Embed Size (px)
Citation preview

Charakterisierung des XIAP-Gens bei zwei Familien mit X-chromosomalem lymphoproliferativem Syndrom
Dissertation zur Erlangung des akademischen Grades
Dr. med. an der Medizinischen Fakultät
der Universität Leipzig Eingereicht von: Peter Christian Horn geboren am 04.08.1982 in Freiburg Angefertigt an: Der Universitätsklinik und Poliklinik für Kinder und Jugendliche Leipzig und dem Zen-trum für familiären Brust- und Eierstockkrebs der technischen Universität München Betreuer: Prof. Dr. med. Volker Schuster (Universität Leipzig) und Prof. Dr. rer. nat. Alfons Meindl (TU München) Beschluss über die Verleihung des Doktorgrades vom:

Inhaltsverzeichnis 2
Inhaltsverzeichnis
Inhaltsverzeichnis ........................................................................................................... 2
Abbildungsverzeichnis.................................................................................................... 4
Tabellenverzeichnis......................................................................................................... 4
Bibliographische Beschreibung ..................................................................................... 5
Abkürzungsverzeichnis .................................................................................................. 6
Vorwort ............................................................................................................................ 8
1 Einleitung................................................................................................................... 9 1.1 Ziele / Fragestellung ................................................................................................ 9 1.2 Wissensstand zu Beginn der Arbeit ....................................................................... 10 1.2.1 Begriffserklärung: Lymphoproliferative Erkrankung ......................................... 10 1.2.2 Begriffserklärung: Mononukleose ....................................................................... 11 1.2.3 Begriffserklärung: Hämophagozytische Lymphhistiozytose (HLH) .................. 11 1.2.4 X-chromosomale lymphoproliferative Syndrome – Erstbeschreibung ............... 13 1.2.5 X-chromosomale lymphoproliferative Syndrome - Entdeckung der XIAP-
Defizienz.............................................................................................................. 18 1.2.6 X-chromosomale lymphoproliferative Syndrome – Klinik der XIAP-
Defizienz.............................................................................................................. 22 1.3 Aufgabenstellung ................................................................................................... 22
2 Material und Methoden.......................................................................................... 25 2.1 Auswahl der zu untersuchenden Proben ................................................................ 25 2.2 Genetische Untersuchung ...................................................................................... 25 2.2.1 Isolierung der DNA ............................................................................................. 26 2.2.2 Herstellung der Primer......................................................................................... 27 2.2.3 Polymerase- Kettenreaktion (PCR) und Sequenzierung ..................................... 29 2.3 Auswertung ............................................................................................................ 31
3 Resultate................................................................................................................... 32 3.1 Genveränderungen ................................................................................................. 32 3.1.1 Single Nucleotide-Polymorphismen.................................................................... 32 3.1.2 Mutationen........................................................................................................... 32 3.2 Klinik der Patienten ............................................................................................... 33 3.2.1 Familie I: S347Lfs5x-Mutation ........................................................................... 33 3.2.2 Familie I: Mutter und zwei Onkel der Betroffenen ............................................. 34 3.2.3 Familie I: 27 Jahre alter, positiv getesteter Erwachsener .................................... 35

Inhaltsverzeichnis 3
3.2.4 Familie I: 29 Jahre alter, positiv getester Erwachsener ....................................... 35 3.2.5 Familie II: 17 Jahre alter Knabe mit R222X-Mutation ....................................... 37 3.3 Übersicht über die Klinik der Patienten................................................................. 38
4 Diskussion ................................................................................................................ 41 4.1 Hauptthese: Im Patientenkollektiv können XIAP-Gen Mutationsträger
identifiziert werden.............................................................................................. 41 4.1.1 Prozentualer Anteil positiv getesteter Proben und Vergleich mit der Literatur .. 42 4.1.2 Vergleich der erhobenen klinischen Daten mit der Literatur .............................. 44 4.2 Nebenthese: Bei XIAP-Defizienz werden im Gegensatz zur SAP-Defizienz
keine Lymphome beobachtet............................................................................... 46 4.3 Nebenthese: Bei der Therapie kann ein konservativer Ansatz einer
Knochenmarkstransplantation vorzuziehen sein ................................................. 47
5 Zusammenfassung der Arbeit................................................................................ 52
Anhang: Einwilligungen............................................................................................... 54
Literaturverzeichnis ..................................................................................................... 56
Erklärung über die eigenständige Abfassung der Arbeit ......................................... 62
Lebenslauf...................................................................................................................... 63
Verzeichnis eigener Veröffentlichungen ..................................................................... 64
Danksagung ................................................................................................................... 65

Abbildungsverzeichnis 4
Abbildungsverzeichnis Abbildung 1: Beispiel für Hämophagozytose durch mehrere Makrophagen im
Blutausstrich........................................................................................................ 12 Abbildung 2: Genlocus von SAP.. .................................................................................. 15 Abbildung 3: Bindung von SAP am SLAM-Rezeptor.. ................................................. 17 Abbildung 4: Lage von XIAP und SAP auf dem Chromosomenabschnitt Xq25........... 19 Abbildung 5: Die drei Aktivierungspfade der Apoptose. ............................................... 20 Abbildung 6: XIAP hemmt die Apoptose an drei kritischen Stellen.............................. 21 Abbildung 7: Stammbaum der Familie mit S347LfsX-Mutation. .................................. 34 Abbildung 8: Hautaffektion bei XIAP-defizientem Patienten........................................ 36 Abbildung 9: Vorlage für die Einwilligung für genetische Untersuchungen der
Ludwig-Maximilians-Universität München........................................................ 55
Tabellenverzeichnis Tabelle 1: Auswertung der Daten der ersten 100 identifizierten XLP-Patienten ........... 14 Tabelle 2: Verteilung der Phänotypen von XLP............................................................. 15 Tabelle 3: Verwendete Primer. ....................................................................................... 28 Tabelle 4 Übersicht über die klinischen Symptome der Patienten. ................................ 39 Tabelle 5: Übersicht über die Klinik bei XIAP-Defizienz-Patienten. ............................ 44 Tabelle 6: Vergleich der Klinik von XLP-1 und XLP-2, sowie Übersicht über alle
veröffentlichten klinischen Angaben über XLP-2 Patienten. ............................. 45 Tabelle 7: Retrospektive Mortalität in den verschiedenen Behandlungsgruppenbei
XIAP Defizienz................................................................................................... 48 Tabelle 8.: Überleben und Todesursache bei 19 knochenmarkstransplantierten
XIAP-Defizienz Patienten................................................................................... 50

Bibliographische Beschreibung 5
Bibliographische Beschreibung Name: Horn, Peter Christian.
Titel: Charakterisierung des XIAP-Gens bei zwei Familien mit X-chromosomalem lymphoproliferativem Syndrom.
Universität Leipzig, Dissertation
S.: 65 Seiten, 82 zitierten Quellen, 9 Abb., 8 Tab.
Referat:
Im Rahmen dieser Arbeit wurde bei einem Patientenkollektiv mit lymphoproliferativem Syndrom untersuch, ob Mutationen im XIAP-Gen Auslöser der Erkrankung sind. Es wurde eine molekulargenetische Sequenzierung und Auswertung des XIAP-Gens bei 36 Verdachtsfällen eines X-chromosomal vererbten lymphoproliferativen Syndroms durchgeführt. In allen Fällen wurde der gesamte exonische Abschnitt des Gens sequen-ziert und auf Polymorphismen und Mutationen untersucht. Bei zwei Proben wurden Mutationen im XIAP-Gen gefunden. Weiter konnten Mutter und Bruder eines der Be-troffenen untersucht werden, so dass insgesamt drei Knaben mit XIAP-Gendefekt sowie eine heterozygote Konduktorin identifiziert wurden. Nach der Identifikation der Mutati-onsträger erfolgte eine Auswertung der Krankengeschichte und ein Vergleich mit den verfügbaren Beschreibungen von XIAP-Defizienz. Die beiden neu identifizierten Muta-tionen verursachen ein Krankheitsbild, das mit den wenigen verfügbaren Beschreibun-gen von XIAP-Defizienz vereinbar ist. Die Auswertung der Klinik der drei Betroffenen unterstützt die Hypothese, dass bei XIAP-Defizienz keine Lymphome auftreten. Diese Arbeit konnte die ersten Patienten mit XIAP-Defizienz im deutschsprachigen Raum identifizieren und zwei bisher nicht beschriebene Mutationen des XIAP-Gens beim Menschen aufdecken.

Abkürzungsverzeichnis 6
Abkürzungsverzeichnis
ALPS Autoimmune lymphoproliferative syndrome
BIRC4 Baculoviral IAP repeat-containing protein 4, entspricht ! XIAP
CD (#) Cluster of Differentiation; Nomenklatur der Oberflächenepitope von immunologisch aktiven Zellen
CVID Kombiniertes variables Immundefizienz-Syndrom
DNA Desoxyribonukleinsäure. Im Text synonym verwandt für die informationskodierenden Moleküle, die den genetischen Code konstituieren
EBV Epstein-Barr Virus
HCT Hematopoetic Stemcell Transplantation = Knochenmarks-transplantation
HLH hämophagozytische Lymphhistiozytose
fHLH familäre hämophagozytische Lymphhistiozytose
Ig Immunglobulin
IVIG Intravenöse Gabe von Immunglobulinpräparaten aus dem gepoolten Plasma gesunder Spender
LDH Laktatdehydrogenase
LPD Lymphoproliferative Disorder / Lymphoproliferative Erkran-kung
MAC Myeloablative conditioning (Standardchemotherapie mit Hochdosis-Busulfan zur Knochenmarksabtragung vor Kno-chenmarkstransplantation)
MIM (#) Zuordnungsnummer im weltweiten Register für Gendefekte, dem Mendelian Inheritance in Man
NCBI National Center for Biotechnology Information
PTLD Post-transplant lymphoproliferative disorder; Transplantati-onsassoziierte Lymphoproliferative Erkrankung

Abkürzungsverzeichnis 7
RIC Reduced intensity conditioning (Chemotherapie mit unter-schiedlichen Chemotherapeutika zur Knochenmarksabtragung vor Knochenmarkstransplantation, weniger toxisch als ! MAC)
SAP SLAM-associated Protein
SCID Severe combined Immonodeficiency; Schweres kombiniertes Immundefizienz-Sydrom
SH2-Domäne Src-homology 2-Domäne; ca. 100 Aminosäuren lange Prote-indomäne die in einer Vielzahl von Proteinen die Bindung von Peptiden mit phosphoryliertem Tyrosin ermöglicht
SLAM Signalling Lymphocyte Activation Molecule, bindet an SAP
SNP Single nucleotide polymorphism
TNF Tumor necrosis factor
TRAIL TNF-related apoptosis-inducing ligand; Synonym: CD 253; Erstes Glied der TNF-alpha vermittelten Apoptosekaskade
XIAP X-linked inhibitor of apoptosis
XLP X-linked lymphoproliferative syndrome
XLP-1 X-linked lymphoproliferative disorder aufgrund einer Mutation im SAP-Gen (nicht mehr gebräuchlich)
XLP-2 X-linked lymphoproliferative disorder aufgrund einer Mutation im XIAP-Gen (nicht mehr gebräuchlich)
Xq25 Genlocus von SAP und XIAP; beide liegen ca. 1,8 Megabasenpaare auseinander
Anmerkung: Da ein Großteil der heutigen Forschungsergebnisse in englischer Sprache veröffentlicht werden, sind die Abkürzungen in dieser Arbeit von den englischen Fach-begriffen abgeleitet. Dies soll den Vergleich zur englischen Fachliteratur erleichtern.

Vorwort Die vorgelegte Doktorarbeit ist 2008 an der Technischen Universität München begon-nen worden. Der Idee der Untersuchung des XIAP-Gens bei unseren Probanden war gemeinschaftlich von Professor B. Belohradsky, Professor A. Meindl und Professor V. Schuster entwickelt worden. Das Thema wurde von Professor A. Meindl vorgeschlagen, von dem auch die Materialien, Räumlichkeiten, finanziellen Mittel und Genproben zur Verfügung gestellt wurden. Die Einarbeitung erfolgte durch die Mitarbeiter des Labors für experimentelle Onkologie der Frauenklinik der TU München, insbesondere durch Heide Hellebrand und Marko Bertone. Der experimentelle Teil der Arbeit wurde Ende 2009 abgeschlossen und 2010 (digitale Ausgabe) bzw. 2011 (Printausgabe) im Journal of Allergy and Clinical Immunology veröffentlicht. Nach Abschluss des experimentel-len Teils wurde die Korrektur und Betreuung der Doktorarbeit freundlicherweise von Professor Schuster übernommen, der auch an der im JACI veröffentlichten Arbeit mit-gewirkt hatte.

1 Einleitung Vorliegende Arbeit hatte zum Ziel, Mutationsträger in einem Gen auf dem X-Chromosom zu identifizieren, welches für den Apoptose-Inhibitor XIAP (X-linked In-hibitor of Apoptosis) kodiert. XIAP-Mutationen bei Menschen wurden 2006 erstmals von Rigaud et al beschrieben (Rigaud, 2006). Aus historischen Gründen wurden für die daraus resultierende Krankheit mehrere Namen verwendet:
" Der Name XLP-2 oder X-linked lymphoproliferative syndrome 2 wurde von Rigaud et al benutzt, um die Krankheit von XLP-1 bzw. SAP-Defizienz abzugrenzen (siehe 1.2.5). SAP-Defizienz ist ein anderes Syndrom, von dem ursprünglich gedacht wur-de, dass es mit XIAP-Defizienz verwandt ist. Die Namensgebung „XLP-1/XLP-2“ wurde in Folge viel kritisiert und ist irreführend, da die beiden Erkrankungen, von Erbgang und einzelnen Symptomen abgesehen, nicht verwandt zu sein scheinen.
" Die Namen SH2D1A- bzw. BIRC4-Mutation beziehen sich auf den Genlokus bzw. die aktive Domäne des geschädigten Proteins (siehe 1.2.5) und waren vor allem in wissenschaftlichen Grundlagenarbeiten gebräuchlich.
In dieser Arbeit wird durchgehend der unmissverständliche Name XIAP-Defizienz be-nutzt.
Die Arbeit verfolgt drei Ziele:
1. Identifikation von Patienten mit XIAP-Defizienz
2. Beschreibung nicht bekannter Mutationen im XIAP-Gen
3. Zusammenstellung von Klinik sowie Therapieerfahrung bei den betroffenen Patien-ten
Ausgehend von diesen Zielen, die im nachfolgenden Kapitel 1.1 im Detail aufgeführt sind, und unter Betrachtung des Wissenstands zu Beginn der Arbeit (Kapitel 1.2) wird der eingeschlagene Lösungsansatz in Kapitel Error! Reference source not found. des Einleitungsteils beschrieben. Die Hauptteile der Arbeit sind im Methoden- und Ergeb-nisteil dargelegt. Ein Diskussionsteil, welcher die Ergebnisse der Arbeit unter Berück-sichtigung der aktuellen Literatur zum Thema beschreibt, bildet den Abschluss.
1.1 Ziele / Fragestellung
Erstes Ziel dieser Arbeit war es, bei einem Patientenkollektiv mit klinischem Verdacht auf ein X-chromosomal vererbtes lymphoproliferatives Syndrom (XLP) eine vollständi-ge Sequenzierung des XIAP-Gens durchzuführen. Von allen Patienten lag eine gültige

1 Einleitung 10
Einwilligung in die Untersuchung krankheitsrelevanter Genabschnitte und in die wis-senschaftliche Auswertung der Ergebnisse vor (siehe Anhang A). Alle Proben hatten gemeinsam, dass zuvor eine Mutation im SAP-Gen ausgeschlossen worden war.
Zu Beginn der Arbeit wurden drei Thesen formuliert, die durch die Arbeit überprüft werden sollten. Die Hauptthese besagte, dass
" bei Patienten, bei denen ein klinischer Verdacht auf ein X-chromosomales lympho-proliferatives Syndrom bestand und keine SAP-Mutation nachgewiesen werden konnte, XIAP-Mutationen für die Erkrankung ursächlich sein könnten. Es bestand darüberhinaus die Möglichkeit, bislang nicht beschriebene Mutationen im XIAP-Gen zu finden.
Falls Mutationsträger indentifiziert würden, sollte die Klinik und die bisherige Therapie der Patienten beschrieben und mit der Literatur verglichen werden. Zwei Nebenthesen besagten, dass
" XIAP-defiziente Patienten im Unterschied zu Patienten mit SAP-Defizienz nicht zur Entwicklung von Lymphomen neigen würden
" Knochenmarkstransplantation bei XIAP-Defizienz gefährlicher als bei anderen Er-krankungen sein könnte, und aufgrund eines im Vergleich zur SAP-Defizienz milde-ren Verlaufs möglicherweise nicht notwendig sei.
1.2 Wissensstand zu Beginn der Arbeit
Gegenstand der Arbeit ist eine familiär erbliche Erkrankung, die sich durch überschie-ßende Immunreaktion nach bestimmten Virusinfektionen äußern kann. Zunächst sollen daher drei Begriffe erläutert werden, die für die Arbeitgrundlegend sind: Lymphoproli-ferative Erkrankung (LPD), fulminante Mononukleose und hämophagozytische Lymph-histiozytose (HLH). Anschließend soll in Grundzügen die Entdeckung X-chromosomaler lymphoproliferativer Syndrome bis zur Entdeckung der XIAP-Defizienz nachgezeichnet werden, und der Wissensstand zu Beginn der Arbeit, aus dem sich die Fragestellung ergibt, dargelegt werden.
1.2.1 Begriffserklärung: Lymphoproliferative Erkrankung
Als lymphoproliferative Erkrankung (lymphoproliferative disorder, LPD) bezeichnet man die klonale Expansion von Lymphozyten im Knochenmark und anderen Geweben. Meist sind B-Zelllininien betroffen, und es handelt sich fast ausschließlich um immun-supprimierte Patienten. Die Expansion der Zellen bei LPD an sich ist meist nicht mali-gne. Allerdings treten bei Betroffenen mit LPD gehäuft sekundäre maligne Lymphome auf. Das Sterblichkeitsrisiko von Kindern mit LPD durch Krebserkrankungen ist im Vergleich zu immunkompetenten Kindern deutlich erhöht (Spector, Perry, & Kersey,

1 Einleitung 11
1978). LPD kann erblich bedingt, erworben oder iatrogener Ursache sein. Sowohl in erblichen als auch in erworbenen Fällen wird eine starke, aber nicht exklusive Assozia-tion zu einer Infektion mit dem Epstein-Barr-Virus (EBV) beobachtet (Reddy, Rezvani, Barrett, & Savani, 2011; Sullivan et al., 1981). Man geht davon aus, dass es in diesen Fällen der Erkrankung zu einer Immortalisierung von B-Zellen durch EBV kommt. Bei immunsupprimierten Patienten können diese immortalisierten B-Zellen aufgrund unzu-reichender zellulärer Kontrolle ungehindert proliferieren. In der Folge kann es zu einer sekundären Infiltration unterschiedlicher lymphoider Organe kommen, wobei andere Zelllinien in ihrer Ausreifung gestört werden und es zu unterschiedlichen pathologi-schen Zusammensetzungen des lymphozytären Zellbildes kommen kann (Spector et al., 1978).
1.2.2 Begriffserklärung: Mononukleose
Mononukleose wird durch das Epstein-Barr-Virus (EBV) ausgelöst. EBV gehört zur Familie der Herpesviren und zählt zu den am häufigsten beim Menschen auftretenden Viren. Weltweit sind über 90% der Bevölkerung infiziert. Eine Infektion mit EBV im Kindesalter verläuft in der Regel symptomarm oder -los, während eine Erstinfektion im Jugend- und Erwachsenenalter häufig das Krankheitsbild der infektiösen Mononukleose auslöst (Cepok, 2004). Infektiöse Mononukleose ist eine meist gutartig verlaufende, akute Erkrankung, die durch Fieber, Lymphknotenschwellung (insbesondere im oropha-ryngealen Bereich) und Hepatosplenomegalie gekennzeichnet ist. Der Verlauf ist nor-malerweise selbstlimitierend. In sehr seltenen Fällen kann die infektiöse Mononukleose jedoch eine hämophagozytische Lymphhistiozytose (HLH, siehe 1.2.3) mit fulminanter Hepatitis auslösen (Okano & Gross, 2012). Die Kombination aus HLH und fulminanter Hepatitis wird als schwere (je nach Verlauf auch fatale) infektiöse Mononukleose be-zeichnet; tritt HLH ohne Hepatitis auf, spricht man von EBV-assoziierter hämophago-zytischer Lymphhistiozytose (Okano & Gross, 2012).
Neben der schweren infektiösen Mononukleose gibt es weitere mit dem Epstein-Barr Virus assoziierte, z.T. lebensbedrohliche Erkrankungen. Zu diesen zählen das Burkitt-Lymphom, das undifferenzierte nasopharyngeale Karzinom, Hodgkin-Lymphom, Lymphoproliferation der B-/T- und natürlichen Killerzell (NK)-Linien, Magenkarzino-me und chronisch aktive EBV-Infektion bei Immunsupprimierten (Okano & Gross, 2012). Für diese Arbeit sind insbesondere die lymphoproliferativen Verläufe sowie HLH von Bedeutung.
1.2.3 Begriffserklärung: Hämophagozytische Lymphhistiozytose (HLH)
HLH ist ein seltenes Syndrom, das mit anhaltendem Fieber, Hepatosplenomegalie, Zy-topenie und neurologischen Symptomen einhergeht. Das kennzeichnende pathologische Merkmal ist der Nachweis von Hämophagozytose im Knochenmark oder anderen

1 Einleitung 12
lymphatischen Geweben (Abbildung 1). Hierbei finden sich aktivierte Makrophagen, welche Erythrozyten, Leukozyten, Thrombozyten und Vorläuferzellen phagozytieren (Fisman, 2000). HLH kann sowohl genetisch bedingt als auch sekundär auftreten. Von der familiären Form (fHLH) sind mindestens fünf genetische Subgruppen mit teils un-terschiedlichem Ausprägungsgrad bekannt (Chandrakasan & Filipovich, 2013; Völkel, 2005; Zhang, Filipovich, Johnson, Marsh, & Villanueva, 2010). Sekundäre (bzw. er-worbene) HLH tritt nach starker immunologischer Aktivierung, beispielsweise durch bestimmte Infektionen (z.B. EBV), Immundefekte oder maligne Tumoren auf (Fisman, 2000). Unbehandelt ist HLH in der Regel tödlich, die klinische Ausprägung ist variabel (Altschul, Gish, Miller, Myers, & Lipman, 1990; Zhang et al., 2010). Der Nachweis von Hämophagozytose im Blut- oder Knochenmarksausstrich gelingt nicht immer, und ein negativer Ausstrich schließt HLH nicht aus, die Diagnose der familiären Form kann durch einen Mutationsnachweis gesichert werden. (Benoit, Rose, Schmidt, & Zillikens, 2003; Janka, 2012).
Abbildung 1: Beispiel für Hämophagozytose durch mehrere Makrophagen im Blutausstrich. In den Ma-krophagen oberhalb des Megakaryozyten zeigt sich viel phagozytiertes Hämochromatin. Der gezeigte Blutausstrich wurde bei Patient III/1 aus Familie 1 angefertigt (siehe Abschnitt Resultate, Kapitel 3.2.3). Fotografie von Prof. V. Schuster.

