rilascio
Impulso nervoso
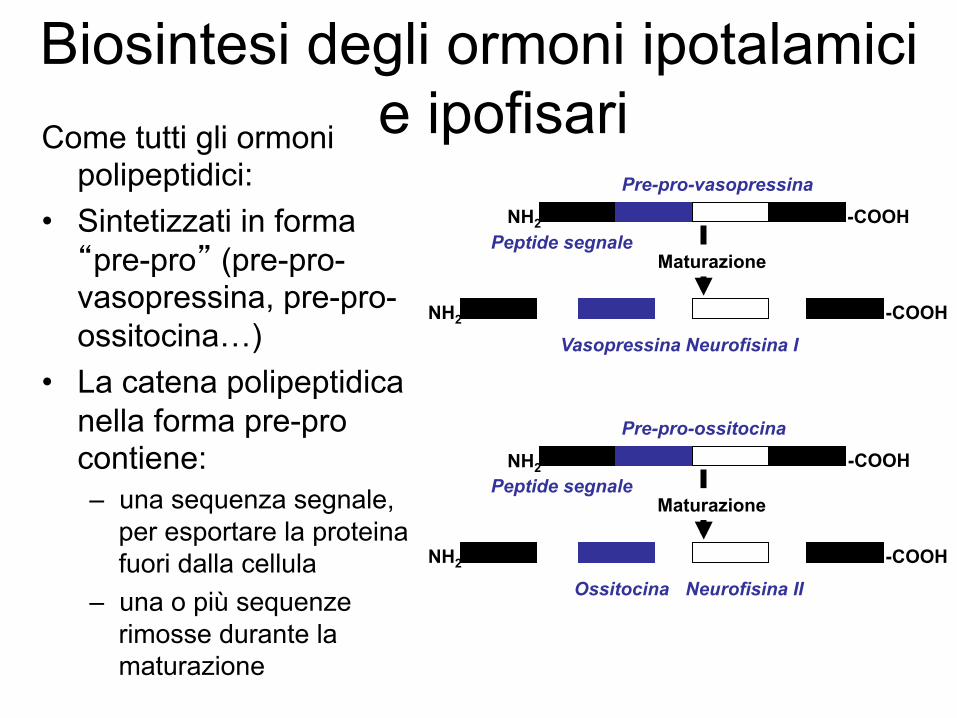
Biosintesi degli ormoni ipotalamici e ipofisari Come tutti gli ormoni
polipeptidici: • Sintetizzati in forma “pre-pro” (pre-pro-vasopressina, pre-pro-ossitocina…)
• La catena polipeptidica nella forma pre-pro contiene: – una sequenza segnale,
per esportare la proteina fuori dalla cellula
– una o più sequenze rimosse durante la maturazione
NH2- -COOH
NH2- -COOH
Pre-pro-vasopressina
Vasopressina
Peptide segnale
Neurofisina I
Maturazione
NH2- -COOH
NH2- -COOH
Pre-pro-ossitocina
Ossitocina
Peptide segnale
Neurofisina II
Maturazione

