Embed Size (px)
Citation preview

INTRODUCTION
Enclosed within all eukaryotic cells is a set of membrane-bound compartments that constitute the secretory andendocytic pathways. Communication between thesecompartments is mediated by transport vesicles that bud fromone compartment and deliver their contents to anothercompartment. Transport vesicles are employed in the export ofsoluble and membrane cargo through the secretory pathway, inthe import of molecules through the endocytic pathway, and inother intra- and intercompartmental transport pathways. Inorder to maintain organelle identity in the presence of aconstant flux of transport vesicles, the mechanisms underlyingmembrane trafficking are highly regulated and specific(Bennett, 1995; Pelham, 1996; Pryer et al., 1992; Rothman,1994).
Insights into the molecular mechanisms contributing tothe specificity of vesicular transport have been provided by
biochemical and genetic dissections of the secretory andendocytic pathways (Bennett, 1995; Hay and Scheller, 1997;Pelham, 1999). A general model known as the SNARE (SNAPreceptor) hypothesis has been proposed to explain thespecificity of targeting and fusion reactions (Sollner et al.,1993b). This model postulates that the correct pairing ofaddressing proteins on the transport vesicle (termed v-SNAREs) with the appropriate binding partners on the targetmembrane (termed t-SNAREs) mediates specific vesiculartrafficking events. The prototypic t- and v-SNAREs wereidentified through the biochemical characterization of synapticvesicle membrane trafficking. These studies demonstratedthat two t-SNARE proteins on the presynaptic plasmamembrane, syntaxin 1A (Bennett et al., 1992) and SNAP-25(synaptosome-associated protein of 25 kDa) (Oyler et al.,1989), and a v-SNARE on synaptic vesicles, VAMP (vesicle-associated membrane protein; also known as synaptobrevin)(Baumert et al., 1989; Elferink et al., 1989; Trimble et al.,
4291Journal of Cell Science 112, 4291-4304 (1999)Printed in Great Britain © The Company of Biologists Limited 1999JCS0539
The syntaxins are a large protein family implicated in thetargeting and fusion of intracellular transport vesicles. Asubset of proteins of this family are the four syntaxin 2splice variants, syntaxins 2A (2), 2B (2′), 2C (2′′ ) and 2D.Each syntaxin 2 variant contains an identical, or nearlyidentical, amino-terminal cytoplasmic domain followedby a distinct hydrophobic (syntaxins 2A and 2B) orhydrophilic (syntaxins 2C and 2D) carboxyl-terminaldomain. To investigate whether the difference among thesyntaxin 2 variants is functionally important, we haveexamined comparatively their RNA transcript and proteinexpression patterns, membrane associations, protein-protein interactions and intracellular localizations.Analysis of the RNA transcript and protein expressionpatterns demonstrated that syntaxins 2A, 2B and 2C arebroadly, but not uniformly, expressed while syntaxin 2Dexpression is restricted to the brain. Subcellularfractionation studies showed that syntaxins 2A and 2Bbehave as integral membrane proteins while syntaxin 2C is
only partially associated with membranes. In vitrobiochemical assays demonstrated that the syntaxin 2variants exhibit similar yet distinct interactions with otherproteins implicated in vesicular trafficking, includingSNAP-25, SNAP-23, VAMP-2 and n-sec1. In a variety ofnonpolarized cell types, syntaxins 2A and 2B localized toboth the plasma membrane and endosomal membranes.However, in two polarized epithelial cell lines, MDCK andCaco-2, syntaxin 2A localized predominantly to the apicalplasma membrane while syntaxin 2B was associated withboth the apical and the basolateral membranes. Theseobservations indicate that the distinct carboxyl-terminaldomains of the syntaxin 2 variants influence theirbiochemical and localization properties and may thereforeconfer upon these variants different functional roles in theregulation of intracellular membrane trafficking.
Key words: Syntaxin, SNARE, Membrane trafficking, Alternativesplicing, Polarized cell, Plasma membrane, Transmembrane domain
SUMMARY
Syntaxin 2 splice variants exhibit differential expression patterns,
biochemical properties and subcellular localizations
Beatriz Quiñones1,*, Kirsi Riento2, Vesa M. Olkkonen2, Stephen Hardy3,‡ and Mark K. Bennett1,§
1Department of Molecular and Cell Biology, 355 LSA, University of California, Berkeley, CA 94720, USA2Department of Biochemistry, National Public Health Institute, Mannerheimintie 166, FIN-00300, Helsinki, Finland3Cell Genesys, Foster City, CA 94404, USA*Present address: Department of Plant and Microbial Biology, 111 Koshland Hall, University of California, Berkeley, CA 94720, USA (e-mail: [email protected])§Present address: Chiron Corporation, 45560 Horton Street, Emeryville, CA 94608, USA¶Author for correspondence at present address: Rigel, Inc., 240 East Grand Ave., South San Francisco, CA 94080, USA (e-mail: [email protected])
Accepted 1 September; published on WWW 17 November 1999

4292
1988) form a highly stable ternary complex commonly referredto as the synaptic SNARE complex (Hayashi et al., 1994;Sollner et al., 1993a,b). The synaptic SNARE complex servesas a receptor for two general membrane trafficking factors, α-SNAP (soluble NSF attachment protein) and NSF (NEM-sensitive factor), which mediate its ATP-dependentdisassembly. The fact that each of the synaptic SNAREproteins is a target for the proteolytic action of Clostridialneurotoxins, which irreversibly inhibit neurotransmitter releasein vivo, highlights the importance of the SNARE complex insynaptic vesicle exocytosis (Jahn and Niemann, 1994;Montecucco, 1998).
The generality of the SNARE-based trafficking mechanismis illustrated by the fact that each of the synaptic SNAREproteins is a member of large v- and t-SNARE families thathave been identified in both yeast and animal cells (Bennettand Scheller, 1993; Ferro-Novick and Jahn, 1994; Hay andScheller, 1997; Pelham, 1999). The observation that distinct v-and t-SNAREs are required at different stages of the secretoryand endocytic pathways indicates that the SNARE proteins area component of the machinery that promotes the specificity oftransport vesicle targeting and fusion. Several properties of theSNARE proteins may contribute to such a role. First, the abilityof v- and t-SNAREs to form complexes in vitro is selective,although not absolutely specific. For example, syntaxin 1A andVAMP2 form a binary complex that is selectively potentiatedby the presence of SNAP-25 (Calakos et al., 1994; Pevsner etal., 1994a). However, a recent study utilizing the minimalinteracting domains from a variety of v- and t-SNAREsdemonstrated that the assembly of the SNARE complex coreis minimally selective (Yang et al., 1999). Thus, SNAREcomplex assembly is clearly not sufficient to account for thespecificity of vesicle targeting and fusion. Another propertythat likely influences SNARE function in vivo is intracellularlocalization. In most cases, SNARE proteins appear to belocalized to a restricted number of cellular membranes. Thus,the combination of selective localization and biochemicalproperties place constraints on the number of functionalSNARE complexes that can form within the cell. The fact thatrestricted combinations of complexed SNARE proteins can berecovered from cell extracts by coimmunoprecipitation isconsistent with this notion (Brennwald et al., 1994; Hay et al.,1997; Sogaard et al., 1994). Finally, two other protein families,the n-sec1/munc18 and rab/ypt families, have been implicatedin the regulation of membrane trafficking pathways (Novickand Zerial, 1997; Pevsner, 1996). These families modulateSNARE complex assembly, either directly (n-sec1 family) orindirectly (rab family), and may thereby further increase thecombinatorial mechanisms regulating the specificity of vesicletargeting and fusion.
The syntaxin family of t-SNAREs consists of at least 17gene products in animal cells that are localized tocompartments throughout the secretory and endocyticpathways, indicating that these proteins regulate variousmembrane trafficking pathways (Advani et al., 1998; Bock etal., 1997; Bock and Scheller, 1997; Prekeris et al., 1998;Steegmaier et al., 1998; Tang et al., 1998a,b,c,d; Wong et al.,1998). Additional diversity within the syntaxin family isgenerated by alternative splicing. Alternatively spliced formsof syntaxins 1A (Jagadish et al., 1997), 2 (Bennett et al., 1993),3 (Ibaraki et al., 1995) and 16 (Simonsen et al., 1998) have
been identified in mammalian cells. An alternatively splicedsyntaxin homologue in C. elegans (unc64) has also beendescribed (Ogawa et al., 1998; Saifee et al., 1998).Interestingly, the diversity generated by alternative splicingcommonly includes the membrane-proximal domain ofsyntaxin (H3), the region required for SNARE complexassembly, and/or the carboxyl-terminal hydrophobicmembrane anchor.
Although alternative splicing appears to represent a commonmechanism for generating structural diversity within the H3and membrane-anchoring domains of syntaxin isoforms, thefunctional consequences of this diversity remain largelyunknown. Therefore, we have comparatively analyzed theproperties of four variants of syntaxin 2 generated byalternative splicing. The properties that we have investigatedinclude: RNA transcript and protein expression patterns,membrane association, protein-protein interactions andintracellular localization. Our results demonstrate that,although the syntaxin 2 variants exhibit many similarities, theyalso display differences, suggesting that these proteins maydifferentially contribute to the regulation of intracellularmembrane trafficking events.