1 Einleitung 13
1.2.4 X-chromosomale lymphoproliferative Syndrome – Erstbeschreibung
Für das Verständis der Fragestellung dieser Arbeit ist es sinnvoll, den Weg zu der Ent-deckung der XIAP-Defizienz kurz zu skizzieren.
1974 veröffentlichte der Pathologe und Mikrobiologe David T. Purtilo mit dem Stamm-baum der Duncan-Familie die Klinik eines X-chromosomal vererbten lymphoprolifera-tiven Syndroms (Benoit et al., 2003; Purtilo, Cassel, & Yang, 1974). In dieser Familie war bei 6 von 18 Jungen eine tödliche lymphoproliferative Erkrankung aufgetreten. Mädchen waren nicht betroffen. Die Ausprägung reichte von benigner bis hin zu malig-ner Proliferation; insbesondere traten maligne B-Zell Lymphome meist vom Burkitt-Typ auf. Bei der Autopsie betroffener Knaben waren Thymus und thymusabhängige Bereiche der Lymphknoten nekrotisch und zelldepletiert. In hämatopoetischen Organen und im zentralen Nervensystem fanden sich diffuse Infiltrate aus Lymphozyten, Plas-mazellen und Histiozyten. Einige der Histiozyten enthielten phagozytierte Erythrozyten. Bei mindestens drei der Knaben ging dem tödlichen Ereignis eine Infektion mit dem Epstein-Barr Virus (EBV) voraus (Purtilo, Cassel, Yang, & Harper, 1975).
Purtilos Arbeitsgruppe hatte bis dahin vor allem an erblichen Tumorprädispositionssyn-dromen geforscht. Obwohl auch bei dieser neu entdeckten, X-chromosomal vererbten Erkrankung bei mehreren Jungen Lymphome auftraten, vermutete Purtilo früh, dass die Malignome sekundär entstanden. Er schrieb 1977:
„In (..) XLP (..) at least 20 boys succumbed from a variety of B-cell lymphomas. Under-lying immunodeficiencies probably permitted fatal lymphoproliferation to occur in XLP. In contrast, intrinsic cellular defects, rather than immunological defects, (are) partially responsible in (other carcinogenesis-Syndromes).“
Bei der Duncan-Familie lag demnach ein Gendefekt vor, der über einen Defekt der Im-munabwehr eine unkontrollierte Lymphoproliferation gestattete und hierdurch ein Ent-artungsrisiko darstellte (D T Purtilo, 1977). Aufgrund seiner klinischen und pathologi-schen Beobachtungen mutmaßte Purtilo, dass die fatale Proliferation bei den Duncan-Knaben durch das Epstein-Barr-Virus (oder andere Viren) getriggert wurde und dass ein progressiver Rückgang der T-Zell-Populationen eine unkontrollierte B-Zell-Lymphoproliferation ermöglicht hatte (Purtilo et al., 1975). Die Erkrankung erhielt den Namen “X-linked lymphoproliferative Disease” und die Abkürzung „XLP.“
1978 wurde das David T. Purtilo-Patientenregister eingeführt, um weltweit Daten über betroffene Familien zu sammeln (Hamilton et al., 1980).
Bereits 1980 waren sieben Familien mit 59 betroffenen Knaben im XLP-Registry ver-zeichnet, 1982 erschien der bis dato größte Report des Registers über 100 Knaben aus 25 Familien (Purtilo et al., 1982). Durch die Auswertung der 100 bekannten Patienten
1 Einleitung 14
von 1982 wurden vier verschiedene, meist überlappende Phänotypen der Erkrankung definiert:
1. Schwere, häufig fatale Infektiöse Mononukleose
2. Maligne B-Zell Lymphome
3. Aplastische Anämie
4. Hypogammaglobulinämie
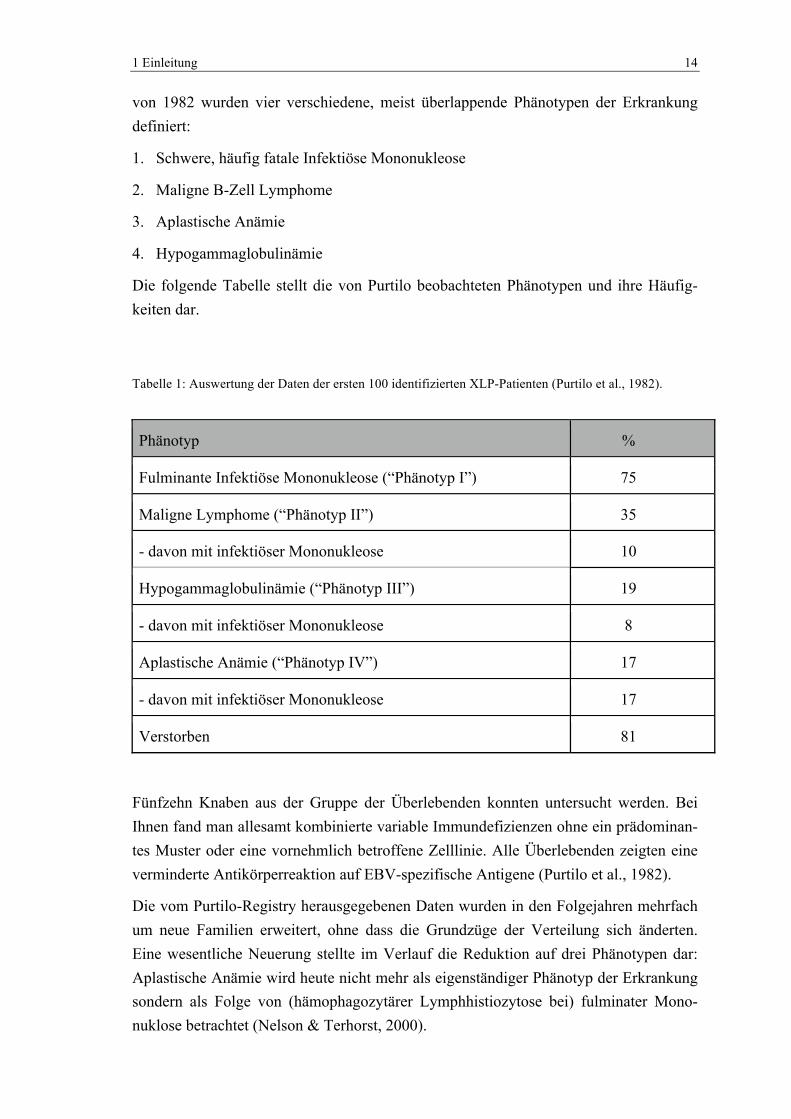
Die folgende Tabelle stellt die von Purtilo beobachteten Phänotypen und ihre Häufig-keiten dar.
Tabelle 1: Auswertung der Daten der ersten 100 identifizierten XLP-Patienten (Purtilo et al., 1982).
Phänotyp %
Fulminante Infektiöse Mononukleose (“Phänotyp I”) 75
Maligne Lymphome (“Phänotyp II”) 35
- davon mit infektiöser Mononukleose 10
Hypogammaglobulinämie (“Phänotyp III”) 19
- davon mit infektiöser Mononukleose 8
Aplastische Anämie (“Phänotyp IV”) 17
- davon mit infektiöser Mononukleose 17
Verstorben 81
Fünfzehn Knaben aus der Gruppe der Überlebenden konnten untersucht werden. Bei Ihnen fand man allesamt kombinierte variable Immundefizienzen ohne ein prädominan-tes Muster oder eine vornehmlich betroffene Zelllinie. Alle Überlebenden zeigten eine verminderte Antikörperreaktion auf EBV-spezifische Antigene (Purtilo et al., 1982).
Die vom Purtilo-Registry herausgegebenen Daten wurden in den Folgejahren mehrfach um neue Familien erweitert, ohne dass die Grundzüge der Verteilung sich änderten. Eine wesentliche Neuerung stellte im Verlauf die Reduktion auf drei Phänotypen dar: Aplastische Anämie wird heute nicht mehr als eigenständiger Phänotyp der Erkrankung sondern als Folge von (hämophagozytärer Lymphhistiozytose bei) fulminater Mono-nuklose betrachtet (Nelson & Terhorst, 2000).
1 Einleitung 15
2004 wurde die Verteilung der Phänotypen folgendermaßen angegeben:
Tabelle 2: Verteilung der Phänotypen von XLP (Ehl & Schuster, 2004; Seemayer et al., 1995)
Symptom %
Fulminate akute infektiöse Mononukleose ca. 60
- davon mit Hämophagozytosesyndrom 90
Maligne Lymphome ca. 30
Hypo- und Dysgammaglobulinämie bis 30
Fulminate akute infektiöse Mononukleose ca. 60
Verstorben (bei Spontanverlauf) > 90
Die Zahlen basieren auf dem letzten Report des XLP-Registers vor der Entdeckung der genetischen Ursachen des Syndroms; dieser wurde 1995 aus den Daten von nunmehr 272 Betroffenen erstellt.
Nach Purtilos Tod 1992 liefen die Bemühungen weiter, den der Krankheit zu Grunde liegenden Gendefekt zu entschlüsseln (Saxon, 1992). Durch Haplotypen-Kopplungsanalysen war es bis 1995 gelungen, das Kandidatengen für XLP auf einen kleinen Bereich des q-Arms von Chromosom X einzugrenzen (Seemayer et al., 1995)(Abbildung 2).
Abbildung 2: Genlocus von SAP. Das ursächliche Gen für die Erkrankung wurde auf dem langen Arm des X-Chromosoms im Sektor 25 vermutet; hier waren letztlich auch die Klonierungsbemühungen von Coffey et. al erfolgreich.

1 Einleitung 16
1998, fünf Jahre nach Purtilos Tod, gelang europäischen und amerikanischen Arbeits-gruppen die Identifikation und Klonierung des Gens (Coffey et al., 1998; Nichols et al., 1998; Sayos et al., 1998). Das Genprodukt besteht aus einer singulären SH2-Domäne und das Gen erhielt den Namen SH2D1A beziehungsweise SAP-Gen (Coffey et al., 1998). Das Genprodukt von SH2D1A, „SAP“ wird in allen großen Subklassen der T-Zellen exprimiert, weniger in B-Zellen (Sayos et al., 1998). SAP ist ein kleines Mole-kül, das an das große, membrangebundene Glykoprotein SLAM (CD 150) und die ver-wandten Glykoproteine CD84 und CD244 binden kann (siehe Abb. 3). Daher auch der Name des Proteins: SAP steht für „SLAM-associated protein“.
SLAM (Signalling Lymphocyte Activation Molecule) kommt in membrangebundener Form (mSLAM) sowohl auf T- als auch auf B-Lymphozyten vor und wird in zytoplas-matischer (cSLAM) und löslicher (sSLAM) Form von B-Zellen exprimiert (Punnonen et al., 1997).
Wenn SAP an SLAM bindet, blockiert es dort die Bindungsstellen für die SH-2 Domä-ne. Dies verhindert ein Binden der SH2 domain bearing tyrosine phosphatase (SHP-2) (siehe Abbildung 3). Obwohl die betroffenen Patienten eine ungehinderte Expansion von B-Zellen zeigen, scheint SAP für die Regulierung der humoralen Abwehr insbeson-dere durch T-Zellen, nicht aber B-Zellen grundlegend. „SAP expression in T cells, not in B cells, is required for humoral immunity“ (Veillette et al., 2008).
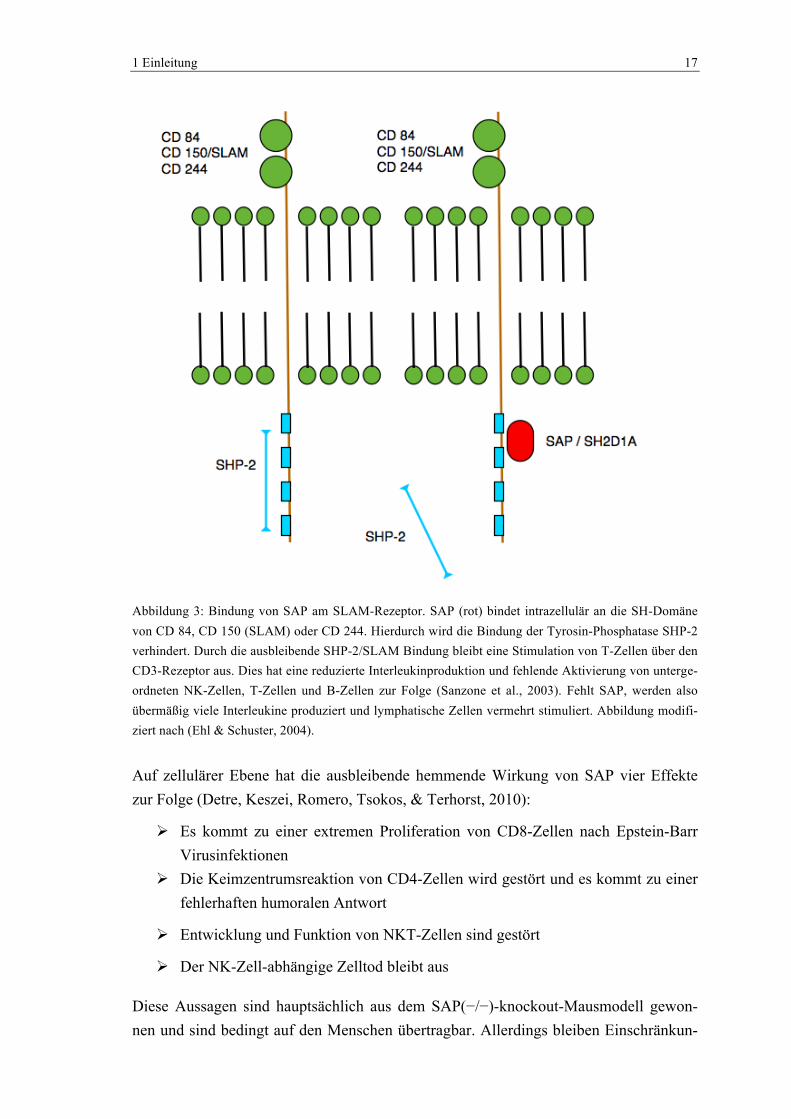
1 Einleitung 17
Abbildung 3: Bindung von SAP am SLAM-Rezeptor. SAP (rot) bindet intrazellulär an die SH-Domäne von CD 84, CD 150 (SLAM) oder CD 244. Hierdurch wird die Bindung der Tyrosin-Phosphatase SHP-2 verhindert. Durch die ausbleibende SHP-2/SLAM Bindung bleibt eine Stimulation von T-Zellen über den CD3-Rezeptor aus. Dies hat eine reduzierte Interleukinproduktion und fehlende Aktivierung von unterge-ordneten NK-Zellen, T-Zellen und B-Zellen zur Folge (Sanzone et al., 2003). Fehlt SAP, werden also übermäßig viele Interleukine produziert und lymphatische Zellen vermehrt stimuliert. Abbildung modifi-ziert nach (Ehl & Schuster, 2004).
Auf zellulärer Ebene hat die ausbleibende hemmende Wirkung von SAP vier Effekte zur Folge (Detre, Keszei, Romero, Tsokos, & Terhorst, 2010):
" Es kommt zu einer extremen Proliferation von CD8-Zellen nach Epstein-Barr Virusinfektionen
" Die Keimzentrumsreaktion von CD4-Zellen wird gestört und es kommt zu einer fehlerhaften humoralen Antwort
" Entwicklung und Funktion von NKT-Zellen sind gestört
" Der NK-Zell-abhängige Zelltod bleibt aus
Diese Aussagen sind hauptsächlich aus dem SAP("/")-knockout-Mausmodell gewon-nen und sind bedingt auf den Menschen übertragbar. Allerdings bleiben Einschränkun-

1 Einleitung 18
gen – an vorderster Stelle beispielsweise, dass Mäuse nicht mit EBV infizierbar sind und deshalb mit Mausherpes- oder Influenza- Viren als Trigger gearbeitet werden muss (Detre et al., 2010).
1.2.5 X-chromosomale lymphoproliferative Syndrome - Entdeckung der XIAP-Defizienz
Die grundlegende Arbeit von Sayos konnte bereits drei Muationationsträger im SAP-Gen unter XLP-Betroffenen identifizieren. Durch den Einsatz verbesserter Sequenzie-rungsmethoden, insbesondere durch Kapillarflusssequenziergeräte, war es in den Folge-jahren möglich, den entsprechenden Genabschnitt eines Großteils der bekannten Gen-träger zu analysieren.
Zur Jahrtausendwende waren innerhalb des XLP-Registries 33 Familien mit Mutations-trägern identifiziert (Sumegi et al., 2000) und 28 verschiedene Mutationen bekannt. Al-lerdings konnte nicht bei allen Familien im XLP-Register eine Mutation des SAP-Gens nachgewiesen werden.
Bei 15 bis 20% der Familien, bei denen eine X-chromosomal vererbte, lymphoprolifera-tive Erkrankung, häufig getriggert durch EBV aufgetreten war, ließ sich keine Mutation im SAP-Gen finden.
Diese „Nachweislücke“ erklärte man sich durch genetische Modifaktoren oder Mutatio-nen auf einem gänzlich anderen Genabschnitt. Der Nachweis des letzteren sollte der Arbeitsgruppe um Sylvain Latour und Stéphanie Rigaud 2006 gelingen.
Das Gen für XIAP befindet sich ebenfalls im Sektor 25 des X-chromosomalen q-Arms (Rigaud et al., 2006), 1.8 Mb-Paare entfernt vom SAP-Gen (Siehe Abbildung 2: Genlo-cus von SAP. Das ursächliche Gen für die Erkrankung wurde auf dem langen Arm des X-Chromosoms im Sektor 25 vermutet; hier waren letztlich auch die Klonierungsbemü-hungen von Coffey et. al erfolgreich. und Abbildung 4) . Dort codiert es für den X-linked inhibitor of apoptosis (XIAP, MIM # 300079(“OMIM Entry - *300079 - INHI-BITOR OF APOPTOSIS, X-LINKED; XIAP,” n.d.)). Das Gen war bereits 1996 von Duckett et al kloniert worden (Duckett et al., 1996). Erst Rigaud et al stellten jedoch den Zusammenhang zu dem „vermissten“ XLP-Gen her.

