Embed Size (px)
Citation preview

Summary ___________________________________________________________________________________________
Characterization of the Novel Photosynthetic
Protein PPP7 involved in
Cyclic Electron Flow around PSI
Dissertation
zur Erlangung des Doktorgrades der Fakultät für Biologie
der Ludwig-Maximilians-Universität München
Vorgelegt von
Giovanni Dal Corso
aus Verona
Italien
München
Dezember 2007
Erstgutachter: Prof. Dr. Dario Leister Zweitgutachter: Prof. Dr. Jürgen Soll Tag der mündlichen Prüfung: 18 Dezember 2007

Summary ___________________________________________________________________________________________
Summary Photosynthetic organisms are able to convert light energy into chemical energy by the operation of the two photosystems, the cytochrome b6/f complex and the ATPase. The two photosystems operate in series during linear electron flow to split H2O and to generate NADP+. During electron transport, a pH gradient is generated across the thylakoid membrane which is used for the generation of ATP. In addition to the linear electron transport mode, ATP can also be produced via cyclic electron flow around photosystem I (CEF). The physiological role of CEF in vascular plants with C3-type photosynthesis is still not solved. Potential functions of CEF are (i) the dissipation of excessive light energy by increasing non-photochemical quenching (NPQ); (ii) ATP synthesis during steady-state photosynthesis; (iii) the regulation of the stromal oxidation state under stress conditions and under conditions when the Calvin cycle is not available as a sink for NADPH. With exception of the thylakoid NADPH-dehydrogenase complex and the stromal protein PGR5, the components that contribute to CEF are still unknown. Obscure is also the regulation that controls the switch from linear to cyclic flow. We have identified a novel transmembrane protein, named PPP7, which is located in thylakoids of photoautotrophic eukaryotes. Mutants lacking PPP7 exhibit the same phenotype as plants missing PGR5. These mutants show reduced NPQ, decreased P700 oxidation and perturbation of ferredoxin-dependent CEF. The work described in this thesis demonstrates that PPP7 and PGR5 interact physically, and that both co-purify with photosystem I. PPP7 does also interact in yeast assays with the cytochrome b6/f complex, as well as with the stromal proteins ferredoxin (Fd) and ferredoxin-NADPH oxido-reductase (FNR), but PPP7 is not a constitutive component of any of the major photosynthetic complexes. In consequence, the existence of a PPP7/PGR5 complex integrated in the thylakoid membrane and facilitating CEF around PSI in eukaryotes, possibly by shuttling electrons together with ferredoxin and the FNR from photosystem I to the cytochrome b6/f complex, is proposed. Moreover, CEF is enhanced in the Arabidopsis psad1 and psae1 mutants with a defect in photosystem I oxidation in contrast to the cyanobacterial psae mutant which exhibits an decreased CEF, pointing to fundamental mechanistic differences in the cyclic electron flow of cyanobacteria and vascular plants. The Arabidopsis psad1 and psae1 mutants also show higher contents of ferredoxin and of the PPP7/PGR5 complex, supporting a role of PPP7 and PGR5 in the switch from linear to cyclic electron flow depending on the redox state of the chloroplast.
i

Zusammenfassung _____________________________________________________________________________
Zusammenfassung Photosynthetisch aktive Organismen sind in der Lage mit Hilfe der Photosysteme, des Cytochrom b6/f-Komplexes und der ATPase Lichtenergie in chemische Energie umzuwandeln. Wenn die beiden Photosysteme während des linearen Elektronentransports in Serie arbeiten wird Wasser gespalten und NADPH erzeugt. Der durch den Elektronentransport über die Thylakoidmembran aufgebaute Protonengradient dient schließlich der Synthese von ATP. Neben dem linearen Elektronentransport trägt auch der zyklische Elektronenfluß um das Photosystem I zur Produktion von ATP bei. Die physiologische Funktion des zyklischen Elektronentransports bei Gefäßpflanzen mit C3-Photosynthese ist jedoch noch immer nicht eindeutig geklärt. Potentielle Funktionen des zyklischen Elektronentransportes könnten sein: (i) die Ableitung von überschüssiger Lichtenergie durch Erhöhung des nicht photochemischen Quencheffektes (NPQ); (ii) die ATP Synthese während der Photosynthesereaktion im stationären Zustand; oder (iii) die Regulation des stromalen Redoxzustandes unter Stressbedingungen und Bedingungen, unter denen der Calvin-Zyklus nicht als Akzeptor von NADPH zur Verfügung steht. Mit Ausnahme des in der Thylakoidmembran lokalisierten NADPH-Dehydrogenase Komplexes und des stromalen Proteins PGR5 sind bisher keine weiteren Komponenten bekannt, die zum zyklischen Elektronentransport beitragen. Unklarheit herrscht auch über die Mechanismen, die ein Umschalten von linearem zu zyklischem Elektronentransport kontrollieren. In dieser Arbeit wird die Identifizierung eines Transmembranproteins mit der Bezeichnung PPP7 beschrieben, welches in der Thylakoidmembran von photoautotrophen Eukaryoten lokalisiert ist. Mutanten, denen PPP7 fehlt, zeigen den gleichen Phänotyp wie Pflanzen, die kein PGR5 exprimieren. Beide Mutanten weisen reduzierten NPQ, erniedrigte Oxidation von P700 als auch eine Störung des Ferredoxin-abhängigen zyklischen Elektronentransport auf. Die Arbeiten, die in dieser Dissertation beschrieben werden, zeigen, dass PPP7 und PGR5 direkt miteinander interagieren und dass beide mit Photosystem I aufgereinigt werden können. Das PPP7 Protein interagiert im Hefesystem auch mit dem Cytochrom b6/f-Komplex und mit den stromalen Proteinen Ferredoxin und Ferredoxin-NADPH-Oxido-Reduktase (FNR), ist jedoch kein konstitutiver Bestandteil der Photosysteme oder von Cytochrom b6/f oder der ATPase. Wir postulieren daher das Vorhandensein eines in die Thylakoidmembran integrierten Komplexes aus PPP7 und PGR5, der den zyklischen Elektronenfluß um PSI bei Eukaryoten unterstützt, indem er mit Ferredoxin und FNR interagiert und somit möglicherweise als Elektronentransporter zwischen PSI und dem Cytochrom b6/f-Komplex fungiert. Zudem ist bei den Arabidopsis Mutanten psad1 und psae1 mit gestörter Oxidation von Photosystem I ein verstärkter zyklischer Elektronentransport messbar, ganz im Gegensatz zu der cyanobakteriellen psae Mutante mit erniedrigtem zyklischen Elektronentransport; dies impliziert grundlegende Unterschiede zwischen Cyanobakterien und Pflanzen beim zyklischen Elektronentransport. Die psae1 und psad1 Mutanten bei Arabidopsis haben zudem einen erhöhten Gehalt an Ferredoxin und an PPP7-PGR5, was auf eine Beteiligung von PPP7 und PGR5 am Umschalten von linearem auf zyklischen Elektronentransport in Abhängigkeit vom Redox-Zustand des Chloroplasten hinweist.
ii

Contents ________________________________________________________________________________________
Contents
Summary i
Zusammenfassung ii
Abbreviations 6
1. Introduction 8
1.1 Photosynthesis 8
1.2 Novel putative photosynthetic proteins 9
1.3 Cyclic electron flow: from PSI to PSI 9
1.4 Measuring cyclic electron flow 11
1.5 Models of cyclic electron flow 13
1.6 The role of cyclic electron flow in C3 plants 14
1.7 The role of cyclic electron flow in C4 plants 16
1.8 The role of cyclic electron flow in cyanobacteria
and unicellular eukaryotes 17
1.9 Cyclic electron flow: a genetic approach 18
1.10 Aim of the thesis 22
2. Material and Methods 23
2.1 Plant materials and growth conditions 23
2.2 Complementation of ppp7ab mutant 23
2.3 Nucleic acid analysis 24
2.4 Synthesis of antibodies against the PPP7 protein 25
2.5 Chlorophyll fluorescence and spectroscopic measurements 25
2.6 Measurements of the redox state of P700 and of CEF to LEF transition 25
2.7 Pigment analysis 26
2.8 Immunoblot analyses 26
2.9 Total protein preparation 27
2.10 Isolation of intact chloroplasts 27
2.11 PSI isolation 28
2.12 In vitro assay of ferredoxin-dependent plastoquinone reduction 28
4

Contents ________________________________________________________________________________________
2.13 In vitro import in pea chloroplasts 28
2.14 Intracellular localization of dsRFP fusion in Arabidopsis protoplasts 30
2.15 2D Blue Native/SDS PAGE for thylakoids protein analysis 31
2.16 Yeast two hybrid and split ubiquitin assay 31
3. Results 34
3.1 Gene structure and homologies 34
3.2 PPP7A and PPP7B are targeted to the thylakoid membrane 36
3.3 Topological orientation of PPP7 in the thylakoid membrane 38
3.4 ppp7a and ppp7b mutations 40
3.5 Complementation of the mutations 42
3.6 The mutations do not cause any loss in the content of thylakoid proteins 43
3.7 Mutants lacking PPP7s are impaired in photosynthesis 44
3.8 Mutants lacking PPP7s are impaired in cyclic electron flow around PSI 46
3.9 PPP7 interacts in vitro with PGR5, Fd, FNR, PSI and cyt b6/f 49
3.10 PPP7 interacts in planta with PSI and PGR5 51
3.11 PPP7/PGR5-PSI interaction and PSI organization 53
3.12 CEF and abundance of Fd, FNR, PPP7 and PGR5 55
3.13 Synthetic phenotype of the ppp7ab psad1-1 triple mutant 56
4. Discussion 59
4.1 PPP7: a novel component of CEF around PSI 59
4.2 PPP7 and PGR5 proteins are involved in the same pathway of CEF 60
4.3 A new model for CEF around PSI 62
4.4 Cyanobacteria do not have PPP7 64
4.5 Independent accumulation of PPP7 from PGR5 65
4.6 Link between LEF and CEF 66
4.7 CEF and state transition 68
5. References 69
Acknowledgement 83
Curriculum vitae 84
Ehrenwörtliche Versicherung 86
5

Abbreviations ________________________________________________________________________________________
Abbreviations
°C Degree Celsius
35SCaMV 35S promoter of the Cauliflower Mosaic virus
ATP Adenosine triphosphate
BN Blue native
cDNA Complementary deoxyribonucleic acid
CEF Cyclic electron flow
Chl Chlorophyll
cyt b6/f Cytochrome b6/f complex
Da Dalton
DNA Deoxyribonucleic acid
ETR Electron transport rate
Fd Ferredoxin
FNR Ferredoxin NADP oxido-reductase
g Gram
h Hour
HPLC High performance liquid chromatography
L Liter
LB Left border
LEF Linear electron flow
LHC Light-harvesting complex
Lu Lutein
m Meter
M Molarity
min Minute
mol Mole
NADP+/H Nicotinamide adenine dinucleotide phosphate
NDH NADPH deydrogenase complex
or NAD(P)H-plastoquinone-oxidoreductase complex
6

Abbreviations ________________________________________________________________________________________
NPQ Non-photochemical quenching
Nx Neoxanthin
OEC Oxygen evolving complex
P700 PSI reaction centre
PAGE Polyacrylamide gel electrophoresis
PAM Pulse amplitude modulation
PCR Polymerase chain reaction
PFD Photon flux density
PGR5 Proton gradient regulation 5
PPP7 Putative photosynthetic protein 7
PQ Plastoquinone
PS Photosystem
qP Photochemical quenching
qE ΔpH-dependent NPQ
RB Right border
RFP Red fluorescent protein
RNA Ribonucleic acid
ROS Reactive oxygen species
RT-PCR Reverse transcriptase-polymerase chain reaction
s Second
SD Standard deviation
SDS Sodium dodecyl sulphate
T-DNA Transfer-DNA V/V Volume per volume
VAZ Xanthophyll cycle pigments W/V Weight per volume
WT Wild-type
β-Car β-carotene
β-DM n-dodecyl β-D-maltoside
ΦII Effective quantum yield of photosystem II
7

Introduction ________________________________________________________________________________________
1. Introduction
1.1 Photosynthesis
Life on Earth ultimately depends on the energy coming from the sun and photosynthesis is
the only known biological process that can utilise this energy and convert it into chemical
energy and organic substances. Photosynthetic organisms (from photosynthetic prokaryotes
to flowering plants) use solar energy to synthesize carbon compounds from carbon dioxide
and water. In photosynthetic organisms, the photosynthetic reactions take place in a
specialized organelle derived from ancient photosynthetic prokaryotes, the chloroplast. In
the chloroplast, light energy is converted into chemical energy by a number of different
complexes working together, all embedded in the thylakoid membranes. As described
schematically in Figure 1.1 light energy drives the electron transfer from photosystem II
(PSII) via photosystem I (PSI) to the final electron acceptor NADP+. Coupled with the
linear electron transport, protons (H+) are transported into the thylakoid lumen by the Q-
cycle at the cytochrome b6/f complex. The activity of the oxygen evolving complex (OEC)
located at the PSII contributes to increase the H+ amount in the lumen. These protons then
diffuse to the ATP synthase, where their diffusion down the electrochemical potential
gradient is used to synthesize ATP in the stroma. ATP and NADPH are subsequently used
in the stroma as an energy source and reducing power for the biosynthesis of organic
compounds like synthesis of amino acids, nucleotides, fatty acids and lipids, vitamins,
hormones and assimilation of sulphur and nitrogen.
Fig 1.1. Scheme of electron transport chain in thylakoids. From Jon Nield, Imperial College London, 2000.
8

Introduction ________________________________________________________________________________________
The electron transfer from H2O to NADP+ through the PSII - cyt b6/f - PSI complexes is
termed linear electron flow (LEF). Already in 1955 Arnon et al. described a pathway of
cyclic electron flow (CEF) around PSI responsible for the generation of a ΔpH across the
thylakoid membrane without accumulation of NADPH. In this pathway, electrons of the
PSI go back to the cytochrome b6/f complex and than again to PSI via plastocyanin.
1.2 Novel putative photosynthetic proteins
Over the past few decades, knowledge about the organization and function of all the major
super-complexes forming the photosynthetic machinery has been accumulated, but many of
the minor components or proteins embedded or simply associated to the thylakoid
membrane have not been identified yet. A powerful tool to attribute a function to an
unknown gene is the so called “transcriptomics” approach in which the expression patterns
of genes are compared. This approach is based on the assumption that genes with
correlated expression patterns are most likely to have related functions. Those groups of
genes are known as regulons (cluster of genes of co-expressed profiles). In 2005, the
definition and characterization of regulons of a number of nuclear genes of Arabidopsis
thaliana have been reported (Biehl et al., 2005). The mRNA expression of 3292 nuclear
genes, most of them coding for chloroplast proteins (Richly et al., 2003) was determined
under a total of 101 different environmental and genetic conditions. This analysis showed
that many photosynthetic genes and genes for proteins of the plastid ribosome can be
grouped together on the basis of their very similar expression pattern. In other words, the
co-regulation of the expression of genes for photosynthetic proteins and of genes for plastid
ribosome can be interpreted as a mechanism to ensure that the subunits of those multi-
protein complexes are synthesized in matching amounts (Biehl et al., 2005). One of the
unknown proteins being co-regulated with photosynthetic genes is the subject of this thesis
and has been named PPP7 (Putative Photosynthetic Protein 7).
1.3 Cyclic electron flow: from PSI to PSI
Although the in vivo occurrence of CEF has been subject of controversy, it is well
established in isolated systems and in vitro models (Johnson, 2005). Most of the still
unsolved questions about CEF concern the components that play a role as donors, sinks and
9

Introduction ________________________________________________________________________________________
carriers of electrons. Since it was observed that CEF enhances the formation of a ΔpH
(Arnon, 1965), it was postulated that there must exist an enzyme that transfers electrons to
the plastoquinone pool which then would supply the cytochrome b6/f complex with
electrons, assuming that the plastoquinone pool is the ultimate electron acceptor prior to
cytochrome b6/f.
Once the electrons are transferred to the oxidised plastoquinone pool, they most probably
follow the normal way through cytochrome f and palstocyanin to P700 of PSI (Johnson,
2005). Regarding the route that electrons follow up-stream of PQ, it has been reported that
ferredoxin is required as a cofactor for cyclic photophosphorylation (Tagawa et al., 1963)
and that this pathway is sensitive to antimycin A, an inhibitor known to interact with the Qi
binding pocket on the stromal side of the cytochrome b6/f complex. However, Moss and
Bendall (1984) showed that HQNO (2-(n-heptyl)-4-hydroxyquinoline N-oxide, an inhibitor
of cytochrome b) does not affect CEF, leading to the idea that a distinct enzyme should be
involved. Unfortunately, until now there exists no biochemical evidence for the presence of
this putative ferredoxin-plastoquinone oxidoreductase enzyme (FQR). One of the possible
pathways that electrons could follow from the acceptor side of PSI to the PQ pool is known
as the “ferredoxin dependent-antimycin sensitive” pathway. In 2001, Zhang et al. could
show that ferredoxin-NADPH-oxidoreductase (FNR) binds tightly to the cytochrome b6/f
complex suggesting the formation of a complex together with ferredoxin, which would
then create a bridge for the electron transfer to the PQ pool.
Evidences also exist for another pathway that is insensitive to antimycin A. It involves the
NDH enzyme (NADPH dehydrogenase) located in the thylakoid membrane. This enzyme
is homologous to the NADH dehydrogenase complex (complex I) of mitochondria. The
first evidences that the NDH enzyme is involved in CEF have been found in the
cyanobacterium Synechocystis (PCC6803). It was found that a mutant defective in ndhB
has an impaired CEF around PSI (Mi et al., 1995). Also the plastome of higher plants
contains at least 11 genes that code for subunits of the NDH complex (reviewed in
Shikanai, 2007). The plastidic NDH has a higher similarity to the cyanobacterial complex
than to the ones of the mitochondrial complex I within the same species, suggesting an
evolutionary and functional similarity for the NDH complexes of cyanobacteria and
chloroplasts. By using chloroplast transformation techniques, it has been shown that mutant
10

Introduction ________________________________________________________________________________________
plants defective in the NDH complex had a slightly reduced CEF. This suggests a role of
NDH in the electron transport from the stromal electron pool to PQ (Burrows et al., 1998,
Kofer et al., 1998, Shikanai et al., 1998). Many authors criticise that the detectable
concentration of the NDH enzyme is too low to sustain a significant rate of CEF (<1% per
photosynthetic electron chain, Sazanov et al., 1998, Joet et al., 2002), whereas others
argue that the rapidly induced cyclic flow is not compatible with the rather low activity of
this enzyme (Breyton et al., 2006). Answering the questions about mechanisms and
pathways followed by electrons to cycle around PSI is one of the last major tasks to be
solved in this research field of photosynthesis.
Fig 1.2. Possible routes followed by electrons from the PSI acceptor side to the PSI donor side. (Johnson, 2005).
1.4 Measuring cyclic electron flow
One of the main problems studying cyclic electron flow is the fact that the electron carriers
of the cyclic pathway are also involved in linear electron transport (i.e. cytochrome b6/f
complex, PSI, Fd and NADP+). For this reason, all developed techniques, both in vitro and
in vivo, try to determine the relative contribution of cyclic flow to the total electron flux
through the shared components. Some of the most commonly used techniques are briefly
explained in the following sections.
1. Chlorophyll fluorescence analysis. Using a PAM fluorometer, the electron flow can
be monitored in vivo by a change in chlorophyll fluorescence. In particular, the electron
flow from NADPH to the intersystem chain can be roughly estimated by transient
increase in chlorophyll fluorescence after a period of illumination with actinic light
11

Introduction ________________________________________________________________________________________
(Schreiber et al., 1986). The fact that far-red light, which activates predominantly PSI
photochemistry, is able to quench this increase in fluorescence demonstrates that
NADPH transfers electrons to the plastoquinone pool (Shikanai et al., 1998).
2. Fd-dependent PQ reduction assay. The reduction of plastoquinone can be monitored
as an increase in chlorophyll fluorescence emitted after the exposure to light of a very
low intensity (1 µEm-2s-1). At such low light intensities, the fluorescence reflects the
reduction of plastoquinone by cyclic electron transport from ferredoxin, not by PSII
photochemistry (Munekage et al., 2002). This system can be applied to in vitro
ruptured chloroplasts, using exogenous ferredoxin and NADPH as an electron source.
3. Photoacoustic techniques. The photoacoustic method is based on the ability of
modulated light to produce a sound upon absorption (Malkin and Canaani, 1994). It
was developed to quantitatively evaluate the storage of photosynthetic energy by cyclic
electron flow in intact leaves or algae, as well as in isolated thylakoids (Herbert et al.,
1990). In brief, photoacoustic signals are used to quantify the conversion of absorbed
light to heat in a sample. If no photochemistry is performed by the sample, the
conversion is 100%. However, if some of the light energy is stored as photochemistry
products, e.g. in leaf or algal sample performing photosynthesis, it is not available for
conversion to heat anymore and changes the thermal photoacoustic signal in a way that
can be quantified. Using far red light as an exciting source, only PSI is functioning and
the photoacoustic signal reflects the cyclic electron flow (Joet et al., 2002). Using this
technique, energy storage due to CEF could be observed in cyanobacteria, algae and C4
plants, and to a lesser extent also in C3 plants (Herbert et al., 1990; Joet et al., 2002).
4. P700 re-reduction measurements. The illumination of a leaf with far red light (λ >
700nm) excites only PSI and not (or only partially) PSII. Under those conditions, the
rate of electron flow from PSII to PSI is negligible (Joliot and Joliot, 2005). After a
period of illumination, the light is switch off and the P700 redox state can be measured
as a change in absorbance at 700 nm or at the near infrared range (810-860 nm)
(Johnson, 2005). The reduction rate of P700 immediately after the cessation of light is
thought to be proportional to the rate of electron flow occurring in light through PSI. A
general accepted approach is to assume that the proportion of P700 in the reduced state
is a value for the quantum efficiency of PSI (Harbinson and Woodward, 1987). The re-
12

