Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM GENÉTICA, CONSERVAÇÃO E BIOLOGIA
EVOLUTIVA
HEIDI LUZ BONIFÁCIO
MANAUS-AM
2011
CITOGENÉTICA CLÁSSICA E MOLECULAR DO BOTO-VERMELHO
Inia geoffrensis (Blainville, 1817)

HEIDI LUZ BONIFÁCIO
ORIENTADORA: Dra. Eliana Feldberg
COORIENTADORA: Dra. Vera Maria Ferreira da Silva
Dissertação apresentada ao Programa de Pós-
Graduação em Genética, Conservação e Biologia
Evolutiva como parte dos requisitos para obtenção do
título de Mestre em Genética, Conservação e Biologia
Evolutiva.
MANAUS-AM
2011
CITOGENÉTICA CLÁSSICA E MOLECULAR DO BOTO-VERMELHO
Inia geoffrensis (Blainville, 1817)

FICHA CATALOGRÁFICA
iii
B715 Bonifácio, Heidi Luz Citogenética clássica e molecular do boto-vermelho Inia geoffrensis (Blainville, 1817) / Bonifácio Heidi Luz. --- Manaus : [s.n.], 2011. xvii, 57 f. : il. color.
Dissertação (mestrado) -- INPA, Manaus, 2011 Orientador : Eliana Feldberg Co-orientador : Vera Maria Ferreira da Silva Área de concentração : Genética, Conservação e Biologia Evolutiva
1. Boto (mamífero aquático). 2. Citogenética molecular. 3. FISH (técnica). 4. Golfinho da Amazônia. 5. Cariótipo. I. Título. CDD 19. ed. 599.530415
Sinopse
Uma ampla caracterização cromossômica foi realizada em uma das espécies de golfinhos da
Amazônia: o boto-vermelho Inia geoffrensis, por meio de análises de citogenética clássica
(coloração convencional, detecção da heterocromatina, das regiões organizadoras do nucléolo e
bandeamento G) e citogenética molecular (hibridização fluorescente in situ com sondas de
DNAr 5S, DNAr 18S, sequência telomérica e retrotransposon LINE-L1). A heterocromatina no
cariótipo do boto-vermelho localiza-se na região terminal e intersticial, com a presença de
polimorfismo e a RON é simples. Homeologias cromossômicas foram estabelecidas entre o
boto-vermelho e o golfinho Tursiops truncatus e pela primeira vez as sequências do
retrotransposon L1 e do DNAr 5S foram mapeadas no genoma de cetáceos.
Palavras-chave: Golfinho da Amazônia, cariótipo, bandeamentos cromossômicos,
polimorfismo banda C, FISH, DNAr, retrotransposon LINE-L1

Dedico aos meus pais e à minha avó que sempre me apoiaram durante toda minha vida acadêmica e me fizeram persistir na busca pelos meus sonhos.
iv

“Nada é fixo para aquele que alternadamente pensa e sonha...”
Gaston Bachelard
v

A realização deste projeto foi possível devido:
Ao Programa de Pós-Graduação em Genética, Conservação e Biologia Evolutiva do INPA.
Ao Laboratório de Genética Animal do INPA, Coordenação de Pesquisas em Biologia
Aquática (CPBA), onde grande parte da atividade laboratorial foi realizada, com o
financiamento do Projeto Caracterização Genética (cromossomos, proteínas e DNA) de
vertebrados amazônicos (INPA/MCT - PRJ 12.01) e Projeto Centro de Estudos de
Adaptações da Biota Aquática da Amazônia - ADAPTA (INCT ADAPTA, FAPEAM/CNPq
Proc. Nº 573976/2008-2)
Ao Laboratório de Mamíferos Aquáticos do INPA em conjunto com a AMPA, pelo apoio à
realização das coletas e a participação em eventos científicos, com o financiamento da
Petrobras por meio do Programa Petrobras Ambiental (PRJ 12.86) e Projetos de Pesquisas
Institucionais (INPA/PPI PRJ 12.16).
Ao Projeto Boto através de cooperação entre o Instituto Nacional de Pesquisas da Amazônia
INPA/MCT e a Reserva de Desenvolvimento Sustentável Mamirauá – MSDR- OS/MCT.
À Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela concessão
da bolsa durante a realização deste trabalho.
vi

Agradecimentos
Agradeço a Deus pelo dom da vida e pela oportunidade de aprendizado.
À minha família pelas orações, pelo amor incondicional, pelo apoio e torcida durante
toda a minha jornada acadêmica.
À minha mãezinha, Rosilene pelo exemplo de fortaleza e por não me deixar desistir dos
meus sonhos.
Ao meu paizinho, Paulo por toda dedicação e por me dar ânimo e força para suportar a
saudade e a distância.
Ao meu irmão Gúbio, por ser forte quando necessário e por segurar a barra enquanto
estive longe.
À minha vó Joana pelo seu carinho e apoio desde sempre.
À grande equipe do Laboratório de Genética Animal-INPA: Eliana, Jorge, Carlos,
Claudia, Leandra, Leila, Valentim, Cacá, Eduardo, Natália, Thatiana, Brenda, Érica, Denise,
Paola, Heidy Alana, Eliane, Masseo e Tássia agradeço pelo chá de casa nova, pelas festinhas,
pela convivência pacífica no laboratório e por toda ajuda.
Ao Carlos Schneider por pacientemente me orientar em toda parte de citogenética
molecular, por fazer questões pertinentes e me ajudar a solucionar vários perrengues durante
este trabalho.
À Claudia Gross pelos sábios conselhos e por sempre estar disposta a ajudar e sanar
dúvidas.
À Leandra Terencio por toda ajuda, dicas e sugestões no sequenciamento do 5S do boto.
Polacos a ajuda de vocês foi imprescindível para execução deste trabalho.
Ao Dr. Jorge Porto pelos conselhos, dicas e por prontamente assinar vários documentos
na ausência das minhas orientadoras.
À Érica e ao Cacá pelas dicas e sugestões e todos os nossos colóquios sobre a banda G.
Meu agradecimento muito especial à minha orientadora Dra. Eliana Feldberg, por me
acolher de braços abertos com seu jeito maternal, por entender mesmo a contra gosto minha
opção por mamíferos, por salvar meu Natal em Manaus, por pacientemente e rapidamente ler
e discutir todos meus documentos do mestrado. Por acreditar e apoiar todas as minhas idéias
mirabolantes e pelo carinho, conselhos, questionamentos e ensinamentos.
vii

À Natália Carvalho, por ser minha guia em Manaus e companheira de horas e horas no
laboratório, de visitas ao centro da cidade, de idas ao supermercado e de entrega de currículos.
Obrigada pela sua amizade e apoio nesta trajetória.
À Thatiana de Lira minha caloura conterrânea, sua presença e da D. Joana me fez sentir
mais perto de casa. Obrigada pela amizade, pela comidinha de mãe, pelas horas de divagações
sobre a vida na porta de casa, pela companhia certa para o cinema e por prontamente me
ajudar inúmeras vezes.
À Jaqueliny Zocca por toda sua amizade, conselhos e apoio durante estes dois anos. Por
pacientemente me escutar nas minhas horas de estresse e de empolgação! Pelo abrigo em sua
casa, pelas nossas horas de debates sobre evolução. Por ser companheira de almoços, festas e
passeios e por me arrastar pra academia.
À minha companheira de casa: Eurizângela obrigada pela paciência nos meus
momentos de estresse e por toda ajuda. Os amigos são a família que podemos escolher.
Meninas vocês tornaram minha vida aqui mais feliz.
À equipe do Laboratório de Mamíferos Aquáticos do INPA. Em especial a todos
envolvidos na Captura de Botos de 2009 e também na coleta e transporte de material de outras
localidades: Vera, Tony, Nívia, Diogo, Deise, Glória, Virginia, Waleska, Rodrigo, Nicole,
Nildon, Louzinha e a todos os pescadores Soldado, seu João Pena, seu Ney por todos seus
ensinamentos sobre a arte de capturar botos.
À Nívia, Diogo, Deise por toda ajuda no processamento do material em Mamirauá,
mesmo achando esta parte de pipetar muita chata! Valeu pela ajuda.
À minha coorientadora Dra. Vera da Silva, por toda orientação e apoio durante a coleta
em Mamirauá e em outras localidades. Pela oportunidade de trabalho e aprendizado sobre este
animal fantástico, boto-vermelho, por todos os ensinamentos, conselhos e constante
questionamento.
Aos meus orientadores da graduação Dr. Ricardo Gunski e Dra. Anália del Valle, por
todos os ensinamentos e conselhos e por me instigarem o gosto pela vida cientifica.
Às minhas amigas desde sempre: Paula Cristina, Aline, Liana e Geanny, obrigada pelos
conselhos, pelas horas ao telefone e no Messenger pelas festinhas quando ia para casa e por
escutarem anos a fio sobre o meu mestrado.
À minha turma de mestrado, Natália, Jaque Zocca, Carol, Alessandro, Diana, Luciana,
Jaque Fortuna, Tatiana Bacry, Ramon e Gilson pelos debates científicos ou não durante as
viii

disciplinas e festinhas, e em especial aos que toparam a empreitada do curso de verão da
Genética.
Ao coordenador do PPG-GCBev Dr. Jorge Porto, e às secretárias GCBev, Alessandra,
Elci e Tamara, obrigada pela ajuda e pela resolução rápida de toda a parte burocrática durante
o mestrado e pelo apoio ao Curso de Verão.
A todos que tornaram este trabalho possível e me apoiaram durante esta jornada meu
muito obrigada.
ix

Resumo
O golfinho da Amazônia ou boto-vermelho (Iniidae, Inia geoffrensis) é um golfinho fluvial
endêmico das bacias Amazônica e do Orinoco. Para compreender a organização genômica
desta espécie e gerar marcadores para investigar as relações evolutivas deste golfinho em
relação a outros cetáceos, foram realizadas análises de citogenética clássica e molecular
(hibridização fluorescente in situ com sondas de DNAr 5S, DNAr 18S, sequência telomérica e
retrotransposon LINE-L1). Foram analisados 24 espécimes (11♀ e 13♂) que apresentaram
2n=44 cromossomos (12m+14sm+6st+10t+XX/XY) e NF= 72. A região organizadora do
nucléolo está localizada em único sítio, no par 21t. A heterocromatina distribuiu-se
terminalmente nos pares metacêntricos, submetacêntricos e subtelocêntricos e na região
pericentromérica dos telocêntricos. Blocos intersticiais foram observados em alguns pares
submetacêntricos e subtelocêntricos, com a presença de polimorfismo no par 9sm.
Homeologias cromossômicas foram estabelecidas através do padrão de banda G entre o boto-
vermelho e o golfinho-nariz-de-garrafa-comum Tursiops truncatus. Sequências teloméricas
intersticiais não foram observadas. Localização do retrotransposon L1 não foi aleatória no
cariótipo do boto-vermelho. Nos autossomos o L1 esteve preferencialmente localizado em
regiões de banda G positiva e o maior acúmulo desta sequência esteve no cromossomo X. O
DNAr 5S e L1 estão colocalizados na região centromérica do par 5m. Além disso, é possível
que o retrotransposon L1 esteja envolvido no polimorfismo de banda C encontrado para
quatro indivíduos da Reserva de Desenvolvimento Sustentável Mamirauá, Amazonas-Brasil.
Embora a estrutura cariotípica seja considerada conservada em cetáceos, características
cariotípicas do boto-vermelho mostram que variações existem em relação à fórmula
cariotípica, distribuição da heterocromatina e do organizador nucleolar comparativamente a
outras espécies de cetáceos.
x

Abstract
Amazon river dolphin or boto or (Iniidae, Inia geoffrensis) is a fluvial dolphin endemic to the
Amazon and Orinoco basins. In order to understand the genomic organization of this species
and to produce markers to investigate the evolutional relationship of this dolphin with others
cetaceans, classical and molecular cytogenetic (fluorescence in situ hybridization with probes
for 5S rDNA, 18S rDNA and telomeric sequence retrotransposon LINE-L1) studies were
carried out. Twenty four specimens were analyzed (11♀ e 13♂) which presented 2n=44
chromosomes (12m+14sm+6st+10t) e FN= 76. The nucleolar organizer region was situated at
single site on pair 21t. The heterochromatin was terminally distributed on metacentric,
submetacentric and subtelocentric pairs and on the pericentromeric region of telocentric.
Interstitial blocks were observed in some submetacentric pairs and subtelocentric as well,
with presence of polymorphism on pair 9sm. Chromosomal homeologies has been settled
throughout the G-band pattern between the boto and Atlantic bottlenose dolphin Tursiops
truncatus. Interstitial telomeric sequences were not observed. Location of retrotransposon L1
was not random on the boto karyotype. At the autosomes L1 had been preferably situated on
G-positive-band regions, and the largest accumulation of this sequence was on chromosome
X. The 5S rDNA and L1 are colocalized in the centromeric region of pair 5m. Beyond this, it
is possible that the L1 retrotransposon could be involved on band C polymorphism found in
four dolphins at Mamiraua Sustainable Development Reserve in Amazonas-Brazil. Although
karyotype structure is being considered as conserved in cetaceans, karyotype characteristics of
the boto indicate that variations may exist in relation to karyotype formula, heterochromatin
distribution and nucleolar organizer compared to other cetacean species.
xi

Sumário1. Introdução..........................................................................................................................................1
1.1 Considerações sobre o boto-vermelho.........................................................................................1
1.2 Estudos Citogenéticos em Cetáceos.............................................................................................4
1.3 Objetivos.....................................................................................................................................12
2. Material e métodos..........................................................................................................................13
2.1 Material e área de estudo...........................................................................................................13
2.2.1 Coleta do Material...................................................................................................................14
2.2.2 Obtenção dos cromossomos mitóticos....................................................................................15
2.2.3 Detecção das regiões de heterocromatina constitutiva: Banda C (CBG) .................................16
2.2.4 Bandeamento G (GTG).............................................................................................................16
2.2.5 Detecção das regiões organizadoras do nucléolo (Ag-RON)....................................................17
2.2.6 Extração de DNA......................................................................................................................17
2.2.7 Obtenção das sondas via PCR (Polymerase Chain Reaction)...................................................18
2.2.8 Marcação das sondas...............................................................................................................19
2.2.9 Hibridização in situ fluorescente (FISH) ..................................................................................20
2.2.10 Análise cromossômica...........................................................................................................21
2.2.11 Caracterização da sondas de DNAr 5S ..................................................................................22
2.2.11.1 Construção de vetores plasmidiais recombinantes contendo fragmentos de DNAr 5S obtidos por PCR ...............................................................................................................................22
2.2.11.2 Transformação ...................................................................................................................22
2.2.11.3 Sequenciamento das sondas de DNA .................................................................................23
3. Resultados........................................................................................................................................25
4. Discussão..........................................................................................................................................34
4.1- O boto-vermelho dentro da carioevolução dos cetáceos..........................................................34
4.2 Padrão de Distribuição das Sequências de DNA repetitivo ........................................................36
5. Conclusões........................................................................................................................................42
7. Referências Bibliográficas.................................................................................................................44
xii

Lista de Figuras
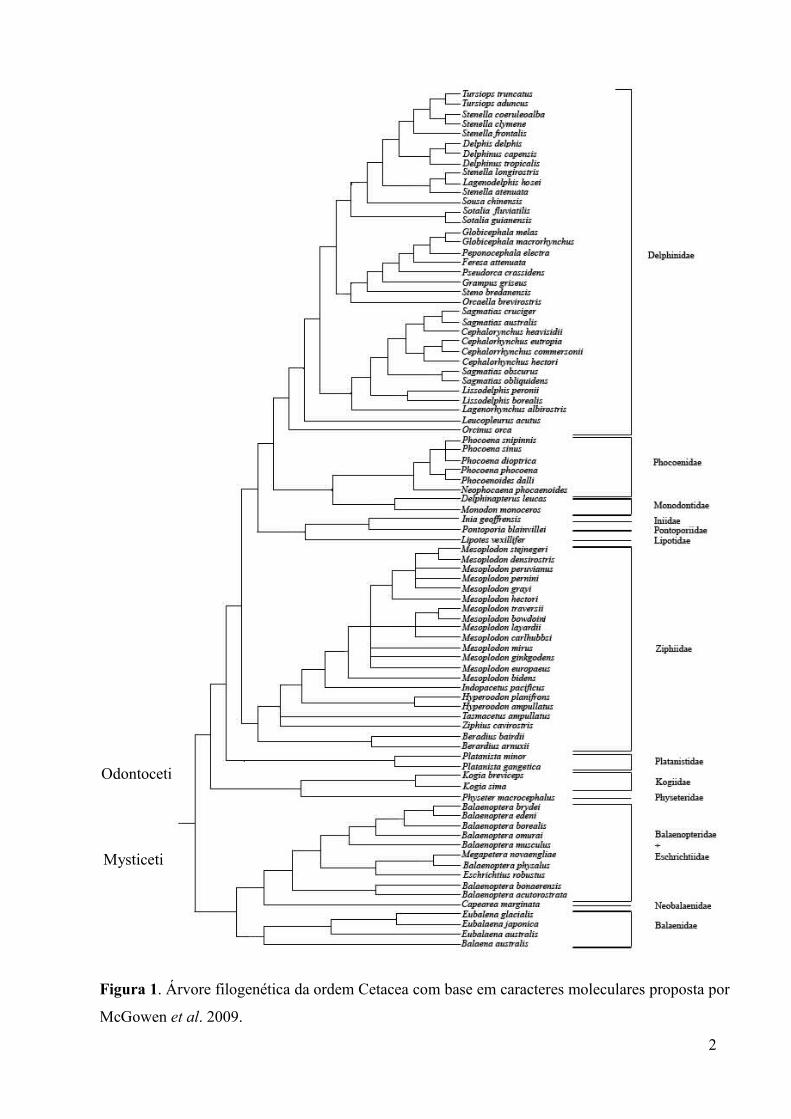
Figura 1. Árvore filogenética da ordem Cetacea com base em caracteres moleculares
proposta por McGowen et al. 2009.............................................................................................2
Figura 2. Mapa da distribuição do gênero Inia (da Silva et al. 2008)........................................3
Figura 3. Exemplar de Inia geoffrensis capturado na Reserva de Desenvolvimento
Sustentável Mamirauá. Foto: Diogo Souza...............................................................................13
Figura 4. Mapa geográfico em cinza a distribuição do gênero Inia e em destaque os locais de
coleta de Inia geoffrensis: a) Rio Branco AC, b) Reserva de Desenvolvimento Sustentável
Mamirauá AM e c) Aruanã. Mapa adaptado de da Silva et al. 2008.......................................14
Figura 5. Obtenção de amostra de sangue da nadadeira caudal do boto-vermelho. Foto: Heidi
Luz............................................................................................................................................15
Figura 6. a) Cariótipo de um macho Inia geoffrensis após coloração com Giemsa; b)
cromossomos sexuais de uma fêmea; c) cariótipo de uma fêmea após o bandeamento C d)
cromossomos sexuais de um macho; e) par 9sm evidenciando além da marcação terminal, um
bloco de heterocromatina intersticial em um dos homólogos. Barra igual a 10
µm.............................................................................................................................................27
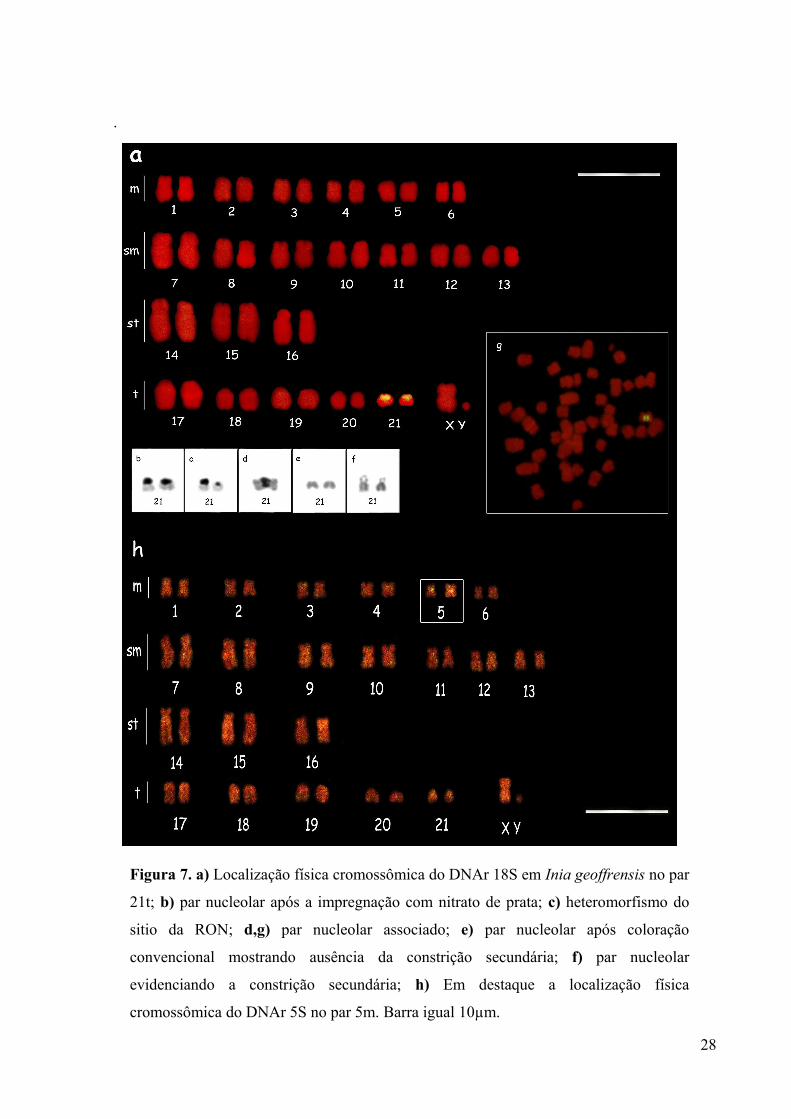
Figura 7. a) Localização física cromossômica do DNAr 18S em Inia geoffrensis no par 21t;
b) par nucleolar após a impregnação com nitrato de prata; c) heteromorfismo do sitio da
RON; d,g) par nucleolar associado; e) par nucleolar após coloração convencional mostrando
ausência da constrição secundária; f) par nucleolar evidenciando a constrição secundária; h)
Em destaque a localização física cromossômica do DNAr 5S no par 5m. Barra igual
10µm.........................................................................................................................................28
Figura 8. a) Padrão de bandeamento G de Inia geoffrensis seguido pelo ideograma da banda
G e bandeamento sequencial G/C. b) Cariótipo de um macho evidenciando o padrão de
distribuição do LINE 1(L1). c) Em destaque o cromossomo X, em banda G, banda C
sequencial e a distribuição do LINE (L1). Note que no braço curto existe maior acúmulo desta
xiii