1 Einleitung 19
Abbildung 4: Lage von XIAP und SAP auf dem Chromosomenabschnitt Xq25 (nach (Rigaud et al., 2006). Die Zahlen in Klammern beziehen sich auf die Entfernung zum p-Telomer in Megabasen. Zwi-schen den Genen für XIAP und SAP liegen also 0.4 Megabasen, und es befindet sich ein weiterer für ein Protein (STAG 2) kodierender Abschnitt zwischen beiden Genen.
Der exonisches Teil des XIAP Gens ist 54 Kilobasenpaare lang, es ist damit doppelt so lang wie SAP (27 kbp). Es wurde vorgeschlagen, XLP, das auf SAP-Mutationen basiert, als XLP-1 zu bezeichnen. XLP aufgrund von XIAP-Mutationen wurde von Rigaud et al XLP-2 genannt. Um Mißverständisse zu vermeiden werden in dieser Arbeit die Begriffe „SAP-Defizienz“ und „XIAP-Defizienz“ verwendet.
XIAP ist ein Apoptose-Inhibitor. Unter Apoptose versteht man die Fähigkeit von mehr-zelligen Organismen, gezielt den Untergang einzelner Zellen einzuleiten. Während die Endstrecke bis zum Tod der jeweiligen Zelle in Ihrer Signalübertragung immer gleich abläuft, gibt es drei verschiedene Aktivierungswege, um die Apoptose in Gang zu set-zen: Einen extrinsischen, der über Interleukine und Tumornekrosefaktoren an Mem-branrezeptoren ausgelöst wird (Abbildung 5 und Abbildung 6), einen intrinsischen, der in der Zelle bei Zellschäden ausgelöst wird, und den Perforin/Granzym Pathway, der einen Kontakt mit zytotoxischen T-Zellen voraussetzt (Abbildung 5) (Elmore, 2007). Die gemeinsame Endstrecke aller drei Apoptosewege beginnt mit der Aktivierung von Kaspase 3 (“execution pathway”) und hat eine Fragmentierung der DNA, Verkümmern des Zellkerns, Auflösung der Zellstruktur und Formierung von Apoptosekörperchen zur Folge.

1 Einleitung 20
Abbildung 5: Die drei Aktivierungspfade der Apoptose. Alle Pfade laufen bei der Aktivierung von Kaspase 3 zusammen; hier beginnt der „execution pathway“ beziehungsweise die Effektor-Kaskade. Bild übernommen mit Genehmigung von Elmore, 2007. XIAP inhibiert vor allem die extrinsische Kaskade, wirkt allerdings auch direkt inhibitorisch auf Kaspase 3.
Der X-linked Inhibitor of Apoptosis XIAP ist vornehmlich ein Inhibitor in der extrinsi-schen Kaskade (Abbildung 5 und Abbildung 6). XIAP gehört zur größeren Familie der baculoviral IAP repeat-containing (BIRC)-Gene, der noch andere Inhibitoren der Apop-tose (IAP) angehören. Allen IAPs ist gemeinsam, dass sie entweder die Aktivierung von Kaspasen verhindern oder die Verbindung von Kaspasen mit deren Zielproteinen bloc-kieren. Sie sind somit potente Inhibitoren der Apoptose.

1 Einleitung 21
Abbildung 6: XIAP hemmt die Apoptose an drei kritischen Stellen. Signalwege unterschiedlicher apopto-sestimulierender extrazellulärer Signalmoleküle laufen im Bereich der sogennanten “Effektorkaspasen” zusammen. Hier wirkt XIAP inhibierend und ist somit in der Lage, die apoptotische Wirkung mehrerer Signalkaskaden zu unterbinden. Es wird diskutiert ob XIAP neben den dargestellten Wirkungen auch in der Lage ist, aktiviertes APAF-1 (Apoptosom) an der Bindung an Kaspase 9 zu hindern (Bratton, Lewis, Butterworth, Duckett, & Cohen, 2002).
XIAP wirkt direkt inhibierend auf die Aktivierung der Kaspasen 3, 7 und 9 und hemmt somit sowohl den Initiationsweg als auch den Effektorweg der Apoptosekaskade (Ma-ródi & Notarangelo, 2007).
Es ist wichtig zu sehen, dass die Apoptosekaskade in Abwesenheit von XIAP ein selbstverstärkender Kreislauf ist. Nachdem einmal die Kaspasenkette angelaufen ist, triggert Kaspase 3 über Ceramide in der Mitochondrienmembran die Freisetzung von Cytochrom C (Riedl & Shi, 2004). Cytochrom C aktiviert wiederum den Apoptose-Protease-Aktivierungs-Faktor 1 (APAF1), der sich mit freiem ATP zum sogenannten Apoptosom verbindet. Das Apoptosom aktiviert dauerhaft Kaspase 9, und der Zyklus beginnt von neuem (Riedl & Shi, 2004). Siehe hierzu Abbildung 6.
Die wichtigste Botschaft der obigen Abbildungen ist:

1 Einleitung 22
" XIAP ist der zentrale Inhibitor der Apoptose. Die Ab- oder Anwesenheit von XIAP ändert nichts daran ob Apoptose auftritt. Wenn sie jedoch in Abwesenheit von XIAP auftritt, beginnt ein sich selbstverstärkender, unkontrollierter Zyklus.
Es bleibt festzuhalten, dass bei XIAP-Defizienz ein Apoptoseinhibitor fehlt, während bei SAP-Defizienz ein deutlicher Anstieg an lymphatischer Zellaktivität und Proliferti-on auftritt.
1.2.6 X-chromosomale lymphoproliferative Syndrome – Klinik der XIAP-Defizienz
Als diese Arbeit begonnen wurde, waren die einzigen identifizierten XIAP-Mutationsträger die zwölf Patienten aus drei Familien, die Rigaud et al (2006) publiziert hatten. Über Klinik und Verlauf von XIAP-Defizienz oder Unterschiede im Vergleich zu XLP-1 war entsprechend wenig bekannt. Die 2009 von der „International Union of Immunological Societies Expert Committee on Primary Immunodeficiencies“ heraus-gegebene Übersichtsarbeit über primäre Immundefekte lieferte eine fast identische Be-schreibung für XIAP-Defizienz wie für SAP-Defizienz:
SAP-Defizienz wurden dort „klinische und immunologische Veränderungen, ausgelöst durch EBV-Infektion, Hepatitis, aplastische Anämie und Lymphome“ zugeordnet, bei XIAP-Defizienz waren „klinische und immunologische Veränderungen, ausgelöst durch EBV-Infektion, Splenomegalie, Hepatitis, hämophagozytäres Syndrom und Lympho-me“ aufgeführt (International Union of Immunological Societies Expert Committee on Primary Immunodeficiencies et al., 2009). Es war daher zu erwarten, dass neu entdeckte Patienten wichtige Informationen zum Phänotyp und Verlauf der Erkrankung beitragen würde.
1.3 Aufgabenstellung
Hauptthese dieser Arbeit war, dass die von Rigaud et. al entdeckte Erkrankung auch in anderen Patientenkollektiven auftreten sollte. Es sollte bewiesen werden, dass unter Blutproben, die in der Arbeitsgruppe von Professor Meindl Arbeitsgruppe negativ auf SAP-Defizienz getestet worden waren, ein gewisser Anteil an XIAP-Mutationen ver-borgen lag. Möglicherweise würde dies auch eine schärfere Abgrenzung zur SAP-Defizienz und in zukünftigen Verdachtsfällen eine gezieltere Testung ermöglichen. Auch für die Patienten in der Untersuchung wären Vorteile absehbar: Zwar war über das Krankheitsbild zum Untersuchungszeitpunkt noch wenig bekannt, doch zeichneten sich theoretische Unterschiede zur SAP-Defizienz ab. Es war anzunehmen, dass mit

1 Einleitung 23
wachsendem Kenntnisstand für Betroffene womoglich unterschiedliche Therapien er-folgsversprechend werden könnten.
Im Falle eines positiven Mutationsnachweises sollten die behandelten Ärzte informiert, und falls durch die Einwilligung gedeckt, die Information und Beratung der Betroffenen durchgeführt werden.
Zum Nachweis der Hauptthese wurde folgender Lösungsansatz festgelegt:
1. Da eine komplette Gensequenzierung des mit 54 kbp im Vergleich zum SAP-Gen doppelt so langen XIAP-Gens sehr aufwendig und kostenintensiv war, soll-ten in einem ersten Schritt aussichtsreiche Kanditen für eine Sequenzierung aus-gewählt werden (siehe auch: Material & Methoden).
2. Nach Auswahl der Patienten sollten Primer hergestellt werden, die eine Sequen-zierung des kompletten XIAP-Gens bei diesen Patienten ermöglichen würden.
3. An einem Wildtypgenom (Genom des Doktoranden) sollten eine Probe-PCR und Sequenzierung des XIAP-Gens mit den Primern erfolgen und abgeglichen werden, ob eine vollständige, auslesbare Abbschifft aller Exone des XIAP-Gens möglich ist.
4. Sodann sollte diese Vorgehensweise auf die DNA der ausgewählten Patienten angewandt werden.
5. Alle gefundenen Unterschiede der untersuchten DNA sollten mit SNP-Datenbanken abgeglichen werden. Echte Mutationen sollten beschrieben wer-den.
6. Wenn Mutationen gefunden würden, sollte eine möglichst exakte Beschreibung der Klinik der betroffenen Patienten erfolgen, und diese zur Ergänzung der ver-fügbaren Information per Publikation behandelnden Ärzten weltweit bereitge-stellt werden.
Neben der Hauptthese ergaben sich aus der Funktion von XIAP als Inhibitor der Apop-tose zwei Nebenthesen:
" Die erhöhte Apoptose-Wahrscheinlichkeit bei Verlust von XIAP sprach dagegen, dass die bei der SAP-Defizienz beobachteten Malignome auch bei XIAP-Defizienz auftreten würden. Es war anzunehmen, dass bei XI-AP-Defizienz Lymphome nicht auftreten würden.
" Vor dem Empfang einer Knochenmarkstransplantation müssen beim Empfänger mit einer Apoptose-induzierenden Chemotherapie die eige-nen blutbildenden Zellen abgetötet werden (Myeloablation). Da in Ab-wesenheit von XIAP eine überschießende Apoptoserate beobachtet wird, besagte die These, dass eine medikamentöse Myeloablation bei Patienten

1 Einleitung 24
mit XIAP-Defizienz gefährlicher sein könnte als bei Patienten mit nor-maler Apoptoserate.
Einen statistischen Beweis zu führen, dass bei XIAP-Defizienz keine Lymphome auftre-ten ist aufgrund der geringen Fallzahlen nicht möglich. Auch definitive Aussagen zur Therapie wären nur mit einer prospektiven randomisierten Studie zu gewinnen, die aus dem gleichen Grund vermutlich nie durchgeführt werden kann. Die Ergebnisse der Ar-beit sollten die Nebenthesen daher entweder unterstützen oder widerlegen.

2 Material und Methoden Die Datenakquise gliederte sich in zwei Teile, namentlich die Auswahl geeigneter Kan-didaten (Abschnitt 2.1) und die Amplifikation und Sequenzierung des XIAP-Gens in deren Genmaterial (Abschnitt 2.2).
2.1 Auswahl der zu untersuchenden Proben
Zwischen 1998 und 2008 waren im gentechnischen Labor der Frauenklinik der Techni-schen Universität München 150 Proben von Patienten mit Einverständniserklärung auf Mutationen im SAP-Gen untersucht worden.
Von den 150 vorliegenden Proben stammten 71 von Familienmitgliedern von auf SAP-Defizienz positiv getesteten Patienten, so dass 79 genetisch unverwandte Proben vorla-gen. Unter diesen 79 Proben war bei insgesamt vierzehn eine SAP-Mutation nachgewie-sen worden. Es lagen also 65 Proben vor, die keine nachweisbare SH2D1A-Mutation trugen und nicht untereinander verwandt waren.
Auf Basis der vorliegenden Krankengeschichten wurden nur solche Proben eingeschlos-sen, bei denen eine Kurzbeschreibung der Klinik der Patienten vorlag, der Verdacht auf ein familiäres Syndrom hierdurch nachvollziehbar war und im Falle eines positiven Er-gebnisses eine vollständige Krankengeschichte anforderbar gewesen wäre.
Ausgeschlossen wurden auch zwei verwandte weibliche Patienten, bei denen ein ähnli-ches Syndrom vorlag. Die Analyse dieser Proben durch eine andere Arbeitsgruppe führ-te zur Entdeckung eines autosomalen Syndroms mit vergleichbarer Klinik (Huck et al., 2009), siehe hierzu Kapitel 4.1.1. Nach Ausschluss der ungeeigneten Einsendungen blieben 36 DNA-Proben, die im Rahmen dieser Arbeit auf Mutationen des XIAP-Gens untersucht wurden.
2.2 Genetische Untersuchung
Grundlage der genetischen Untersuchung ist die Polymerase-Kettenreaktion (PCR, ‚po-lymerase-chain-reaction’). Die PCR ist ein Verfahren zur Vervielfältigung von DNA in vitro. Die PCR wurde 1983 von Kary Mullis entwickelt, die Basis für die Kettenab-bruchs-Sequenzierung bereits um 1975 von Frederick Sanger. Die von Mullis und San-ger entwickelten Verfahren sind Standardmethoden, wenn Gene aufgeschlüsselt werden sollen; zu besseren Lesbarkeit des Text werden im folgenden Abschnitt die expliziten Literaturverweise weggelassen, beziehen sich jedoch- wo nicht anders gekennzeichnet auf Mullis (PCR) und Sanger (Sequenzierung).

2 Material und Methoden 26
Um DNA beinahe unbegrenzt zu vervielfältigen, werden bei einer PCR unter Laborbe-dingungen einer Lösung, welche die abzuschreibende DNA (‚Matrix’) enthält, alle Nu-kleinsäuren (Adenin, Guanin, Cystin und Thymin) beigegeben, die benötigt werden, um DNA herzustellen. Die beiden Stränge der DNA werden durch Erhitzen voneinander getrennt. Mit kurzen DNA-Fragmenten, sogenannten ‚Primern’, markiert man die Stel-le, die abgeschrieben werden soll. Nun kann das Enzym Polymerase hinzugegeben wer-den, das an der markierten Stelle beginnt, eine Abschrift zu erstellen.
Im Labor kommt hier meist die Polymerase des Bakteriums Thermus aquaticus („Taq-Polymerase“) zum Einsatz. Diese ist im Gegensatz zur menschlichen Polymerase hitze-stabil, überlebt das ‚Schmelzen’ der doppelsträngigen DNA und synthetisiert nach Ab-kühlung erneut die komplementären Stränge zur Doppelhelix. So kann man die DNA mehrfach denaturieren, ohne Polymerase erneut hinzugeben zu müssen. Da bei jedem Zyklus auch alle Kopien wieder zum Doppelstrang ergänzt werden, vermehrt man die DNA exponentiell (2n): Beginnt man z.B. mit zwei Kopien, entstehen im nächsten Zy-klus vier, dann acht, dann sechzehn, zweiunddreißig... Kopien. So können bis zum voll-ständigen Aufbrauchen des pipettierten Ansatzes viele Millionen Kopien erstellt wer-den, wenn beispielsweise 30 Zyklen von Erwärmung und Abkühlung durchlaufen wer-den.
Dieser Schritt der Vervielfältigung wurde mit der jeweiligen Matrix-DNA der unter-suchten Patienten durchgeführt. Um die genaue Basenfolge im untersuchten Ge-nabschnitt zu bestimmen, mussten die Kopien ‚sequenziert’, ihre Basenfolge bestimmt werden. Die unten beschriebene Methode basiert auf dem Prinzip, dass vom zu untersu-chenden Abschnitt erneute PCR-Zyklen durchgeführt werden; allerdings in einer Lö-sung, der neben den vier Standard-Nukleotiden auch einzelne ‚Abbruch-Basen’ beige-mischt werden. Diese farbmarkierten Basen werden von der Polymerase unterschiedslos an Stelle eines Nukleotids eingebaut, können jedoch nicht weiter verknüpft werden, wodurch ein Fragment statt einer vollständigen Kopie entsteht. Über die Länge des Fragments kann nun die Position der Base ermittelt werden. In unserem Fall wurden die Basen nach der Diffusion durch eine Mikrokapillare farbcodiert mit Laserlicht abgele-sen.
2.2.1 Isolierung der DNA
Zuerst wurde die DNA der Patienten aus deren Blut „isoliert“. Dieser Schritt war bei den meisten Proben in den Jahren zuvor im Zuge der Sequenzierung des SAP-Gens durch die medizinisch-technischen Assistenten des Labors erfolgt.
In Blutproben, die in EDTA-Röhrchen konserviert vorlagen, kam das Qiamp DNA Blood Mini Kit zum Einsatz (Qiagen).
Die einzelnen Schritte umfassten:

2 Material und Methoden 27
1. Gründliches Vortexen der eingesandten Blutproben
2. 100µl Vollblut in ein 1,5 ml Reaktionsgefäß (Eppendorff-Tube) überpipettieren
3. Nacheinander +100µl hochreines (HPLC-) Wasser, +200µl Pufferlösung (Qiagen) und +30µl lyophylisierte Protease K („Qiagen Proteinase“) hinzugeben
4. den Ansatz 30 sec vortexen, danach für 1 Minute bei 2000 rpm abzentrifugieren
5. Inkubation im Wasserbad bei 65°C für mind. 15-20min
6. +200µl Ethanol 96% hinzugeben, 30 sec schütteln/vortexen, danach für 1 Minute bei 2000 rpm abzentrifugieren
7. den flüssigen Inhalt des Gefäßes, der nun die gereinigte DNA enthält auf eine Qia-gen-Säule überführen
8. Zentrifugation bei 6000 rpm für 4 min
9. Eluat verwerfen, neues Auffanggefäß montieren
10. +500µl AW1-Puffer (Qiagen) auf den Filter geben
11. Zentrifugation 6000rpm für 2min
12. Eluat verwerfen, neues Auffanggefäß montieren
13. Erneut +500µl AW2-Puffer (Qiagen) auf den Filter geben
14. Zentrifugation bei 6000 rpm für 2 min
15. Eluat verwerfen, Montage der Säule auf ein Reaktionsgefäß
16. 100 µl AE-Puffer35 (Qiagen) auf die Säule pipettieren
17. Inkubation bei Raumtemperatur für 1min
18. Zentrifugation bei 14000 rpm für 2 min
19. Einfrieren des Eluats bei -20°C (Völkel, 2005)
Bei zwei Proben war die DNA degeneriert und es konnte auch nach Amplifikationsver-suchen mit dem AmpliKit F13A (Fa. AmpliKit) keine lesbare Matrix mehr hergestellt werden, so dass insgesamt 34 Proben untersucht werden konnten.
2.2.2 Herstellung der Primer
Das XIAP Gen liegt auf dem plus-Strang von Chromosom X im Bereich der Bande q25, zwischen den Basen 122 993 662 bis 123 047 829. Es umschließt somit 54.168 einzelne Basen. Die exonische DNA ist in sechs Exons aufgeteilt, die für eine mRNA von insge-samt 8.413 Basen codieren; das fertige Proteinprodukt besteht aus nur noch 497 Amino-säuren (NCBI GeneLib).