MATERIALS AND METHODS
MaterialsAntibodies used in the present study were from the following sources:anti-syntaxin 2C rabbit polyclonal antiserum (Gaisano et al., 1996;Peng et al., 1997), Berkeley Antibody (Richmond, CA); anti-ZO-1mouse monoclonal antibody mAbT8-754 (Itoh et al., 1991), a kindgift from Shoichiro Tsukita (Kyoto University, Kyoto, Japan); anti-ZO-1 rat monoclonal antibody (Stevenson et al., 1986), Chemicon(Temecula, CA); anti-FLAG mouse monoclonal antibody M2, Sigma(St Louis, MO); horseradish peroxidase (HRP)-conjugated secondaryantibodies, Kirkegaard and Perry Laboratories (Gaithersburg, MD);125I-labeled secondary antibody, ICN Pharmaceuticals, Inc. (CostaMesa, CA); fluorophore-conjugated secondary antibodies andfluorescein-conjugated anti-human IgG antibody, JacksonImmunochemical Labs (West Grove, PA). Cell culture media,antibiotics and solutions were obtained from Gibco BRL LifeTechnologies (Grand Island, NY) and Mediatech Cellgro (Herndon,VA.). Fetal bovine serum (FBS) was purchased from Gemini Bio-Products (Calabasas, CA) and Gibco BRL Life Technologies (GrandIsland, NY). Horse serum (HS) was obtained from SummitBiotechnologies (Fort Collins, CO). Transwell polycarbonate filters(12 mm, 0.4 µm pore size) were purchased from Corning CoStar(Cambridge, MA). Brefeldin A was purchased from Alexis (SanDiego, CA). Molecular biology reagents were purchased from NewEngland Biolabs (Beverly, MA), Promega (Madison, WI), andBoehringer Mannheim Biochemicals (Indianapolis, IN). Reagentgrade chemicals were obtained from BioRad (Hercules, CA), FisherScientific (Pittsburg, PA) and Sigma (St Louis, MO).
Total RNA isolation and reverse transcriptase-polymerasechain reaction (RT-PCR) analysisTissues from Sprague Dawley rats were harvested and immediatelyfrozen in liquid nitrogen. RNA samples were prepared byhomogenizing the rat tissue or cell lines in buffer containing 6 Mguanidine isothiocyanate, 5 mM sodium citrate pH 7.0, 0.1 Mmercaptoethanol, 0.5% sarcosyl, using a Potter-Elvehjemhomogenizer. The total RNA was isolated by centrifugation througha cesium chloride gradient (Sambrook, 1989). First-strand cDNA wassynthesized from 10 µg of total RNA using AMV reverse transcriptaseand oligo d(T) as primer. PCR amplification was performed using
B. Quiñones and others

4293Characterization of syntaxin 2 variants
three specific sequences: primer #1 hybridized to a sequence presentin the coding region of all syntaxin 2 variants (nucleotides 636-665of GenBank #L20823); primer #2 hybridized with the sequence thatcodes for the carboxyl-terminal domain of syntaxin 2C and is alsopresent in the 3′-untranslated region of the other syntaxin 2 variants(nucleotides 794-812 of GenBank #L20888); primer #3 hybridized tothe sequence that codes for the carboxyl-terminal domain of syntaxin2A and is present in the 3′-untranslated region of syntaxin 2D(nucleotides 819-840 of GenBank #L20823). PCR-amplifiedfragments were resolved by electrophoresis on a 2.5% agarose gel anddetected by ethidium bromide staining.
Recombinant adenovirus constructionVectors encoding syntaxin 2 variants were constructed by insertingeach cDNA downstream of a tetracycline operator and minimal CMVpromoter in the pAdlox/Tet shuttle plasmid (Hardy et al., 1997). Therecombinant adenoviruses containing each syntaxin 2 variant cDNAunder control of the tetracycline-controlled transactivator (tTA) wereproduced as described elsewhere (Hardy et al., 1997). Additionally, arecombinant adenovirus containing the cDNA for the tTA wasconstructed as previously described (Jin et al., 1998).
Cell culture, adenovirus infection and cDNA transfectionHela cells stably expressing the chimeric tTA (Hela-tTA) (Gossen andBujard, 1992) were kindly provided by Drs M. Gossen and H. Bujard(Zentrum für Molekulare Biologie, Heidelberg, Germany). Cos7 cellsstably expressing human α-2,6-sialyltransferase (Cos7-ST) or ratcellubrevin (Cos7-Cb) with lumenal human IgG (Fc domain) epitopetags were a kind gift of Drs K. Teter and H.-P. Moore (University ofCA, Berkeley, CA). The cDNA encoding cellubrevin was PCR-amplified from an RBL cell library while the sialyltransferase cDNAwas provided by Brian Seed (Massachusetts General Hospital,Boston, MA). Caco-2 cells were cultured as previously described(Riento et al., 1998). The remaining cell types were cultured at 37°Cin culture medium containing 100 U/ml penicillin and 100 µg/mlstreptomycin under the following conditions. For NRK, Hela-tTA,Cos7, Cos7-ST and Cos7-Cb cells: DMEM supplemented with 10%FCS under 5% CO2/95% air; MDCK strain II cells: MEM-alphamodification medium supplemented with 10% FCS under 5%CO2/95% air; AtT20-NIH cells: DMEM supplemented with 10% FCSunder 15% CO2/85% air; PC12 cells: DMEM supplemented with 5%FCS and 5% HS under 10% CO2/90% air. Infections withrecombinant syntaxin 2 adenoviruses were carried out in Hela-tTAcells at a multiplicity of infection (MOI) of 5-10 plaque-forming units(PFU) per cell. MDCK, Cos7, Cos7-ST and Cos7-Cb cells wereinfected with adenoviruses encoding tTA and either syntaxin 2A or2B each at an MOI of 100 PFU/cell. Adenovirus infections inpolarized MDCK cells that were plated on Transwell polycarbonatefilters were performed as previously described (Altschuler et al.,1998). Trypsinized Caco-2 cells that were suspended in the completegrowth medium were transfected with plasmids containing the cDNAsencoding either syntaxins 2A or 2B using the Superfect transfectionreagent (Qiagen, Chatsworth, CA), according to the manufacturer’sinstructions. The cell suspension was plated on Transwellpolycarbonate filters and analyzed after 4 days.
Subcellular fractionationTissues were excised from Sprague Dawley rats and immediatelyhomogenized in 0.32 M sucrose, 10 mM Hepes-KOH, pH 7.5, 1 mMEGTA, 0.1 mM EDTA and 0.3 mM PMSF, using a Potter-Elvehjemhomogenizer. Hela-tTA cells, 15 hours after infection withrecombinant syntaxin 2 adenoviruses, were rinsed twice withphosphate-buffered saline (PBS), trypsinized with 0.05% trypsin-EDTA solution, harvested by centrifugation, and resuspended in 10mM Hepes-KOH, pH 7.4, 5 mM EDTA, 2 µg/ml pepstatin, 10 µg/mlleupectin, 0.3 mM PMSF. Cells were lysed by passing the suspension6 times through a 23-gauge syringe needle. Homogenates from
infected Hela-tTA and rat tissues were centrifuged at 5000 g for 10minutes to obtain a postnuclear supernatant that was furthercentrifuged at 100,000 g for 1 hour at 4°C to obtain membrane andsoluble fractions. Fractions from rat tissues (100 µg each) weresolubilized in SDS-PAGE sample buffer (50 mM Tris-HCl pH 6.8, 5%glycerol, 1.5% SDS, 0.02% bromophenol blue). Fractions from Hela-tTA extracts (1 mg total) were solubilized in 1% NP-40, 0.1% bovineserum albumin, 0.1% gelatin and immunoprecipitated with anti-syntaxin 2 antibody coupled to protein A-Sepharose beads for 2 hoursat 4°C. After three washes in 50 mM Tris, pH 8, 150 mM NaCl, 1%Nonidet P-40, 0.5% deoxycholate, 0.1% SDS, the immunoprecipitatewas eluted in SDS-PAGE sample buffer. Rat tissue and Hela-tTAsamples were resolved on a 12.5% polyacrylamide gel according tothe method of Laemmli (1970).
Immunofluorescence microscopyPrior to the immunostaining procedure, AtT20 cells were grown onlaminin-coated coverslips while NRK and adenovirus-infected Hela-tTA, Cos7, Cos7-ST, Cos7-Cb and subconfluent MDCK cells wereplated on untreated coverslips. PC12 cells were grown on poly-D-lysine-coated coverslips and treated with nerve growth factor (50ng/ml) for 24 hours prior to fixation. Coverslip-grown cells wereprocessed for immunofluorescence microscopy as previouslydescribed (Bucci et al., 1992), with the exception that the pre-fixationstep was omitted. Antibodies used were a fluorescein-conjugated anti-human IgG (diluted 1:50) and/or rabbit anti-syntaxin 2 (diluted 1:200).Polarized, filter-grown MDCK and Caco-2 cells were processed aspreviously described (Low et al., 1998; Riento et al., 1998), with themodification that the blocking and antibody incubation steps in MDCKcells were performed in PBS containing 0.7% gelatin and 0.025%saponin. The primary antibodies used were rabbit anti-syntaxin 2(1:300) and, in some experiments, anti-ZO-1 antibody (for MDCKcells, rat mAb diluted 1:100; for Caco-2 cells, mouse mAb diluted1:300). Endogenous syntaxin 2 immunoreactivity in NRK, AtT20 andPC12 cells was detected with an HRP-conjugated secondary antibodyusing a fluorescent signal amplification system (ReinassanceTSA-Direct reagent; NEN Life Science Products, Boston, MA). In all otherexperiments, syntaxin 2 immunoreactivity was detected with afluorophore-conjugated secondary antibody. All samples weremounted on slides in an anti-bleach medium (Kirkegaard and PerryLaboratories, Gaithersburg, MD) and observed by confocalmicroscopy. Caco-2 cells were examined with a Leica TCS NTconfocal microscope (Riento et al., 1998) while all other cell lines wereexamined by using a Bio-Rad MRC1000 confocal microscope aspreviously described (Teter et al., 1998). Horizontal optical sectionsseparated by 0.36 µm were collected sequentially for each fluorophore.
Preparation of bacterially expressed proteinsFull-length sequences encoding syntaxins 2A (amino acids 1-289), 2B(amino acids 1-288) and 2D (amino acids 1-237) were inserted intothe vector pGEX-KG (Guan and Dixon, 1991) for expression in E.coli as glutathione S-transferase (GST) fusion proteins. A constructencoding VAMP2 with an amino-terminal FLAG-tag was generatedby insertion of an oligonucleotide coding for the FLAG epitope(DYKDDDDK) into the BamHI site of a previously described pGEX-KG plasmid encoding the cytoplasmic domain of VAMP2 (aminoacids 1-94) (Hao et al., 1997). Previously described pGEX-KGplasmids encoding syntaxin 2C (amino acids 4-264), n-sec1, SNAP-25 and SNAP-23 were also used (Calakos et al., 1994; Hao et al.,1997; Pevsner et al., 1994a; Risinger and Bennett, 1999). GST fusionproteins were expressed in the AB1899 strain of E. coli and purifiedon glutathione-agarose beads as described previously (Hao et al.,1997; Poirier et al., 1998a). For recovery of the recombinant proteinsin a soluble form, the immobilized GST-fusion proteins weresubjected to thrombin cleavage. Protein concentrations were estimatedby Coomassie Blue staining of protein bands after SDS-PAGE withbovine serum albumin as a standard.