sequência na região de heterocromatina, na região centromérica e no braço longo na região de
banda G positiva. Barra igual 10µm........................................................................................31
Figura 9. Cromossomos homeólogos entre o cariótipo de Inia geoffrensis e Tursiops
truncatus. Os números cromossômicos são apresentados abaixo de cada cromossomo. *
Cromossomos de I. geoffrensis. ** Cromossomos de T. truncatus (CRES, 2006). Barra igual
10µm.........................................................................................................................................32
Figura 10. Comparação do padrão de banda G do cromossomo X entre cinco espécies de
cetáceos. Eg Eubalaena glacialis baleia-franca-do-norte (Pause et al. 2006) ; Bb
Balaenoptera borealis baleia-sei (Nash 2006); Ig Inia geoffrensis boto-vermelho (presente
trabalho); Pp Phocoena phocoena boto-do-porto (Árnason 1980); Tt Tursiops truncatus
golfinho-nariz-garrafa-comum (CRES 2006). Note no braço curto do cromossomo X do boto-
vermelho a ocorrência de uma inversão paracêntrica. Barra igual 10µm.................................32
Figura 11. Cariótipo de um macho do Inia geoffrensis evidenciando a distribuição da
sequência telomérica. Barra igual 10µm...................................................................................33
xiv

Lista de Tabelas
Tabela 1. Formas cromossômicas descritas para as espécies da ordem Cetacea, subordem
Odontoceti compiladas da literatura (2n= número diploide; m=metacêntrico;
sm=submetacêntrico; st=subtelocêntrico; t=telocêntrico; P=puntiforme; XY=cromossomos
sexuais; p=braço curto; q=braço longo RON= Região organizadora do nucléolo (número do
par cromossômico, tipo cromossômico e braço);(*) Fórmulas cromossômicas diferentes para
uma mesma espécie; (-) Informações não determinadas nos trabalhos; (**) Nome atualizado
conforme World Cetacea Database http://www.marinespecies.org e Rice
1998.............................................................................................................................................6
Tabela 2. Formas cromossômicas descritas para as espécies da ordem Cetacea, subordem
Mysticeti compiladas da literatura (2n= número diploide; m=metacêntrico;
sm=submetacêntrico; st=subtelocêntrico; t=telocêntrico; P=puntiforme; XY=cromossomos
sexuais; p=braço curto; q=braço longo; RON=Região organizadora do nucléolo (número do
par cromossômico, tipo cromossômico e braço);(*) Fórmulas cromossômicas diferentes para
uma mesma espécie; (-) Informações não determinadas nos
trabalhos......................................................................................................................................9
Tabela 3. Média do comprimento total e da razão de braços dos cromossomos do boto-
vermelho, Inia geoffrensis, baseado na medida de três células em metáfase de 10 indivíduos.
Morfologia cromossômica relacionada a razão entre os braços (p/q) de acordo com Levan et
al. 1964. (-) Medidas não aplicadas ao cromossomo Y, considerado
puntiforme.................................................................................................................................25
Tabela 4. Alinhamento das sequências de DNAr 5S de mamíferos que apresentaram alta
similaridade com a sequência do boto-vermelho. Fonte: GenBank
(http://www.ncbi.nlm.nih.gov/)................................................................................................29
xv

Lista de Abreviações e Termos
µL Microlitros2n Número diploide Ag-RON Região organizadora de nucléolo argenteofílicaágua mili-Q Água ultra pura (deionizada e dessalinizada)AMPA Associação de Amigos do Peixe-boiBanda C Técnica de detecção da heterocromatina constitutivaBanda G Técnica de diferenciação das bandas eucromáticasBLAST Ferramenta de busca e alinhamento de sequências (basic local
alignment research tool)DNA Ácido desoxirribonucléicoDNAr DNA ribossomaldNTP DesoxirribonucleotídeoEDTA Ácido etilenodiaminotetracéticoFISH Hibridização fluorescente in situFITC Fluoresceina isotiocianato (fluorescein isothiocyanate)GenBank Banco de sequências públicas de DNAHCl Ácido ClorídricoHoméologo Mesmos cromossomos presentes em espécies distintasFISH heteróloga hibridização utilizando sonda de uma espécie em cromossomos
de outra espécieFISH homóloga hibridização utilizando sondas da própria espécieIPTG Isopropil-β-D-1-tiogalactopiranosídeo
(Isopropyl β-D-1-thiogalactopyranoside)KCl Cloreto de potássioLINE Elemento nucleolar intercalado longom Cromossomo metacêntricoMgCl2 Cloreto de magnésiomL MililitroNaCl Cloreto de sódioNCBI National Centre for Biotechnology InformationNF Número fundamental Nick translation Deslocamento de CorteNTS Espaçador não transcritopb Pares de basesPBD Tampão fosfato dextrano (phosphate-buffered dextran)PBS Tampão de bloqueioPCR Reação em cadeia da polimerase (polymerase chain reaction)Primer Oligonculeotídeos iniciadoresRDSM Reserva de Desenvolvimento Sustentável MamirauáRNAse Enzima que degrada o Ácido Ribonucléicorpm Rotações por minutoRPMI-1640 Meio de cultura completo para linfócitosSDS Dodecil sulfato de sódioSintenia Quando ocorre a presença física dos mesmos genes em um dado
cromossomo ou parte dele
xvi

SINE Elemento nucleolar intercalado curtosm Cromossomo submetacêntricoSSC Solução salina de citrato padrãost Cromossomo subtelocêntricoStop Buffer Tampão de paradat Cromossomo telocêntricoTaq Enzima termoestável derivada da bactéria Thermus aquaticusX-Gal 5 bromo-4-cloro-3-indolyl-β-D-galactoside
xvii

1. Introdução
1.1 Considerações sobre o boto-vermelho
O boto-vermelho é um golfinho fluvial endêmico das bacias Amazônica e do Orinoco e
pertence à família Iniidae, subordem Odontoceti (Best e Silva 1993) (Figura 1). Alguns
autores reconhecem a existência de uma única espécie, Inia geoffrensis com três subespécies
I .g. humboldtiana (distribuída na bacia do rio Orinoco), I. g. geoffrensis (rio Amazonas) e I.
g. boliviensis (na bacia dos rios Beni-Mamoré) (Trebbau e van Bree 1974; Pilleri e Gihr 1977;
Best e da Silva 1993; Rice 1998) (Figura 2). Entretanto, estudos morfométricos do crânio e de
sequências de DNA mitocondrial vêm legitimando a existência de duas espécies I. geoffrensis
(drenagem do rio Amazonas e Orinoco, exceto acima das cachoeiras do rio Madeira) e I.
boliviensis (acima das cachoeiras do rio Madeira e rio Beni-Mamoré) (da Silva 1994;
Hamilton et al. 2001; Banguera-Hinestroza et al. 2002).
Os ancestrais de Inia possivelmente adentraram na bacia Amazônica através do oceano
Pacífico durante o Mioceno. Neste período a drenagem da bacia do rio Amazonas corria em
direção ao Pacífico. Com o soerguimento dos Andes ocorreu a inversão do sentido da
drenagem do Amazonas e a ruptura de um grande sistema fluvial, resultando no isolamento de
três populações de Inia, que hoje são limitadas em sua distribuição por cachoeiras, corredeiras
e rios muito pequenos ou superficiais (Grabert 1984; Best e da Silva 1993; Fordyce e Barnes
1994; Lundberg et al. 1998; Hamilton et al. 2001).
Ao contrário da maioria dos cetáceos que vivem em ambientes oceânicos com pouca
variação entre uma estação e outra, o boto-vermelho ocorre em um ambiente aquático
dinâmico, com mudanças drásticas em relação ao nível da água ao longo do ano. Durante a
estação seca, quando o nível das águas está mais baixo, podem ficar restritos a lagos grandes e
profundos ou aos canais dos rios. Quando o nível da água dos rios sobe, eles se dispersam nos
igapós e nas planícies alagadas (várzea). O boto-vermelho possui o rostro longo e as
nadadeiras peitorais grandes. Além disso, as vértebras cervicais não são fusionadas, o que
proporciona uma grande flexibilidade do pescoço e do corpo. Somadas, estas adaptações
morfológicas tornam o boto-vermelho apto a perseguir e capturar presas entre os galhos e
raízes da floresta alagada (Best e da Silva 1993; da Silva 1994; Martin e da Silva 2004a).
1
2
Mysticeti
Odontoceti
Figura 1. Árvore filogenética da ordem Cetacea com base em caracteres moleculares proposta por
McGowen et al. 2009.

Figura 2. Mapa da distribuição do gênero Inia (da Silva et al. 2008).
Para esta espécie de golfinho de rio o período gestacional estimado é de 10 a 11
meses, e as fêmeas dão a luz a um único filhote. É possível que o intervalo entre os
nascimentos seja de três anos, com as fêmeas sexualmente maduras por volta dos sete anos de
idade (da Silva et al. 2008).
A degradação dos ecossistemas aquáticos coloca os golfinhos de rio entre os mais
ameaçados de todos os cetáceos (Hamilton et al. 2001). Atualmente, o boto-vermelho está
classificado como espécie “insuficientemente conhecida” de acordo com a “Lista Vermelha
de Espécies Ameaçadas” da União Internacional para a Conservação dos Recursos Naturais
(IUCN 2010), pois os dados sobre sua distribuição e abundância são incipientes e limitados a
algumas localidades. No Brasil o status do boto-vermelho é considerado “Vulnerável” pelo
Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (IBAMA 2001),
devido a inúmeras ameaças impostas à espécie como: a alteração do hábitat pelo represamento
dos rios para construção de barragens com fins hidrelétricos e pelo desflorestamento e pela
contaminação ambiental com inseticidas organoclorados utilizados na agricultura e com
mercúrio usado para amalgamar o ouro (Best e da Silva 1989; Rosas e Lehti 1996; Lailson-
Brito et al. 2008; Torres et al. 2009; da Silva 2009; da Silva e Martin 2010).
Os pescadores comerciais e de subsistência têm se tornado menos tolerantes com o
boto-vermelho, aumentando os conflitos que ocorrem quando estes retiram os peixes das
3

redes de pesca ou devido aos danos que causam nas mesmas. Mortes acidentais também
podem acontecer quando o boto-vermelho fica preso nas redes de espera (da Silva e Best
1996; Loch et al. 2009). Além disso, uma implacável mortandade de espécimes de boto-
vermelho tem acontecido, para o uso desta espécie como isca para a captura de um peixe
conhecido como piracatinga ou mota (Calophysus macropterus). Estima-se que em 2005, na
Amazônia Central aproximadamente 1600 botos foram mortos e usados como isca (da Silva e
Martin 2007; da Silva et al. 2008).
O boto-vermelho e o tucuxi (Sotalia fluviatilis) são duas espécies simpátricas (Best e da
Silva 1993), com grande importância no ecossistema amazônico, pois assim como outros
predadores, contribuem para a manutenção da saúde e equilíbrio das populações de peixes, já
que os indivíduos mais fracos ou doentes são presas fáceis, imediatamente comidas. Os
golfinhos da Amazônia também têm papel relevante na fertilização dos lagos de várzea, pois
possuem um metabolismo alto e ingerem grandes quantidades de presas, e como estão em
constante movimento suas fezes auxiliam na fertilização desses lagos (da Silva et al. 2008).
1.2 Estudos Citogenéticos em Cetáceos
Em mamíferos, a análise do complemento cromossômico é utilizada na resolução de
problemas taxonômicos (Machado et al. 2005), na detecção de espécies crípticas (Silva et al.
2006) no manejo e na conservação de espécies silvestres (Vianna et al. 2006; Rubes et al.
2008). Em cetáceos, também é empregada na sexagem de animais com hermafroditismo
(Tarpley et al. 1995), no estudo da viabilidade de espermatozóides criopreservados (Watanabe
et al. 2007) e na avaliação de danos causados ao material genético por poluentes no ambiente
aquático (Wise et al 2008).
Para as cinco espécies de mamíferos aquáticos, encontradas na Amazônia brasileira,
dados sobre a constituição cromossômica ainda são incipientes. Caracterizações iniciais já
foram realizadas para o peixe-boi amazônico Trichechus inunguis (Assis et al. 1988), para
ariranha Pteronura brasiliensis (Franco-de-Sá et al. 2007) e para lontra Lontra longicaudis
(Freitas et al. 1975). Entre os golfinhos somente o número diploide do boto-vermelho foi
determinado (Kulu et al. 1971). Já para o tucuxi Sotalia fluviatilis nenhuma informação
acerca de seus cromossomos ainda é conhecida.
Atualmente, existem 86 espécies de cetáceos descritas para a ordem Cetartiodactyla
(Perrin et al. 2009; Di Beneditto et al. 2010), e estudos citogenéticos realizados em 37
espécies (Tabelas 1 e 2) indicam que a macroestrutura cromossômica pode ser considerada
4

conservada (Árnason 1972; 1974). Embora, variações em relação ao número diploide, às
fórmulas cariotípicas, à distribuição das regiões organizadoras do nucléolo (Tabelas 1 e 2) e
ao padrão de heterocromatina constitutiva já foram observadas (Árnason 1974; Duffield et al.
1978).
Para golfinhos e baleias o número diploide varia entre 42 e 44 cromossomos, com
sistema de determinação sexual simples XX/XY (Makino 1948; Árnason 1969). A
heterocromatina constitutiva está geralmente localizada na posição terminal ou/e intersticial
dos cromossomos, mas não exclusivamente, podendo apresentar blocos pequenos
evidenciados na região centromérica ou pericentromérica (Árnason 1974; Árnason et al.
1978). Em misticetos, como exemplo na baleia-cinza Eschrichtius robustus (Árnason 1981b)
e na baleia-sei Balaenoptera borealis (Árnason 1974) os blocos de heterocromatina
centromérica ocorrem principalmente nos cromossomos metacêntricos e submetacêntricos.
5

