Embed Size (px)
Citation preview

azhvmwcvogbtespptsbpw(mt(cappwcspom
dTcm
0CA
Experimental Cell Research 249, 22–32 (1999)Article ID excr.1999.4460, available online at http://www.idealibrary.com on
Coordinate Induction of the Actin Cytoskeletal Regulatory ProteinsGelsolin, Vasodilator-Stimulated Phosphoprotein, and Profilin
during Capillary Morphogenesis in VitroRene Salazar, Scott E. Bell, and George E. Davis1
Department of Pathology and Laboratory Medicine, Texas A&M University Health Science Center, College Station, Texas 77843-1114
m
sdpbsrmaTtdl
tmiaibaek5tmcrel
sdVt
The formation of capillaries during developmentnd tissue repair is likely to involve active reorgani-ation of the actin cytoskeleton, although few studiesave addressed this issue. Here, we have utilized an initro model of capillary morphogenesis whereby hu-an umbilical vein endothelial cells are suspendedithin three-dimensional type I collagen gels. The
ells undergo dramatic morphogenic changes to de-elop capillary lumens, tubes, and networks over 72 hf culture. Western blots using cell extracts of theseels over this time frame were performed using anti-odies directed to various proteins associated withhe actin cytoskeleton. Three proteins showed alteredxpression during the time course, and they were gel-olin, which increased fivefold; vasodilator-stimulatedhosphoprotein (VASP), which increased twofold; androfilin, which increased threefold in expression be-ween the 24- and the 72-h time points. Reverse tran-criptase-polymerase chain reaction and Northernlot analysis revealed a similar increase in mRNA ex-ression of the three proteins. After the onset of net-ork formation, the differentiated endothelial cells
dECs) undergoing capillary morphogenesis were re-oved from collagen gels at 48 h of culture to compare
heir properties with untreated endothelial cellsuECs). These dECs showed two- to threefold in-reased spontaneous migration in Boyden chamberssays compared to uECs. The dECs also displayed arominent spindle-shaped morphology and the novelresence of intranuclear gelsolin compared to uECshen both cell types were replated on type I collagen-
oated microwells and glass coverslips. These datauggest that increased gelsolin, VASP, and profilin ex-ression may play an important role in the regulationf capillary tube and network formation in three-di-ensional extracellular matrix. © 1999 Academic Press
1 To whom correspondence and reprint requests should be ad-ressed at the Department of Pathology and Laboratory Medicine,exas A&M University Health Science Center, 208 Reynolds Medi-al Bldg., College Station, TX 77843-1114. Fax: (409) 862-1299. E-
fail: [email protected].
22014-4827/99 $30.00opyright © 1999 by Academic Pressll rights of reproduction in any form reserved.
Key Words: gelsolin; VASP; profilin; capillaryorphogenesis.
INTRODUCTION
Reorganization of the actin cytoskeleton has beenhown to occur within cells during processes such asifferentiation, development, migration, and wound re-air [1–8]. Cells frequently need to change shape andecome more motile to facilitate these processes. The cellhape changes, as well as increases in motility, requireeorganization of the actin cytoskeleton. In addition,odulation of cell shape itself regulates gene expression
nd, thus, can induce cellular differentiation [9, 10].hree actin cytoskeletal regulatory proteins that appearo play a role in cytoskeletal reorganization and signalinguring these processes are gelsolin, vasodilator-stimu-ated phosphoprotein (VASP), and profilin.
Gelsolin is an 82-kDa actin filament-severing proteinhat functions both as a nucleating agent for actinonomers and as a capping protein for the fast-grow-
ng ends of actin filaments [11, 12]. It is an evolution-rily conserved protein found in a wide range of organ-sms, from lower eukaryotes to mammals [13]. Theiologic activity of gelsolin requires calcium ions [14]nd is inhibited by phosphoinositides [15]. Gelsolinxpression has also been shown to be induced duringeratinocyte, myeloid, and myocyte differentiation [2,, 7], while its expression appears to be reduced inumorigenic cell lines [16, 17]. Gene transfer experi-ents have shown that overexpression of gelsolin in-
reased fibroblast migratory ability [18]. Also, a recenteport strongly suggests that gelsolin is a downstreamffector of rac activation, which is known to induceamellipodia formation [19].
VASP is a 45- to 50-kDa phosphoprotein that is as-ociated with areas of the cell membrane undergoingynamic remodeling such as lamellipodia [20, 21].ASP is known to bind profilin, a small 13-kDa protein
hat caps and sequesters G-actin [21], as well as the
ocal adhesion proteins vinculin and zyxin [22, 23].
Vaa
lIptt2apte
aapsRcrcms[cvcdtnatrf
ppTmsintpCpssnwtpt
tv
(bcwtcr
t((LGtjCa
awwm4was(Na
wibsehitvmibSwb1Bntw3jbspat
l
23ACTIN CYTOSKELETAL REGULATION IN CAPILLARY FORMATION
ASP is also known to be phosphorylated by cAMP-nd cGMP-dependent protein kinases in response toctivation of second-messenger pathways [20].Profilin is a 13-kDa ubiquitous protein which regu-
ates actin filament assembly and disassembly [24, 25].t is known to have affinity for actin monomers, phos-hoinositides, and polyproline sequences [24]. This lat-er affinity is responsible for the ability of VASP andhe rho effector protein, p140mDia, to bind profilin [21,6]. The interaction of profilin with these two proteinsnd phosphoinositides are examples of how profilinarticipates in signal transduction pathways and linkshese pathways to the regulation of the actin cytoskel-ton.The formation of blood vessels during development
nd tissue repair [27, 28] is a process that may involven intricate regulation of the actin cytoskeleton. Thisresumption is due to the marked endothelial cellhape changes that occur during morphogenic events.ecently, we developed an in vitro system to study
apillary morphogenesis, whereby the relevant stepsegulating the formation of endothelial cell-lined tubesould be examined in a three-dimensional extracellularatrix. In this system, human endothelial cells are
uspended in type I collagen gels over a 3-day period29]. The endothelial cells undergo dramatic shapehanges initiated by the formation of intracellularacuoles. These vacuoles coalesce to form the lumenalompartment within individual or small groups of en-othelial cells. The endothelial cells then interconnecthrough cell–cell contacts to form capillary tubes andetworks. These marked changes in cell shape implyn active reorganization of the actin cytoskeleton, al-hough few studies to date have examined the potentialole of actin-regulatory proteins in endothelial cellunction [30] or morphogenic events.