2 Material und Methoden 28
Die Qualität einer PCR ist abhängig von den Primersequenzen, die bei einer bestimmten Optimaltemperatur am besten binden, um eine gute Ausbeute zu erreichen, gleichzeitig aber keine ähnlichen Sequenzen im Genom binden. Zur Vorbereitung einer Polymerase-Kettenreaktion mussten entsprechende Primer angefertigt werden. Die Berechung der Primer erfolgte nach der „nearest neighbor thermodynamic theory” (siehe unten). Für fünf Exons wurden insgesamt sechs ‚Forward’ und sechs ‚Reverse’ Primer angefertigt Dies war notwendig, weil Exon 2 mit mehr als 450 Basenpaaren zu lang für eine sichere durchgängige Sequenzierung war. Deshalb wurde es für die Anfertigung der Primer in zwei Unterabschnitte aufgeteilt, die eigene Primerassays erhielten und in einzelnen Ab-schnitten analysiert wurden.
Alle verwendeten Primer wurden jeweils mindestens 30 Basenpaare über das Start- oder Stoppcodon hinaus gelegt. Bei Exon 2 wurde “überlappend” gearbeitet. Die Primer in-nerhalb des Exons überschnitten sich um knapp 80 Basenpaare.
Ebenso wurde der Abschnitt vor dem Initiationscodon auf Exon 1 um 100 Basenpaare verlängert und mituntersucht, mit dem Ziel, auch Splicevarianten zu entdecken. Alle verwendeten Primer wurden für die Versuche individuell angefertigt (Premier BioSoft International, PaloAlto, USA). Eine Übersicht der verwendeten Primer ist in Tabelle 3 dargestellt.
Tabelle 3: Verwendete Primer. Für Exon 2 wurden aufgrund der Länge zwei überlappende Primerpaare verwendet, um die komplette Sequenz abzudecken.
Exon Forward-Primer Reverse-Primer
E1 5ʼ-TACCGTGCGGTGCTTTAGTT-3ʼ 5ʼ-TTCTGACCAGGCACGATCAC-3ʼ
E2.1 5ʼ-TGATCGTGCCTGGTCAGAAC-3ʼ
5ʼ-CGCCTTAGCTGCTCTTCAGT-3ʼ
E2.2 5ʼ-CATTTGATCGGTTTAAACAGT-3ʼ
5ʼ-TAGATTTCATTTATAGGACTA-3ʼ
E3 5ʼ-GTGATCGTGCCTGGTCAGAA-3ʼ
5ʼ-GGTCTTCACTGGGCTTCCAA-3ʼ
E4 5ʼ-AGTGCCACGCAGTCTACAAA-3ʼ
5ʼ-CGGCCCAAAACAAAGAAGCA-3ʼ
E5 5ʼ-TGCTTCTTTGTTTTGGGCCG-3ʼ 5ʼ-CAAGGGTCTTCACTGGGCTT-3ʼ
Die Schmelztemperaturen von Primerpaar und Anlagerungsprodukt in Grad Celsius wurden entsprechend der „nearest neighbor thermodynamic theory“ nach folgender Formel errechnet:

2 Material und Methoden 29
Tm(°C) = {#H/ #S + R ln(°C)} - 273.15
Hier ist:
Tm die Schmelztemperatur,
#H der Enthalpieunterschied, errechnet anhand des nächsten Nachbarn;
#S der Entropieunterschied; bei der verwendeten salzhaltigen Pufferlösung entsprach dieser (1M NaCl) + 0.368 x N x ln([Na+])
Alle verwendeten Primer hatten eine errechnete Schmelztemperatur zwischen 54°C und 60°C. Die Bindungstemperatur (annealing Temperature, Ta) wurde anschließend nach folgender Formel berechnet:
Ta = 0.3 x Tm(Primer) + 0.7 Tm (Produkt) – 14.9
Wobei
- Tm (Primer) die Schmelztemperatur des Primers und
- Tm (Produkt) die Schmelztemperatur des Produkts ist (“Primer Design Guide for PCR :: Learn Designing Primers for PCR,” online abgerufen).
Die Bindungstemperatur lag bei allen Primerpaaren zwischen 48°C und 60°C. Der Cy-tosin/Guanin-Anteil aller Primer war größer als 50%. Das Design der Primer wurde mit Hilfe des Basic Local Alignment Search Tools (BLAST) auf Spezifität überprüft, um unspezifische Bindungen auszuschließen (BLAST basiert auf der Arbeit von (Altschul et al., 1990)).
2.2.3 Polymerase- Kettenreaktion (PCR) und Sequenzierung
Alle PCR wurden in 96-Well Platten (MicroAmp; Applied Biosystems), die mit opti-schen Versiegelungsfolien (Applied Biosystems; Cat No. 4313663) verschlossen wur-den, durchgeführt. Zuerst unterliefen alle Proben einen Amplifikationszyklus. Pro Probe wurde eingesetzt:
- PCR Buffer 2,6 $l
- PCR Nucleotid-Mix (1.25 mM) 2,2 $l
- MgCl2-Lösung (25 mM) 0,4 $l
- Primer fwd 0,2 $l
- Primer rev 0,2 $l
- TaqMan Gold 0,2 $l
- DNA-Lösung 1,0 $l
- Aqua ad injectabilia 4 $l

2 Material und Methoden 30
Zur Farbmarkierung für den Kapillarsequenzer wurde das ABI PRISM BigDye Termi-nator v3.1 Kit (Applied Biosystems) verwendet. Hier werden vor einem weiteren Zy-klus farbmarkierte Abbruchbasen beigemischt, welche die Längenbestimmung der Fragmente im Kapillarsequenzer ermöglichen. Der Ansatz wurde folgendermaßen pi-pettiert:
- Big Dye Premix 4 $l
- Primer (forward oder reverse je nach Ansatz) 0,2 $l
- Matrizen DNA 2$l
- TaqMan Gold 0,2 $l
- Aqua a.i. 3,6 $l
Anschließend erfolgte ein erneuter Zyklus, jeweils spezifisch für den verwendeten Pri-mer nach folgendem Schema: Prä-Denaturierung bei 96°, anschließend 28 Amplifikati-onszyklen mit Denaturierung, Annealing nach spezifischer Primertemperatur, Extending bei 60° und Abkühlung auf 4°C.
Die Proben wurden ein letztes Mal aufgereinigt (per Sephadex-Säule bei 4°C, Zentrifu-gation für 10 Minuten bei 2000 U/min).
Nun erfolgte die elektrophoretische Auftrennung auf dem ABI Prism 3100 Analyzer, einem automatischen Kapillarsequenziergerät mit 16 Kapillaren. Mithilfe der Data Col-lection Software 40100 (Version für Apple Macintosh OS 8) wurde aus den registrierten Emisionswerten ein Chromatogramm der Reihenfolge der Nukleotide erstellt.
Die automatische Auslesung des Chromatogramms ist fehleranfällig. Daher wurden alle Chromatogramme manuell ausgelesen (Sequencing Analysis Software, Version für Mac OS 8) und die Basensequenz der Matrizen-DNA notiert.
Insgesamt wurden alle 34 Proben für je 6 Exons mit beiden Primern (forward und rever-se) per PCR amplifiziert, sequenziert und manuell analysiert. Es wurden also 408 An-sätze (34 x 6 x 2) sowie für jeden Versuch Negativkontrollen pipettiert und untersucht.
Um die Kosten und den zeitlichen Einsatz abzuschätzen, muss man zu den 408 manuell ausgelesenen Genabschnitten noch - vor allem bei Beginn der Experimente –die Zahl der Ansätze hinzurechnen, die aufgrund von Verunreinigungen oder trotz aller Sorgfalt vorkommenden Fehlern verworfen werden mussten.

2 Material und Methoden 31
2.3 Auswertung
Alle gewonnenen DNA-Sequenzen wurden mit der Sequencing Analysis Software, Ver-sion für Mac OS 8 mit einer in Vorversuchen sequenzierten Wildtyp-Sequenz vergli-chen. Wenn Abweichungen zur Wildtyp-Sequenz gefunden wurden, wurden diese erst mit der single nucleotide polymorphism database (SNPdb) des NCBI abgeglichen, um Einzelnukleotidpolymorphismen (SNP) von Mutationen zu unterscheiden. Eine statisti-sche Übersicht über gefundene SNPs in der Studiengruppe findet sich im Abschnitt Re-sultate.
Für Abweichungen, die nicht in der SNPdb erfasst waren, wurden theoretische Überle-gungen angestellt, wie die mRNA-Sequenz (bzw. Expression) durch die Mutation be-einflusst wurde und ob sich Abweichungen für die Proteinstruktur ergeben würden.

3 Resultate Bei zwei der 34 untersuchten Proben wurden Mutationen im XIAP-Gen gefunden. Dies entspricht 5.8% der untersuchten Patienten. Beide Mutationen waren in der Literatur noch nicht beschrieben.
Von einem Patienten konnten auch der Bruder und die Mutter für eine XIAP-Gensequenzierung rekrutiert werden. Auch bei ihnen ließ sich die Mutation nachweisen. Somit wurden drei XIAP-defiziente Kinder und eine heterozygote Trägerin identifiziert.
3.1 Genveränderungen
3.1.1 Single Nucleotide-Polymorphismen
Bei 26 (76.5%) der untersuchten Patienten fand sich mindestens ein single nucleotide Polymorphismus im Bereich der untersuchten Exone. Alle SNPs befanden sich zwi-schen den Exonen vier bis sechs. Exon eins bis drei waren (mit Ausnahme einer der beiden Mutationen) frei von Wildtypabweichungen.
Eine Auflistung der Polymorphismen mit Häufigkeit erfolgt hier nach zugehöriger SNPdb-Nummer:
- rs 28382732: 9 Probanden
- rs 5956583: 13 Probanden
- rs 5958338: 3 Probanden
- rs 28382739: 9 Probanden
- rs 28382740: 10 Probanden
- rs 17334739: 5 Probanden
- rs 12838858: 2 Probanden
Die Verteilung der Polymorphismen innerhalb der Studengruppe zeigte im Abgleich mit der SNP database keine statistisch signifikanten Abweichungen im Vergleich zur Nor-malbevölkerung.
3.1.2 Mutationen
Zwei neuartige Mutationen im XIAP Gen wurden gefunden. In der ersten Familie fand sich eine vier Basenpaare lange Deletion an Position 347 der Basenfolge. Dies führt zu

3 Resultate 33
einem Austausch von Serin durch Leucin. Da es durch die Deletion von vier Basen auch zu einer Rasterverschiebung kommt, sind die nachfolgenden Triplets ebenfalls betrof-fen, und es kommt fünf Triplets später zum Einbau eines Stoppcodons. In Kurzschreib-weise: S347Lfs5X.
Bei der zweiten Familie lag eine R222X-Nonsense Mutation in der BIR2-Region des Gens vor. Hierbei wird durch einen Einzelnukleotid-Austausch von Cytosin durch Thymidin ein frühzeitiges Abbruchcodon erzeugt. Aus der Sequenz CGA, welche im Wildtyp-Protein für Arginin als 222te Aminosäure codiert, wird das Stoppcodon TGA und es kommt zum Abbruch der Aminosäurenkette an dieser Position.
3.2 Klinik der Patienten
Nach Identifikation der Mutationen wurden wie im Methodenteil beschrieben die be-handelnden Ärzte in Kenntnis gesetzt und mit deren Hilfe die Krankengeschichte so genau wie möglich rekonstruiert. Die betroffenen Patienten wurden durch die behan-delnden Ärzte zum Beratungs- und Aufklärungsgespräch eingeladen. Bei Familie I war dies Professor Behloradsky, LMU München, bei Familie II Professor Urban, Universi-tätskinder- und Jugendklinikklinik Graz.
3.2.1 Familie I: S347Lfs5x-Mutation
Die S347Lfs5X-Mutation wurde bei zwei Brüdern der Familie sowie deren Mutter mo-lekulargenetisch nachgewiesen. Bei der Mutter der beiden Knaben lag die Mutation heterozygot vor (Carrier-Status).
Anamnestisch war bei zwei Onkeln mütterlicherseits ebenfalls ein ähnliches Syndrom aufgetreten; diese waren bereits verstorben, so dass kein Gentest durchgeführt werden konnte. Für den Stammbaum der Familie siehe Abbildung 7.

3 Resultate 34
Abbildung 7 Stammbaum der Familie mit S347LfsX-Mutation. Die Mutation wurde zuerst bei Patient III/1 nachgewiesen. Schwarze Boxen zeigen betroffene Knaben; Punkte im Kreis kennzeichnen Konduk-torinnen; gestrichene Boxen stehen für verstorbene Verwandte.
3.2.2 Familie I: Mutter und zwei Onkel der Betroffenen (II/5, II/3 und II/2 im Stammbaum)
Bei der Mutter der beiden betroffenen Brüder (II/2 auf dem Stammbaum) fand sich die Mutation in heterozygoter Form. Die Krankengeschichte der Mutter war bis zum Ab-schluss der Untersuchung unauffällig.
Ein Bruder der Mutter (Patient II/3 auf dem Stammbaum) war vermutlich ebenfalls Genträger. 1974, als der Patient ein Jahr alt war, wurde bei ihm ein mit hohem Fieber einhergehender „Virusinfekt“ diagnostiziert. Im Anschluss entwickelte sich eine sekun-däre aplastische Anämie mit ausgeprägter Splenomegalie. Der Patient wurde damals mit einer Splenektomie und Intervallgaben von Kortikosteroiden behandelt, es konnte je-doch keine dauerhafte Verbesserung seines Zustandes erreicht werden. Im Alter von vier Jahren verstarb er an einer Meningitis mit Nebenniereninsuffizienz. Möglicherwei-se lag hier ein Waterhouse-Friderichsen-Syndrom vor.
Ein Cousin der Mutter (Patient II/5 auf dem Stammbaum) litt, seit er zweieinhalb Jahre alt war, unter ausgeprägter Hepatosplenomegalie. Bei diesem Patienten wurde im Alter

3 Resultate 35
von neun Jahren eine Splenektomie durchgeführt, woraufhin sich sein Zustand deutlich verbesserte. Er verstarb im Alter von 25 Jahren an einem blutenden Magenulkus. Die Diagnose XLP-2 ist bei beiden Patienten wahrscheinlich; es lag jedoch keine genomi-sche DNA vor, so dass die Verdachtsdiagnose nicht verifiziert werden konnte. Auf-grund der Klinik wurden beide Patienten in der Veröffentlichung beschrieben.
3.2.3 Familie I: 27 Jahre alter, positiv getesteter Erwachsener (Patient III/1 im Stammbaum)
Im Alter von sechs Jahren trat bei dem Knaben eine akute infektiöse Mononukleose mit hohem Fieber, Ausschlag, Pharyngitis, Schwellung der Halslymphknoten, Perimyocar-ditis und Hepatosplenomegalie auf. Die Hepatomegalie lag in der Ausprägung zwischen 8 und 10 cm unterhalb der Rippengrenze. Im Blutausstrich zeigte sich das Bild einer Panzytopenie; die Hämoglobinkonzentration lag bei 8.8 g/dL, die Leukozytenzahl bei 2400/$L, Thrombozyten bei 76.000/$L. Sowohl in der Knochenmarkbiopsie als auch im Blutausstrich konnte die Phagozytose von Erythroblasten und Thrombozyten nach-gewiesen werden (siehe Abbildung 1 im Einleitungsteil).
Neben der Diagnose des hämophagozytären Syndroms wiesen serologische Studien auf eine primär persistierende EBV-Infektion hin: Anti-Virus-Capsid-Antigen (VCA)-IgG lag bei 1:1024, anti-VCA IgM bei 1:60, anti-VCA IgA bei 1:32; anti early Antigen (EA) bei 1:16 und anti nuclear Antigen (EBNA) war negativ.
In den quantitativen Serumimmunglobulinmessungen zeigten sich zu Beginn Serum-werte von IgG, IgM und IgA im unteren Normbereich. In den folgenden Monaten fiel IgG jedoch auf auf 382 mg/dL (normal für Sechsjährige: 528 - 1064 mg/dL), IgM auf 18 mg/dL (normal 75 - 161 mg/dL) und IgA auf 23 mg/dL (normal 62 - 192 mg/dL). Daraufhin wurden monatliche intravenöse Immunglobulingaben (IVIG) begonnen; in der Akutsituation wurde mit Aciclovir und Kortikosteroiden behandelt. Nach initial gutem Ansprechen kam es fünf Monate später zu einem Rezidiv mit Panzytopenie, hä-mophagozytärem Syndrom, Fieber und Splenomegalie. Eine erneute EBV-Serologie zeigte einen sekundären Verlust von EBNA-Antikörpern und einen signifikanten An-stieg der EA-Antikörper (1:64), was als insuffiziente Kontrolle der EBV-Infektion ge-wertet wurde. Erneute Kortikosteroidgabe führte zu einer raschen und bleibenden Bes-serung, mit Ausnahme einer persistierenden Splenomegalie.
3.2.4 Familie I: 29 Jahre alter, positiv getester Erwachsener (Patient III/2 im Stammbaum)
Beim älteren Bruder des Indexpatienten (Patient III/2, derzeit 29 Jahre) war eine Sple-nomegalie vom ersten Lebensjahr an bekannt. Es wurde berichtet, dass die Milzgröße während Infektionen wie Erkältungen vorübergehend zunahm. Im Alter von drei Jahren

3 Resultate 36
entwickelte dieser Patient eine akute idiopathische thrombozytopenische Purpura (ITP), die ohne Behandlung wieder abklang. Im Knochenmarksaspirat im Alter von vier Jah-ren fand man Erythrophagozytose. Zu diesem Zeitpunkt war eine EBV-Serologie kom-plett negativ, und die Serumimmunglobulinwerte im Normalbereich. Im Alter von fünf Jahren machte der Patient eine schwere akute Mononukleose mit hohem Fieber, Hals- und Bauchlymphknotenschwellung und Ausschlag, aber ohne Mandelentzündung durch. Eine Serologie zu diesem Zeitpunkt zeigte ein ähnliches Bild wie bei seinem Bruder (anti-VCA-IgG 1:128, anti-VCA-IgM 1:32, anti-EA 1:32). Im Differentialblut-bild fand man eine milde Anämie (Hämoglobin 9.6 g/dL) und Leukopenie (3100/$L) bei niedrignormalen Thrombozytenwerten. Serum-Transaminasen waren erhöht; LDH lag mit 1311 U/L bei ähnlich hohen Werten wie bei dem Patienten mit R222X-Mutation (siehe unten). Die Serumimmunglobuline waren jetzt im unteren Normalbereich (IgG 573 mg/dL; IgM 118 mg/dL; IgA 42 mg/dL).
Wie sein Bruder (III/1) sprach der Patient sehr gut auf eine Behandlung mit Kortikoste-roiden an. Allerdings fielen die Immunglobulinwerte wie bei seinem Bruder auch trotz gutem klinischen Verlauf stetig weiter ab. Da er unter rezidivierenden Infekten litt, wurden auch bei ihm monatliche IVIG-Gaben begonnen. Hierunter war der Patient symptomfrei bis zum 21. Lebensjahr. Zu diesem Zeitpunkt entwickelte er ein schweres papulopustulöses Exanthem des Gesichts (siehe Abb. 7 a und b). Nach Biopsie wurde die Diagnose einer bakteriell superinfizierten Demodex-Follikulitis gestellt (Benoit et al., 2003). Demodex Follikulorum ist eine fakultativ pathogene Hautmilbe, die bei Er-wachsenen ubiquitär ist; die Krankheitsentstehung wird vom Immunzustand des Befal-lenen und der Dauer der Erkrankung maßgeblich beeinflusst. In diesem Fall wurde die Hautreaktion in Zusammenhang mit der verminderten Immunkompetenz des Patienten gebracht. Die Hautentzündungen sind bis heute rezidivierend; davon abgesehen ist auch dieser Patient unter Therapie symptomfrei (Abb. 7c).
Abbildung 8 A und B: Schuppige, zu Plaques konfluierende Papulopusteln an Gesicht, Hals und Ohren. C: Resterythem im Gesichtsbereich nach viermonatiger systemischer Behandlung mit Metronidazol und Isotretinoin (Benoit et al., 2003)

3 Resultate 37
3.2.5 Familie II: 17 Jahre alter Knabe mit R222X-Mutation
Der Patient mit der R222X Mutation war ein Einzelkind; wir konnten die Mutation he-terozygot auch bei seiner asymptomatischen Mutter nachweisen (Konduktorin). Ana-mnestisch waren keine Familienmitglieder mit einem ähnlichen Syndrom bekannt, al-lerdings gab es kaum männliche Verwandte auf Seite der Mutter.
Die Mutation wurde bei einem seinerzeit 17 Jahre alten Jungen nachgewiesen. Er war im Alter von vier Jahren mit Fieberschüben bis 40°C, Verwirrtheit, mildem Husten und Rhinorrhoe über einen Zeitraum von zwei Wochen in das Kinderkrankenhaus überwie-sen worden. Dort wurde eine Panzytopenie (Hämatokrit 26%, 2900 Leukozyten/$L, 126.000 Thrombozyten/$L) diagnostiziert und der Patient stationär behandelt. Bei der Aufnahmeuntersuchung zeigte sich ein akut kranker, blass wirkender, aber normal ent-wickelter Vierjähriger (das Körpergewicht lag mit 17 kg auf der 50ten Perzentile, die Größe mit 105cm auf der 75ten Perzentile). Die Vitalparameter waren von einer mäßi-gen Tachykardie (130/min) abgesehen unauffällig, die rektal gemessene Temperatur lag bei 37.8°C. CRP war auf 16.1 mg/L erhöht (normal bis 8 mg/L). Albumin, Kreatinin, Harnstoff und Harnsäure lagen alle im Normbereich. Bei den Leberwerten war die Se-rum Aspartat-Aminotransferase (AST/GOT) mit 62 U/L erhöht (normal bis 43 U/L), die Alanin-Aminotransferase (ALT/GPT) mit 31 U/L (normal bis 45 U/L) im Normalbe-reich; die Cholestaseparameter gamma-Glutamyltransferase und alkalische Phosphatase waren ebenfalls erhöht (gamma-GT 45 U/L, normal bis 28 U/L; AP 336 U/L, normal bis 250U/L). Die Lactatdehydrogenase (LDH) lag bei 1881 U/L (normal bis 240 U/L); auch Ferritin war drastisch erhöht (7147$g/L; normal bis 140$g/L). Fibrinogen als weiteres akute-Phase-Protein war hingegen auf 120 mg/dL erniedrigt (altersangepasster Nor-malwert zwischen 170 und 400 mg/dL). Die Triglyzeride lagen bei 311 mg/dL (normal bis zu 150 mg/dL).
Eine Lumbalpunktion mit Analyse des cerebrospinalen Liquors war unauffällig. Ein EBV-monospot-Test erwies sich als positiv. Es wurde eine EBV-Serologie veranlasst. Serum IgM- und IgG Antikörper gegen Epstein Barr Virus Capsid Antigen (VCA) wa-ren postiv. Antikörper gegen early antigen (EA) und nuclear antigen (EBNA) waren nicht nachweisbar. Eine EBV-DNA PCR aus peripheren Blutzellen war negativ, jedoch postiv in einer Knochenmarksbiopsie.
Quantitative Serum-Immunglobulinmessungen ergaben folgende Werte: IgM leicht er-höht (212 mg/dL, normal für diese Altersgruppe 52 – 200 mg/dl) IgG und IgA im Normbereich. Der lösliche Interleukin-2-Rezeptor (IL-2R) war deutlich erhöht (3546, normal bis zu 300 U/mL).
Flusszytometrieuntersuchungen der peripheren Lymphozyten zeigten eine relative Ver-minderung der CD4+ Zellen (1000/$L, 18% der T- Lymphozytenzahl; Normalwert 32 - 62%) und eine absolute wie relative Vermehrung der CD8+ Zellen (4226/$L, 76% der