Tabela 1. Formas cromossômicas descritas para as espécies da ordem Cetacea, subordem Odontoceti compiladas da literatura (2n= número diploide;
m=metacêntrico; sm=submetacêntrico; st=subtelocêntrico; t=telocêntrico; P=puntiforme; XY=cromossomos sexuais; p=braço curto; q=braço longo RON= Região
organizadora do nucléolo (número do par cromossômico, tipo cromossômico e braço);(*) Fórmulas cromossômicas diferentes para uma mesma espécie; (-)
Informações não determinadas nos trabalhos; (**) Nome atualizado conforme World Cetacea Database http://www.marinespecies.org e Rice 1998.
Família Espécie 2n Fórmula cariotípica X Y Banda G Banda C RON ReferênciaDelphinidae Stenella longirostris 44 12m+16sm+4st+10t sm P - X 3tp,5tp Stock 1981
Stenella attenuata**44 12m+18sm+4st+8t
12m+16sm+4st+10t*
sm
m
P
P
X X 3tp,5tp Árnason 1974; Árnason 1980; *Stock 1981
Stenella clymene 44 12m+18sm+4st+8t m t X X 9smp Árnason 1980Steno bredanensis 44 - - - - - - Kulu 1972Delphinus delphis 44 14m+16sm+4st+8t m t - - - Árnason 1974
Tursiops truncatus
44 12m+18sm+4st+8t sm t X X 9smp,
3tp
Walen e Madin 1965; Duffield et al. 1967;
Prasad et al. 1970; Árnason 1974; Bielec et
al. 1997; CRES 2006Lagenorhynchus
albirostris
44 12m+18sm+4st+8t m - X X 9smp Árnason 1980; Árnason 1981a
Lagenorhynchus
obliquidens
44 8m+14sm+10st+10t sm t - - - Duffield et al. 1967
Orcinus orca44 12m+18sm+4st+8t
20m+12sm+10st*
sm t X X 9smp,
4 tp
Carr et al. 1966; Horrall et al. 1968, Kulu et
al. 1971*; Kulu 1972; Árnason et al. 1980 Globicephala
macrorhynchus
44 12m+16sm+6st+8t sm - X - Walen e Madin 1965; Árnason 1974
6
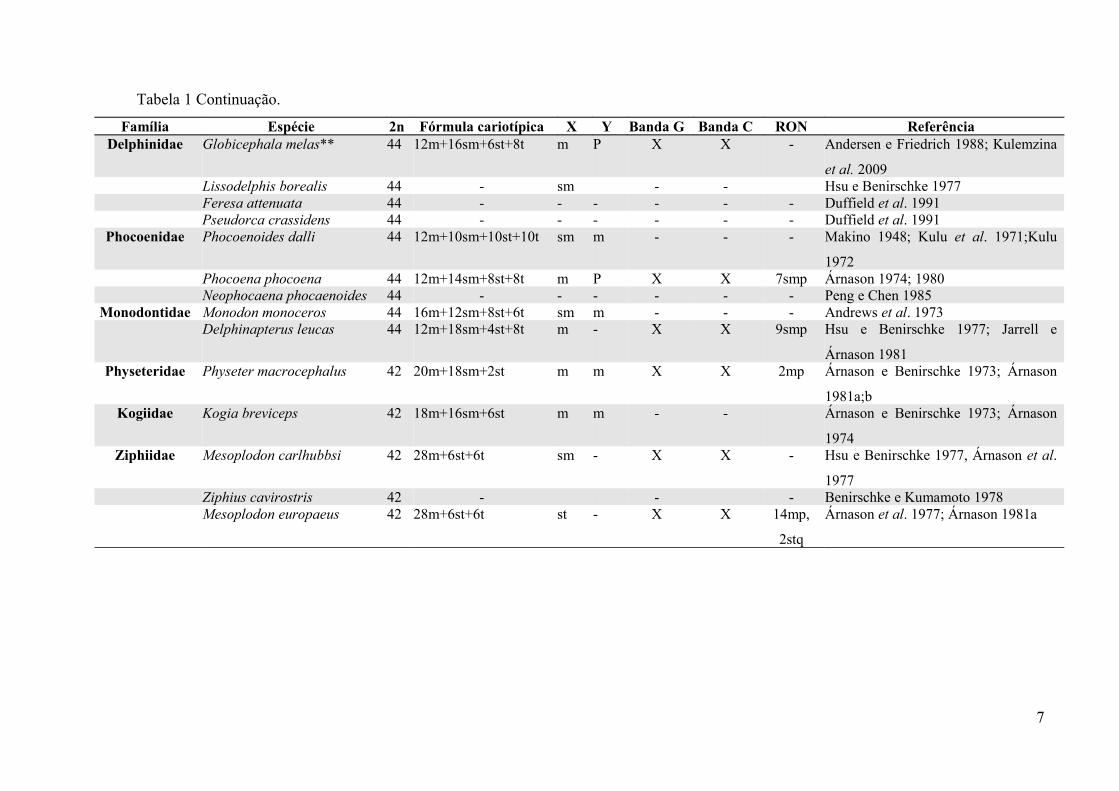
Tabela 1 Continuação.
Família Espécie 2n Fórmula cariotípica X Y Banda G Banda C RON ReferênciaDelphinidae Globicephala melas** 44 12m+16sm+6st+8t m P X X - Andersen e Friedrich 1988; Kulemzina
et al. 2009Lissodelphis borealis 44 - sm - - Hsu e Benirschke 1977Feresa attenuata 44 - - - - - - Duffield et al. 1991
Pseudorca crassidens 44 - - - - - - Duffield et al. 1991Phocoenidae Phocoenoides dalli 44 12m+10sm+10st+10t sm m - - - Makino 1948; Kulu et al. 1971;Kulu
1972 Phocoena phocoena 44 12m+14sm+8st+8t m P X X 7smp Árnason 1974; 1980Neophocaena phocaenoides 44 - - - - - - Peng e Chen 1985
Monodontidae Monodon monoceros 44 16m+12sm+8st+6t sm m - - - Andrews et al. 1973Delphinapterus leucas 44 12m+18sm+4st+8t m - X X 9smp Hsu e Benirschke 1977; Jarrell e
Árnason 1981Physeteridae Physeter macrocephalus 42 20m+18sm+2st m m X X 2mp Árnason e Benirschke 1973; Árnason
1981a;bKogiidae Kogia breviceps 42 18m+16sm+6st m m - - Árnason e Benirschke 1973; Árnason
1974Ziphiidae Mesoplodon carlhubbsi 42 28m+6st+6t sm - X X - Hsu e Benirschke 1977, Árnason et al.
1977Ziphius cavirostris 42 - - - Benirschke e Kumamoto 1978Mesoplodon europaeus 42 28m+6st+6t st - X X 14mp,
2stq
Árnason et al. 1977; Árnason 1981a
7
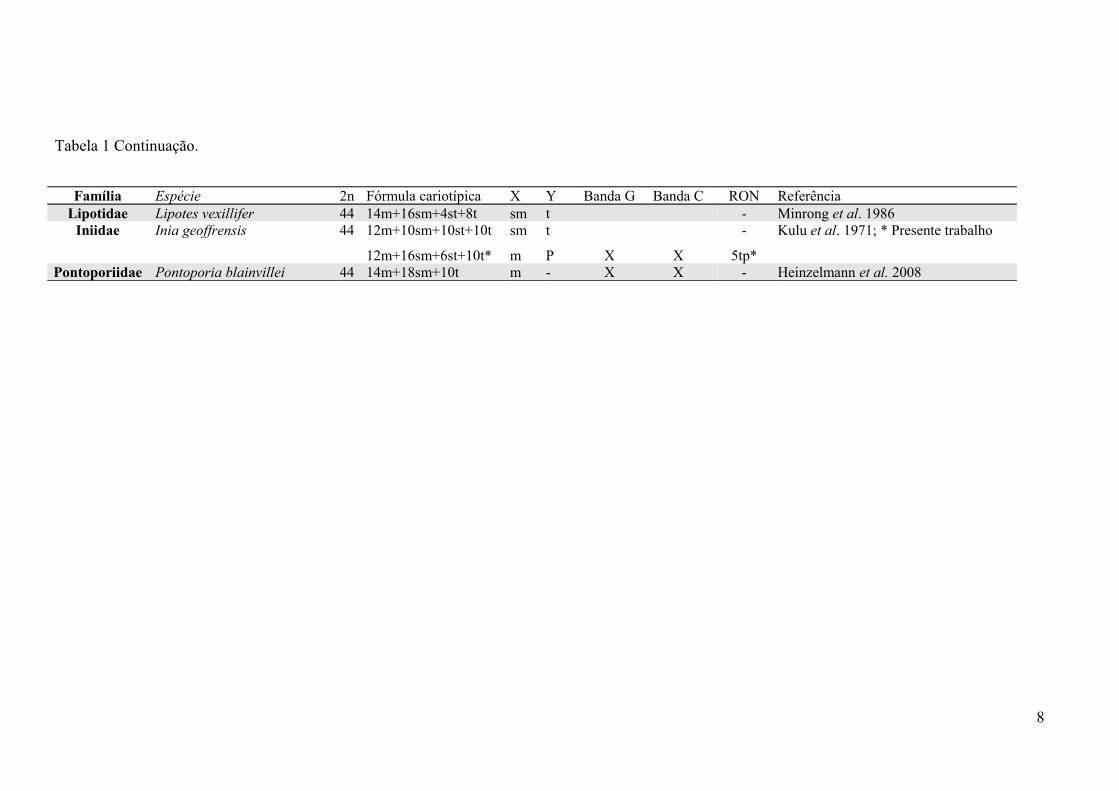
Tabela 1 Continuação.
Família Espécie 2n Fórmula cariotípica X Y Banda G Banda C RON ReferênciaLipotidae Lipotes vexillifer 44 14m+16sm+4st+8t sm t - Minrong et al. 1986
Iniidae Inia geoffrensis 44 12m+10sm+10st+10t
12m+16sm+6st+10t*
sm
m
t
P X X
-
5tp*
Kulu et al. 1971; * Presente trabalho
Pontoporiidae Pontoporia blainvillei 44 14m+18sm+10t m - X X - Heinzelmann et al. 2008
8

Tabela 2. Formas cromossômicas descritas para as espécies da ordem Cetacea, subordem Mysticeti compiladas da literatura (2n= número diploide; m=metacêntrico;
sm=submetacêntrico; st=subtelocêntrico; t=telocêntrico; P=puntiforme; XY=cromossomos sexuais; p=braço curto; q=braço longo; RON=Região organizadora do
nucléolo (número do par cromossômico, tipo cromossômico e braço);(*) Fórmulas cromossômicas diferentes para uma mesma espécie; (-) Informações não
determinadas nos trabalhos.
Família Espécie 2n Fórmula cariotípica X Y Banda G Banda C RON ReferênciaEschrichtiidae Eschrichtius gibbosus 44 18m+10sm+6st+8t m - - - - Árnason 1974
Eschrichtius robustus 44 18m+10sm+6st+8t m - X X 5smp Árnason 1981a, bBalaenopteridae Balaenoptera acutorostrata 44 18m+10sm+6st+8t m st X X 5smp Árnason 1974; Árnason et al.
1977; Árnason 2006Balaenoptera borealis 44 14m+14sm+6st+8t m m X X - Kasuya 1966; Árnason 1970;
Árnason 1974; Árnason 2006 Balaenoptera physalus 44 14m+14sm+6st+8t
16m+12sm+6st+8t*
st
st
m
m
X X 6smp Árnason 1969*; 1974; 1981a
Balaenoptera musculus 44 14m+14sm+6st +8t sm m X X - Árnason et al. 1985; Árnason
e Widegren 1989Megaptera novaeangliae 44 14m+14sm+6st+8t sm- t- - X - Duffield et al. 1991; Árnason
2006Neobalaenidae Caperea marginata 44 - - - - - - Árnason 2006 Balaenidae Balaena mysticetus 42 18m+ 10sm+ 6st+6t m m X - 7m Jarrel 1979
Eubalaena glacialis 42 12m + 22sm + 6st sm t X - - Pause et al. 2006
9

Recentemente, análises mais acuradas empregando técnicas de citogenética molecular
para mapeamento físico dos genes, têm sido aplicadas a cetáceos, permitindo comparações
cariotípicas com espécies distantemente relacionadas do ponto de vista filogenético (Bielec et
al. 1998a; Kulemzina et al. 2009). Estas análises permitem detectar homeologias
cromossômicas, identificar grupos sintênicos, localizar genes específicos e identificar
polimorfismos e/ou variabilidade não detectados por ferramentas da citogenética clássica.
Além disso, ampliam o conhecimento sobre a evolução cromossômica (Yonenaga-Yassuda,
2004; Martins et al. 2010).
No cariótipo do golfinho-nariz-de-garrafa-comum Tursiops truncatus já foi observado
que 36 segmentos sintênicos são conservados entre humanos e golfinhos. Também foram
mapeados vários genes como o do DNA ribossomal 28S, gene da β-glucocerebrosidase
relacionado à doença de Gaucher, gene da interleucina-8 proteína ligada ao processo
inflamatório e o gene do cristalino CRYBA1 associado à formação de catarata em humanos
(Bielec et al. 1997; Bielec et al. 1998a; 1998b; 1998c; Bielec et al. 2000).
Um amplo estudo do genoma de cinco espécies da ordem Cetartiodactyla, a qual inclui
os cetáceos e os artiodátilos, foi realizado por meio da construção de mapas cromossômicos.
Estes mapas obtidos por pintura cromossômica utilizaram sondas de cromossomos humanos e
de dromedários, com posterior hibridização in situ heteróloga em metáfases do hipopótamo
(Hippopotamus amphibius), do ocapi (Okapia johnstoni), do cervo-almiscarado-siberiano
(Moschus moschiferus) e da baleia-piloto-de-peitorais-longas (Globicephala melas). Com a
comparação destes mapas cromossômicos foi possível reconstruir o caminho evolutivo e
determinar as taxas de rearranjos cromossômicos ocorridos na evolução dos cetartiodátilos. O
cariótipo da baleia foi considerado o mais conservado entre os cetartiodátilos analisados,
mostrando que a taxa de evolução cromossômica em cetáceos é extremamente lenta
(Kulemzima et al. 2009).
Além disso, o emprego da citogenética molecular, utilizando sequências de DNA
repetitivo, tem possibilitado uma melhor compreensão sobre os processos evolutivos que
estão associados aos rearranjos cromossômicos e às alterações de sequências específicas do
genoma (Martins et al. 2010). Os elementos repetitivos formam uma grande fração do
genoma dos mamíferos (Acosta et al. 2008). Algumas destas sequências são organizadas em
tandem, como as famílias multigênicas, o DNA satélite e a sequência telomérica, enquanto
outras se encontram dispersas ao longo do genoma como os elementos transponíveis
(Charlesworth et al. 1994).
10

Existem duas famílias multigênicas de DNA ribossomal, DNAr 45S e DNAr 5S cujos
genes apresentam taxa de repetição moderada. O DNAr 45S correspondente à região
organizadora do nucléolo, codifica os RNAr 18S, 5,8S, 28S e dentre estes o RNAr 18S
compõe a subunidade menor dos ribossomos. O DNAr 5S é formado por múltiplas cópias
altamente conservadas de uma sequência de 120 pares de bases, separadas por um espaçador
não transcrito. Este gene não está relacionado à formação do nucléolo, situando-se em um
ambiente cromossômico distinto. O RNAr 5S é processado fora do nucléolo e posteriormente
se junta ao RNAr 5,8S e 28S para a formação da subunidade maior do ribossomo (Long e
Dawid 1980; Martins et al. 2010). A caracterização quanto à identidade nucleotídica e o
mapeamento dos genes de DNAr têm sido utilizados em estudos evolutivos de mamíferos
(Zijlstra et al. 1997; Mäkinen et al. 1997; Christensen et al. 1998; Tanaka et al. 2000;
Pienkowska et al. 2002).
Outra classe de DNA repetitivo inclui as sequências teloméricas, formadas por unidades
de repetições de TTAGGG, predominantemente encontradas nas extremidades dos braços
cromossômicos para protegê-las e manter a estabilidade cromossômica (McEachern et al.
2000; Blackburn 2005; Castiglia et al. 2006). A localização destas sequências em regiões
intersticiais pode evidenciar a ocorrência de rearranjos cromossômicos como translocação
e/ou fusão em tandem (Castiglia et al. 2006).
Os elementos transponíveis são uma importante classe de sequências repetitivas, devido
a sua habilidade de transpor de uma região cromossômica para outra, inserindo-se em novos
sítios, próximos, ou até mesmo dentro de sequências gênicas, promovendo rearranjos
cromossômicos, mutações, alteração da estrutura e função de genes, mudanças na regulação
gênica e aumento do tamanho do genoma. Desta forma, contribuem para originar diversidade
alélica e para a criação de novos genes (Biémont e Vieira 2006; Feschotte e Pritham 2007;
O’Donnel e Burns 2010). Considera-se que a invasão dos elementos móveis e sua
subsequente ação de amplificação seja o principal fator morfogenético que está assegurando a
adaptação das populações de diferentes organismos às mudanças ambientais e em alguns
casos, causando rápida especiação (Evgen’ev 2007).
Em mamíferos, a família do retroelemento LINE1 compõe grande porção do genoma de
humanos (16,9%), camundongos (18,9%) e do marsupial Monodelphis domesticus (20%)
(Gentles et al. 2007) e tem um papel fundamental no remodelamento do genoma (Lee et al.
2010). Diversas funções são atribuídas ao LINE, entre elas prover a maquinaria enzimática
para a retrotransposição de elementos não autônomos da família SINE e formação de
11

pseudogenes processados, além de modular a expressão gênica fornecendo sinais regulatórios
da transcrição (Esnalt et al. 2000; Nigumann et al. 2002; Lee et al. 2010). Além disso, o papel
do LINE na inativação do cromossomo X tem sido investigado (Lyon 2005). Entretanto
poucos estudos que enfoquem o mapeamento físico cromossômico deste retroelemento vêm
sendo realizados em mamíferos.
Inúmeras espécies têm sido estudadas com ferramentas citogenômicas, entretanto para o
boto-vermelho as análises genéticas ainda são incipientes. Marcadores moleculares foram
desenvolvidos para estudos da estrutura populacional e relações de parentesco (Gravena et al.
2009), entretanto estudos citogenéticos determinaram apenas seu número diploide (Kulu et al.
1971).
Desta forma, uma caracterização cariotípica do boto-vermelho, por meio de marcadores
de citogenética clássica e molecular, com o mapeamento físico cromossômico de sequências
repetitivas de DNA, pode propiciar uma melhor visão da sua organização genômica e gerar
marcadores úteis para investigar as relações evolutivas deste golfinho em relação a outros
cetáceos, além de inferir sobre a evolução cromossômica deste grupo.
1.3 Objetivos
• Caracterizar o cariótipo do boto-vermelho (Inia geoffrensis) através de técnicas de
citogenética clássica e molecular.
• Verificar se existe polimorfismo cromossômico na população do boto-vermelho na
Reserva de Desenvolvimento Sustentável Mamirauá, AM-Brasil.
• Situar o boto-vermelho dentro da carioevolução dos cetáceos.
12

2. Material e métodos
2.1 Material e área de estudo
Foram estudados citogeneticamente 24 espécimes (11 fêmeas e 13 machos) de Inia
geoffrensis (Figura 3), dos quais amostras de tecido epitelial e sangue foram retiradas
Figura 3. Exemplar de Inia geoffrensis capturado na Reserva de Desenvolvimento
Sustentável Mamirauá. Foto: Diogo Souza
No âmbito do Projeto-Boto, Sub-Projeto: “Marcação e Monitoramento de botos, Inia
geoffrensis e Sotalia fluviatilis” foram capturados 21 animais em novembro de 2009, na
Reserva de Desenvolvimento Sustentável Mamirauá (RDSM), situada próxima à confluência
dos rios Japurá e Solimões (3º20’S, 64º54’W), cerca de 500 km a oeste da cidade de Manaus
AM. A captura foi realizada com autorização do IBAMA (licença nº 13462-3), emitida em
21/10/2009.
Ainda, amostras de tecido e sangue de dois indivíduos capturados no Lago dos Cavalos,
rio Vermelho (14º 93’S, 51º09’ W) próximo a Aruanã (GO) e de um indivíduo no Igarapé
Caipora, rio Acre (10º 09' S, 67º 54' W) próximo à Rio Branco (AC) foram gentilmente
cedidas pelo Laboratório de Mamíferos Aquáticos do INPA (Figura 4).
13

2.2 Métodos
2.2.1 Coleta do Material
Os animais foram capturados com uso de redes de malha 120-150 mm, colocadas
durante a madrugada na entrada dos canais da Reserva de Desenvolvimento Sustentável
Mamirauá, durante o mês de novembro, período em que o nível do rio está baixo e os canais
estreitos. Ao se avistar um grupo de golfinhos nadando em direção à rede, uma segunda rede
foi colocada por trás do grupo cruzando o canal para prevenir o seu escape. Posteriormente, a
área foi subdividida com outras redes para a retirada dos animais individualmente. Os animais
foram conduzidos um a um para a margem do rio, retirados da água em uma maca e levados
até o flutuante, selecionando apenas um animal por vez, com exceção de mãe com filhote que
foram mantidos sempre juntos. Durante o tempo em que cada animal esteve fora d’água
14
Figura 4. Mapa geográfico, mostrando a distribuição do gênero Inia em cinza e em destaque
os locais de coleta de Inia geoffrensis: a) Rio Branco, AC, b) Reserva de Desenvolvimento
Sustentável Mamirauá, AM e c) Aruanã, GO. Mapa adaptado de da Silva et al. 2008.

monitorou-se a frequência respiratória e a pele e os olhos foram molhados constantemente
para prevenir a dessecação ou superaquecimento (da Silva e Martin 2000).
Cada animal foi pesado e medido. Amostra de sangue foi retirada da veia caudal que
drena a parte ventral do pedúnculo caudal, utilizando scalps 19G e acondicionada em tubo
vacutainer heparinizado (Figura 5). O tecido epitelial foi retirado da nadadeira dorsal e
armazenado em tubos de 1,5 mL com álcool etílico 100%.
Figura 5. Obtenção de amostra de sangue da nadadeira
caudal do boto-vermelho. Foto: Heidi Luz
2.2.2 Obtenção dos cromossomos mitóticos
As amostras de sangue coletadas dos animais da RDSM foram semeadas em meio
completo para cariótipo no mesmo dia de coleta, no flutuante base do Projeto Boto. Já, as
amostras de Rio Branco (AC) e Aruanã (GO) foram acondicionadas e transportadas em isopor
com gelo e processadas até trinta horas após a coleta no Laboratório de Genética Animal do
INPA em Manaus (AM).
Para obtenção das preparações cromossômicas foi realizada a cultura de linfócitos de
sangue periférico (Moorhead et al. 1960). Foi adicionado ao frasco de meio de cultura
completo para cariótipo (RPMI) 0,5 mL de sangue total, misturando-se delicadamente. Em
seguida a cultura foi colocada no banho-maria a 37 ºC por 72 horas. Ao completar 71 horas,
foi acrescentado à cultura 0,3 mL de colchicina a 0,0125%. O frasco foi agitado levemente e
recolocado no banho-maria. Finalizado o período de 60 minutos, o frasco foi retirado do
banho-maria e a suspensão foi homogeneizada e transferida para um tubo falcon, o qual foi
15

centrifugado a 800 rpm por oito minutos. Posteriormente, o sobrenadante foi descartado e foi
acrescentado 7 mL de solução hipotônica de KCl, 0,075 M, que foi colocado na estufa a 37 ºC
por 25 min. Após este período foi centrifugado a 800 rpm por 8 minutos e o sobrenadante foi
descartado. Realizou-se uma fixação e três lavagens com fixador Carnoy (metanol: ácido
acético - 3:1). A suspensão celular foi armazenada em tubo de 1,5 mL e acondicionada em
frezzer (-10 ºC).
Citogenética Clássica
2.2.3 Detecção das regiões de heterocromatina constitutiva: Banda C (CBG)
Para a caracterização do padrão de heterocromatina constitutiva utilizou-se a técnica de
banda C descrita por Sumner (1972). As lâminas contendo a preparação cromossômica foram
tratadas durante 2 minutos com HCl 0,2N a 45 ºC, lavadas em água destilada à temperatura
ambiente e secas ao ar. Em seguida foram incubadas em solução de hidróxido de bário a 5%,
filtrada e recém preparada a 45 ºC, por 1 minuto e 40 segundos. A ação do hidróxido de bário
foi interrompida imergindo-se rapidamente a lâmina em solução de HCl 0,2N à temperatura
ambiente, sendo posteriormente lavada em água destilada. Após secas, as lâminas foram
incubadas em solução 2xSSC (cloreto de sódio 0,3M e citrato trisódico 0,03M, pH 6,8), em
estufa a 60 ºC, por um período de 15 minutos, lavadas em água destilada e secas ao ar.
Posteriormente, as lâminas foram coradas com Giemsa 5% em tampão fosfato 0,06M e pH
6,8 por 4 minutos, lavadas em água corrente e secas ao ar.
2.2.4 Bandeamento G (GTG)
Para obtenção do padrão de bandas G foi utilizada a técnica de Seabright (1971), com
modificações. As lâminas contendo a suspensão celular foram envelhecidas por 4 dias em
estufa a 37 ºC e tratadas por 1 hora em solução de 2XSSC a 60 °C (cloreto de sódio 0,3M e
citrato trissódico 0,06M e pH 6,8), lavadas em água destilada e deixadas secar ao ar. Em
seguida, estas foram imersas em uma solução de tripsina a 37 °C (0,024g de tripsina em 40
mL de tampão fosfato pH 6,8) por 17segundos. A ação da solução de tripsina foi interrompida
em uma solução gelada de tampão fosfato. As lâminas foram lavadas em água destilada, secas
ao ar e coradas com solução de Giemsa, diluída a 5% em tampão fosfato pH 6,8 por 4 minutos
e depois lavadas em água destilada e secas ao ar.
16