l’asse ipotalamo-ipofisi-surrene (HPAA) • Il CRH è un neurotrasmettitore (recettori
nell’amigdala, locus ceruleus, stria terminale), prodotto dalle cellule neuroendocrine del NPV ipotalamico, attraverso il circolo portale arriva alle cellule corticotrope dell’eminenza mediana dell’adenoipofisi. Queste rilasciano nel circolo sistemico corticotropina (ACTH), sintetizzata dal precursore proteico Pro-opio-melanocortina (POMC), insieme ad altre sostanze biologiche . L’ACTH presenta un ritmo circadiano di secrezione (con massima produzione al mattino 5AM) e di conseguente stimolazione dei glucocorticoidi corticosurrenalici (cortisolo), aldosterone e androgeni.
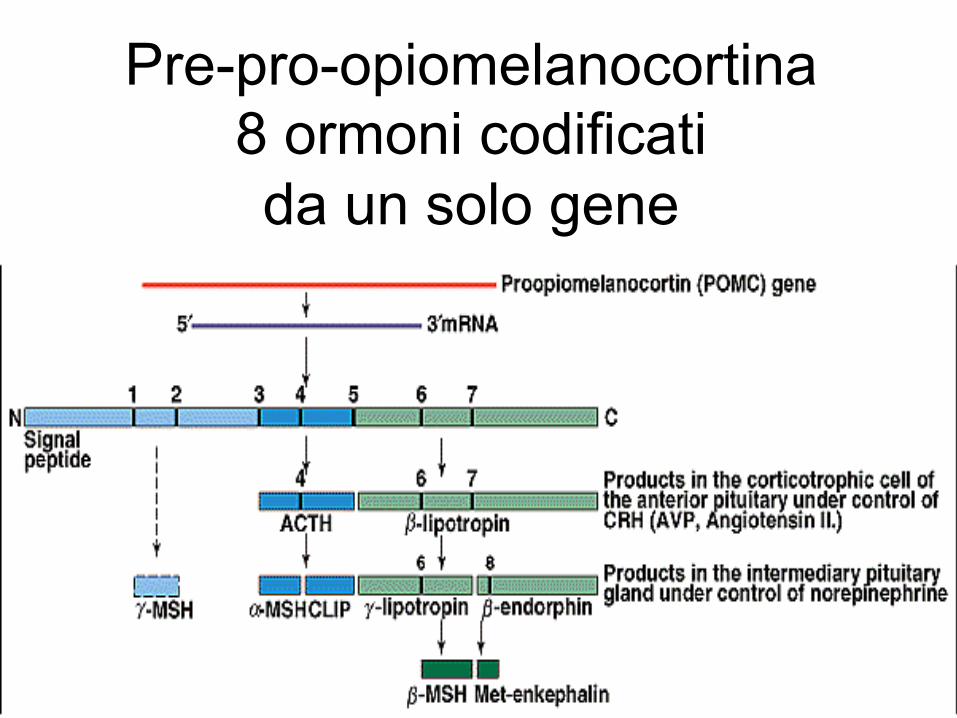
Pre-pro-opiomelanocortina 8 ormoni codificati da un solo gene

Asse ipotalamo-ipofisi-surrene • Ipotalamo → Ormone di rilascio della
corticotropina (CRH) e vasopressina (ADH) – cAMP
• Ipofisi anteriore → Ormone adrenocorticotrofico (ACTH) – Derivato dalla pro-opiomelanocortina – Implicato in bioritmi (picco max 5 AM) – T1/2=10 min – Agisce tramite cAMP come 2° messaggero
• Surrenali – Zona glomerulare: mineralocorticoidi – Zona fascicolata: glucocorticoidi – Zona reticolare: ormoni sessuali – Zona midollare: adrenalina e noradrenalina

• La corteccia surrenalica è distinguibile in • Zona glomerulare (cellule parenchimali
sintetizzano e secernono ormoni mineralocorticoidi per il mantenimento dell'equilibrio idrico)
• Zona fascicolata : sintetizza e secerne i glucocorticoidi e controlla il metabolismo dei carboidrati (↓ l'utilizzo del glucosio, ↑gluconeogenesi), grassi e proteine
• Zona reticolata (produce gli ormoni sessuali come androgeni, estrogeni e progesterone).
• Zona midollare completamente ricoperta dalla corticale, sintetizza vari ormoni tra i quali le catecolamine. Comprende due popolazioni di cellule parenchimali: cellule cromaffini e cellule gangliari simpatiche.

Ormoni steroidei della corteccia surrenalica
• Tutti gli ormoni steroidei derivano dal colesterolo
• Catena laterale al C17: – Accorciata a 2 C → progestinici,
corticosteroidi – Rimossa completamente →
androgeni, estrogeni • Aggiunta di gruppi -OH o chetonici • Mineralocorticoidi: aldeide al C18 • Estrogeni: anello A aromatico

Colesterolo
Mineralocorticoidi, 21C Ritenzione di Na+ Escrezione di K+
Pregnenolone
Progesterone
11-deossi corticosterone
Corticosterone
Aldosterone
Estrogeni, 18C Estradiolo
Glucocorticoidi, 21C Stimolatori della gluconeogenesi
Adattamento dell’organismo allo stress Antiinfiammatori
17-OH pregnenolone
17-OH progesterone
11-deossi cortisolo
Cortisolo
Androgeni,19C Differenziazione sessuale
Oncogenesi
deidroepiandrosterone
Androstenedione
Testosterone
Biosintesi degli ormoni steroidei