Introduction ________________________________________________________________________________________
reduction kinetics of P700 in the dark accelerates when electrons coming from the
stroma are donated to PSI. This fast reduction has been observed in algae,
cyanobacteria and C4 plants. This reduction takes place also in C3 plants with a reduced
rate, probably due to a lower NDH activity (Munekage and Shikanai, 2005).
5. Use of inhibitors. Methylviologen is well known to accept electrons from PSI (Ivanov
et al., 2007) forcing the photosynthetic chain to operate only in linear electron flow
(Joliot and Joliot, 2005). The involvement of PQ and the cytochrome b6/f complex in
CEF around PSI was shown by studies using inhibitors like DCMU (3-(3,4-
dichlorophenyl)-1,1-dimethiylurea) which binds to the QB site of PSII or HQNO (2-
heptyl-4-hydroxyquinolines) which binds to the cytochrome b6/f complex (Munekage
and Shikanai, 2005). In these studies, DCMU was shown not to inhibit cyclic electron
transport (Bendall and Manasse, 1995). Finally, a vexing question is still open about the
activity of antimycin A, an inhibitor supposed to bind the Qi binding site of the
cytochrome b6/f complex interrupting the Fd-dependent CEF pathway (Okegawa et al.,
2005).
1.5 Models of cyclic electron flow
CEF and LEF share a number of common electron carriers, from plastoquinone to
ferredoxin (Breyton et al., 2006). In the following, different models on the organization of
the “cyclic” machinery are discussed:
1. The diffusion model. It is generally accepted that the different multi-protein
complexes are heterogeneously distributed in the thylakoid membrane; in particular,
PSII is mostly located in the grana stacks while PSI is more abundant in the non-
compressed stromal lamellae, in the margin and end membranes of the grana stacks
(Allen and Forsberg, 2001). The cytochrome b6/f complex, on the other hand, is
homogenously distributed over the thylakoid membranes (Albertsson 2001). In this
model, only the cytochrome b6/f complexes located in the non-appressed membranes
would receive electrons from PSI resulting in CEF, while in the grana stacks the
plastoquinone pool is exclusively reduced by PSII (Albertsson 2001; Bukhov and
Carpentier, 2004).
13

Introduction ________________________________________________________________________________________
2. The super-complex model. Already in 1967 Boardman and Anderson found super-
complexes of PSI and cytochrome b6/f in Chlamydomonas and vascular plants. FNR is
also known to associate with cytochrome b6/f (Zhang et al., 2001). These findings led
to the postulation of a tightly associated complex of FNR, PSI and cytochrome b6/f
which would take over electrons from ferredoxin, fuelling the cyclic pathways (Joliot
and Joliot, 2002). Recent biochemical approaches aimed to confirm an eventual FNR-
PSI-cytochrome b6/f super-complex failed, however, to prove its existence (Breyton et
al., 2006).
3. The FNR model. FNR has been found to be associated with both PSI (Scheller et al.,
2001) and the cytochrome b6/f complex (Zhang et al., 2001). Joliot and Joliot (2005)
proposed that PSI-FNR complexes transfer electrons from Fd to NADP+, via linear
electron flow, while the population of FNR associated with the cytochrome b6/f
complex is responsible for the cyclic electron pathway from ferredoxin (reduced by
those PSI not associated with the FNR) to plastoquinone (Figure 1.3).
Fig 1.3. Possible routes followed by electrons from the PSI acceptor side to the PSI donor side.
(SII: PSII and SI: PSI), from Joliot and Joliot (2002).
1.6 The role of cyclic electron flow in C3 plants
There are two major conditions under which CEF plays a significant role in photosynthesis:
1. during the transition dark-to-light. During this brief period of time (within minutes),
the majority of the enzymes involved in the Calvin cycle have to be newly synthesized
or activated, most importantly Rubisco, fructose-1,6-bisphostphate phosphatase,
sedoheptulose-1,7-bisphospate phosphatase, ribulose-5-phosphate kinase and NADP-
glyceraldehyde-3-phosphate dehydrogenase. Moreover, the main intermediate of the
14

Introduction ________________________________________________________________________________________
Calvin cycle (ribulose-1,5-bisphosphate) has to be synthesized. As an example, the
activation of Rubisco requires ATP and an increase in the stromal pH (Campbell and
Ogren, 1992)). The activation of the other enzymes listed before requires reduced
ferredoxin, which then can activate the thioredoxin regulatory system. Reduced
ferredoxin, increase in stromal pH and ATP production are all factors influenced by
cyclic electron flow (Munekage et al., 2002). Indeed, during the first minutes of
illumination of dark adapted leaves, high rate of CEF can be measured (Laisk et al.,
2005; Joliot and Joliot, 2002).
2. during steady state of photosynthesis. In contrast to LEF, CEF can generate a ΔpH
without accumulation of NADPH. It can also modify the ratio between proton
translocation and electron transport and eventually the ratio of ATP/NADPH
production (Munekage and Shikanai, 2005). Owing to only linear electron flow, the
ATP/NADPH ratio is 1.29. On the other hand, the requirement for the Calvin Cycle is
1.5 to 1.66 depending on the amount of photorespiration (Osmond, 1981) that
corresponds to roughly 20% more of ATP. It has been proposed that CEF could provide
this ATP portion needed for the activity of the Calvin cycle during steady state
photosynthesis. This could explain the decreased fitness of mutants defective in cyclic
electron flow (see below in this section).
To prevent photoinhibition caused by excessive absorption of light energy, plants
developed different mechanisms. The most effective is thermal dissipation, which is
induced by the generated ΔpH across the thylakoid membrane (Müller et al., 2001). In this
respect, CEF may regulate the induction of thermal dissipation modifying the generation of
the ΔpH (Heber and Walker, 1992). Supporting this hypothesis, in 1999 the Arabidopsis
thaliana mutant pgr5 has been discovered as a high chlorophyll fluorescence mutant
impaired in CEF (Shikanai el al., 1999).
The finding that in ruptured chloroplasts, exogenous electron donors like ferredoxin or
NADPH, trigger cyclic electron transport independently from PSI photochemistry (Mills et
al., 1979), suggests that CEF is promoted by reducing power within the chloroplast. In
other words, CEF might be regulated by the redox state of the chloroplast (Bukhov et al.,
2002; Nandha et al., 2007).
Under stress conditions, such as high light, the partial pressure of available CO2 for the
15

Introduction ________________________________________________________________________________________
Rubisco activity is reduced, while the NPQ increases. In tobacco, for example, the activity
of CEF relative to that of LEF is enhanced under stress conditions, contributing to the
induction of NPQ by generating a ΔpH across the membrane (Miyake et al., 2005). Indeed,
tobacco mutants defective in the NDH complex are more sensitive to high light stresses
(Endo et al., 1999). As already confirmed by in vitro experiments (Endo et al., 1998),
reduced forms of Fd and NADPH lead to the activation of CEF pathways.
Other critical conditions for a plant’s performance are drought and high temperatures,
similar conditions under which the stomatal closure and a higher transpiration rate create a
reduced availability of CO2. Furthermore, high temperatures decrease the Rubisco activity
leading to an over-reduction of the stromal environment (Crafts-Brandner and Salvucci,
2000). Under such conditions, CEF is activated, generating a proton gradient that induces
NPQ which is involved in dissipating excessive energy (Golding and Johnson, 2004).
CEF could be considered as a “safety valve” in which electrons are (re)cycled around PSI
and thus, oxygen reduction and consequently reactive oxygen species (ROS) production are
minimized. According to this hypothesis, PSI of the pgr5 mutant is highly sensitive to
photoinhibition even at low light intensities (Munekage et al., 2002). This cannot be
attributed to a lack of NPQ, since this effect does not occur in mutants with blocked qE
(e.g. npq4, Munekage and Shikanai, 2005).
Recently Arabidopsis mutant, pgr5 crr, has been generated defective in both the Fd-
dependent (pgr5 mutation) as well as in the NDH dependent (crr mutation) pathways. The
mutant plants show a dramatic reduction in photosynthetic growth (Munekage et al., 2004),
suggesting that cyclic electron transport around PSI is required for efficient photosynthesis
and autotrophic growth. However, the lack of a generally accepted technique to measure
CEF in vivo and the still limited knowledge of the molecular mechanisms that drive CEF
result in contrary hypotheses about the role of CEF around PSI in C3 plants especially
under environmental growth conditions.
1.7 The role of cyclic electron flow in C4 plants
In the bundle-sheath cells of C4 plants, cyclic electron flow is thought to play an important
role. It is well known that C4 plants can generate a high CO2 concentration by minimizing
16

Introduction ________________________________________________________________________________________
the dissipating oxygenase activity of Rubisco, but such alternative C fixation system costs
them about two additional molecules of ATP to fix one molecule of CO2 compared with C3
plants. Moreover, in the bundle-sheath cells of C4 plants, the reduced packing of the
thylakoids in grana stacks gives a higher PSI/PSII ratio than in C3 plants (Takabayashi et
al., 2005). This thylakoid organization determines that in the bundle-sheath cells, PSII
amounts and activity are highly reduced compared to the mesophyll cells (Romanowska et
al., 2006). This strongly suggests that no significant linear electron transport occurs in
bundle-sheath thylakoids (Bassi et al., 1995). An enhancement of CEF would provide the
extra ATP that could be used for CO2 fixation reactions. Recently it has been found that in
the C4 plants belonging to the NADP-ME family (e.g. Sorghum bicolor and Zea mays) the
amount of the NDH complex is significantly higher in bundle-sheath cells in comparison to
the mesophyll’s and at least ten times higher than in the thylakoids of the C3 plant tobacco.
Interestingly, the homologue of the Arabidopsis PGR5 protein accumulates rather
uniformly in mesophyll and bundle-sheath cells and the amounts are comparable with those
of tobacco or Arabidopsis. These findings indicate that CEF plays a role in the production
of additional ATP in the C4 metabolism (Takabayashi et al., 2005).
1.8 The role of cyclic electron flow in cyanobacteria and unicellular eukaryotes
NDH activity has been found in both cyanobacteria (Mi et al., 1992, 1994) and algae
(Seidel-Guyenot et al., 1996). So far eleven genes coding for the subunits of this complex
have been identified in the genomes of both types of organisms.
a) Cyanobacteria. The first aspect that has to be considered is that the diazotrophic
cyanobacteria need to separate nitrogen fixation from photosynthesis, since that the
nitrogenase enzyme is highly sensitive to molecular oxygen (Berman-Frank et al., 2003).
To prevent inhibition, the nitrogenase complex is confined in specialized cells called
heterocysts. Since heterocysts lack O2-producing photosystem II (Wolk et al., 1994) they
generate ATP through CEF around photosystem I (Ernst et al., 1983). Moreover,
Synechococcus and Synechocystis mutants lacking or being defective in the NDH complex,
show impaired inorganic carbon assimilation (Bukhov and Carpentier, 2004) and show
impaired CEF around PSI. Additionally, in Synechocystis the homologue of PGR5
17

Introduction ________________________________________________________________________________________
(ssr2016) has been reported (Yeremenko et al., 2005). As in flowering plants, this protein
is thought to play a role in an antimycin-A sensitive pathway of CEF.
Thus, it has been suggested that the ssr2016 pathway is not a major contributor to CEF but
has a regulatory function, sensing the redox balance of the cytoplasm. In fact it is strongly
induced by high light intensities or salt stress (Allakhverdiev et al., 2002; Kanesaki et al.,
2002). Under such circumstances, the most of the required ATP, needed for repair and
stress-activated mechanisms, could be supplied by CEF (Thomas et al., 2001). It has been
found that subunits of PSI (for instance PsaE) are involved in CEF (Zhao et al., 1993; Yu
et al., 1993) and the ndh/psaE double knock out does not show any CEF around PSI (Yu et
al., 1993). Besides the NDH- and the PGR5-dependent pathways, there is evidence for the
existence of further pathways of PSI-mediated CEF, differentially induced by
environmental conditions (Cooley et al., 2000; Matthijs et al., 2002).
b) Algae. Unlike in vascular plants, in unicellular green algae (e.g. Chlamydomonas) a
large fraction of the LHCII (about 80%) migrates to PSI under light conditions favouring
PSII (Allen, 1992). This phenomenon, known as state transition, causes a switch between
linear and cyclic electron flow. The finding that the injection of electrons in the
cytochrome b6/f complex is insensitive to the addition of PSII inhibitors during state 2
confirmed that electrons re-cycle (Finazzi et al., 1999, Finazzi and Forti, 2004). The
plastome of Chlamydomonas does not code for the 11 genes of the subunits of the NDH
complex (Shikanai, 2006), but its genome codes for a homologue of the Arabidopsis PGR5
protein, supporting an involvement of this pathway in this switch between linear and cyclic
flow, even though the precise role and contribution of it is still not clear.
1.9 Cyclic electron flow: a genetic approach
As mentioned before, it is generally accepted that in flowering plants exist two partially
redundant routes that electrons can follow to cycle around PSI. One is via the thylakoid
NDH complex (Shikanai et al., 1998), and the other is probably involving ferredoxin and
the protein PGR5 (Munekage et al., 2002).
The NADPH dehydrogenase complex
The thylakoidal NDH complex is a multi-protein complex whose structure has not been
elucidated yet. The plastome contains 11 genes (ndhA to ndhK) that are homologues to the
18

Introduction ________________________________________________________________________________________
bacterial ndh genes. Further three subunits are present in bacteria, but not encoded by the
plastome of higher plants. These missing subunits are involved in the formation of the
docking site for NAD(P)H and its oxidation (Rumeau et al., 2005). The nature of the
“electron input subunit” of the plastidial NDH is still unclear. It has been proposed that
FNR could interact with the NDH complex supplying electrons derived from NADPH
reduction. Recently it has been suggested that also reduced ferredoxin could represent an
electron source, in which case the FNR enzyme would play a significant role as an electron
shuttle (Shikanai, 2007). Alternatively, in plastidial NDH a new and still unidentified
electron input module could be present even if the attempts to purify the plastidial complex
did not reveal any missing subunits involved in electron input (Rumeau et al., 2005). In the
1990s by means of plastome transformation techniques (Svab and Maliga, 1993), almost all
of the 11 plastidial ndh genes were inactivated. The different knockout lines displayed an
alteration in electron transport in chloroplasts and an inhibited reduction of the
plastoquinone pool. In particular, lines with an altered or missing thylakoid NDH complex
showed reduced levels of a transient increase in fluorescence (see Figure 1.4) which could
be attributed to the electron transfer from the reduced stromal electron pool to the PQ pool
(Burrows et al., 1998; Kofer et al., 1998; Shikanai et al., 1998). Recently, the identification
of three novel nuclear-encoded subunits for the plastidic NDH (NDH-M, -N and -O) has
been reported. The corresponding mutants show a phenotype comparable to the other ndh-
mutants (Rumeau et al., 2005).
Fig 1.4. Typical fluorescence phenotype of a mutant without functional plastidial NDH: the
transient increase of the fluorescence after switching off the light (AL off) is reduced. (Adapted
from Kofer et al. (1998); ML: measuring light, AL: actinic light; Fo: base fluorescence).
WT ndh
19

Introduction ________________________________________________________________________________________
The antimycin A sensitive pathway
Given that cyclic electron flow is involved in ΔpH generation, it should play a role in the
induction of non-photochemical quenching (Heber and Walker, 1992). In case a mutant is
defective in CEF it should be identifiable due to its defective NPQ phenotype. As already
discussed, the amount of NDH is too low for a significant contribution of this complex to
an increase in NPQ (Shikanai et al., 1998). Thus, any mutant defective in CEF with a
marked NPQ reduction should be defective in a NDH-independent pathway. Using
chlorophyll fluorescence imaging, the Arabidopsis mutant pgr5 (proton gradient
regulation) has been identified based on its NPQ phenotype (Shikanai et al., 1999). In
pgr5, the ratio of P700 oxidised to P700 reduced is lowered at high light intensities, in
contrast to the WT. The WT phenotype can be restored by infiltration with methylviologen
(electron acceptor of PSI) indicating that electron transport is limited at the acceptor side of
PSI (Munekage and Shikanai, 2005). LEF was not affected in isolated thylakoids of pgr5,
while it is reduced in vivo (Munekage et al., 2005). The ferredoxin-dependent
plastoquinone reduction was also assayed in ruptured chloroplasts of pgr5, and it has been
shown that this pgr5 phenotype can be “simulated” in WT chloroplasts by antimycin A
treatment. PGR5 encodes for a 10 kDa protein associated with the thylakoid membrane
(Munekage et al., 2002). PGR5 does not contain any known metal-binding or
transmembrane motif and is stable in mutant backgrounds lacking PSII, PSI, the
cytochrome b6/f complex or ATPase, suggesting that PGR5 is not a constituent of any of
these major complexes (Munekage and Shikanai, 2005). The exact localisation of PGR5 in
the thylakoid membrane is still not clear and its role in CEF is still under debate.
A knock-out mutant entirely lacking CEF
Recently, the group of Shikanai created a series of double mutants by crossing mutants
missing or defective in the NDH complex (crr mutants) and pgr5 (Munekage et al., 2004).
The double mutants showed a retarded development and growth, pale green colour and
defects in linear electron flow (Figure 1.5).
20

Introduction ________________________________________________________________________________________
Fig 1.5. Growth phenotypes of WT, crr2-2, crr3, crr4-2 and pgr5 single mutants as well as the
crr4-2pgr5, crr3pgr5 and crr2-2pgr5 double mutants. From Munekage et al. (2004).
The plastoquinone reduction activity in ruptured chloroplasts of the double mutants is
completely abolished (see Figure 1.6). The treatment of ruptured chloroplast of the crr2-2
mutants with antimycin A mimics the effect of the double mutation. On the other hand, the
inhibitor does not have any effects on pgr5 mutant. This result clearly shows that CRR2
and PGR5 are involved in two different pathways of CEF and that the antimycin A is
affecting only the “PGR5 pathway” (Munekage et al., 2004).
Fig 1.6. Electron transfer to plastoquinone in ruptured chloroplasts measured as an increase in
chlorophyll fluorescence after the addition of NADPH and ferredoxin under illumination with weak
actinic light. From Munekage et al. (2004).
These findings, together with the fact that mutants lacking the entire CEF apparatus (crr
pgr5) are defective in photoautotrophic growth, show that CEF is important for plant
fitness and plays a crucial role in preventing stroma over-reduction (Munekage et al.,
21

Introduction ________________________________________________________________________________________
2004). In WT plants, this is achieved by adjusting the concentration of ATP and hence
keeping a correct ratio ATP/NADPH.
1.10 Aim of the thesis
In this thesis a new putative photosynthetic protein (PPP7) identified on the basis of its
transcriptional co-regulation with known photosynthetic genes was functionally
characterized. The first aim of the thesis was to clarify whether PPP7 is truly a new
thylakoid protein. The second aim was to characterize the function of the PPP7 protein by
identifying lines which lack the PPP7 protein in a reverse genetics approach and by
characterizing those loss-of-function lines on the physiological level. When it became clear
in the course of the thesis that PPP7 is involved in cyclic electron flow, an in-depth
biochemical and genetic characterization of the role of PPP7 in cyclic electron flow was
initiated, including the identification of its interaction partners. In a final step the entire set
of data was combined to develop a new model on how cyclic electron flow functions in
flowering plants.
22

Material and Methods _____________________________________________________________________________________
2. Materials and Methods
2.1 Plant materials and growth conditions
An At4g11960 (PPP7B) insertion mutant line was identified in the SALK collection
(http://signal.salk.edu/; Alonso et al., 2003) which is made up of flank-tagged ROK2 T-
DNA lines (ecotype Columbia-0). The At4g22890 (PPP7A) insertion mutant line derives
from the Sail collection (Session et al., 2002), which is made up of a flank-tagged DAP101
T-DNA lines (ecotype Columbia-0). Both lines were identified by searching the insertion
flanking database SIGNAL (http://signal.salk.edu/cgi-bin/tdna express).
pgr5 mutant seeds were kindly provided by Toshiharu Shikanai (Graduate School of
Agriculture, Kyushu University Fukuoka, Japan). Arabidopsis thaliana wild-type (ecotype
Columbia 0) and mutant seeds were sown in Petri dishes on water soaked Whatman paper
and incubated three days at 4°C in the dark to break dormancy. Plants were gown on soil
under greenhouse controlled conditions (PDF: 70-90µEm-2s-1, 16h light: 8h dark cycles).
Fertilization with “Osmocote Plus” (Scotts Deutschland GmbH, Nordon Germanz) was
performed according to manufacturer’s instructions.
2.2 Complementation of ppp7ab mutant
The PPP7A and PPP7B complete coding regions (primers: P7AF: 5’- ATGGGTAGCA
AGATGTTGTT-3’; P7AR: 5’-TTAAGCTTGGCTTCCTTCTGG-3’; P7BF: 5’-
ATGGCTTTTACTCTAACAAT -3’; P7BR: 5’- TTAAGCTTTCCCTCCTTCTG -3’)
were ligated into the plant expression vector pH2GW7 (Invitrogen) under the control of a
single Cauliflower Mosaic Virus 35S promoter and the constructs were used to transform
flowers of ppp7ab mutant plants by the floral dipping technique as described in Clough and
Bent (1998). Hygromycin resistant plants, selected in vitro, were transferred into the
greenhouse and seeds were collected after 3 weeks. Successful complementation was
confirmed by chlorophyll fluorescence measurement and P700 oxidation state analysis (see
below). In addition, the integration of the transgene in the genome of the complemented
mutant plants was confirmed by PCR using specific primers (22890F: 5’-
CTAAAGCCACAACAGAGCAATC-3’ and 22890R: 5’-TGTAATGTCGTCCAGGAA-
3’; 11960F: 5’-TACTCTAACAATCCCTAGATT-3’ and 11960R: 5’-CCTCCT
23