Após a realização do bandeamento G, as lâminas com as melhores metáfases foram
fotografadas e posteriormente colocadas em uma cubeta contendo álcool 100% por 20
minutos para retirada do óleo de imersão e do corante. Em seguida foram lavadas em água
destilada e secas ao ar. Após este procedimento, utilizou-se o protocolo de detecção da
heterocromatina constitutiva para realização do bandeamento C sequencial, conforme descrito
anteriormente.
2.2.5 Detecção das regiões organizadoras do nucléolo (Ag-RON)
Para a caracterização da região organizadora de nucléolo utilizou-se a técnica descrita
por Howell e Black (1980), que consistiu em pingar sobre a lâmina contendo a preparação
cromossômica, 1 gota de uma solução coloidal obtida com 0,25 g de gelatina comercial sem
sabor dissolvida em 12,5 mL de água destilada e acrescida de 0,125 mL de ácido fórmico. Em
seguida, foram adicionadas sobre a solução coloidal 2 gotas de solução aquosa de AgNO3
(nitrato de prata) a 50%. A lâmina foi coberta com lamínula, colocada em câmara úmida e
levada à estufa a 60 ºC por 3 minutos e 40 segundos. Após a lâmina adquirir uma coloração
marrom, esta foi lavada em água destilada e seca ao ar.
Citogenética Molecular
2.2.6 Extração de DNA
O DNA total do boto-vermelho foi extraído de fragmentos de tecido epitelial da
nadadeira dorsal, utilizando o kit comercial QIAmp DNA (QIAGEN) conforme especificações
do fabricante.
Além disso, também foi extraído o DNA total do marsupial Caluromys philander a
partir de fragmentos de tecido muscular, com o intuito de se obter sondas para realização de
hibridização in situ fluorescente (FISH) heteróloga, utilizada no mapeamento físico
cromossômico dos genes ribossomais. A extração foi realizada conforme o protocolo descrito
por Sambrook e Russell (2001) com algumas modificações. Para tanto, adicionou-se a um
tubo de 1,5 mL 0,02 g do tecido muscular e 500μL de tampão de lise (8 mL de Tris-HCl 1M
pH 8,0, 1 mL NaCl 0,3 M, 2 mL EDTA 0,5M e 20 mL de SDS 10% e água para completar
100 mL) de Estoup et al. (1993) e com acréscimo de uréia 4M segundo Asahida et al. (1996).
Em seguida foi acrescentado 15 μL de proteinase K (10 mg/mL) e as amostras foram
17

incubadas a 60 ºC por aproximadamente 1 hora com agitação. Posteriormente as amostras
foram incubadas 37 ºC acrescentando-se 15 μL de RNAse (10 mg/mL).
Após a digestão total do tecido foram feitas lavagens sucessivas com fenol-clorofórmio
(1:1) e clorofórmio. O DNA foi separado das proteínas por precipitação salina com
centrifugação a 14000 rpm e o sobrenadante foi precipitado com 600 μL de isopropanol 100%
gelado também com o auxílio de centrifugação. Ao final, o DNA precipitado foi lavado com 1
mL de etanol 70% com centrifugação a 14000 rpm e hidratado com aproximadamente 100 μL
de água milli-Q, dependendo do tamanho do pellet formado.
Para possibilitar a análise da quantidade e integridade do material, o DNA extraído foi
quantificado por comparação com marcador Lambda DNA (Invitrogen), em eletroforese
padrão com tampão Tris-Borato (EDTA 1X e corrida a 70 V por 40 minutos) em gel de
agarose 0,8% e corado com 3 μL GelRed Acid Gel Stain (Biotium 1:500) e utilizando 3 μL de
DNA. A visualização dos géis e análise do DNA foi feita no fotodocumentador Easy Doc 100
(BioAgency) com transluminador de luz ultravioleta (260 nM).
2.2.7 Obtenção das sondas via PCR (Polymerase Chain Reaction)
As sondas do DNAr 5S e 18S, e do elemento transponível LINE L1 foram obtidas a
partir da amplificação por Reação em Cadeia da Polimerase (PCR - Saiki et al. 1988) em um
volume total de 15 μL [1 μL de DNA genômico( ~100 ng); 1,5 μL Tampão 10X; 0,15 de Taq
DNA Polimerase (5 U/uL) ; 3,0 μL de dNTP (1 mM); 0,6 μL de cada primer (5 pmol) e 8,15
μL água mili-Q)]. Para as reações de amplificação das sequências teloméricas, a mesma
concentração dos reagentes foi mantida, porém sem a presença de DNA molde, devido à
característica de auto anelamento de dupla fita dos primers. As reações foram processadas em
termociclador (Applied Biosystems, Vereti) e os primers utilizados para a amplificação destas
sequências alvos, bem como os perfis dos ciclos de amplificação foram os seguintes:
DNAr 5SPrimer: A 5’-TACGCCCGATCTCGTCCGATC-3’ e B (5’-
CAGGCTGGTATGGCCGTAAGC-3’ (Komiya e Takemura 1979). Ciclos de amplificação: 1
minuto a 94 ºC (desnaturação), 30 ciclos de 1 minuto a 94 ºC, 1 minuto de 57 ºC (anelamento
dos primers) e 1 minuto de 72 ºC (extensão); 5 minutos a 72 ºC (extensão final).
18

DNAr 18SPrimer: IpF 5’-CCGCTTTGGTGACTCTTGAT-3’ e IpR 5’-
CCGAGGACCTCACTAAACCA 3’ (Gross et al. 2010). Ciclo de amplificação: 1 minuto a
95 ºC (desnaturação); 35 ciclos de 1 minuto a 94 ºC, 1 minuto a 56 ºC (anelamento dos
primers) e 1 minuto e 30 segundos a 72 ºC (extensão); 3 minutos a 72 ºC (extensão final).
Sequência Telomérica
Primer: F (5’ TTAGGG-3’)5 e R (5’ CCCTAA-3’)5 (Ijdo et al. 1991). Ciclos de amplificação:
10 ciclos de 1 minuto a 94 ºC (desnaturação), 30 segundos a 55 ºC (anelamento dos primers),
1 minuto a 72 ºC (extensão); 30 ciclos de 1 minuto a 94 ºC (desnaturação), 30 segundos a 60
ºC (anelamento dos primers), 1 minuto e 30 segundos a 72 ºC (extensão); de 5 minutos 72 ºC
(extensão final).
LINE L1Primer: L1-F 5’ CCATGCTCATSGATTGG- 3’ e L1-R 5’ ATTCTRTTCCATTGGTCTA-3’,
(Waters et al. 2004; Rebuzzini et al. 2009). Ciclos de amplificação: 2 minutos a 94 ºC
(desnaturação), 30 ciclos de 1 minuto a 94 ºC (desnaturação); 1 minuto a 51 ºC (anelamento
dos primers), 1 minuto e 30 segundos a 72 ºC (extensão); 5 minutos a 72 ºC (extensão final).
Depois de amplificados, os produtos da PCR foram verificados e quantificados por
comparação com o marcador Low Mass DNA Ladder (Invitrogen) em eletroforese em gel de
agarose 1,0%, corado com GelRed Acid Gel Stain (Biotium 1:500). A visualização e
documentação dos géis de agarose foram realizadas no fotodocumentador Easy Doc 100
(BioAgency). Os produtos de PCR foram utilizados como sondas para mapeamento desta
sequências nos cromossomos do boto-vermelho.
2.2.8 Marcação das sondas
As sondas foram marcadas pelo método Nick translation utilizando o Kit BioNickTM
Labeling System (Invitrogen). Para a preparação do mix adicionou-se, em um tubo de 1,5 mL
no gelo, os seguintes componentes: 1µL de DNA (200 µg/µL); 1µL de dNTP mix; 1µL de
enzima; e 6 µL de água. Este tubo foi centrifugado brevemente e incubado por 45 minutos a
16 ºC. A reação foi interrompida com a adição de 1µL de Stop Buffer. Para verificar se o
DNA foi clivado realizou-se uma eletroforese em gel de agarose 1,0% e o gel foi visualizado
no fotodocumentador Easy Doc 100 (BioAgency).
19

2.2.9 Hibridização in situ fluorescente (FISH)
A hibridização in situ fluorescente homóloga e heteróloga foram realizadas em
condições de alta estringência (77 %) conforme a técnica descrita por Pinkel et al. (1986),
com alguns ajustes.
Tratamento das lâminas
As lâminas foram lavadas por 5 minutos em tampão PBS 1X [7,58 g de cloreto de sódio
1,36 M; 0,993g de fosfato de sódio dibásico (Na2HPO4); 0,414g de fosfato de sódio
monobásico (NaH2PO4); completando para um volume de 1000 mL com água destilada] com
agitação e desidratadas em série alcoólica gelada 70%, 85% e 100% durante 5 minutos cada.
Em seguida cada lâmina foi tratada com 90μl da solução de RNAse (5 μL de RNase 10
mg/mL e 975 μL de 2XSSC – cloreto de sódio 0,3M e citrato trisódico 0,03M; pH 6,8) por 1
hora em câmara úmida a 37 ºC, depois lavadas três vezes em 2xSSC e uma vez em PBS 1X
por 5 minutos cada, fixadas por 10 minutos em solução de formaldeído (10 mL de PBS 10x, 5
mL de MgCl2 e 1 mL de formaldeído, completar com água destilada para o volume final de
100 mL). Posteriormente cada lâmina foi lavada novamente em PBS 1X por 5 minutos com
agitação e desidratadas em série alcoólica gelada (70%, 85% e 100%) por 5 minutos. Após
este processo as lâminas foram secas ao ar.
Desnaturação
O DNA cromossômico foi desnaturado em solução de formamida 70% (70 mL de
formamida e 30 mL de 2xSSC) a 72 ºC por 5 minutos, em seguida as lâminas foram
desidratadas em série alcoólica gelada e secas ao ar. Simultaneamente à desidratação em série
alcoólica, a solução de hibridização (6 μL de sonda, 15 μL de formamida, 6 μL de sulfato de
dextrano 50% e 3 μL de 20xSSC) foi desnaturada a 99 ºC por 10 minutos e logo em seguida
colocada no gelo.
Hibridização
Sobre cada lamínula foi colocado 30 μL da solução de hibridização e a lâmina foi
invertida sobre a lamínula. As lâminas, com o material voltado para baixo, foram mantidas
em câmara úmida (2xSSC) a 37 ºC por cerca de 16 horas.
20

LavagensApós a remoção das lamínulas, as lâminas foram colocadas em 2xSSC pH 7,0 a 72 ºC
por 5 minutos, em banho-maria sem agitação e em seguida foram transferidas para o tampão
de bloqueio 1xPBD (20 mL de 20xSSC, 1 mL de TritonX 100 e 1 g de leite em pó desnatado
diluído em água destilada completando para o volume final de 100 mL) à temperatura
ambiente.
DetecçãoSobre cada lamínula foi adicionado 30 μL de avidina-FITC 0,07% em tampão C (0,1 M
de bicarbonato de sódio, pH 8,5 e 0,15 M de NaCl). As lâminas foram invertidas sobre as
lamínulas e colocadas por 30 minutos, em câmara úmida a 37 ºC, e depois lavadas em tampão
a 45 oC, por 5 minutos, por 3 vezes com agitação. Após a lavagem, foi colocado sobre cada
lamínula 20 μL de anti-avidina biotina-conjugada 2,5% (1 μL de anti-avidina estoque em 19
μL de tampão de bloqueio). As lâminas foram invertidas sobre as lamínulas, deixadas em
câmara úmida com 2xSSC a 37 ºC por 10 minutos e lavadas em tampão de bloqueio por 3
vezes (5 minutos cada) com agitação. Novamente foi adicionado sobre cada lamínula a
solução de detecção (30 μL de avidina-FITC 0,07% em tampão C) e a lâmina foi invertida
sobre a lamínula. As lâminas, com o material voltado para baixo, foram mantidas em câmara
úmida 37 ºC por 10 minutos. Em seguida as lamínulas foram retiradas e as lâminas lavadas
em tampão de bloqueio por 3x (5 minutos cada) com agitação.
Montagem das lâminas
A cada lâmina foi adicionado 20 μL de antifading (VectaShield Vector) com 0,7 μL de
solução de iodeto de propídio, na proporção de 50 μL/mL.
2.2.10 Análise cromossômica
As lâminas, contendo as preparações cromossômicas, submetidas às técnicas de
citogenética clássica foram analisadas em microscópio óptico com objetiva de imersão. Já, as
lâminas utilizadas para a FISH foram analisadas em microscópio de fluorescência Olympus
BX51. As imagens foram capturadas através do software Image-PRO MC 6.3.
As melhores metáfases foram editadas no programa Adobe Photoshop CS4. A média do
comprimento total dos cromossomos e da razão entre os braços cromossômicos (q/p) foram
calculadas, com auxílio do programa livre Image J analisando-se três células em metáfase de
dez indivíduos. O cariótipo foi montado pareando os cromossomos com base na razão entre
21

os braços cromossômicos e na posição do centrômero (Levan et al. 1964). Os cromossomos
foram separados em quatro categorias: metacêntrico, submetacêntrico, subtelocêntrico e
telocêntrico, seguindo a metodologia utilizada na maioria das publicações com cetáceos
(Árnason 1974). Para o par portador da região organizadora do nucléolo, a constrição
secundária e os satélites não foram incluídos na medida da relação de braços cromossômicos
(p/q) (Gray et al. 2002).
O idiograma da banda G foi construído com o auxílio do programa Adobe Photoshop
CS4, levando em consideração a intensidade e o tamanho de cada banda.
2.2.11 Caracterização da sondas de DNAr 5S
As sequências de DNAr 5S, utilizadas como sondas para a hibridização homóloga,
foram caracterizadas quanto à sua identidade nucleotídica para confirmar se as sequências
mapeadas correspondiam ao gene de DNAr 5S.
2.2.11.1 Construção de vetores plasmidiais recombinantes contendo fragmentos de DNAr 5S
obtidos por PCR
A construção de vetores plasmidiais recombinantes contendo fragmentos de DNAr 5S
(produtos de PCR) foi realizada para a caracterização destes segmentos de DNA, utilizando o
kit de ligação pGEM-T Easy Vector System I (Promega). Para a ligação dos fragmentos de
interesse ao plasmídeo pGEM-T para cada amostra adicionou-se em um tubo de 1,5 mL: 5 μL
de Tampão, 1 μL de plasmídeo, 3 μL de produto de amplificação e 1 μL de T4 DNA ligase,
sendo esta solução incubada a 4 ºC por 12 horas.
2.2.11.2 Transformação
As células de Escherichia coli DH5α (Promega) foram utilizadas para a transformação e
obtenção dos fragmentos de DNA recombinante. As células competentes foram retiradas do
freezer -70 ºC e colocadas em recipiente com gelo mantido no fluxo laminar para descongelar.
Em um tubo de 1,5 ml com 25 μL de bactérias competentes foram adicionados 5 μL de
plasmídios recombinantes, seguido de leve agitação. A mistura foi deixada no gelo por 30
minutos. Transcorrido este tempo, a amostra de transformação foi colocada em banho-maria a
42 ºC por um minuto, em seguida voltou ao gelo por dois minutos o que possibilitou o choque
térmico e permitiu que os poros da membrana se abrissem mais e internalizassem o DNA
plasmidial.
22

Posteriormente foram adicionados e misturados com muito cuidado 950 μL de meio
líquido LB (peptona 1%; cloreto de sódio 0,17M; extrato de levedura 0,5%, pH 7,5), sendo a
mistura incubada em um agitador (shaker) 225 rpm a 37 ºC por uma hora para que as
bactérias começassem a recuperar a parede celular. Faltando 15 minutos para acabar a
incubação, as placas com meio LB sólido (peptona 1%; cloreto de sódio 0,17M; extrato de
levedura 0,5%, ágar 1,5%, pH 7,5) contendo 50 μL de amplicilina foram retiradas da
geladeira e foram adicionados 35 μL de X-Gal e 35 μL de IPTG , deixando-as semi-abertas.
Ao término da incubação os tubos foram centrifugados por 10 minutos a 10.000 rpm e o
sobrenadante foi descartado por inversão. O meio de cultura líquido (200 μL) restante no tubo
foi utilizado para cuidadosamente ressuspender as bactérias recombinantes que em seguida
foram adicionadas à placa e espalhadas com alça de vidro esterilizada.
Após secas, as placas foram incubadas de maneira invertida em estufa a 37 ºC, cerca de
12 horas para o crescimento das colônias. Cada colônia recombinante (coloração branca) foi
retirada com uma ponteira e colocada em uma solução de PCR padrão (14 μL de água, 5 μL
de dNTP (1 mM), 1 μL de cada primer (5 pmol), 2,5 μL de tampão (10X), 0,19 μL de Taq (5
U/uL), utilizando os primers universais T7F 5’-TAATACGACTCACTATAGGGCGA; SP6R
5’-ATTTAGGTGACACTATAGAATAC -3’, com os seguintes ciclos de amplificação: 3
segundos a 95 ºC (desnaturação), 35 ciclos de 3 segundos a 95 ºC (desnaturação), 1 minuto a
50 ºC (anelamento dos primers), e 2 minutos a 72 ºC (extensão); 5 minutos a. 72 ºC (extensão
final). Depois de amplificado, os produtos da PCR foram verificados e quantificados por
comparação com o marcador Low Mass DNA Ladder (Invitrogen) em eletroforese em gel de
agarose 1,0%, corado com GelRed Acid Gel Stain (Biotium 1:500). A visualização e
documentação dos géis de agarose foram realizadas no fotodocumentador Easy Doc 100
(BioAgency).
Em seguida os produtos de PCR clonados foram purificados utilizando o kit GFX (GE
Healthcare) seguindo protocolo do fabricante, verificados e quantificados por comparação
com o marcador Low Mass DNA Ladder em eletroforese em gel de agarose 1,0%.
2.2.11.3 Sequenciamento das sondas de DNA
Para verificar se o fragmento amplificado por PCR correspondia às regiões de interesse,
o sequenciamento do DNAr 5S foi realizado pelo método de Sanger et al. (1977), com
terminadores marcados com fluorescência. Para a realização das reações de sequenciamento
foi utilizado o kit DYEnamic ET Terminator Cycle Sequencing (GE Healthcare), no qual cada
23