Il CRF modula la risposta di adattamento allo stress, attraverso la secrezione di ormoni ipofisari e connessioni nervose dirette con neurotrasmettitori ( noradrenalina e adrenalina), glucosio e glucagone. Anche la serotonina partecipa alla risposta adattativa, ma solo per deplezione dopo stimolo intenso. Il CRF correlato anche agli oppiodi endogeni: dinorfina, prodinorfina ed encefalina (peptidi connessi con la riduzione della percezione del dolore, della fatica, della sensazione di benessere e di energia ecc.).

Glucocorticoidi e cortisolo • Sintesi e secrezione nella fascicolata surrenale sotto controllo di
ACTH – Soggette a bioritmi (mattina > sera) – Proteina di trasporto: Cortisol-binding globulin (CBG) – t1/2=100 min
• EFFETTI: – Aumento di gluconeogenesi, glicogeno epatico, lipolisi, biosintesi
proteica (come il glucagone, ma più lento) ma infine prevale il catabolismo. Può causare diabete
– Soppressione di risposta immunitaria ed infiammatoria (cortisone) – Incremento dell’ultrafiltrazione renale
• PATOLOGIE PRINCIPALI – Morbo di Addison: insufficienza surrenalica → ipoglicemia, intolleranza
a stress, debolezza, ipotensione – Sindrome di Cushing: eccesso di glucorticoidi (somministrazione
farmacologica o adenoma delle surrenali, o aumento di ACTH) → iperglicemia, catabolismo delle proteine, perdita di massa corporea, “faccia a luna piena” (accumulo di tessuto adiposo sul viso)

Catecolamine (ormoni) della midollare surrenale che mediano l’adattamento allo stress • Tyr: precursore (anche Phe)
– Tyr idrossilasi: Reazione limitante • DOPA, diidrossifenilalanina (carente in
Parkinson) – Passa la barriera emato-encefalica e funge
da farmaco • Dopamina: non passa la barriera emato-
encefalica – In caso di deficit: DOPA – In caso di eccesso: metil-DOPA
• Adrenalina e noradrenalina – Accumulate nei granuli cromaffini – Rilasciate con stimolazione β-adrenergica – Metabolizzate rapidamente nel plasma
• catecolo metiltransferasi (COMT) e monoamina ossidasi (MAO)
• Inibitori di COMT e MAO: eccitatori nervosi

• La noradrenalina è sintetizzata da una serie di passaggi enzimatici nella midollare surrenale a partire dall'aa. tirosina. La prima reazione è l'ossidazione in diidrossifenilalanina (L-DOPA), seguita dalla decarbossilazione nel neurotrasmettitore dopamina, e infine dalla β-ossidazione in noradrenalina. Questa può essere ulteriormente metilata dalla feniletanolamina N-metiltransferasi (PNMT) in adrenalina (epinefrina in USA).
Tirosina L-DOPA DOPAMINA
Noradrenalina Fenilalanina

• Nell'alimentazione umana, essenziali i seguenti aminoacidi che l’organismo NON sintetizza:
• H Istidina • I Isoleucina • L Leucina • K Lisina • M Metionina + C Cisteina • F Fenilalanina + Y Tirosina (aromatici)* • T Treonina • W Triptofano** • V Valina • Arginina, cisteina, istidina e tirosina sono essenziali durante
l'infanzia e lo sviluppo. * La fenilalanina abbondante in aringa sott'aceto, avocado, banane, fagioli Lima,
formaggio, formaggio casereccio, latte non grasso in polvere, mandorle, noccioline, semi di sesamo, semi di zucca gialla. Fabbisogno OMS di F+Y 25mg/kg
** Il triptofano abbondante nel cioccolato, nell'avena, nelle banane, nei datteri, nelle arachidi, nel latte e nei latticini. Fabbisogno raccomandato dall'OMS 4mg/kg.