Material and Methods _____________________________________________________________________________________
TCTGGTAATGTGATC-3’) and the presence of the PPP7 protein was tested probing with
specific antibodies thylakoid isolations as described below.
2.3 Nucleic acid analysis
Arabidopsis genomic DNA was isolated grinding fresh material in grinding buffer (200
mM Tris (pH 7.5), 250 mM NaCl, 25 mM EDTA, 0.5% SDS) followed by isopropanol
precipitation. T-DNA insertion junction sites were recovered by polymerase chain reaction
(PCR) with the use of combinations of insertion-specific and gene-specific primers, and
then sequenced by means of the sequencing service at the LMU München
(http://www.genetik.biologie.uni-muenchen.de/sequencing). T-DNA primers specific for
ROK2 were LBb1 (5’-GCGTGGACCGCTTGCTGCAACTC-3’) and RBb1 (5’-
TCAGTGACAACGTCGAGCAC-3’). T-DNA primer specific for DAP101 was LB1 (5’-
GCCTTTTCAGAAATGGATAAATAGCCTTGCTTCC-3’). Primers specific for
At4g22890 were PPP7A1F (5’-CAAGATGCAGTCTCCGTAGT-3’) and PPP7A1R (5’-
GCTGGAGATTGACAGAATTGT-3’); for At4g11960, PPP7B1F (5’-TAACTCATCGT
TATGTGATCGA-3’) and PPP7B1R (5’-GTCCAAATTACACATGTAACAAG-3’); for
PGR5 (At2g05620), pg5841F (5’-AGGTGATCACTGAGTTTTGC-3’) and pg5961R (5’-
ATCAGACACAAGCAGAGAG-3’). For the latter, the PCR products were digested with
DdeI in order to discriminate between WT and mutant plants.
To determine gene expression, total leaf or root RNA was extracted from fresh tissues
using the TRIzol reagent (Invitrogen, Karlsruhe, Germany). First-strand cDNA synthesis
was performed using the SuperScript™ III Reverse Transcriptase (Invitrogen, Karlsruhe,
Germany), and reverse-transcriptase-mediated PCRs (RT-PCR) were performed, using
primers specific for At4g22890 (sense primer 5’-ATGGGTAGCAAGATGTTGTTTA-3’,
antisense primer 5’-CAACGGTTGCTGGAACATTC-3’) and At4g11960 (sense primer 5’-
TTTACTCTAACAATCCCTAGATTT-3’, antisense primer 5’-CCTCCTTCTG
GTAATGTGATCA-3’) as well as ACTIN1-specific primers as a control (sense primer 5'-
TGCGACAATGGAACTGGAATG-3', antisense primer 5'-GGATAGCATGTGG
AAGTGCATACC-3'). To determine the difference in the level of gene expression, Real
Time PCR has been performed on cDNA using specific primers on PPP7A, PPP7B and
PGR5 genes (ppp7aF: 5’-TCCTGGACGACATTACAG-3’ and ppp7aR: 5’-
24

Material and Methods _____________________________________________________________________________________
TGATTGATAAAGCCAGATAT-3’; ppp7BF: 5’-GGGAAGAAGTTGATAGCA-3’ and
ppp7bR: 5’-ATCGCTTTCGCTTCGTAATA-3’; pgr5F: 5’-AGTTCCAATGATGAA
GAATG-3’ and pgr5R 5’-GCAAAACTCAGTGATCACCT-3’).
2.4 Synthesis of antibodies against the PPP7 protein
Antibodies recognising both proteins PPP7A and PPP7B have been raised against the N
terminal domain of PPP7A. The cDNA regions corresponding to the N-Terminus of
PPP7A (from aa 61 to aa 200, primers: IG7F: 5’-CACCGCCACAACAGAGCAATCAG-
3’ and IG7R: 5’-TTATTTGAAATAATCTACAGCGAG-3’) was cloned in the pET151-
Topo vector (Invitrogen). The expression of the protein in E. coli cells (BL21-star) was
induced with 1 mM IPTG at 16°C overnight in LB medium (Sambrook et al., 1989). The
expressed truncated PPP7A protein was purified by means of its His-Tag using a Ni-
Agarose resin under denaturating conditions (Invitrogen) according to manufacturers’
instructions. The purified protein was then injected into the rabbits. Probing them on
thylakoids isolated from single ppp7a and ppp7b mutants, proved that the antibodies
recognized both proteins as expected from the high homology of the protein sequences.
2.5 Chlorophyll fluorescence and spectroscopic measurements
Chlorophyll fluorescence was measured in vivo on single leaves, using a PAM (pulse
amplitude modulation) 101/103 fluorometer (Walz, Germany) as described before by
Varotto et al. (2000b). Saturating pulses (800 ms) of white light (4500 µE m-2 s-1) were
used to determine the maximum fluorescence in the dark (FM) and after the illumination
(FM’) and the ratio (FM - F0)/FM corresponds to FV/FM. A 20-min illumination with actinic
light (80 µE m-2 s-1) served to drive electron transport between PSII and PSI before ΦII and
qP were measured according to the formulas (FM’-FS)/FM’ and (FM’-FS)/(FM’-F0). NPQ was
measured after 20 min illumination with actinic light of different light intensity, from 80 to
2000 µE m-2 s-1, as described in Results, and it was calculated as (FM-FM’)/FM’.
2.6 Measurements of the redox state of P700 and of CEF to LEF transition
Redox changes of P700 were measured by monitoring the absorbance at 810 nm and 860 nm
with a PAM 101/103 chlorophyll fluorometer (Walz) connected to a Dual Wavelength
25

Material and Methods _____________________________________________________________________________________
ED_P700DW emitter-detector unit as described by Schreiber et al. (1988). Oxidized P700
levels (ΔA) were recorded in vivo during actinic light illumination under different light
intensities (from 70 to 800 µE m-2 s-1). The maximum level of oxidised P700 (ΔAMAX) was
recorded during far red light illumination (720 nm, 50 µE m-2 s-1). The P700 oxidation state
was then calculated as ΔA/ΔAMAX.
CEF-to-LEF transitions were measured as P700 redox kinetics in intact leaves with a flash
spectrophotometer as described before (Nandha et al., 2007). Actinic light driving LEF was
provided by a green LED peaking at 520 nm, and P700 oxidation was measured at 820 nm
(Breyton et al., 2006). P700 was specifically excited by far-red light and the maximum
extent of P700+ was estimated from the kinetics of P700 oxidation as described (Joliot and
Joliot, 2005).
2.7 Pigment analysis
Pigment content was analysed by reverse-phase HPLC as previously described in Färber et
al. (1997). Leaf discs were weighted, frozen in liquid nitrogen and ground to powder.
Pigments were extracted with 95% acetone. After short centrifugation, pigment extracts
were filtered through a 0.2 µm membrane filter and used directly for HPLC analysis
performed in collaboration with Prof. Dr. Peter Jahns (Düsseldorf, Germany).
2.8 Immunoblot analyses
Leaves from 4-week-old plants were harvested and grinded in buffer 1 (0.4 M sorbitol, 0.1
M Tricine (pH 7.8-KOH) and 0.5% milk). After sieving the material through two layers of
Miracloth (Calbiochem) to remove cellular debris, chloroplasts were collected centrifuging
at 2,450 g for 5 min at 4°C in a Ja-25.50 (Beckman) rotor. The chloroplasts were
resuspended and lysed in buffer 2 (20 mM Hepes (pH 7.8-KOH) and 10 mM EDTA) and
thylakoids were collected by centrifugation at 12,000 g at 4°C for 10 min. Thylakoids were
resuspended in storage buffer (10 mM Hepes (pH 7.5-KOH), 1 mM EDTA and 50%
glycerol) and the chlorophyll concentration was determined measuring the absorbance at
different wavelengths (A750, A663.6 and A646.6) after acetone precipitation as described in
Porra, 2002. Thylakoids or total chloroplasts (see paragraph 2.10) were resuspended in two
volumes of SDS-loading buffer (6 M Urea, 50 mM Tris-Cl (pH 6.8), 100 mM
26

Material and Methods _____________________________________________________________________________________
dithiothreitol, 2% SDS, 10% glycerol, 0.1% bromophenol blue) and subsequently loaded
on acrylamide Tris-Tricine SDS-PAGE gradient gels (10 to 16% acrylamide) and
fractionated according to Schaegger and Jagow (1987). After overnight run at 100 V (12-20
mA, anode buffer: 0.2 M Tris-Cl (pH 8.9), cathode buffer: 0.1 M Tris-Cl (pH 8.9), 0.1 M
Tricine (pH 8.9), 0.1% SDS, 1 mM EDTA) the proteins were transferred to PVDF
membranes by means of semi-dry blotting apparatus (Biorad) according to Towbin et al.
(1979) using a current corresponding to 1 mA cm-2 in transfer buffer (96 mM Glycine, 10
mM Tris, 10% V/V methanol).
Filters were then probed with antibodies against individual subunits of PSI, PSII and cyt
b6/f according to standard protocols (Sambrook et al., 1989) and signals were detected by
enhanced chemo-luminescence (ECL kit, Amersham Biosciences). The PGR5 protein was
detected using antibodies gently provided by Toshiharu Shikanai (Fukuoka, Japan).
Both PPP7A and PPP7B proteins were detected by antibodies raised against the N terminal
part of PPP7A protein (see paragraph 2.4).
2.9 Total protein preparation
Leaves from 4-week-old plants were harvested and homogenized in solubilisation buffer
(100 mM Tris (pH 8), 50 mM EDTA (pH 8), 0.25 M NaCl, 1 mM DTT and 0.7% SDS).
The homogenate was heated at 65°C for 10 min and centrifuged at 16,000 g 10 min (at RT)
to remove cellular debris. Prior electrophoresis fractionations, proteins were precipitated
with ice-cold acetone following standard protocols (Sambrook et al., 1989) and
resuspended in SDS-loading buffer (6 M Urea, 50 mM Tris-Cl (pH 6.8), 100 mM
dithiothreitol, 2% SDS, 10% glycerol, 0.1% bromophenol blue).
2.10 Isolation of intact chloroplasts
Leaves of 4- to 5-week-old plants were homogenized in the homogenization buffer (330
mM sorbitol, 20 mM Tricine (pH 7.6), 5 mM EGTA, 5 mM EDTA, 10 mM NaHCO3, 0.1%
BSA and 330 mg/L ascorbate) and the homogenate was filtrated through two layers of
Miracloth (Calbiochem). Chloroplasts were collected by a centrifugation at 2,000 g for 5
min at 4°C. The pellet was carefully resuspended in the washing buffer (330 mM sorbitol,
20 mM HEPES/KOH (pH 7.6), 5 mM MgCl2 and 2.5 mM EDTA). Chloroplasts were
27

Material and Methods _____________________________________________________________________________________
loaded on a two-step Percoll gradient as described in Aronsson and Jarvis (2002). Intact
chloroplasts at the interface between the two Percoll phases were broken by incubation for
30 min on ice in four volumes lysis buffer (20 mM HEPES/KOH (pH 7.5), 10 mM EDTA)
and used for subsequent experiments. To separate thylakoids and stroma phases, ruptured
chloroplasts were centrifuged at 42,000 g, 30 min at 4°C.
2.11 PSI isolation
Leaves from 4-week-old plants were harvested and thylakoids were prepared following the
protocol described in paragraph 2.8. Thylakoids were washed twice in 5 mM EDTA (pH
7.8) and dilute to 2 mg mL-1 chlorophyll concentration. An equal volume of 2% n-dodecyl-
β-D-maltoside (β-DM) was added to the thylakoids and solubilisation was carried out on
ice for 10 min. The un-solubilised part was pelleted by centrifugation at 16,000 g for 5 min
at 4°C and the supernatant was loaded on a sucrose gradient (prepared directly into the
centrifuge tubes after a freezing-thawing cycle of a 0.4 M sucrose, 20 mM Tricine (pH
7.5), 0.06% β-DM solution). The gradients were centrifuge at 191,000 g (SW40 rotor) for
21 h at 4°C. The PSI migrated as a distinct band on the bottom of the gradient. PSI
isolations were separated in 16% to 23% acrylamide Tris-Glycine SDS-PAGE following
standard protocols (Sambrook et al., 1989).
2.12 In vitro assay of ferredoxin-dependent plastoquinone reduction
Ferredoxin-dependent plastoquinone reduction was measured in ruptured chloroplasts
diluted in lysis buffer (see paragraph 2.10) to 10 µg Chl mL-1 and immediately used for the
measurements of chlorophyll fluorescence with a PAM fluorometer 101/103 (Walz,
Germany). The fluorescence increase after the addition of 5 µM spinach ferredoxin
(Sigma) and 0.25 mM NADPH (Sigma) was recorded under measuring light corresponding
to 1 µE m-2 s-1.
2.13 In vitro import in pea chloroplasts
Coding regions for PPP7A and PPP7B were amplified by PCR (using the primers
ppp7aFimp: 5’- ATGGGTAGCAAGATGTTGTT-3’, ppp7aRimp: 5’-TTAAGCTTGGC
TTCCTTCTG-3’; ppp7bFimp: 5’-ATGGCTTTTACTCTAACAATCCC-3’, ppp7bRimp:
28

Material and Methods _____________________________________________________________________________________
5’-TTAAGCTTTCCCTCCTTCTGGTA-3’) and cloned into pGEM-Teasy vector
(Promega, Madison, USA) under control of the T7 promoter. The constructs were verified
by DNA sequencing (http://www.genetik.biologie.uni-muenchen.de/sequencing). mRNA
was obtained by transcription with T7-RNA polymerase according to manufacturer’s
instructions (MBI Fermentas, St. Leon-Rot, Germany) and used for translation in wheat
germ (Wheat Germ Extract System, Promega, Madison, USA) in the presence of
[35S]methionine at 30°C for 1 h. Transcription of pSSU and pOE33 constructs (gift from
Jürgen Soll, Germany) was performed from the SP6 promoter and proteins were
synthesized in Reticulocyte Extract System (Flexi®, Promega, Madison, USA) as outlined
above. All translation mixtures were centrifuged at 50,000 g for 1 h at 4°C prior to import
experiments.
Intact chloroplasts were isolated from 10-day-old pea leaves (Pisum sativum, var. Golf) and
purified through Percoll density gradients as described (Waegemann and Soll, 1991).
Import assays were performed with chloroplasts equivalent to 20 µg of chlorophyll in 100
µl of import buffer (10 mM methionine, 10 mM cysteine, 20 mM potassium gluconate, 10
mM NaHCO3, 330 mM sorbitol, 50 mM HEPES/KOH (pH 7.6), 5 mM MgCl2) (Nada and
Soll, 2004). The amount of translation product never exceeded 10% of the total reaction
volume. The import was carried out at 25°C for 30 min. Chloroplasts were subsequently re-
purified over a 40% Percoll cushion in import buffer. If no additional treatment was
intended, the chloroplasts were washed twice (30 mM sorbitol, 50 mM HEPES (pH 7.6),
0.5 mM CaCl2) and samples for SDS-PAGE were prepared adding two volumes of SDS-
loading buffer (described in paragraph 2.9). For thermolysin treatment, chloroplasts were
washed in 330 mM sorbitol, 50 mM HEPES (pH 7.6), 0.5 mM CaCl2 and incubated with
20 µg/mL thermolysin (Calbiochem, Darmstadt, Germany) for 20 min on ice. The reaction
was stopped by addition of EDTA (pH 8) to a final concentration of 5 mM and chloroplasts
were washed once again. The treatment with trypsin (10 µg/mL) was performed for 30 min
on ice and the reaction was stopped by adding two volumes SDS sample buffer (6 M Urea,
50 mM Tris-Cl (pH 11), 100 mM dithiothreitol, 2% SDS, 10% glycerol, 0.1%
bromophenol blue). When indicated, the chloroplasts were incubated with 10% β-DM
before trypsin treatment.
29

Material and Methods _____________________________________________________________________________________
To obtain membrane fraction, chloroplasts were hypotonically lysed in 50 mM
HEPES/KOH (pH 7.6) and membranes were collected after centrifugation at 10,000 g for
10 min at 4°C. Treatment with 6 M urea was carried out in 50 mM HEPES/KOH (pH 7.6)
for 15 min at 25°C (Nada and Soll, 2004). Solubilised proteins were separated from the
insoluble fraction after centrifugation at 10,000 g for 10 min at 4°C.
To fractionate envelope, stroma and thylakoids, after import and thermolysin treatment,
chloroplasts were washed twice as before and resuspend in 20 mM Tricine (pH 7.6), 5 mM
MgCl2. The lysis was conduced on ice for 30 min and the lysate was loaded on a two-step
sucrose gradient (15% and 35% sucrose, prepared in Tricine buffer) and centrifuged for 3 h
at 134,000 g at 4°C. Stromal fraction (upper part) was precipitated with ice-cold TCA
(final concentration: 10%), washed twice with ice-cold acetone and resuspended in SDS-
PAGE loading buffer. The envelope fractions, distributed in the middle of the gradient,
were centrifuged at 280,000 g, 4°C for 30 min and resuspended in SDS-PAGE loading
buffer. The thylakoid fraction (pelletted at the bottom of the gradient) was directly
resuspended in SDS loading buffer.
Radiolabelled proteins were separated by SDS-PAGE and detected with a PhosphoImager
(FujiFilm FLA-3000).
2.14 Intracellular localization of dsRFP fusion in Arabidopsis protoplasts
Coding regions of PPP7A and PPP7B genes were amplified by PCR (using the primers
ppp7aFRFP: 5’- CACCATGGGTAGCAAGATGTTGTT-3’, ppp7aRRFP: 5’-AGCTTG
GCTTCCTTCTG-3’; ppp7bFRFP: 5’-CACCATGGCTTTTACTCTAACAA-3’, ppp7bR
RFP: 5’-AGCTTTCCCTCCTTCTGGTA-3’) and cloned upstream of the dsRED sequence
in the pGJ1425 vector by digestion with NcoI (Jach et al., 2001). Sterile cotyledons of 2-
week-old Arabidopsis plants (ecotype Col_Gl-1) were cut and incubated for 16 h at 24°C
in the dark in a protoplasting solution 1 (10 mM MES, 20 mM CaCl2, 0.5 M mannitol, pH
5.8, 0.1 g/mL macerozyme (Duchefa), 0.1 g/mL cellulase (Duchefa)). Protoplasts were
collected by centrifugation at 50 g for 10 min. Protoplasts were resuspended in 8 mL of
solution 2 (10 mM MES, 20 mM CaCl2, 0.5 M mannitol, 120 g/L sucrose, pH 5.8). 2 mL of
solution 3 (10 mM MES, 10 mM CaCl2, 10 mM MgSO4, 0.5 M mannitol, pH 5.8) were
added on top of the protoplasts and intact protoplasts were recovered at the interface
30

Material and Methods _____________________________________________________________________________________
between the two solutions after centrifugation at 70 g for 10 min (described in Dovzhenko
et al., 2003). 40 µg of plasmid DNA were introduced into protoplasts by PEG transfection
in the solution (40 % PEG solution, 70 mM Ca(NO3)2). PEG solution was prepared
dissolving 0.413 g Ca(NO3)2 • 4H2O, 1.275 g mannitol and 10 g PEG 1500 (Merck,
Germany) in 17.5 ml H2O (described in Koop et al., 1996). Microscopy analysis (with
Fluorescence Axio Imager microscope in ApoTome mode (Zeiss)) was conducted after 16
h of incubation at 23°C in the dark. Fluorescence was excited with the X-Cite Series 120
fluorescence lamp (EXFO) and images were collected in the 565-620 nm (dsRED
fluorescence) and 670-750 nm (chlorophyll auto-fluorescence) ranges.
2.15 2D Blue Native/SDS PAGE for thylakoids protein analysis
Leaves from 4-week-old plants were harvested and thylakoids were prepared as described
in paragraph 2.8. For the first dimension of Blue Native PAGE analysis, protein amounts
equivalent to 100 μg of chlorophyll for each genotype were washed with 10 mM Tris-HCl
(pH 6.8), 10 mM MgCl2 and 20 mM KCl, and subsequently solubilised in 750 mM ε-
aminocaproic acid, 50 mM Bis-Tris (pH 7.0), 5 mM EDTA (pH 7.0), 50 mM NaCl and
1.5% W/V β-DM. Solubilized samples were then incubated for 1 h on ice and afterwards
centrifuged for 10 min at 21,000 g and 4°C. Supernatants were supplemented with 5% W/V
Coomassie-blue in 750 mM ε-aminocaproic acid, and directly loaded onto BN gels (4-12%
acrylamide gels, containing 0.5 M ε-aminocaproic acid, 50 mM Bis-Tris (pH 7.0) and 10%
glycerol). One dimensional BN-PAGE was carried out at 750 V and 12 mA with cathode
buffer (50 mM Tricine, 15 mM Bis-Tris (pH 7.0) and 0,02% Coomassie G) and anode
buffer (50 mM Bis-Tris (pH 7.0)) as described by Schägger and von Jagow (1991). The gel
slices corresponding to the first dimension were treated in denaturing buffer (0.125 M Tris-
Cl (pH 6.8), 4% SDS and 1% β-mercaptoethanol) for 30 min at room temperature and 5
min at 70°C before the second dimension run. Second dimension was performed as
described in paragraph 2.8.
2.16 Yeast two hybrid and split ubiquitin assay
For yeast two hybrid assay, coding sequences of mature bait proteins (without cTP, for a
list of primers that have been used, refer to Table 2.1) were cloned into pGBKT7 carrying
31