3 Resultate 38
T-Lymphozytenzahl; Normalwert 13-45%) mit einer resultierenden Verminderung des CD4/CD8 Quotienten auf 0.24 (normal 1-1.4). Natürliche Killerzellen waren deutlich vermindert (35/$L, 2% der absoluten Lymphozytenzahl; normal 8-22%) und die NK-Zell-Aktivität war beinahe vollständig erloschen.
In der Knochenmarkbiopsie zeigten sich eine erhöhte und etwas dysplastische Granulo- und Erythropoese und aktive hämophagozytäre Zellen, so dass die Diagnose eines hä-mophagozytären Syndroms gestellt wurde.
In der Bauchsonografie fanden sich eine grenzwertig vergrößerte, homogene Leber, eine mittelgradig vergrößerte Milz und etwas freie intraperitoneale Flüssigkeit.
In den ersten acht Tagen nach der Aufnahme litt der Patient unter Fieberspitzen von mehr als 40°C. Nach der Diagnosestellung eines EBV-assozierten hämophagozytären Syndroms wurde eine medikamentöse Therapie begonnen. Der Patient erhielt wieder-holte intravenöse Hochdosis-Immunglobulingaben (5 x 18g im ersten Monat, dann eine Einzeldosis von 6g, anschließend 6 x 3g über die folgenden sechs Wochen) und intrave-nöses Ganciclovir (150mg täglich über die erste Woche, anschließend jeden dritten Tag für die nächsten 3 Monate; eine orale Gabe war nicht möglich, da sich der Patient wei-gerte Tabletten zu schlucken). Unter Therapie klang das Fieber ab und der Zustand des Patienten besserte sich drastisch. Bei Entlassung war er klinisch erholt und die Panzyto-penie, die Leberwerte, Triglyzeride, Fibrinogen und Ferritin hatten sich vollständig normalisiert. Die Verdachtsdiagnose XLP wurde geäußert und Blut zur Genanalyse des SH2D1A-Gens eingeschickt, aber keine Mutation gefunden.
Dreizehn Jahre nach der ersten Aufnahme wurde der Patient in unsere Studie einge-schlossen und oben genannte Mutation gefunden. Bei einer erneuten Untersuchung zeigte sich ein normalentwickelter 17 Jahre alter junger Mann. Von einer milden persi-stierenden Splenomegalie im Ultraschall (12 x 5cm) abgesehen gab es keine klinischen Auffälligkeiten.
3.3 Übersicht über die Klinik der Patienten
Die klinischen Daten aus Kapitel 3.2 sind in der Tabelle auf der nächsten Seite zusam-mengefasst.

Mutation
Stammbaum
Familie II
R222X
-
Familie I
S347Lfs5X
III-2
Familie I
S347Lfs5X
III-1
Familie I
nicht untersucht
II-3
Familie I
nicht untersucht
II-6
Derzeitiges Alter 18 29 27 Tod mit 4 Jahren (anno 1968) Tod mit 25 Jahren (anno 1976)
Schwere Mononukleose Ja (4) Ja (6) Ja (5) Möglich, “Schwere Virusin-fektion”
Nicht bekannt
Fatale Mononukleose Nein Nein Nein Nein Nein
Hämophagozytäres Syndrom Ja (4) Ja (6) Ja (4 – vor einer nachweisli-chen EBV-Infektion)
Nicht bekannt Nicht bekannt
Aplastische Krise Nein Ja Nein Nicht bekannt, “Aplastische Anämie”
Nicht bekannt
Splenomegalie Ja (4) Mittelgradig Ja (6) 10cm Ja (ab der frühen Kindheit, vor EBV-Infektion)
Ja (1) Ja (2)
Hepatomegalie Ja (4)
Mild
Ja (6) 8 cm Nein Nicht bekannt Ja (2)
Persistierende Hypogamma-globulinämie
Nein Ja (6) IgG 382; IgA23; IgM 18
Nein; während EBV-Infektion IgG 573, IgA 42, IgM 118
Nicht bekannt Nicht bekannt
Superinfizierte Hautläsionen Ja Nein Ja Nicht beschrieben Nicht beschrieben
XIAP-Mutation Nonsense, R222X (CGA->TGA)
S347L, fs5X S347L, fs5X Nicht getestet Nicht getestet
Todesursache - - - Meningitis und Nebennieren-versagen
Blutendes Magenulkus
Lymphom Nein Nein Nein Nein Nein
Tabelle 4 Übersicht über die klinischen Symptome der Patienten. Die Nummern in Klammern geben das Alter des Patienten bei der Manifestation des jeweiligen Symptoms an.

4 Diskussion In der Einleitung (Kapitel Ziele, 1.1 und Eingeschlagener Lösungsansatz, Error! Refe-rence source not found.) wurden drei Thesen formuliert, deren Gültigkeit in diesem Abschnitt anhand der gefundenen Ergebnisse überprüft werden soll. Die Ergebnisse werden im Kontext der Literatur diskutiert.
Die drei Thesen lauteten:
1. (Hauptthese): Hauptthese dieser Arbeit war, dass die von Rigaud et. al entdeckte Erkrankung auch im untersuchten Patientenkollektiv nachgewiesen werden kann. Es sollte bewiesen werden, dass im genetischen Labor der Frauenklinik der Technischen Universität München, in dem von Professor Meindls Arbeits-gruppe Untersuchungen auf SAP-Defizienz durchgeführt wurden, unter den hierfür negativ getesteten Patienten ein gewisser Anteil an XIAP-Defizienten verborgen lag.
2. (Nebenthese): Bei XIAP-Defizienz werden im Gegensatz zur SAP-Defizienz keine Lymphome beobachtet
3. (Nebenthese): Bei der Therapie von XIAP-Patienten kann aufgrund der erhöhten Apoptoserate ein konservativer Ansatz günstiger als eine Knochenmarkstrans-plantation sein.
4.1 Hauptthese: Im Patientenkollektiv können XIAP-Gen Mutationsträger identifiziert werden
Die Hauptthese konnte bewiesen werden. Durch die Untersuchungen wurden drei Pati-enten mit XIAP-Defizienzsyndrom identifiziert. Zwei weitere Patienten, die verstorben waren, litten möglicherweise ebenfalls an dem Syndrom. Bis dato waren nur die zwölf Patienten aus der Orginalarbeit von Rigaud et al sowie ein weiterer, von Zhao beschrie-bener Patient mit XLP-2 bekannt (Rigaud et al., 2006; Zhao et al., 2010). Die hier be-schriebenen Patienten waren also unter den ersten Fünfzehn, die mit dem Syndrom dia-gnostiziert wurden. Nach dem Erscheinen der dieser Promotion zu Grunde liegenden wissenschaftlichen Veröffentlichung wurden noch weitere Patientendaten von gesicher-ten Mutationsträgern veröffentlicht, die ebenfalls in die Diskussion einfließen sollen. Bisher haben vier Arbeitsgruppen Mutationsträger im XIAP-Gen identifizieren können und deren Krankengeschichte veröffentlicht, so dass heute weltweit 54 Patienten be-kannt sind. Diese Patienten setzen sich wie folgt zusammen:
- Paris, Frankreich, Arbeitsgruppe um Latour, Rigaud: 30 Patienten (*)

4 Diskussion 42
- Cincinnati, USA, Arbeitsgruppe Marsh et al.: 10 Patienten
- Toyama, Japan, Arbeitsgruppe um Yang, Zhao: 9 Patienten
- München/Leipzig/Graz, (unsere Arbeitsgruppe): 3/5 Patienten (*)
* In der Patientengruppe aus Paris ist eine nicht näher genannte Anzahl der Patienten nicht genetisch, sondern über den Stammbaum identifiziert worden. Von den amerikani-schen und japanischen Arbeitsgruppen liegen bei allen Patienten positive Gentests vor. In unserer Gruppe sind drei Patienten über einen positiven Test identifiziert, während zwei über den Stammbaum identifiziert wurden (Horn et al., 2011; Marsh et al., 2010; Pachlopnik Schmid et al., 2011; X. Yang, Miyawaki, & Kanegane, 2012). Diese unter-schiedlichen Angaben über die Patienten können dazu führen, dass nicht bei allen Ver-gleichen alle 54 Patienten berücksichtigt werden.
Es war zu Beginn der Arbeit nicht abzusehen, ob die Hypothesen verifiziert werden können. Der folgende Absatz soll anhand der Literatur helfen abzuschätzen, wie hoch die Erfolgsaussichten waren und ob ein ähnlicher Ansatz für andere Arbeitsgruppen sinnvoll ist.
4.1.1 Prozentualer Anteil positiv getesteter Proben und Vergleich mit der Literatur
Bei 34 untersuchten Proben fanden sich zwei Mutationen (5,9%) Die Frage, weshalb nicht alle Proben Genmutationen zeigten lässt sich in drei Punkten beantworten:
1. Auswahlbias der Proben durch die Einsender: Es wurden für die Arbeit nur Pro-ben untersucht, die zuvor negativ auf eine Mutation im SAP-Gen getestet hatten. Alle Proben waren demnach eingesandt worden, weil die behandelnden Ärzte den Verdacht auf XLP-1 geäußert hatten. XIAP-Defizienz war zum Zeitpunkt der Einsendungen nicht bekannt bzw. charakterisiert. Die Auswahl durch die Einsendenden litt entsprechend unter einem Selektionsbias hin zu einer anderen Erkrankung. Weiter wurde nicht nach festgelegten Regeln wie beispielsweise ei-nem entsprechendem Stammbaum bei bestimmten klinischen Kriterien unter-sucht, sondern war der klinische Verdacht der behandelnden Ärzte für die Einsendung entscheidend.
2. Auswahlbias der Proben durch den Doktoranden: Wie in Abschnitt 2.1, Auswahl der zu untersuchenden Proben beschrieben, wurden nicht alle SAP-Mutations-negativen Probanden untersucht. Nur aus Fällen mit vorliegender Einwillung und ausreichender klinischer Beschreibung wurden aufgrund der damals vorlie-genden Daten zur XIAP-Defizienz erfolgversprechende Fälle ausgewählt. Geht man davon aus, dass die Selektion gut erfolgt ist, wird der Anteil positiver Pro-ben möglicherweise überschätzt.

4 Diskussion 43
3. Weitere Syndrome mit ähnlichem Krankheitsbild sind denkbar: XIAP-Defizienz war entdeckt worden, nachdem in vielen Verdachtsfällen mit eindeutigem Stammbaum kein XLP-1 Syndrom nachgewiesen werden konnte. Genauso wäre denkbar, dass Muationen auf anderen Genen ein ähnliches Syndrom verursa-chen, die bis heute noch nicht entdeckt worden sind. Beispielsweise fanden sich unter den Proben, die auf SAP-Defizienz untersucht wurden auch die von zwei Schwestern, die eine tödliche lymphoproliferative Reaktion auf eine EBV-Infektion enwickelt hatten. Die Untersuchung dieser Proben durch eine andere Arbeitsgruppe führte zur Entdeckung einer Mutation im IL-2-inducible T cell kinase (ITK)-Gen (Huck et al., 2009). Bei Verdacht auf ein autosomal vererbtes Syndrom oder wenn der Erbgang nicht erhoben werden kann, sollte neben SAP- und XIAP- auch ITK-Defizienz differentialdiagnostisch in Betracht gezogen werden.
Zur Auswahl durch den Doktoranden ist anzumerken, dass eine Mutation bei derjenigen Familie gefunden wurde, bei der vor der Untersuchung der dringendste klinische Ver-dacht auf XIAP-Defizienz bestanden hatte. Die andere Mutation fand sich bei einem Probanden, der sich klinisch nicht wesentlich von den anderen untersuchten Probanden abgehoben hatte. Es ist anzunehmen, dass die prozentuale Angabe der „Trefferquote“ von ca. 6% in anderen Patientenpopulationen deutlich abweicht und sich mit der Vor-auswahl entsprechend ändert, je besser die Klinik der XIAP-Defizienz charakterisiert wird.
In der Literatur findet sich nur eine weitere Studie mit vergleichbarem Ansatz und von vergleichbarer Größe aus China (Zhizhuo et al., 2012). Hier wurde ebenfalls versucht, bei Kindern, die mit EBV-assoziierter hämophagozytärer Lymphhistiozytose auffällig geworden waren, Mutationen zu finden. Es wurden neben SH2D1A, XIAP und ITK auch Gene für familiäre hämophagozytäre Lymphhistiozytose (fHLH) wie PRF1, UNC13D und STX11 gescreent. Daher waren auch Mädchen eingeschlossen. Eine Un-tersuchung auf XIAP-Mutation erfolgte bei insgesamt 32 männlichen Probanden, auf SAP-Mutation bei 38. In dieser Population wurde ein Patient mit XIAP-Mutation und keiner mit SAP-Mutation identifiziert (Zhizhuo et al., 2012). Dies entspricht einem Pro-zentsatz von 3.1% für XLP-2. Zu bedenken ist, dass diese Studie keine (vermutete) ge-netische Komponente als Einschlusskriterium benutzte, sondern einzig Kinder unter-suchte, die mit einem EBV-reaktiven klinischen Syndrom auffällig geworden waren. Weitere Erklärungen für den Unterschied zu der vorgelegten Arbeit könnten sein, dass
- die chinesische Studie auf den westlichen Raum nicht ohne weiteres übertragbar ist, da der unterschiedliche Genpool bei seltenen genetischen Erkrankungen stark ins Gewicht fallen kann
- es sich bei den Fallzahlen um statistische Schwankungen handelt

4 Diskussion 44
Genau wie in der vorgelegten Arbeit konnte jedoch auch in der Untersuchung aus China nur ein geringer Prozentsatz der Verdachtsfälle bestätigt werden. Dies war zu Beginn der Arbeit bereits erwartet worden. Die Identifikation von zwei nicht beschriebenen Mutationen im XIAP-Gen sowie die Identifikation von damals drei aus weniger als zwanzig weltweit bekannten Patienten kann als großer Erfolg gewertet werden.
4.1.2 Vergleich der erhobenen klinischen Daten mit der Literatur
Wie in der Einleitung beschrieben, war zu Beginn der Arbeit keine klare Abgrenzung zwischen SAP-Defizienz und XIAP-Defizienz (damals XLP-1 und XLP-2) möglich, was maßgeblich an der geringen Zahl der identifizierten Mutationsträger lag. Beim Ein-reichen der Veröffentlichung handelte es sich um die 14. bis 18 Patienten weltweit. Während des Review-Prozesses wurden noch die Daten von zehn weiteren Patienten aus den USA veröffentlicht (Marsh et al., 2010), so dass diese in die Auswertung mit einfließen konnten. Insgesamt waren zum Zeitpunkt der Publikation also 28 Patienten bekannt; für die statistische Auswertung rechneten wir nur 26 Patienten ein, da bei den beiden verstorbenen Patienten aus unserer Gruppe mehrere klinische Angaben nachträglich nicht zu erheben waren. Tabelle 5 ist der Veröffentlichung im JACI entnommen und fasst die Klinik der zum Erscheinen der Veröffentlichung bekannten Patienten zusammen.
Tabelle 5: Aus der dissertationsbegleitenden Publikation: Übersicht über die Klinik aller bisher bekannten XIAP-Defizienz-Patienten.
Vergleicht man die klinischen Merkmale unserer Patientengruppe mit dem Gesamtkol-lektiv aller bekannten Patienten zeigt sich eine deutliche Übereinstimmung vor allem im Auftreten von HLH und Splenomegalie. Beide scheinen wesentliche Charakteristika der XIAP-Defizienz zu sein. Eine EBV-Assoziation trat in unserem Kollektiv häufiger auf als in der Gesamtgruppe (100% vs. 58%), was möglicherweise an der Auswahl der Pati-enten (klinischer Verdacht auf SAP-Defizienz) lag. Die von Rigaud et al. beobachtete Colitis wurde von unserer und den weiteren Arbeitsgruppen nicht beobachtet.
Die nächste Tabelle (
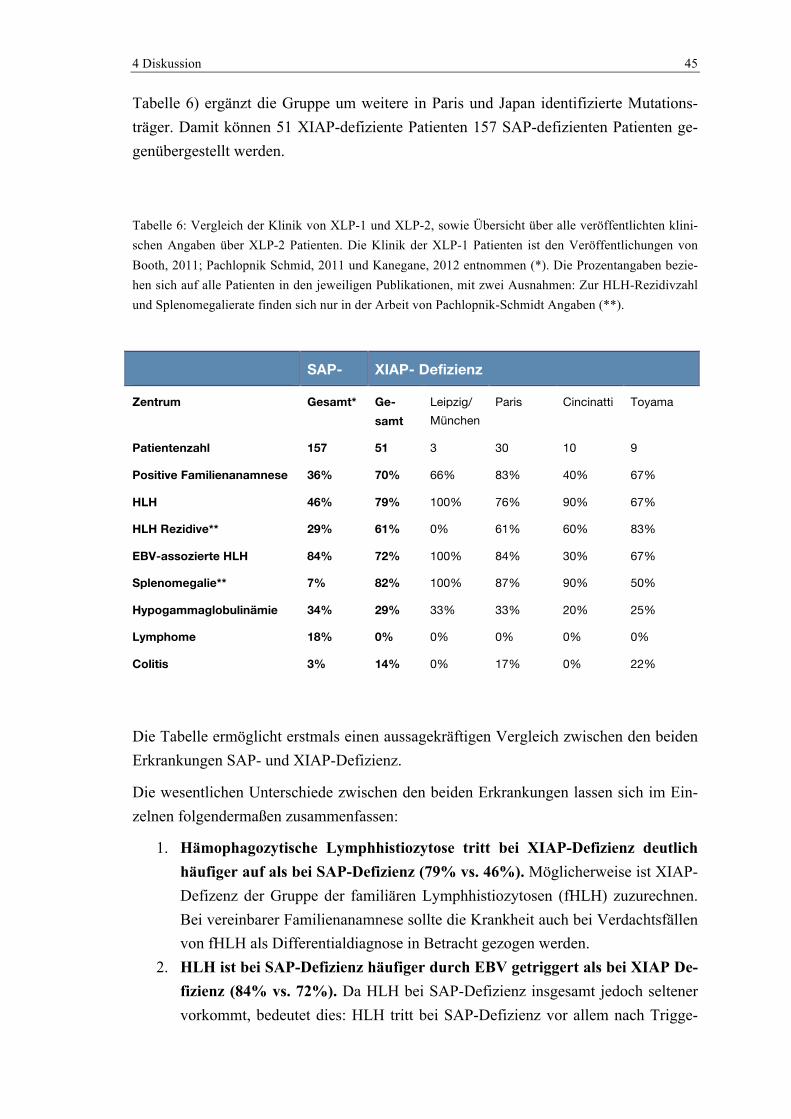
4 Diskussion 45
Tabelle 6) ergänzt die Gruppe um weitere in Paris und Japan identifizierte Mutations-träger. Damit können 51 XIAP-defiziente Patienten 157 SAP-defizienten Patienten ge-genübergestellt werden.
Tabelle 6: Vergleich der Klinik von XLP-1 und XLP-2, sowie Übersicht über alle veröffentlichten klini-schen Angaben über XLP-2 Patienten. Die Klinik der XLP-1 Patienten ist den Veröffentlichungen von Booth, 2011; Pachlopnik Schmid, 2011 und Kanegane, 2012 entnommen (*). Die Prozentangaben bezie-hen sich auf alle Patienten in den jeweiligen Publikationen, mit zwei Ausnahmen: Zur HLH-Rezidivzahl und Splenomegalierate finden sich nur in der Arbeit von Pachlopnik-Schmidt Angaben (**).
Die Tabelle ermöglicht erstmals einen aussagekräftigen Vergleich zwischen den beiden Erkrankungen SAP- und XIAP-Defizienz.
Die wesentlichen Unterschiede zwischen den beiden Erkrankungen lassen sich im Ein-zelnen folgendermaßen zusammenfassen:
1. Hämophagozytische Lymphhistiozytose tritt bei XIAP-Defizienz deutlich häufiger auf als bei SAP-Defizienz (79% vs. 46%). Möglicherweise ist XIAP-Defizenz der Gruppe der familiären Lymphhistiozytosen (fHLH) zuzurechnen. Bei vereinbarer Familienanamnese sollte die Krankheit auch bei Verdachtsfällen von fHLH als Differentialdiagnose in Betracht gezogen werden.
2. HLH ist bei SAP-Defizienz häufiger durch EBV getriggert als bei XIAP De-fizienz (84% vs. 72%). Da HLH bei SAP-Defizienz insgesamt jedoch seltener vorkommt, bedeutet dies: HLH tritt bei SAP-Defizienz vor allem nach Trigge-
SAP- XIAP- Defizienz
Zentrum Gesamt* Ge-
samt Leipzig/
München Paris Cincinatti Toyama
Patientenzahl 157 51 3 30 10 9
Positive Familienanamnese 36% 70% 66% 83% 40% 67%
HLH 46% 79% 100% 76% 90% 67%
HLH Rezidive** 29% 61% 0% 61% 60% 83%
EBV-assozierte HLH 84% 72% 100% 84% 30% 67%
Splenomegalie** 7% 82% 100% 87% 90% 50%
Hypogammaglobulinämie 34% 29% 33% 33% 20% 25%
Lymphome 18% 0% 0% 0% 0% 0%
Colitis 3% 14% 0% 17% 0% 22%

