Embed Size (px)
Citation preview

[CANCER RESEARCH 44, 909-914, March 1984]
Comparison of 5-Fluorouracil Metabolism in Two Human GastrointestinalTumor Cell Lines1
Wendy L. Washtien2
Department of Pharmacology and the Cancer Center, Northwestern University Medical School, Chicago, Illinois 60611
ABSTRACT
HuTu 80 and HT 29 are two human gastrointestinal tumor celllines which differ 20-fold in their sensitivity to growth inhibitionby 5-fluorouracil (FUra) [concentration of inhibitor necessary to
inhibit cell growth 50% as compared with control cells grownunder identical conditions (except that inhibitor was omitted):HuTu 80, 6 /¿M;HT 29, 0.3 /UM].Drug response in the two celllines is further distinguished by the ability of thymidine to reversethe growth inhibition produced by FUra in HT 29 cells but not inHuTu 80 cells. Furthermore, the presence of deoxyinosine potentiates FUra action in HuTu 80 cells, while having no effect ongrowth inhibition by FUra in HT 29 cells.
The metabolism of [6-3H]FUra was examined in these cell lines
using techniques which allow simultaneous measurement of: (a)the amount of 5-fluoro-2'-deoxyuridylate bound to thymidylate
synthetase; (b) the level of free thymidylate synthetase; (c) FUraincorporation into RNA; and (d) acid-soluble metabolites of FUra.
A comparison of FUra metabolism in the two cell lines, followinga 24-hr exposure of the cells to equitoxic concentrations of FUra,
demonstrated that such exposure led to thymidylate synthetaseinhibition in both HuTu 80 and HT 29 cells. This inhibition,however, was accompanied by a 6-fold greater level of incorpo
ration of FUra into RNA in HuTu 80 cells than in HT 29 cells, andsuggests that drug incorporation into RNA may be the dominantdeterminant of growth inhibition by FUra in HuTu 80 cells.
Deoxyinosine, which potentiates FUra action in HuTu 80 cells,was found to alter FUra metabolism in the cells so that thymidylate synthetase inhibition was now achieved under conditionsof reduced FUra incorporation into RNA. Thymidine was able toreverse growth inhibition of HuTu 80 cells by the FUra-deoxyi-
nosine combination, suggesting a change in the mechanism ofgrowth inhibition.
INTRODUCTION
The pyrimidine analogue FUra3 is an antitumor agent which is
widely used in the treatment of a variety of neoplasms. Thisdrug, following activation to nucleotide derivatives, is currentlythought to exert its major cytotoxic and chemotherapeutic effects by 2 mechanisms: (a) through inhibition of dTMP synthetase(EC 2.1.1.45) by the metabolite FdUMP; and (b) by incorporation(at the level of FUTP) into cellular RNA (14, 23). The former,which occurs through formation of a ternary complex, containing
'This study was supported in part by Grant CA 277021 from the National
Cancer Institute.2To whom requests for reprints should be addressed.3The abbreviations used are: FUra, 5-fluorouracil; CHjFH4, 5,10-methylene-
tetrahydrofolate; dTMP synthetase, thymidylate synthetase; FdUrd, 5-fluoro-2'-deoxyuridine; FdUMP, 5-fluoro-2'-deoxyuridine 5'-monophosphate; FUTP, fluo-
rouridine triphosphate; dThd, thymidine; EC«,,that concentration of inhibitor necessary to inhibit cell growth 50% as compared with control cells grown underidentical conditions, except that the inhibitor was omitted.
Received September 1,1983; accepted November 21,1983.
the enzyme, FdUMP, and the CH2FH4 cofactor (7, 28), results indepletion of cellular thymidylate pools and a cessation of DNAsynthesis. FUra incorporation into RNA has been shown to affectRNA maturation (36). Recent reports have highlighted yet another possible cytotoxic mechanism for FUra, the incorporationof fluorodeoxyuridine triphosphate into cellular DNA (6, 20).
The variable response rate which is seen clinically with FUrais mirrored by the variability in sensitivity observed among cultured cells and experimental tumors (1, 8, 15, 17, 18, 23, 26,30). In an attempt to understand the biochemical basis fordifferences in drug sensitivity, investigators have examined manybiochemical parameters which might contribute to the sensitivityof cells and tumors to FUra. Studies have considered enzymeactivities necessary for FUra activation (1,18, 25, 27), inhibitionof DNA synthesis following drug exposure (9,17, 22), expansionand maintenance of FdUMP pools (1,17, 24), and levels of FUraincorporation into cellular RNA (3, 9, 21). Such studies havedemonstrated measurable differences among cells in many ofthese parameters and, in some cases, have shown correlationsbetween the measured parameters and cell response. Thesestudies, however, have often focused on only one or 2 parameters which are related to FUra activity, and it has been suggestedthat a more comprehensive measurement of FUra metabolismmight be useful in better defining the biochemical reasons fordifferential sensitivity of cells to FUra (23, 26, 35). Technicaladvances have made such measurements feasible (22, 26, 35),although such techniques have not, to date, been used tocompare FUra metabolism in cell lines of differing sensitivity toFUra.
In this paper, we use such a comprehensive approach tocompare FUra metabolism in 2 human gastrointestinal tumor celllines and examine the relationship between the measured parameters and the observed sensitivity of these cells to FUra.
MATERIALS AND METHODS
Chemicals
[6-3H]FUra (specific activity, 25 Ci/mmol) was purchased from Mora-
vek Biochemicals, City of Industry, CA, and was used within 1 month ofreceipt. Tetrahydrofolic acid was prepared from folie acid (Sigma Chemical Co., St. Louis, MO) by the procedure of Hatefi ef al. (13). Solutionsof CH2FH< (pH 9.5) were stored under vacuum at -80° in 0.12 M ß-
mercaptœthanol and 30 HIM formaldehyde. The actual concentration ofCH2FH4 was determined spectrophotometrically (2). All other chemicalswere purchased from Sigma unless otherwise specified.