In this study, we show that gelsolin, VASP, androfilin are coordinately induced during capillary mor-hogenesis in a three-dimensional collagen matrix.his induction occurs at both the protein and theRNA level. In contrast, the expression levels of a
eries of other cytoskeletal regulatory proteins, includ-ng actin, a-actinin, paxillin, and focal adhesion ki-ase, are not altered during this process. Furthermore,he increased levels of these three proteins are accom-anied by changes in cell morphology and behavior.ells expressing elevated levels of gelsolin, VASP, androfilin (differentiated endothelial cells or dECs) as-ume a more spindle-shaped morphology and increasedpontaneous migratory ability. These dECs show theovel appearance of gelsolin antigen within nuclei,hile untreated endothelial cells (uECs) do not show
his staining. These data implicate the potential im-ortance of gelsolin, VASP, and profilin in the regula-
ion of capillary morphogenesis, and to our knowledge, dhis is the first report indicating their potential rele-ance in the control of capillary formation.
MATERIALS AND METHODS
Cells and tissue culture. Human umbilical vein endothelial cellsClonetics Corp., San Diego, CA) were propagated as described [29]etween passages 2 and 10. Endothelial cells were removed fromulture plastic flasks using trypsin–EDTA. Trypsin was inactivatedith fetal calf serum and the cells were allowed to recover from
rypsinization for 20 min in Medium 199 containing 10% serum. Theells were then centrifuged and washed to remove serum and wereesuspended in a concentrated form in Medium 199.Antibodies, reagents, and chemicals. Antibodies directed to cy-
oskeletal proteins that were used in this study included gelsolinclone GS-2C4) [31] (Sigma Chemical Co., St. Louis, MO), a-actininChemicon, Temecula, CA), paxillin, p125fak, VASP (Transductionaboratories, Lexington, KY), profilin (immunoGlobe, Grossostheim,ermany), and actin (Amersham, Arlington Heights, IL). Other an-
ibodies used include rabbit anti-mouse horseradish peroxidase-con-ugated IgG and goat anti-rabbit fluorescein-conjugated IgG (DAKO,arpinteria, CA). Type I collagen was obtained from rat tails usingcetic acid extraction as described [32].Capillary morphogenesis system. Purified rat tail collagen was
dded to tubes to which 103 concentrated Medium 199 and NaOHere added in a mixture at 0°C. After thorough mixing, the cellsere added at 106/ml with a final collagen concentration of 3.75g/ml. The cell–collagen mix was then added at 25 ml per well in
.5-mm-diameter microwells. The collagen was allowed to gel andas equilibrated for 30 min at 37°C in a CO2 incubator prior to theddition of culture medium. The serum-free culture medium con-isted of Medium 199 with the reduced-serum II supplement, bFGF40 ng/ml), VEGF (40 ng/ml) (Upstate Biotechnology, Lake Placid,Y), phorbol ester (50 ng/ml) (Sigma), and 50 mg/ml ascorbic acid in100-ml volume per 4.5-mm well, as previously described [29].Preparation of cellular protein and Western blots. Collagen gelsere removed from each well with fine forceps and placed directly
nto boiling SDS sample buffer at 100°C or were digested withacterial collagenase (Sigma) for 10 min at 37°C (10 ml of 10 mg/mlolution per well). The cells were centrifuged and solubilized inlectrophoresis sample buffer plus mercaptoethanol. Previous workas shown that the number of viable endothelial cells changes min-
mally in the collagen matrices over the 3-day time course using aetrazolium assay for cell viability (G. E. Davis, unpublished obser-ations). Thus, cell extracts from different times during capillaryorphogenesis are derived from equal numbers of cells extracted
nto a given volume of SDS–PAGE sample buffer. Samples were thenoiled for 3 min and stored at 220°C until use. Samples were run onDS–PAGE (8.5 or 4–20% running gel) and proteins from these gelsere transferred to PVDF membranes (450 mA for 30 min followedy 500 mA for 1 h). Membranes were then blocked at 25°C for 1 h in0 mM Tris–HCl, pH 7.5, containing 0.1% Tween 20 saline and 1%SA solution on a shaker. Primary antibodies were incubated over-ight in Tris–Tween 20–BSA solution at 4°C, and the next morning,his solution was further incubated for 1 h at 25°C. The blots wereashed with multiple changes of Tris–Tween 20 saline buffer over a0-min period. The secondary antibody, horseradish peroxidase-con-ugated rabbit anti-mouse IgG (DAKO; 1:2000 dilution), was incu-ated for 1 h at 25°C and was followed by multiple Tris–Tween 20aline washes over a 30-min period. Blots were then developed foreroxidase activity using chemiluminescence reagents (Amersham)nd then exposed to X-ray film for various times. Band intensities onhe films were quantitated using an IS-1000 Digital Imager.
Reverse transcriptase polymerase chain reaction (RT-PCR). Col-agen gels at different time points (0, 8, 24, 48, and 72 h) were
igested with collagenase and the cells were pelleted and extracted
wdtap(upCGG5C(TG(GG4nucEmSutp
pmf6NlecRolgiw
gpbrmAtwat1ptirBafmfl
m
wreoAhdwp
I
lIoecpFmtitactd1ppap
piicassbwfiVdcsttiiiD
24 SALAZAR, BELL, AND DAVIS
ith Trizol (Gibco BRL, Grand Island, NY) (1 ml/5 3 106 cells) asescribed [33] to obtain total RNA. RNA was quantitated by spec-rophotometry and 5 mg of total RNA from each time point was useds a template for production of cDNA. Reverse transcription waserformed using random primers for first-strand cDNA synthesisStratagene, La Jolla, CA). Polymerase chain reaction was performedsing samples from each time point (0, 8, 24, and 48 h) to showatterns of expression with gene-specific primers for gelsolin (59-TTTCCAGCCATATCGCCAAC-39, 59-TTCTCTGCCTCGCTG-CTC-39), VASP (59-GGAAAGTCAGCAAGCAGG-39, 59-TGTGCG-AAAGGAGAAGC-39), profilin (59-GGCCAGAAATGTTCGGTG-39,9-ACGGGAGGGATATGGGTA-39), G3PDH (59-GCCAAAAGGGT-ATCATCTC-39, 59-GTAGAGGCAGGGATGATGTTC-39), p125fak
FAK) (59-GACTCACCTGGGTACTGGTATG-39, 59-ATCGCTCT-CACCTGTTGATAG-39), paxillin (59-CAGTCGCCAAAGGA-TCTG-39, 59-GTAGTCCTTGCGACAGTAGGC-39), and a-actinin