4294
In vitro binding assaysFor in vitro binding assays, GST-fusion proteins (syntaxin 2 variants,SNAP-23 or SNAP-25) bound to glutathione-agarose beads wereincubated with soluble proteins (n-sec1, SNAP-23, SNAP-25 orFLAG-VAMP2) for 2 hours at 4°C in 50 mM Tris-HCl, pH 8.0, 150mM NaCl, 2.5 mM CaCl2, 0.5% Triton X-100, 0.1% bovine serumalbumin. The beads were then washed three times and proteins boundto the beads were recovered by boiling in SDS-PAGE sample buffer.The samples were then subjected to SDS-PAGE and immunoblottingfor specific detection of bound soluble proteins.
ImmunoblottingImmunoblotting was used to detect syntaxin 2 variants expressed inrat tissues and adenovirus-infected Hela-tTA cells, and to monitor theoutcome of in vitro binding assays. After SDS-PAGE, proteins weretransferred to Protran nitrocellulose filters (Schleicher and Schuell,Keene, NH) and the filters stained with Ponceau S. Syntaxin 2 variantswere detected by probing the blots with an anti-syntaxin 2 antibody(1:1000). To minimize cross-reaction with syntaxin 1 (Gaisano et al.,1996), the syntaxin 2 antibody was pre-adsorbed with immobilizedGST-syntaxin 1A. This treatment effectively eliminated all cross-reaction with syntaxin 1A on immunoblots (data not shown). FLAG-VAMP2 was detected with a mouse monoclonal against theFLAG epitope (1:1000). Immunoreactive proteins were detectedby incubation with either HRP-conjugated secondary antibodies
followed by chemiluminescence visualization (NEN Life ScienceProducts, Boston, MA) or, for quantitative analysis, 125I-conjugatedsecondary antibodies followed by phosphorimaging (MolecularDynamics, Sunnyvale, CA).
RESULTS
Syntaxin 2 splice variantsIn the original isolation of syntaxin 2 cDNAs (Bennett et al.,1993), four variants were described that encode proteins thatare identical throughout much of their amino-terminalcytoplasmic domains but differ in their carboxyl-terminalsequences (Fig. 1A). Three of the four syntaxin 2 variants (2,2′ and 2′′ ; now designated 2A, 2B and 2C, respectively, forconsistency with the naming of other syntaxin isoformvariants) are identical through amino acid 264 but then havedivergent carboxyl-terminal domains: syntaxins 2A and 2Bterminate with distinct 22-24 amino acid hydrophobicsequences while syntaxin 2C has a 10-amino-acid, relativelyhydrophilic, carboxyl-terminal sequence. The fourth syntaxin2 variant, here named syntaxin 2D, was described as beingidentical to syntaxin 2A with the exception of a 27-base-pair
B. Quiñones and others
Fig. 1. RT/PCR analysis of syntaxin 2 variantexpression patterns. (A) Schematic diagramillustrating the organization of syntaxin 2 transcripts.The position/orientation of PCR primer hybridizationsites and the expected sizes of PCR products aredepicted at the top and right, respectively. Filled boxesindicate the sequences coding for the unique carboxyl-terminal domains of syntaxins 2A (vertical lines;sequence: KKWIIAAVVVAVIAVLALIIGLSVGK),2B (crosshatched; sequence:KVMFVLICVVTLLVILGIILATALS), 2C (diagonallines; sequence: GVLCALGRQC) and 2D (horizontallines; sequence: TLSPPGRDPG). The positions of thestop codon for each variant are marked by theasterisks. (B) Expression patterns of syntaxin 2variants among different tissues and cell lines.RT/PCR was performed as described in Materials andMethods on total RNA isolated from the indicated rattissues (left) and cell lines (right). PCR-amplifiedfragments were resolved on a 2.5% agarose gel andvisualized by staining with ethidium bromide.Products generated with primer set [1+2] (upper gels)were used to monitor the presence of syntaxin 2B and2C transcripts, while products generated with primerset [1+3] (lower gels) were used to monitor thepresence of syntaxin 2A and 2D transcripts. Thepositions of DNA size standards run on parallel lanesof the same gels are indicated.
4295Characterization of syntaxin 2 variants
insert (coding for 9 amino acids) between the codons encodingamino acids 227 and 228 (Bennett et al., 1993). However,resequencing the syntaxin 2D cDNA and independent isolatesthat were amplified by PCR (see below) indicated that syntaxin2D differs from syntaxin 2A by a 28-base-pair insert. Thisinsert generates a frameshift in the downstream sequence suchthat a stop codon is encountered soon after the insert. Thus, thesyntaxin 2D cDNA encodes a truncated protein of 237 aminoacids (compared with 274-289 for the other syntaxin 2variants) with a distinct hydrophilic carboxyl-terminal domain.The truncated nature of syntaxin 2D was confirmed byexpression of recombinant forms of each variant (see Fig. 7).
Syntaxin 2 variant expression patternsAlthough syntaxin 2 genomic clones have not beencharacterized, the most likely explanation for the generation ofthe syntaxin 2 variants is alternative splicing. Indeed, thesequences encoding the distinct carboxyl-terminal domains ofsyntaxins 2B and 2C are found in the 3′-untranslated region ofsyntaxin 2A (Fig. 1A). To extend a previous northern blotanalysis of syntaxin 2 expression which did not discriminatebetween splice variants (Bennett et al., 1993), and toinvestigate the potential differential regulation of splicing, wehave performed RT-PCR analysis of syntaxin 2 varianttranscript expression. Two primer pairs were utilized (Fig. 1A):primer pair [1+2] resulted in the amplification of products fromall four syntaxin isoforms which, due to limited gel resolution,were separated into bands corresponding to syntaxins 2A/2D,2B and 2C (Fig. 1B); primer pair [1+3] yielded products foronly syntaxins 2A and 2D but whose 28-base-pair differencein size was more readily resolved (Fig. 1B). RT-PCR analysisof a variety of rat tissues demonstrated that transcriptsencoding syntaxins 2A, 2B and 2C are all broadly but notuniformly expressed. In contrast, the syntaxin 2D transcriptwas only detected in brain. To examine the expression patternsin individual cell types, the RT-PCR procedure was performedon three cell lines: AR42J, AtT20 and PC12 cells. Syntaxin 2Awas expressed in each of the cell lines assayed. In contrast,syntaxin 2B was expressed in AtT20 cells but not in PC12 orAR42J cells while syntaxin 2C was expressed in PC12 andAR42J but not in AtT20 cells. Together, these findings indicatethat the broad but variable expression of syntaxins 2B and 2Cat the tissue level may result from heterogeneous expressionamong the cell types that comprise each tissue and that thesyntaxin 2 variants may contribute to both general andspecialized membrane-trafficking pathways.
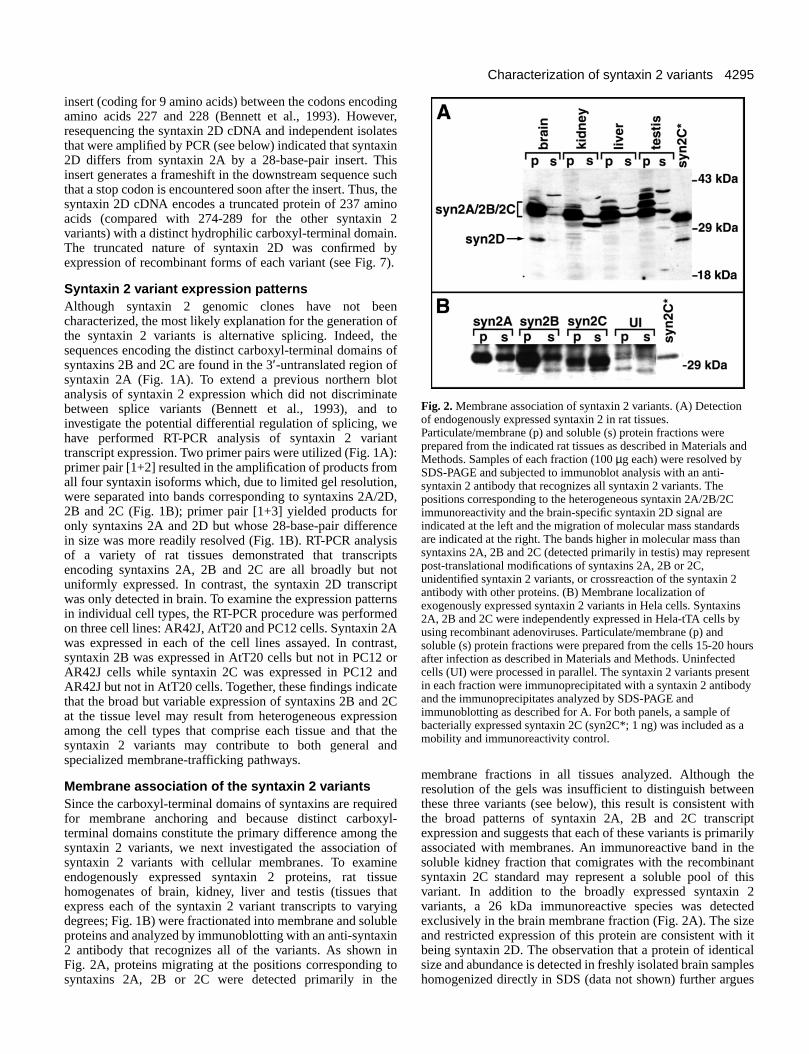
Membrane association of the syntaxin 2 variantsSince the carboxyl-terminal domains of syntaxins are requiredfor membrane anchoring and because distinct carboxyl-terminal domains constitute the primary difference among thesyntaxin 2 variants, we next investigated the association ofsyntaxin 2 variants with cellular membranes. To examineendogenously expressed syntaxin 2 proteins, rat tissuehomogenates of brain, kidney, liver and testis (tissues thatexpress each of the syntaxin 2 variant transcripts to varyingdegrees; Fig. 1B) were fractionated into membrane and solubleproteins and analyzed by immunoblotting with an anti-syntaxin2 antibody that recognizes all of the variants. As shown inFig. 2A, proteins migrating at the positions corresponding tosyntaxins 2A, 2B or 2C were detected primarily in the
membrane fractions in all tissues analyzed. Although theresolution of the gels was insufficient to distinguish betweenthese three variants (see below), this result is consistent withthe broad patterns of syntaxin 2A, 2B and 2C transcriptexpression and suggests that each of these variants is primarilyassociated with membranes. An immunoreactive band in thesoluble kidney fraction that comigrates with the recombinantsyntaxin 2C standard may represent a soluble pool of thisvariant. In addition to the broadly expressed syntaxin 2variants, a 26 kDa immunoreactive species was detectedexclusively in the brain membrane fraction (Fig. 2A). The sizeand restricted expression of this protein are consistent with itbeing syntaxin 2D. The observation that a protein of identicalsize and abundance is detected in freshly isolated brain sampleshomogenized directly in SDS (data not shown) further argues
Fig. 2. Membrane association of syntaxin 2 variants. (A) Detectionof endogenously expressed syntaxin 2 in rat tissues.Particulate/membrane (p) and soluble (s) protein fractions wereprepared from the indicated rat tissues as described in Materials andMethods. Samples of each fraction (100 µg each) were resolved bySDS-PAGE and subjected to immunoblot analysis with an anti-syntaxin 2 antibody that recognizes all syntaxin 2 variants. Thepositions corresponding to the heterogeneous syntaxin 2A/2B/2Cimmunoreactivity and the brain-specific syntaxin 2D signal areindicated at the left and the migration of molecular mass standardsare indicated at the right. The bands higher in molecular mass thansyntaxins 2A, 2B and 2C (detected primarily in testis) may representpost-translational modifications of syntaxins 2A, 2B or 2C,unidentified syntaxin 2 variants, or crossreaction of the syntaxin 2antibody with other proteins. (B) Membrane localization ofexogenously expressed syntaxin 2 variants in Hela cells. Syntaxins2A, 2B and 2C were independently expressed in Hela-tTA cells byusing recombinant adenoviruses. Particulate/membrane (p) andsoluble (s) protein fractions were prepared from the cells 15-20 hoursafter infection as described in Materials and Methods. Uninfectedcells (UI) were processed in parallel. The syntaxin 2 variants presentin each fraction were immunoprecipitated with a syntaxin 2 antibodyand the immunoprecipitates analyzed by SDS-PAGE andimmunoblotting as described for A. For both panels, a sample ofbacterially expressed syntaxin 2C (syn2C*; 1 ng) was included as amobility and immunoreactivity control.