Cells and Media
The human carcinoma cell lines, HuTu 80 (30) and HT 29 (11), wereobtained from the Naval Biosciences Laboratory (Berkeley, CA). All cellswere maintained in Dulbecco's modified Eagle's medium supplemented
with 10% fetal calf serum (KC Biologicals, Kansas City, MO) at 37°in
an atmosphere of 92% air:8% CO2. Doubling times for the cells underthese conditions were: HuTu 80, 21 hr; and HT 29, 28 hr. Cells were
MARCH 1984 909
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

W. L. Washtien
monitored semimonthly for the absence of Mycoplasma using theHoechst staining method (5).
Growth Inhibition Studies
Determinations of the growth of tumor cells in the presence of drugswere conducted in Costar (Cambridge, MA) 24-well plates as described
previously (33). Experiments were begun by the addition of 1 ml ofmedium containing 0.7 to 1 x 105 cells to wells in which 0 to 50 ^\ ofdrug had been placed. Following incubation at 37°for 72 to 96 hr, cell
number was determined using a Coulter Counter (Model ZBI) and maximally increased 10- to 15-fold over that inoculated.
Cellular Metabolism of [6-3H]FUra
Cells (0.25 to 1 x 106) were seeded in 75-sq cm tissue culture flasks
and allowed to attach and enter exponential growth (3 days; final density,1 to 3 x 10s cells/sq cm). At this time, medium was replaced with freshmedium containing [6-3H]FUra, and the flasks were placed at 37°in a
humidified 8% CO2 atmosphere for 24 hr. Cells were then removed fromthe flask by gentle trypsinization, collected by centrifugation (1000 x gfor 3 min at 4°),and resuspended in 2 ml of ice-cold phosphate-buffered
saline (0.137 M NaCI; 2.7 rriM KCI; 8 HIM Na2HPO4; 1.47 ITIM KH2PO4;0.68 mM CaCI2; 0.49 rtiM MgCI2).
Intracellular Metabolites and FUra-RNA. One ml of the suspension
of treated cells was extracted as described by Pogolotti ef a/. (26), andthe acid-soluble metabolites of [6-3H]FUra obtained were analyzed by
high-pressure liquid chromatography in the presence of unlabeled standards, using the system described below. The acid-insoluble pellet was
washed with cold trichloroacetic acid and solubilized in tissue solubilizer,and the amount of radioactivity in the solubilized material was determinedby liquid scintillation counting. This radioactivity represents both FUraincorporated into RNA and any FdUMP bound to dTMP synthetase. Thelatter quantity is determined (see below) and is subtracted to provide afinal value for FUra present in cellular RNA. The acid-insoluble pellet may
also include radioactivity present in DNA (6, 20) or protein. Alkalinehydrolysis (32) of acid-insoluble pellets from treated cells indicates that
less than 4% of the radioactivity in the pellet is derived from either ofthese 2 cellular components, and no correction has been made toaccount for these components.
FdUMP Bound to dTMP Synthetase and Free dTMP Synthetase.The remaining 1 ml of FUra-treated cells was used to determine theamount of [3H]FdUMP bound to dTMP synthetase and the amount of
free dTMP synthetase present in the cells, using Sephadex G-25 chromatography and high-pressure liquid chromatography (34). The cellswere centrifugea and resuspended in ice-cold 20 mw sodium phosphate(pH 7.3) containing 10 mw 0-mercaptoethanol and 8 mM formaldehyde.The cell suspension was sonicated (Branson sonifier; setting 4°,3 sec),
and cell debris were removed by centrifugation (15,000 x g for 5 min at4°).Sonicate (0.6 ml) was combined with 0.2 ml of a charcoal suspension
(charcoal, 100 mg/ml; bovine serum albumin, 10 mg/ml; dextran, 0.5mg/ml) to remove endogenous nucleotides (21, 30). Following centrifugation to remove the charcoal, an aliquot (0.2 ml) of sonicate wassubjected to chromatography on Sephadex G-25 (1 x 30 cm).
The void volume of the column (3 to 4 ml), containing the dTMPsynthetase:FdUMP:CH2FH4 complex (the ternary complex formed between these 3 compounds), was pooled and heated at 65°for 15 min
to dissociate FdUMP from the complex (35). Following cooling and theaddition of a known amount of [14C]dThd (5 //I; 50,000 dpm) to serve as
an internal standard for recovery, the solution was brought to 5% intrichloroacetic acid by the addition of an appropriate volume of 50% ice-cold trichloroacetic acid, and the acid-precipitable material was removedby centrifugation (4°;1000 x g for 5 min). The acidic supernatant was
removed and added to an equal volume of cold Freon containing 0.5 Mtri-n-octylamine according to the method of Khym (16). The mixture wasvortexed and, after centrifugation (4°;1000 x g for 5 min), the aqueous
upper phase was removed and concentrated using a Savant Speed Vacconcentrator. The lyophilized sample was taken up in 0.1 ml of water
containing 20 nmol each of unlabeled FUra, 5-fluorouridine, FdUrd, 5-fluorouridine-5'-monophosphate, and FdUMP and analyzed by high-
pressure liquid chromatography using an Altex gradient liquid Chromatograph equipped with a fixed-wavelength (254 nm) UV detector.Separations were performed on a RP-C18 column (Alltech Associates,Deerfield, IL) using an ion-pairing gradient with 1 rnw (n-Bu)4N+HSO4~
and 5 mw potassium phosphate, pH 7, as the low concentration eluentand the same buffer, containing 40% methanol, as the high concentrationeluent. The elution consisted of an isocratic period of 5 min, followed bya 10-min linear gradient to a new isocratic period consisting of 25% high-
concentration buffer, which was maintained for 32 min. At this time, a15-min linear gradient to 100% high-concentration eluent was used to
complete the run. The entire gradient was run at a flow rate of 2 ml/min.Fractions (1 ml) were collected directly into minivials and analyzed forradioactivity (on 3H channel) by liquid scintillation counting. Such mea
surements were performed using 3a70b counting fluid (Research Products International, Elk Grove Village, IL) on a Beckman LS 6800 liquidscintillation counter. Those fractions containing [14C]dThd (which containno 3H) were recounted on a 14C channel. The retention volumes for
standards in this system are: FUra, 9 ml; 5-fluorouridine, 21 ml; FdUrd,25 ml; dThd, 31 ml; 5-fluorouridine 5'-monophosphate, 52 ml; and
FdUMP, 63 ml.The amount of 3H associated with the FdUMP standard, which repre
sents the amount of [3H]FdUMP complexed to dTMP synthetase, was
corrected for recovery during extraction and chromatography, as determined by the amount of [14C]dThd recovered. Recovery was routinely
between 65 and 72%.A second aliquot of charcoal-treated sonicate was incubated with [6-
3H]FdUMP and CH2FH4 for 1 hr (33) to allow binding of [6-3H]FdUMP toany free dTMP synthetase present in the cells. The amount of [3H]-
FdUMP bound to dTMP synthetase was then determined as describedabove. The amount of free dTMP synthetase was calculated by subtracting the amount of [3H]FdUMP found bound to dTMP synthetase insonicates which were not incubated with [6-3H]FdUMP from the valuesobtained for [3H]FdUMP bound to dTMP synthetase in sonicates whichhad been incubated with [6-3H]FdUMP.