4 Diskussion 46
rung durch eine EBV-Infektion auf. Bei XIAP-Defizienz wird HLH ebenfalls häufig durch eine EBV-Infektion getriggert, tritt jedoch auch spontan auf. Ent-sprechend ist auch die Rezidivrate von HLH bei XIAP-Defizienz höher (61% vs. 29%).
3. Splenomegalie ist ein charakteristisches Merkmal der XIAP-Defizienz und wird bei SAP-Defizienz nur selten beobachtet (82% vs 7%).
4. Hypogammaglobulinämie wird bei beiden Erkrankungen etwa gleich häu-fig (34% vs 29%) beobachtet und ist möglicherweise sekundäre Folge der im-munologischen Veränderungen.
5. Lymphome treten nur bei SAP-Defizienz auf (18%). Siehe hierzu auch Ab-satz 4.2.
6. Autoimmuncolitis kann bei beiden Krankheitsbildern beobachtet werden, scheint jedoch nicht häufig zu sein. Direkt nach der Veröffenltichung von Ri-gaud et al. war gemutmaßt worden, dass eine Automimmuncolitis charakteri-stisch für die XIAP-Defizienz sei. In unserem und dem amerikanischen Patien-tenkollektiv war eine Autoimmuncolitis jedoch nie aufgetreten, in der asiati-schen Gruppe etwa gleich häufig wie in der französischen. Dass Autoimmunco-litis ein Alleinstellungsmerkmal von XIAP-Defizienz sei, wurde in einem Re-view von Yang et al. 2012 beschrieben (X. Yang et al., 2012). Dies kann jedoch wiederlegt werden, es sind Fälle der SAP-Defizienz mit Colitis, Gastritis, Ga-stroenterocolitis und postentzündlicher intestinaler Metaplasie beschrieben (Booth et al., 2011; Filipovich et al., 2010; Horn & Schuster, 2012; Rougemont et al., 2008).
Die aufgeführten Punkte verdeutlichen, dass es sich bei XIAP-Defizienz und SAP-Defizienz um zwei unterschiedliche Erkankungen handelt. Die Nomenklatur XLP-1 und XLP-2 ist irreführend und sollte vermieden werden.
4.2 Nebenthese: Bei XIAP-Defizienz werden im Gegensatz zur SAP-Defizienz keine Lymphome beobachtet
Eine Nebenthese der Arbeit lautete, dass die bei SAP-Defizienz beobachteten Lympho-me bei der XIAP-Defizienz nicht auftreten. Bei den ersten zwölf von Rigaud et al publi-zierten Patienten waren keine Malignome beobachtet worden und auch Beobachtungen am Tiermodell legten die Annahme nahe, dass XIAP-Defizienz einer Entartung entge-gen wirkt. Beispielsweise konnte für diffus großzellige B-Zell-Lymphome nachgewie-sen werden, dass XIAP-Antagonisten die Apoptose wieder in Gang setzen können und zu einer Verkleinerung des Tumors führen (Cillessen et al., 2008). Nodal diffuse groß-zellige B-Zell-Lymphome zeigen eine erhöhte Apoptoserate und werden sensibler ge-

4 Diskussion 47
genüber Chemotherapie, wenn XIAP aufgrund äußerer oder zellulärer Einflüsse unte-rexprimiert ist (Muris et al., 2005). Versuche an der Zellkultur für das Hodgkin-Lymphom demonstrieren, dass auch hier eine XIAP-Herabregulierung - in diesem Fall durch Bortozemib - die Tumorzellen empfänglicher sowohl für die Lyse durch T-Zellen (Kashkar et al., 2006) als auch für die Apoptoseinduktion durch Zytotherapeutika macht (Kashkar et al., 2007).
Bis jetzt ist in der Literatur kein Patient mit XIAP-Mutation beschrieben, der ein Lymphom entwickelt hätte. Auch wenn eine Tumorentwicklung aufgrund von Persi-stenz von EBV oder reaktiven Veränderungen des Immunsystems nicht ausgeschlossen werden kann, ist diese sehr viel unwahrscheinlicher als bei einer SAP-Mutation, und nicht Teil des Krankheitsbildes. Diese Feststellung ist klinisch bedeutsam, da Sie die Entscheidung für oder gegen eine Knochenmarkstransplantation beinflussen kann. Dies leitet über zur zweiten Nebenthese, welche im peer-review der Publikation für die größ-te Kontroverse sorgte: Die Annahme, dass XIAP-Defizienz mit konservativer Therapie ausreichend kontrollierbar und eine Stammzelltransplantation nicht zwingend notwen-dig sei.
4.3 Nebenthese: Bei der Therapie kann ein konservativer Ansatz einer Knochenmarkstransplantation vorzuziehen sein
Bei SAP-Defizienz ist eine Knochenmarkstransplantation die Therapie der Wahl. Die Mortalität liegt bei 29% (Ochs, Smith, & Puck, 2007). Aufgrund der schweren Konse-quenzen bei SAP-Defizienz ist sie jedoch klar indiziert und erfolgt prophylaktisch bei Genträgern.
Wie die Therapie bei XIAP-Defizienz aussehen sollte, ist noch umstritten. Prinzipiell kommt eine supportive, medikamentöse Therapie oder eine Stammzelltransplantation in Frage.
Als unsere Arbeit in den Review-Prozess ging, hatten wir aufgrund der Erfahrungen mit unseren Patienten und der verfügbaren Literatur den Verdacht geäußert, dass XIAP-Defizienz/XLP-2 einen milderen Verlauf haben könnte als SAP-Defizienz/XLP-1; ins-besondere dann, wenn HLH bereits aufgetreten war und die Patienten – falls notwendig - eine Immunglobulinsubstitution erhielten. Verbunden war dies mit dem Hinweis auf die hohe Komplikationsrate und Mortalität einer Knochenmarktransplantation.
Dies wurde im Reviewprozess kritisch aufgenommen. Einer der drei Erstgutachter be-stand auf einer Löschung der Aussagen; zu diesem Zeitpunkt waren die Patienten aus Cincinnati noch nicht veröffentlicht. Als diese hinzukamen und ein vierter Gutachter hinzugezogen wurde, konnten wir uns darauf einigen, eine objektive Zusammenfassung
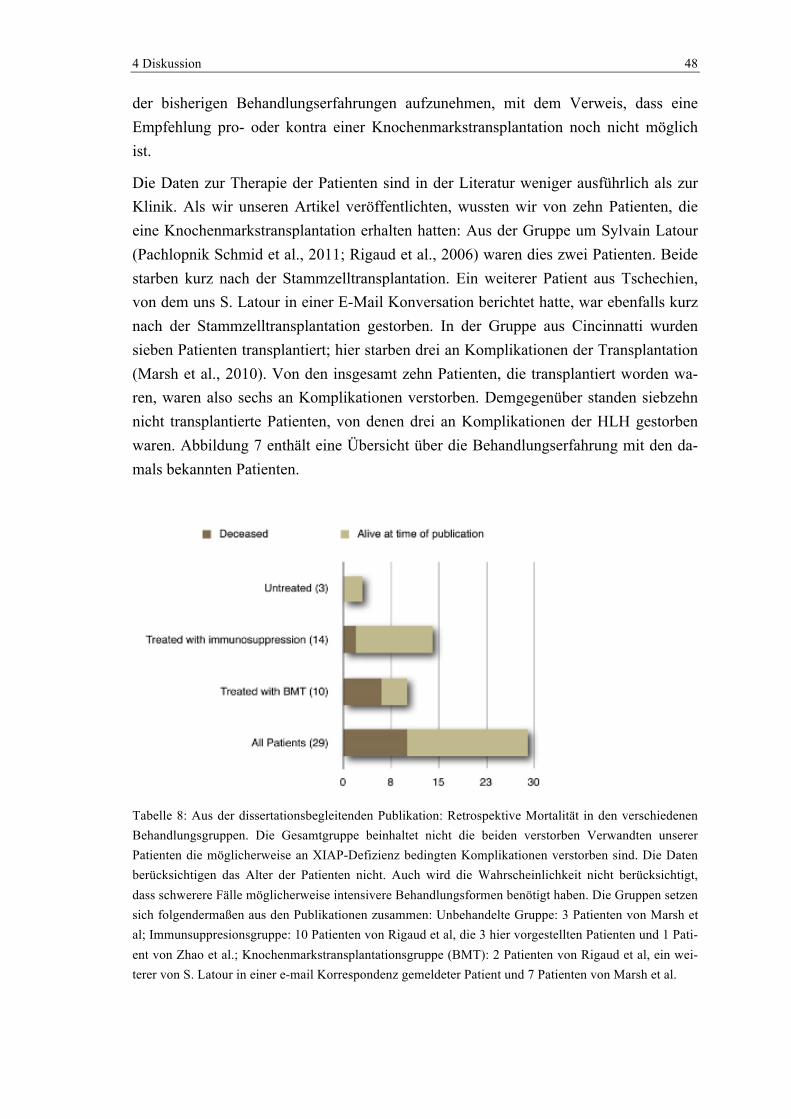
4 Diskussion 48
der bisherigen Behandlungserfahrungen aufzunehmen, mit dem Verweis, dass eine Empfehlung pro- oder kontra einer Knochenmarkstransplantation noch nicht möglich ist.
Die Daten zur Therapie der Patienten sind in der Literatur weniger ausführlich als zur Klinik. Als wir unseren Artikel veröffentlichten, wussten wir von zehn Patienten, die eine Knochenmarkstransplantation erhalten hatten: Aus der Gruppe um Sylvain Latour (Pachlopnik Schmid et al., 2011; Rigaud et al., 2006) waren dies zwei Patienten. Beide starben kurz nach der Stammzelltransplantation. Ein weiterer Patient aus Tschechien, von dem uns S. Latour in einer E-Mail Konversation berichtet hatte, war ebenfalls kurz nach der Stammzelltransplantation gestorben. In der Gruppe aus Cincinnatti wurden sieben Patienten transplantiert; hier starben drei an Komplikationen der Transplantation (Marsh et al., 2010). Von den insgesamt zehn Patienten, die transplantiert worden wa-ren, waren also sechs an Komplikationen verstorben. Demgegenüber standen siebzehn nicht transplantierte Patienten, von denen drei an Komplikationen der HLH gestorben waren. Abbildung 7 enthält eine Übersicht über die Behandlungserfahrung mit den da-mals bekannten Patienten.
Tabelle 8: Aus der dissertationsbegleitenden Publikation: Retrospektive Mortalität in den verschiedenen Behandlungsgruppen. Die Gesamtgruppe beinhaltet nicht die beiden verstorben Verwandten unserer Patienten die möglicherweise an XIAP-Defizienz bedingten Komplikationen verstorben sind. Die Daten berücksichtigen das Alter der Patienten nicht. Auch wird die Wahrscheinlichkeit nicht berücksichtigt, dass schwerere Fälle möglicherweise intensivere Behandlungsformen benötigt haben. Die Gruppen setzen sich folgendermaßen aus den Publikationen zusammen: Unbehandelte Gruppe: 3 Patienten von Marsh et al; Immunsuppresionsgruppe: 10 Patienten von Rigaud et al, die 3 hier vorgestellten Patienten und 1 Pati-ent von Zhao et al.; Knochenmarkstransplantationsgruppe (BMT): 2 Patienten von Rigaud et al, ein wei-terer von S. Latour in einer e-mail Korrespondenz gemeldeter Patient und 7 Patienten von Marsh et al.

4 Diskussion 49
Aus diesen Daten konnte nicht abgeleitet werden, dass eine Knochenmarkstransplanta-tion zwangsläufig zu schlechteren Ergebnissen führt, die Tendenz ist jedoch eindrück-lich. Im Reviewprozess wurde diskutiert, dass vermutlich ein Behandlungsbias vorliegt. Fast alle primären Immundefizienzen zeigen ein „Spectrum of disease“, was bedeutet dass es mildere und schwerere Verläufe gibt. Patienten mit schwereren Verläufen wer-den vermutlich eher transplantiert als diejenigen mit von Natur aus benignerem Verlauf.
Der vierte Gutachter stellte unseren Erfahrungen entgegen, es gebe keinen theoretischen Hintergrund dafür, dass XIAP-Defizienz eine schlechtere Prognose bei der Transplanta-tion habe als andere PID. Dafür spreche auch, dass Marsh et al an vier Patienten demon-stieren konnten, dass eine Knochenmarkstransplantation (HCT) prinzipiell durchführbar ist.
Ob der Reviewer Recht hat, dass es keinen theoretischen Hintergrund dafür gibt, dass XIAP-Defizienz eine schlechtere Prognose bei einer HCT hat als andere PID, ist frag-lich. Man kann zumindest einwerfen, dass XIAP ein Apoptoseinhibitor ist und bei sei-nem Fehlen die präoperative Myeoloablation überschießend ablaufen könnte. Muris et al haben an Biopsien von B-Zell-Lymphomen gezeigt, dass Zellen um so widerstands-fähiger gegenüber Chemotherapeutika sind, je höher ihre XIAP-Konzentration ist (Muris et al., 2005). Im Umkehrschluss könnte das bedeuten, dass bei Fehlen von XIAP eine erhöhte Empfänglichkeit gegenüber Apoptose-stimulierenden Substanzen wie Chemotherapeutika besteht.
Aufgrund der vorliegenden Daten entschieden wir uns bei allen von uns betreuten Pati-enten gegen eine Knochenmarktransplantation; an anderen Zentren wurde jedoch weiter transplantatiert. Inzwischen liegen weitere Daten vor, die unsere These stützen. In einer internationalen Umfrage wurden die behandelnden Zentren befragt. Insgesamt kamen so 19 Patienten zusammen, die eine Stammzelltransplantation erhalten hatten (Marsh et al., 2013). Hierbei kamen 12 Patienten aus den USA, sechs aus Europa und einer aus Japan.
Die Umfrage teilt die Patienten in zwei Gruppen ein: Patienten, bei denen ein Standard-myeloablatives Verfahren mit Busulfan (MAC) zur Anwendung kam, und Patienten, bei denen ein myeloablatives Verfahren mit reduzierter Intensität (RIC) zur Anwendung kam. Bei einem weiteren Patienten (Nummer 8 in der folgenden Tabelle), der von die-sen beiden Gruppen ausgeschlossen wurde, wird das Verfahren als „intermediate“ be-zeichnet. Hier herrschte Unklarheit, welches Verfahren zum Einsatz gekommen war (Marsh 2013).
Die Zahlen sind deutlich: In der MAC-Gruppe hatte von sieben Behandelten nur ein Kind überlebt, bei einer Nachbeobachtungszeit von 414 Tagen. Das Kind leidet unter einer auf die Haut limitierten Graft Versus Host Disease und muss permanent immun-supprimierend therapiert werden.

4 Diskussion 50
In der RIC Gruppe sind fünf von elf Patienten verstorben, bei einer mittleren Nachbeob-achtungszeit von 570 Tagen.
In der Gesamtpopulation sind also 12 von 19 Transplantierten verstorben – dies ent-spricht 63 %. Zum Vergleich: In unserer Veröffentlichung 2011 hatten wir die Mortali-tät bei Knochmarkstransplantation mit 60% angegeben, was diesem Wert nahekommt. Tabelle 8 gibt einen Überblick über die 19 knochenmarkstransplantierten Patienten.
Tabelle 9.: Überleben und Todesursache bei 19 knochenmarkstransplantierten XIAP-Defizienz Patienten (Marsh et al, 2013)
Bedauerlicherweise werden in der Veröffentlichung von Marsh et al keine Vergleiche zum Überleben nichttransplantierter XLP-2 Erkrankter aufgeführt. Die drei lebenden Patienten, die wir identifiziert haben, führen unter Immunglobulingaben ein relativ normales Leben. In der größten Zusammenfassung der französischen Gruppe (Pachlop-nik, Schmid et al, 2011) ist von 19 Patienten die diagnostiziert und therapiert wurden, jedoch keine Stammzelltransplantation erhalten haben, ein Patient verstorben. Dies ent-spricht einer Mortalität von 5% unter medikamentöser Therapie.
Die vorliegende Arbeit war als retrospektive Fallstudie nicht darauf ausgelegt, die bei-den Behandlungsmöglichkeiten zu vergleichen. Die Mortalitätsrate von 63% in der mit Knochenmarkstransplantation behandelten Gruppe gegenüber 5% in der medikamentös behandelten Gruppe zeigt jedoch einen klaren Trend, der gegen eine prophylaktische Stammzelltherapie bei XIAP-Defizienz spricht. In anderen Zentren werden möglicher-weise weiter nach RIC-Schema Myeloablationen durchgeführt. Laut Marsh et al (2013) hätte RIC eine Mortalität von nur 16 %, solange im HLH-freien Intervall therapiert wird. Dies liegt jedoch immer noch weit über der Mortalität bei IVIG-Gabe. Bedenkt man die nichtletalen Komplikationen wie Graft-versus-host-Disease, ist auch nicht si-cher gegeben, dass nach einer Knochenmarkstransplantation eine bessere klinische Ausgangslage für die Patienten geschaffen wird.

4 Diskussion 51
Die Therapieentscheidung bleibt einzelfallabhängig. Bei den von uns betreuten Patien-ten hat uns die vorliegende Erfahrung darin bestätigt, keine Transplantation zu empfeh-len. Wären die Knaben positiv für eine SAP-Mutation getestet worden, wäre die Ent-scheidung vermutlich anders ausgefallen.
Unklar bleibt weiterhin, wie man Genträger behandeln soll, die diagnostiziert werden, bevor es zur Erstmanifestation kommt. Diese Konstellation wird wahrscheinlicher, je mehr Familien bekannt werden.
Da HLH an sich eine hohe Mortalität aufweist, die Penetranz fast vollständig zu sein scheint und die Folgeschäden für das Immunsystem, wenn es zu Knochenmarksnekro-sen kommt, unberechenbar sind, ist in diesen Fällen möglicherweise eine RIC-abladierte Transplantation Therapie der Wahl.