4296
that this species does not correspond to a brain-specificproteolytic fragment derived from syntaxins 2A, 2B or 2C.
Because syntaxins 2A, 2B and 2C exhibit similarelectrophoretic mobilities and reactivities with availableantibodies, we were unable to individually characterize themembrane associations of these variants in rat tissues.Therefore, we examined the membrane associations ofsyntaxins 2A, 2B and 2C following heterologous expression inHela-tTA cells using recombinant adenoviruses (see Materialsand Methods). As shown in Fig. 2B, syntaxins 2A and 2Bfractionated primarily with membrane proteins while syntaxin2C was more abundant in the soluble protein fraction.However, a significant fraction of the syntaxin 2C was alsoassociated with the membrane protein fraction. In contrast tothe other syntaxin 2 variants, syntaxin 2D expressed in Hela-tTA cells fractionated exclusively with soluble proteins (datanot shown). The portion of syntaxins 2A, 2B and 2C associatedwith the membrane protein fraction was not altered bytreatment with NaCl (1.5 M) or urea (2 M) (data not shown).This observation indicates that syntaxins 2A and 2B behave asintegral membrane proteins, asexpected from their primary sequence,and that some syntaxin 2C istightly membrane-associated. Therecovery of a portion of theexogenously expressed syntaxin 2immunoreactivity in the solublefraction could be a consequence ofoverexpression. Alternatively, therecould be cell-type differences in thelevel of membrane association orfractionation characteristics comparedto the tissues used for analysis of theendogenous syntaxin 2.
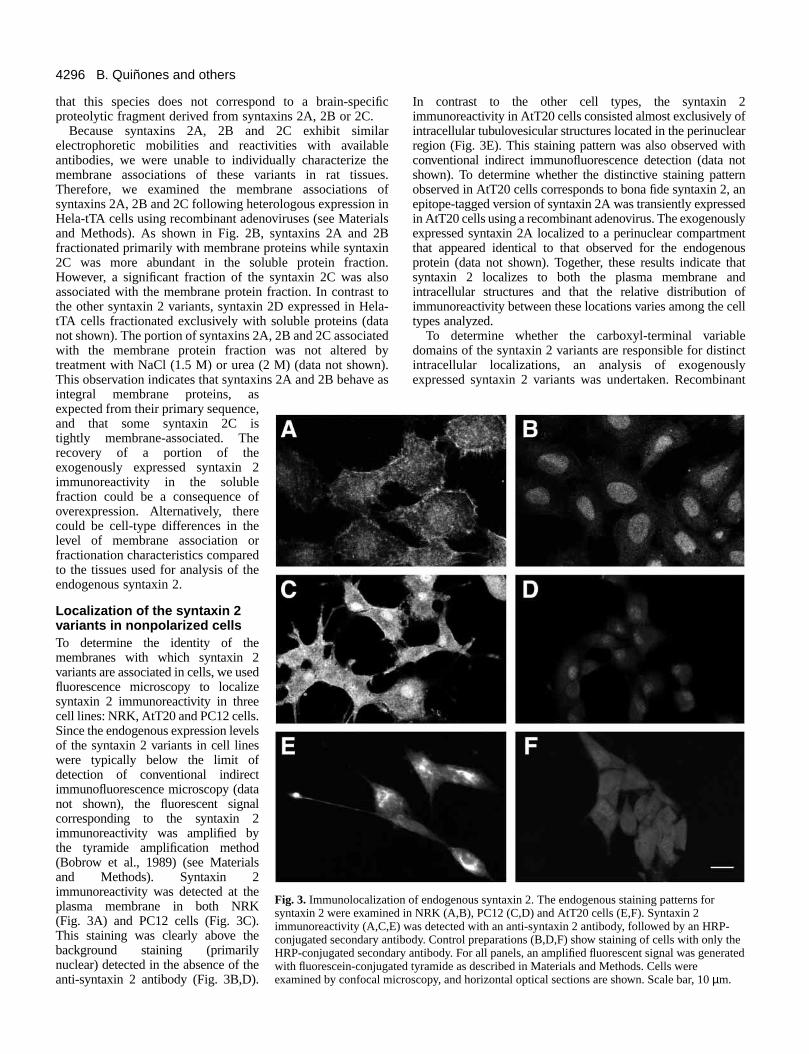
Localization of the syntaxin 2variants in nonpolarized cellsTo determine the identity of themembranes with which syntaxin 2variants are associated in cells, we usedfluorescence microscopy to localizesyntaxin 2 immunoreactivity in threecell lines: NRK, AtT20 and PC12 cells.Since the endogenous expression levelsof the syntaxin 2 variants in cell lineswere typically below the limit ofdetection of conventional indirectimmunofluorescence microscopy (datanot shown), the fluorescent signalcorresponding to the syntaxin 2immunoreactivity was amplified bythe tyramide amplification method(Bobrow et al., 1989) (see Materialsand Methods). Syntaxin 2immunoreactivity was detected at theplasma membrane in both NRK(Fig. 3A) and PC12 cells (Fig. 3C).This staining was clearly above thebackground staining (primarilynuclear) detected in the absence of theanti-syntaxin 2 antibody (Fig. 3B,D).
In contrast to the other cell types, the syntaxin 2immunoreactivity in AtT20 cells consisted almost exclusively ofintracellular tubulovesicular structures located in the perinuclearregion (Fig. 3E). This staining pattern was also observed withconventional indirect immunofluorescence detection (data notshown). To determine whether the distinctive staining patternobserved in AtT20 cells corresponds to bona fide syntaxin 2, anepitope-tagged version of syntaxin 2A was transiently expressedin AtT20 cells using a recombinant adenovirus. The exogenouslyexpressed syntaxin 2A localized to a perinuclear compartmentthat appeared identical to that observed for the endogenousprotein (data not shown). Together, these results indicate thatsyntaxin 2 localizes to both the plasma membrane andintracellular structures and that the relative distribution ofimmunoreactivity between these locations varies among the celltypes analyzed.
To determine whether the carboxyl-terminal variabledomains of the syntaxin 2 variants are responsible for distinctintracellular localizations, an analysis of exogenouslyexpressed syntaxin 2 variants was undertaken. Recombinant
B. Quiñones and others
Fig. 3. Immunolocalization of endogenous syntaxin 2. The endogenous staining patterns forsyntaxin 2 were examined in NRK (A,B), PC12 (C,D) and AtT20 cells (E,F). Syntaxin 2immunoreactivity (A,C,E) was detected with an anti-syntaxin 2 antibody, followed by an HRP-conjugated secondary antibody. Control preparations (B,D,F) show staining of cells with only theHRP-conjugated secondary antibody. For all panels, an amplified fluorescent signal was generatedwith fluorescein-conjugated tyramide as described in Materials and Methods. Cells wereexamined by confocal microscopy, and horizontal optical sections are shown. Scale bar, 10 µm.