dideoxinucleotídeo (terminador) é marcado com fluoresceína (responsável pela ativação do
segundo elemento) e um dos quatro tipos de rodamina, cuja fluorescência é captada pelo
sequenciador automático de DNA. As reações de sequenciamento foram realizadas em placas
de 96 poços, utilizando-se entre 150 e 300 ng do produto de PCR purificado; 5 pmol de cada
primer em reações separadas; 4 μL do premix (kit) e água mili-Q para completar o volume de
10 μL. As reações de sequenciamento foram feitas no termociclador Applied Biosystems,
Vereti , com o perfil padrão de 30 ciclos (desnaturação a 95 ºC por 20 segundos, anelamento a
50 ºC por 15 segundos e extensão a 72 ºC por 1 minuto e 30 segundos). Os primers utilizados
no sequenciamento foram os mesmos utilizados para o PCR. Depois da reação de
sequenciamento, os fragmentos foram submetidos a um tratamento de precipitação para a
eliminação dos produtos não incorporados durante a reação. Neste processo adicionou-se à
placa 1 μL de acetato de amônia 7,5M (GE Healthcare) e 25 μL de etanol 100% (gelado).
Após 15 minutos em temperatura ambiente, a placa foi centrifugada por 45 minutos a 4000
rpm a 25 ºC. Em seguida foi adicionado à placa 100 μL de etanol 70% recém-preparado e esta
foi centrifugada por 15 minutos a 4000 rpm e colocada para secar protegida da luz por 15
minutos. Posteriormente foi acrescentado 10 μL de formamida (GE Healthcare).
Após a precipitação foi realizada a leitura automática dos fragmentos no sequenciador
automático de DNA MegaBACE 1000 (GE Healthcare), nas condições de injeção e corrida
recomendadas pelo fabricante. As amostras sequenciadas foram salvas (formato abd) e
transferidas do sequenciador para o computador. As sequências geradas foram comparadas
com sequências depositadas no banco de dados público na Internet, GenBank
(http://www.ncbi.nlm.nih.gov), utilizando o programa BLASTN. Este procedimento foi
realizado para determinar se o fragmento sequenciado realmente correspondia ao gene de
interesse. Depois de conferidas, todas as sequências foram alinhadas utilizando-se o programa
de alinhamento múltiplo Clustal W (Thompson et al. 1994), incluso no BioEdit v. 5.0.6 (Hall
1999). Após o alinhamento automático, as sequências foram conferidas e editadas
manualmente no mesmo programa.
24
3. Resultados
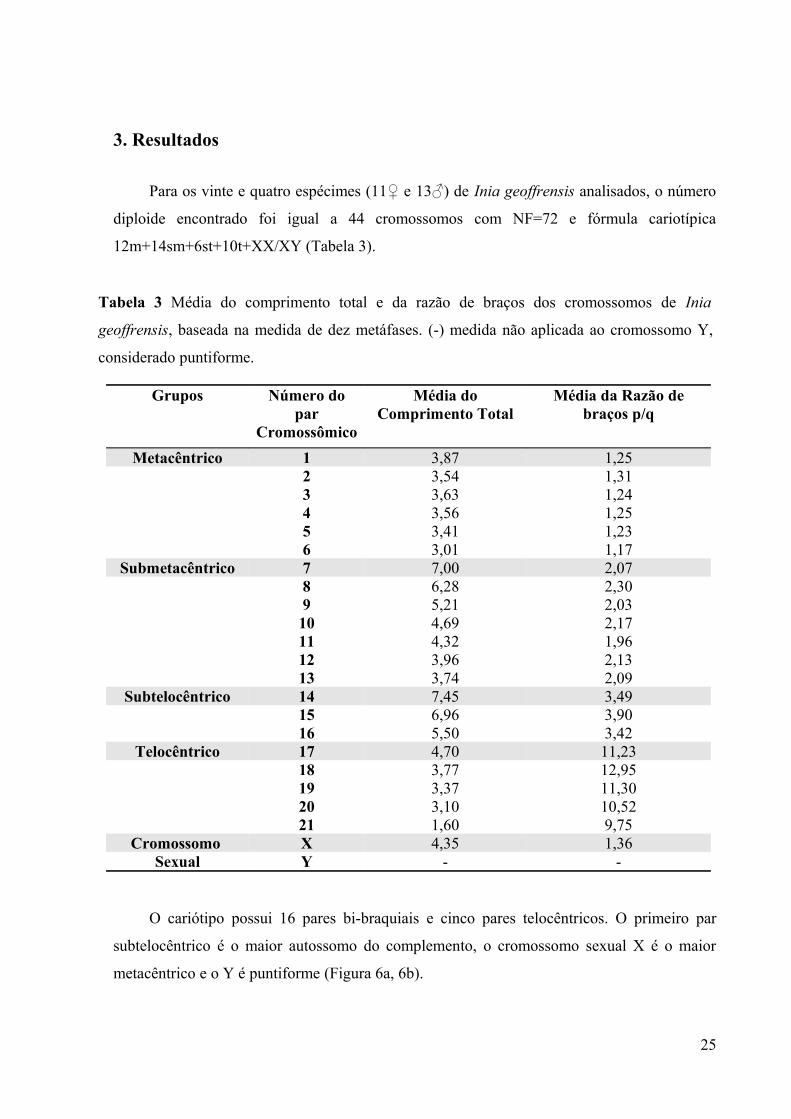
Para os vinte e quatro espécimes (11♀ e 13♂) de Inia geoffrensis analisados, o número
diploide encontrado foi igual a 44 cromossomos com NF=72 e fórmula cariotípica
12m+14sm+6st+10t+XX/XY (Tabela 3).
O cariótipo possui 16 pares bi-braquiais e cinco pares telocêntricos. O primeiro par
subtelocêntrico é o maior autossomo do complemento, o cromossomo sexual X é o maior
metacêntrico e o Y é puntiforme (Figura 6a, 6b).
25
Grupos Número do par
Cromossômico
Média do Comprimento Total
Média da Razão de braços p/q
Metacêntrico 1 3,87 1,252 3,54 1,313 3,63 1,244 3,56 1,255 3,41 1,236 3,01 1,17
Submetacêntrico 7 7,00 2,078 6,28 2,309 5,21 2,0310 4,69 2,1711 4,32 1,9612 3,96 2,1313 3,74 2,09
Subtelocêntrico 14 7,45 3,4915 6,96 3,9016 5,50 3,42
Telocêntrico 17 4,70 11,2318 3,77 12,9519 3,37 11,3020 3,10 10,5221 1,60 9,75
Cromossomo X 4,35 1,36Sexual Y - -
Tabela 3 Média do comprimento total e da razão de braços dos cromossomos de Inia
geoffrensis, baseada na medida de dez metáfases. (-) medida não aplicada ao cromossomo Y,
considerado puntiforme.

Quanto ao padrão de banda C, marcações tênues na região terminal dos braços curtos
foram visualizadas nos pares metacêntricos (exceto no par 2m e 3m). Nos cromossomos
submetacêntricos (pares 7 a 12) e em todos subtelocêntricos foram observadas marcações
terminais conspícuas nos braços curtos. No par 13 esta marcação não foi evidenciada em
todas as metáfases. Já, nos telocêntricos a heterocromatina constitutiva foi visualizada na
região pericentromérica. Os pares 7sm, 8sm e 14st possuem uma marcação proximal no braço
longo (Figura 6c, 6d) Indivíduos de diferentes localidades apresentaram o mesmo padrão de
banda C, com exceção de quatro indivíduos da RDSM, que apresentaram uma variação
interindividual, com uma marcação intersticial em um dos homólogos do par 9sm (Figura 6e).
A região organizadora do nucléolo localizou-se nos braços curtos do menor par
telocêntrico (21t), com a presença de heteromorfismo em relação ao tamanho do sítio (Figura
7b, 7c). A hibridização in situ fluorescente heteróloga corroborou o padrão obtido pela
impregnação com nitrato de prata, confirmando a existência de um único locus deste gene
para I. geoffrensis tanto em machos como em fêmeas (Figura 7a). A RON foi observada em
associação em várias metáfases (Figura 7d, 7g). Além disso, a constrição secundária do par
nucleolar nem sempre foi visualizada em coloração convencional (Figura 7e), só sendo
possível observá-la em metáfases com cromossomos mais distendidos (Figura 7f). O par
nucleolar não mostrou nenhuma marcação para a banda C.
A amplificação do DNAr 5S do boto-vermelho resultou em um fragmento com
aproximadamente 1100 pb, que quando hibridizado revelou resultado similar à hibridização
heteróloga (sondas de Caluromys philander), ambas realizadas em condições de alta
estringência (77%). O DNAr 5S foi detectado em um único par, localizando-se na região
centromérica do quinto par metacêntrico (5m) e na maioria das metáfases analisadas um dos
homólogos foi mais intensamente marcado (Figura 7h). O fragmento de DNAr 5S
amplificado do boto-vermelho foi clonado e sequenciado e a sequência resultante corresponde
ao NTS (espaçador não transcrito) com 620 pb e uma sequência gênica do DNAr 5S com 120
pb. Os clones sequenciados apresentaram 93% de homologia com outras sequências do gene
5S de mamíferos depositadas no GenBank como: Cavia porcellus, Mus musculus e Bos
taurus (Tabela 4).
26

27
Figura 6. a) Cariótipo de um macho Inia geoffrensis após coloração com Giemsa; b)
cromossomos sexuais de uma fêmea; c) cariótipo de uma fêmea após o bandeamento C d)
cromossomos sexuais de um macho; e) par 9sm evidenciando além da marcação terminal, um
bloco de heterocromatina intersticial em um dos homólogos. Barra igual a 10 µm
.
28
Figura 7. a) Localização física cromossômica do DNAr 18S em Inia geoffrensis no par
21t; b) par nucleolar após a impregnação com nitrato de prata; c) heteromorfismo do
sitio da RON; d,g) par nucleolar associado; e) par nucleolar após coloração
convencional mostrando ausência da constrição secundária; f) par nucleolar
evidenciando a constrição secundária; h) Em destaque a localização física
cromossômica do DNAr 5S no par 5m. Barra igual 10µm.
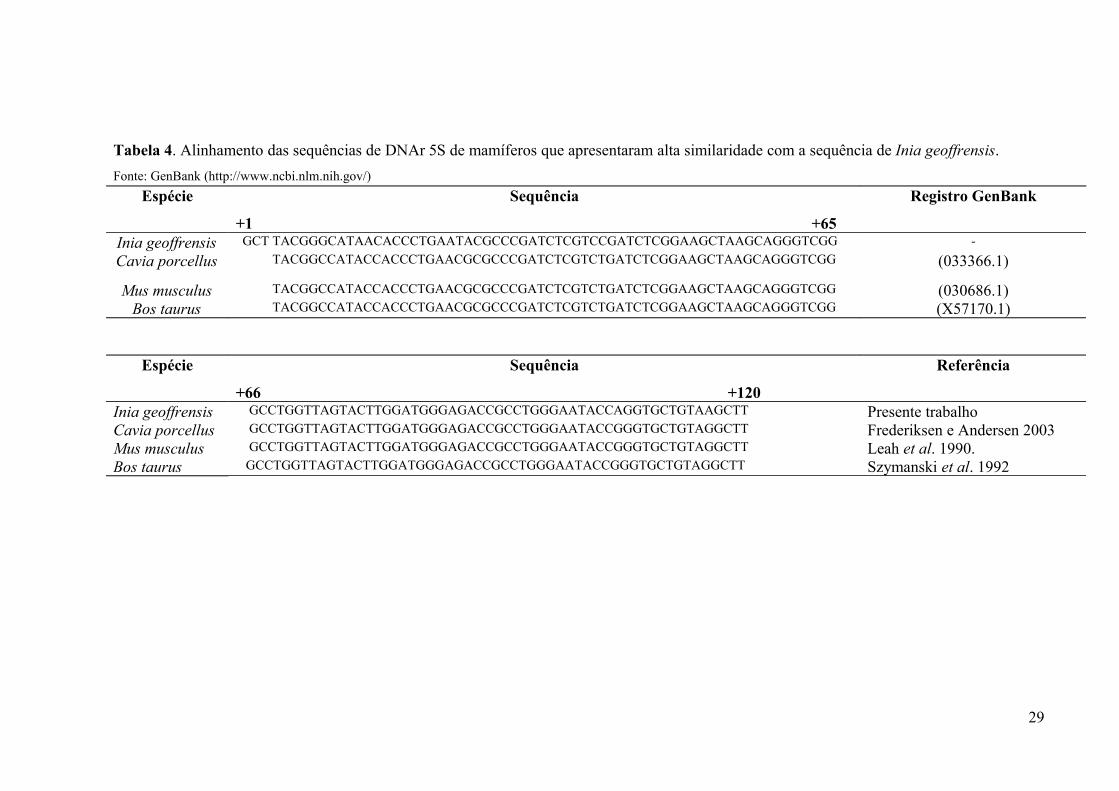
Tabela 4. Alinhamento das sequências de DNAr 5S de mamíferos que apresentaram alta similaridade com a sequência de Inia geoffrensis.
Fonte: GenBank (http://www.ncbi.nlm.nih.gov/)
29
Espécie Sequência
+1 +65
Registro GenBank
Inia geoffrensis GCT TACGGGCATAACACCCTGAATACGCCCGATCTCGTCCGATCTCGGAAGCTAAGCAGGGTCGG -
Cavia porcellus TACGGCCATACCACCCTGAACGCGCCCGATCTCGTCTGATCTCGGAAGCTAAGCAGGGTCGG (033366.1)
Mus musculus TACGGCCATACCACCCTGAACGCGCCCGATCTCGTCTGATCTCGGAAGCTAAGCAGGGTCGG (030686.1)Bos taurus TACGGCCATACCACCCTGAACGCGCCCGATCTCGTCTGATCTCGGAAGCTAAGCAGGGTCGG (X57170.1)
Espécie Sequência
+66 +120
Referência
Inia geoffrensis GCCTGGTTAGTACTTGGATGGGAGACCGCCTGGGAATACCAGGTGCTGTAAGCTT Presente trabalhoCavia porcellus GCCTGGTTAGTACTTGGATGGGAGACCGCCTGGGAATACCGGGTGCTGTAGGCTT Frederiksen e Andersen 2003Mus musculus GCCTGGTTAGTACTTGGATGGGAGACCGCCTGGGAATACCGGGTGCTGTAGGCTT Leah et al. 1990.Bos taurus GCCTGGTTAGTACTTGGATGGGAGACCGCCTGGGAATACCGGGTGCTGTAGGCTT Szymanski et al. 1992

O bandeamento G e a construção do idiograma permitiram o correto pareamento dos
cromossomos homólogos e a identificação do cromossomo sexual X, que possui quatro
bandas conspícuas. O cromossomo Y apresentou uma pequena banda, fracamente corada, em
uma extremidade. Na região centromérica dos pares metacêntricos 2, 3 e 6 foram observadas
marcações positivas para banda G, como também na região pericentromérica do par nucleolar,
entretanto no sítio da RON a banda G mostrou-se negativa. O bandeamento sequencial
(Banda G e Banda C) mostrou um padrão de distribuição da heterocromatina constitutiva
semelhante ao observado em metáfases em que foi realizado somente o bandeamento C,
considerando somente as marcações mais escuras. O cromossomo X apresentou uma
marcação relacionada à banda C intersticial no braço curto, não observada em metáfases
tratadas somente com o bandeamento C. Foi observado também que a heterocromatina na
maioria dos pares cromossômicos está localizada nas regiões de banda G negativas com
exceção dos pares 5m e 16sm (Figura 8a). Além disso, baseado na comparação do padrão de
banda G observou-se quatro pares homeólogos entre o cariótipo do boto-vermelho e do
Tursiops truncatus (Figura 9), e a ocorrência de uma inversão paracentromérica no braço
curto do cromossomo X do boto-vermelho (Figura 10).
A amplificação com os primers para L1 resultou em um fragmento de
aproximadamente 500 pb. A distribuição do LINE1 (L1) ao longo dos cromossomos é
dispersa, mas não é aleatória. Nos cromossomos metacêntricos está na região do centrômero e
também na região proximal do braço longo. Na região centromérica do par 5m o LINE está
colocalizado com o DNAr 5S. Nos maiores cromossomos submetacêntricos subtelocêntricos e
também nos telocêntricos, regiões com sinal de hibridização mais evidente foram observadas
em toda extensão do braço longo intimamente relacionada às regiões de banda G positiva. O
cromossomo X mostrou o maior acúmulo desta sequência, tanto nas regiões proximais dos
braços curtos e longos quanto na região centromérica. No cromossomo Y e no par nucleolar
as marcações foram tênues ou quase ausentes (Figura 8b).
A hibridização em situ fluorescente com a sonda telomérica mostrou a distribuição
esperada das sequências teloméricas, localizadas nas extremidades de todos os cromossomos.
A intensidade do sinal variou em relação aos homólogos e nenhum sitio intersticial foi
detectado (Figura 11).
30

31
Figura 8. a) Padrão de bandeamento G de Inia geoffrensis seguido pelo
ideograma da banda G e bandeamento sequencial G/C. b) Cariótipo de um
macho evidenciando o padrão de distribuição do LINE 1(L1). c) Em destaque o
cromossomo X, em banda G, banda C sequencial e a distribuição do LINE (L1).
Note que no braço curto existe maior acúmulo desta sequência na região de
heterocromatina, na região centromérica e no braço longo na região de banda G
positiva. Barra igual 10µm.

32
Figura 9. Cromossomos homeólogos entre o cariótipo de Inia geoffrensis e Tursiops
truncatus. Os números cromossômicos são apresentados abaixo de cada cromossomo. *
Cromossomos de I. geoffrensis . ** Cromossomos de T. truncatus (CRES 2006). Barra igual
10µm.
Figura 10. Comparação do padrão de banda G do cromossomo X entre cinco espécies de
cetáceos. Eg Eubalaena glacialis baleia-franca-do-norte (Pause et al. 2006) ; Bb Balaenoptera
borealis baleia-sei (Nash 2006); Ig Inia geoffrensis boto-vermelho (presente trabalho); Pp
Phocoena phocoena boto-do-porto (Árnason 1980); Tt Tursiops truncatus golfinho-nariz-
garrafa-comum (CRES 2006). Note no braço curto do cromossomo X do boto-vermelho a
ocorrência de uma inversão paracêntrica. Barra igual 10µm.

33
Figura 11. Cariótipo de um macho do Inia geoffrensis evidenciando a distribuição da
sequência telomérica. Barra igual 10µm.