59-GGAGCCGAAGAAATCGTG-39, 59-CTGCTCGTTCTCCTG-TTG-39 and 59-TGCTGCTGGAGGTCATCTC-39, 59-CAACACCTT-CAGATGCG-39); fragment sizes were 617 (gelsolin), 482 (VASP),04 (profilin), 350 (G3PDH), 406 (FAK), 350 (paxillin), 492 (a-acti-in), and 599 bp (a-actinin). Samples were obtained from reactionssing 25 cycles for all genes except gelsolin and profilin, for which 30ycles were required to obtain sufficient signal for visualization.ach cycle consisted of 94°C for 30 s, 60°C for 30 s, and 72°C for 2in and was performed in a 2400 Perkin–Elmer thermal cycler.amples were run on 1% agarose minigels and DNA was visualizedsing ethidium bromide. Each gene-specific primer set was used onhree different collagen assay time courses to confirm the expressionattern.RNA preparation and Northern blots. RNA for the different time
oints (0, 8, 24, 48, and 72 h) was obtained as above. Total RNA (3g) was loaded per lane on a 1.2% agarose gel containing Mops and
ormaldehyde and the RNA was transferred to nylon membranes forh using a Turboblotter apparatus (Schleicher & Schuell, Keene,H). The RNA was UV-crosslinked to the membrane using a Strata-
inker (Stratagene) and was then stained with methylene blue tonsure equivalent loading of RNA in each lane. A 617-bp gelsolinDNA probe was obtained by RT-PCR with human dermal fibroblastNA, while a 482-bp VASP and a 350-bp G3PDH cDNA probe werebtained using RT-PCR and endothelial cell RNA. The probes wereabeled with [a-32P]CTP using a random-primer labeling kit (Strata-ene) and were hybridized to the blots at 42°C in formamide hybrid-zation buffer [34]. The blots were exposed to autoradiographic filmith an intensifying screen for varying times at 270°C.Immunofluorescent staining of endothelial cells. After being di-
ested out of the gels with collagenase, endothelial cells were resus-ended in medium containing 20% fetal calf serum, heparin, andovine hypothalamic extract in Medium 199 [35]. These cells wereeseeded onto glass coverslips that were previously coated with 100g/ml type I collagen in phosphate-buffered saline (PBS) for 30 min.fter an overnight incubation at 37°C, endothelial cells were fixed on
he coverslips for 20 min in 2% paraformaldehyde in PBS. Coverslipsere blocked by rinsing several times with a Tris–glycine solutionnd incubated for 15 min in 0.5% TX-100 in PBS. Coverslips werehen incubated overnight in 0.1% TX-100 in PBS containing 1% BSA,% goat serum, and 0.2% sodium azide at 4°C. The following dayrimary antibody was added in the same solution for a 1-h incuba-ion at 25°C. Coverslips were washed 43 with PBS for 10 min andncubated for 1 h in secondary antibody (goat anti-mouse IgG–fluo-escein) at a 1:25 dilution in PBS containing 0.1% TX-100 and 1%SA. Several groups of both cell types were treated with secondaryntibody alone as controls. Coverslips were then washed 43 in PBSor 10 min and mounted onto glass slides using a glycerol-basedounting medium. Slides were visualized using a Nikon Labphotuorescence microscope.Boyden chamber assays. Cells were seeded (15,000/well) in basal
edia for 6 h at 37°C on membranes (8-mm pore) that were coated iith 100 mg/ml type I collagen in a 48-well Boyden chamber appa-atus (Neuroprobe, Cabin John, MD). Lower chambers containedither basal medium alone (M199 1 Reduced Serum II supplement)r basal medium containing 40 ng/ml VEGF as a chemoattractant.fter 6 h of incubation, membranes were fixed in 2% paraformalde-yde for 30 min and stained in 0.5% toluidine blue 1 3.7% formal-ehyde for 2–3 min. The nonmigrated cells on the upper surface wereiped off and the migrated cells were visualized and counted usinghase microscopy and an ocular grid at 2003 magnification.
RESULTS
ncreased Gelsolin, VASP, and Profilin Expressionduring Capillary Morphogenesis in Vitro
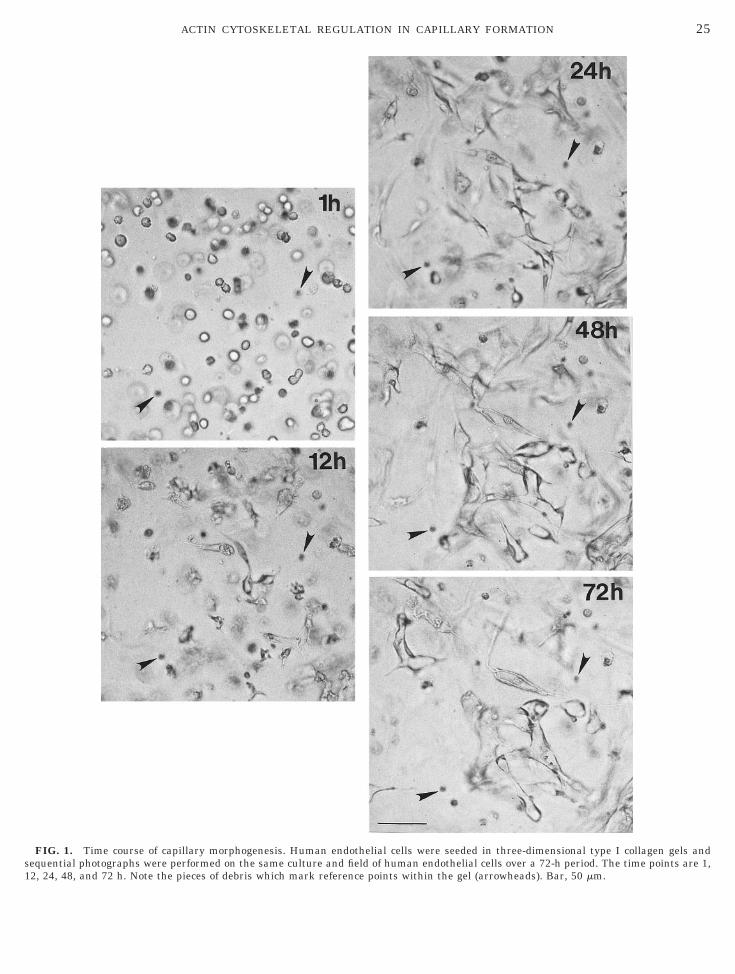
Our previous work has shown that human endothe-ial cells, when resuspended in three-dimensional typecollagen gels, will rearrange to form capillary tubes
ver a 3-day period [29]. In Fig. 1, the same field ofndothelial cells is shown at 1, 12, 24, 48, and 72 h ofulture. The arrows show debris in the field to align theositions of the cells at each time point. As shown inig. 1, the pattern of capillary tubes and networksarkedly changes. This is particularly evident from 24
o 72 h of culture during which considerable remodel-ng of the capillary networks is seen. We hypothesizedhat such changes in capillary morphogenesis would beccompanied by alterations in the expression of actinytoskeletal regulatory proteins. In order to addresshis issue, we prepared endothelial cell extracts atifferent times during capillary formation (0, 2, 4, 8,2, 24, 48, and 72 h). These extracts were then used toerform Western blots to determine if the expression ofroteins known to be important in the regulation of thectin cytoskeleton was altered during capillary mor-hogenesis.As shown in Fig. 2, gelsolin, VASP, and profilin ex-
ression were upregulated in similar temporal fash-ons during this process. The results from multiplendependently prepared time courses are shown. Inontrast, the expressions of actin, a-actinin, paxillin,nd focal adhesion kinase were unchanged during theame time period. Each of the antibodies used in thistudy immunoreacted with only a single band on thelots, which was consistent with the known moleculareight of each protein. As shown in Fig. 3, there is avefold induction of gelsolin, a twofold induction ofASP, and a threefold induction of profilin expressionuring the time course. Actin expression over the timeourse was stable, while an increase in gelsolin expres-ion began at the 24-h time point and continued up tohe 72-h time point. A similar expression pattern overime was seen with VASP, although it appeared to benduced somewhat earlier than gelsolin. Profilin wasnduced in a pattern most similar to gelsolin with anncrease observed between 24 and 72 h of culture.uring the time frame of gelsolin, VASP, and profilin
nduction (i.e., 24 to 48 h of culture), lumens have
s1
25ACTIN CYTOSKELETAL REGULATION IN CAPILLARY FORMATION
FIG. 1. Time course of capillary morphogenesis. Human endothelial cells were seeded in three-dimensional type I collagen gels andequential photographs were performed on the same culture and field of human endothelial cells over a 72-h period. The time points are 1,
2, 24, 48, and 72 h. Note the pieces of debris which mark reference points within the gel (arrowheads). Bar, 50 mm.