Material and Methods _____________________________________________________________________________________
the GAL4 DNA-binding domain and the pGADT7 vector (described in Harper et al.,
1993), carrying the GAL4 activation domain, was used to express the prey proteins. YPAD
(6.0 g yeast extract, 12 g peptone, 12 g glucose, 6 g adenine hemisulphate: dissolve in 600
ml distilled water), SC (4 g yeast nitrogen base, 12 g glucose, 0.5 g synthetic complete drop
out mix: dissolve in 600 ml distilled water) and synthetic complete drop out mix (mix 2 g
adenine hemisulfate, 2 g arginine-HCl, 2 g histidine-HCl, 2 g isoleucine, 2 g leucine, 2 g
lysine-HCl, 2 g methionine, 3 g phenylalanine, 6 g homoserine, 3 g tryptophan, 2 g
tyrosine, 1.2 g uracil, 9 g valine) have been described previously by Sherman (1991). The
yeast two hybrid assay was performed as described in James et al. (1996) using the yeast
strain AH109 supplied by Clontech (Palo Alto, CA).
For split ubiquitin assay, the coding sequence of the mature PPP7A protein was cloned in
pAMBV4 and used as bait in interaction studies with prey proteins which were generated
by cloning the coding sequences of mature thylakoid proteins into pADSL (for a list of
primers, refer to Table 2.2). Interaction studies were performed using the Dual-Membrane
kit (Dualsystems Biotech AG) according to manufacturer’s instruction and as described by
Pasch et al. (2005).
Table 2.1 List of primers (5’ to 3’ orientation) used for cloning pray and bait sequences for yeast
two hybrid assay.
32
Material and Methods _____________________________________________________________________________________
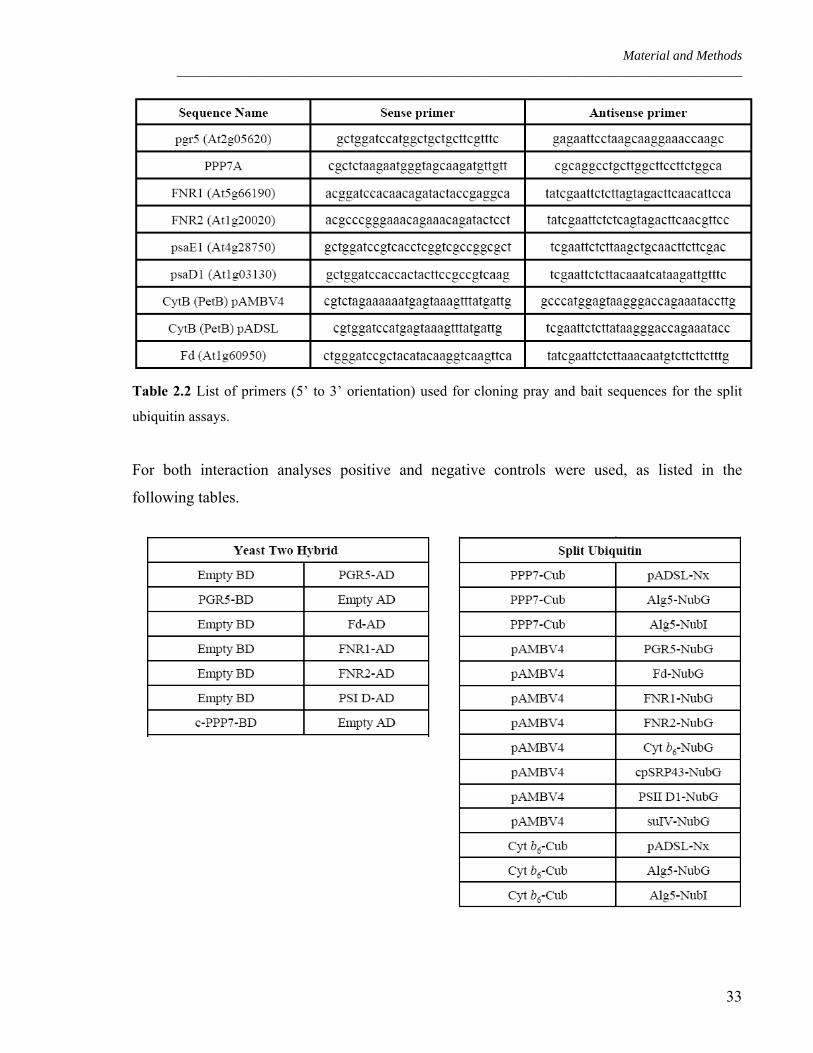
Table 2.2 List of primers (5’ to 3’ orientation) used for cloning pray and bait sequences for the split
ubiquitin assays.
For both interaction analyses positive and negative controls were used, as listed in the
following tables.
33

Results ____________________________________________________________________________________________
3. Results
3.1 Gene structure and homologies
Previous work identified groups of transcriptionally co-regulated nuclear genes in Arabidopsis
thaliana, which were enriched for photosynthetic genes (Biehl et al., 2005). These
photosynthetic regulons contain genes of yet unknown function, the products of which
represent putative photosynthetic proteins (PPPs). One of them, PPP7, is the subject of this
thesis.
PPP7 is encoded by the two highly homologous genes At4g22890 (PPP7A) and At4g11960
(PPP7B), that are located on a duplicated region of chromosome 4 of A. thaliana. The PPP7
proteins do not share any common motive with proteins of known function, but are conserved
among different species. Orthologous genes can be found in flowering plants both in
monocotyledons like Oryza sativa and Zea mays, and in dicotyledons for instance Populus
trichocarpa and Lycopersicon esculentum (Figure 3.2). Also the eukaryote algae
Chlamydomonas reinhardtii and the moss Physcomitrella patens possess an orthologue of
PPP7.
When analyzing the publicly available microarray data of A. thaliana (https://www.geneve-
stigator.ethz.ch/) both genes, PPP7A and PPP7B, showed an increased expression in green
tissues. This could be confirmed by Real-Time PCR on cDNA extracted from leaves and roots
of wild type (WT) plants (Figure 3.1). Additionally, it can be observed that the expression
level of PPP7A is considerably higher than the expression level of PPP7B.
Leaves Roots
PPP7BPPP7A
Fig 3.1. Real-Time PCR analysis on cDNA from leaves and roots of WT plants using specific primers on At4g22890 and At4g11960. The expression levels are shown as arbitrary units, normalized on the expression levels of ACTIN 1 which was used as a reference.
34

Results ____________________________________________________________________________________________
PPP7A MGSKMLFSLTSPRLFSAVSR--KPSSSFSPSPPSPSSRTQWTQ-LSPGKSISLRRR 53 PPP7B ----MAFTLTIPR-FSAISR--KPITCSSSRTQCPAP-------FTHGRSISLRRR 42 Os08g0526300 ----MASLLPSPVFPTAAAAAASTSSSSRCRCRCRITTSSSAHGWSNCQGWRLHHR 52 Os03g0857400 ------MLMASEAWLSPASRLFHPPRSSTAAFFSASGLPRLPRRSSCALATSTRGR 50 P. trichocarpa gw1 MAGKLAFTLTSPRVLIAPIQ--KPFISSSSSLPPPSC-SSSTRVHFNVKQFSLRRR 53 P. trichocarpa gr3 MASKLAFNLTSPRVFTAPIQ--KPIISSSSSLPSLSSPSCSTRVQLNGKQFSLRGR 54 C. reinhardtii ----MQTTVLRSSTRVGCVR---PVGLARALPVRP-------------VVFGSRTP 36 PPP7A VFLLPAKAT----TEQS-GPVG-GDNVDSNVLPYCSINKA----EKKTIGEMEQEFLQALQSFYY 108 PPP7B LTLLPLKAS----TDQS-GQVG-GEEVDSKILPYCSINKN----EKRTIGEMEQEFLQAMQSFYY 97 Os08g0526300 VWAAQAADQQGGVQQQQQQEENEDGVVDSNVLPYCSINRK----EKKTIGEMEQEFLQALQAFYY 113 Os03g0857400 RLRLGATDQ-----QQQQGKQDGDEVVDSNVLQYCSIDGKGKKAEKRSLGEMEQEFLQALQAFYY 110 P. trichocarpa gw1 MLLPPTKAT----ADQQAGLX--DEMVDGKILQYCSIDKR----GKKSIGEMEQEFLQALQAFYY 108 P. trichocarpa gr3 MLFLPTKAT----ADQQTDQVQEDDMDDGKILQYCSIDKK----GKKSLGEMEQDFLQALQAFYY 111 C. reinhardtii MLLRSAKKD--------DGYISEDEGLGNVAADYCAIDGAG-KKAKRSLGEMEQEFLAAMTSWYY 92 * ** * ** * ** * *** * PPP7A DGKAIMSNEEFDNLKEELMWEGSSVVMLSSDEQRFLEASMAYVSGNPILNDEEYDKLKLKLKIDG 173 PPP7B EGKAIMSNEEFDNLKEELMWEGSSVVMLSSDEQRFLEASMAYVSGNPILSDEEYDKLKMKLKMDG 162 Os08g0526300 DKKAVMSENEFDNLKEELMWEGSSVVMLSPDEQRLLEASMAYVAGNPIMTDAEFDELKLRLRKEG 178 Os03g0857400 DQKAIMSNEEFDNLKEELMWEGSSVVMLSPDEQRLLEASMAYAAGNPIMSDAEFDQLKLRLKKDG 175 P. trichocarpa gw1 EGKAIMSNEEFDNLKEELMWEGSSVVMLSSDEQKFLEASMAYVSENPIMSDKEYDELKMKLKSEG 173 P. trichocarpa gr3 EGKAIMSNEEFDNLKEELMWQGSSVVMLSSDEQKFLEASLAYVSGNPIMNDEEFDKLKIKLKTEG 176 C. reinhardtii EGKPTMSDEEFSLLKEELIWSGSMVAVLSSDEQRFLEASMAYAKGKPIMTDEDYDALKAELRNKS 157 ┌───── TM1 ──────┐ PPP7A SDIVSEGPRCSLRSKKVYSDLAVDYFKMLLLNVPATVVALGLFFFLDDITGFEITYIMELPEPYS 238 PPP7B SEIVCEGPRCSLRSKKVYSDLAIDYFKMFLLNVPATVVALGLFFFLDDITGFEITYLLELPEPFS 227 Os08g0526300 SEIVQEGPRCSLRSRKVYSDLTVDYFKMFLLNVPAAVLALTLFFFLDDLTGFEITYLLELPEPFS 243 Os03g0857400 SDIVTEGPRCSLRSRKVYSDLTVDYLKMFLLNVPATTLALGLFFFIDELTGFEIN-IFQLPEPFG 239 P. trichocarpa gw1 SEIVVEGPRCSLRSRKVYSDLYVDYLKMFLLNVPATVTALGLFFFLDDLTGFEITYLLELPEPFS 238 P. trichocarpa gr3 SEIVVEGPRCSLRSRKVYSDLSVDYLKMFLLNVPATVVALGLFFFLDDLTGFEITYLLELPEPFS 241 C. reinhardtii SIVTAQGPRCSIRSKKMYADAEPDYLRMTALNLPGVLFVLGLVFAVDYSTGFGVTKLVELPAPYG 222 └─────────────────┘ ┌──────── TM2 ────────┐ PPP7A FIFTWFAAVPVIVYLALSITKLIIKDFLILKGPCPNCGTENTSFFGTILSISSGGKTNTVKCTNC 303 PPP7B FIFTWFAAVPAIVYLALSLTKLILKDFLILKGPCPNCGTENVSFFGTILSIPNDSNTNNVKCSGC 292 Os08g0526300 FIFTWFAALPLIFWVAQAITSAIVKDFLILKGPCPNCGNENLSFFGTILSVPSGGARNSVKCANC 308 Os03g0857400 FIFTWFAALPLILFIAQSITNAIVNDFLILKGPCPNCGAENLSFFGTILSVSSGGETNKVKCADC 304 P. trichocarpa gw1 FLFTWFAAVPLIVWLALTLTNAIVKDFLILKGPCPNCGTENGSFFGTILSISSGGTTNTLKCSNC 303 P. trichocarpa gr3 FLFTWFAAVPLIVWLALTLTNAIVKDFLILKGPCPNCGTENGSFFGTILSISSGGASNTLKCSNC 306 C. reinhardtii PILLWGLLLPSLFTVAYALTQVGFKDNLILKAPCPSCGSENFSYFGDVFTVAGARGQNLVECPNC 287 └──────────────────────┘ PPP7A GTAMVYDSGSRLITLPEGSQA------------------------- 324 PPP7B GTEMVYDSGSRLITLPEGGKA------------------------- 313 Os08g0526300 SSSLVYDSASRLITLPETAEA------------------------- 329 Os03g0857400 STEMVYDSKSRLITLPEPSQ-------------------------- 324 P. trichocarpa gw1 STELVYNSKTRLITLPEGSEA------------------------- 324 P. trichocarpa gr3 STELVYDSKTRLITLPEGSEA------------------------- 327 C. reinhardtii KADMIFDEYKRVVVVAETSEVKQEKLAAAAAKKAAAAAKKKAKAAA 333
Fig 3.2. Sequence alignment of the PPP7 proteins in A. thaliana and other plant and algal species. Chloroplast transit peptides are indicated in italics, predicted transmembrane domains are boxed. A putative negatively charged region is indicated by asterisks. The alignment has been created with the program Clustal W (http://www.ebi.ac.uk/clustalw/) and sequences have been obtained from the genome sequences available at http://genome.jgi-psf.org/ and http://www.ncbi.nlm.nih.gov/.
35

Results ____________________________________________________________________________________________
3.2 PPP7A and PPP7B are targeted to the thylakoid membrane
Already a few years ago, Peltier et al. (2002) identified PPP7A as a component of the
chloroplast proteome by using mass spectrometry. In 2004, Kleffmann et al. confirmed the
chloroplast localization of PPP7A, while Friso et al. (2004) found PPP7A to be a thylakoid
transmembrane protein. To analyze the localization of PPP7 proteins, three different
techniques have been applied.
A first in silico approach clearly showed that the gene products of both PPP7A and PPP7B are
predicted to be targeted to the chloroplasts by at least five of seven publicly available predictor
algorithms (Table 3.1).
ChloroP TargetP Predotar Mitoprot iPSORT PCLR PSORT
PPP7A C (0.598) C (0.956) C (0.770) M (0.985) M C (0.999) C (0.888)
PPP7B C (0.557) C (0.904) C (0.670) M (0.993) C C (0.874 M (0.845)
Table 3.1. In silico prediction of subcellular targeting of PPP7A and PPP7B. In parentheses the highest output scores for each program are reported. C: chloroplast; M: mitochondrion.
The second approach was the in vivo localization of PPP7A and PPP7B in Arabidopsis
protoplasts: the full-length coding sequences of both genes have been fused 5’ to the sequence
coding for the red fluorescence protein of the coral Discosoma (dsRFP; described in Jach et al.,
2001). The obtained constructs were used to transiently transfect Arabidopsis thaliana (ecotype
C24) protoplasts. The signal of both RFP-fusion proteins is clearly associated with the
chloroplasts and overlaps with the signal of chlorophyll auto-fluorescence (Figure 3.3).
PPP7A
PPP7B
DIC Auto dsREDFig 3.3. Subcellular localization of the PPP7 proteins. The PPP7A and PPP7B-dsRED fusions were transiently expressed in Arabidopsis protoplasts and protein localization was analyzed using fluorescence microscopy. “DIC”: differ-ential interference contrast image; “Auto”: chloroplasts revealed by chlorophyll auto-fluorescence; “dsRED”: dsRED protein-fluorescence. Scale bars = 50 µm.
36
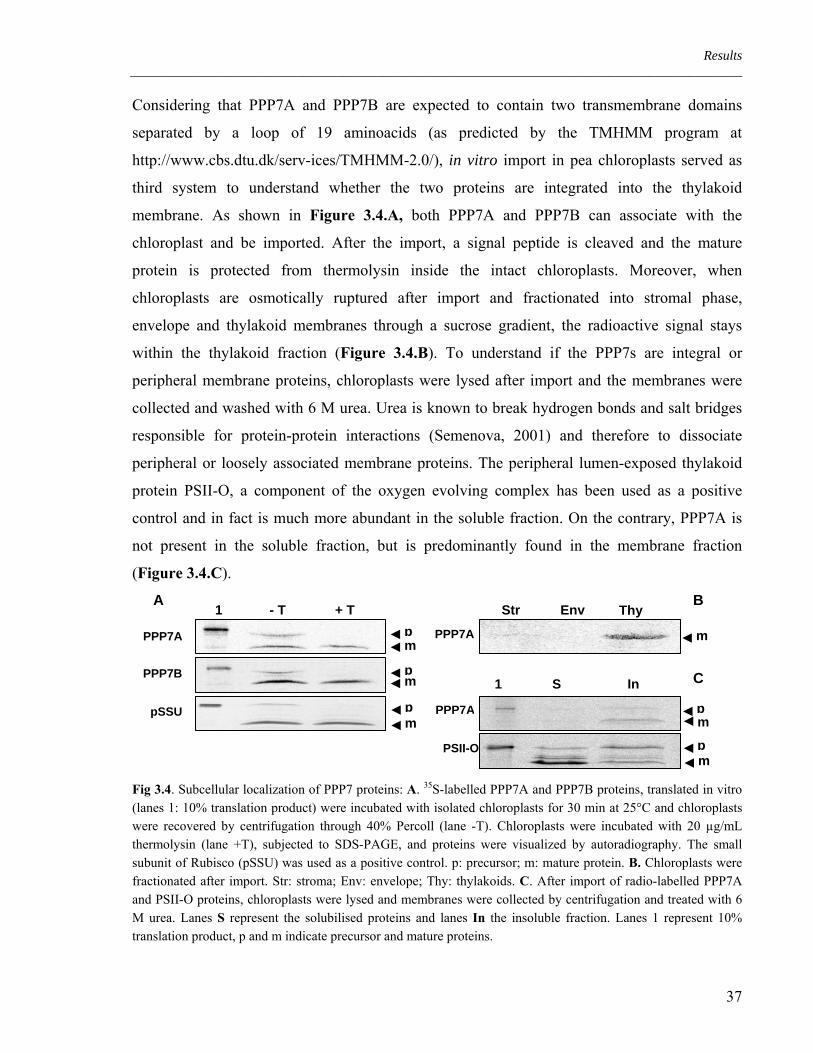
Results ____________________________________________________________________________________________
Considering that PPP7A and PPP7B are expected to contain two transmembrane domains
separated by a loop of 19 aminoacids (as predicted by the TMHMM program at
http://www.cbs.dtu.dk/serv-ices/TMHMM-2.0/), in vitro import in pea chloroplasts served as
third system to understand whether the two proteins are integrated into the thylakoid
membrane. As shown in Figure 3.4.A, both PPP7A and PPP7B can associate with the
chloroplast and be imported. After the import, a signal peptide is cleaved and the mature
protein is protected from thermolysin inside the intact chloroplasts. Moreover, when
chloroplasts are osmotically ruptured after import and fractionated into stromal phase,
envelope and thylakoid membranes through a sucrose gradient, the radioactive signal stays
within the thylakoid fraction (Figure 3.4.B). To understand if the PPP7s are integral or
peripheral membrane proteins, chloroplasts were lysed after import and the membranes were
collected and washed with 6 M urea. Urea is known to break hydrogen bonds and salt bridges
responsible for protein-protein interactions (Semenova, 2001) and therefore to dissociate
peripheral or loosely associated membrane proteins. The peripheral lumen-exposed thylakoid
protein PSII-O, a component of the oxygen evolving complex has been used as a positive
control and in fact is much more abundant in the soluble fraction. On the contrary, PPP7A is
not present in the soluble fraction, but is predominantly found in the membrane fraction
(Figure 3.4.C).
PPP7A
PPP7B p
p
m
m
◄◄
◄◄
pm
◄◄
pSSU
1 - T + T Str Env Thy
PPP7A
C
PPP7A
PSII-O
1 S In
pm
◄◄
pm
◄◄
m◄
A B
Fig 3.4. Subcellular localization of PPP7 proteins: A. 35S-labelled PPP7A and PPP7B proteins, translated in vitro (lanes 1: 10% translation product) were incubated with isolated chloroplasts for 30 min at 25°C and chloroplasts were recovered by centrifugation through 40% Percoll (lane -T). Chloroplasts were incubated with 20 µg/mL thermolysin (lane +T), subjected to SDS-PAGE, and proteins were visualized by autoradiography. The small subunit of Rubisco (pSSU) was used as a positive control. p: precursor; m: mature protein. B. Chloroplasts were fractionated after import. Str: stroma; Env: envelope; Thy: thylakoids. C. After import of radio-labelled PPP7A and PSII-O proteins, chloroplasts were lysed and membranes were collected by centrifugation and treated with 6 M urea. Lanes S represent the solubilised proteins and lanes In the insoluble fraction. Lanes 1 represent 10% translation product, p and m indicate precursor and mature proteins.
37

Results ____________________________________________________________________________________________
Further evidence for the thylakoid localization of the PPP7s was provided by western analysis
using antibodies against PPP7 on WT and ppp7ab chloroplasts, thylakoid and stromal fractions
(Figure 3.5).
chl thy str chl thy str
WT ppp7ab
PPP7
Fig 3.5. Subcellular localization of PPP7 proteins: chl lanes correspond to 5µg chlorophyll of intact chloroplasts; thy lanes correspond to the thylakoid fraction coming from the osmotic lysis of 5µg chlorophyll of chloroplasts and str lanes correspond to the stromal fraction of the same lysis reaction. In the lower panel, the Coomassie staining of the portion of the gel, where the large subunit of the Rubisco runs, is presented to confirm the purity of the fractions.
3.3 Topological orientation of PPP7 in the thylakoid membrane
According to predictions (see Figure 3.2), PPP7 proteins probably fold with two
transmembrane helices separated by a loop of 19 aminoacids. Both N- and C- terminal
domains of the protein represent hydrophilic regions of 141 and 63 aminoacids, respectively.
As a result, two topological orientations of PPP7 can be assumed:
thylakoid lumen
stroma
Topology 1
NH2 COOH
Topology 2
NH2 COOH
To understand the real topology of PPP7, mild trypsin digestion was applied to intact
thylakoids. Trypsin hydrolyzes the peptidic bonds at the carboxylic groups of aminoacids
arginine and lysine and is not permeable to membranes, for that it is digesting the part of the
protein exposed to the stromal side of the thylakoids. Both the C-terminus and the N-terminus
of the protein contain multiple trypsin cleavage sites (PeptideCutter,
http://expasy.ch/tools/peptidecutter/), while the loop between the two does not carry any
sequences predicted to be cleaved by trypsin. If the native PPP7 conformation is the topology
38

Results ____________________________________________________________________________________________
model 1, the N- and C- terminal domains are exposed to the stroma and they will therefore be
digested by trypsin. In this case, a fragment of 6.7 kDa will remain protected, comprising the
loop and the two transmembrane domains. If the protein is folded according to topology model
2, it would remain insensitive to the trypsin treatment, since the loop between the two
membrane helices does not contain a single trypsin cleavage site.
The quality of the preparations was monitored by digestion pattern of the intermediate and
mature form of the PSII-O subunit of the lumenal Oxygen Evolving Complex.
TL TL Th Tr Th Tr TL TL Th Tr Th Tr
32.5
25
16.5
6.5*
m m
p p i ► ► ►
PSII-O PPP7A
32.5
25
16.5
6.5
m m
p p i ►
► ►
PSII-O PPP7A
Fig 3.6. Left: Trypsin treatment of the thylakoid membrane with incorporated 35S-PSII-O and 35S-PPP7A. Pea chloroplasts corresponding to 20 µg of chlorophyll were incubated with 35S-labeled pPSII-O and pPPP7A for 30 min. Intact chloroplasts were re-purified after import and treated with thermolysin to remove the precursor proteins (lane Th). Thylakoids were isolated and trypsin was added to 10 µg/mL (lane Tr). Right: Trypsin treatment of thylakoids after solubilisation with β-DM. Thermolysin treatment of chloroplasts after import (lane Th). Thylakoids were incubated with β-DM prior to trypsin treatment in order to release protein complexes from the membrane (lane Tr). Lanes TL represent 1/10 of the translation product. On the right side of both panels, the corresponding protein MWs are indicated in KDa.
Concerning the PSII-O control (Figure 3.6) the stromal intermediate protein, present in intact
chloroplasts (Th lane), completely disappeared in the trypsin-treated thylakoids whereas the
mature form of PSII-O remains intact and no partial proteolysis can be detected (Tr lane).
Quite the opposite is the case for PPP7A. Its mature form is digested into three low-molecular-
weight products (*), the smallest of them being approximately 7 kDa in size while the other
two are ranging from 9 to 14 kDa. The appearance of a 7 kDa digestion product is in
agreement with the topology 1 folding. The appearance of the two additional fragments of
larger size may indicate higher resistance of certain PPP7 domains from trypsin digestion,
possibly implying tight interactions of PPP7 with other protein constituents on the stromal side
of the thylakoid membrane.
39