Intracellular Accumulation of [3H]FUra
Cells were seeded at 1 x 10s cells/60 mw tissue culture dish 3 days
prior to the day of the experiments. At 1 hr before the beginning of anexperiment, the medium was replaced with 2 ml of fresh medium.Experiments were begun with the addition of [6-3H]FUra (specific activity,
1.8 Ci/mmol; final concentration, 0.5 ^M) to each plate. The time courseof [3H]FUra accumulation was measured as described by Schilsky ef a/.
(29).
RESULTS
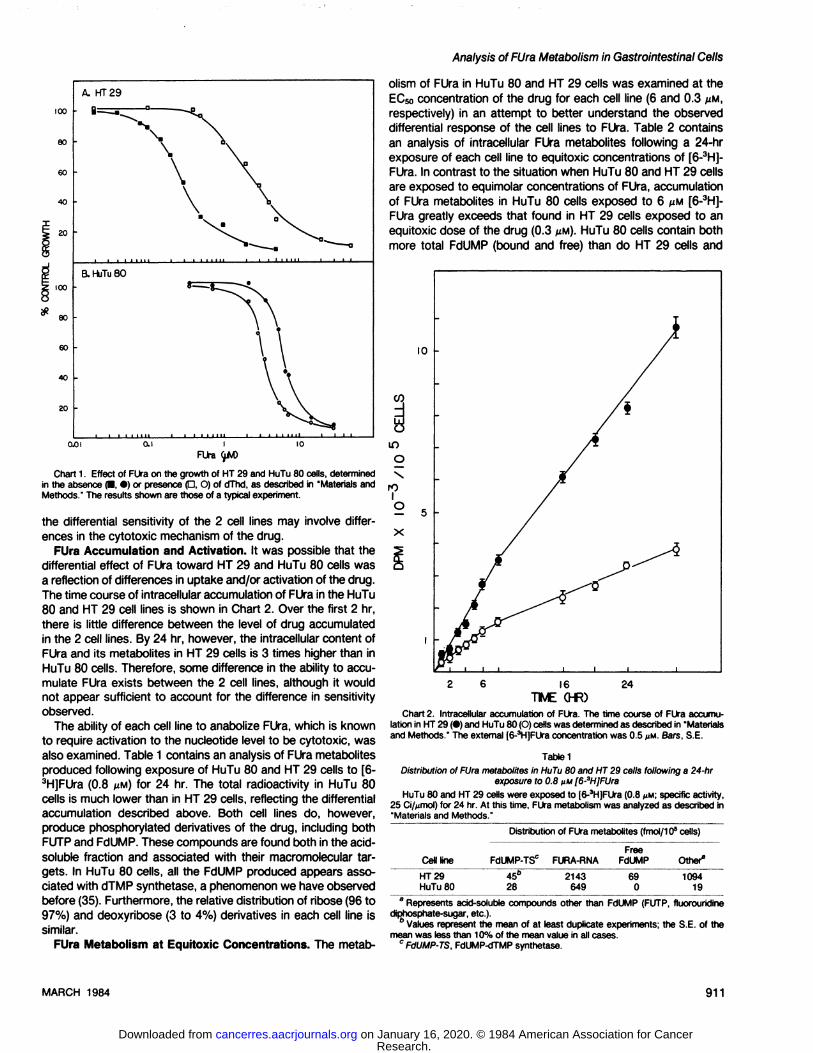
Growth Inhibition by FUra. Chart 1 shows the inhibition ofgrowth of 2 cultured human gastrointestinal tumor cell lines,HuTu 80 and HT 29, by the fluoropyrimidine FUra. HuTu 80 cellsare 20-fold less sensitive to this compound than are HT-29 cells.
The EC50 for HuTu 80 cells was 6 MM, and that of HT 29 cellswas 0.3 fiM.
We had previously shown a similar differential sensitivity (10-
fold) to FdUrd, the deoxynucleoside analogue of FUra, in these2 cells lines (33). In that study, the cytotoxicity of FdUrd in bothcell lines could be directly correlated with the cellular level ofdTMP synthetase and was reversed by dThd, consistent withinhibition of thymidylate biosynthesis as the site of FdUrd action.As shown in Chart 1, while dThd can partially reverse growthinhibition by FUra in HT 29 cells, it is ineffective in reversing theeffects of this drug in HuTu 80 cells and actually causes a 2-fold
increase in the cytotoxicity of FUra. This suggests that, for FUra,
910 CANCER RESEARCH VOL. 44
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
Analysis ofFUra Metabolism in Gastrointestinal Cells
100
20
A. HT29
a HuTu80
(XOIFUra (¡tß
Chart 1. Effect of FUra on the growth of HT 29 and HuTu 80 cells, determinedin the absence (•.•)or presence p, O) of dThd, as described in "Materials andMethods." The results shown are those of a typical experiment.
the differential sensitivity of the 2 cell lines may involve differences in the cytotoxic mechanism of the drug.