Mineralocorticoidi e aldosterone • Sotto controllo del sistema renina-angiotensina
– Stimolato da ipotensione, dieta priva di sali – Inibito da ipertensione, dieta ricca di sali
• EFFETTI: – ↑riassorbimento di Na+ nel rene (riassorbimento di
H2O) – ↑secrezione di K+ e H+ (urine acide)
• PATOLOGIE: – Sindrome di Conn (aldosteronismo primario) → ↑riassorbimento di Na+, ↑volume extracellulare, ipertensione, ↓K+
– Insufficienza surrenalica →↓gluconeogenesi, ↑sensibilità a insulina, ipoglicemia, ipotensione

Fegato
Aumenta il glucosio ematico
Diminuisce il glucosio ematico
↑ Glicolisi ↓ Gluconeogenesi
Insulina Glucagone
↓ Glicolisi ↑ Gluconeogenesi
Pancreas
Cellule β Cellule α
↓Glucosio ↑ Glicogeno
↑ Glucosio ↓ Glicogeno
Ormoni pancreatici: insulina e glucagone

Insulina • Cellule β delle isole di Langerhans
– Agonisti α-adrenergici inibiscono – Agonisti β-adrenergici e i derivati della
sulfonilurea (tolbutammide) stimolano • Fortemente conservata: insulina bovina e
suina usabili nell’uomo • Pro-insulina dà reazioni crociate con
insulina • Livello di peptide C discrimina insulina eso-
ed endogena
Reticolo endoplasmico
Pre-pro-insulina
Apparato di Golgi
Pro-insulina
Insulina + C
Circolazione
Pro-insulina
Insulina + C 95%
5%

Recettore per insulina • Legame con insulina attiva recettore Tyr
chinasi (autofosforilazione) • Attivazione di cascata kinasica • Risposte metaboliche veloci (tendenti a
diminuire la glicemia) – Aumento di:
• Trasporto di glucosio (GLUT-4) • Sintesi di glicogeno • Glicolisi
– Inibizione di: • Gluconeogenesi • Glicogenolisi • Lipolisi
• Risposte biosintetiche lente o a lungo termine
– Aumento di: • Sintesi proteica • Replicazione cellulare
• Regolazione dell’attività del recettore e del livello di ormone circolante tramite endocitosi (desensibilizzazione)

Glucagone • Cellule α delle isole di Langerhans
– Secreto come pro-glucagone – Agisce tramite recettori e cAMP
• Inattivato nel fegato – Breve t1/2 in circolo
• Stimolatore di: – Gluconeogenesi – Glicogenolisi – Chetogenesi – Assunzione di aminoacidi
• Inibitore di: – Glicolisi – Glicogenogenesi

Insulina vs glucagone

Asse dell’ormone della crescita (GH)
• Ipotalamo → Ormone di rilascio di GH (GHRH) – cAMP, Ca++-CaM – Inibito da somatostatina
• Ipofisi anteriore → GH – Proteina di trasporto specifica – Rilascio in picchi ogni 3-4 h (specie durante il sonno)
• Fegato – Stimolazione di lipolisi, induce resistenza all’insulina – Produzione di insulin-like growth factor (IGF-I)
• Proteina di trasporto specifica • Marker dell’attività di GH
• Tessuti – Stimolazione dell’utilizzo di acidi grassi con risparmio di glucosio – Stimolazione dell’utilizzo di aminoacidi con sintesi proteica
• Patologie: – ↓GH (ipopituitarismo): difetti di crescita – ↑GH (tumore dell’ipofisi): gigantismo, acromegalia

Somatostatina • Cellule γ delle isole di Langerhans • Secreta come precursore • Targets:
– Pancreas: Inibisce la secrezione di insulina e glucagone
– Stomaco e intestino: Diminuisce la secrezione di gastrina
• Prolunga il tempo di svuotamento gastrico • Rallenta l’assorbimento intestinale degli zuccheri • Riduce il passaggio dei nutrienti in circolo
– Cervello e ipofisi: inibisce secrezione di GH