fn
elsaG4cTsc
G
et42ufpt
tsfif
tmipswct
D
dVtpdfi(nIip
deadp
gcSgctgpfLt
26 SALAZAR, BELL, AND DAVIS
ormed in individual endothelial cells and capillaryetworks are assembling.To address whether the marked increase in gelsolin
xpression was global or limited to subsets of endothe-ial cells, immunofluorescent staining of three-dimen-ional collagen gel cultures was performed at 24, 48,nd 72 h, using anti-gelsolin and control antibodies.elsolin immunostaining increased between 24 and8 h (as shown earlier); however, its cellular and sub-ellular distributions appear uniform (data not shown).hus, it appeared that the increase in gelsolin expres-ion is present in the overall population of endothelialells rather than in subsets of cells.
elsolin, VASP, and Profilin mRNAs Are Inducedduring Capillary Morphogenesis
RT-PCR analysis was performed to examine the lev-ls of mRNA expression of cytoskeletal-associated pro-eins at 0, 8, 24, and 48 h of culture. As shown in Fig., VASP mRNA expression increased and peaked at4 h, while gelsolin and profilin expression increasedp to the 48-h time point. In contrast, the expression ofocal adhesion kinase, paxillin, and glyceraldehyde-3-hosphate dehydrogenase remained stable throughout
FIG. 2. Cytoskeletal protein expression during capillary morpho-enesis. Human endothelial cells were seeded in three-dimensionalollagen gels and were extracted at various times of culture withDS–PAGE sample buffer. The extracts were run on SDS–PAGEels, were blotted to PVDF membranes, and were probed with mono-lonal or polyclonal antibodies specific for different cytoskeletal pro-eins. The results for three different time courses are shown forelsolin and two different time courses for vasodilator-stimulatedhosphoprotein (VASP), while the others show representative blotsor profilin, actin, a-actinin, paxillin, and focal adhesion kinase.anes are labeled with the time (hours) of the endothelial cell ex-ract.
he time course. The actin filament-crosslinking pro- t
ein, a-actinin, showed a pattern of decreasing expres-ion. This decreasing expression pattern was con-rmed using two different sets of PCR primers specificor a-actinin.
As shown in Fig. 5, Northern blot analysis confirmedhe RT-PCR data indicating that gelsolin and VASPRNAs are induced in a pattern similar to that shown
n Fig. 4. The gelsolin mRNA peaked at the 48-h timeoint and then decreased (Fig. 5). VASP mRNA expres-ion showed a pattern similar to that found in RT-PCR,ith the mRNA appearing to peak at 24 h, while the
ontrol mRNA, G3PDH, showed stable expression overhe time course (Fig. 5).
ifferentiated Endothelial Cells Expressing HigherLevels of Gelsolin, VASP, and Profilin Exhibit aSpindle-Shaped Morphology and IncreasedSpontaneous Migration in Boyden Chamber Assays
To examine in more detail the properties of the en-othelial cells expressing elevated levels of gelsolin,ASP, and profilin, the cells were removed from the
hree-dimensional collagen matrix and either werelated on collagen-coated wells or were tested in Boy-en chamber migration assays using collagen-coatedlters. Cells incubated in type I collagen gels for 48 hdECs) were digested out of the gels and replated over-ight on microwells or glass coverslips coated with typecollagen. As seen in Fig. 6, these cells show a prom-
nent spindle-shaped morphology compared to uECslated under the same conditions. In contrast, uECs
FIG. 3. Expression levels of gelsolin, VASP, profilin, and actinuring capillary morphogenesis. Western blots were visualized usingnhanced chemiluminescence reagents and an IS-1000 digital im-ger. Bands from multiple time courses were quantitated using theensitometer. The values were normalized to 1 using the 2-h timeoint and are expressed as the mean values 6 SD (n 5 4 separate
ime courses for gelsolin, VASP, and actin; n 5 3 for profilin).
tcolse(ecWocpl
VdpBwaVdwptesnm
A
g
aibaAtwsTcspsit
c
R((Rd
cttp(afcltA
27ACTIN CYTOSKELETAL REGULATION IN CAPILLARY FORMATION
hat had not been exposed to three-dimensional matrixonditions showed a typical endothelial cell morphol-gy. The dECs and uECs, when treated with both col-agenase and trypsin to rule out effects of enzymes,howed the same morphological and migratory differ-nces as did those cells treated with only one enzymedata not shown). Despite the marked differences inndothelial cell morphology exhibited by the dECs, theells retained strong immunoreactivity with anti-vonillebrand factor antibodies (data not shown). Previ-
us studies investigating endothelial phenotypichanges resembling a transition to a mesenchymal-likehenotype revealed a prominent decrease in von Wil-ebrand factor staining [36].