FUra Accumulation and Activation. It was possible that thedifferential effect of FUra toward HT 29 and HuTu 80 cells wasa reflection of differences in uptake and/or activation of the drug.The time course of intracellular accumulation of FUra in the HuTu80 and HT 29 cell lines is shown in Chart 2. Over the first 2 hr,there is little difference between the level of drug accumulatedin the 2 cell lines. By 24 hr, however, the intracellular content ofFUra and its metabolites in HT 29 cells is 3 times higher than inHuTu 80 cells. Therefore, some difference in the ability to accumulate FUra exists between the 2 cell lines, although it wouldnot appear sufficient to account for the difference in sensitivityobserved.
The ability of each cell line to anabolize FUra, which is knownto require activation to the nucleotide level to be cytotoxic, wasalso examined. Table 1 contains an analysis of FUra metabolitesproduced following exposure of HuTu 80 and HT 29 cells to [6-3H]FUra (0.8 U.M)for 24 hr. The total radioactivity in HuTu 80
cells is much lower than in HT 29 cells, reflecting the differentialaccumulation described above. Both cell lines do, however,produce phosphorylated derivatives of the drug, including bothFUTP and FdUMP. These compounds are found both in the acid-
soluble fraction and associated with their macromolecular targets. In HuTu 80 cells, all the FdUMP produced appears associated with dTMP synthetase, a phenomenon we have observedbefore (35). Furthermore, the relative distribution of ribose (96 to97%) and deoxyribose (3 to 4%) derivatives in each cell line issimilar.
FUra Metabolism at Equitoxic Concentrations. The metab
olism of FUra in HuTu 80 and HT 29 cells was examined at theEC50concentration of the drug for each cell line (6 and 0.3 ^M,respectively) in an attempt to better understand the observeddifferential response of the cell lines to FUra. Table 2 containsan analysis of intracellular FUra metabolites following a 24-hrexposure of each cell line to equitoxic concentrations of [6-3H]-
FUra. In contrast to the situation when HuTu 80 and HT 29 cellsare exposed to equimolar concentrations of FUra, accumulationof FUra metabolites in HuTu 80 cells exposed to 6 UM [6-3H]-
FUra greatly exceeds that found in HT 29 cells exposed to anequitoxic dose of the drug (0.3 MM).HuTu 80 cells contain bothmore total FdUMP (bound and free) than do HT 29 cells and
to
— 5
16TIME (HR)
24
Chart 2. Intracellular accumulation of FUra. The time course of FUra accumulation in HT 29 (•)and HuTu 80 (O) cells was determined as described in "Materialsand Methods." The external [6-3H]FUra concentration was 0.5 »M.Bars. S.E.
Table 1Distribution of FUra metabolites in HuTu 80 and HT 29 cells following a 24-hr
exposure to 0.8 ¡IM[6-3H]FUra
HuTu 80 and HT 29 cells were exposed to [6-3H]FL)ra (0.8 utt; specific activity,
25 Ci/^mol) for 24 hr. At this time, FUra metabolism was analyzed as described in"Materials and Methods."
Distribution of FUra metabolites (fmol/105 cells)
Cell lineFree
FdUMP-TSc FURA-RNA FdUMP Other*
HT 29HuTu 80
45"
282143
64969
01094
19a Represents acid-soluble compounds other than FdUMP (FUTP, ftuorouridine
diphosphate-sugar, etc.)." Values represent the mean of at least duplicate experiments; the S.E. of the
mean was less than 10% of the mean value in all cases.c FdUMP-TS, FdUMP-dTMP synthetase.
MARCH 1984 911
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from
W. L Washtien
Table 2Distribution of FUra metabolites in HuTu 80 and HT 29 cells following a 24-hr
exposure to equitoxic concentrations of FUraHuTu 80 and HT 29 cells were each exposed to equitoxic concentrations of [6-
3H]FUra (6 and 0.3 JIM, respectively) for 24 hr. At this time, FUra metabolism wasanalyzed as described in "Materials and Methods."
Distribution of FUra metabolites (fmol/105cells)Cell
lineHT
29HuTu 80FdUMP-TSc41*157FURA-RNA10756585Free
FdUMP20
117Other8378 2307' Represents acid-soluble compounds other than FdUMP (FUTP, fluorouridine
diphosphate-sugar, etc.)." Values represent the mean of at least duplicate experiments; the S.E. of the
mean was less than 10% of the mean value in all cases.cFdUMP-rs, FdUMP-dTMP synthetase.
more FUra incorporated into cellular RNA.Thymidylate synthetase inhibition by FdUMP can play a role in
FUra cytotoxicity in either cell line only if all the enzyme in thecell is bound to FdUMP. We have previously demonstrated that,as FdUMP is generated within a cell, it becomes bound to dTMPsynthetase and, thus, may not be detected as free intracellularFdUMP until all the enzyme in the cell has been titrated (35). Thepresence of free FdUMP, therefore, might be considered asevidence that all the dTMP synthetase in the cell has beencomplexed to FdUMP. However, increasing levels of dUMP orinadequate folate pools can result in the coexistence of freeintracellular FdUMP and unbound enzyme (15, 24). Therefore,we determined whether any free dTMP synthetase could bedetected in either cell line following FUra exposure. In both HT29 and HuTu 80 cells, no free enzyme was observed, indicatingthat, in both cell lines, equitoxic concentrations of FUra result incomplete inhibition of dTMP synthetase.
In an earlier study, we determined the level of dTMP synthetase in both HT 29 (7 fmol/105 cells) and HuTu 80 cells (72 fmol/105 cells) and can therefore compare the amount of enzyme-
bound FdUMP measured following exposure to FUra with thelevels determined previously; since no free dTMP synthetase isobserved, one would expect the amounts to be equivalent. Theamount of FdUMP found bound to dTMP synthetase in each cellline (HT 29, 41 fmol/105 cells; HuTu 80, 157 fmol/105 cells), in
fact, exceeds the value of enzyme determined in untreated cells(see Table 2). We have observed this phenomenon in a numberof cell lines following exposure to FUra and have found it to bedue to an increased in vivo stability of dTMP synthetase complexed to FdUMP as compared to uncomplexed enzyme (34).