4. Discussão
4.1- O boto-vermelho dentro da carioevolução dos cetáceos
Quase todas as espécies de golfinho de rio já foram estudadas citogeneticamente, com
exceção do tucuxi Sotalia fluviatilis e do golfinho-do-rio-Ganges Platanista gangetica. Os
golfinhos fluviais como o boto-vermelho Inia geoffrensis, baiji Lipotes vexillifer e toninha
Pontoporia blainvillei compartilham o número diploide de 44 cromossomos, que também é o
padrão predominante para outros cetáceos. Além disso, entre os odontocetos estes golfinhos
são as espécies basais com 2n=44 (Tabelas 1 e 2).
Algumas hipóteses têm sido levantadas sobre o cariótipo ancestral dos cetáceos, entre
estas Jarrel (1979) e Duffield et al. (1991) sugerem que os cariótipos da baleia-da-
Groenlândia Balaena mysticetus e do cachalote-pigmeu Kogia breviceps foram originados a
partir de um cariótipo ancestral com 44 cromossomos, com a fusão de um par telocêntrico
reduzindo o número diploide para 42 cromossomos (Rice 1998). Entretanto, ao se sobrepor os
números diploides descritos para cetáceos na proposta filogenética mais recente (McGowen et
al. 2009) é possível observar que 2n=42 está restrito às espécies das famílias basais, por
exemplo nos misticetos a família Balaenidae (Pause et al. 2006; Jarrel 1979) e entre os
odontocetos, as famílias Physeteridae (Árnason 1981b), Kogiidae (Árnason e Benirschke
1973) e Ziphidae (Hsu e Benirschke 1977; Árnason et al. 1977; Benirschke e Kumamoto
1978).
A hipótese mais parcimoniosa descrita por Heinzelmann et al. (2008), sugere que o
cariótipo ancestral para os cetáceos possuía 42 cromossomos, e que em dois momentos
rearranjos cromossômicos ocorreram independentemente na evolução das duas subordens e
levaram ao aumento do número diploide para 44 cromossomos. Este número diploide é
compartilhado pela linhagem das famílias derivadas nos misticetos: Neobalaenidae e
Balaenopteridae + Eschrichtiidae e nos odontocetos: Lipotidae, Pontoporidae, Iniidae,
Monodontidae, Phocoenidae e Delphinidae (Tabelas 1 e 2).
A comparação das fórmulas cariotípicas mostra a presença de mais pares bibraquiais
(m-sm) em cetáceos com 2n=42, como observado no cachalote Physeter macrocephalus com
19 pares m-sm e nenhum par telocêntrico, em relação ao cariótipo do boto-vermelho com
2n=44, que possui 13 pares m-sm e cinco pares telocêntricos. O que corrobora a hipótese de
que a partir de um cariótipo ancestral com 42 cromossomos, rearranjos cromossômicos do
34

tipo fissão levaram ao aumento do número diploide com mais cromossomos com um braço,
nas espécies com 2n=44 (Heinzelmann et al. 2008).
Kulu et al. (1971) descreveram para o boto-vermelho um cariótipo com 44
cromossomos com 12m+10sm+10st+10t+XY. No presente estudo o número diploide foi
corroborado, entretanto a fórmula cariotípica encontrada foi 12m+14sm+6st+10t+XY, pois
dois pares considerados subtelocêntricos pela autora apresentaram a razão de braços
condizentes a cromossomos submetacêntricos (Tabela 3). Além disso, o cromossomo X que é
enriquecido por sequências de L1 é o maior metacêntrico (Figura 8b) e não submetacêntrico
como proposto anteriormente. Tais divergências podem estar associadas às diferenças do
padrão de condensação em algumas metáfases, como também à realização de medições dos
braços cromossômicos, que fornece maior acurácia na determinação dos tipos
cromossômicos.
A baixa taxa de rearranjos cromossômicos observada em cetáceos é determinada por
características próprias do grupo: como tempo de geração longo com maturidade sexual tardia
e nascimento de somente um filhote por gestação, que somados à alta vagilidade tornam rara a
possibilidade de indivíduos com rearranjos cariotípicos idênticos se encontrarem para serem
fundadores de uma subpopulação com um novo cariótipo (Árnason 1972; Burt et al. 1999;
Fordyce 2009; Kulemzina et al. 2009).
Tais características também são observadas na população do boto-vermelho na
Amazônia Central. Estudos realizados por Martin e da Silva (2004b) demonstraram um sítio
de fidelidade, com aproximadamente 270 animais considerados visitantes e 130 residentes, no
sistema de lagos do Setor Mamirauá. Observaram também que há ocorrência de movimento
desses animais ao longo dos rios, e nenhuma sugestão de isolamento entre indivíduos
residentes de sistemas de lagos diferentes. Desta forma, existe um potencial considerável para
a ocorrência de fluxo gênico entre as áreas, o que pode contribuir para a manutenção da
estabilidade cariotípica nesta espécie.
A diferença observada quanto ao número fundamental e à fórmula cariotípica entre o
boto-vermelho e a toninha, espécies de golfinhos de rio mais intimamente relacionadas do
ponto de vista filogenético e morfológico (Cassens et al. 2000; Hamilton et al. 2001;
McGowen et al. 2009), pode ser resultado de diferentes critérios de medida e organização dos
cromossomos já que para a toninha a existência de cromossomos subtelocêntricos não foi
considerada. Em relação à distribuição da heterocromatina, as duas espécies se diferenciam
35

porque a toninha apresenta alguns pares submetacêntricos com braços longos totalmente
heterocromáticos (Heinzelmann et al. 2008).
Além disso, homeologia cromossômica estabelecida com base no padrão de banda G,
entre quatro pares cromossômicos do boto-vermelho e do golfinho-nariz-de-garrafa-comum
(Figura 9) mostra que pelo menos alguns pares cromossômicos se mantiveram conservados
entre a linhagem que originou uma das espécies de golfinhos fluviais mais antiga I.
geoffrensis e uma das espécies de golfinhos marinhos mais recente da família Delphinidae
Tursiops truncatus (Bielec et al. 1997; Cassens et al. 2000; McGowen et al. 2009). Ambas as
espécies compartilham 2n=44, mas diferenciam-se quanto à fórmula cariotípica, quanto ao par
organizador nucleolar e pela distribuição da heterocromatina na região centromérica (Árnason
1974; Bielec et al. 1997).
Ainda, ao se comparar o padrão de banda G do cromossomo X do boto-vermelho com
os da baleia-franca-do-norte Eubalaena glacialis (Pause et al. 2006), da baleia-sei
Balaenoptera borealis (Nash 2006), do boto-do-porto Phocoena phocoena (Árnason 1980),
do golfinho-nariz-garrafa-comum Tursiops truncatus (CRES 2006) é possível observar que o
o padrão de bandas do X é conservado entre estas espécies de cetáceos, entretanto fica
evidente que no braço curto do cromossomo X do boto-vermelho ocorreu uma inversão
paracêntrica (Figura 10).
4.2 Padrão de Distribuição das Sequências de DNA repetitivo
A heterocromatina é uma estrutura dinâmica que pode permanecer condensada durante
todo o ciclo celular, abrigando sequências de DNA repetitivo, como DNA satélite e elementos
transponíveis ou até mesmo genes, distribuindo-se na região centromérica, telomérica ou
intersticial dos cromossomos (Dilon 2004; Grewal e Jia 2007).
No boto-vermelho, blocos conspícuos de heterocromatina foram evidenciados na região
terminal dos braços curtos de quase todos os pares do complemento. Além disso, os maiores
pares apresentaram blocos heterocromáticos na região intersticial dos braços longos (Figura
6c), como observado no golfinho-pintado-pantropical Stenella attenuata e golfinho-de-
clymene Stenella clymene (Árnason 1980; 2006). No par sexual do boto-vermelho, a
heterocromatina só foi visualizada após o bandeamento sequencial G/C com uma marcação
intersticial no braço longo do cromossomo X e o cromossomo Y não apresentou marcações de
heterocromatina.
36

A heterocromatina centromérica tem um papel fundamental na segregação
cromossômica (Bernard e Allshire 2002), entretanto esta nem sempre é observada em
odontocetos. Por exemplo, a heterocromatina centromérica está ausente no cariótipo do boto-
vermelho (Figura 6c) e do golfinho-pintado-pantropical (Árnason 2006).
Nestas espécies, a região centromérica deve abrigar outras sequências de DNA não
reveladas pelo bandeamento C, por exemplo nos pares 2m, 3m, 5m e 6m do cariótipo do boto-
vermelho a presença do retrotransposon LINE L1 foi evidenciada (Figura 8b). Ainda em
odontocetos, outras espécies como o golfinho-de-bico-branco Lagenorhynchus albirostris
apresentam marcações de heterocromatina centroméricas tênues (Árnason 1980) e somente a
baleia-bicuda-de-Gervais Mesoplodon europaeus apresenta blocos conspícuos de banda C
centromérica (Árnason et al. 1977).
Nas espécies de misticetos das famílias Balaenopteridae e Balaenidae duas sequências
de DNA satélite foram localizadas na heterocromatina: DNA satélite altamente repetitivo rico
em GC, com metade do comprimento desta repetição formada pela sequência TTAGGG,
preferencialmente localizada na região terminal e o DNA satélite, moderadamente repetitivo
rico em AT, limitado aos centrômeros (Árnason et al. 1978; Widegren et al. 1985; Árnason e
Widegren 1989; Adegoke et al. 1993). A ausência destas sequências de DNA satélite em
odontocetos resulta em marcações de banda C menos conspícuas (Árnason 2006) e revela a
presença de diferentes tipos de DNA satélite no genoma dos cetáceos.
Os elementos transponíveis não estão distribuídos aleatoriamente no genoma dos
mamíferos. Por exemplo, as sequências de L1 são mais abundantes nos cromossomos sexuais
(Graham e Boissinot 2005; Acosta et al. 2008) como observado no cromossomo X do boto-
vermelho: com marcações conspícuas na região centromérica, na região de heterocromatina
do braço curto e na região positiva para banda G no braço longo (Figura 8b, 8c). Em alguns
mamíferos o retrotransposon L1 está distribuído somente nas regiões de eucromatina do
cromossomo X, como observado no cariótipo do elefante africano Loxodonta africana, do
peixe-boi marinho Trichechus manatus, do musaranho-elefante Elephantulus edwardii e do
tamanduá-mirim Tamandua tetradactyla (Waters et al. 2004). Já em espécies como o roedor
Microtus agrestis este elemento está acumulado nas regiões de heterocromatina (Acosta et al.
2008).
Em mamíferos, o gene Xist atua na nucleação heterocromática durante a inativação de
um dos cromossomos sexuais em fêmeas, para promover o silenciamento gênico e garantir
dosagem gênica do par sexual quando comparado com os machos. Contudo algumas regiões
37

gênicas são propensas a escapar desta inativação e o L1, induzido pelo Xist facilita a
inativação destes genes por meio da heterocromatinização facultativa ou associação com RNA
de interferência. (Lyon 2000; Graham e Boissinot 2005; Chow et al. 2010).
Nos autossomos do boto-vermelho o L1 está preferencialmente localizado em regiões
eucromáticas, especificamente em regiões de banda G positivas, caracterizadas como ricas em
AT, com replicação tardia, baixo nível de quebras cromossômicas e pobres de genes (Graham
e Boissinot 2005). Esse padrão também é observado nos cromossomos do rato doméstico Mus
musculus domesticus (Boyle et al. 1990, Rebuzzini et al. 2009), do musaranho-elefante
Elephatulus edwardii (Waters et al. 2004) e do morcego Carollia brevicauda (Parish et al.
2002).
Entre os indivíduos estudados neste trabalho, todos apresentaram as mesmas
características cromossômicas, com exceção de quatro espécimes da RSDM que apresentaram
polimorfismo em relação à heterocromatina constitutiva, com um bloco heterocromático
intersticial em heterozigose no par 9sm (Figura 6e). O tamanho e a distribuição dos blocos de
heterocromatina são características instáveis no genoma dos cetáceos onde heteromorfismos
intraespecíficos são bem acentuados, principalmente em relação à heterocromatina intersticial
(Árnason 1974; Árnason 1980; Árnason et al. 1980; 1985). Uma das consequências da
presença de blocos heteromórficos de banda C seria a redução da recombinação meiótica nas
proximidades dessa região, já que diferentes quantidades de DNA altamente repetitivo dos
blocos heteromórficos podem colocar a eucromatina adjacente fora do pareamento
cromossômico durante a meiose (Árnason et al. 1978).
Diversos mecanismos podem contribuir para o surgimento dos polimorfismos de banda
C: heterocromatização das regiões eucromáticas, adição de heterocromatina, emparelhamento
não-homólogo na meiose, resultando em permuta desigual e ação de elementos transponíveis
(Kasahara 2009). Interessantemente, a relação entre o L1 e as sequências de DNA satélite
presentes na heterocromatina já foi estabelecida em cetáceos. Acredita-se que um fragmento
rearranjado de L1 tenha originado o DNA satélite “comum” nos cetáceos que apresenta 63%
de similaridade com este retrotransposon. Este tipo de sequência repetitiva constitui 15% de
todo o genoma dos cetáceos e localiza-se no cariótipo de espécies de odontocetos e misticetos
na heterocromatina intersticial (Widegren et al. 1985; Kapitonov et al. 1998).
Possivelmente para o boto, o L1 tenha contribuído para o polimorfismo observado no
par 9sm já que esta sequência também está localizada intersticialmente neste par (Figura 6b).
Este retroelemento pode ocasionar a permuta desigual entre as repetições de DNA satélite,
38

presentes nas regiões heterocromáticas. Consequentemente, o crossing-over desigual mediado
pelo L1 pode levar à ocorrência de rearranjos cromossômicos deletérios ou ainda ao
remodelamento do genoma de uma determinada espécie (Fitch et al. 1991; Segal et al. 1999;
Graham e Boissinot 2005).
Neste trabalho, a família multigênica de DNA ribossomal 5S foi caracterizada pela
primeira vez no genoma dos cetáceos, sendo que no cariótipo do boto-vermelho foi mapeada
em apenas um locus no par 5m, na posição centromérica (Figura 5i). Diversos estudos estão
sendo realizados para compreender a dinâmica evolutiva desta sequência no genoma dos
vertebrados (Martins e Wasko 2004). Em mamíferos, o DNAr 5S pode estar situado em um
único par cromossômico (Mäkinen et al. 1997; Puerma et al 2008), como observado nos
roedores do gênero Apodemus (Matsubara et al. 2004; Gornung et al 2009) e em cães
(Mäkinen et al. 1997; Pienkowska et al. 2002) ou até em nove pares como detectado no
morcego Rhinolophus hipposideros (Puerma et al. 2008).
Na região centromérica do par 5m estão colocalizados o DNAr 5S e sequências de L1
(Figura 7h e 8b). Em alguns organismos a relação entre estas duas sequências de DNA
repetitivo já foi estabelecida. Muitos pseudogenes de DNAr 5S foram gerados pelo processo
de retrotransposição utilizando as proteínas codificadas pelo L1 (Matsutani 2005). Em peixes,
morcegos e roedores foi descoberto que o DNAr 5S originou famílias de SINEs, um
retrotransposon, com sequências curtas não autônomas (Kapitonov e Jurka 2003; Gogolevsky
et al. 2008 e 2009), que necessitam da maquinaria enzimática do retrotransposon LINE para a
sua transposição no genoma (Kajikama e Okada 2002). Contudo para o boto-vermelho, o
retroelemento L1 parece não influenciar o DNAr 5S pois o número de sítios permanece
constante no cariótipo.
O gene ribossomal 5S do boto-vermelho apresentou 93% de similaridade com outras
espécies de mamíferos, entretanto a sequência do NTS não possui homologia com outras
sequências depositadas no GenBank. Segundo Suzuki et al. (1994), a sequência do espaçador
não transcrito do DNAr 5S não é conservada e varia em relação ao tamanho e composição
nucleotídica entre espécies e Martins e Wasko (2004) sugerem que esta região tem uma
evolução rápida.
Em cetáceos, o conhecimento sobre a organização das regiões organizadoras do
nucléolo ainda é incipiente. No boto-vermelho o sítio de DNAr 18S foi mapeado tanto pela
impregnação com nitrato de prata, quanto pela hibridização in situ fluorescente heteróloga
localizando esta sequência em apenas um par, o 21t, em posição terminal dos braços curtos
39

(Figura 7a,7b). A utilização das duas metodologias se fez necessário, pois o nitrato de prata
impregna apenas as proteínas nucleolares associadas ao RNAr anteriormente transcrito,
revelando apenas as regiões organizadoras do nucléolo que estiveram ativas na última
intérfase (Goodpasture e Bloom 1975; Roussel et al. 1996; Raska et al. 2004), enquanto a
FISH mapeia estes genes no genoma, independente de seu estado de expressão. Em cetáceos
até o momento, apenas o cariótipo do golfinho-nariz-de-garrafa-comum (Tursiops truncatus)
tinha sido mapeado utilizando sondas heterólogas para localizar o DNAr 28S, revelando que
estes genes estão situados em dois pares cromossômicos distintos (Bielec et al. 1997).
A RON localizada somente em um par cromossômico é um caráter ancestral em
mamíferos (Hsu et al. 1975), compartilhado também por várias espécies de odontocetos,
como o boto-vermelho, o golfinho-de-Clymene Stenella clymene, o golfinho-de-bico-branco
Lagenorhynchus albirostris, o boto-do-porto Phocoena phocoena, a beluga Delphinapterus
leucas, o cachalote Physeter macrocephalus e o cachalote-pigmeu Kogia breviceps (Árnason
1980; Árnason 1981a). Entretanto, nos odontocetos rearranjos cromossômicos modificaram o
par nucleolar tanto na baleia-bicuda-de-Gervais Mesoplodon europaeus da família Ziphiidae,
cujo organizador nucleolar está situado em dois pares cromossômicos, como em Delphinidae
na qual se encontram espécies com RON simples e outras com RON múltiplas como a orca
Orcinus orca (Árnason 1981a) e o golfinho-nariz-de-garrafa-comum Tursiops truncatus
(Bielec et al. 1997).
A associação do par nucleolar pelos braços curtos como observada no boto-vermelho
(Figura 7d, 7g) também foi visualizada em espécies das duas subordens de cetáceos (Árnason
1974; Árnason et al. 1978; Bielec et al. 1997). Em 70-80% das metáfases do golfinho-nariz-
de-garrafa-comum, o par nucleolar estava em associação (Duffield et al. 1978), que segundo
Árnason (1981a) ocorre devido a participação destes cromossomos na formação do nucléolo.
A morfologia variável do par nucleolar pode ser devido a um fenômeno epigenético
denominado anfiplastia, que é a habilidade dos cromossomos nucleolares metafásicos
apresentarem-se sob diferentes formas relacionadas à atividade das RONs e determinar a
presença ou não da constrição secundária (Pikaard 2000). Este fenômeno já foi descrito em
plantas como no girassol Helianthus annuus (Schrader et al. 1997), em animais como no
cetáceo Monodon monoceros (Adrews et al. 1973), em peixes como Symphysodon spp (Gross
2009) e no marsupial Caluromys philander (Souza et al. 2009).
A formação da constrição secundária ocorre devido à permanência dos fatores de
transcrição da RNA Polimerase I associados aos genes ativos de RNAr, proporcionando uma
40

estrutura de cromatina diferenciada que resiste à condensação. Os fatores de transcrição
permanecem ligados à RON durante a metáfase, quando a constrição secundária está aparente
e estão ausentes nas RONs inativas, que consequentemente não formam constrição secundária
e não são evidenciadas pela impregnação com nitrato de prata (Sullivan et al. 2001; McStay
2006; Preuss e Pikaard 2007; McStay e Grummt 2008).
O heteromorfismo de tamanho da RON evidenciado por nitrato de prata no par
nucleolar do boto-vermelho pode estar refletindo apenas diferenças na atividade transcricional
do organizador nucleolar, já que após a hibridização in situ fluorescente com a sonda de
DNAr 18S, nenhuma diferença quanto ao tamanho entre os sítios foi observada (Figura 7a, 7b
e 7c). Esta variação na atividade gênica pode revelar necessidades diferentes de RNAr em
algumas células (Zurita et al. 1999), ou que algumas sequências de DNAr estavam silenciadas
anteriormente na intérfase. Alguns estudos têm sugerido que apenas uma fração dos genes de
DNAr é ativa, por exemplo no milho a maioria dos genes de DNAr estão inativos e
densamente compactados em heterocromatina (Pikaard 2000). Desta forma, a associação do
DNAr com a heterocromatina, impediria a recombinação homóloga entre as sequências de
DNAr, contribuindo para manutenção da estrutura nucleolar e estabilidade do DNAr (Carmo-
Fonseca 2000; Sulivan et al. 2001; McStay e Grummt 2008). No cariótipo da maioria dos
cetáceos como o golfinho-de-bico-branco o sítio da RON é totalmente heterocromático
(Árnason 1980; 1981a), entretanto no boto-vermelho o par nucleolar não apresenta blocos de
heterocromatina, o que poderia facilitar a ocorrência de rearranjos cromossômicos neste par
(Figura 6a e 7b).
O presente trabalho foi uma inovação no estudo de mamíferos aquáticos amazônicos,
visto que diferentes marcadores cromossômicos foram utilizados para a descrição cariotípica
do boto-vermelho. O mapeamento de diferentes tipos de DNA repetitivo na população de
boto-vermelho encontrada na RDSM servirá de parâmetros para determinar se existem
populações cariotipicamente diferenciadas ao longo da distribuição desta espécie, com
inferências para o manejo e conservação da mesma. Ainda estudos posteriores poderão revelar
possíveis alterações no genoma desta espécie, em decorrência de modificações no ambiente.
Além disso, a caracterização cromossômica realizada é o passo inicial para a construção de
mapas genéticos do boto-vermelho.
41