Results ____________________________________________________________________________________________
As an additional control, isolated thylakoids were treated with 10% n-dodecyl- β-D-maltoside
(β-DM) prior to trypsin treatment in order to extract protein complexes from the thylakoid
membrane. Both imported PSII-O and PPP7A were entirely degraded under those conditions
(Figure 3.6), confirming that the observed protease resistance was due to the membrane
barrier.
Eventually, the topology of the native PPP7s was studied by means of immunoblotting
analysis. Anti-PPP7 antibodies were raised against the first 140 N-terminal amino acids of the
mature PPP7A protein and recognize both PPP7A and PPP7B (Figure 3.9). If the conclusion
that the N-terminal region is exposed to the stromal side was correct, PPP7 should be no longer
detectable by anti-PPP7 antibody after trypsin treatment of thylakoid membrane. Indeed, the
PPP7 signal was barely visible after trypsin treatment (Figure 3.7).
PSII-O PPP7A
Thy Thy Tr Tr Fig 3.7. Immunoblotting of the thylakoid membrane with α PsbO and α PPP7 antisera before (lane Thy) and after (lane Tr) trypsin treatment.
3.4 ppp7a and ppp7b mutations
The gene structures of PPP7A and PPP7B are depicted in Figure 3.8.A. The predicted intron-
exon organization was confirmed by RT-PCR for both genes and the sequences obtained were
compared with the coding sequences available on public websites (MIPS:
http://mips.gsf.de/proj/plant/jsf/athal/searchjsp/index.jsp and TAIR: http://www.arabid-
opsis.org/). T-DNA insertion mutants for PPP7A and PPP7B were identified by browsing a T-
DNA insertion flanking database (http://signal.salk.edu/cgi-bin/tdnaexpress). For each of the
two genes, a single mutant line could be identified, the SAIL line Sail_443E10 mutated in
PPP7A and the Salk line Salk_059233 for PPP7B. For both lines the insertion site has been
determined using gene- and T-DNA-specific primers, followed by sequencing of the PCR
products. To confirm that both lines were knock-outs and thus, that the T-DNA insertions
effectively interrupted gene transcription, total RNA was isolated from WT and mutant leaves
and Reverse Transcriptase PCR (RT-PCR) was performed using gene-specific primers. Neither
ppp7a nor ppp7b showed residual transcription of the respective interrupted gene (Figure
3.8.B). Due to the high homology between PPP7A and PPP7B and to analyze their likely
40

Results ____________________________________________________________________________________________
redundant function, the double mutant ppp7ab was created crossing homozygous ppp7a plants
with ppp7b plants. The F2 generation was then screened by PCR for homozygous double
mutants.
B
Fig 3.8. Mutations of PPP7A and PPP7B genes and their effects on transcript accumulation. A: The translated parts of exons (boxes), as well as intron sequences (lines) are depicted. The T-DNA insertion sites and orientation are provided and drawn not in scale. B: Transcript levels were detected by RT-PCR. Amplicons were selected 5’ of the insertion for ppp7a and 3’ for ppp7b. PCR was carried out for 40 cycles using gene- and Actin1-specific primer (the latter as a control).
To investigate the expression level of PPP7A and PPP7B, antibodies were raised against PPP7
proteins (see Material and Method). The protein level of PPP7A and PPP7B was different: in
ppp7a mutant backgrounds PPP7B is only weakly expressed compared with the homologue
PPP7A, detected in ppp7b single mutant. In the double mutant ppp7ab, PPP7 proteins are
completely missing (Figure 3.9).
Fig 3.9. Western analysis of PPP7 content in thylakoid membrane of WT and mutant plants. Lanes were loaded with 5 µg of chlorophyll except lane 0.5xWT and probed with αPPP7 antibodies. Coomassie staining of the same gel serves as a loading control
Both, single and double mutants could germinate and grow photoautotrophycally on soil
(Figure 3.10). Only ppp7ab double mutant shows a relatively pale green leaf coloration and
reduced biomass under controlled greenhouse growth conditions. Chlorophyll content analysis
(Table 3.2) showed a reduced amount of total chlorophyll while the chl a/b ratio was
unchanged.
41

Results ____________________________________________________________________________________________
ppp7ab
WT ppp7b
ppp7a
Fig 3.10. Growth phenotype of 3-week-old WT, ppp7a and ppp7b single mutants as well as the ppp7ab double mutant. Plants were grown in greenhouse under long-day conditions.
Sample Pigments in pmol per mg fresh weight
VAZ Nx Lut Car Chl a Chl b Chl a+b Chl a/b
WT 59 ± 5 83 ± 6 288 ± 24 183 ± 8 1722 ± 94 521 ± 38 2243 ± 132 3.31 ± 0.08
ppp7ab 46 ± 5 65 ± 10 217 ± 37 141 ± 18 1300 ± 187 399 ± 56 1699 ± 243 3.26 ± 0.08
Table 3.2. Pigment composition of WT and ppp7ab double mutant leaves. Pigment content was determined by HPLC of leaves from four different plants each genotype. Mean values ±SD are shown. VAZ: xanthophylls cycle pigments (violaxanthin + antheraxantin + zeaxanthin); Nx: Neoxanthin; Lu: Lutein; Car: β-carotene; Chl: chlorophyll.
3.5 Complementation of the mutations
Since for each gene only one line could be identified in the available mutant collections, it has
been necessary to complement the phenotype of the double mutant ppp7ab with both genes
PPP7A and PPP7B. To achieve this, the entire coding sequence of both genes has been cloned
under the control of the 35SCaMV promoter and the constructs were used to transform ppp7ab
double mutant plants. Positive transformants were selected by hygromycin resistance and the
insertion was confirmed by PCR with specific primers (refer to Material and Methods for a
list). The presence of PPP7 proteins was eventually detected by Western analysis (Figure
3.11). The complementation of the phenotype was also analyzed in terms of photosynthetic
42

Results ____________________________________________________________________________________________
43
performance confirming that under the control of the 35SCaMV promoter both genes can
complement the mutation and indeed have redundant functions.
WT
PPP7
PGR5
35S:: PPP7A
ppp7ab
35S:: PPP7B
Fig 3.11. Complementation of ppp7ab with both PPP7A and PPP7B. All lanes are loaded with thylakoid extraction corresponding to 5 µg of chlorophyll.
3.6 The mutations do not cause any loss in the content of thylakoid proteins
PPP7 proteins are integrated into the thylakoid membrane and their loss could have deleterious
effects on the organization of the photosynthetic machinery. To test this possibility, native one-
and two-dimensional gels were performed using β-DM to solubilise thylakoid proteins of WT
and ppp7ab double mutant plants. β-DM is a mild detergent known to preserve the supra-
structure of complexes by isolating them from the lipid-films. One dimensional blue native
gels showed that there are no differences between WT and double mutant regarding the
organization of the photosynthetic machinery in super-complexes (Figure 3.12).
ppp7ab
WT
B A
47 kDa 34 kDa
26 kDa 20 kDa
47 kDa 34 kDa
26 kDa 20 kDa
Fig 3.12. Separation of complexes and super-complexes of the photosynthetic machinery. A. One dimension BN-PAGE using 100 µg of chlorophyll each lane. B. Lanes of the first dimension BN-PAGE gel were then loaded on an SDS-PAGE acrylamide gel for the second dimension run and stained with non-quantitative silver staining.

Results ____________________________________________________________________________________________
3.7 Mutants lacking PPP7s are impaired in photosynthesis
Photosynthetic electron transport was characterized in detail by comparing the light intensity
dependence of chlorophyll fluorescence parameters between mutant and WT plants using a
Pulse Amplitude Modulated chlorophyll fluorometer (PAM). Both single mutants behaved
indistinguishable from WT, still expressing the PPP7 protein to varying degrees because of
residual expression of PPP7 transcript from the non-mutated locus (Figure 3.9). In the ppp7ab
double mutant the maximum quantum yield (FV/FM) is similar to WT (Table 3.3). The fraction
of the primary electron receptor of PSII (QA) present in the reduced state, estimated by the
parameter 1-qP, is increased in the double mutant plants. Moreover, the effective quantum
yield of PSII (ФII) is reduced. Taken together, these findings suggest an unaltered electron flow
through PSII, while a downstream step of the electron transfer could be affected.
In addition, under the control of the 35SCaMV promoter, both genes can restore the WT
photosynthetic phenotype in the double mutant background, confirming the redundancy of
their function.
Parameter ppp7a ppp7b ppp7ab WT 35S::PPP7A 35S::PPP7B
FV/FM 0.83 ± 0.01 0.83 ± 0.01 0.83 ± 0.01 0.83 ± 0.01 0.83 ± 0.02 0.83 ± 0.01
ФII 0.76 ± 0.02 0.74 ± 0.02 0.66 ± 0.02 0.75 ± 0.01 0.74 ± 0.02 0.76 ± 0.02
1-qP 0.05 ± 0.02 0.07 ± 0.02 0.13 ± 0.02 0.06 ± 0.01 0.08 ± 0.02 0.07 ± 0.03
Table 3.3. Spectroscopic data for mutant and WT leaves. Mean values ± SD (5 plants each genotype) are shown. For the double mutants transformed with the 35S::PPP7 constructs, the data of only one complemented line of each gene are listed.
Supporting previous results, the electron transport rate (ETR) was not affected at low light
intensities in ppp7ab mutants (Figure 3.13) but remains lower than in WT at higher light
intensities. Non photochemical quenching was induced with increasing light intensities but this
induction was impaired in comparison to WT (Figure 3.13). Both of these findings resemble
the behaviour of the pgr5 mutant with a defective CEF around PSI (Munekage et al., 2002).
For this reason, pgr5 has been included as an additive control in the following measurements.
44

Results ____________________________________________________________________________________________
Fig 3.13. Light intensity dependence of the electron transport rate (ETR) and of non-photochemical quenching. ETR is depicted relative to the maximal value in WT; NPQ is calculated from chlorophyll fluorescence in WT and mutant leaves. White circles: WT plants; black circles: ppp7ab; white squares: pgr5. Each value represents the mean ± standard deviation (5 plants each genotype).
NPQ is measured mainly as thermal dissipation of excitation energy from PSII (qE) triggered
by luminal acidification. When dark adapted leaves are exposed to light, immediate
acidification of the lumen drives a rapid and transient increase of NPQ within the first minute.
After further two minutes a relaxation of NPQ can be observed. This relaxation is due to the
fact that the intensity of 80 µE m-2 s-1 is not high enough to induce thermal dissipation during
steady-state photosynthesis. As it was already described for the pgr5 mutant (Munekage et al.,
2002), also in ppp7ab this transient increase in NPQ is reduced when compared with WT
(Figure 3.14). This indicates that in both pgr5 and ppp7ab the thylakoid lumen was not
acidified enough to induce transient thermal dissipation.
Fig 3.14. Time course of induction and relaxation of NPQ. Induction and relaxation of NPQ were monitored for 4 min at 80 µE m-2s-1 (white bar) and 2 min in the dark (black bar) on dark adapted leaves of WT plants (white circles), ppp7ab (black circles) and pgr5 mutants (white squares). Each value represent the mean ± standard deviation (5 plants each genotype).
45

Results ____________________________________________________________________________________________
Considering that pgr5 and ppp7ab mutants could be impaired in the same pathway, the absence
of the PPP7 protein should have an effect on the oxidation ratio of the PSI reaction centre, as
well as on the redox state of the plastoquinone, similar to pgr5.
Fig 3.15. Light intensity dependence of the P700 oxidation ratio (ΔA/ ΔAMAX) in dark adapted leaves of WT plants (white circles), ppp7ab (black circles) and pgr5 mutants (white squares). Each value represents the mean ± standard deviation (5 plants each genotype).
In order to monitor the redox state of P700, the changes in the difference of absorbance at 810
and 860 nm have been evaluated. P700 is oxidised to P700+ under different light intensities. The
level of oxidation is measured as ΔA. The oxidation is followed by a reduction in the dark and
re-oxidation, to a maximum level, under far-red light (measured as ΔAMAX). The light
dependence of the P700 oxidation ratio can then be calculated as ΔA/ ΔAMAX. In the ppp7ab
double mutant (as in pgr5) the P700 stays in a reduced state already at relatively low light
intensities (Figure 3.15).
3.8 Mutants lacking PPP7 are impaired in cyclic electron flow around PSI
It has been shown that in pgr5 mutant the ferredoxin-dependent plastoquinone reduction is
decreased. To examine this phenotype also in ppp7ab, plastoquinone reduction was monitored
in vitro as an increase in chlorophyll fluorescence. Under illumination of ruptured chloroplasts
with light of low intensity (1 µE m-2 s-1) the detectable chlorophyll fluorescence reflects the
reduction of plastoquinone by cyclic electron transport from ferredoxin. In this assay, NADPH
is used as an electron source via the reverse reaction of ferredoxin-NADPH reductase enzyme
(Myake and Asada, 1994; Munekage et al., 2002). Also mutants defective in linear electron
flow were tested to determine the CEF level when linear flow is slightly impaired. This was
done to clarify whether the presumable defects in CEF in ppp7ab and pgr5 are just secondary
effects due to an altered linear electron flow. As shown in Figure 3.16, in ppp7ab as well as in
the pgr5 mutant, the ferredoxin-dependent plastoquinone reduction is impaired indicating that
46

Results ____________________________________________________________________________________________
PPP7 and PGR5 operate in the same pathway associated with Fd-dependent CEF. Interestingly,
under the tested conditions, linear electron flow defective Arabidopsis mutants psad1-1
(Ihnatowicz et al., 2004) and psae1-3 (Ihnatowicz et al., 2007) show a slightly higher reduction
of the plastoquinone pool, which is already induced by the addition of the NADPH (in WT,
pgr5 and ppp7ab, NADPH addition do not cause any significant increase in chlorophyll
fluorescence). These findings clearly suggest that impaired linear electron flow does not
necessarily result in a defective cyclic electron transfer.
NADPH Fd 30 sec
WT psae1-3
pgr5
psad1-1
ppp7ab
Chl
Flu
ores
cenc
e
Fig 3.16. Increase of chlorophyll fluorescence by addition of NADPH (0.25 mM) and ferredoxin (Fd, 5 µM) under weak measuring light (1 µE m-2 s-1) was monitored in osmotically ruptured chloroplasts (20 µg chlorophyll/mL) of WT, psae1-3, psad1-1, pgr5 and ppp7ab mutants. To measure the occurrence of CEF around PSI in intact leaves, another method was applied as
already described (Joliot et al., 2004; Joliot and Joliot, 2002; Nandha et al., 2007). During
these measurements the P700 oxidation kinetics is tracked under illumination with far-red light
in a way that only PSI photochemistry is active. The slow P700 oxidation observed in dark-
adapted leaves is supposed to represent the occurrence of CEF, whereas the fast P700 oxidation
observed in light-adapted leaves should reflect the rate of LEF because of the activated Calvin
cycle (Joliot and Joliot, 2005).
Without pre-illumination, ppp7ab, pgr5 and WT plants exhibit similar P700 oxidation kinetics
(Figure 3.17). In leaves that were first illuminated for 5 min with green light to activate linear
electron flow, oxidation of P700 occurred as rapidly in ppp7ab plants as in the pgr5 mutant and
WT plants. These results imply that, under those conditions, LEF replaces CEF with equal
efficiency in all three genotypes. However, after shorter periods of priming with green light,
both ppp7ab and pgr5 showed faster P700 oxidation than WT, clearly suggesting that the CEF-
to-LEF transition occurs more rapidly in the two mutants. In the same assay, P700 oxidation is
47

Results ____________________________________________________________________________________________
suppressed in dark-adapted leaves of psad1-1 mutants (Figure 3.17), indicating that they show
increased rates of CEF. This is in agreement with the results obtained from the in vitro assay as
depicted in Figure 3.16.
Fig 3.17. Quantification of CEF in intact leaves. P700 oxidation was induced by illumination with far-red light. Dark-adapted leaves from WT (open circles in the left and right panel), ppp7ab (filled circles, left panel), pgr5 (squares, left panel) and psad1-1 (filled diamonds, right panel) plants were illuminated for different time intervals with green light to induce LEF (black color: 0 s and red color: 20 s), and P700 oxidation was recorded 200 ms after the green light was switched off. The P700 oxidation after 5-min exposure to green light, followed by 2 min in the dark is indicated by green curves. The data are presented as relative values.
In conclusion the applied in vitro and in vivo assays indicate that CEF is disturbed to similar
extents in both ppp7ab and pgr5 mutants. However, the relative severity of the measured CEF
defects differs between the two assays: the intact-leaf assay detects an imbalanced CEF-to-LEF
transition and a slight reduction in the maximum rates of CEF while the in vitro assay suggests
that the CEF capacity is markedly reduced.
Considering that pgr5 and ppp7ab mutants show a defective CEF which results in a reduced
capacity to induce NPQ during the dark-to-light transition, it could be concluded that increased
CEF can result in a higher reduction of the plastoquinone pool.
Ultimately this stronger reduction of the plastoquinone pool could lead to an increase in the
induction of transient NPQ. Indeed, this could be confirmed for the psad1-1 and psae1-3
mutants (Figure 3.18).
48
Results ____________________________________________________________________________________________
Fig 3.18. Time course of induction and relaxation of NPQ. Induction and relaxation of NPQ were monitored for 4 min at 80 µE m-2s-1 (white bar) and 2 min in the dark (black bar) on dark adapted leaves of WT plants (white circles), psad1-1 (dark diamonds) and psae1-3 mutants (white diamonds). Each value represents the mean ± standard deviation (5 plants each genotype).
3.9 PPP7 interacts in vitro with PGR5, Fd, FNR, PSI and cyt b6/f
To identify possible interaction partners of PPP7, proteins that were shown to be involved in
CEF around PSI or to mediate linear electron transfer through the stromal sides of PSI and the
cyt b6/f complexes were taken into consideration.
Since PPP7A is integrated in the thylakoid membrane, the Split Ubiquitin Assay was chosen,
which has been comprehensively described in Pasch et al. (2005). For the interaction study, the
coding sequence of mature PPP7A has been cloned in the pAMBV4 vector fused to the C
terminus of the Ubiquitin gene (Cub), whereas the to be tested interactors (Fd, PSI-D, Cyt b6,
and the two leaf isoforms of FNR, FNR1 and FNR2) were cloned into the pADSL vector fused
to the sequence coding for the N terminus of the Ubiquitin protein (Nub).
The yeast strain DSY-1 was co-transformed with various combinations of bait and prey
plasmids (see Material and Methods) and the ability to grow on selective media was tested to
confirm the success of the co-transformation (medium lacking leucine and tryptophan). The
ability to grow on medium lacking leucine, tryptophan and histidine was a proof for the
interaction between the two putative interactors. Negative controls were also included in the
interaction assay (see Material and Methods). The interaction of PPP7A with proteins not
involved in CEF was also tested. In particular, the PSII core protein D1, the integral subunit of
the chloroplast ATPase CF0-IV (both transmembrane proteins) and stromal components
involved in shuttling LHC proteins from the envelope to the thylakoid membrane (cpSRP54
and 43) were chosen as preys.
49

Results ____________________________________________________________________________________________
According to the results of the split ubiquitin assay, PPP7A interacts with PGR5 and with the
proteins involved in electron transfer (Fd and FNR) or involved in CEF (Cyt b6 and PSI-D).
There was no detectable interaction with subunits of PSII or the ATPase complex (Table 3.4).
To investigate the interaction of PGR5 with the other PPP7 interactors, the yeast two hybrid
assay was applied. The PGR5 coding sequence corresponding to the mature protein form, was
cloned in the vector pGADT7 carrying the GAL4 DNA activation domain, whereas the
sequences encoding the prey proteins were cloned in the pGBKT7, carrying the GAL4 DNA
binding domain.
The yeast strain AH109 was co-transformed with the indicated combinations of bait and prey
plasmids and its ability to grow on selective media was tested to confirm the success of the co-
transformation (medium lacking tryptophan and leucine) and the interaction between the two
putative interactors (β-galactosidase activity). Negative controls were performed as described
in Material and Methods.
In the yeast two hybrid assay, PGR5 was found to interact only with ferredoxin. Also the N-
and the C-terminal part of PPP7A were tested via the Y2H assay for interaction with PGR5,
but only weak interactions could be detected between Fd and the C-terminal part of PPP7A
(Table 3.4). No further interactions could be detected, implying that both loops might be
required for the correct folding structure necessary for such interactions.
PGR5 FNR1 FNR2 Fd PSI-D Cyt b6 PSII-D1 ATPaseIV cpSRP
PPP7A +a +a +a +a +a + a - a - a - a
PGR5 - b - b - b + b - b + a nd nd nd
Table 3.4. protein-protein interactions between PPP7 or PGR5 and other thylakoid proteins. a split ubiquitin
assay; b yeast two hybrid assay. -: negative interaction; +: positive interaction; nd: not determined.
Given that the topology of PPP7 proteins allows an efficient protein-protein interaction only
between the stromal exposed N- and C-terminal domains, it could be concluded that also PGR5
is localized on the stromal side of the thylakoid membrane. This stromal localisation of PGR5
was confirmed by immunoblot analysis with PGR5-specific antibodies after mild trypsin
treatment of intact thylakoids (Figure 3.19). The abundance of negatively charged amino acids
within the N-terminal loop of PPP7 (see Figure 3.2) suggests a possible electrostatic
interaction with the positively charged PGR5 protein (Munekage et al., 2002) in this region.
50