The mechanism by which FUra incorporation into RNA produces cytotoxicity is unclear and, thus, one does not, as withdTMP synthetase, have a defined cytotoxic endpoint which mustbe achieved. Studies within a cell line have demonstrated acorrelation between the level of drug incorporation into RNA anddrug cytotoxicity and would suggest that FUra cytotoxicity mediated through RNA effects is more likely as higher levels of drugincorporation into RNA are produced (3, 4). Equitoxic concentrations of FUra result in a 6-fold higher level of incorporation of
FUra into the RNA of HuTu 80 cells as compared with HT 29cells and, along with the inability of dThd to reverse the effectsof FUra in HuTu 80 cells (Chart 1), suggest the possible importance in HuTu 80 cells of RNA effects of FUra.
Effect of Deoxyinosine on Growth Inhibition and FUra Metabolism. In most cells, FUra is metabolized mainly throughpathways involving initial conversion of FUra to ribose derivatives(23). However, it can be converted directly to FdUrd by the action
of dThd phosphorylase. This pathway appears limited in vivo bythe availability of deoxyribose donors, and deoxyinosine hasbeen used to boost intracellular deoxyribose levels (9, 12).
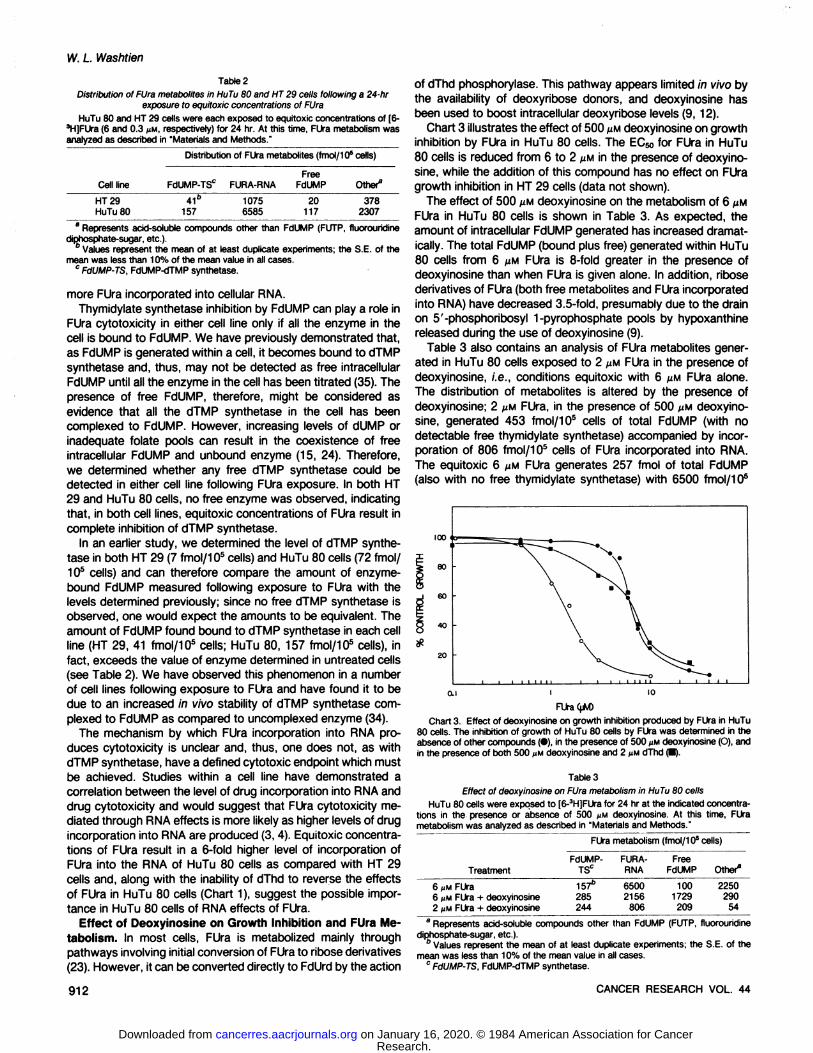
Chart 3 illustrates the effect of 500 ^M deoxyinosine on growthinhibition by FUra in HuTu 80 cells. The EC50for FUra in HuTu80 cells is reduced from 6 to 2 U.Min the presence of deoxyinosine, while the addition of this compound has no effect on FUragrowth inhibition in HT 29 cells (data not shown).
The effect of 500 UMdeoxyinosine on the metabolism of 6 U.MFUra in HuTu 80 cells is shown in Table 3. As expected, theamount of intracellular FdUMP generated has increased dramatically. The total FdUMP (bound plus free) generated within HuTu80 cells from 6 U.MFUra is 8-fold greater in the presence of
deoxyinosine than when FUra is given alone. In addition, ribosederivatives of FUra (both free metabolites and FUra incorporatedinto RNA) have decreased 3.5-fold, presumably due to the drainon 5'-phosphoribosyl 1-pyrophosphate pools by hypoxanthine
released during the use of deoxyinosine (9).Table 3 also contains an analysis of FUra metabolites gener
ated in HuTu 80 cells exposed to 2 U.MFUra in the presence ofdeoxyinosine, i.e., conditions equitoxic with 6 U.MFUra alone.The distribution of metabolites is altered by the presence ofdeoxyinosine; 2 ^M FUra, in the presence of 500 UM deoxyinosine, generated 453 fmol/105 cells of total FdUMP (with no
detectable free thymidylate synthetase) accompanied by incorporation of 806 fmol/105 cells of FUra incorporated into RNA.
The equitoxic 6 MM FUra generates 257 fmol of total FdUMP(also with no free thymidylate synthetase) with 6500 fmol/105
O.I I 10
FUra<t*D
Chart 3. Effect of deoxyinosine on growth inhibition produced by FUra in HuTu80 cells. The inhibition of growth of HuTu 80 cells by FUra was determined in theabsence of other compounds (•),in the presence of 500 UMdeoxyinosine (O), andin the presence of both 500 JIMdeoxyinosine and 2 ¿iMdThd (•).
Table 3
Effect of deoxyinosine on FUra metabolism in HuTu 80 cellsHuTu 80 cells were exposed to [6-3H]FUra for 24 hr at the indicated concentra
tions in the presence or absence of 500 ^M deoxyinosine. At this time, FUrametabolism was analyzed as described in "Materials and Methods."