4297Characterization of syntaxin 2 variants
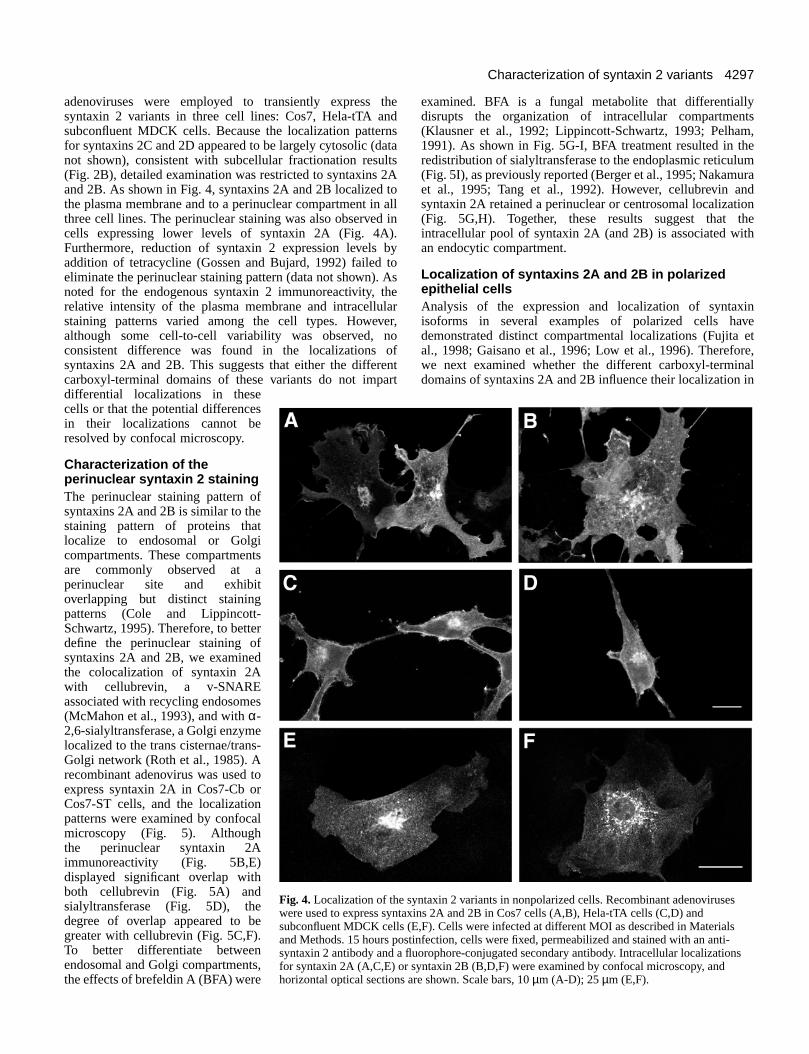
adenoviruses were employed to transiently express thesyntaxin 2 variants in three cell lines: Cos7, Hela-tTA andsubconfluent MDCK cells. Because the localization patternsfor syntaxins 2C and 2D appeared to be largely cytosolic (datanot shown), consistent with subcellular fractionation results(Fig. 2B), detailed examination was restricted to syntaxins 2Aand 2B. As shown in Fig. 4, syntaxins 2A and 2B localized tothe plasma membrane and to a perinuclear compartment in allthree cell lines. The perinuclear staining was also observed incells expressing lower levels of syntaxin 2A (Fig. 4A).Furthermore, reduction of syntaxin 2 expression levels byaddition of tetracycline (Gossen and Bujard, 1992) failed toeliminate the perinuclear staining pattern (data not shown). Asnoted for the endogenous syntaxin 2 immunoreactivity, therelative intensity of the plasma membrane and intracellularstaining patterns varied among the cell types. However,although some cell-to-cell variability was observed, noconsistent difference was found in the localizations ofsyntaxins 2A and 2B. This suggests that either the differentcarboxyl-terminal domains of these variants do not impartdifferential localizations in thesecells or that the potential differencesin their localizations cannot beresolved by confocal microscopy.
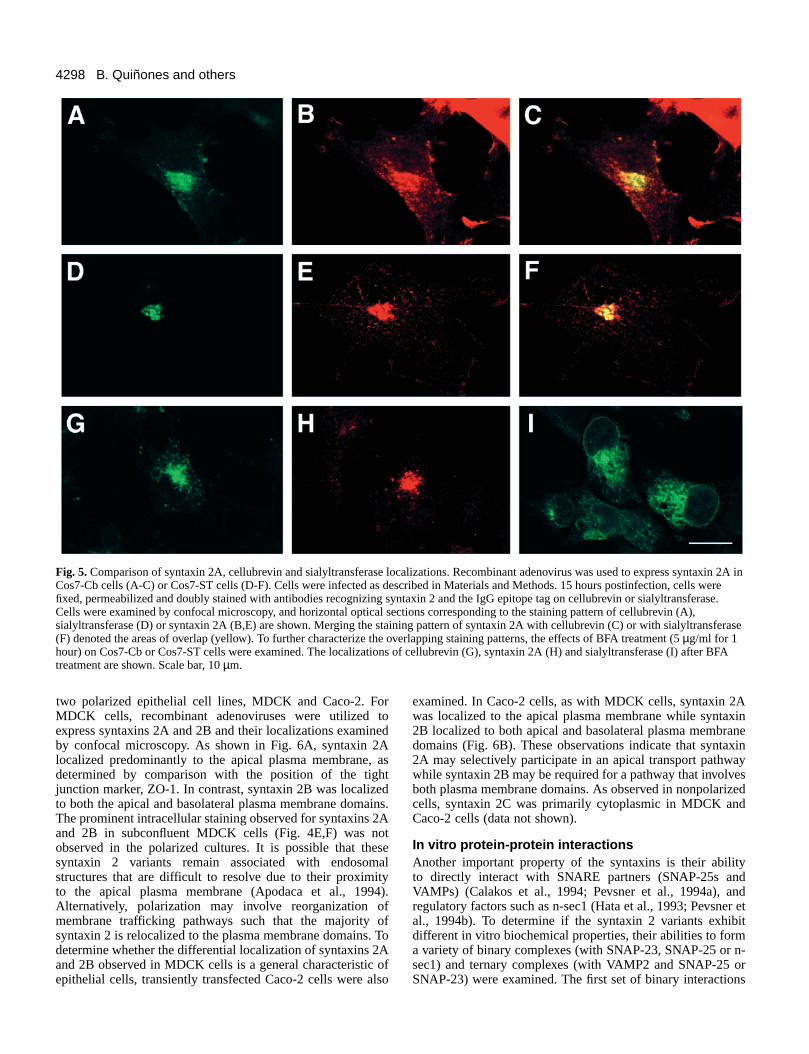
Characterization of theperinuclear syntaxin 2 stainingThe perinuclear staining pattern ofsyntaxins 2A and 2B is similar to thestaining pattern of proteins thatlocalize to endosomal or Golgicompartments. These compartmentsare commonly observed at aperinuclear site and exhibitoverlapping but distinct stainingpatterns (Cole and Lippincott-Schwartz, 1995). Therefore, to betterdefine the perinuclear staining ofsyntaxins 2A and 2B, we examinedthe colocalization of syntaxin 2Awith cellubrevin, a v-SNAREassociated with recycling endosomes(McMahon et al., 1993), and with α-2,6-sialyltransferase, a Golgi enzymelocalized to the trans cisternae/trans-Golgi network (Roth et al., 1985). Arecombinant adenovirus was used toexpress syntaxin 2A in Cos7-Cb orCos7-ST cells, and the localizationpatterns were examined by confocalmicroscopy (Fig. 5). Althoughthe perinuclear syntaxin 2Aimmunoreactivity (Fig. 5B,E)displayed significant overlap withboth cellubrevin (Fig. 5A) andsialyltransferase (Fig. 5D), thedegree of overlap appeared to begreater with cellubrevin (Fig. 5C,F).To better differentiate betweenendosomal and Golgi compartments,the effects of brefeldin A (BFA) were
examined. BFA is a fungal metabolite that differentiallydisrupts the organization of intracellular compartments(Klausner et al., 1992; Lippincott-Schwartz, 1993; Pelham,1991). As shown in Fig. 5G-I, BFA treatment resulted in theredistribution of sialyltransferase to the endoplasmic reticulum(Fig. 5I), as previously reported (Berger et al., 1995; Nakamuraet al., 1995; Tang et al., 1992). However, cellubrevin andsyntaxin 2A retained a perinuclear or centrosomal localization(Fig. 5G,H). Together, these results suggest that theintracellular pool of syntaxin 2A (and 2B) is associated withan endocytic compartment.
Localization of syntaxins 2A and 2B in polarizedepithelial cellsAnalysis of the expression and localization of syntaxinisoforms in several examples of polarized cells havedemonstrated distinct compartmental localizations (Fujita etal., 1998; Gaisano et al., 1996; Low et al., 1996). Therefore,we next examined whether the different carboxyl-terminaldomains of syntaxins 2A and 2B influence their localization in
Fig. 4. Localization of the syntaxin 2 variants in nonpolarized cells. Recombinant adenoviruseswere used to express syntaxins 2A and 2B in Cos7 cells (A,B), Hela-tTA cells (C,D) andsubconfluent MDCK cells (E,F). Cells were infected at different MOI as described in Materialsand Methods. 15 hours postinfection, cells were fixed, permeabilized and stained with an anti-syntaxin 2 antibody and a fluorophore-conjugated secondary antibody. Intracellular localizationsfor syntaxin 2A (A,C,E) or syntaxin 2B (B,D,F) were examined by confocal microscopy, andhorizontal optical sections are shown. Scale bars, 10 µm (A-D); 25 µm (E,F).