The different morphologies and elevated gelsolin,ASP, and profilin expression levels exhibited by theECs isolated from collagen gels prompted us to com-are their migratory activity to that of control uECs inoyden chamber assays. As seen in Fig. 7, with orithout VEGF as a chemoattractant, dECs migratedcross the membrane in similar numbers. In contrast,EGF stimulated uEC migration two- to threefold. TheECs migrated two- to threefold better than uECshen no chemoattractant was added to the lower com-artment of the Boyden chamber. Cell attachment ofhe dECs vs uECs to the collagen-coated filters wasquivalent (data not shown). Thus, the dECs showed atatistically significant (P , 0.01) increase in sponta-eous migratory ability (i.e., without the added che-oattractant) compared to the uECs.
ppearance of Gelsolin within Nuclei ofDifferentiated Endothelial Cells
After overnight plating on glass coverslips, both
FIG. 4. Analysis of cytoskeletal protein mRNA expression usingT-PCR. RT-PCR was performed using primers specific for gelsolin
617 bp), VASP (482 bp), profilin (404 bp), paxillin (350 bp), p125fak
406 bp), a-actinin (492 and 599 bp), and G3PDH (350 bp) using totalNA obtained from HUVEC cultures at 0, 8, 24, and 48 h during theevelopment of capillary tubes and networks.
roups of endothelial cells were immunostained with m
nti-gelsolin, anti-VASP, and/or anti-profilin antibod-es to assess possible differences in the cellular distri-ution of the antigens. Controls treated with secondaryntibody alone showed no staining (data not shown).s seen in Fig. 8, dECs showed a localization of gelsolin
o the actin cytoskeleton as well as to the nucleus,hereas the control endothelial cells showed gelsolin
taining of the actin cytoskeleton, but not the nucleus.he nuclear gelsolin staining was confirmed by confo-al microscopy and optical sections of the cells (data nothown). Western blot analysis of cell extracts of thelated dECs using anti-gelsolin antibodies revealed aingle immunoreactive 82-kDa band consistent withntact gelsolin. No obvious differences were observed inhe distribution of VASP or profilin antigens.
DISCUSSION
Relatively limited information is currently availableoncerning the molecular regulation of blood vessel
FIG. 5. Analysis of gelsolin and VASP mRNA expression duringapillary morphogenesis. Total RNA was isolated from HUVEC cul-ures at 0, 4, 8, 24, 48, and 72 h during the development of capillaryubes and networks. Northern blots were loaded with 3 mg of RNAer lane. Blots were probed with a-32P-labeled cDNAs for gelsolin617 bp), VASP (482 bp), or G3PDH (350 bp). The blots were washednd exposed to autoradiographic film at 270°C. Two different blotsor gelsolin are shown using RNAs isolated from two different timeourses of capillary formation. Equal loads of RNA were seen in eachane by staining the blot with methylene blue prior to the addition ofhe probes. The positions of the 28S and 18S rRNAs are indicated.rrowheads indicate the positions of the gelsolin, VASP, or G3PDH
RNAs.
28 SALAZAR, BELL, AND DAVIS

fcaiescnuccvapp
G
ptgpfiVadtwtcoiatqctfwl
eecTtftbfssaBtgmnwdrip
dpea
tmmueut2mTgphaod(u
29ACTIN CYTOSKELETAL REGULATION IN CAPILLARY FORMATION
ormation. This is particularly evident in the area con-erning the mechanisms required for capillary lumennd tube formation. To this end, we have developed ann vitro model of capillary morphogenesis using humanndothelial cells resuspended within three-dimen-ional collagen matrices [29]. Over a 72-h period, theells rearrange from single cells to an interconnectingetwork of capillary tubes. Our general interest is totilize this system to identify endothelial cell mole-ules that are required for morphogenic events such asapillary lumen, tube, and network formation. An ad-antage of our system, as illustrated in the currentrticle, is its utility in identifying differentially ex-ressed genes that may be relevant to capillary mor-hogenesis.
FIG. 6. Morphology of differentiated endothelial cells during caimensional collagen gels and at 48 h of culture, the cells were digestelastic wells overnight at 37°C in endothelial cell growth medium. Dndothelial cells (uECs, bottom), which were similarly seeded overn
FIG. 7. Boyden chamber migration assays comparing the migra-ory ability of differentiated versus untreated endothelial cells. Hu-an endothelial cells were seeded in three-dimensional collagenatrices and at 48 h of culture, the cells were digested from the gels
sing collagenase. These differentiated cells (dECs) and untreatedndothelial cells (uECs) were seeded at 15,000 cells per well into thepper chamber of a 48-well Boyden chamber apparatus. The migra-ion filter (8-mm pores) was coated with 100 mg/ml type I collagen for
h at room temperature. Both the upper and the lower chamberedia consisted of Medium 199 and Reduced Serum Supplement II.he lower chamber contained either 40 ng/ml vascular endothelialrowth factor or no added growth factor as indicated. The assay waserformed for 6 h at 37°C followed by fixation with paraformalde-yde. Cell migration was quantitated by counting the migrated cellst 2003 using an ocular grid. The data indicate the average numberf migrated cells per well 6 SD (n 5 24 total wells counted from threeifferent experiments). The asterisk indicates statistical significanceP , 0.01) compared to uEC migration without chemoattractantsing the Student t test.
fter plating. Bar, 50 mm.
elsolin, VASP, and Profilin Are CoordinatelyUpregulated during Capillary Morphogenesis inVitro
In this study, we hypothesized that differential ex-ression of components that regulate the actin cy-oskeleton may be important during capillary morpho-enesis in vitro. Here, we show that the actin-severingrotein, gelsolin, is markedly upregulated (approx.vefold) during capillary morphogenesis in vitro. Also,ASP and profilin, two proteins that are known to formcomplex within cells [21], were also coordinately in-
uced (two- to threefold) along with gelsolin duringhese morphogenic events. This increased expressionas observed at both protein and mRNA levels. The
ime course of their induction correlated with a stage ofapillary formation at which dramatic reorganizationf capillary tubes and networks was observed. Thenduction of three distinct proteins that participate inctin cytoskeletal regulation and signaling implieshat a cascade of actin-regulatory genes may be re-uired for capillary tube and network formation. Inontrast, the expressions of other cytoskeletal regula-ory proteins such as actin, a-actinin, paxillin, andocal adhesion kinase were not differentially regulated,hile a-actin in may be downregulated at the mRNA
evel.Previous work has shown that increased gelsolin
xpression occurs during other types of cellular differ-ntiation. This has been demonstrated during leuko-yte, keratinocyte, and myocyte differentiation [5–7].his work coupled with the current study indicateshat gelsolin expression is altered during cellular dif-erentiation and morphogenesis in a variety of con-exts. Very recent studies implicate a close relationshipetween the GTPase rac, which induces lamellipodiaormation, and gelsolin, which appears to be a down-tream effector of rac. In fibroblasts isolated from gel-olin knockout mice, rac overexpression occurs and isccompanied by a defect in lamellipodia formation [19].oth of these cellular changes can be corrected by
ransfecting the normal gelsolin gene back into theelsolin-knockout fibroblasts. The gelsolin-knockoutice also appear to have a defect in wound healing and
eutrophil migration speed [37]. One possibility whichas not mentioned in this work is that endothelial cellefects could play a role in the delayed wound healingesponse. Additional work implicates a role for gelsolinn the regulation of apoptosis [38, 39]. Gelsolin can beroteolytically cleaved with the apoptotic protease,
ary morphogenesis. Human endothelial cells were seeded in three-rom the gels using collagenase and were seeded onto collagen-coatedrentiated endothelial cells (dECs, top) were compared to untreatedt on collagen-coated wells. Both cell types were photographed 24 h
pilld fiffeigh
civpitpi
cpaVdsVam
rfiiasspscpttcVmrtspts
E
eidtssehadwtqaccItentiwiwsa
awtwicstcfld
30 SALAZAR, BELL, AND DAVIS
aspase-3, generating an NH2-terminal fragment thatnduces apoptosis [38]. During the process of bloodessel formation and maturation, both capillary mor-hogenesis and regression occur concurrently [40]. Annduction of gelsolin might be relevant in either one ofhese phenomena, and its elevated endothelial cell ex-ression may facilitate the process of apoptosis in cap-llaries that are to undergo regression.