FUra metabolism (fmol/10scells)Treatment6
jiM FUra6 MMFUra + deoxyinosine2 /<MFUra + deoxyinosineFdUMP-
TSC157"
285244FURA-
RNA6500
2156806Free
FdUMP100
1729209Other*2250
29054
a Represents acid-soluble compounds other than FdUMP (FUTP, fluorouridine
diphosphate-sugar, etc.).6 Values represent the mean of at least duplicate experiments; the S.E. of the
mean was less than 10% of the mean value in all cases.c FdUMP-TS, FdUMP-dTMP synthetase.
912 CANCER RESEARCH VOL. 44
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

Analysis of FUra Metabolism in Gastrointestinal Cells
cells of FUra ¡ncoporatedinto RNA.The increased cytotoxicity of FUra in the presence of deoxyi-
nosine, accompanied as it is by a decrease in drug incorporationinto RNA, suggested a change in the cytotoxic mechanism ofFUra in HuTu 80 cells in the presence of deoxyinosine. To furthertest this hypothesis, we examined the ability of dThd to reversethe growth inhibition produced by FUra in the presence ofdeoxyinosine. As shown in Chart 3, dThd is able to reversegrowth inhibition produced by the combination of FUra anddeoxyinosine, in contrast to the results described previously forFUra alone (Chart 1).
In HT 29 cells, in which the combination of deoxyinosine andFUra is no more growth inhibitory than FUra alone, deoxyinosinehas a similar effect on metabolism of FUra via ribose derivatives.Surprisingly, however, the amount of FdUMP (free plus bound)is not significantly different in the presence of this deoxyribosedonor (17 versus 20 fm). As with FUra alone, there is no detectable free dTMP synthetase in the HT 29 cells treated with FUraand deoxyinosine.
DISCUSSION
We have examined the metabolism of FUra in 2 human gastrointestinal tumor cell lines, HuTu 80 and HT 29, which differ20-fold in their sensitivity to growth inhibition by FUra. We haveanalyzed [6-3H]FUra metabolism within these cell lines using
methods which permit simultaneous determination of: (a) theinteraction of FdUMP and dTMP synthetase; (b) the presence offree dTMP synthetase; (c) the extent of FUra incorporation intoRNA; and (d) the distribution of acid-soluble metabolites of the
drug.Following exposure to equitoxic concentrations of FUra, both
HuTu 80 and HT 29 cells generate sufficient FdUMP to bind allthe dTMP synthetase within the cells, as evidenced by the lackof detectable free dTMP synthetase. This would suggest that,for both cell lines, conditions are achieved after 24 hr of drugexposure which would allow dTMP synthetase inhibition to begrowth limiting. Variability in responsiveness to FUra amongsome cells has been correlated with differences in the rate ofintracellular clearance of FdUMP (1, 17, 24). However, becausethe conditions used in the current study involve continuousexposure of cells to drug, the levels of FdUMP achieved followingexposure to FUra for 24 hr should be maintained and shouldrepresent the level achieved during determinations of growthinhibition.
In HT 29 cells, dThd can reverse the effects of FUra, as wouldbe expected if dTMP synthetase inhibition is growth limiting (8,21). The inability of dThd to reverse FUra-mediated growthinhibition in HuTu 80 cells, however, argues against dTMP synthetase inhibition as the dominant site of action for FUra in thesecells and suggests that, for HuTu 80 cells, growth inhibition byFUra is related to RNA-directed effects of the drug. Comparisonof the levels of FUra present in the RNA of each cell line followingdrug exposure illustrates that HuTu 80 cells contain 6 times asmuch FUra incorporated into their RNA as do HT 29 cells at anequitoxic dose of drug, and provides a possible explanation forthe dominance of RNA-directed cytotoxicity in HuTu 80 but not
HT 29 cells.HuTu 80 cells are less efficient than HT 29 cells in accumulating
FUra. In addition, more FdUMP is required to produce dTMPsynthetase inhibition in these cells, since they contain 10 times
as much enzyme as do HT 29 cells (72 versus 7 fm/1 05 cells)
(33). These 2 factors result in the necessity for high FUraconcentrations to achieve dTMP synthetase inhibition in HuTu80 cells. FUra metabolism through the ribose pathway generatesboth FdUMP and FUTP (23). In HuTu 80 cells, drug concentrations which produce sufficient FdUMP for dTMP synthetaseinhibition also appear to result in levels of FUTP incorporationinto RNA which are cytotoxic; hence, the inability of dThd toreverse FUra cytotoxicity in these cells. The observed enhancement of FUra cytotoxicity in HuTu 80 cells by dThd is puzzling,since we have found that the uptake and disposition of FUrametabolites in the presence of dThd4 is essentially the same as
that seen with FUra alone. HT 29 cells, more efficient in drugaccumulation and containing lower dTMP synthetase levels,achieve adequate FdUMP levels following exposure to muchlower concentrations of FUra. However, even in these cells,exposure to higher concentrations of FUra (3 MM) producesgrowth inhibition which is refractory to dThd and which resultsin increased (10-fold) levels of FUra incorporation into RNA.4
As mentioned, HuTu 80 cells are less efficient in FUra accumulation than are HT 29 cells. One approach to increasing theefficiency of FUra use in such cells is to combine FUra withagents such as methotrexate or A/-(phosphonacetyl)-i_-asparticacid. A/-(phosphonacetyl)-L-aspartic acid reduces the endogenous uridine pools which normally compete with FUra for cosub-
strates and enzymes (19), while the inhibition of purine synthesisby methotrexate leads to an increased availability of ¿'-phos-
phoribosyl 1-pyrophosphate for FUra anabolism (3). Such com
pounds, while potentiating FUra cytotoxicity, are unlikely to alterits mechanism of action within a cell. Instead, they produce thesame internal profile of FUra metabolism, but at lower externalFUra concentrations. Deoxyinosine, by providing a source ofdeoxyribose within the cell, affords the possibility of changingthe predominant route of FUra metabolism from a ribose todeoxyribose pathway (9). In such a situation, adequate FdUMPlevels might be generated under conditions which do not lead toa parallel production of FUTP.