5 Zusammenfassung der Arbeit 52
5 Zusammenfassung der Arbeit
Dissertation zur Erlangung des akademischen Grades Dr. med.
Charakterisierung des XIAP-Gens bei zwei Familien mit X-chromosomalem lymphoproliferativem Syndrom
Eingereicht von:
Peter Christian Horn geboren am 04.08.1982 in Freiburg
Angefertigt an:
Der Universitätsklinik und Poliklinik für Kinder und Jugendliche Leipzig und dem Zentrum für familiären Brust- und Eierstockkrebs der technischen Universität München
Betreuer:
Prof. Dr. med. Volker Schuster (Universität Leipzig) Prof. Dr. rer. nat. Alfons Meindl (TU München)
Eingereicht im Dezember 2014
Das X-linked-inhibitor-of-apoptosis Protein XIAP nimmt eine zentrale Rolle in der Hemmung von Apoptoseprozessen beim Menschen ein. In Abwesenheit von XIAP kann eine defekte Immunabwehr gegenüber viralen Infektionen beobachtet werden. In Folge können lebensbedrohliche Immunreaktionen wie hämophagozytäre Lymphhistiozytose, aplastische Anämie und persistierende Hypogammaglobulinämie auftreten.
Im Rahmen dieser Arbeit wurde untersucht, ob bei einem Patientenkollektiv mit lymphoproliferativem Syndrom Mutationen im XIAP-Gen Auslöser der Erkrankung sind. Es wurde eine molekulargenetische Sequenzierung und Auswertung des XIAP-

5 Zusammenfassung der Arbeit 53
Gens bei 36 Verdachtsfällen eines X-chromosomal vererbten lymphoproliferativen Syndroms durchgeführt. In allen Fällen wurde der gesamte exonische Abschnitt des Gens sequenziert und auf Polymorphismen und Mutationen untersucht. Bei zwei Proben wurden Mutationen im XIAP-Gen gefunden. Weiter konnten Mutter und Bruder eines der Betroffenen untersucht werden, so dass insgesamt drei Knaben mit XIAP-Gendefekt sowie eine heterozygote Konduktorin identifiziert wurden.
Nach der Identifikation der Mutationsträger erfolgte eine Auswertung der Krankenge-schichte und ein Vergleich mit den verfügbaren Beschreibungen von XIAP-Defizienz. Die beiden neu identifizierten Mutationen verursachen ein Krankheitsbild, das mit den wenigen verfügbaren Beschreibungen von XIAP-Defizienz vereinbar ist. Die Auswer-tung der Klinik der drei Betroffenen unterstützt die Hypothese, dass bei XIAP-Defizienz keine Lymphome auftreten.
Die Therapieentscheidung bei XIAP-Defizienz ist einzelfallabhängig. Die Arbeit ver-gleicht den Krankheitsverlauf der neu beschriebenen Patienten mit der Literatur und unterstreicht, dass die Entscheidung zu einer Knochenmarkstransplantation gut begrün-det werden muss. Es sind inzwischen mehr Patienten bekannt, die ohne Therapie oder unter IVIG-Gabe asymptomatisch sind, als solche, die eine Knochenmarkstransplantati-on überlebt haben. Alle hier beschriebenen Patienten konnten ohne Transplantation be-handelt werden. Eine Stammzelltransplantation kann jedoch erfolgreich sein. Bei kon-servativ behandelten Patienten können Rezidive auftreten.
Kritisch zu sehen sind im Licht der Erfahrung mit den XIAP-defizenten Patienten An-strengungen, XIAP-Inhibitoren zur Tumortherapie am Menschen zu entwickeln – dies könnte als Nebenwirkung paradoxe Folgen wie Lymphoproliferation oder HLH haben.
Die in der Arbeit beschriebenen Patienten waren die ersten Patienten mit XIAP-Defizienz, die im deutschsprachigen Raum identifiziert wurden. Die beiden Mutationen die bei den Familien gefunden wurden, waren bisher nicht beschrieben.

Anhang: Einwilligungen 54
Anhang: Einwilligungen Bei Probeneingang im genetischen Labor musste eine unterschriebene Einwilligung zur genetischen Untersuchung vorliegen. Ein Exemplar der Einwilligung zeigt Abbildung 9.
Die Einwilligung bot den Untersuchten verschiedene Optionen. Es wurden nur Proben zur Untersuchung herangezogen, die im Labor asserviert worden waren. Für die Asser-vierung musste folgendem Passus zugestimmt worden sein:
"Das GenDG schreibt die Vernichtung des Untersuchungsmaterials nach Abschluss der Untersuchung vor. Mit Ihrer Einwilligung darf es jedoch länger aufbewahrt werden. Dies kann dann wichtig sein, wenn aufgrund neuer wissenschaftlicher Erkenntnisse eine Überprüfung oder Erweiterung der Untersuchung möglich wird, oder wenn für spätere genetische Untersuchungen in der Familie Vergleichsmaterial benötigt wird.
Ich bin mit der Aufbewahrung und Verwendung meines Untersuchungsmaterials über die gesetzliche Frist hinaus zum Zwecke der Nachprüfbarkeit oder Erweiterung der Er-gebnisse oder eventueller späterer Untersuchungen in meiner Familie einverstanden."
Bei den positiv getesteten Patienten wurde durch diese Untersuchungen nachgewiesen, dass die neu entdeckten Mutationen im XIAP-Gen vorlagen.
Die behandelnden Ärzte der betroffenen Familien wurden vom Labor kontaktiert und führten die Information und Aufklärung der Patienten durch. Dies war durch folgenden Passus der Aufklärung gedeckt:
"Bei genetischen Untersuchungen können sich Zufallsbefunde ergeben, die nicht im Zusammenhang mit der o.g. Fragestellung stehen aber trotzdem gesundheitliche Bedeu-tung haben. Ich möchte über Zufallsbefunde informiert werden."
Die behandelnden Ärzte der Patienten - Professor Urban, Graz und Professor Behlo-radsky, München - wurden ab dem Nachweis der Mutation bei ihren Patienten über die Forschungsergebnisse informiert und maßgeblich mit in die Auswertung und Abschät-zung der klinischen Auswirkungen eingebunden. Entsprechend wurden beide Mitauto-ren der folgenden wissenschaftlichen Veröffentlichung.

Anhang: Einwilligungen 55
Abbildung 9: Vorlage für die Einwilligung für genetische Untersuchungen der Ludwig-Maximilians-Universität München
Einwilligung zur genetischen Untersuchung
Patientendaten (ggf. Aufkleber)
____________________________________ _________________________________________ Name Vorname
_______________________ ! männlich ! weiblich geb.
_________________________________________________________________________________ Strasse
________________ _______________________________________________________ PLZ Ort
Bitte zurücksenden an:
Klinikum der LMU Institut für Humangenetik Goethestrasse 29 D-80336 München
Telefon +49 (0)89 5160 - 3683 Telefax +49 (0)89 5160 - 4468
E-mail: [email protected] Internet: www.klinikum.uni-muenchen.de
02/2010
Das Gendiagnostikgesetz (GenDG) schreibt vor, dass vor jeder genetischen Untersuchung eine ausführliche Auf-klärung des Patienten erfolgt und eine genetische Beratung angeboten wird. Die Untersuchung darf erst nach Vor-liegen der schriftlichen Einwilligung des Patienten begonnen werden. Bei vorgeburtlichen und prädiktiven (vorher-
sagenden) genetischen Untersuchungen muss zusätzlich vorher eine genetische Beratung angeboten werden. Bitte lesen Sie diese Einwilligungserklärung sorgfältig durch und kreuzen Sie die zutreffenden Antworten an:
Gewünschte genetische Untersuchung:
…………………………………………………………………………… Über die genetischen Grundlagen der Erkrankung sowie die Aussagekraft, Grenzen und möglichen Konsequenzen der geplanten genetischen Untersuchung einschließlich der mit der Blut-/Gewebe-entnahme verbundenen Risiken bin ich hinreichend aufgeklärt worden. Alle meine Fragen wurden mir beantwortet.
Ja Nein
Das GenDG erlaubt die Versendung des Befundes nur an den Arzt, welcher Sie über die genetische Untersuchung aufgeklärt hat. Wenn Sie möchten, dass auch weitere, mitbehandelnde Ärzte den Befund bekommen, brauchen wir hierfür Ihre Zustimmung. Ich bin damit einverstanden, dass die Befunde der genetischen Untersuchung auch an folgende mitbehandelnde Ärzte geschickt werden:
Frau/Herrn Dr.: ……………………………………………………………………………………………………………
………………………………………………………………………………………………………………………………
Ja Nein
Bei genetischen Untersuchungen können sich Zufallsbefunde ergeben, die nicht im Zusammenhang mit der o.g. Fragestellung stehen aber trotzdem gesundheitliche Bedeutung haben. Ich möchte über Zufallsbefunde informiert werden.
Ja Nein
Das GenDG schreibt die Vernichtung des Untersuchungsmaterials nach Abschluss der Untersuchung vor. Mit Ihrer Einwilligung darf es jedoch länger aufbewahrt werden. Dies kann dann wichtig sein, wenn aufgrund neuer wissenschaftlicher Erkenntnisse eine Überprüfung oder Erweiterung der Untersuchung möglich wird, oder wenn für spätere genetische Untersuchungen in der Familie Vergleichsmaterial benötigt wird. Ich bin mit der Aufbewahrung und Verwendung meines Untersuchungsmaterials über die gesetzliche Frist hinaus zum Zwecke der Nachprüfbarkeit oder Erweiterung der Ergebnisse oder eventueller späterer Untersuchungen in meiner Familie einverstanden.
Ja Nein
Genetische Proben können als Vergleichsmaterial für die Qualitätssicherung im Labor wichtig sein. Ich bin mit der Aufbewahrung und anonymisierten (Unkenntlichmachung persönlicher Daten) Verwendung meines Untersuchungsmaterials zum Zwecke der Qualitätssicherung einverstanden.
Ja Nein
Das GenDG schreibt vor, dass genetische Untersuchungsergebnisse nach 10 Jahren vernichtet werden. Häufig sind diese Ergebnisse aber auch später noch wichtig für Ihre Kinder oder Enkelkinder. Ich bin einverstanden, dass meine Untersuchungsergebnisse über die gesetzliche Frist hinaus zum Zwecke eventueller späterer Beratungen/Untersuchungen in meiner Familie aufbewahrt werden.
Ja Nein
Bei Bedarf dürfen die Ergebnisse dieser genetischen Untersuchung für die Beratung/Untersuchung meiner Angehörigen genutzt werden.
Ja Nein
Ich wurde darauf hingewiesen, dass ich meine Einwilligung jederzeit ohne Angaben von Gründen ganz oder teilweise zurückziehen kann, ohne dass mir daraus Nachteile entstehen und dass ich das Recht habe, Untersuchungsergebnisse nicht zu erfahren (Recht auf Nichtwissen). Mir ist bekannt, dass ich die eingeleitete Untersuchung jederzeit stoppen und die Vernichtung des Untersuchungsmaterials sowie aller bis dahin erhobenen Ergebnisse und Befunde verlangen kann.
Ja Nein
Mit meiner Unterschrift gebe ich meine Einwilligung zur genetischen Untersuchung für die o.g. Fragestellung und zu der dafür erforderlichen Blut-/Gewebeentnahme. ……….………………………………………………………………………………………………………………………………………………………………………………. Ort, Datum Unterschrift des zu Untersuchenden Stempel und Unterschrift der gemäß GenDG bzw. des gesetzlichen Vertreters verantwortlichen ärztlichen Person (aufklärender und die Untersuchung veranlassender Arzt)

Literaturverzeichnis 56
Literaturverzeichnis Al-Herz, W., Bousfiha, A., Casanova, J.-L., Chapel, H., Conley, M. E., Cunningham-
Rundles, C., Etzioni, A., et al. (2011). Primary immunodeficiency diseases: an up-date on the classification from the international union of immunological societies expert committee for primary immunodeficiency. Frontiers in Immunology, 2, 54.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., & Lipman, D. J. (1990). Basic local alignment search tool. Journal of molecular biology, 215(3), 403–410.
Benoit, S., Rose, C., Schmidt, E., & Zillikens, D. (2003). Persistierende Papulopusteln im Gesicht eines 21-Jährigen. Journal der Deutschen Dermatologischen Ge-sellschaft, 1(11), 907–909.
Booth, C., Gilmour, K. C., Veys, P., Gennery, A. R., Slatter, M. A., Chapel, H., Heath, P. T., et al. (2011). X-linked lymphoproliferative disease due to SAP/SH2D1A de-ficiency: a multicenter study on the manifestations, management and outcome of the disease. Blood, 117(1), 53–62.
Bratton, S. B., Lewis, J., Butterworth, M., Duckett, C. S., & Cohen, G. M. (2002). XIAP inhibition of caspase-3 preserves its association with the Apaf-1 apoptosome and prevents CD95- and Bax-induced apoptosis. Cell Death and Differentiation, 9(9), 881–892.
Bruton, O.C. (1952). Agammaglobulinemia. Pediatrics, 9(6), 722–728. Cepok, S. (2004). Charakterisierung der humoralen Immunantwort bei der Multiplen
Sklerose. (M. Bölker & B. Hemmer, Eds.) Dissertationsschrift im Fachbereich Bi-ologie der Philipps-Universität Marburg, 83–90.
Chaganti, S., Ma, C. S., Bell, A. I., Croom-Carter, D., Hislop, A. D., Tangye, S. G., & Rickinson, A. B. (2008). Epstein-Barr virus persistence in the absence of conven-tional memory B cells: IgM+IgD+CD27+ B cells harbor the virus in X-linked lym-phoproliferative disease patients. Blood, 112(3), 672–679.
Chandrakasan, S., & Filipovich, A. H. (2013). Hemophagocytic lymphohistiocytosis: advances in pathophysiology, diagnosis, and treatment. The Journal of Pediatrics, 163(5), 1253–1259.
Cillessen, S. A. G. M., Reed, J. C., Welsh, K., Pinilla, C., Houghten, R., Hooijberg, E., Deurhof, J., et al. (2008). Small-molecule XIAP antagonist restores caspase-9 me-diated apoptosis in XIAP-positive diffuse large B-cell lymphoma cells. Blood, 111(1), 369–375.
Coffey, A. J., Brooksbank, R. A., Brandau, O., Oohashi, T., Howell, G. R., Bye, J. M., Cahn, A. P., et al. (1998). Host response to EBV infection in X-linked lymphopro-liferative disease results from mutations in an SH2-domain encoding gene. Nature Genetics, 20(2)
Conley, M. E., & Howard, V. (2002). Clinical findings leading to the diagnosis of X-linked agammaglobulinemia. The Journal of Pediatrics, 141(4), 566–571.
Cooper, N., Rao, K., Goulden, N., Webb, D., Amrolia, P., & Veys, P. (2008). The use of reduced-intensity stem cell transplantation in haemophagocytic lymphohistiocy-tosis and Langerhans cell histiocytosis. Bone Marrow Transplantation, 42, S47–S50.
Detre, C., Keszei, M., Romero, X., Tsokos, G. C., & Terhorst, C. (2010). SLAM family receptors and the SLAM-associated protein (SAP) modulate T cell functions. Semi-nars in Immunopathology, 32(2), 157–171.

Literaturverzeichnis 57
Duckett, C. S., Nava, V. E., Gedrich, R. W., Clem, R. J., Van Dongen, J. L., Gilfillan, M. C., Shiels, H., et al. (1996). A conserved family of cellular genes related to the baculovirus iap gene and encoding apoptosis inhibitors. The EMBO Journal, 15(11), 2685–2694.
Ehl, S., & Schuster, V. (2004). X-chromosomale lymphoproliferative Erkrankung (XLP). Allergologie, 27(2), 72–74.
Elmore, S. (2007). Apoptosis: A Review of Programmed Cell Death. Toxicologic Pa-thology, 35(4), 495–516.
Engel, P., Eck, M. J., & Terhorst, C. (2003). The SAP and SLAM families in immune responses and X-linked lymphoproliferative disease. Nature Reviews Immunology, 3(10),
Filipovich, A. H. (2001). Unrelated cord blood transplantation for correction of genetic immunodeficiencies. The Journal of Pediatrics, 138(4), 459–461.
Filipovich, A. H., Zhang, K., Snow, A. L., & Marsh, R. A. (2010). X-linked lymphopro-liferative syndromes: brothers or distant cousins? Blood, 116(18), 3398–3408.
Filipovich, A., Johnson, J., Zhang, K., & Marsh, R. (n.d.). Lymphoproliferative Disease, X-Linked - GeneReviews™ - NCBI Bookshelf. Zugriff Juni 2013.
Fisman, D. (2000). Hemophagocytic Syndromes and Infection. Emerging Infectious Diseases, 6(6), 601–608.
Galbán, S., & Duckett, C. S. (2009). XIAP as a ubiquitin ligase in cellular signaling. Cell Death and Differentiation, 17(1), 54–60.
Gross, T. G., Filipovich, A., Conley, M. E., Pracher, E., Schmiegelow, K., Verdirame, J., Vowels, M., et al. (1996). Cure of X-linked lymphoproliferative disease (XLP) with allogeneic hematopoietic stem cell transplantation (HSCT): report from the XLP registry. Bone Marrow Transplantation, 741–744.
Geha, R. S., Hyslop, N., Alami, S., Farah, F., Schneeberger, E. E., & Rosen, F. S. (1979). Hyper Immunoglobulin M Immunodeficiency (Dysgammaglobulinemia). The Journal of Clinical Investigation, 64(2), 385–391.
Hamilton, J. K., Paquin, L. A., Sullivan, J. L., Maurer, H. S., Cruzi, F. G., Provisor, A. J., Steuber, C. P., et al. (1980). X-linked lymphoproliferative syndrome registry re-port. The Journal of Pediatrics, 96(4), 669–673.
Horn, P. C., & Schuster, V. (2012). Gastritis and colitis can be associated with XLP-1 (SAP deficiency). Pediatrics International, 54(6), 964–author reply 964–5. doi:10.1111/ped.12000
Horn, P. C., Belohradsky, B. H., Urban, C., Weber-Mzell, D., Meindl, A., & Schuster, V. (2011). Two new families with X-linked inhibitor of apoptosis deficiency and a review of all 26 published cases. The Journal of Allergy and Clinical Immunology, 127(2), 544–546.
Huck, K., Feyen, O., Niehues, T., Rüschendorf, F., Hübner, N., Laws, H.-J., Telieps, T., et al. (2009). Girls homozygous for an IL-2-inducible T cell kinase mutation that leads to protein deficiency develop fatal EBV-associated lymphoproliferation. The Journal of Clinical Investigation, 119(5), 1350–1358.
Janka, G. E. (2012). Familial and acquired hemophagocytic lymphohistiocytosis. An-nual Review of Medicine, (63), 223–246.
Kainulainen, L., Nikoskelainen, J., Vuorinen, T., Tevola, K., Liippo, K., & Ruuskanen, O. (1999). Viruses and Bacteria in Bronchial Samples from Patients with Primary Hypogammaglobulinemia. American Journal of Respiratory and Critical Care Medicine, 159(4), 1199-1204.
Kashkar, H., Deggerich, A., Seeger, J.-M., Yazdanpanah, B., Wiegmann, K., Haubert, D., Pongratz, C., et al. (2007). NF-kappaB-independent down-regulation of XIAP by bortezomib sensitizes HL B cells against cytotoxic drugs. Blood, 109(9), 3982–

Literaturverzeichnis 58
3988. Kashkar, H., Seeger, J.-M., Hombach, A., Deggerich, A., Yazdanpanah, B., Uter-
möhlen, O., Heimlich, G., et al. (2006). XIAP targeting sensitizes Hodgkin lym-phoma cells for cytolytic T-cell attack. Blood, 108(10), 3434–3440.
Kildebeck, E., Checketts, J., & Porteus, M. (2012). Gene therapy for primary immu-nodeficiencies. Current Opinion In Pediatrics. 24(6), 731-738
Lankester, A. C., Visser, L. F. A., Hartwig, N. G., Bredius, R. G. M., Gaspar, H. B., van der Burg, M., van Tol, M. J. D., et al. (2005). Allogeneic stem cell transplantation in X-linked lymphoproliferative disease: two cases in one family and review of the literature. Bone Marrow Transplantation, 36(2), 99–105.
Ma, C. S., Nichols, K. E., & Tangye, S. G. (2007). Regulation of Cellular and Humoral Immune Responses by the SLAM and SAP Families of Molecules. Annual Review of Immunology, 25(1), 337–379.
Maródi, L., & Notarangelo, L. D. (2007). Immunological and genetic bases of new pri-mary immunodeficiencies. Nature Reviews Immunology, 7(11), 851–861.
Marsh, R. A., Madden, L., Kitchen, B. J., Mody, R., McClimon, B., Jordan, M. B., Bleesing, J. J., et al. (2010). XIAP deficiency: a unique primary immunodeficiency best classified as X-linked familial hemophagocytic lymphohistiocytosis and not as X-linked lymphoproliferative disease. Blood, 116(7), 1079–1082.
Marsh, R. A., Rao, K., Satwani, P., Lehmberg, K., Muller, I., Li, D., Kim, M. O., et al. (2013). Allogeneic hematopoietic cell transplantation for XIAP deficiency: an in-ternational survey reveals poor outcomes. Blood, 121(6), 877–883.
Marsh, R. A., Villanueva, J., Zhang, K., Snow, A. L., Su, H. C., Madden, L., Mody, R., et al. (2009). A rapid flow cytometric screening test for X-linked lymphoprolifera-tive disease due to XIAP deficiency. Cytometry. Part B, Clinical Cytometry, 76(5), 334–344.
Muris, J. J. F., Cillessen, S. A. G. M., Vos, W., van Houdt, I. S., Kummer, J. A., van Krieken, J. H. J. M., Jiwa, N. M., et al. (2005). Immunohistochemical profiling of caspase signaling pathways predicts clinical response to chemotherapy in primary nodal diffuse large B-cell lymphomas. Blood, 105(7), 2916–2923.
Nelson, D. L., & Terhorst, C. (2000). X-linked lymphoproliferative syndrome. Clinical and Experimental Immunology, 122(3), 291–295.
Nichols, K. E., Harkin, D. P., Levitz, S., Krainer, M., Kolquist, K. A., Genovese, C., Bernard, A., et al. (1998). Inactivating mutations in an SH2 domain-encoding gene in X-linked lymphoproliferative syndrome. Proceedings of the National Academy of Sciences of the United States of America, 95(23), 13765–13770.
Nichols, K. E., Ma, C. S., Cannons, J. L., Schwartzberg, P. L., & Tangye, S. G. (2005). Molecular and cellular pathogenesis of X-linked lymphoproliferative disease. Im-munological Reviews, 203(1), 180–199.
Notarangelo, L. D., Fischer, A., Geha, R. S., Casanova, J.-L., Chapel, H., Conley, M. E., Cunningham-Rundles, C., et al. (2009). Primary immunodeficiencies: 2009 up-date. The Journal of Allergy and Clinical Immunology, 124(6), 1161–1178.
Ochs, H. D., & Hitzig, W. H. (2012). History of primary immunodeficiency diseases. Current Opinion in Allergy and Clinical Immunology, 12(6), 577–587.
Ochs, H. D., Smith, C. I. E., & Puck, J. (2007). Primary Immunodeficiency Diseases. A Molecular and Genetic Approach (p. 726). Oxford University Press, USA.
Okano, M., & Gross, T. G. (2012). Acute or chronic life-threatening diseases associated with Epstein-Barr virus infection. The American Journal of the Medical Sciences, 343(6), 483–489.
OMIM Entry - #300635 - LYMPHOPROLIFERATIVE SYNDROME, X-LINKED, 2; XLP2. (n.d.). OMIM Entry - #300635 - LYMPHOPROLIFERATIVE SYN-