5. Conclusões
Com base nos resultados obtidos no presente estudo pode-se concluir que:
• As características cromossômicas do boto-vermelho são similares entre os indivíduos
cariotipados, exceto em relação à distribuição da heterocromatina.
• A localização da heterocromatina no cariótipo do boto-vermelho em relação a outros
cetáceos é um marcador espécie-específico.
• O boto-vermelho compartilha o caráter plesiomórfico com outros cetáceos em relação à
localização da região organizadora do nucléolo em único par cromossômico.
• No cariótipo do boto-vermelho a localização do L1 não é aleatória e é possível que este
retrotransposon esteja envolvido no polimorfismo de banda C, encontrado para quatro
indivíduos da RDSM.
• No cariótipo do boto-vermelho o retrotransposon L1 está colocalizado com DNAr 5S no
par 5m.
• Homeologias cromossômicas observadas pelo bandeamento G entre o boto-vermelho e
Tursiops truncatus revelam que associações sintênicas devem ter sido mantidas durante a
evolução dos golfinhos fluviais e marinhos.
• No cromossomo X do boto-vermelho foi verificada uma inversão paracêntrica no braço
curto.
• Embora a estrutura cariotípica seja considerada conservada em cetáceos, características
cariotípicas do boto-vermelho mostram que variações existem em relação à fórmula
cariotípica, distribuição da heterocromatina e do organizador nucleolar quando comparado
a outras espécies de cetáceos.
42

6. Perspectivas
• Contribuir com mais informações para a resolução da incerteza taxonômica ainda
presente no gênero Inia caracterizando cariotipicamente Inia boliviensis.
• Determinar o papel das sequências de DNA repetitivo na carioevolução dos golfinhos
da Amazônia.
• Determinar se o retroelemento L1 está sendo expresso no genoma do boto-vermelho.
• Determinar quais danos, os poluentes encontrados no ambiente aquático podem causar
no material genético dos golfinhos da Amazônia.
• Ampliar o conhecimento sobre o genoma do boto-vermelho determinando as
associações sintênicas existentes entre o genoma humano e do boto-vermelho.
43

7. Referências Bibliográficas
Acosta, M.J.; Marchal, J.A.; Fernández-Espartero, C.H.; Bullejos, M.; Sánchez, A. 2008.
Retroelements (LINEs and SINEs) in vole genomes: Differential distribution in the
constitutive heterochromatin. Chromosome Research, 16: 949-959.
Adegoke, J.A.; Árnason, U.; Widegren, B. 1993. Sequence organization and evolution, in all
extant whalebone whales, of a DNA satellite with terminal chromosome localization.
Chromosoma, 102: 382-388.
Andersen, L.W.; Friedrich, U. 1988. The karyotype of the long-finned pilot whale,
Globicephala melaena. Hereditas, 109: 245-51.
Andrews, J.C.; Dill, F.J.; Masui, S.; Fisher, H.D. 1973. The chromosome complement of the
Narwhal (Monodon monoceros). Canadian Journal Genetic Cytology, 15: 349-353.
Árnason, U. 1969. The karyotype of the Fin Whale. Hereditas, 62: 273-284.
Árnason, U. 1970. Karyotypes of a male sperm whale (Physeter catodon L.) and a female sei
whale (Balaenoptera borealis Less). Hereditas, 64: 291–293.
Árnason, U. 1972. The role of chromosomal rearrangements in mammalian speciation with
special reference to Cetacean and Pinnipedia. Hereditas, 70: 113-118.
Árnason, U.; Benirschke K. 1973. Karyotypes and idiograms of sperm and pygmy sperm
whales. Hereditas, 75: 7-73.
Árnason, U. 1974. Comparative chromosome studies in Cetacea. Hereditas, 77: 1-36.
Árnason, U.; Benirschke, K.; Mead, J.G.; Nichols, W.W. 1977. Banded karyotypes of three
whales: Mesoplodon europaeus, M. carlhubbsi and Balaenoptera acutorostrata.
Hereditas, 87: 189-200.
Árnason, U. Purdom, I.F.; Jones, K.W. 1978. Conservation and Chromosomal Localization of
DNA Satellites in Balenopterid Whales. Chromosoma, 66: 141-159.
Árnason, U. 1980. C-and G-banded karyotypes of three delphinids: Stenella clymene,
Lagenorhynchys albirostris and Phocoena phocoena. Hereditas, 92: 179-187.
Árnason, U.; Sandholt, B.; Lutley, R. 1980. Banding studies on six killer whales: an account
of C-band polymorphism and G-band patterns. Cytogenet Cell Genetic, 28(1-2): 71-8.
Árnason, U. 1981a. Localization of NORs in cetacean karyotypes. Hereditas, 95: 269-275.
Árnason U. 1981b. Banding studies on the gray and sperm whale karyotypes. Hereditas,
95(2): 277-81.
44

Árnason, U.; Bellamy, H.; Eypórsson, P.; Lutley, R.; Sigurjónsson, J.; Widegren, B. 1985.
Conventionally stained and C-banded karyotypes of a female blue whale. Hereditas, 102:
251-253.
Árnason, U.; Widegren, B. 1989. Composition and chromosomal localization of cetacean
highly repetitive DNA with special reference to the blue whale, Balaenoptera musculus.
Chromosoma, 98: 323-329.
Árnason, U. 2006. Order Cetartiodactyla: Cetacea. p 541-546. In: O’Brien, S.J.; Menninger,
J.C.; Nash, W.G. (Eds). Atlas of Mammalian Chromosomes. Wiley-Liss: USA.
Asahida, T.; Kobayashi, T.; Saitoh, K.; Nakayama, I. 1996. Tissue preservation and total
DNA extraction from fish stored at ambient temperature using buffers containing high
concentration of urea. Fisheries Science, 62(5): 727-730.
Assis, M.F.L.; Best, R.C.; Barros, R.M.S.; Yonenaga-Yassuda, Y. 1988. Cytogenetic study of
Trichechus inunguis (Amazonian manatee). Brazilian Journal of Genetics, 11: 41-50.
Banguera-Hinestroza, E.; Cárdenas, H.; Ruiz-Garcia, M.; Marmontel, M.; Gaitán, E.;
Vázquez, R.; Garcia-Vallejo, F. 2002. Genetic differentiation in populations of the river
dolphins genus Inia (Blainville, 1817) and apports to the taxonomic status with
molecular analysis. Revista de la Academia Colombiana de Ciencias Exatas, Físicas y
Naturales, 26: 575-588.
Benirschke, K.; Kumamoto, A. 1978. The chromosomes of Cuvier’s beaked whale, Ziphius
cavirostris. Mammal Chromossome Newsletter, 19: 70-72.
Bernard, C.; Allshire, R.C. 2002. Centromeres become unstuck without heterochromatin.
Trends in Cell Biology, 12: 419-424.
Best, R.C.; da Silva, V.M.F. 1989. Amazon river dolphin, boto Inia geoffrensis (de Blainville,
1817). p. 1-23. In: Ridgway, S.H.; Harrison, R.J. (Eds.). Handbook of Marine Mammals.
Volume 4. River Dolphins and the Larger Toothed Whales. Academic Press. London.
Best, R.C.; da Silva, V.M.F. 1993. Inia geoffrensis. Mammalian Species, 426: 1-8.
Bielec, P.E.; Riggs, P.K.; Womack, J.E.; Busbee, D.L. 1997. A high resolution GBG-banded
karyotype of the Atlantic bottlenose dolphin, Tursiops truncatus: generation of an
ideogram, and NOR localization by fluorescence in situ hybridization. Cytogenetic Cell
Genetic, 78: 6-11.
Bielec, P.E.; Gallagher, D.S.; Womack, J.E.; Busbee, D.L. 1998a. Homologies between
human and dolphin chromosomes detected by heterologous chromosome painting.
Cytogenetic Cell Genetic, 81: 18–26.
45

Bielec, P.E.; Riggs, P.K.; Womack, J.E.; Taylor, J.; Davis, S.; Busbee, D.L. 1998b. Dolphin
interleukin-8 receptor Map position 18q25-26. Chromosome Research, 6: 661.
Bielec, P.E.; Riggs, P.K.; Womack, J.E.; Taylor, J.; Davis, S.; Busbee, D.L. 1998c. Dolphin
β-glucocerebrosidase, Map position 1q22. Chromosome Research, 6: 425.
Bielec, P.E.; Gallangher, D.; Yang, Y.P.; Womack, J.; Davis, S.; Taylor, S.; Busbee, D. 2000.
Assignment of crystallin beta-polypeptide 1 (CRYBA1) to Atlantic bottlenose dolphin
chromosome band 16p11 by in situ hybridization. Cytogenetic Cell Genetic, 89: 96-97.
Biémont, C.; Vieira, C. 2006. Junk DNA as an evolutionary force. Nature, 443: 521-524.
Blackburn, E.H. 2005. Telomeres and telomerase: their mechanisms of action and the effects
of altering their functions. Federation of European Biochemical Societies Letters, 579:
859-862.
Boyle, A.L.; Ballard, S.G.; Ward, D.C. 1990. Differential distribution of long and short
interspersed element sequences in the mouse genome: Chromosome karyotyping by
fluorescence in situ hybridization. Proceedings of the National Academy of Sciences of
the United States of America, 87: 7757-7761.
Burt, D.W.; Bruley, C.; Dunn, I.C.; Jones, C.T.; Ramage, A.; Law, A.S.; Morrice, D.R.;
Patton, I.R.; Smith, J.; Windsor, D.; Sazanov, A.; Ruedi, F.; Waddington, D. 1999. The
dynamics of chromosome evolution in birds and mammals. Nature, 402: 412-413.
Carmo-Fonseca, M.; Mendes-Soares, L.; Campos, I. 2000. To be or not to be in the nucleolus.
Nature Cell Biology, 2: 107-112.
Carr, D.H.; Sinoh, R.P.; Miller, I.R. ; McGeer, P.L.1966. The chromosome complement of the
Pacific killer whale (Orcinus rectipinna). Mammal Chromosome Newsletter, 22: 208.
Cassens, I.; Vicario, S.; Waddell, V.G.; Balchowsky, H.; Belle, D.V.; Ding, W.; Fan, C.;
Mohan, R.S.L.; Simões-Lopesi, P.C.; Bastida, R.; Meyer, A.; Stanhope, M.J.;
Milinkovitch, M.C. 2000. Independent adaptation to riverine habitats allowed survival of
ancient cetacean lineages. Proceedings National Academy of Science USA, 97: 11343–
11347.
Castiglia, R.; Garagna, S.; Merico, V.; Oguge, N.; Corti, M. 2006. Cytogenetics of a new
cytotype of African Mus (subgenus Nannomys) minutoides (Rodentia, Muridae) from
Kenya: C- and G- banding and distribution of (TTAGGG)n telomeric sequences.
Chromosome Research, 14: 587-594.
Charlesworth, B.; Sniegowski, P.; Wolfgang, S. 1994. The evolutionary dynamics of
repetitive DNA in eukaryotes. Nature, 371: 215:220.
46

Christensen, K.; Lomholt, B.; Hallenberg, C.; Nielsen, K.V. 1998. Mink 5S rRNA genes map
to 2q in three loci suggesting conservation of synteny with human 1q. Hereditas, 128:
17-20.
Chow, J.C.; Ciaudo, C.; Fazzari, M.J.; Mise, N.; Servant, N.; Glass, J.L.; Attreed, M.; Ayner,
P.; Wutz, A.; Barillot, E.; Greally, J.M.; Voinnet, O.; Heard, E. 2010. LINE-1 Activity in
Facultative Heterochromatin Formation during X Chromosome Inactivation. Cell, 141:
956-969.
CRES (Center for Reprodution of Endangered Species), 2006. Tursiops truncatus, p. 552. In:
O’Brien, S.J.; Menninger, J.C.; Nash, W.G. (Eds). Atlas of Mammalian Chromosomes.
Wiley-Liss: USA.
da Silva, V.M.F. 1994. Aspects of the biology of the Amazonian dolphin genus Inia and
Sotalia fluviatilis. PhD dissertation, Cambridge University, Cambridge: 327 pp.
da Silva, V.M.F.; Best, R.C. 1996. Freshwater dolphin/fisheries interactions in the Amazon
region (Brazil). Amazoniana, 14: 165-175.
da Silva, V.M.F.; Martin, A.R. 2000. A study of the boto, or Amazon river dolphin (Inia
geoffrensis), in the Mamirauá Reserve, Brazil: operation and techniques. In: Reeves, R.R.;
Smith, B.D.; Kasuya, T. (Eds). Biology and Conservation of Freshwater Cetaceans in
Asia. IUCN, 23: 121-131.
da Silva, V.M.F.; Martin, A.R. 2007. Impact of human activities upon two species of dolphins
in Amazon flooded forest, Brazil. 17th Biennial Conference on the Biology of Marine
Mammals. Cape Town, S. Africa.
da Silva, V.M.F.; Goulding, M.; Barthem, R. 2008. Golfinhos da Amazônia. INPA, Manaus,
Brasil. 43 pp.
da Silva, V.M.F. 2009. Amazon River Dolphin Inia geoffrensis. In: Perrin, W.F.; Wursig, B.;
Thewissen, J.G.M. (Eds) Encyclopedia of Marine Mammals, Elsevier, 26-28.
da Silva, V.M.F.; Martin, A.R. 2010. Status threats, conservation initiatives and possible
solutions for Inia geoffrensis and Sotalia fluviatilis in Brazil. In: Trujillo, F.; Crespo, E.
Damme, P.A.; Usma, J.S. (Eds) The Action Plan for South American River Dolphins
2010-2020, Solomac, 123-143.
Di Beneditto, A.P.M.; Siciliano, S.; Ramos, R.M.A. 2010. Cetáceos: introdução à biologia e
metodologia básica para o desenvolvimento de estudos. Fundação Oswaldo Cruz, Escola
Nacional de Saúde Pública, Rio de Janeiro. 13 p.
Dillon, N. 2004. Heterochromatin structure and function. Biology of the Cell, 96: 631-637.
47

Duffield, D.A; Ridgway, S.H.; Parkes, R.S. 1967. Cytogenetic studies of two species of
porpoise. Nature, 213: 189-190.
Duffield, D.A.; Spence, M.A.; Paquin, C. 1978. Evaluation of metaphase association patterns
for acrocentric chromosomes in the order Cetacea. Cytologia, 43: 541-549.
Duffield, D.A.; Chamberin-Lea, J.; Sweenwy, J.C.; Odell, D.K.; Asper, E.D.; Gilmartin, W.G.
1991. Use of corneal cell culture for R-Band chromosome studies on stranded cetaceans.
Marine Mammal Strandings, 98: 91-100.
Esnault, C.; Maestre, J.; Heidmann, T. 2000. Human LINE retrotransposons generate
processed pseudogenes. Nature, 24: 363-367.
Estoup, A.; Presa, P.; Krieg, F.; Vaiman, D.; Guyomard, R. 1993. (CT)n and (GT)n
microsatellites: a new class of genetic markers for Salmo truta L. (brown trout). Heredity,
71: 488-496.
Evgen’ev, M.B. 2007. Mobile elements and genome evolution. Molecular Biology, 41(2):
203-213.
Feschotte, C.; Pritham, E.J. 2007. DNA Transposons and the Evolution of Eukaryotic
Genomes. Annual Reviews Genetic, 41: 331-68.
Fitch, D.H.A.; Bailey, W.J.; Tagle, D.A.; Goodman, M.; Sieut, L.; Slightom, J.L. 1991.
Proceedings of the National Academy of Sciences of the United States of America, 88:
7396-7400.
Fordyce, R.E.; Barnes, L.G. 1994. The evolutionary history of whales and dolphins. Annual
Review Earth Planet Science, 22: 419-55.
Fordyce, R.E. 2009. Cetacean Evolution. In: Perrin, W.F.; Wursig, B.; Thewissen, J.G.M.
(Eds) Encyclopedia of Marine Mammals, Elsevier,201-207.
Franco-de-Sá, J.F.O.; Rosas, F.C.W.; Feldberg, E. 2007. Cytogenetic study of the giant otter
Pteronura brasiliensis Zimmermann 1780 (Carnivora, Mustelidae, Lutrinae). Genetics
and Molecular Biology, 30: 1093-1096.
Frederiksen, S.; Andersen, J.H. 2003. The external promoter in the guinea pig 5S rRNA gene
is different from the rodent promoter. Hereditas, 139: 156-160.
Freitas, T.R.O.; Mattevi, M.S.; Oliveira, L.F.B. 1975. Karyotype characterization of lontra
(Lutra longicaudis longicaudis, Mustelidae, Carnivora). Mammalian Chromosomes
Newsletter, 23: 91-96.
48

Gray, B. A.; Zori, R.T.; McGuire, P.M.; Bonde, R.K. 2002. A first generation cytogenetic
ideogram for the Florida manatee (Trichechus manatus latirostris) based on multiple
chromosome banding techniques. Hereditas, 137: 215-223.
Gentles, A.J.; Wakefield, M.J.; Kohany, O.; Gu, W.; Batzer, M.A.; Pollock, D.D.; Jurka, J.
2007. Evolutionary dynamics of transposable elements in the short-tailed opossum
Monodelphis domestica. Genome Research, 17: 992-1004.
Gogolevsky, K.P.; Vassetzky, N.S.; Kramerov, D.A. 2008. Bov-B-mobilized SINEs in
vertebrate genomes. Gene, 407: 75–85.
Gogolevsky, K.P.; Vassetzky, N.S.; Kramerov, D.A. 2009. 5S rRNA-derived and tRNA-
derived SINEs in fruit bats. Genomics, 93: 494-500.
Goodpasture, C.; Bloom S.E. 1975. Visualization of nucleolar organizer regions in
mammalian chromosomes using silver staining. Chromosoma, 53: 37-50.
Gornung, E.; Cristaldi, M.; Castiglia, R. 2009. Comparative cytogenetic analysis of the
“Sylvaemus”group of Apodemus (Rodentia, Muridae): A. sylvaticus from Sicily and A.
flavicollis from the central Apennines. Acta Theriologica, 54: 267–275.
Grabert, H. 1984. Migration and speciation of the South American Iniidae (Cetacea-
Mammalia) Zeitschrift Saugetierkunde. 49(6): 334-341.
Graham, T.; Boissinot, S. 2005. The Genomic Distribution of L1 Elements: The Role of
Insertion Bias and Natural Selection. Journal of Biomedicine and Biotechnology, 2006:
1–5.
Gravena, W.; Hrbek, T.; da Silva, V.M.F.; Astolfi-Filho, S.; Farias, I.P. 2009. Microsatellite
loci for population and parentage analysis in the Amazon River dolphin (Inia geoffrensis
de Blainville, 1817). Molecular Ecology Resources, 9: 600-603.
Grewal, S.I.S.; Jia, S. 2007. Heterochromatin revisited. Nature, 8: 35-46.
Gross, M.C. 2009. Citogenética comparativa das variedades selvagens de acará-disco
(Symphysodon spp., Cichlidae, Perciformes) endêmicos da Amazônia: uma abordagem
clássica e molecular dos cromossomos mitóticos e meióticos. Tese de doutorado.
Instituto Nacional de Pesquisas da Amazônia. Manaus/Amazonas. 114pp.
Gross, M.C.; Schneider, C.H.; Valente, G.T.; Martins, C.; Feldberg, E. 2010. Variability of
18S rDNA locus among Symphysodon fishes: chromosomal rearrangements. Journal of
Fish Biology, 76: 1117–1127.
Hall, T.A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis
program for Windows 95/96/NT. Nucleic Acids Symposium Series, 41: 95-98.
49