In HuTu 80 cells, the presence of 500 MMdeoxyinosine potentiates growth inhibition by FUra 3-fold. While this decrease inEC50 is accompanied by a 2-fold increase in the amount of
FdUMP (free plus bound) generated from equitoxic concentrations of FUra, exposure to either 6 MMFUra or 2 MMFUra in thepresence of deoxyinosine results in complete binding of cellulardTMP synthetase. More significant, perhaps, is the 8-fold de
crease in the level of FUra incorporated into RNA when HuTu 80cells are exposed to 2 MMFUra in the presence of deoxyinosineas compared to the levels achieved with 6 MMFUra alone. In thepresence of deoxyinosine, HuTu 80 cells are able to achieveFdUMP levels adequate to produce dTMP synthetase inhibitionin the absence of cytotoxic levels of drug incorporation into RNA,and the dominant mechanism of FUra action is changed. Thischange in site of action is illustrated both by the ability of dThdto reverse the effects of the FUra-deoxyinosine combination and
by the fact that growth inhibition produced by this combinationcan be further potentiated by folinic acid, which is known toaffect the interaction of FdUMP with dTMP synthetase (9, 15).Folinic acid has no effect on growth inhibition in HuTu 80 cellsproduced by FUra alone.4
In HT 29 cells, deoxyinosine does not potentiate growth
4W. L. Washtien, unpublished observations.
MARCH 1984 913
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

IV. L. Washtien
inhibition by FUra. The level of total FdUMP generated from FUrain the presence of deoxyinosine is not significantly different fromthat achieved with FUra alone, while the level of FUra incorporation into RNA is reduced. These data are consistent withinhibition of dTMP synthetase as the primary site of action forFUra in HT 29 cells in the presence or absence of deoxyinosine.The level of FUra incorporation into RNA when FUra is usedalone is not growth-inhibitory (until concentrations higher than
the EC50 are used) and is reduced further in the presence ofdeoxyinosine. It is somewhat surprising that the presence of adeoxyribose donor does not increase FdUMP levels in this cellline. It could be that deoxyribose levels are already sufficient inHT 29 cells to allow maximal utilization of this pathway or thatthe results reflect a serendipitous equality in the level of FdUMPgenerated from FUra in the absence and presence of deoxyinosine.
The conditions used in this study for measurement of growthinhibition involve continuous exposure of cells to FUra and, thus,resemble the conditions achieved during infusion therapy. Theresults suggest that, under such conditions, the use of deoxyinosine to direct FUra action toward dTMP synthetase mightincrease the toxicity of FUra toward cells such as HuTu 80,which can normally only achieve this inhibition in concert withRNA-directed cytotoxicity. Whether such a combination would
provide an improved therapeutic index is unknown and woulddepend on FUra metabolism in normal tissues.
The results in this study demonstrate the utility of an approachwhich allows simultaneous measurement of 2 major cytotoxicendpoints of FUra metabolism in defining the dominant mechanism of FUra action within a cell line and in evaluating the effectof other agents on FUra metabolism and mechanism. We feelthat such an approach will be useful in evaluating the interactionof FUra with such agents as methotrexate, which has beenproposed to affect FUra cytotoxicity at both of those sites ofaction (3,10), and in further delineating the relationship betweenFUra metabolism and cell sensitivity to this drug. Our studieshave not addressed the possible role of FUra incorporation intoDNA, but it should not be difficult to expand the methodologyused to also examine this possibility.
ACKNOWLEDGMENTS
The author wishes to thank Veronica Janik for her excellent technical assistance.
REFERENCES
1. Ardalan, B., Cooney, D. A., Jayaram, H. N.. Carneo, C. K., Glazer, R. l.,MacDonald, J., and Schein, P. S. Mechanisms of sensitivity and resistance ofmurine tumors to 5-fluorouracil. Cancer Res., 40: 1431-1437,1980.
2. Blakely, R. L. Crystalline dihydropteroylglutamic acid. Nature (Lond.). 188:231-232, 1960.
3. Cadman, E., Heimer, R., and Benz, C. The influence of methotrexate pretreatment on 5-fluorouracil metabolism in L1210 cells. J. Biol. Chem., 256: 1695-1704, 1981.
4. Cadman, E., Heimer, R., and Davis, L. Enhanced 5-fluorouracil nucleotideformation after methotrexate administration: explanation for drug synergism.Science (Wash. D. C.), 205: 1135-1137,1979.
5. Chen, T. R. In situ detection of Mycoplasma contamination in cell cultures byfluorescent Hoechst 33258 stain. Exp. Cell Res., 704: 255-262, 1977.
6. Cheng, Y-C., and Nakayama, K. Effects of 5-fluoro-2'-deoxyuridine on DNA
metabolism in HeLa cells. Mol. Pharmacol., 23. 171-174,1983.
7. Danenberg, P. V., Langenbach, R. J., and Heidelberger, C. Structures ofreversible and irreversible complexes of thymidylate synthetase and fluorinatedpyrimidine nucleotides. Biochemistry, 13: 926-933,1974.
8. Evans, R. M., Laskin, J. D., and Hakala, M. T. Assessment of growth-limitingevents caused by 5-fluorouracil in mouse cells and in human cells. CancerRes., 40.: 4113-4122,1980.
9. Evans, R. M., Laskin, J. D., and Hakala, M. T. Effect of excess folates anddeoxyinosine on the activity and site of action of 5-fluorouracil. Cancer Res.,47:3288-3295, 1981.
10. Femandes, D. J., and Berlino, J. R. 5-Fluorouracil-methotrexate synergy:enhancement of 5-fluorodeoxyuridylate binding to thymidylate synthetase bydihydropteroyl-polyglutamates. Proc. Nati. Acad. Sci. U. S. A., 77: 5663-5667,1980.
11. Fogh, J., and Trempe, G. New human tumor cell lines. In: J. Fogh (ed.), HumanCells in Vitro, pp. 115-159. New York: Plenum Press, 1975.
12. Gotto, A. M., Belkhode, M. L., and Touster, 0. Stimulatory effects of inosineand deoxyinosine on the incorporation of uraal-2-14C, 5-fluorouracil-2-'4C, and5-bromouracil-2-"'C into nucleic acids by Ehrlich ascites tumor cells in vitro.