Results ____________________________________________________________________________________________
PGR5
PSII-O
Fig 3.19. Immunoblot analysis with antisera specific for PSII-O and PGR5 proteins before (lane –) and after (lane +) trypsin treatment of intact thylakoid membranes. Lanes were loaded with thylakoids corresponding to 5 µg of chlorophyll.
3.10 PPP7 interacts in planta with PSI and PGR5
It has already been shown that the lack of PPP7 does not affect the organization of the
photosynthetic machinery (see Figure 3.12). In consequence, it is of major interest to
understand whether PPP7 and its interactor PGR5, are part of a specific complex or if they are
only transiently associated with the cyt b6/f complex and/or PSI. To answer this question, the
presence of PPP7 and PGR5 proteins has been investigated in mutants lacking the different
photosynthetic complexes: hcf136 lacks a PSII assembly/stability factor and as a consequence,
the entire PSII (Plücken et al., 2002); psad1psad2 lacks both PSI-D isoforms (Ihnatowicz et
al., 2004) and therefore the PSI complex is missing in this mutant. In petc the gene coding for
the Rieske protein is not functional and the mutant lacks the entire cyt b6/f complex (Maiwald
et al., 2003). Finally, atpd was chosen as a mutant defective in the ATPase due to a mutation in
the gene coding for the ATPase δ subunit (Maiwald et al., 2003). As showen in Figure 3.20
PPP7 and PGR5 proteins are both present in all of the mutants tested. This suggests a stable
integration in the thylakoid membrane independent from the known major complexes. PPP7
and PGR5 seem also not to influence the stability of the photosynthetic complexes since all the
major complexes are present in the ppp7ab and pgr5 mutants.
Fig 3.20. Immunodetection of the different subunits of photosynthetic complexes in WT, hcf136, psad1psad2, petc, atpd, ppp7ab and pgr5 mutants. On the left the used antibodies are indicated. Lanes were loaded with total protein extracts corresponding to 10 µg of proteins.
51

Results ____________________________________________________________________________________________
Interestingly, is that in the absence of PPP7 no accumulation of PGR5 could be observed
(Figure 3.20) whereas PGR5 expression is not essential for accumulation of PPP7 (in the pgr5
mutant, PPP7 is only marginally decreased). To analyse the relationship between PPP7 and
PGR5 in more detail, preparations of total chloroplasts, thylakoids and stroma fraction were
probed with antibodies specific for PGR5 and PPP7 (Figure 3.21). It could be confirmed that
in the absence of PGR5, PPP7 still accumulates in the thylakoid membrane, while no traces of
PGR5 can be detected in chloroplasts of ppp7ab plants. The slightly reduced accumulation of
PPP7 in pgr5 plants can only be attributed to post-translational regulation (e.g. protein
degradation) since on the mRNA level, the amount of transcripts of PGR5 and PPP7 was not
altered in ppp7ab and pgr5 plants (Table 3.5). In case of PPP7 not being present in the
thylakoid membrane, PGR5 is probably synthesized but also rapidly degraded as no traces of
PGR5 can be found in the stroma fraction of ppp7ab.
Fig. 3.21. Accumulation of PPP7 and PGR5 proteins in different chloroplast subfractions of WT, ppp7ab and pgr5 plants. Lanes “chl” correspond to 5 μg chlorophyll of intact chloroplasts. Lanes “Thy” correspond to the thylakoid fraction resulting from the osmotic lysis of chloroplasts corresponding to 5 μg chlorophyll. Lanes “Str” correspond to the stromal fraction of the same lysis reaction. In the bottom panel, as control for the purity of the fractions, a Coomassie stained part of the gel is shown, where the large subunit of Rubisco migrated,
Transcript ppp7a ppp7b ppp7ab pgr5
PPP7A 0 ± 0 96 ± 8 0 ± 0 198 ± 37 PPP7B 60 ± 8 0 ± 0 0 ± 0 96 ± 13 PGR5 85 ± 35 71 ± 18 125 ± 20 1330 ± 799*
Table 3.5. Real Time RT-PCR analyses on the transcript levels of PGR5, PPP7A and PPP7B in pgr5, ppp7a, ppp7b and ppp7ab mutant leaves. Values are in % of WT and ACTIN1 transcript levels were used as an internal control. The values are calculated according to the 2-ΔΔCT method (Livak). *The pgr5 point mutation prevents accumulation of the PGR5 protein although it also results in increased levels of PGR5 mRNA.
52

Results ____________________________________________________________________________________________
Multiple factors could cause the impairment of CEF, which occurs in ppp7ab and pgr5
mutants. One possibility could be that other components of the CEF machinery are defective or
reduced, for example the cyt b6/f complex or the NDH complex, which is responsible for the
reduction of plastoquinone by NADPH. To answer this question, Western analyses have been
carried out on thylakoid preparation of WT, ppp7ab and pgr5 mutants using antibodies against
subunits of both complexes. As shown in Figure 3.22 the defect in CEF in ppp7ab and pgr5
can be entirely explained by the lack of PGR5 and PPP7 proteins in these mutants, because the
cyt b6/f complex and the NDH complex are normally present in both mutant lines..
Fig. 3.22. Accumulation of subunits of the cyt b6/f and NDH complexes in WT, ppp7ab and pgr5 mutant plants, detected by Western analysis. Analyses were performed on thylakoid proteins using specific antibodies raised against Cyt b6, Cyt f and subunit K of the plastid NDH complex. Lower amounts of WT proteins were loaded in lanes 2/3x WT and 1/3x WT, as indicated. In the bottom panel, a replicate gel stained with Coomassie Blue is shown as a loading control.
3.11 PPP7/PGR5-PSI interaction and PSI organization
As previously shown, PGR5 and PPP7 are involved in CEF (Figure 3.14 and 3.16). Still, it
remains to be clarified whether they are part of the PSI complex or “only” transiently
associated with it. PSI was hence isolated from pgr5, ppp7ab and WT leaves and probed with
antibodies against PGR5 and PPP7. As shown in Figure 3.23 both proteins are present in the
PSI fraction. If one compares equal amounts of PSI fractions and thylakoid extraction, it can be
concluded that PPP7 is less abundant in PSI.
With PGR5 this does not seem to be the case. In other words, the interaction of PPP7 with
PGR5 is not necessary for the stability of PPP7 (see also Figure 3.21); rather PPP7 requires
PGR5 to stably interact with PSI. It could also be possible that a fraction of PPP7 present in the
thylakoid membrane, does not bind PGR5.
53

Results ____________________________________________________________________________________________
Fig 3.23. Immunodetection of PGR5 and PPP7 in thylakoids and PSI fractions of WT and mutant leaves. Thylakoids were loaded equal to 3.5µg of chlorophyll, while the PSI fractions were loaded corresponding to 5 µg of chlorophyll. The lower panel shows the AuroDye (GE Healthcare) staining of the membrane after blotting, to confirm equal loading.
Because PGR5 and PPP7 are associated with the PSI complex and ppp7ab is defective in linear
electron flow, it was important to figure out whether the lack of PGR5 and PPP7 is affecting
the composition or structure of PSI and therefore influencing the linear electron flow. For that,
PSI has been isolated from pgr5, ppp7ab and WT leaves via sucrose gradient followed by
SDS-PAGE using 16 to 23% gradient gels. The analyses revealed that absence of PGR5 or
PPP7 does not influence the composition of PSI (Figure 3.24) supporting the idea that these
proteins are not subunits of PSI.
Fig 3.24. PSI subunit composition in wild-type and mutant plants. Equal amounts (corresponding to 5 µg of chlorophyll) of PSI isolated from mutant and WT leaves were fractionated by SDS-PAGE on a 16 to 23% acrylamide gel and visualised by Colloidal Coomassie staining. The positions of subunits were previously identified by immunodetection (Jensen et al., 2002)
54
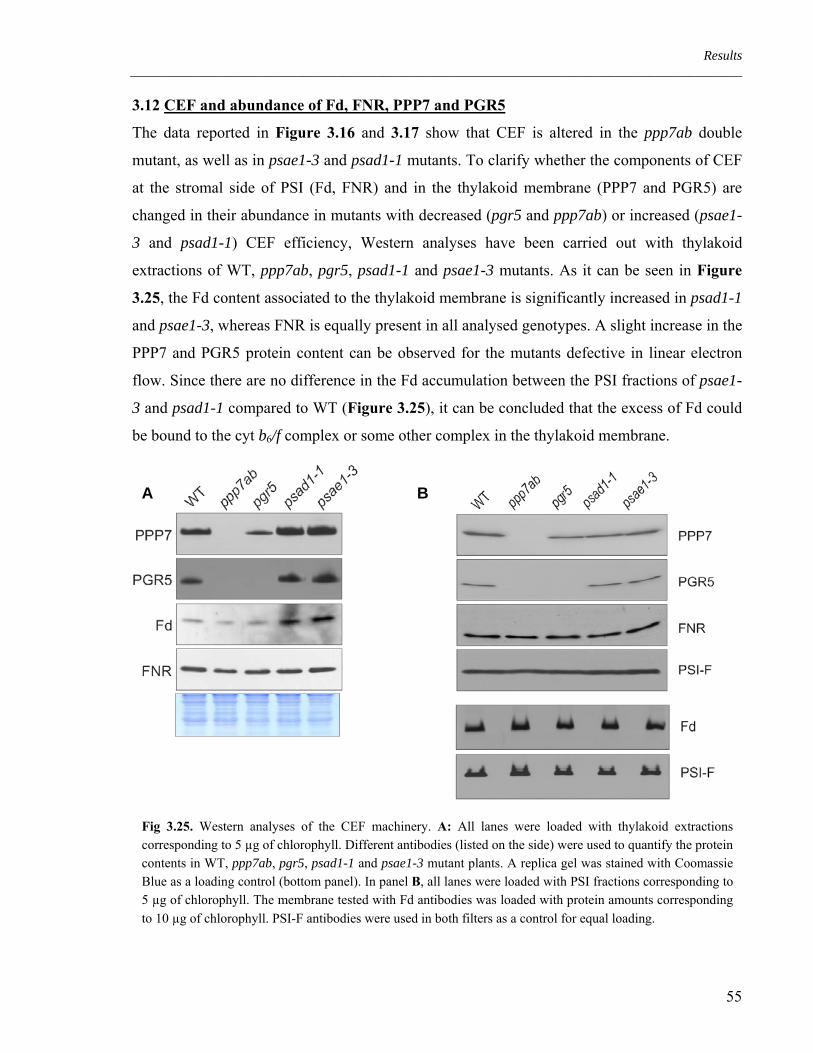
Results ____________________________________________________________________________________________
3.12 CEF and abundance of Fd, FNR, PPP7 and PGR5
The data reported in Figure 3.16 and 3.17 show that CEF is altered in the ppp7ab double
mutant, as well as in psae1-3 and psad1-1 mutants. To clarify whether the components of CEF
at the stromal side of PSI (Fd, FNR) and in the thylakoid membrane (PPP7 and PGR5) are
changed in their abundance in mutants with decreased (pgr5 and ppp7ab) or increased (psae1-
3 and psad1-1) CEF efficiency, Western analyses have been carried out with thylakoid
extractions of WT, ppp7ab, pgr5, psad1-1 and psae1-3 mutants. As it can be seen in Figure
3.25, the Fd content associated to the thylakoid membrane is significantly increased in psad1-1
and psae1-3, whereas FNR is equally present in all analysed genotypes. A slight increase in the
PPP7 and PGR5 protein content can be observed for the mutants defective in linear electron
flow. Since there are no difference in the Fd accumulation between the PSI fractions of psae1-
3 and psad1-1 compared to WT (Figure 3.25), it can be concluded that the excess of Fd could
be bound to the cyt b6/f complex or some other complex in the thylakoid membrane.
A B
Fig 3.25. Western analyses of the CEF machinery. A: All lanes were loaded with thylakoid extractions corresponding to 5 µg of chlorophyll. Different antibodies (listed on the side) were used to quantify the protein contents in WT, ppp7ab, pgr5, psad1-1 and psae1-3 mutant plants. A replica gel was stained with Coomassie Blue as a loading control (bottom panel). In panel B, all lanes were loaded with PSI fractions corresponding to 5 µg of chlorophyll. The membrane tested with Fd antibodies was loaded with protein amounts corresponding to 10 µg of chlorophyll. PSI-F antibodies were used in both filters as a control for equal loading.
55

Results ____________________________________________________________________________________________
3.13 Synthetic phenotype of the ppp7ab psad1-1 triple mutant
From the previous results, it can be suggested that in mutants with a defective linear electron
flow, the CEF is enhanced. This might lead to an increased acidification of the lumen and
consequent NPQ induction. Indeed, opposite to the pgr5 and ppp7ab mutants, the psad1-1 and
psae1-3 mutants show a faster transient induction of NPQ upon dark to light transition.
Because PPP7 appears to be present in large excess in WT plants (in ppp7a single mutant,
PPP7b level of only 10% is sufficient to maintain WT phenotype, see Figure 3.10 and 3.11)
the slight increase of PPP7 and PGR5 protein levels in psae1-3 and psad1-1 could be
responsible for the increase in CEF. If this is the case, severe effects on plant performance
should be detectable in a triple mutant defective in PSI-D or PSI-E and PPP7. For this purpose
ppp7ab psad1-1 triple mutants were generated by crossing psad1-1 single mutant with and
ppp7ab double mutants. The segregating F2 progeny was screened for homozygous triple
mutants via PCR. The lack of PPP7 and PGR5 was confirmed by Western analysis.
As shown in Figure 3.26, the triple mutant is significantly decreased in its growth (compared
to psad1-1) and has a pale green coloration.
Fig 3.26. Growth phenotypes of 3-week-old WT, ppp7ab, psad1-1 and ppp7ab psad1-1 mutants. Plants were grown in the greenhouse under long-day conditions.
The transient increase in non-photochemical quenching, which can be observed in psad1-1
during the transition from dark-to-light, occurs also in the triple mutant ppp7ab psad1-1 though
less prominent than in the psad1-1 single mutant (Figure 3.27). Surprisingly, the ppp7ab
mutation is not inhibiting the NPQ phenotype of psad1-1 in the triple mutant and this could be
explained by an involvement of PPP7 in the regulation of CEF rather than PPP7 being a direct
component of CEF itself. Another explanation could be that the remaining NDH is
56

Results ____________________________________________________________________________________________
compensating the deficiency in the PPP7/PGR5 depending pathway even though its efficiency
is decreased.
1 2 3 4 5 6
time (min)
0.2
0.4
0.6
0.8
1.0
1.2
1.41.6 Fig 3.27. Time course of induction and
relaxation of NPQ. Induction and relaxation of NPQ were monitored for 4 min at 80 µE m-2s-1 (white bar) followed by 2 min in the dark (black bar) using dark adapted leaves of WT plants (white circles), psad1-1 (dark diamonds), ppp7ab psad1-1 (white diamonds) and ppp7ab mutants (dark circles). Each value represent means ± standard deviation (5 plants each genotype).
NPQ
Analysing the CEF activity in the triple mutants, chlorophyll measurements on in vitro
ruptured chloroplasts showed that ppp7ab psad1-1 triple mutants perform CEF at a similar
extent as ppp7ab mutant (Figure 3.28).
NADPH Fd 30 sec
WT
ppp7ab
psad1-1
ppp7ab psad1-1
Chl
Flu
ores
cenc
e
Fig 3.28. Increases of chlorophyll fluorescence by addition of NADPH (0.25 mM) and Ferredoxin (Fd, 5 µM) under the illumination of weak measuring light (1 µE m-2 s-1) was monitored in osmotically ruptured chloroplasts (20 µg chlorophyll/mL) of WT, psad1-1, ppp7ab and ppp7ab psad1-1 mutants.
As it was demonstrated for ppp7ab, also in the case of the triple mutant the measurement of
CEF via the in vivo analysis provides different result. As can be seen in Figure 3.29, the in
vivo estimation of the CEF suggests that even considering the significantly lower signal, the
57

Results ____________________________________________________________________________________________
triple mutant ppp7ab psad1-1 has an intermediate activity in comparison with the two parental
mutants.
A B
C D
0.2
0.4
0.6
0.8
1.0
1.2
0
0.2
0.4
0.6
0.8
1.0
1.2
0
0.2
0.4
0.6
0.8
1.0
1.2
0
0.2
0.4
0.6
0.8
1.0
1.2
0
Fig 3.29. Quantification of CEF in intact leaves. P700 oxidation was induced by illumination with far-red light. Dark-adapted leaves from WT (panel A), psad1-1 (panel B), ppp7ab (panel C) and ppp7ab psad1-1 (panel D) plants were illuminated for different times with green light to induce LEF (black colour 0 s and red colour 20 s), and P700 oxidation was recorded 200 ms after the green light was switched off. A 10-min exposure to green light, followed by 2 min in the dark is indicated by green curves. Data are expressed as relative values.
58

Discussion ____________________________________________________________________________________________
4. Discussion
4.1 PPP7: a novel component of CEF around PSI
Since the discovery of cyclic electron flow (CEF), the pathway(s) taken by the electrons to
flow around PSI remain unclear and under debate. The existence of at least two possible
routes, via ferredoxin (Fd) and via the plastidial NADPH dehydrogenase (NDH), is generally
accepted but their interconnections and interactions are still obscure.
Recently the protein PGR5 has been identified as a component of the antimycin-A sensitive
Fd-dependent pathway and the effects of a lack of PGR5 in chloroplasts have been subject of
extensive studies (Munekage et al., 2002; Munekage et al., 2004). The phenotype of the pgr5
mutant in Arabidopsis suggests that PGR5 plays either a direct role in CEF (Shikanai, 2007) or
acts as a regulator of the switch between linear and cyclic electron transfer (Avenson et al.,
2005; Nandha et al., 2007) with its absence leading to a reduced capability of CEF to compete
with linear electron flow. To date, the exact function of PGR5 and its precise localization in the
thylakoid membrane are still unclear.
This thesis focuses on an Arabidopsis protein that we named PPP7, a novel component of the
thylakoid machinery involved in the electron transfer from ferredoxin to plastoquinone.
Orthologous genes coding for PPP7 proteins can be found in the genomes of higher plants and
eukaryotic algae (Figure 3.2) as well as mosses. Apparently it is not encoded by the genome of
cyanobacteria, leading to the conclusion that it is an eukaryotic invention.
The genome of Arabidopsis thaliana contains two genes (At4g22890 and At4g11960) coding
for homologous proteins with redundant functions, PPP7A and PPP7B. PPP7A is highly
expressed, while PPP7B is only weakly expressed. The mRNA of both genes can be found in
green tissues but not in roots (Figure 3.1). The proteins are targeted to the chloroplasts where
they fold into the thylakoid membranes in a manner that the N- and C-terminal domains are
exposed towards the stroma (Figure 3.3, 3.4 and 3.6).
Our data suggest that due to this topology, PPP7 interacts with PGR5 which is present on the
stromal side (Figure 3.19). Supporting this model, PGR5 has been characterized to be
associated with, but not integrated into, the thylakoid membrane because it does not contain
any transmembrane domain (Munekage et al., 2002). In the Arabidopsis double mutant
ppp7ab, completely devoid of PPP7, the PGR5 gene is normally transcribed but probably the
59

Discussion ____________________________________________________________________________________________
protein is not able to anchor on the membrane via PPP7 and therefore it is most likely rapidly
degraded. On the other hand, if PGR5 is missing, PPP7 is still synthesized and integrated into
the thylakoid membrane but to a lesser extent (Figure 3.20).
The interaction of the PPP7/PGR5 complex with other proteins in the thylakoid membrane is
still not clear. It seems to be transiently but not tightly associated with PSI (Figure 3.20) or
cytb6/f (Table 3.4). The PPP7/PGR5 complex is not a structural component of the major
photosynthetic complexes and most likely it is present and stable by itself in the thylakoid
membrane (Figure 3.20). Confirming these hypotheses, the depletion of PPP7 and
consequently of PGR5 does not affect the organization and composition of the photosynthetic
machinery and mutant plants can grow photo-autotrophically (Figure 3.10 and 3.12).
4.2 PPP7 and PGR5 proteins are involved in the same pathway of CEF
Regarding photosynthetic performance, PPP7 and PGR5 functionally interact and can be
considered as components or, at least, regulators of the ferredoxin-dependent CEF around PSI.
In ppp7ab plants, the reaction centre of the PSI (P700) remains in a reduced state during actinic
light illumination and it is oxidized to a normal level only under far red light, which favours
mainly PSI photochemistry (Figure 3.15). This phenotype may be explained by the
phenomenon called charge recombination in P700, which is caused by an over-reduction of the
stromal electron acceptors from PSI, ferredoxin and NADP+. Under these conditions, electrons
return to P700 from the PSI-acceptor-side electron carriers (i.e. A0, A1, FX and FA/FB). This has
been proven by the fact that the artificial electron acceptor methylviologen, known to acquire
electrons directly from the early acceptors of PSI, is restoring WT P700 oxidation ratio in the
pgr5 mutant (Munekage et al., 2002). The results obtained with the methylviologen treatment
provide an indication that this limited P700 oxidation is not due to a defect in the electron
transfer chain within PSI. Moreover, taking into account that treating isolated thylakoids of
pgr5 with exogenous electron acceptors, linear electron flow (LEF) appears to occur as in WT
plants (Munekage et al., 2002) and it can be concluded that the linear electron flow is not per
se impaired in pgr5 and therefore, most likely also not in ppp7ab.
Hence, the over-reduction of the electron carriers on the stromal side of PSI could be explained
by a defective CEF around the PSI.
60

Discussion ____________________________________________________________________________________________
Up to now there are two techniques that are thought to directly quantify CEF. The first,
developed by Shikanai and co-workers (Endo et al., 1998), is an in vitro evaluation of the
electron transfer from the ferredoxin to the plastoquinone pool. In this assay, exogenous
ferredoxin is used as electron carrier and the NADPH as electron source. NADPH donates
electrons indirectly via reduced ferredoxin generated by the thylakoid-bound ferredoxin
NADP+ reductase (FNR). In this assay only the first steps of CEF are monitored, but the
destiny of the electrons, once loaded on the plastoquinone, is not followed. Applying this
assay, the ppp7ab mutant and the pgr5 mutant show a markedly reduced CEF (Figure 3.16).
On the other hand, the in vivo measurements of CEF designed by Joliot and Joliot (2006) are
estimating the rate of the oxidation of PSI. In this way, only the oxidation rate of P700 is
considered, independently where the electrons go or come from. By this method, ppp7ab and
pgr5 mutants result to be impaired only in the transition between cyclic and linear electron
flow, while they both potentially perform WT level of CEF (Figure 3.17).
It is not easy to clarify which of the two techniques is correct or most effective in measuring
CEF. The differences between the two experiments are responsible for the diverse results
obtained analysing the mutants. One possible conclusion, which is taking into account the
result of both experiments, is that the lack of PPP7 and PGR5 results in a defective regulation
of CEF around PSI. In any case, the redox state of the stroma is unbalanced in ppp7ab with an
over-accumulation of reduced electron carriers (mostly Fd and NADPH).
Due to the reduced availability of oxidized ferredoxin, electrons are leaving PSI less rapidly
and could be therefore responsible for charge recombination. Caused by this impairment of
CEF in ppp7ab mutant, like in pgr5, less protons are pumped into the thylakoid lumen and as a
consequence, the rapid and transient increase in non-photochemical quenching normally
associated to the dark-light transition is decreased (Figure 3.14).
Under high light conditions this effect is even more severe, leading to a reduction of NPQ to
almost 60%. This reduction of non-photochemical quenching is not responsible for those
defects in linear electron flow like the decreased electron transport rate and the decreased PSII
quantum yield (Figure 3.13 and Table 3.3). Indeed, the linear electron transport is only
marginally impaired in mutants completely lacking NPQ, as it is the case for the Arabidopsis
npq4 mutant (Li et al., 2000).
61