VASP expression has been associated with dynami-ally changing areas of the cell periphery, such as inrotruding lamellae of locomoting cells and in cell–cellnd focal contact sites in a variety of cells [20, 21].ASP is known to be a substrate for cAMP- and cGMP-ependent kinases in some cell types, such as vascularmooth muscle cells and endothelial cells [41, 42].ASP is known to interact with profilin, which bindsctin monomers and thus regulates actin microfila-ent assembly/disassembly [21, 24, 25].
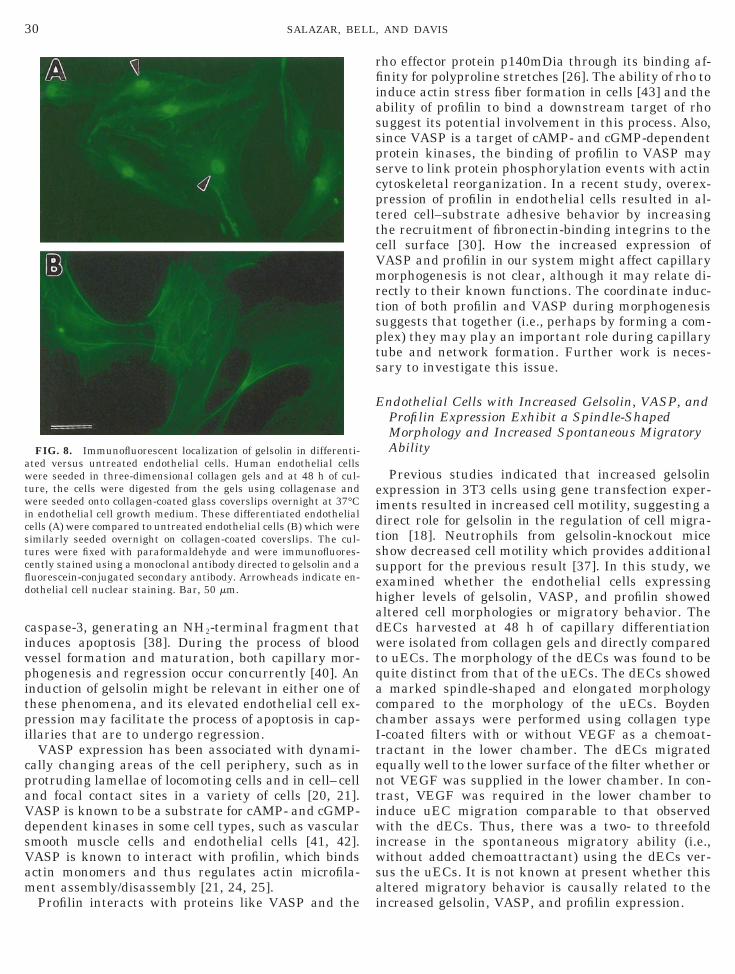
FIG. 8. Immunofluorescent localization of gelsolin in differenti-ted versus untreated endothelial cells. Human endothelial cellsere seeded in three-dimensional collagen gels and at 48 h of cul-
ure, the cells were digested from the gels using collagenase andere seeded onto collagen-coated glass coverslips overnight at 37°C
n endothelial cell growth medium. These differentiated endothelialells (A) were compared to untreated endothelial cells (B) which wereimilarly seeded overnight on collagen-coated coverslips. The cul-ures were fixed with paraformaldehyde and were immunofluores-ently stained using a monoclonal antibody directed to gelsolin and auorescein-conjugated secondary antibody. Arrowheads indicate en-othelial cell nuclear staining. Bar, 50 mm.
Profilin interacts with proteins like VASP and the i
ho effector protein p140mDia through its binding af-nity for polyproline stretches [26]. The ability of rho to
nduce actin stress fiber formation in cells [43] and thebility of profilin to bind a downstream target of rhouggest its potential involvement in this process. Also,ince VASP is a target of cAMP- and cGMP-dependentrotein kinases, the binding of profilin to VASP mayerve to link protein phosphorylation events with actinytoskeletal reorganization. In a recent study, overex-ression of profilin in endothelial cells resulted in al-ered cell–substrate adhesive behavior by increasinghe recruitment of fibronectin-binding integrins to theell surface [30]. How the increased expression ofASP and profilin in our system might affect capillaryorphogenesis is not clear, although it may relate di-
ectly to their known functions. The coordinate induc-ion of both profilin and VASP during morphogenesisuggests that together (i.e., perhaps by forming a com-lex) they may play an important role during capillaryube and network formation. Further work is neces-ary to investigate this issue.
ndothelial Cells with Increased Gelsolin, VASP, andProfilin Expression Exhibit a Spindle-ShapedMorphology and Increased Spontaneous MigratoryAbility
Previous studies indicated that increased gelsolinxpression in 3T3 cells using gene transfection exper-ments resulted in increased cell motility, suggesting airect role for gelsolin in the regulation of cell migra-ion [18]. Neutrophils from gelsolin-knockout micehow decreased cell motility which provides additionalupport for the previous result [37]. In this study, wexamined whether the endothelial cells expressingigher levels of gelsolin, VASP, and profilin showedltered cell morphologies or migratory behavior. TheECs harvested at 48 h of capillary differentiationere isolated from collagen gels and directly compared
o uECs. The morphology of the dECs was found to beuite distinct from that of the uECs. The dECs showedmarked spindle-shaped and elongated morphology
ompared to the morphology of the uECs. Boydenhamber assays were performed using collagen type-coated filters with or without VEGF as a chemoat-ractant in the lower chamber. The dECs migratedqually well to the lower surface of the filter whether orot VEGF was supplied in the lower chamber. In con-rast, VEGF was required in the lower chamber tonduce uEC migration comparable to that observedith the dECs. Thus, there was a two- to threefold
ncrease in the spontaneous migratory ability (i.e.,ithout added chemoattractant) using the dECs ver-
us the uECs. It is not known at present whether thisltered migratory behavior is causally related to the
ncreased gelsolin, VASP, and profilin expression.