Literaturverzeichnis 59
DROME, X-LINKED, 2; XLP2. omim.org. Abgerufen am 2. Juni 2012, unter http://www.omim.org/entry/300635
OMIM Entry - #308240 - LYMPHOPROLIFERATIVE SYNDROME, X-LINKED, 1; XLP1. (n.d.). OMIM Entry - #308240 - LYMPHOPROLIFERATIVE SYN-DROME, X-LINKED, 1; XLP1. omim.org. Abgerufen am 2. Juni 2012 unter http://omim.org/entry/308240?search=308240
OMIM Entry - *300079 - INHIBITOR OF APOPTOSIS, X-LINKED; XIAP. (n.d.). OMIM Entry - *300079 - INHIBITOR OF APOPTOSIS, X-LINKED; XIAP. omim.org. Abgerufen am 2. Juni 2012 unter http://www.omim.org/entry/300079
OMIM Entry - *300490 - SH2 DOMAIN PROTEIN 1A; SH2D1A. (n.d.). OMIM Entry - *300490 - SH2 DOMAIN PROTEIN 1A; SH2D1A. omim.org. Abgerufen am 2. Juni 2012 unter http://omim.org/entry/300490
Pachlopnik Schmid, J., Canioni, D., Moshous, D., Touzot, F., Mahlaoui, N., Hauck, F., Kanegane, H., et al. (2011). Clinical similarities and differences of patients with X-linked lymphoproliferative syndrome type 1 (XLP-1/SAP deficiency) versus type 2 (XLP-2/XIAP deficiency). Blood, 117(5), 1522–1529.
Primer Design Guide for PCR :: Learn Designing Primers for PCR. premierbiosoft.com. Abgerufen am 4. Juni 2012, unter http://www.premierbiosoft.com/tech_notes/PCR_Primer_Design.html
Punnonen, J., Cocks, B. G., Carballido, J. M., Bennett, B., Peterson, D., Aversa, G., & De Vries, J. E. (1997). Soluble and membrane-bound forms of signaling lympho-cytic activation molecule (SLAM) induce proliferation and Ig synthesis by activated human B lymphocytes. The Journal of Experimental Medicine, 185(6), 993–1004.
Purtilo, D. T., Cassel, C. K., Yang, J. P., & Harper, R. (1975). X-linked recessive pro-gressive combined variable immunodeficiency (Duncan's disease). Lancet, 1(7913), 935–940.
Purtilo, D. T., Cassel, C., & Yang, J. P. (1974). Letter: Fatal infectious mononucleosis in familial lymphohistiocytosis. The New England Journal of Medicine, 291(14), 736.
Purtilo, DT, J. (1977). Immunological disorders and malignancies in five young broth-ers. Archives of Disease in Childhood, 52(4), 310.
Purtilo, D. T., Sakamoto, K., Barnabei, V., Seeley, J., Bechtold, T., Rogers, G., Yetz, J., et al. (1982). Epstein-Barr virus-induced diseases in boys with the X-linked lym-phoproliferative syndrome (XLP): update on studies of the registry. The American Journal of Medicine, 73(1), 49–56.
Reddy, N., Rezvani, K., Barrett, A. J., & Savani, B. N. (2011). Strategies to Prevent EBV Reactivation and Posttransplant Lymphoproliferative Disorders (PTLD) after Allogeneic Stem Cell Transplantation in High-Risk Patients. Biology of Blood and Marrow Transplantation, 17(5), 591–597.
Riedl, S. J., & Shi, Y. (2004). Molecular mechanisms of caspase regulation during apoptosis. Nature Reviews. Molecular Cell Biology, 5(11), 897–907. doi:10.1038/nrm1496
Rigaud, S., Fondanèche, M.-C., Lambert, N., Pasquier, B., Mateo, V., Soulas, P., Galicier, L., et al. (2006). XIAP deficiency in humans causes an X-linked lymphop-roliferative syndrome. Nature, 444(7115), 110–114.
Rougemont, A.-L., Fournet, J.-C., Martin, S. R., de Saint Basile, G., Latour, S., Pri-meau, M.-N., Rubbia-Brandt, L., et al. (2008). Chronic active gastritis in X-linked lymphoproliferative disease. The American Journal of Surgical Pathology, 32(2), 323–328.
Sanzone, S., Zeyda, M., Saemann, M. D., Soncini, M., Holter, W., Fritsch, G., Knapp, W., et al. (2003). SLAM-associated Protein Deficiency Causes Imbalanced Early

Literaturverzeichnis 60
Signal Transduction and Blocks Downstream Activation in T Cells from X-linked Lymphoproliferative Disease Patients. Journal of Biological Chemistry, 278(32), 29593–29599.
Sayos, J., Wu, C., Morra, M., Wang, N., Zhang, X., Allen, D., van Schaik, S., et al. (1998). The X-linked lymphoproliferative-disease gene product SAP regulates sig-nals induced through the co-receptor SLAM. Nature, 395(6701), 462–469.
Schuster, V., Dohrmann, E., & Kreth, H. W. (1991). Epstein-Barr virus-associated lym-phoproliferative syndromes: studies in two European families. Cancer Detection and Prevention, 15(1), 65–67.
Schuster, V., Grimm, T., Kress, W., Seidenspinner, S., Belohradsky, B. H., Müller, P., & Kreth, H. W. (1995). Die X-chromosomal-rezessive Lymphoproliferative Er-krankung (XLP): Molekulargenetische Untersuchungen. Klinische Pädiatrie, 207(5), 271–276.
Schuster, V., Seidenspinner, S., Grimm, T., Kress, W., Zielen, S., Bock, M., & Kreth, H. W. (1994). Molecular genetic haplotype segregation studies in three families with X-linked lymphoproliferative disease. European Journal of Pediatrics, 153(6), 432–437.
Schuster, V., & Terhorst, C. (2007). X-linked Lymphoproliferative Disease (XLP) due to Defects of SH2D1A; (H. D. Ochs, C. I. E. Smith, & J. Puck, Eds.) Primary Im-munodeficency Syndromes. A Molecular and Genetic Approach (p. 523-527). Ox-ford University Press, USA.
Seemayer, T. A., Gross, T. G., Egeler, R. M., Pirruccello, S. J., Davis, J. R., Kelly, C. M., Okano, M., et al. (1995). X-linked lymphoproliferative disease: twenty-five years after the discovery. Pediatric Research, 38(4), 471–478.
Speckmann C., Rohr J., Ehl, S.: Genetic disorders of Immune Regulations (2008); in Rezaei, N., Notarangelo, L., & Aghamohammadi, A. (Hrsg). Primary Immunodefi-ciency Diseases. Definition, Diagnosis, and Management (168-171 und 175-178). Springer Verlag, 2te Auflage (Berlin/Heidelberg).
Spector, B. D., Perry, G. S., & Kersey, J. H. (1978). Genetically determined immunode-ficiency diseases (GDID) and malignancy: report from the immunodeficiency--cancer registry. Clinical Immunology and Immunopathology, 11(1), 12–29.
Sullivan, J. L., Medveczky, P., Forman, S. J., Baker, S. M., Monroe, J. E., & Mulder, C. (1981). Epstein–Barr-Virus Induced Lymphoproliferation. The New England Jour-nal of Medicine, 311(18), 1163–1167.
Sumegi, J., Huang, D., Lanyi, A., Davis, J. D., Seemayer, T. A., Maeda, A., Klein, G., et al. (2000). Correlation of mutations of the SH2D1A gene and epstein-barr virus infection with clinical phenotype and outcome in X-linked lymphoproliferative dis-ease. Blood, 96(9), 3118–3125.
Veillette, A., Zhang, S., Shi, X., Dong, Z., Davidson, D., & Zhong, M. C. (2008). SAP expression in T cells, not in B cells, is required for humoral immunity. Proceedings of the National Academy of Sciences, 105(4), 1273–1278.
Völkel, I. (2005). Untersuchungen zur molekulargenetischen Rassendifferenzierung bei Canis familiaris. Dissertationschrift an der Humboldt-Universität Berlin, 1–154.
Veillette, A., Zhang, S., Shi, X., Dong, Z., Davidson, D., & Zhong, M. C. (2008). SAP expression in T cells, not in B cells, is required for humoral immunity. Proceedings of the National Academy of Sciences, 105(4), 1273–1278.
Worth, A., Thrasher, A. J., & Gaspar, H. B. (2006). Autoimmune lymphoproliferative syndrome: molecular basis of disease and clinical phenotype. British Journal of Haematology, 133(2), 124–140.
Yang, X., Miyawaki, T., & Kanegane, H. (2012). SAP and XIAP deficiency in hemo-phagocytic lymphohistiocytosis. Pediatrics International, 54(4), 447-454.

Literaturverzeichnis 61
Zhang, K., Filipovich, A. H., Johnson, J., Marsh, R. A., & Villanueva, J. (2010). Hemo-phagocytic Lymphohistiocytosis, Familial. Gene Reviews (Internet). Abgerufen am 4. April 2013 unter ncbi.nlm.nih.gov/books/NBK1444/
Zhao, M., Kanegane, H., Ouchi, K., Imamura, T., Latour, S., & Miyawaki, T. (2010). A novel XIAP mutation in a Japanese boy with recurrent pancytopenia and sple-nomegaly. Haematologica, 95(4), 688–689. doi:10.3324/haematol.2009.018010
Zhizhuo, H., Junmei, X., Yuelin, S., Qiang, Q., Chunyan, L., Zhengde, X., & Kunling, S. (2012). Screening the PRF1, UNC13D, STX11, SH2D1A, XIAP, and ITK gene mutations in Chinese children with Epstein-Barr virus-associated hemophagocytic lymphohistiocytosis. Pediatric blood & cancer, 58(3), 410–414. doi:10.1002/pbc.23216
Bemerkung zur Formatierung der Arbeit: Als Word-Dokumentvorlage wurde „Ab-schlussarbeiten u.a. wissenschaftliche Arbeiten“ der Hochschule für Bibliotheks- und Informationswesen Stuttgart benutzt. http://v.hdm-stuttgart.de/~riekert/theses/. (Datum des Zugriffs: 04.04.2014).

Erklärung über die eigenständige Abfassung der Arbeit 62
Erklärung über die eigenständige Abfassung der Arbeit Hiermit erkläre ich, dass ich die vorliegende Arbeit selbständig und ohne unzulässige Hilfe oder Benutzung anderer als der angegebenen Hilfsmittel angefertigt habe. Ich versichere, dass Dritte von mir weder unmittelbar noch mittelbar geldwerte Leistungen für Arbeiten erhalten haben, die im Zusam-menhang mit dem Inhalt der vorgelegten Dissertation stehen, und dass die vorgelegte Arbeit weder im Inland noch im Ausland in gleicher oder ähnlicher Form einer anderen Prüfungsbehörde zum Zweck einer Promotion oder eines anderen Prüfungsverfahrens vorgelegt wurde. Alles aus anderen Quellen und von anderen Personen übernommene Material, das in der Arbeit verwendet wurde oder auf das direkt Bezug genommen wird, wurde als solches kenntlich gemacht. Insbesondere wurden alle Personen genannt, die direkt an der Ent-stehung der vorliegenden Arbeit beteiligt waren. ................................. ....................................................
Datum Unterschrift

Lebenslauf 63
Lebenslauf
! PROFIL! Geboren am 04.08.1982 in Freiburg. Assistenzarzt für Augenheilkunde. Staats-examen mit 2,0 abgelegt. Platzierung im schriftlichen Examen nach Punktzahl unter den besten zehn Prozent der deutschen Medizinstudenten.
" AUSBILDUNG! MAX PLANCK-GYMNASIUMLahr, Baden-Württemberg! 1993-2002Abitur mit Notendurchschnitt 1,2. Preis für herausragende Leistungen im Fach Englisch. Im Schuljahr 98/99 Schülersprecher.
" " ZIVILDIENST AM INSTITUT FÜR HUMANGENETIKLudwig-Maximillians Universität München! 2002-2003Erlernen von PCR, Sequenzierung, Isolation von RNA und Herstellen von cDNA.
! ! MEDIZINSTUDIUM,LMU München, TU München! 2003-2009Vorlesungsassistent für Professor Gänsbacher (Hämatologie/Onkologie). Erfol-greiches Ablegen der ersten beiden Teile des amerikanischen Staatsexamens, USMLE Step 1 und Step 2 (96 von 99 Punkten).
! ! FACHARZTAUSBILDUNGCharité Berlin, Universitätsaugenklinik Freiburg ! 2010-2015Erwarteter Abschluss September 2015
!AUSLANDSERFAHRUNG! STUDIUM! ! !2004 Auslandssemester an der Universidad San Miguel Hernandez de Elche, Spanien, 2005 Famulatur in der Neurolgie in Wien,
! ! PRAKTISCHES JAHR ! ! !Wahlfach Augenheilkunde an der Universität Toronto, Kanada! !Innere Medizin an der Weill Cornell Medical School, New York, USA!!Chirurgie an der University of Cape Town, Südafrika
VERÖFFENTLICHUNGEN! Horn, Lohmann, Lanzl: Optikusneuropathie bei Schmidt-Carpenter Syndrom. Der Ophthalmologe, 2010! ! ! ! ! ! !Horn, Lohmann, Harder, Lanzl: Beidseitiger Visusabfall bei einer 11-Jährigen. Der Ophthalmologe, 2010! ! ! ! ! ! !Horn, Behloradsky, Urban, Weber-Mzell, Schuster: Two new families with X-linked inhibitor of apoptosis deficiency and a review of all 26 published cases Journal of Allergy and Immunology, 2011! ! ! ! !Horn und Schuster: Gastritis and colitis can be associated with XLP-1 (SAP de-ficiency). Pediatrics International, 2012
" SONSTIGES" Teilnahme am Mikrochirurgie-Kurs der Plastischen Chirurgie bei Professor Biemer. Hervorragende Englischkenntnisse (APIEL-Score: 5/5), gutes Spanisch. DSV-zertifizierter Snowboardlehrer. Gitarre, akustisch und elektrisch, zu Schul-zeiten in diversen Bands.
T 0761 48992061" PETER C. HORN" M 0179 157 672 2""
ADELHAUSERSTR. 9
79098 FREIBURG

Verzeichnis eigener Veröffentlichungen 64
Verzeichnis eigener Veröffentlichungen
Dissertationsbegleitende Veröffentlichungen:
Horn, P. C., Belohradsky, B. H., Urban, C., Weber-Mzell, D., Meindl, A., & Schuster, V. (2011). Two new families with X-linked inhibitor of apoptosis deficiency and a re-view of all 26 published cases. The Journal of allergy and clinical immunology, 127(2), 544–546.
Horn, P. C., & Schuster, V. (2012). Gastritis and colitis can be associated with XLP-1 (SAP deficiency). Pediatrics international : official journal of the Japan Pediatric So-ciety, 54(6), 964–author reply 964–5.
Impact Factor 2011 für JACI: 11.003 Impact Factor 2012 für Pediatrics international: 0.731 Kombinierter Impact Factor der dissertationsbegleitenden Veröffentlichungen: 11.734
Weitere, eigene Veröffentlichungen: Horn, Lohmann, Lanzl: Optikusneuropathie bei Schmidt-Carpenter Syndrom. Der Ophthalmologe, 2010 Horn, Lohmann, Harder, Lanzl: Beidseitiger Visusabfall bei einer 11-Jährigen. Der Ophthalmologe, 2010 Horn, Reinhard, Böhringer: Versuchsaufbau für die blitzlichtfreie Aufnahme von Stan-dard Dynamic Range und High Dynamic Range (HDR) Epithelfotografien, Ophthalmo-logische Nachrichten, 2012 Vorträge Augenbeteiligung bei Neurofibromatose; Von Recklinghausen-Gesellschaft, Berlin, 2010 Ectrodactyly-ectodermal dysplasia-clefting syndrome - an ophthalmologic approach to treatment. Poster-Vortrag, Deutsche Gesellschaft für Ophthalmologie, 2011 Blitzlichtfreie Aufnahme von Standard-Dynamic-Range- und High-Dynamic- Range-Epithelfotografien, Poster-Vortrag, Deutsche Gesellschaft für Ophthalmologie, 2012

Danksagung 65
Danksagung Für die Betreuung dieser Arbeit möchte ich Professor Alfons Meindl und Professor Volker Schuster sehr danken. Bei Professor Meindl durfte ich bereits als Zivildienstlei-stender meine Begeisterung für wissenschaftliches Arbeiten und Molekulargenetik ent-decken. Ich danke Professor Meindl dafür, dass er mir als Doktorand dieses interessante Thema zur Bearbeitung überlassen hat. Großer Dank gebührt auch der von Ihm geleite-ten Arbeitsgruppe, in der ich freundlich und kooperativ aufgenommen wurde. Beson-ders Heide Hellebrand und Marko Bertone möchte ich für die Einarbeitung und Hilfe bei der Mutationsanalyse danken.
Ebenso großer Dank gilt Professor Schuster, der den medizinischen Aspekt der Arbeit betreut hat. Das Überlassen seiner Habilitationsschrift, in der die Klinik einer der beiden untersuchten Familien beschrieben ist, diente mir als Leitfaden. Seine Korrekturen für den JACI-Artikel und Anleitung und Unterstützung während des Review-Prozess haben maßgeblich dazu beigetragen, dass die Publikation in dieser hochrangingen Zeitschrift gelang. Ebenfalls möchte ihm für die Korrektur dieser Arbeit, seine Geduld sowie seine Ermutigungen danken, auch für den freundlichen Empfang in Leipzig und die zahlrei-chen telefonischen Besprechungen. Ich habe ich mich von Professor Schuster und Pro-fessor Meindl trotz der zuletzt räumlichen Entfernung sehr gut betreut gefühlt.
Dank gilt auch Professor Belohradsky, der das Thema mit meinen Beteuern angeregt hatte. Zusammen mit Professor Urban übernahm er das Erläutern der Untersuchungser-gebnisse mit den Patienten sowie deren Therapie und half, Krankheitsgeschichte und Verlauf zu ergänzen.
Meinen Eltern, Anita und Rudolf Horn möchte ich für das häufige Probelesen der Ar-beit danken. Meiner Mutter danke ich gesondert für die sprachliche Korrektur der engli-schen Publikation, was sicher zu deren Annahme beigetragen hat, meinem Vater für Diskussionen und interssiertes Zuhören. Gewidmet ist die Arbeit meiner Frau Stefanie, mit großem Dank für ihr Verständis und Ihre Unterstützung bei vor dem Laptop ver-brachten Ferientagen und in Erinnerung an gegenseitiges Probelesen. Meine Familie war mir immer der größte Rückhalt - und ich freue mich unheimlich, dass diese in we-nigen Monaten weiter wachsen wird.





