Cancer Res., 29: 807-811, 1969.13. Hatefi, Y., Talbert, P. T., Osbom, M. T., and Huennekens, F. M. Tetrahydrofouc
acid. Biochem. Prep., 7: 89-92, 1960.14. Heidelberger, C., Danenberg, P. V., and Moran, R. G. Fluorinated pyrimidines
and their nucteosides, Adv. Enzymol. 54: 58-119,1983.15. Houghton, J. A., Maroda, S. J., Philips, J. 0., and Houghton, P. J. Biochemical
determinants of responsiveness to 5-fluorouracil and its derivatives in xeno-grafts of human colorectal adenocarcinomas in mice. Cancer Res., 47: 144-149, 1981.
16. Khym, J. X. An analytical system for rapid separation of tissue nucleotides atlow pressure on conventional ion exchangers. Clin. Chem., 27: 1245-1252,1975.
17. «lubes,P., Connelly, K., Cerna, I., and Mandel, H. G. Effects of 5-fluorouracilon 5-fluorodeoxyuridine-5'-monophosphate and 2-deoxyuridine-5'-monophos-
phate pools, and DNA synthesis in solid mouse L1210 and rat Walker 256tumors. Cancer Res., 36: 2325-2331, 1978.
18. Laskin, J. D., Evans, R. M., Slocum, H. K., Burke, D., and Hakala, M. T. Basisfor natural variation in sensitivity to 5-fluorouracil in mouse and human cells inculture. Cancer Res., 39: 383-390,1979.
19. Liang, C-M., Donehower, R. C., and Chabner, B. A. Biochemical interactionsbetween N-<phosphonacetyl)-L-aspartate and 5-fluorouracil. Mol. Pharmacol.,27:224-230, 1982.
20. Major, P. P., Egan, E., Herrick, D., and Kufe, D. W. 5-Fluorouracil incorporationinto DNA of human breast carcinoma cells. Cancer Res., 42: 3005-3009,1982.
21. Maybaum. J., Ullman. B., Mandel, H. G . Day, J. L., and Sadee, W. Regulationof RNA- and DNA-directed actions of 5-fluoropyrimidines in mouse T-lymphoma(S-49) cells. Cancer Res., 40: 4209-4215, 1980.
22. Moran, R. G., Spears, C. P., and Heidelberger, C. Biochemical determinantsof tumor sensitivity to 5-fluorouracil: ultrasensitive methods for the determination of 5-fluoro-2'-deoxyuridylate, 2'-deoxyuridylate, and thymidylate syn
thetase, Proc. Nati. Acad. Sei. U. S. A., 76: 1456-1460,1979.23. Myers, C. E. The pharmacology of the fluoropyrimidines. Pharmacol. Rev., 33:
1-15, 1981.24. Myers, C. E., Young, R. C., and Chabner, B. A. Biochemical determinants of
5-fluorouradl response in vivo. J. Clin. Invest., 56: 1231-1238, 1975.25. Nahas, A., Savlov, E. D., and Hall, T. C. Phosphoribosyltransferase in colon
tumor and normal mucosa as an aid to adjuvant chemotherapy with 5-fluorouracil. Cancer Chemother. Rep., 58: 909-912, 1974.
26. Pogolotti, A. L., Nolan, P. A., and Santi, D. V. Methods for complete analysisof 5-fluorouracil metabolites in cell extracts. Anal. Biochem., 777: 178-186,1981.
27. Reyes, P., and Hall, T. C. Synthesis of 5-fluorouridine-5'-phosphate by a
pyrimidine phosphoribosyltransferase of mammalian origin. II. Correlation between tumor levels of the enzyme and the 5-fluorouracil-promoted increase insurvival of tumor bearing mice. Biochem. Pharmacol., 78: 2587-2590,1969.
28. Santi, D. V., McHenry, C. S., and Sommer, H. Mechanism of interaction ofthymidylate synthetase with 5-fluorodeoxyuridine. Biochemistry, 73:471 -481,
1974.29. Schilsky, R. L., Bailey, B. D., and Chabner, B. A. Characteristics of membrane
transport of methotrexate by cultured human breast cancer cells. Biochem.Pharmacol., 30: 1537-1542,1981.
30. Schmidt, M., and Good, R. A. Transplantation of human cancers to nude miceand effects on thymus growth. J. Nati. Cancer Inst, 55: 81-84,1975.
31. Spears, C. P., Shahinian, A. H., Moran, R. G., Heidelberger, C., and Corbe«,T. H. In vivo kinetics of thymidylate synthetase inhibition in 5-fluorouracil-sensitive and -résistantmurine colon adenocarcinomas. Cancer Res., 42:450-
456, 1982.32. Trakatellis, A. C., and Axelrod, A. E. Effect of pyridoxine deficiency on nucleic
acid metabolism in the rat. Biochem. J., 95. 344-349, 1965.33. Washtien, W. L. Thymidylate synthetase levels as a factor in 5-fluoro-deox-
yuridine and methotrexate cytotoxicity in gastrointestinal tumor cells. Mol.Pharmacol., 27: 723-728, 1982.
34. Washtien, W. L.. Increased levels of thymidylate synthetase in cells exposedto 5-fluorouracil. Mol. Pharmacol., in press, 1984.
35. Washtien, W. L., and Santi, D. V. Assay of intracellular free and macromolec-ular-bound metabolites of 5-fluorodeoxyuridine and 5-fluorouracil. Cancer Res.,39:3397-3404, 1979.
36. Wilkinson, D. S., and Pilot, H. C. Inhibition of ribosomal ribonucleic acidmaturation in Novikoff hepatoma cells by 5-fluorouracil and 5-fluorouridine, J.Bid. Chem., 248: 63-68, 1973.
914 CANCER RESEARCH VOL. 44
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from

1984;44:909-914. Cancer Res Wendy L. Washtien Gastrointestinal Tumor Cell LinesComparison of 5-Fluorouracil Metabolism in Two Human
Updated version
http://cancerres.aacrjournals.org/content/44/3/909
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/44/3/909To request permission to re-use all or part of this article, use this link
Research. on January 16, 2020. © 1984 American Association for Cancercancerres.aacrjournals.org Downloaded from





