P
taTtesnTpcr(dcmpetnnwnnsgptsl
R
gpmfc(ietaisd3pt
f
fm(
1
1
1
1
1
1
1
1
31ACTIN CYTOSKELETAL REGULATION IN CAPILLARY FORMATION
resence of Gelsolin within Differentiated EndothelialCell Nuclei
Although usually associated with the cytoplasm,here is accumulating evidence that components of thectin cytoskeleton are present in the nucleus [44, 45].he significance of these findings is not clear althoughhey may play a role in regulating the nuclear cytoskel-ton and thereby influence gene expression. In thistudy, we show that gelsolin can be detected withinuclei of the plated dECs but not in the control uECs.o our knowledge, this is the first report showing theresence of gelsolin within nuclei (Fig. 8). Nuclear lo-alization was confirmed using confocal immunofluo-escence microscopy and optical sections of the cellsdata not shown). Thus far, we have been unable toetect gelsolin using immunofluorescence within nu-lei of the endothelial cells while they are undergoingorphogenesis in three-dimensional collagen gels (un-
ublished observations). The nuclear translocationvent appears to occur following removal of the endo-helial cells from collagen gels and plating them over-ight on collagen-coated glass coverslips. This phe-omenon is not dependent on treatment of the cellsith trypsin or collagenase, either alone or in combi-ation (data not shown). A motif search using knownuclear localization signals has failed to reveal anyuch sequences in gelsolin [46]. One possibility is thatelsolin is translocated by interacting with anotherrotein that regulates its nuclear entry. Interestingly,he macrophage actin-binding protein, CapG, whichhows sequence homology to gelsolin, is also known toocalize to nuclei [47].
ole of Gelsolin, VASP, and Profilin in CapillaryMorphogenesis
What are the potential mechanisms whereby increasedelsolin, VASP, or profilin may regulate capillary mor-hogenesis? Possibilities include: (i) an increased require-ent for actin microfilament reorganization necessary
or proper alignment and arrangement of endothelialells within developing capillary tubes and networks andii) alterations in the cytoskeletal scaffolding regulatingntracellular signaling pathways or gene expression, nec-ssary to control endothelial cell differentiation. At thisime, we cannot distinguish between these possibilities,lthough both may be involved. The known direct andndirect links of gelsolin, VASP, and profilin with majorignal transduction pathways regulating morphogenesis,ifferentiation, and apoptosis in other systems [2, 20, 24,8] further implicate their likely role in capillary mor-hogenesis. Additional work is necessary to elucidateheir specific role in capillary morphogenesis.
The authors thank Dr. Nancy Dawson for her help with the con-
ocal microscopy, Drs. Julian Leibowitz and Steve Maxwell for help-ul discussions, and Kayla Bayless for helpful comments on theanuscript. This work was supported by NIH Grant HL 59373
G.E.D.)
REFERENCES
1. Ankenbauer, T., Kleinschmidt, J. A., Vanderkerckhove, J., andFranke, W. (1988). Proteins regulating actin assembly in oogen-esis and early embryogenesis of Xenopus laevis: Gelsolin is themajor cytoplasmic actin-binding protein. J. Cell Biol. 107,1489–1498.
2. Kwiatkowski, D. J. (1988). Predominant induction of gelsolinand actin-binding protein during myeloid differentiation.J. Biol. Chem. 263, 13857–13862.
3. Dieffenbach, C. W., SenGupta, D. N., Krause, D., Sawzak, D.,and Silverman, R. H. (1989). Cloning of murine gelsolin and itsregulation during differentiation of embryonal carcinoma cells.J. Biol. Chem. 264, 13281–13288.
4. Grinnell, F. (1992). Wound repair, keratinocyte activation andintegrin modulation. J. Cell Sci. 101, 1–5.
5. Schwartz, S. B., Higgins, P. J., Ayyappan, K. R., and Staiano-Coico, L. (1994). Gelsolin expression in normal human keratin-ocytes is a function of induced differentiation. Adv. Exp. Med.Biol. 358, 169–181.
6. Watts, R. G. (1995). Role of gelsolin in the formation and orga-nization of triton-soluble F-actin during myeloid differentiationof HL-60 cells. Blood 85, 2212–2221.
7. Scholz, A., and Hinssen, H. (1995). Biphasic pattern of gelsolinexpression and variations in gelsolin-actin interactions duringmyogenesis. Exp. Cell Res. 219, 384–391.
8. Arora, P. D., and McCulloch, C. A. G. (1996). Dependence offibroblast migration on actin severing activity of gelsolin.J. Biol. Chem. 217, 20516–20523.
9. Mooney, D. J., Langer, R., and Ingber, D. E. (1995). Cytoskel-etal filament assembly and the control of cell spreading andfunction by extracellular matrix. J. Cell Sci. 108, 2311–2320.
0. Chen, C. S., Mrksich, M., Huang, S., Whitesides, G. M., andIngber, D. E. (1997). Geometric control of life and death. Science276, 1425–1428.
1. Yin, H. L., and Stossel, T. P. (1979). Control of cytoplasmic actingel-sol transformation by gelsolin, a calcium-dependent regula-tory protein. Nature 281, 583–586.
2. Kwiatkowski, D. J., Janmey, P. A., and Yin, H. L. (1989).Identification of critical functional and regulatory domains ingelsolin. J. Cell Biol. 108, 1717–1726.
3. Kwiatkowski, D. J., Stossel, T. P., Orkin, S. H., Mole, J. E.,Colten, H. R., and Yin, H. L. (1986). Plasma and cytoplasmicgelsolins are encoded by a single gene and contain a duplicatedactin-binding domain. Nature 323, 455–458.
4. Yin, H. L., Zaner, K. S., and Stossel, T. P. (1980). Ca21 controlof actin gelation. J. Biol. Chem. 255, 9494–9500.
5. Janmey, P. A., and Stossel, T. P. (1989). Gelsolin–polyphosphoi-nositide interaction: Full expression of gelsolin-inhibiting func-tion by polyphosphoinositides in vesicular form and inactiva-tion by dilution, aggregation, or masking of the inositol headgroup. J. Biol. Chem. 264, 4825–4831.
6. Banyard, M. R., Medveczyk, C. J., and Tellam, R. L. (1990).Microfilament organization correlates with increased cellularcontent of gelsolin. Exp. Cell Res. 187, 180–183.
7. Tanaka, M., Mullauer, L., Ogiso, Y., Fujita, H., Moriya, S.,Furuuchi, K., Harabayashi, T., Shinohara, N., Koyanagi, T.,and Kuzumaki, N. (1995). Gelsolin: A candidate for suppressor
of human bladder cancer. Cancer Res. 55, 3228–3232.
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
4
4
4
4
4
4
4
RR
32 SALAZAR, BELL, AND DAVIS
8. Cunningham, C. C., Stossel, T. P., and Kwiatkowski, D. J.(1991). Enhanced motility in NIH3T3 fibroblasts that overex-press gelsolin. Science 251, 1233–1236.
9. Azuma, T., Witke, W., Stossel, T. T., Hartwig, J. H., and Kwi-atkowski, D. J. (1998). Gelsolin is a downstream effector of racfor fibroblast motility. EMBO J. 17, 1362–1370.