Asse ipotalamo-ipofisi-tiroide • Ipotalamo → Ormone di rilascio della tirotropina (TRH)
– Fosfolipasi C e Ca++
• Ipofisi anteriore → TSH • glicoproteina cAMP, PKA • Iodio
– >150 µg/giorno di I2 (dieta) – I2 → I- nell’intestino
• Tiroide → triiodiotironina (T3), triiodiotironina inversa (rT3), tiroxina (T4) – rT3 inattiva, T4 preormone, T3 metabolicamente più attiva – Attività sotto controllo di deiodinasi (tiroide e tessuti target):
T4 → T3
• Targets: Gran numero (tutti?) di tessuti

Gli ormoni tiroidei tiroxina (T4) e triiodotironina (T3) regolano il metabolismo di tutti i tessuti dell'organismo. Essi aumentano il metabolismo basale, incrementando il consumo di ossigeno e la produzione di calore, attraverso l’aumento degli enzimi cellulari, soprattutto quelli dei mitocondri coinvolti nella respirazione cellulare. Una diminuzione del T3 e T4 nel sangue ed un abbassamento della temperatura corporea, stimolano l'ipotalamo a produrre TRH , il quale agisce nell'ipofisi anteriore e provoca il rilascio di prolattina, GH e TSH (ormone stimolante la tiroide), che ripristina la concentrazione di T3 e T4 nel sangue aumentando la temperatura corporea.

• L’eccesso di T3 ha effetto adrenergico inotropo e cronotropo positivo sul cuore. La deficienza di T3 nelle fasi iniziali della vita causa ritardo di sviluppo del SNC seguito da un deficit cognitivo e motorio (cretinismo): il flusso sanguigno nel cervello e il metabolismo cerebrale di glucosio sono più bassi. L’ormone tiroideo aumenta anche la velocità e l'ampiezza dei riflessi nervosi periferici, la vigilanza e reattività a diversi stimoli, la memoria e la capacità di apprendimento. La T3 partecipa al rimodellamento osseo negli adulti e il suo eccesso può causare osteoporosi

• La T3 inibisce la sintesi e aumenta la degradazione dei glicosaminoglicani , della matrice dermale, aumenta la produzione di eritropoietina e l’emopoiesi. L’ormone aumenta anche i livelli di 2-3 difosfoglicerato negli eritrociti, promuovendo la dissociazione dell’ossigeno dall’emoglobina, elevando quindi la disponibilità di ossigeno libero ai tessuti. Promuove la maturazione dei pre-adipociti ad adipociti. La T3 abbassa i livelli plasmatici di colesterolo (LDL), aumentando la sintesi di recettori LDL sulla membrana cellulare. L’ormone aumenta anche la lipolisi degli acidi grassi nel tessuto adiposo.

• L’azione termogenica obbligatoria della T3 avviene grazie all’attivazione dell’espressione delle proteine UCP che promuovono il ritorno di protoni attraverso lo spazio intermembranoso della membrana mitocondriale, senza attivare l’ ATP sintetasi e, quindi, senza produrre ATP. Questo flusso di protoni dissipa la loro energia potenziale chimica sotto la forma di calore. La T3 aumenta anche la lipolisi, fondamentale per questo processo di generazione di calore che è denominato disaccoppiamento.

• Nella termogenesi obbligatoria può esserci anche l’idrolisi dell’ATP da parte delle ATPasi, promossa dalla T3. Negli ambienti più freddi, è necessaria l’attivazione di un altro meccanismo, la termogenesi facoltativa; questa è attivata da parte del sistema nervoso simpatico grazie a stimoli esterni, ma subisce un'influenza importante da parte della T3. Una carenza di T3 rende incapaci di sostenere la risposta noradrenergica per la generazione di calore supplementare (la T3 e le catecolamine hanno effetti sinergici).


Asse ipotalamo-ipofisi-gonadi • Ipotalamo → Ormone di rilascio della
gonadotropina (GnRH) – Ca++, PIP2 e PKC
• Ipofisi anteriore → Ormone follicolo stimolante (FSH), ormone luteinizzante (LH)
• FSH e LH (glicoproteine) hanno funzioni relativamente simili in entrambi i sessi

• L' GnRH, prodotto nel nucleo arcuato e nell’area preottica ipotalamica induce il rilascio di FSH, LH dall‘ipofisi anteriore. Dopo un picco in età prenatale ed uno minore subito dopo la nascita, la secrezione di GnRH rimane bassa fino all'avvento della pubertà (tra gli 8 e i 13 anni nelle femmine; tra i 9 e i 14 anni nei maschi). Due le ipotesi più accreditate sulla ripresa della secrezione di GnRH puberale : a)La precoce degenerazione dall’epifisi con progressiva diminuzione della secrezione di melatonina, inibitoria sui nuclei ipotalamici che producono e secernono GnRH.
b)Un troppo basso peso, tramite le leptine ritarderebbe la pubertà.