Discussion ____________________________________________________________________________________________
Because CEF is thought to enhance ATP synthesis balancing the ATP/NADPH ratio, in
ppp7ab the deficient ATP production could lead to an over-reduction of the stroma because of
the slower consumption of reducing power in the carbon fixing reactions. This could ultimately
turn in a reduced linear electron flow, reflected by a slow-growth phenotype (Avenson et al.,
2005; Kramer et al., 2004).
4.3 A new model for CEF around PSI
To clarify the localization of the PPP7/PGR5 complex, interaction assays have been developed
studying the interaction of both PPP7 and PGR5 with different subunits of the photosynthetic
machinery, belonging to the PSII, PSI, cyt b6/f complex and the ATP-synthase. Combining
split ubiquitin- and yeast two hybrids-assays it has been found that the PPP7/PGR5 complex
probably interacts with some of the proteins that play a role in CEF: the Fd/FNR stromal
system, at least one subunit of the cyt b6/f complex and of PSI (Table 3.4). In addition, a
further biochemical indication is the co-purification of PPP7/PGR5 complex with PSI (Figure
3.23).
As already discussed, PPP7 is a membrane integrated thylakoid protein and this feature allows
lateral migration and/or interaction with other multi-protein complexes. Considering these
findings together, a model could be proposed for the function of the PPP7/PGR5 complex in
the thylakoid membranes. In the following pictures, the possible localization of the
PPP7/PGR5 complex within the thylakoid membrane and its activity are depicted.
As a first thought, proven by the fact that PGR5 and PPP7 co-purify with the PSI complex
(Figure 3.23), the complex could be imagined as tightly associated with PSI (LEF1) implying
that during CEF the cyt b6/f complex is in close contact with PSI and PPP7/PGR5 (CEF1).
62

Discussion ____________________________________________________________________________________________
Considering that both PPP7 and PGR5 can accumulate to WT levels in mutants depleted of PSI
or cyt b6/f (Figure 3.20) and that until now the existence of a PSI- cyt b6/f super-complex could
not be experimentally confirmed (Breyton et al., 2006) this first model has to be revised.
More likely, the stability of the PPP7/PGR5 complex in the absence of PSI or cyt b6/f suggests
that during linear electron flow the PPP7/PGR5 complex is independent and free to migrate in
the thylakoid membrane (LEF2) and it can interact alternatively with PSI and cyt b6/f possibly
shuttling FNR-Fd or creating an alternative CEF-favouring conformation (CEF2). This
hypothesis is compatible with previous findings of FNR association with cyt b6/f (Zhang et al.,
2001).
Once that FNR and Fd are bound to the PPP7/PGR5 complex and interact with the cyt b6/f
complex, the electrons could be directly injected into the plastoquinone pool without binding to
any other protein (CEF2a) or more likely, by means of the still unidentified FQR enzyme
(CEF2b).
63

Discussion ____________________________________________________________________________________________
Regarding the physical way taken by electrons to reach the oxidized plastoquinone two points
have to be taken into account:
- on one hand PGR5 apparently lacks known domains appointed to bind electron carrier
elements (i.e. metal binding motifs or NAD+ or FAD+ co-enzymes) on the other hand, the
biochemical properties of PPP7 are still unknown;
- the involvement of a putative FQR could not be excluded, even if the conformational
arrangement offered to the Fd and FNR by binding with the PPP7/PGR5 complex could be
sufficient for electron transfer.
The question is still unsolved whether the electrons are transferred directly to the oxidized
forms of the plastoquinone or if auxiliary elements are involved. One of these could be
represented by the novel haem (ci) on the stromal side at the Qi site of the cyt b6/f complex
which is a typical feature of the plastidic cytochrome complex (Kurisu et al., 2003 and
Stroebel et al., 2003) but the actual involvement of this group in transferring electrons is still
under debate. The discovery that the pgr1 mutation (point mutation of the Rieske protein),
which causes a defective Q-cycle (Okegawa et al., 2005), does not affect Fd-dependent CEF,
might suggest that ferredoxin directly donates electrons to oxidized plastoquinone (PQ) and not
to the cyt b6/f complex. Alternatively, it is also possible that the pgr1 mutation does not affect
electron transport from Fd to PQ through the cyt b6/f complex. In this last case, PGR5 together
with PPP7 could be factors facilitating the access of Fd to the cyt b6/f complex.
4.4 Cyanobacteria do not have PPP7
PPP7 does not have homologues in cyanobacteria. This could be puzzling at first sight, since
the PGR5 protein is conserved among these species. It has to be taken into account that
cyanobacterial cyt b6/f preparations lack the FNR, in contrast to plant preparation (Zhang et al.,
64

Discussion ____________________________________________________________________________________________
2001). This indicates that the formation of the complex depicted in CEF2 model could be a
specific feature of eukaryotic thylakoids, implying that cyanobacteria developed slightly
different pathways and regulatory mechanisms of cyclic flow around PSI. Supporting this
hypothesis is that in Synechococcus sp. PCC7002 the PSI subunit E is required for CEF in vivo
(Yu et al., 1993) and in fact mutants lacking this subunit show defective CEF.
In vascular plants, PSI-E is not essential for CEF and moreover, the A. thaliana psae1-3 mutant
seems to perform even higher CEF around PSI (Figure 3.16 and 3.17). Probably PSI-E is
involved in the binding during the formation of an hypothetical “cyclic flow super-complex”
and therefore its lack results in a less stable or malfunctioning complex.
It appears possible that in cyanobacteria, PGR5 alone is sufficient to stabilize an interaction
between PSI (or Fd) and cyt b6/f or the complex responsible for the electron transfer from
ferredoxin to plastoquinone. Furthermore, differently from the eukaryotic ferredoxin, the
cyanobacterial one has a short C-terminal helix extending from the vicinity of its Fe-S cluster
(Fukuyama et al., 1980). It might be considered that this carboxy-terminal helix could
penetrate into the cyt b6/f complex binding to the haem ci (Cramer et al., 2006). Also in this
case, PGR5 alone could be responsible for the protein rearrangement appointed to switch from
linear to cyclic flow.
4.5 Independent accumulation of PPP7 from PGR5
Regarding the stoichiometry between PPP7 and PGR5 it is interesting to notice that probably
not the entire pool of PPP7 found in the thylakoid membrane is bound to PGR5. As showed in
Figure 3.23, the ratio PPP7 to PGR5 in the thylakoid membranes is lower than the ratio
between the two proteins bound to the PSI. It is reasonable that PPP7 is explicating more than
one function in the thylakoid membranes. Unfortunately, until now all transgenic lines carrying
PPP7A or PPP7B genes under control of the 35S promoter failed in over-expressing PPP7 and
therefore a direct control of the effect of over-expression of PPP7 is still missing. On the other
hand the over-expression of PGR5 resulted in plants with defective chloroplast development
(Okegawa et al., 2007). Moreover, the stromal redox state is unbalanced by the over-
accumulation of PGR5.
It is well documented that ferredoxin and NADP+/NADPH play a crucial role as redox
regulator in the chloroplasts. These molecules are involved in regulating enzymes of the Calvin
65

Discussion ____________________________________________________________________________________________
cycle via the thioredoxin system and as electron source for nitrogen and sulphur metabolism
and the general plastidic anabolism. Therefore, fine adjustment of the amount of reduced
ferredoxin and of the NADP+/NADPH ratio needs to be rigorously controlled. In this way the
abundance of PGR5 and PPP7 could be an important factor adjusting the competition between
linear and CEF, according to the environmental conditions and specific requirements of
reducing powers or electrons donors.
4.6 Link between LEF and CEF
The discussion about the cause-effect relationship between impaired CEF and decreased LEF
is still open. Our data support the idea that a defect in CEF results in a defective LEF, but the
opposite seems not to be the case. It has been extensively reported that Arabidopsis mutants
psad1-1 and psae1-3 display a reduced linear electron flow (Varotto et al., 2000; Ihantowicz et
al., 2004; Ihnatowicz et al., 2007). Interestingly they seem to have increased CEF (Figure 3.16
and 3.17), a fact that is confirmed by the higher NPQ induction during the dark-to-light
transition (Figure 3.18). This can be explained if we assume that a reduced content of PSI-E or
PSI-D leads to a decreased amount of FNR that transfer electrons from Fd to NADP+. Under
these circumstances, an alternative route could be followed by electrons that leave PSI to be
promptly re-injected into the plastoquinone pool.
In accordance with this hypothesis, psad1-1 and psae1-3 show a higher amount of thylakoid-
bound Fd, PPP7 and PGR5 (Figure 3.25). Interestingly, a closer look of Figure 3.16 shows
that the addition of NADPH to psad1-1 and psae1-3 ruptured chloroplasts induces a slight
increase in chlorophyll fluorescence, which is not found in WT and in the other analysed
genotypes. This could be explained when considering that the amount of ferredoxin bound to
the thylakoid membranes in psad1-1 and psae1-3 is higher than in WT. It seems plausible that
this higher level of endogenous ferredoxin is responsible for the electron transfer from
NADPH to the reduced plastoquinone pool. In the case of WT and ppp7ab (as well as pgr5)
the smaller amount of ferredoxin bound to the thylakoids could result in a negligible increase
in chlorophyll fluorescence.
An alternative explanation of the higher CEF displayed by psad1-1 and psae1-3 could be that
under genetic conditions limiting the transfer of electrons from Fd to NADP+, CEF is activated
66

Discussion ____________________________________________________________________________________________
as protection mechanism, dissipating excessive energy of the active PSII through increased
luminal acidification that turns in induction of NPQ.
Since the amount of required PPP7 is probably relatively low (considering that roughly just
10% of the protein is enough to give WT phenotype, see Figure 3.9 and 3.10) the increase in
the level of the PPP7/PGR5 complex in psae1-3 and psad1-1 mutants could be sufficient for
the increase in CEF. Interestingly, when both linear electron flow and this safety valve are shut
down (as in the triple mutant ppp7ab psad1-1), the plant is still able to survive even if
decreased in size and paler in leaf coloration (Figure 3.26).
Regarding the activity of CEF, preliminary results based on the in vitro techniques show that
CEF activity in the ppp7ab psad1-1 triple mutant is comparable to that of the ppp7ab mutant
(Figure 3.28). On the other hand, if CEF is measured according to the in vivo measurement
developed by Joliot and colleagues, ppp7ab psad1-1 appears to have an intermediate
phenotype between ppp7ab and psad1-1 (Figure 3.29). In any case, it is important to notice
that the lack of the PPP7/PGR5 complex cause a defective CEF even in a mutant that is
genetically prompt to higher cyclic activity.
As a consequence, the transient NPQ normally generated during the dark-to-light transition is
decreased. Noticeably, as showed in Figure 3.27, the NPQ of the triple mutant is reduced if
compared to psad1-1, but remains higher than in WT leaves. A possible explanation for this
phenotype could be represented by the alternative cyclic pathways through the NDH complex.
This alternative route probably compensates the defective ferredoxin-dependent pathway even
though it is supposed to be less efficient (Sazanov et al., 1998; Joet et al., 2002).
Alternatively, we could consider that PPP7 and PGR5 are rather involved in the switch
between linear to CEF, rather than in CEF itself. In this case, the psad1-1 genetic background
favourites cyclic flow upon linear. In the triple mutant, the lack of the PPP7/PGR5 complex
denies the possibility for the enhanced switch from linear to cyclic. As a result, the depletion of
one of those mechanisms enabling plants to cope with the fluctuating environmental conditions
is ultimately influencing the plant fitness.
67

Discussion ____________________________________________________________________________________________
4.7 CEF and state transition
The trigger of the switch between linear electron flow and CEF is another issue that needs to
be discussed when comparing green algae to vascular plants. It has been first hypothesised and
then demonstrated in Chlamydomonas that CEF is activated upon induction of state transition
(Finazzi and Forti, 2004).
LHCII migration along the thylakoids is responsible for a membrane rearrangement in the
chloroplast (Georgakopulos and Argyroudi-Akoyunoglu, 1994). During state transition some
grana lamellae disassemble and convert into stroma lamellae. Due to his membranes
reorganization, the accessibility of the cyt b6/f to the PPP7/PGR5 complex binding Fd-FNR
could be higher, increasing the ferredoxin pool that contributes to cyclic flow (Finazzi and
Forti, 2004). It has been shown that when the cells switch from state 1 to state 2 the electron
flow pathway switches from linear to cyclic (Finazzi et al., 1999).
A rationale for this switch could be the fact that when PSI is equipped with LHCII and its
activity increases, it could be necessary to generate a secure electron source to compensate the
PSII reduced activity. On the other hand, if PSI is more active, the risk of reduction of
molecular oxygen is higher. Under these conditions, a safety system that re-directs electrons to
the plastoquinone pool preventing oxygen reduction would support photosynthetic flexibility.
Whether this mechanism would appreciably modify photosynthetic electron flow in higher
plants is still unknown. The population of LHCII antennae that move from PSII to PSI during
the state transition is highly different considering algae and higher plants. While in the first
species around 80% of the total antennae complex goes under state transition, in higher plants
only 20% of the LHCII population is thought to migrate to PSI (Allen, 1992; Delosome et al.,
1996). Concerning this aspect, the availability of mutants defective in state transition as well as
mutants with increased or decreased CEF, will definitely stimulate further research
understanding the regulation of the mechanisms responsible for the adjustment of chloroplast’s
redox state.
68

References __________________________________________________________________________________________
5. References
- Albertsson P.A. (2001) A quantitative model of the domain structure of the photosynthetic
membrane. Trends Plant Sci 6: 349-354
- Allakhverdiev S.I., Nishiyama Y., Miyairi S., Yamamoto H., Inagaki N., Kanesaki Y. and
Murata, N. (2002) Salt stress inhibits the repair of photodamaged photosystem II by
suppressing the transcription and translation of psbA genes in Synechocystis. Plant Physiol
130: 1443-1453
- AllenJ. F. (1992) Protein phosphorylation in regulation of photosynthesis. Biochim Biophys
Acta 1098: 275-335
- Allen J. F. and Forsberg J. (2001) Molecular recognition in thylakoid structure and function.
Trends Plant Sci 6: 317-326
- Alonso J.M., Stepanova A.N., Leisse T.J., Kim C.J., Chen H., Shinn P., Stevenson D.K.,
Zimmerman J., Barajas P., Cheuk R., Gadrinab C., Heller C., Jeske A., Koesema E., Meyers
C.C., Parker H., Prednis L., Ansari Y., Choy N., Deen H., Geralt M., Hazari N., Hom E.,
Karnes M., Mulholland C., Ndubaku R., Schmidt I., Guzman P., Aguilar-Henonin L., Schmid
M., Weigel D., Carter D.E., Marchand T., Risseeuw E., Brogden D., Zeko A., Crosby W.L.,
Berry C.C., Ecker J.R. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana.
Science 301: 653-657
- Arnon D.I., Whatley F.R. and Allen M.B. (1955) Vitamin K as a cofactor of photosynthetic
phosphorylation. Biochim Biophys Acta 16: 607-608
- Aronsson H. and Jarvis P. (2002) A simple method for isolating import-competent
Arabidopsis chloroplasts. FEBS Letters 529: 215-220
69

References __________________________________________________________________________________________
- Bassi R., dal Belin Peruffo A., Barbato R. and Ghisi R. (1985) Differences in chlorophyll-
protein complexes and composition of polypeptides between thylakoids from bundle sheaths
and mesophyll cells in maize. Eur J Biochem 146: 589-595
- Bassi R., Marquardt J. and Lavergne J. (1995) Biochemical and functional properties of
photosystem II in agranal membranes from maize mesophyll and bundle sheath chloroplasts.
Eur J Biochem 233: 709-719
- Bendall D.S. and Manasse R.S. (1995) Cyclic photophosphorilation and electron transport.
Biochim Biophys Acta 1229: 23-38
- Berman-Frank I., Lundgren P. and Falkowski P. (2003) Nitrogen fixation and photosynthetic
oxygen evolution in cyanobacteria. Res Microbiol 154: 157-164
- Biehl A., Richly E., Noutsos C., Salamini F. and Leister D. (2005) Analysis of 101 nuclear
transcriptomes reveals 23 distinct regulons and their relationship to metabolism, chromosomal
gene distribution and co-ordination of nuclear and plastid gene expression. Gene 3: 33-41
- Boardman N. K. and Anderson Jan M. (1967) Fractionation of the photochemical systems of
photosynthesis. II. Cytochrome and carotenoid contents of particles isolated from spinach
chloroplasts Biochim Biophys Acta 143: 187-203
- Breyton C., Nandha B., Johnson GN., Joliot P. and Finazzi G. (2006) Redox modulation of
cyclic electron flow around photosystem I in C3 plants. Biochemistry 45: 13465-13475
- Bukhov N. and Carpentier R. (2004) Alternative photosystem I-driven electron transport
routes: mechanisms and functions. Photosynth Res 82: 17-33
- Bukhov N., Egorova E. and Carpentier R. (2002) Electron flow to photosystem I from
stromal reductants in vivo: the size of the pool of stromal reductants controls the rate of
70

References __________________________________________________________________________________________
electron donation to both rapidly and slowly reducing photosystem I units. Planta 215: 812-
820
- Burrows P.A., Sazanov L.A., Svab Z., Maliga P. and Nixon PJ. (1998) Identification of a
functional respiratory complex in chloroplasts through analysis of tobacco mutants containing
disrupted plastid ndh genes. EMBO Journal 17: 868-876
- Campbell W.J. and Ogren W.L. (1992) Light activation of Rubisco by Rubisco activase and
thylakoid membranes. Plant Cell Physiol 33: 751-756
- Clough S. J. and Bent A. F. (1998) Floral dip: a simplified method for Agrobacterium-
mediated transformation of Arabidopsis thaliana. Plant J 16: 735-743
- Cooley J. W., Howitt C. A. and Vermans W. F. F. (2002) Succinate quinol oxidoreductases in
the cyanobacterium Synechocystis sp. strain PCC 6803: presence and function in metabolism
and electron transport. J Bacteriol 182: 714-722
- Crafts-Brandner S.J. and Salvucci M.E. (2000) Rubisco activase constrains the
photosynthetic potential of leaves at high temperature and CO2. PNAS-USA 97: 13430-13435
- Cramer W.A., Zhang H., Yan J., Kurisu G. and Smith J.L. (2006) Transmembrane traffic in
the cytochrome b6f complex. Annu Rev Biochem 75: 769-790
- Delosome R., Olive J. and Wollman F. A. (1996) Changes in light energy distribution upon
state transition: an in vivo photoacoustic study of the wild type and photosynthesis mutants
from Chlamydomonas reinhardtii. Biochim Biophys Acta 1273: 150-158
- Dovzhenko A., Dal Bosco C., Meurer J. and Koop H. U. (2003) Efficient regeneration from
cotyledon protoplasts in Arabidopsis thaliana. Protoplasma 222: 107-111
71

References __________________________________________________________________________________________
- Endo T., Shikanai T., Takabayashi A., Asada K. and Sato F. (1999) The role of chloroplastic
NAD(P)H dehydrogenase in photoprotection. FEBS Letters 457: 5-8
- Endo T., Shikanai T., Sato F. and Asada K. (1998) NAD(P)H Dehydrogenase-Dependent,
antimycin A-sensitive electron donation to plastoquinone in tobacco chloroplasts. Plant Cell
Physiol 39: 1226-1231
- Ernst A., Böhme H. and Böger P. (1983) Phosphorylation and nitrogenase activity in isolated
heterocysts from Anabaena variabilis. Biochim Biophys Acta 723: 83-90
- Färber A., Young A. J., Ruban A. V., Horton P. and Jahns P. (1997): Dynamics of
xanthophyll-cycle activity in different antenna subcomplexes in the photosynthetic membranes
of higher plants - The relationship between zeaxanthin conversion and nonphotochemical
fluorescence quenching. Plant Physiol 115: 1609-1618
- Finazzi G., Furia A., Barbagallo R.P. and Forti G. (1999) State transition, cyclic and linear
electron transport and photophosphorylation in Chlamydomonas reinhardtii. Biochim Biophys
Acta 1413: 117-129
- Finazzi G. and Forti G. (2004) Metabolic flexibility of the green alga Chlamydomonas
reinhardtii as revealed by the link between state transition and cyclic electron flow. Photosynth
Res 82: 327-338
- Friso G., Giacomelli L., Ytterberg A.J., Peltier J.B., Rudella A., Sun Q. and Wijk K.J. (2004)
In-depth analysis of the thylakoid membrane proteome of Arabidopsis thaliana chloroplasts:
new proteins, new functions, and a plastid proteome database. Plant Cell 16: 478-499
- Fukuyama K., Hase T., Matsumoto S., Tsukihara T., Katsube Y., Tanaka N., Kakudo M.,
Wada K. and Matsubara H. (1980) Structure of Spirulina platensis [2Fe-2S] ferredoxin and
evolution of chloroplast-type ferredoxins. Nature 286: 522-524
72