4298
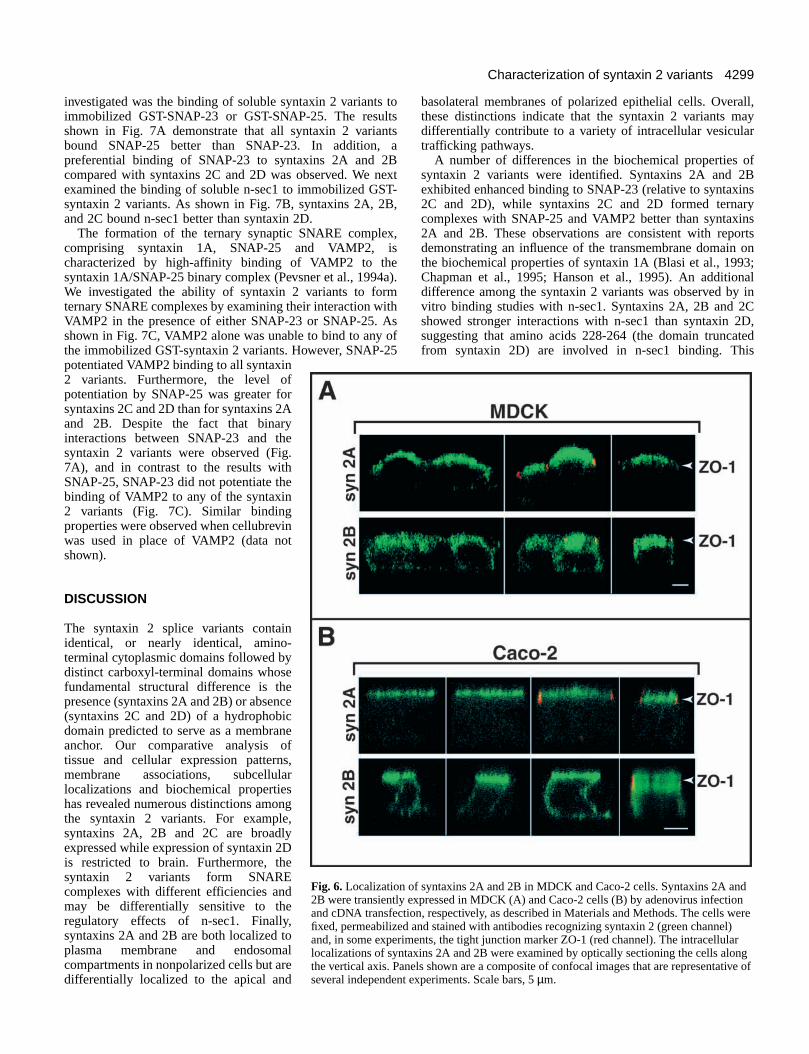
two polarized epithelial cell lines, MDCK and Caco-2. ForMDCK cells, recombinant adenoviruses were utilized toexpress syntaxins 2A and 2B and their localizations examinedby confocal microscopy. As shown in Fig. 6A, syntaxin 2Alocalized predominantly to the apical plasma membrane, asdetermined by comparison with the position of the tightjunction marker, ZO-1. In contrast, syntaxin 2B was localizedto both the apical and basolateral plasma membrane domains.The prominent intracellular staining observed for syntaxins 2Aand 2B in subconfluent MDCK cells (Fig. 4E,F) was notobserved in the polarized cultures. It is possible that thesesyntaxin 2 variants remain associated with endosomalstructures that are difficult to resolve due to their proximityto the apical plasma membrane (Apodaca et al., 1994).Alternatively, polarization may involve reorganization ofmembrane trafficking pathways such that the majority ofsyntaxin 2 is relocalized to the plasma membrane domains. Todetermine whether the differential localization of syntaxins 2Aand 2B observed in MDCK cells is a general characteristic ofepithelial cells, transiently transfected Caco-2 cells were also
examined. In Caco-2 cells, as with MDCK cells, syntaxin 2Awas localized to the apical plasma membrane while syntaxin2B localized to both apical and basolateral plasma membranedomains (Fig. 6B). These observations indicate that syntaxin2A may selectively participate in an apical transport pathwaywhile syntaxin 2B may be required for a pathway that involvesboth plasma membrane domains. As observed in nonpolarizedcells, syntaxin 2C was primarily cytoplasmic in MDCK andCaco-2 cells (data not shown).
In vitro protein-protein interactionsAnother important property of the syntaxins is their abilityto directly interact with SNARE partners (SNAP-25s andVAMPs) (Calakos et al., 1994; Pevsner et al., 1994a), andregulatory factors such as n-sec1 (Hata et al., 1993; Pevsner etal., 1994b). To determine if the syntaxin 2 variants exhibitdifferent in vitro biochemical properties, their abilities to forma variety of binary complexes (with SNAP-23, SNAP-25 or n-sec1) and ternary complexes (with VAMP2 and SNAP-25 orSNAP-23) were examined. The first set of binary interactions
B. Quiñones and others
Fig. 5. Comparison of syntaxin 2A, cellubrevin and sialyltransferase localizations. Recombinant adenovirus was used to express syntaxin 2A inCos7-Cb cells (A-C) or Cos7-ST cells (D-F). Cells were infected as described in Materials and Methods. 15 hours postinfection, cells werefixed, permeabilized and doubly stained with antibodies recognizing syntaxin 2 and the IgG epitope tag on cellubrevin or sialyltransferase.Cells were examined by confocal microscopy, and horizontal optical sections corresponding to the staining pattern of cellubrevin (A),sialyltransferase (D) or syntaxin 2A (B,E) are shown. Merging the staining pattern of syntaxin 2A with cellubrevin (C) or with sialyltransferase(F) denoted the areas of overlap (yellow). To further characterize the overlapping staining patterns, the effects of BFA treatment (5 µg/ml for 1hour) on Cos7-Cb or Cos7-ST cells were examined. The localizations of cellubrevin (G), syntaxin 2A (H) and sialyltransferase (I) after BFAtreatment are shown. Scale bar, 10 µm.
4299Characterization of syntaxin 2 variants
investigated was the binding of soluble syntaxin 2 variants toimmobilized GST-SNAP-23 or GST-SNAP-25. The resultsshown in Fig. 7A demonstrate that all syntaxin 2 variantsbound SNAP-25 better than SNAP-23. In addition, apreferential binding of SNAP-23 to syntaxins 2A and 2Bcompared with syntaxins 2C and 2D was observed. We nextexamined the binding of soluble n-sec1 to immobilized GST-syntaxin 2 variants. As shown in Fig. 7B, syntaxins 2A, 2B,and 2C bound n-sec1 better than syntaxin 2D.
The formation of the ternary synaptic SNARE complex,comprising syntaxin 1A, SNAP-25 and VAMP2, ischaracterized by high-affinity binding of VAMP2 to thesyntaxin 1A/SNAP-25 binary complex (Pevsner et al., 1994a).We investigated the ability of syntaxin 2 variants to formternary SNARE complexes by examining their interaction withVAMP2 in the presence of either SNAP-23 or SNAP-25. Asshown in Fig. 7C, VAMP2 alone was unable to bind to any ofthe immobilized GST-syntaxin 2 variants. However, SNAP-25potentiated VAMP2 binding to all syntaxin2 variants. Furthermore, the level ofpotentiation by SNAP-25 was greater forsyntaxins 2C and 2D than for syntaxins 2Aand 2B. Despite the fact that binaryinteractions between SNAP-23 and thesyntaxin 2 variants were observed (Fig.7A), and in contrast to the results withSNAP-25, SNAP-23 did not potentiate thebinding of VAMP2 to any of the syntaxin2 variants (Fig. 7C). Similar bindingproperties were observed when cellubrevinwas used in place of VAMP2 (data notshown).
DISCUSSION
The syntaxin 2 splice variants containidentical, or nearly identical, amino-terminal cytoplasmic domains followed bydistinct carboxyl-terminal domains whosefundamental structural difference is thepresence (syntaxins 2A and 2B) or absence(syntaxins 2C and 2D) of a hydrophobicdomain predicted to serve as a membraneanchor. Our comparative analysis oftissue and cellular expression patterns,membrane associations, subcellularlocalizations and biochemical propertieshas revealed numerous distinctions amongthe syntaxin 2 variants. For example,syntaxins 2A, 2B and 2C are broadlyexpressed while expression of syntaxin 2Dis restricted to brain. Furthermore, thesyntaxin 2 variants form SNAREcomplexes with different efficiencies andmay be differentially sensitive to theregulatory effects of n-sec1. Finally,syntaxins 2A and 2B are both localized toplasma membrane and endosomalcompartments in nonpolarized cells but aredifferentially localized to the apical and
basolateral membranes of polarized epithelial cells. Overall,these distinctions indicate that the syntaxin 2 variants maydifferentially contribute to a variety of intracellular vesiculartrafficking pathways.
A number of differences in the biochemical properties ofsyntaxin 2 variants were identified. Syntaxins 2A and 2Bexhibited enhanced binding to SNAP-23 (relative to syntaxins2C and 2D), while syntaxins 2C and 2D formed ternarycomplexes with SNAP-25 and VAMP2 better than syntaxins2A and 2B. These observations are consistent with reportsdemonstrating an influence of the transmembrane domain onthe biochemical properties of syntaxin 1A (Blasi et al., 1993;Chapman et al., 1995; Hanson et al., 1995). An additionaldifference among the syntaxin 2 variants was observed by invitro binding studies with n-sec1. Syntaxins 2A, 2B and 2Cshowed stronger interactions with n-sec1 than syntaxin 2D,suggesting that amino acids 228-264 (the domain truncatedfrom syntaxin 2D) are involved in n-sec1 binding. This
Fig. 6. Localization of syntaxins 2A and 2B in MDCK and Caco-2 cells. Syntaxins 2A and2B were transiently expressed in MDCK (A) and Caco-2 cells (B) by adenovirus infectionand cDNA transfection, respectively, as described in Materials and Methods. The cells werefixed, permeabilized and stained with antibodies recognizing syntaxin 2 (green channel)and, in some experiments, the tight junction marker ZO-1 (red channel). The intracellularlocalizations of syntaxins 2A and 2B were examined by optically sectioning the cells alongthe vertical axis. Panels shown are a composite of confocal images that are representative ofseveral independent experiments. Scale bars, 5 µm.

4300
observation is consistent with previous studies demonstratingthat most of the cytoplasmic domain of syntaxin 1A is requiredfor n-sec1 binding (Hata et al., 1993; Hata and Sudhof, 1995;Kee et al., 1995; Pevsner et al., 1994b). Remarkably, n-sec1exhibited threefold greater binding to syntaxins 2A, 2B and 2Cthan to syntaxin 1A (data not shown). A preferential bindingof n-sec1 with syntaxin 2 has also been observed by yeast two-hybrid analysis (Hata and Sudhof, 1995). It is possible that n-sec1 functions as a negative regulator of SNARE complexformation by promoting or stabilizing a conformation in whichsyntaxin’s amino- and carboxyl-terminal domains interact,thereby precluding interactions with other SNAREs (Calakoset al., 1994; Fernandez et al., 1998; Fiebig et al., 1999; Hansonet al., 1995; Pevsner et al., 1994a). The relative stability of sucha conformation of syntaxin 2 (as compared to a similarconformation in syntaxin 1A) could provide an explanation forboth the enhanced binding of syntaxin 2 to n-sec1 and the lackof binary interactions between syntaxin 2 and VAMP2.