• Quando il GnRH si lega a specifici recettori posti sulla membrana di cellule gonadotrope adenoipofisarie, viene attivata una cascata di segnali intracellulari che portano alla secrezione dell’ormone luteinizzante, precedentemente accumulato nella cellula all’interno di granuli secretori e di cui il GnRH è in grado di promuovere l’ulteriore trascrizione e sintesi.

FSH e LH nell’ovaio • Secrezioni ritmiche col ciclo mestruale:
– Secrezione di estradiolo → maturazione del follicolo
– In concerto con progesterone: ciclo mestruale
– In concerto con insulin-like growth factor I (IGF-I): crescita e maturazione dei caratteri secondari femminili
– Turnover dell’osso (osteoporosi senile)

FSH e LH nei testicoli • Secrezione di testosterone (via
proteina G) – Proteine di trasporto
• Albumina aspecifica • Sex-hormone binding globulin
(SHBG) specifica – Targets:
• Organo del Sertoli (spermatogenesi) • Tessuto muscolare: sintesi proteica → anabolismo

• Gli ormoni LH,FSH sono glicoproteine. Nel maschio, l’LH stimola l'attività endocrina delle cellule interstiziali del testicolo con produzione di testosterone. Inoltre, a causa del suo ruolo nella produzione di testosterone, ormone necessario per la maturazione delle cellule germinali, l’ormone luteinizzante stimola anche, in maniera indiretta, la spermatogenesi.
• Nella femmina, invece, l’LH stimola, insieme alla prolattina, l‘ovulazione e la conversione del follicolo ovarico in corpo luteo, L’ormone steroideo prodotto, quindi, porta alla sintesi dei precursori dell’estradiolo. L’LH stimola anche la sintesi di progesterone da parte delle cellule della granulosa .

Asse della prolattina TRH → Ipofisi anteriore → prolattina → ghiandola mammaria → secrezione di
lattoalbumina – Meccanismo simile a quello del GH (citochina) – Neuroni → dopamina (controllo negativo) – cAMP, Ca++-CaM – Inibito da somatostatina
• Patologie: – ↑prolattina (tumore dell’ipofisi o deficit di dopamina):
irregolarità mestruale nelle donne, iperplasia prostatica negli uomini

• La PRL ormone di 198 aa, è secreto dall‘adenoipofisi in modo pulsatile e raggiunge il picco durante il sonno rem tra le 3 e le 5 del mattino. Il suo valore medio è di 20 µg/l. La sua regolazione ipotalamica avviene principalmente tramite un fattore inibitorio il PIF=PIH risultato successivamente essere la DA; stimolato meno dal TRH e da peptidi minori. Essa viene anche prodotta da cellule specializzate dell‘utero, della placenta, delle mammelle ed anche del sistema immunitario, con un ruolo paracrino e autocrino come citochina. La PRL soffre un tono inibitorio costante da parte della DA ipotalamica.

• Durante la gravidanza, gli estrogeni aumentano la sua produzione, promuovendo l‘iperplasia delle cellule lattotrofe dell'ipofisi e l'aumento dell‘espressione del suo gene. La PRL aumenta col riflesso neuroendocrino della suzione, attribuito all'azione della serotonina(5-HT), la cui liberazione è aumentata dalla stimolazione sensoriale fatta dal bambino nella mammella, attraverso il NPV ipotalamico. La serotonina, pur essendo un importante fattore di liberazione di prolattina, dipende per questa funzione da un'altra sostanza: il peptide intestinale vasoattivo (VIP)

• Questa sostanza, avrebbe un'azione paracrina nell'ipofisi, segnalando ai lattotrofi di produrre prolattina. Questo ruolo fondamentale del VIP è stato dimostrato tramite l'iniezione di serotonina e di siero anti-VIP nel cervello di conigli, il cui picco di prolattina nel plasma indotto dalla serotonina era molto più basso di quello presentato dai conigli che non hanno ricevuto il siero anti-VIP.