Hamilton, H.; Caballero, S.; Collins, A.G.; Brownell Jr., R.L. 2001. Evolution of river
dolphins. Proceedings Royal Society of London B, 268: 549-556.
Heinzelmann, L.; Telles, P.C.C.; Danilewicz, D.; Chies, J.A.B.; Andrades-Miranda, J. 2008.
The karyotype of franciscana dolphin (Pontoporia blainvillei). Journal of Heredity,
100(1): 119–122.
Horrall, J.F.; Taylor, B.K.; Taylor, K.M. 1968. Karyotypes of the Pacific killer whale,
Orcinus orca (Linnaeus 1758) and the Brazilian Tapir, Tapirus terrestris (Gray 1843).
Mammal Chromosomes Newsletter, 9: 244-245.
Howell, W.M.; Black, D.A. 1980. Controlled silver-staining of nucleolus organizer regions
with a protective colloidal developer: a 1-step method. Experientia, 36: 1014-1015.
Hsu, T.C.; Spirito, S.E.; Pardue, M.L. 1975. Distribution of 18S+28S Ribosomal Genes in
Mammalian Genomes. Chromosoma, 53: 25-36.
Hsu, T.C.; Benirschke, K.; 1977. An Atlas of Mammalian Chromosomes. New York,
Springer-Verlag, 10: 486.
IBAMA. 2001. Mamíferos Aquáticos do Brasil: Plano de Ação. Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis. 2: 1-102.
Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. 1991. Improved telomere detection using a
telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Research. 19:
4780.
IUCN 2010. Cetacean update of the 2010 IUCN (International Union for Conservation of
Nature) Red List of Threatened Species. Disponível em: <www.iucnredlist.org>. Acesso
em: 06/03/2011.
Jarrell, G.H. 1979. Karyotype of the bowhead whale (Balaena mysticetus). Journal Mammal,
60: 607-610.
Jarrell, G.H.; Árnason, U. 1981. Banded karyotypes of a belukha whale, Delphinapterus
leucas. Hereditas, 95: 37-41.
Kajikama, M.; Okada, N. 2002. LINEs mobilize SINEs in the Eel through a shared
3’sequence. Cell, 111: 433–444.
Kapitonov, V.V.; Holmquist, G.P.; Jurka, J. 1998. L1 repeat is a basic unit of heterochromatin
satellites in Cetaceans. Molecular Biology and Evolution 15(5): 611–612.
Kapitonov, V.V.; Jurka, J. 2003. A novel class of SINE elements derived from 5S rRNA.
Molecular Biology Evolution, 20: 694-702.
50

Kasahara, S. 2009. Introdução à Pesquisa em Citogenética de Vertebrados. Sociedade
Brasileira de Genética, Ribeirão Preto, Brasil. 160 pp.
Kasuya, T. 1966. Karyotype of a sei whale. Scientific Reports of the Whales Research
Institute, 20: 83-88.
Komiya, H.; Takemura, S. 1979. Nucleotide sequence of 5S ribosomal RNA from rainbow
trout (Salmo gairdnerii) liver. The Journal of Biochemistry, 86: 1067-1080.
Kulemzina, A.I.; Trifonoy, V.A.; Perlman, P.L.; Rubtsova, N.V.; Volobuey,V.; Ferguson-
Smith, M.A.; Stanyon, R. Yang, F.; Graphodatsky. 2009. Cross-species chromosome
painting in Cetartiodactyla: Reconstructing the karyotype evolution in key phylogenetic
lineages. Chromosome Research, 17: 419-436.
Kulu, D.D.; Veomett, I.; Sparkes, R.S. 1971. Cytogenetic comparison of four species of
cetaceans. Journal of Mammalogy, 52: 828-832.
Kulu, D.D.1972. Evolution and cytogenetics. p. 503-527. In: Thomas, C.C. (Org.) Mammals
of the Sea- Biology and Medicine. Bannerstone House, Illinous.
Lailson-Brito Jr, J.; Dorneles, P.R.; da Silva, V.M.F.; Martin, A.R.; Bastos, W.R.; Azevedo-
Silva, C.E.; Azevedo, A.F.; Torres, J.P.M.; Malm, O. 2008. Dolphins as indicators of
micropollutant trophic flow in Amazon Basin. Oecologia Brasiliensis, 12: 531-541.
Leah, R.; Frederihsen, S.; Engberg, J.; Sorensen, P.D.; 1990. Nucleotide sequence of a mouse
5S rRNA variant gene. Nucleic Acids Research, 18(24): 7441.
Lee, S.H.; Cho, S.Y.; Shannon, M.F.; Fan, J.; Rangasamy, D. 2010. The impact of CpG Island
on defining transcriptional activation of the mouse L1 retrotransposable elements. Plos
One, 5: 1-14.
Levan, A.; Fredga, K.; Sandberg, A. 1964. Nomenclature for centromeric position on
chromosomes. Hereditas, 52: 201-220.
Loch, C.; Marmontel, M.; Simões-Lopes, P. 2009. Conflicts with fisheries and intentional
killing of freshwater dolphins (Cetacea: Odontoceti) in the Western Brazilian Amazon.
Biodiversity and Conservation, 18: 3979-3988.
Long, E.O.; Dawid, I.B. 1980. Repeated genes in eukaryotes. Annual Review of Biochemistry,
49: 727-764.
Lundberg, J.G.; Marshall, L.G.; Guerrero, J.; Horton, B.; Malabarba, M.C.S.L.; Wesselingh,
F. 1998. The stage for Neotropical fish diversification: A history of a tropical South
American rivers. p 13-48. In: Malabarba, L.R.; Reis, R.E.; Vari, R.P.; Lucena, Z.M.S.;
51

Lucena, C.A.S. (eds). Phylogeny and classification of Neotropical fishes.EDIPUCRS,
Porto Alegre, RS.
Lyon, M.F. 2000. LINE-1 elements and X chromosome inactivation: A function for ‘‘junk’’
DNA? Proceedings of the National Academy of Sciences of the United States of America,
97: 6248-6249.
Lyon, M.F. 2005. No longer ‘all-or-none’. European Journal of Human Genetics, 13: 796-
797.
Machado, T.; Silva, M.J.J.; Leal-Mesquita, E.R.; Carmignotto, A.P.; Yonenaga-Yassuda, Y.
2005. Nine karyomorphs for spiny rats of the genus Proechimys (Echimyidae, Rodentia)
from North and Central Brazil. Genetics and Molecular Biology, 28: 682-692.
Mäkinen, A.; Zijlstra, C.; de Haan, N.A.; Mellink, C.H.M.; Bosma, A.A. 1997. Localization
of 18S+28S and 5S ribosomal RNA genes in the dog by fluorescence in situ
hybridization. Cytogenetics and Cell Genetics, 78: 231-235.
Makino, S. 1948. The chromosomes of the Dall’s porpoise, Phocoenoides dalli (True) with
remarks on the phylogenetic relations of the Cetacean. Chromosoma, 3: 220-231.
Martin, A.R.; da Silva, V.M.F. 2004a. River dolphins and flooded forest: seasonal habitat use
and sexual segregation of botos (Inia geoffrensis) in an extreme cetacean environment.
Journal of Zoology London, 263: 295-305.
Martin, A.R.; da Silva, V.M.F. 2004b. Number, seasonal movements, and residency
chraracteristics of river dolphins in an Amazonian floodplain lake system. Canadian
Journal of Zoology, 82: 1307-1315.
Martins, C.; Cabral-de-Mello, D.C.; Targino, G.V.; Mazzuchelli, J.; Oliveira, S.G. 2010.
Cytogenetic mapping and contribution to the knowledge of animal genomes. In: Urbano,
K.V. (eds) Advances in Genetics Research. Vol. 4. Nova Science Publishers (in press).
Martins, C.; Wasko, A.P. 2004. Organization and evolution of 5S ribosomal DNA in the fish
genome. p. 289-318. In: Williams, C.R. (eds). Focus on Genome Research. Nova
Science Publishers, Hauppauge, NY, USA.
Matsubara, K.; Nishida-Umehara, C.; Tsuchiya, K.; Nukaya, D.; Matsuda, Y. 2004.
Karyotypic evolution of Apodemus (Muridae, Rodentia) inferred from comparative FISH
analyses. Chromosome Research, 12: 383–395.
Matsutani, S. 2005. Links between repeated sequences. Journal of Biomedicine and
Biotechnology, 2006: 1–3.
52

McEachern, M.J.; Krauskopf, A.; Blackburn, E.H. 2000. Telomeres and their control. Annual
Review of Genetics, 34: 331-358.
McGowen, M.R.; Spaulding, M.; Gatesy, J. 2009. Divergence date estimation and a
comprehensive molecular tree of extant cetaceans. Molecular Phylogenetics and
Evolution, 53: 891-906.
McStay, B. 2006. Nucleolar dominance: a model for rRNA gene silencing. Genes &
Development, 20: 1207–1214.
McStay, B.; Grummt, I. 2008. The Epigenetics of rRNA Genes: From Molecular to
Chromosome Biology. The Annual Review of Cell and Developmental Biology, 24: 131–
57.
Minrong, C.; Hanqin, L.; Zhimei, G. 1986. The karyotype and the c-band patterns of the Baiji
dolphin, Lipotes vexillifer. Acta Hydrobiologica Sinica, 20(2): 138-143.
Moorhead, P.S.; Nowell, P.C.; Mellinan, W.J.; Battips, D.M.; Hungerford, D.A. 1960.
Chromosome preparations of leukocytes cultured from human peripheral blood.
Experimental Cell Research, 20: 613-616.
Nash, W.G. 2006. Balaenoptera borealis, p. 561. In: O’Brien, S.J.; Menninger, J.C.; Nash,
W.G. (Eds). Atlas of Mammalian Chromosomes. Wiley-Liss: USA.
Nigumann, P.; Redik, K.; Mätlik, K.; Speek, M. 2002. Many human genes are transcribed
from the antisense promoter of L1 retrotransposon. Genomics, 79: 628-634.
O’Donnell, K.A.; Burns, K.H. 2010. Mobilizing diversity: transposable element insertions in
genetic variation and disease. Mobile DNA, 1: 1-10.
Parish, D.A.; Vise, P.; Wichman, H.A.; Bull, J.J.; Baker, R.J. 2002. Distribution of LINEs and
other repetitive elements in the karyotype of the bat Carollia: implications for X-
chromosome inactivation. Cytogenetic and Genome Research, 96: 191-197.
Pause, K.C.; Bonde, R.K.; Mcguire, P.M.; Zori, R.T.; Gray, B.A. 2006. G-Banded karyotype
and ideogram for the North Atlantic Right Whale (Eubalaena glacialis). Journal of
Heredity, 97(3): 303-306.
Peng, X.; Chen, J. 1985. The study on karyotype of finless porpoise (Neophocaena
phocaenoides). Acta Theriologica Sinica, 5: 161–165.
Perrin, W.F.; Wursig, B.; Thewissen, J.G.M. (Eds). 2009. Marine Mammal Species.
Encyclopedia of Marine Mammals. Elsevier, New York, 2: 1259-1261.
53

Pienkowska, A.; Schelling, C.; Opiola, T.; Rozek, M.; Barciszewski, J. 2002. Canine 5S
rRNA: nucleotide sequence and chromosomal assignment of its gene cluster in four
canid species. Cytogenetic and Genome Research, 97: 187-190.
Pikaard, C.S. 2000. The epigenetics of nucleolar dominance. Trends in Genetics, 16(11): 495-
500.
Pilleri, G.; Gihr, M. 1977. Observations on the Bolivian, Inia boliviensis, (D’Orbigny, 1834)
and the Amazonian bufeo, Inia geoffrensis (Blainville, 1817), with a description of a new
subespecies (Inia geoffrensis humboldtiana). Investigations on Cetacea, 8: 11-76.
Pinkel, D.; Straume, T.; Gray, J.W. 1986. Cytogenetic analysis using quantitative, high-
sensitivity, fluorescence hybridization. Proceedings National Academy of Science Usa,
83: 2934-2938.
Prasad, N.; Numford, D.M.; Barsales, P.B.; Whithman, T.; Wilbur, J.R. 1970. Cytogenetic
studies of dolphin (Tursiops truncatus) by an extended tissue culture technique.
Experientia, 26: 1167-1168.
Preuss, S.; Pikaard, C.S. 2007. rRNA gene silencing and nucleolar dominance: Insights into a
chromosome-scale epigenetic on/off switch. Biochimica et Biophysica Acta, 1769: 383–
392.
Puerma, E.; Acosta, M.J.; Barragán, M.J.L.; Martínez, S.; Marchal, J.A.; Bullejos, M.;
Sánchez, A. 2008. The karyotype and 5S rRNA genes from Spanish individuals of the
bat species Rhinolophus hipposideros (Rhinolophidae; Chiroptera). Genetica, 134: 287–
295.
Raska, I.; Koberna, K.; Malinsky, J.; Fidlerova, H.; Masata, M. 2004. The nucleolus and
transcription of ribosomal genes. Biology of the Cell, 96: 579–594.
Rebuzzini, P.; Castiglia, R.; Negardze, S.G.; Mitsainas, G.; Munclinger, P.; Zuccotti, M.;
Capanna, E.; Redi, C.A.; Garagna, S. 2009. Quantitative variation of LINE-1 sequences
in five species and three subspecies of the subgenus Mus and in five Robertsonian races
of Mus musculus domesticus. Chromosome Research, 17: 65-76.
Rice, D.W. 1998. Marine mammals of the world - Systematics and Distribution. Society
for Marine Mammalogy: Lawrence. 231pp.
Rosas, F.C.W.; Lehti, K.K. 1996. Nutritional and mercury content of milk of the Amazon
River dolphin, Inia geoffrensis. Comparative Biochemistry Physiology, 115A(2): 117-
119.
54

Roussel, P.; André, C.; Comai, L.; Hernandez-Verdum, D. 1996. The rDNA transcription
machinery is assembled during mitosis in active NORs and absent in inactive NORs. The
Journal of Cell Biology, 133: 235-246.
Rubes, J.; Musilova, P.; Mastromonaco, G.F. 2008. Cytogenetic of wild and captive bred non-
domestic animals. Cytogenetic Genome Research, 120: 61-68.
Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K. B.;
Erlich, H.A. 1988. Primer-directed enzymatic amplification of DNA with a thermostable
DNA polymerase. Science, 239: 487-491.
Sambrook, J.; Russell, D.W. 2001. Molecular cloning: a laboratory manual. v. I. Cold Spring
Harbor Press. Cold Spring.
Sanger, F.; Nicklen, S.; Coulson, A.R. 1977. DNA Sequencing with chain terminating
inhibitors. Proceedings of National Academy of Science, 74: 5463-5467.
Schrader, O.; Ahne, R.; Fuchs, J.; Schubert, I. 1997. Karyotype analysis of Helianthus annuus
using Giemsa banding and fluorescence in situ hybridization. Chromosome Research, 5:
451-456.
Seabright, M. 1971. A rapid banding technique for human chromosomes. Lancet, 11: 971-
972.
Segal, Y.; Peissel, B.; Renieri, A.; Marchi, M.; Ballabio, A.; Pei, Y.; Zhou, J. 1999. LINE-1
elements at the sites of molecular rearrangements in Alport Syndrome–Diffuse
Leiomyomatosis. The American Journal of Human Genetics, 64: 62-69.
Silva, M.J.J.; Patton, J.L.; Yonenaga-Yassuda, Y. 2006. Phylogenetic relationships and
karyotype evolution in the Sigmodontine rodent Akodon (2n=10 and 2n=16) from Brazil.
Genetics and Molecular Biology, 29: 469-474.
Souza, E.M.S.; Moraes, B.N.G.; Andrade, R.A.; Gross, M.C.; Feldberg, E. 2009. Anfiplastia
em Caluromys philander (Didelphimorphia, Didelphidae): é a explicação para diferenças
no número fundamental? In: III Encontro de Genética do Norte (ENGENOR), Santarém-
PA.
Stock, A.D. 1981. Chromosomal variation and constitutive heterochromatin in three porpoise
specie (genus Stenella). Cytogenetic Cell Genetic, 31(2): 91-100.
Sullivan, G.J.; Bridger, J.M.; Cuthbert, A.P.; Newbold, R.F.; Bickmore, W.A.; McStay, B.
2001. Human acrocentric chromosomes with transcriptionally silent nucleolar organizer
regions associate with nucleoli. The European Molecular Biology Organization Journal,
20 (11): 2867-2877.
55

Sumner, A.T. 1972. A simple technique for demonstrating centromeric heterochromatin.
Experimental Cell Research, 75: 304-306.
Suzuki, H.; Moriwaki, K.; Sakura, S. 1994. Sequences and Evolutionary Analysis of Mouse
5s rDNAs. Molecular Biology and Evolution, 11: 704-710.
Szymanski, M.; Nalaskowska, M.; Barciszewski, J. 1992. The nucleotide sequence of 5S
rRNA from bovine liver. Acta Biochimica Polinica, 39: 295-299.
Tanaka, K.; Matsuda, Y.; Masangkay, J.S.; Solis, C.D.; Anunciado, R.V.P.; Kuro, M.;
Namikawa, T. 2000. Cytogenetic analysis of Tamaraw (Bubalus mindorensis): A
comparison of R-Banded karyotype and chromosomal distribution of centromeric
satellite DNAs, telomeric sequence, and 18S-28S rRNA genes with domestic water
buffaloes. The Journal of Heredity, 91: 117-121.
Tarpley, R.J.; Jarrel, G.H.; George, J.C.; Cubbage, J.; Stott, G.G. 1995. Male
pseudohermaphroditism in the Bowhead Whale, Balaena mysticetus. Journal of
Mammalogy, 76: 1267-1275.
Thompson, J.D.; Higging, D.G.; Gibson, T.J. 1994. ClustalW: Improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, position specific
gap penalties and matrix choice. Nucleic Acids Research, 22: 4673-4680.
Torres, J.P.M.; Lailson-Brito, J.; Saldanha, G.C.; Dorneles, P.; Silva, C.E.A.; Malm, O.;
Guimarães, J.R.; Azeredo, A.; Bastos, W.R.; da Silva, V.M.F.; Martin, A.R.; Cláudio, L.;
Markowitz, S. 2009. Persistent toxic substances in the Brazilian Amazon: Contamination
of Man and the Environment. Journal of Brazilian Chemical Society, 20(6): 1175-1179.
Trebbau, P.; van Bree, P.J.H. 1974. Notes concerning the freshwater dolphin Inia geoffrensis
(de Blainville, 1817) in Venezuela. Zeitschrift des Säugetierk, 39: 50-57.
Vianna, J.A.; Bonde, R.K.; Caballero, S.; Giraldo, J.P.; Lima, R.P.; Clark, A.; Marmontel, M.;
Morales-Vela, Souza, M.J.; Parr, L.; Rodríguez-Lopez, M.A.; Mignucci-Giannoni, A.A.;
Powell, J.A.; Santos, F.R. 2006. Phylogeography, phylogeny and hybridization in
trichechid sirenians: implications for manatee conservation. Molecular Ecology, 15: 433-
447.
Walen, K.H.; Madin, S.H. 1965. Comparative chromosome analyses of the bootlenosed
dolphin (Tursiops truncatus) and the pilot whale (Globicephala scammoni). American
Naturalist, 99: 349-354.
56

Watanabe, H.; Tateno, H.; Kusaksabe, H.; Matsuoka, T.; Kamiguchi, Y.; Fujise, Y.; Ishikawa,
H.; Ohsumi, S.; Fukui, Y. 2007. Fertibility and chromosomal integrity of frozen-thawed
Bryde’s whale (Baleonoptera edeni) spermatozoa intracytoplasmically injected into
mouse oocytes. Zygote, 15: 9-14.
Waters, P.D.; Dobigny, A.T.P.; Robison, T.J. 2004. LINE-1 distribution in Afrotheria and
Xenarthra: implications for understanding the evolution of LINE-1 in eutherian genomes
Chromosoma, 113: 137–144.
Widegren, B.; Árnason, U.; Akusjarvi, G. 1985. Characteristics of a conserved 1,579 bp highy
repetitive component in the killer whale, Orcinus orca. Molecular Biology and
Evolution. 2(5): 41l-419.
Wise, J.P.; Wise, S.S.; Kraus, S.; Shaffiey, F.; Grau, M.; Chen, T.L.; Perkings, C.; Thompson,
D.; Zheng, T.; Zhang, Y.; Romano, T.; O’Hara, T. 2008. Hexavalent chromium is
cytotoxic and genotoxic to the North Atlantic right whale (Eubalaena glacialis) lung and
testes fibroblasts. Mutation Research, 650: 30-38.
Yonenaga-Yassuda, Y. 2004. Contribuição da FISH à citogenética de mamíferos e répteis. p.
89-114. In: Guerra, M. FISH Conceitos e aplicações na citogenética. Sociedade
Brasileira de Genética, Ribeirão Preto, Brasil.
Zijlstra, C.; Mellink, C.H.M.; Haan, N.A.; Bosma, A.A. 1997. Localization of the 18S, 5.8S
and 28S rRNA genes and the 5S rRNA genes in the babirusa and the white-lipped
peccary. Cytogenetic and Cell Genetics, 77: 273-277.
Zurita, F.; Jimenez, R.; Guardia, R.D.; Burgos, M. 1999. The relative rDNA content of a
NOR determines its level of expression and its probability of becoming active. A
sequential silver staining and in-situ hybridization study. Chromosome Research, 7: 563-
570.
57





