0. Reinhard, M., Halbrugge, M., Scheer, U., Wiegand, C., Jock-usch, B. M., and Walter, U. (1992). The 46/50 kDa phosphopro-tein VASP purified from human platelets is a novel proteinassociated with actin filaments and focal contacts. EMBO J. 11,2063–2070.
1. Reinhard, M., Griehl, K., Abel, K., Haffner, C., Jarchau, T.,Hoppe, V., Jockusch, B. M., and Walter, U. (1995). The proline-rich focal adhesion and microfilament protein VASP is a ligandfor profilins. EMBO J. 14, 1583–1589.
2. Reinhard, M., Jouvenal, K., Tripier, D., and Walter, U. (1995).Identification, purification, and characterization of zyxin-re-lated protein that binds the focal adhesion and microfilamentprotein VASP (vasodilator-stimulated phosphoprotein). Proc.Natl. Acad. Sci. USA 92, 7956–7960.
3. Brindle, N. P. J., Holt, M. R., Davies, J. E., Price, C. J., andCritchley, D. R. (1996). The focal-adhesion vasodilator-stimu-lated phosphoprotein (VASP) binds to the proline-rich domainin vinculin. Biochem. J. 318, 753–757.
4. Sohn, R. H., and Goldschmidt, P. J. (1994). Profilin: At thecrossroads of signal transduction and the actin cytoskeleton.BioEssays 16, 465–472.
5. Schafer, D. A., and Cooper, J. A. (1995). Control of actin assem-bly at filament ends. Annu. Rev. Cell Dev. Biol. 11, 497–518.
6. Watanabe, N., Madaule, P., Reid, T., Ishizaki, T., Watanabe, G.,Kakizuka, A., Saito, Y., Nakao, K., Jockusch, B. M., and Naru-miya, S. (1997). P140mDia, a mammalian homolog of Drosoph-ila diaphanous, is a target protein for Rho small GTPase and isa ligand for profilin. EMBO J. 16, 3044–3056.
7. Vernon, R. B., and Sage, E. H. (1995). Between molecules andmorphology. Am. J. Pathol. 147, 873–883.
8. Senger, D. R. (1996). Molecular framework for angiogenesis—Acomplex web of interactions between extravasated plasma pro-teins and endothelial cell proteins induced by angiogenic cyto-kines. Am. J. Pathol. 149, 1–7.
9. Davis, G. E., and Camarillo, C. (1996). An alpha-2-beta-1 inte-grin-dependent pinocytotic mechanism involving intracellularvacuole formation and coalescence regulates capillary lumenand tube formation in three-dimensional collagen matrix. Exp.Cell Res. 224, 39–51.
0. Moldovan, N. I., Milliken, E. E., Irani, K., Chen, J., Sohn, R. H.,Finkel, T., and Goldschmidt-Clermont, P. J. (1997). Regulationof endothelial cell adhesion by profilin. Curr. Biol. 7, 24–30.
1. Chaponnier, C., Janmey, P. A., and Yin, H. L. (1986). The actinfilament-severing domain of plasma gelsolin. J. Cell Biol. 103,1473–1481.
2. Bornstein, M. B. (1958). Reconstituted rat-tail collagen used asa substrate for tissue cultures on coverslips in Maximov slides
and roller tubes. Lab. Invest. 7, 134–137.3. Chomczynski, P., and Sacchi, N. (1987). Single-step method ofRNA isolation by acid guanidinium thiocyanate–phenol–chlo-roform extraction. Anal. Biochem. 162, 156–159.
4. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Analysisand cloning of eukaryotic genomic DNA. In “Molecular Cloning:A Laboratory Manual” (N. Ford, C. Nolan, and M. Ferguson,Eds.), Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, NY.
5. Maciag, T., Cerundolo, J., Lisley, S., Kelley, P. R., and Forand,R. (1979). An endothelial cell growth factor from bovine hypo-thalamus: Identification and partial characterization. Proc.Natl. Acad. Sci. USA 76, 5674–5678.
6. Lipton, B. H., Bensch, K. G., and Karasek, M. A. (1991). Mi-crovessel endothelial cell transdifferentiation: Phenotypic char-acterization. Differentiation 46, 117–133.
7. Witke, W., Sharpe, A. H., Hartwig, J. H., Azuma, T., Stossel,T. P., and Kwiatkowski, D. J. (1995). Hemostatic, inflamma-tory, and fibroblast responses are blunted in mice lacking gel-solin. Cell 81, 41–51.
8. Kothakota, S., Azuma, T., Reinhard, C., Klippel, A., Tang, J.,Chu, K., McGarry, T. J., Kirschner, M. W., Koths, K., Kwiat-kowski, D. J., and Williams, L. T. (1997). Caspase-3-generatedfragment of gelsolin: Effector of morphological change in apo-ptosis. Science 278, 294–298.
9. Ohtsu, M., Sakai, N., Fujita, H., Kashiwagi, M., Gasa, S.,Shimizu, S., Eguchi, Y., Tsujimoto, Y., Sakiyama, Y., Koba-yashi, K., and Kuzumaki, N. (1997). Inhibition of apoptosis bythe actin-regulatory protein gelsolin. EMBO J. 16, 4650–4656.
0. Folkman, J., and Haudenschild, C. (1980). Angiogenesis invitro. Nature 288, 551–556.
1. Draijer, R., Vaandrager, A. B., Nolte, C., de Jonge, H. R.,Walter, U., and van Hinsbergh, V. W. M. (1995). Expression ofcGMP-dependent protein kinase I and phosphorylation of itssubstrate, vasodilator-stimulated phosphoprotein, in humanendothelial cells of different origin. Circ. Res. 77, 897–905.
2. Markert, T., Krenn, V., Leebman, J., and Walter, U. (1996).High expression of the focal adhesion- and microfilament-asso-ciated protein VASP in vascular smooth muscle and endothelialcells of the intact human vessel wall. Basic Res. Cardiol. 9,337–343.
3. Ridley, A. J., and Hall, A. (1992). The small GTP-binding pro-tein rho regulates the assembly of focal adhesions and actinstress fibers in response to growth factors. Cell 70, 389–399.
4. Ankenbauer, T., Kleinschmidt, J. A., Walsh, M. J., Weiner,O. H., and Franke, W. W. (1989). Identification of a widespreadnuclear actin binding protein. Nature 342, 822–825.
5. Gant, T. M., and Wilson, K. L. (1997). Nuclear assembly. Annu.Rev. Cell Dev. Biol. 13, 669–695.
6. Boulikas, T. (1993). Nuclear localization signals (NLS). Crit.Rev. Eukaryotic Gene Exp. 3, 193–227.
7. Onoda, K., Yu, F. X., and Yin, H. L. (1993). GCap39 is a nuclear
and cytoplasmic protein. Cell Motil. Cytoskeleton 26, 227–238.eceived November 11, 1998evised version received February 25, 1999





