• La DA è il principale inibitore della PRL ma si lega ai recettori D2 dei lattotrofi solo ad alte concentrazioni ematiche. Ciò diminuisce la produzione di cAMP, apre canali del K+ e diminuisce il flusso di Ca+ verso il citoplasma cellulare. Come conseguenza, diminuisce la secrezione e trascrizione genica della PRL.
• Nel circolo portale ipotalamo-ipofisario dell’eminenza mediana, i livelli di DA sono più alti di quelli sistemici, visto che gli assoni del nucleo arcuato ipotalamico che la producono, vi scaricano il loro contenuto direttamente.
• Questo consente all'ipotalamo di generare un tono inibitorio sulla produzione di PRL.

• La PRL presenta un meccanismo feedback negativo con la DA, alti livelli di PRL ne favoriscono la sintesi nei neuroni dopaminergici dell'ipotalamo (n.Arcuato). I lattotrofi presentano recettori per il TRH. il GAP (peptide associato al GnRH), inibisce la secrezione di PRL in antagonismo con il peptide liberatore di prolattina (PrRP), attivato da livelli alti di estrogeni. Altro ormone capace di aumentare in modo blando la secrezione di prolattina è l‘ossitocina. Gli oppioidi esogeni, come pure la beta-endorfina, la neurotensina e l'angiotensina II aumentano la secrezione di prolattina.

• La PRL promuove la captazione di alcuni aa e la trascrizione di geni che producono le proteine del latte: la caseina, la lattoalbumina e la beta-lattoalbumina. La PRL aumenta anche la produzione di enzimi importanti per la sintesi del lattosio (il principale carboidrato del latte). Dopo il parto, i bassi livelli di progesterone, rendono la PRL capace di indurre la lattogenesi,come fa pure in questo periodo l’aumento dei livelli di cortisolo, grazie alla minor produzione di globuline a cui esso si lega. La PRL ha un effetto anche su altri organi dell‘apparato genitale femminile trasmettendo un segnale inducente refrattarietà alla fecondazione.

Aumento della prolattina e caduta del progesterone sembrano essere (nel ratto) le
cause immediate della comparsa del comportamento materno












• Psychoneuroendocrinology (2009) 34, 382—388
• BDNF Val66Met polymorphism is associated with HPA axis reactivity to psychological stress characterized by genotype and gender interactions
• Idan Shalev a, Elad Lerer b, Salomon Israel c, Florina Uzefovsky c,
• Inga Gritsenko d, David Mankuta e, Richard P. Ebstein c,d,*, Marsha Kaitz c
• a Neurobiology, Hebrew University, Jerusalem, Israel • b Human Genetics, Hebrew University, Jerusalem, Israel • c Psychology Department, Hebrew University, Jerusalem, Israel • d S. Herzog Memorial Hospital, Jerusalem, Israel • e Hadassah Medical Organization, Department of Labor and
Delivery, Jerusalem, Israel

• Molecular Psychiatry (2008), 1–8 • Evidence of biologic epistasis between BDNF
and SLC6A4 and implications for depression • L Pezawas1,2,6, A Meyer-Lindenberg1,3,6, AL Goldman1, BA
Verchinski1, G Chen4, BS Kolachana1, • MF Egan1, VS Mattay1, AR Hariri5 and DR Weinberger1 • 1Genes, Cognition and Psychosis Program, National Institute of
Mental Health, National Institutes of Health, Bethesda, MD, • USA; 2Division of Biological Psychiatry, Medical University of
Vienna, Vienna, Austria; 3Department of Psychiatry and • Psychotherapy, Central Institute of Mental Health, Mannheim,
Germany; 4Scientific and Statistical Computing Core, National • Institute of Mental Health, National Institutes of Health,
Bethesda, MD, USA and 5Department of Psychiatry, Western • Psychiatric Institute and Clinic, University of Pittsburgh School
of Medicine, Pittsburgh, PA, USA





