A previous report documenting an inability of SNAP-25 topromote ternary complex formation with syntaxin 2C andVAMP2 (Pevsner et al., 1994a) is not consistent with ourfindings. Although the reason for this discrepancy remains tobe determined, one possibility is that the recombinant VAMP2used in the present study lacked a 26-amino-acid amino-terminal ‘spacer’ sequence (Calakos et al., 1994) and wasincluded at fivefold higher concentrations. In contrast toSNAP-25, SNAP-23 did not promote the formation of a ternary
complex with any syntaxin 2 variant in the presence ofVAMP2. Distinctions in the properties of SNAP-25 and SNAP-23 have also been noted in studies of SNARE complexassembly (Foster et al., 1998) and protein phosphorylation(Risinger and Bennett, 1999).
Subcellular fractionation studies demonstrated that themajority of each of the syntaxin 2 variants is membrane-associated in tissue homogenates. The membrane associationof syntaxins 2C and 2D (variants lacking hydrophobiccarboxyl-termini) could result from an interaction with adistinct membrane-anchored protein (perhaps another SNARE)or through post-translational lipid modification. In the case ofsyntaxin 2C, the following observations are consistent withlipid anchoring: (1) membrane-associated syntaxin 2C was notextracted from Hela-tTA membranes by high salt or urea,conditions expected to disrupt protein-protein interactions; (2)on optimally resolved gels, the mobility of membrane-anchored syntaxin 2C was found to be retarded relative to thesoluble form (data not shown); and (3) the unique 9-amino-acid carboxyl-terminal domain of syntaxin 2C includes twocysteine residues that could potentially serve as palmitoylationsites (Bizzozero et al., 1994; Schlesinger et al., 1993). Sincesyntaxin 2D lacks potential lipid modification motifs and issoluble following expression in Hela-tTA cells, its membraneassociation is more likely mediated by interaction with aneuronal or glial membrane protein.
Syntaxins 2A and 2B differ only in the sequences of their
B. Quiñones and others
Fig. 7. In vitro protein-protein interactions.(A) Interaction of syntaxin 2 variants withGST-SNAP-25 and GST-SNAP-23.Immobilized GST-SNAP-23, GST-SNAP-25and GST (0.5 µM each) were incubated withsoluble syntaxin 2 variants (0.5 µM each) asdescribed in Materials and Methods. Boundsyntaxin 2 variants were detected byimmunoblotting utilizing an antibodyrecognizing syntaxin 2 followed by an 125I-labeled secondary antibody. A phosphorimagerscan of the immunoreactivity is presented.Black arrows indicate the position of syntaxins2A, 2B and 2C while the white arrows indicatethe position of syntaxin 2D. The nature of theband of lower mobility in the syntaxin 2B laneremains to be determined. (B) Interaction of n-sec1 with GST-syntaxin 2 variants.Immobilized GST-syntaxin 2 variants and GST(0.5 µM of each) were incubated with solublen-sec1 (0.75 µM) as described in Materials andMethods. After SDS-PAGE and transfer tonitrocellulose, the bound n-sec1 was readilydetectable by Ponceau S staining. Since thisamount of n-sec1 exceeded that which could bereliably monitored by antibody detection (datanot shown), the n-sec1 binding presentedcorresponds to the Ponceau S-stained blot.(C) Formation of ternary SNARE complexeswith GST-syntaxin 2 variants, VAMP2 andeither SNAP-23 or SNAP-25. ImmobilizedGST-syntaxin 2 variants and GST (0.5 µMeach) were incubated with soluble FLAG-tagged VAMP2 (5 µM) either in the presence (+) or absence (−) of soluble SNAP-23 or SNAP-25 (0.5 µM each). Bound VAMP2 was detectedby immunoblotting with an anti-FLAG antibody and the immunoreactive proteins were visualized by chemiluminescence.

4301Characterization of syntaxin 2 variants
hydrophobic carboxyl-terminal transmembrane domains,suggesting that these domains may have functions beyondmembrane anchoring. One potential secondary function for thedifferent transmembrane sequences is in protein targeting. Forexample, transmembrane domains have been implicated assorting signals in the retention of Golgi resident enzymes(Munro, 1995) and in delivery of certain proteins to the apicalplasma membrane of polarized epithelial cells (Kundu et al.,1996; Scheiffele et al., 1997). When expressed in nonpolarizedcells, syntaxins 2A and 2B were localized to the plasmamembrane and a perinuclear compartment overlapping with theendosomal marker cellubrevin. The pronounced perinuclearlocalization of syntaxin 2A and 2B was unexpected, since aprevious study described a primarily plasma membrane patternfor syntaxin 2A in Cos7 cells (Bennett et al., 1993). It ispossible that overexpression may at least partially contribute tothe observed intracellular staining. However, several lines ofevidence indicate that this perinuclear pattern is not simply aconsequence of syntaxin 2 overexpression. First, theperinuclear staining is observed in cells expressing the syntaxin2 variants at both high and low levels. Second, the endogenoussyntaxin 2 immunoreactivity in AtT20 cells localizes tointracellular tubulovesicular structures with a morphologysimilar to that described for endosomes in these cells (Toozeand Hollinshead, 1991). Third, incubation of Hela-tTA cellsexpressing syntaxin 2A with cycloheximide for up to 3 hours,a treatment previously shown to ‘chase’ a plasma membraneSNARE protein in transit through the secretory pathway (Janttiet al., 1994), fails to eliminate the perinuclear localization ofsyntaxin 2A (data not shown). Finally, in MDCK cells, theprominent perinuclear staining observed in subconfluent cellsis less apparent in polarized filter-grown cells. The duallocalization of syntaxins 2A and 2B suggests that theseproteins may function as t-SNAREs during both exocytic andendocytic trafficking events. Alternatively, the intracellularsyntaxin 2A and 2B may represent a pool of these variants thatdynamically cycle between an intracellular compartment andthe plasma membrane, similar to the pool of syntaxin 1Aassociated with synaptic vesicles (Otto et al., 1997; Walch-Solimena et al., 1995).
In contrast to their overlapping distributions in nonpolarizedcells, a distinction was observed in the localization of syntaxins2A and 2B in polarized epithelial cells. Syntaxin 2A waslocalized to the apical plasma membrane domain whilesyntaxin 2B was found on both the apical and lateralmembranes. Interestingly, studies of the localization ofendogenous syntaxin 2 in epithelial cells (using antibodies thatdo not distinguish between the variants) have also reporteddistributions to either the apical surface (Gaisano et al., 1996)or both the apical and basolateral plasma membrane domains(Fujita et al., 1998). Whether differential expression of thesyntaxin 2 variants is responsible for these observationsremains to be determined. In contrast to our results with bothadenovirus-infected MDCK cells and transiently transfectedCaco-2 cells, syntaxin 2A in stably transfected MDCK cellshas been reported to be partially localized to the basolateralmembrane (Low et al., 1996). One possible explanation for thisdiscrepancy is a difference in syntaxin 2A expression levels.Consistent with this possibility, infection of MDCK cells withhigher amounts of syntaxin 2A adenovirus resulted in anincrease in basolateral immunoreactivity (data not shown).
What mechanisms may be responsible for the differentialtargeting of syntaxins 2A and 2B in polarized cells? Since thesetwo variants differ only in the transmembrane domain, amechanism involving differential association with apically-targeted sphingolipid-cholesterol rafts is an attractive possibility(Aroeti et al., 1998; Simons and Ikonen, 1997). Thetransmembrane domain of influenza virus neuraminidaseconstitutes an apical targeting signal that promotesincorporation into detergent-insoluble lipid microdomains(Kundu et al., 1996). Similarly, the transmembrane domain ofinfluenza hemagglutinin is important for raft association andthis property can be disrupted by mutations that alter thehydrophobic residues in contact with the exoplasmic leaflet ofthe membrane (Scheiffele et al., 1997). Thus, it is possible thatthe different hydrophobic residues comprising thetransmembrane domains of syntaxins 2A and 2B differentiallypromote interaction with lipid rafts. In contrast to a GPI-anchored control protein, syntaxins 2A or 2B fail to associatewith lipid rafts, as assessed by Triton X-100 insolubility (datanot shown). Similarly, lipid raft association was not initiallyidentified for apically targeted syntaxins 2A and 3 in stablytransfected MDCK cells (Low et al., 1996). However, a morerecent study has demonstrated an association of endogenoussyntaxin 3 with sphingolipid-cholesterol rafts in MDCK cells(Lafont et al., 1999). Therefore, a more extensive analysis ofthe lipid raft association of syntaxin 2 variants is warranted.Alternative sorting or retention signals (Drubin and Nelson,1996; Yeaman et al., 1999) could also contribute to thedifferential distribution of syntaxins 2A and 2B in polarizedepithelial cells. However, an involvement of transmembranedomains in such signals has not been described.
Results from a variety of in vitro biochemical (Weber et al.,1998), structural (Poirier et al., 1998b; Sutton et al., 1998), andgenetic studies (Littleton et al., 1998; Schulze et al., 1995)indicate that SNARE proteins may directly participate inmembrane fusion by forming intermembrane complexes thatbring the opposing bilayers into close proximity. This functionis likely to be defined by combinations of expression patterns,intracellular localizations, and biochemical properties of theinteracting SNARE components (Bennett, 1995; Hay andScheller, 1997). The differences we have observed among thesyntaxin 2 variants are consistent with the possibility that thissubset of the syntaxin family of t-SNAREs may differentiallyregulate intracellular membrane trafficking pathways. Anadditional distinction that remains to be investigated is theability of the syntaxin 2 variants to participate in membranefusion reactions. Since the ability of influenza hemagglutininto promote membrane fusion is compromised by replacementof its transmembrane domain with a glycolipid anchor(Melikyan et al., 1995), one would expect that the function ofsyntaxin 2 variants may be influenced by the presence orabsence of a transmembrane anchor. Clearly, further studieswill be required to identify the membrane traffickingpathway(s) mediated by syntaxin 2 and establish the potentialdifferential contribution of the syntaxin 2 variants.