References __________________________________________________________________________________________
- Georgakopoulos J.H. and Argyroudi-Akoyunoglou J.H. (1994) On the question of lateral
migration of LHC II upon thylakoid protein phosphorylation in isolated pea chloroplasts: the
stroma lamellar fraction separated from phosphorylated chloroplasts is not homogeneous.
Biochim Biophys Acta 1188: 380-390
- Golding A.J. and Johnson G.N. (2004) Down-regulation of linear and activation of cyclic
electron transport during drought. Planta 218: 107-114 Erratum in: Planta. 218: 682
- Harbinson J. and Woodward F.I. (1987) The use of light induced absorbency changes at 820
nm to monitor the oxidation state of P700 in leaves. Plant Cell Environ 10: 131-140
- Harper J.W., Adami G.R., Wie N., Keyomarsi K. and Elledge S.J. (1993) The p21 Cdk-
interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 75: 805-816
- Havaux M., Rumeau D. and Ducruet J.M. (2005) Probing the FQR and NDH activities
involved in cyclic electron transport around photosystem I by the 'afterglow' luminescence.
Biochim Biophys Acta 1709: 203-213
- Heber U. and Walker D. (1992) Concerning a dual function of coupled cyclic electron
transport in leaves. Plant Physiol 100: 1621-1626
- Herbert S.K., Fork D.C. and Malkin S. (1990) Photoacoustic measurements in vivo of energy
storage by cyclic electron flow in algae and higher plants. Plant Physiol 94: 926-934
- Hualing M., Endo T., Ogawa T. and Asada K. (1995) Thylakoid membrane-bound, NADPH-
specific pyridine nucleotide dehydrogenase complex mediates cyclic electron transport in the
cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol 36: 661-668
- Ihnatovicz A., Pesaresi P. and Leister D. (2007) The E subunit of photosystem I is not
essential for linear electron flow and photoautotrophic growth in Arabidopsis thaliana. Planta
226: 889-895
73

References __________________________________________________________________________________________
- Ihnatowicz A., Pesaresi P., Varotto C., Richly E., Schneider A., Jahns P., Salamini F. and
Leister D. (2004) Mutants of photosystem I subunit D of Arabidopsis thaliana: effects on
photosynthesis, photosystem I stability and expression of nuclear genes for chloroplast
functions. Plant J 37: 839-852
- Ivanov B., Asada K. and Edwards G.E. (2007) Analysis of donors of electrons to
photosystem I and cyclic electron flow by redox kinetics of P700 in chloroplasts of isolated
bundle sheath strands of maize. Photosynth Res 92: 65-74
- Jach G., Binot E., Frings S., Luxa K. and Schell J. (2001) Use of red fluorescent protein from
Discosoma sp. (dsRED) as a reporter for plant gene expression. Plant J 28: 483-491
- James P., Hallada, J. and Craig E. A. (1996) Genomic libraries and a host strain designed for
highly efficient two-hybrid selection in yeast. Genetics 144: 1425-1436
- Jensen P.E., Bassi R., Boekema E.J., Dekker J.P., Jansson S., Leister D., Robinson C. and
Scheller H.V. (2007) Structure, function and regulation of plant photosystem I. Biochim
Biophys Acta 1767: 335-352
- Jensen P.E., Gilpin M., Knoetzel J. and Scheller H.V. (2000) The PSI-K subunit of
photosystem I is involved in the interaction between light-harvesting complex I and the
photosystem I reaction centre core. J Biol Chem 275: 24701-24708
- Joet T., Cournac L., Peltier G. and Havaux M. (2002) Cyclic electron flow around
photosystem I in C3 plants. In vivo control by the redox state of chloroplasts and involvement
of the NADH-dehydrogenase complex. Plant Physiol 128: 760-769
- Johnson G. (2005) Cyclic electron transport in C3 plants: fact or artefact? J Exp Bot 56: 407-
416
74

References __________________________________________________________________________________________
- Joliot P. and Joliot A. (2002) Cyclic electron transfer in plant leaf. PNAS-USA 99: 10209-
10214
- Joliot P., Beal D. and Joliot A. (2004) Cyclic electron flow under saturating excitation of
dark-adapted Arabidopsis leaves. Biochim Biophys Acta 1656: 166-176
- Joliot P. and Joliot A. (2005) Quantification of cyclic and linear flows in plants. PNAS-USA
102: 4913-4918
- Kanesaki Y., Suzuki I., Allakhverdiev S.I., Mikami K. and Murata N. (2002) Salt stress and
hyperosmotic stress regulate the expression of different sets of genes in Synechocystis sp.PCC
6803. Biochemical and Biophysical Research Community 290: 339-348
- Kleffmann T., Russenberger D., von Zychlinski A., Christopher W., Sjolander K., Gruissem
W. and Baginsky S. (2004) The Arabidopsis thaliana chloroplast proteome reveals pathway
abundance and novel protein functions. Curr Biol 14: 354-362
- Kofer W., Koop H.-U., Wanner G. and Steinmüller K. (1998) Mutagenesis of the genes
encoding subunits A, C, H, I, J and K of the plastid NAD(P)H-plastoquinone-oxidoreductase in
tobacco by polyethylene glycol-mediated plastome transformation. Mol Gen Genet 258: 166-
173
- Koop H. U., Steinmüller K., Wagner H., Rößler C., Eibl C. and Sacher L. (1996) Integration
of foreign sequences into the tobacco plastome via polyethylene glycol-mediated protoplast
transformation. Planta 199: 193-201
- Kramer D. M., Avenson T. J. and Edwards G. E. (2004) Dynamic flexibility in the light
reactions of photosynthesis governed by both electron and proton transfer reactions. Trends
Plant Sci 9: 349-357
75

References __________________________________________________________________________________________
- Kurisu G., Zhang H., Smith J.L. and Cramer W.A. (2003) Structure of the cytochrome b6/f
complex of oxygenic photosynthesis: tuning the cavity. Science 302: 1009-1014
- Laisk A., Eichelmann H., Oja V. and Peterson R. B. (2005) Control of cytochrome b6/f at low
and high light intensity and cyclic electron transport in leaves. Biochim Biophys Acta 1708: 79-
90
- Liu Y.G., Mitsukawa N., Oosumi T. and Whittier R.F. (1995) Efficient isolation and mapping
of Arabidopsis thaliana T-DNA insert junctions by thermal asymmetric interlaced PCR. Plant
J 3: 457-463
- Maiwald D., Dietzmann A., Jahns P., Pesaresi P., Joliot P., Joliot A., Levin J.Z., Salamini F.
and Leister D. (2003) Knock-out of the genes coding for the Rieske protein and the ATP-
synthase delta-subunit of Arabidopsis. Effects on photosynthesis, thylakoid protein
composition, and nuclear chloroplast gene expression. Plant Physiol 133: 191-202
- Malkin S. and Canaani O. (1994) The use and characteristics of the photoacoustic method in
the study of photosynthesis. Annu Rev Plant Phys 45: 493-526
- Matthijs H.C.P., Jeanjean R., Yeremenko N., Huisman J., Joset F. and Hellingwerf K.J.
(2002) Hypothesis: versatile function of ferredoxin-NADP+ reductase in cyanobacteria
provides regulation for transient photosystem I-driven cyclic electron flow. Funct Plant Biol
29: 201–210
- Mi H., Endo T., Schreiber U. and Asada K. (1992) Donation of electrons from cytosolic
components to the intersystem chain in the cyanobacterium Synechococcus sp. 7002 as
determined by the reduction of P700+. Plant Cell Physiol 33: 1099-1105
- Mills J. D., Crowther D., Slovacek RE., Hind G. and McCarty R.E. (1979) Electron transport
pathways in spinach chloroplasts. Reduction of the primary acceptor of photosystem II by
76

References __________________________________________________________________________________________
reduced nicotinamide adenine dinucleotide phosphate in the dark. Biochim Biophys acta 547:
127-137
- Miyake C., Horiguchi S., Makino A., Shinzaki Y., Yamamoto H. and Tomizawa K. (2005)
Effects of light intensity on cyclic electron flow around PSI and its relationship to non-
photochemical quenching of Chl fluorescence in tobacco leaves. Plant Cell Physiol 46: 1819-
1830
- Müller P., Li X-P. and Niyogi K.K. (2001) Non photochemical quenching. A response to
excess light energy. Plant Physiol 125: 1558-1566
- Munekage Y., Hojo M., Meurer J., Endo T., Tasaka M. and Shikanai T. (2002) PGR5 is
involved in cyclic electron flow around photosystem I and is essential for photoprotection in
Arabidopsis. Cell 110: 361-371
- Munekage Y., Hashimoto M., Miyake M., Tomizawa K I., Endo T., Tasaka M. and Shikanai
T. (2004) Cyclic electron flow around photosystem I is essential for photosynthesis. Nature
429: 579-582
- Munekage Y. and Shikanai T. (2005) Cyclic electron transport through photosystem I. Plant
Biotech 22: 361-369
- Nada A. and Soll J. (2004) Inner envelope protein 32 is imported into chloroplasts by a novel
pathway. J Cell Sci 117: 3975-3982
- Nandha B., Finazzi G., Joliot P., Haid S. and Johnson G. (2007) The role of PGR5 in the
redox poising of photosynthetic electron transport. Biochim Biophys Acta 1767:1252-1259
- Ogawa T. and Asada K. (1994) NAD(P)H dehydrogenase-dependent cyclic electron flow
around photosystem I in the cyanobacterium Synechocystis PCC 6803: a study of dark-starved
cells and spheroplasts. Plant Cell Physiol 35: 163-173
77

References __________________________________________________________________________________________
- Okegawa Y., Tsuyama M., Kobayashi Y. and Shikanai T. (2005) The pgr1 mutation in the
Rieske subunit of the cytochrome b6/f complex does not affect PGR5-dependent cyclic electron
transport around photosystem I. J Biol Chem 280: 28332-28336
- Okegawa Y., Long T.A., Iwano M., Takayama S., Kobayashi Y., Covert S.F. and Shikanai T.
(2007) Balanced PGR5 level is required for chloroplast development and optimum operation of
cyclic electron flow around photosystem I. Plant Cell Physiol in press
- Osmond C.B. (1981) Photorespiration and photoinhibition: Some implications for the
energetics of photosynthesis. Biochim Biophys Acta 639: 77-98
- Pasch J. C., Nickelsen J. and Schünemann D. (2005) The yeast split-ubiquitin system to study
chloroplast membrane protein interactions. Appl Microbiol Biot 69: 440-447
- Peltier J.B., Emanuelsson O., Kalume D.E., Ytterberg J., Friso G., Rudella A., Liberles D.A.,
Soderberg L., Roepstorff P., von Heijne G. and van Wijk K.J. (2002) Central functions of the
lumenal and peripheral thylakoid proteome of Arabidopsis determined by experimentation and
genome-wide prediction. Plant Cell 14: 211-236
- Pesaresi P., Lunde C., Jahns P., Tarantino D., Meurer J., Varotto C., Hirtz R.D., Soave C.,
Scheller H.V., Salamini F. and Leister D. (2002) A stable LHCII-PSI aggregate and
suppression of photosynthetic state transitions in the psae1-1 mutant of Arabidopsis thaliana.
Planta 215: 940-948
- Plücken H., Müller B., Grohmann D., Westhoff P. and Eichacker L.A. (2002) The HCF136
protein is essential for assembly of the photosystem II reaction center in Arabidopsis thaliana.
FEBS Letters 532: 85-90
- Porra R. J. (2002) The chequered history of the development and use of simultaneous
equations for the accurate determination of chlorophylls a and b. Photosynth Res 73: 149-156
78

References __________________________________________________________________________________________
- Richely E., Dietzmann A., Biehl A., Kurth J., Laloi C., Apel K., Slamini F. and Lesiter D.
(2003) Covariations in the nuclear chloroplast transcriptome reveal a regulatory master-switch.
EMBO Rep 4: 491-498
- Romanowska E., Drozak A., Pokorska B., Shiell B.J. and Michalski W.P. (2006)
Organization and activity of photosystems in the mesophyll and bundle sheath chloroplasts of
maize. J Plant Physiol 163: 607-618
- Rumeau D., Bécuwe-Linka N., Beyly A., Louwagie M., Garin J. and Peltier G. (2005) New
subunits NDH-M, -N, and -O, encoded by nuclear genes, are essential for plastid Ndh complex
functioning in higher plants. Plant Cell 17: 219-232
- Sambrook J., Fritsch E. F. and Maniatis T. (1989) Molecular cloning. New York: Cold
Spring Harbor Laboratory Press
- Schagger H. and von Jagow G. (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel
electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal Biochem
166: 368-379
- Schägger H. and von Jagow G. (1991) Blue native electrophoresis for isolation of membrane
protein complexes in enzymatically active form. Anal Biochem 199: 223-231
- Scheller H.V., Jensen P.E., Haldrup A., Lunde C. and Knoetzel J. (2001) Role of subunits in
eukaryotic Photosystem I. Biochim Biophys Acta 1507: 41-60
- Scherer S. (1990) Do photosynthetic and respiratory electron transport chains share redox
proteins? Trens Biochem Sci 15: 458-462
79

References __________________________________________________________________________________________
- Schreiber U., Schliwa U. and Bilger W. (1986) Continuous recording of photochemical
chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth
Res 10: 51-62
- Schreiber U., Klughammer C. and Neubauer C. (1988) Measuring P700 absorbance changes
around 830 nm with a new type of pulse modulation system. Z. Naturforsch. 43c: 686-698
- Seidel-Guyenot W., Schwabe C. and Büchel C. (1996) Kinetic and functional
characterization of a membrane-bound NAD(P)H dehydrogenase located in the chloroplasts of
Pleurochloris meiringensis (Xanthophyceae). Photosynth Res 49: 183-193
- Semenova G.A. (2001) Effect of urea and distilled water on the structure of the thylakoid
system. J Plant Physiol 158: 1041-1050
- Sherman F. (1991) Getting started with yeast. Method Enzymol 194: 3-21
- Shikanai T. (2007) Cyclic electron transport around Photosystem I: genetic approaches. Annu
Rev Plant Biol 58: 199-217
- Shikanai T., Endo T., Hashimoto T., Yamada Y., Asada K. and Yokota A. (1998) Directed
disruption of the tobacco ndhB gene impairs cyclic electron flow around photosystem I. PNAS-
USA 95: 9705-9709
- Shikanai T., Munekage Y., Shimizu K., Endo T. and Hashimoto T. (1999) Identification and
characterization of Arabidopsis mutants with reduced quenching of chlorophyll fluorescence.
Plant Cell Physiol 40: 1134-1142
- Stroebel D., Choquet Y., Popot J.L. and Picot D. (2003) An atypical haem in the cytochrome
b(6)f complex. Nature 426: 413-418
80

References __________________________________________________________________________________________
- Svab Z. and Maliga O. (1993) High-frequency plastid transformation in tobacco by selection
for a chimeric aadA gene. PNAS-USA 90: 913-17
- Avenson T. J., Cruz J. A., Kanazawa A. and Kramer D. M. (2005) Regulating the proton
budget of higher plant photosynthesis. PNAS-USA 102: 9709-9713
- Tagawa K., Tsujimoto H.Y. and Arnon D.I. (1963) Role of chloroplast ferredoxin in the
energy conversion process of photosynthesis. PNAS-USA 49: 567-572
- Taiz L., Zeiger E. (2006) Plant Physiology, Fourth Edition, Sinauer Associates INC
Publishers, Sunderland, Massachusetts
- Takabayashi A., Kishine M., Asada K., Endo T. and Sato F. (2005) Differential use of two
cyclic electron flows around photosystem I for driving CO2-concentration mechanism in C4
photosynthesis. PNAS-USA 102: 16898-16903
- Thomas D.J., Thomas J., Youderian P.A. and Herbert S.K. (2001) Photoinhibition and light-
induced cyclic electron transport in ndhB(-) and psaE(-) mutants of Synechocystis sp. PCC
6803. Plant Cell Physiol 42: 803-12
- Towbin H., Staehelin T. and Gordon J. (1979) Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets: procedure and some applications. PNAS-USA 76:
4350-4354
- Varotto C., Pesaresi P., Maiwald D., Kurth J., S., Salamini F. and Leister D. (2000b)
Identification of photosynthetic mutants of Arabibidopsis by automatic screening for altered
effective quantum yield of photosystem II. Photosynthetica 38: 497-504
- Varotto C., Pesaresi P., Meurer J., Oelmuller R., Steiner-Lange S., Salamini F. and Leister D.
(2000) Disruption of the Arabidopsis photosystem I gene PSA-E1 affects photosynthesis and
impairs growth. Plant J 22: 115-124
81

References __________________________________________________________________________________________
- Waegemann K. and Soll J. (1991) Characterization of the import apparatus in isolated outer
envelopes of chloroplasts. Plant J 1: 149-159
- Wolk C.P., Ernst A. and Elhai J. (1994) Heterocyst metabolism and development. The
Molecular Biology of Cyanobacteria ed. DA Bryant: 769-823
- Yeremenko N., Jeanjean R., Prommeenate P., Krasikov V., Nixon P.J., Vermaas W.F.,
Havaux M. and Matthijs H.C. (2005) Open reading frame ssr2016 is required for antimycin A-
sensitive photosystem I-driven cyclic electron flow in the cyanobacterium Synechocystis sp.
PCC 6803. Plant Cell Physiol 46: 1433-1436
- Yu L., Zhao J., Mühlenhoff U., Bryant D. and Golbeck J.H. (1993) PsaE is required for in
vivo cyclic electron flow around photosystem I in the cyanobacterium Synechococcus sp. PCC
7002. Plant Physiol 103: 171-180
- Zhang H.M., Whitelegge J.P. and Cramer W.A. (2001) Ferredoxin:NADP+ ossidoreductase is
a subunit of the chloroplast cytochrome b6f complex. J Biol Chem 276: 38159-38165
- Zhao J., Snyder W.B., Mühlenhoff U., Rhiel E., Warren P.V., Golbeck J.H. and Bryant D.A.
(1993) Cloning and characterization of the psaE gene of the cyanobacterium Synechococcus
sp. PCC 7002: Characterization of a psaE mutant and overproduction of the protein in
Escherichia coli. Mol Microbiol 9: 183-194
82

Acknowledgments __________________________________________________________________________________________
Acknowledgments
I would like to thank my supervisor, Prof. Dr. Dario Leister, who gave me the possibility to
develop my way to do science during all these years of PhD, discussing about my projects and
supporting me.
I thank everybody who contributed with experiments and discussions to this project: Dr. Elena
Aseeva for the in vitro import; Prof. Dr. Peter Jahns for the pigment analysis; PD Dr. Danja
Schünemann for the split ubiquitin assay; Dr. Simona Masiero for the yeast two hybrid assay;
Prof. Dr. Roberto Barbato and Dr. Paolo Pesaresi for the helpful discussion and the
contribution to the antibodies biosynthesis and Alexander Hertle for the PPP7A protein
expression and purification. Last but not least, I thank Prof. Dr. Pierre Joliot and Dr. Giovanni
Finazzi for their willingness in measuring in vivo cyclic electron flow.
A special thank to Prof. Dr. Toshiharu Shikanai and PD Dr. Jörg Meurer for discussion and
helpful suggestions for the interpretation of my results.
Many thanks to my lab colleagues, who taught me how to cope with a different culture and
how to stay in a foreign country. Special thanks to those who helped me to write this thesis
reading it and giving me advices.
I am glad and thankful to have met friends, which, with their presence and sustain, supported
me and stood my behaviour, making the time here lighter.
Finally, I am grateful to my family and to Maria Teresa, who have always been there for me,
encouraging and helping me whenever I needed it.
É stato bello!
83

Curriculum vitae ___________________________________________________________________________________________
Curriculum vitae PERSONAL DATA
Date of birth: 12 October 1979 Place of birth: Verona, Italy Citizenship: Italian RESEARCH EXPERIENCES
October 2005 – December 2007 Ludwig-Maximillians-Universität München (Germany) Department Biology I, Botanic - PhD continuation. Supervisor Prof. Dr. Dario Leister July 2004 – September 2005 Max Planck Institut für Züchtungsforschung (Köln, Germany) - PhD in “Characterization of the novel photosynthetic protein PPP7 involved in Cyclic Electron Flow around PSI”. Supervisor Prof. Dr. Dario Leister Summer 2004 Universitá degli Studi di Verona (Italy) Dipartimento Scientifico e Tecnologico - Technical assistant 2003 – 2004 Universitá degli Studi di Verona (Italy) Dipartimento Scientifico e Tecnologico - Diploma thesis in “Characterization of a bZIP transcriptional factor induced by Cadmium in Brassica juncea (L.) Czern”. Supervisor Prof. Dr. Antonella Furini EDUCATION
1998 – 2004 Universitá degli Studi di Verona, Italy - Degree in Plant and Microbial Biotechnology (graduated summa cum laude)
84

Curriculum vitae ___________________________________________________________________________________________
1993 – 1998 Liceo Scientifico Tecnologico, Galileo Galilei, Verona, Italy - Diploma, high school PUBLICATIONS
- Dal Corso G., Pesaresi P., Masiero S., Aseeva E., Schünemann D., Finazzi G., Joliot P., Barbato R. and Leister D. (2008) A complex of PGRL1 and PGR5 is involved in the switch between linear and ferredoxin-dependent cyclic electron flow. Cell: accepted - Noutsos C., Kleine T., Armbruster U., Dal Corso G. and Leister D. (2007) Nuclear insertions of organellar DNA can create novel patches of functional exon sequences. Trends Genet Nov 2; (Epub ahead of print) - Dal Corso G., Borgato L. and Furini A. (2005) In vitro plant regeneration of the heavy metal tolerant and hyperaccumulator Arabidopsis halleri (Brassicaceae). Plant Cell, Tissue and Organ Culture 82: 267-270 - Fusco N., Micheletto L., Dal Corso G., Borgato L. and Furini A. (2005) Identification of Cadmium-regulated genes by cDNA-AFLP in the heavy metal accumulator Brassica juncea. Journal of Experimental Botany 56: 3017-3027 POSTERS
- Fusco N., Dal Corso G., Borgato L, Furini A: Transcription profile of genes induced by cadmium in Brassica juncea for phytoremediation. SIGA meeting Verona (Italy), 24-27 September 2003 - Furini A., Fusco N., Dal Corso G., Micheletto L. and Borgato L.: Transcription profile of genes induced by cadmium in Brassica juncea. Phytoremediation: OECD Environmental and Molecular Biological Aspects. Mátraháza (Hungary), 9-12 September 2004 LANGUAGES
- Italian, mother tongue - English, fluent - German, basic
85

Ehrenwörtliche Versicherung __________________________________________________________________________________________
Ehrenwörtliche Versicherung
Hiermit versichere ich, dass ich die vorliegende Arbeit selbständig und nur unter Verwendung
der angegebenen Hilfsmittel und Quellen angefertigt habe. Der Autor hat zuvor nicht versucht,
anderweitig eine Dissertation einzureichen oder sich einer Doktorprüfung zu unterziehen. Die
Dissertation wurde keiner weiteren Prüfungskommission weder in Teilen noch als Ganzes
vorgelegt.
München,
Dezember 2007
Giovanni Dal Corso
86