This work was supported by National Institutes of Health grantGM-51313. We wish to thank Duncan Stuart for assistance withconfocal microscopy, Shoichiro Tsukita for anti-ZO-1 antibody, andcurrent and former members of our laboratory for helpful discussionsthroughout the course of this work.

4302
REFERENCES
Advani, R. J., Bae, H. R., Bock, J. B., Chao, D. S., Doung, Y. C., Prekeris,R., Yoo, J. S. and Scheller, R. H. (1998). Seven novel mammalian SNAREproteins localize to distinct membrane compartments. J. Biol. Chem. 273,10317-10324.
Altschuler, Y., Barbas, S. M., Terlecky, L. J., Tang, K., Hardy, S., Mostov,K. E. and Schmid, S. L. (1998). Redundant and distinct functions fordynamin-1 and dynamin-2 isoforms. J. Cell Biol. 143, 1871-1881.
Apodaca, G., Katz, L. A. and Mostov, K. E. (1994). Receptor-mediatedtranscytosis of IgA in MDCK cells is via apical recycling endosomes. J. CellBiol. 125, 67-86.
Aroeti, B., Okhrimenko, H., Reich, V. and Orzech, E. (1998). Polarizedtrafficking of plasma membrane proteins: emerging roles for coats,SNAREs, GTPases and their link to the cytoskeleton. Biochim. Biophys.Acta 1376, 57-90.
Baumert, M., Maycox, P. R., Navone, F., De Camilli, P. and Jahn, R.(1989). Synaptobrevin: an integral membrane protein of 18,000 daltonspresent in small synaptic vesicles of rat brain. EMBO J. 8, 379-384.
Bennett, M. K. (1995). SNAREs and the specificity of transport vesicletargeting. Curr. Opin. Cell Biol. 7, 581-586.
Bennett, M. K., Calakos, N. and Scheller, R. H. (1992). Syntaxin: a synapticprotein implicated in docking of synaptic vesicles at presynaptic activezones. Science 257, 255-259.
Bennett, M. K., Garcia-Arraras, J. E., Elferink, L. A., Peterson, K.,Fleming, A. M., Hazuka, C. D. and Scheller, R. H. (1993). The syntaxinfamily of vesicular transport receptors. Cell 74, 863-873.
Bennett, M. K. and Scheller, R. H. (1993). The molecular machinery forsecretion is conserved from yeast to neurons. Proc. Natl. Acad. Sci. USA 90,2559-2563.
Berger, E. G., Burger, P., Hille, A. and Bachi, T. (1995). Comparativelocalization of mannose-6-phosphate receptor with 2,6sialyltransferase inHepG2 cells: an analysis by confocal double immunofluorescencemicroscopy. Eur. J. Cell Biol. 67, 106-111.
Bizzozero, O. A., Tetzloff, S. U. and Bharadwaj, M. (1994). Overview:protein palmitoylation in the nervous system: current views and unsolvedproblems. Neurochem. Res. 19, 923-933.
Blasi, J., Chapman, E. R., Yamasaki, S., Binz, T., Niemann, H. and Jahn,R. (1993). Botulinum neurotoxin C1 blocks neurotransmitter release bymeans of cleaving HPC-1/syntaxin. EMBO J. 12, 4821-4828.
Bobrow, M. N., Harris, T. D., Shaughnessy, K. J. and Litt, G. J. (1989).Catalyzed reporter deposition, a novel method of signal amplification.Application to immunoassays. J. Immunol. Methods 125, 279-285.
Bock, J. B., Klumperman, J., Davanger, S. and Scheller, R. H. (1997).Syntaxin 6 functions in trans-Golgi network vesicle trafficking. Mol. Biol.Cell 8, 1261-1271.
Bock, J. B. and Scheller, R. H. (1997). A fusion of new ideas. Nature 387,133-135.
Brennwald, P., Kearns, B., Champion, K., Keranen, S., Bankaitis, V. andNovick, P. (1994). Sec9 is a SNAP-25-like component of a yeast SNAREcomplex that may be the effector of Sec4 function in exocytosis. Cell 79,245-258.
Bucci, C., Parton, R. G., Mather, I. H., Stunnenberg, H., Simons, K.,Hoflack, B. and Zerial, M. (1992). The small GTPase rab5 functions as aregulatory factor in the early endocytic pathway. Cell 70, 715-728.
Calakos, N., Bennett, M. K., Peterson, K. E. and Scheller, R. H. (1994).Protein-protein interactions contributing to the specificity of intracellularvesicular trafficking. Science 263, 1146-1149.
Chapman, E. R., Hanson, P. I., An, S. and Jahn, R. (1995). Ca2+ regulatesthe interaction between synaptotagmin and syntaxin 1. J. Biol. Chem. 270,23667-23671.
Cole, N. B. and Lippincott-Schwartz, J. (1995). Organization of organellesand membrane traffic by microtubules. Curr. Opin. Cell Biol. 7, 55-64.
Drubin, D. G. and Nelson, W. J. (1996). Origins of cell polarity. Cell 84,335-344.
Elferink, L. A., Trimble, W. S. and Scheller, R. H. (1989). Two vesicle-associated membrane protein genes are differentially expressed in the ratcentral nervous system. J. Biol. Chem. 264, 11061-11064.
Fernandez, I., Ubach, J., Dulubova, I., Zhang, X., Sudhof, T. C. and Rizo,J. (1998). Three-dimensional structure of an evolutionarily conserved N-terminal domain of syntaxin 1A. Cell 94, 841-849.
Ferro-Novick, S. and Jahn, R. (1994). Vesicle fusion from yeast to man.Nature 370, 191-193.
Fiebig, K. M., Rice, L. M., Pollock, E. and Brunger, A. T. (1999). Foldingintermediates of SNARE complex assembly. Nat. Struct. Biol. 6, 117-123.
Foster, L. J., Yeung, B., Mohtashami, M., Ross, K., Trimble, W. S. andKlip, A. (1998). Binary interactions of the SNARE proteins syntaxin-4,SNAP23, and VAMP- 2 and their regulation by phosphorylation.Biochemistry 37, 11089-11096.
Fujita, H., Tuma, P. L., Finnegan, C. M., Locco, L. and Hubbard, A. L.(1998). Endogenous syntaxins 2, 3 and 4 exhibit distinct but overlappingpatterns of expression at the hepatocyte plasma membrane. Biochem. J. 329,527-538.
Gaisano, H. Y., Ghai, M., Malkus, P. N., Sheu, L., Bouquillon, A., Bennett,M. K. and Trimble, W. S. (1996). Distinct cellular locations of the syntaxinfamily of proteins in rat pancreatic acinar cells. Mol. Biol. Cell 7, 2019-2027.
Gossen, M. and Bujard, H. (1992). Tight control of gene expression inmammalian cells by tetracycline-responsive promoters. Proc. Natl. Acad.Sci. USA 89, 5547-5551.
Guan, K. L. and Dixon, J. E. (1991). Eukaryotic proteins expressed inEscherichia coli: an improved thrombin cleavage and purification procedureof fusion proteins with glutathione S-transferase. Anal. Biochem. 192, 262-267.
Hanson, P. I., Otto, H., Barton, N. and Jahn, R. (1995). The N-ethylmaleimide-sensitive fusion protein and alpha-SNAP induce aconformational change in syntaxin. J. Biol. Chem. 270, 16955-16961.
Hao, J. C., Salem, N., Peng, X. R., Kelly, R. B. and Bennett, M. K. (1997).Effect of mutations in vesicle-associated membrane protein (VAMP) on theassembly of multimeric protein complexes. J. Neurosci. 17, 1596-1603.
Hardy, S., Kitamura, M., Harris-Stansil, T., Dai, Y. and Phipps, M. L.(1997). Construction of adenovirus vectors through Cre-lox recombination.J. Virol. 71, 1842-1849.
Hata, Y., Slaughter, C. A. and Sudhof, T. C. (1993). Synaptic vesicle fusioncomplex contains unc-18 homologue bound to syntaxin. Nature 366, 347-351.
Hata, Y. and Sudhof, T. C. (1995). A novel ubiquitous form of Munc-18interacts with multiple syntaxins. Use of the yeast two-hybrid system tostudy interactions between proteins involved in membrane traffic. J. Biol.Chem. 270, 13022-13028.
Hay, J. C., Chao, D. S., Kuo, C. S. and Scheller, R. H. (1997). Proteininteractions regulating vesicle transport between the endoplasmic reticulumand Golgi apparatus in mammalian cells. Cell 89, 149-158.
Hay, J. C. and Scheller, R. H. (1997). SNAREs and NSF in targetedmembrane fusion. Curr. Opin. Cell Biol. 9, 505-512.
Hayashi, T., McMahon, H., Yamasaki, S., Binz, T., Hata, Y., Sudhof, T. C.and Niemann, H. (1994). Synaptic vesicle membrane fusion complex:action of clostridial neurotoxins on assembly. EMBO J. 13, 5051-5061.
Ibaraki, K., Horikawa, H. P., Morita, T., Mori, H., Sakimura, K., Mishina,M., Saisu, H. and Abe, T. (1995). Identification of four different forms ofsyntaxin 3. Biochem. Biophys. Res. Commun. 211, 997-1005.
Itoh, M., Yonemura, S., Nagafuchi, A., Tsukita, S. and Tsukita, S. (1991).A 220-kD undercoat-constitutive protein: its specific localization atcadherin-based cell-cell adhesion sites. J. Cell Biol. 115, 1449-1462.
Jagadish, M. N., Tellam, J. T., Macaulay, S. L., Gough, K. H., James, D.E. and Ward, C. W. (1997). Novel isoform of syntaxin 1 is expressed inmammalian cells. Biochem. J. 321, 151-156.
Jahn, R. and Niemann, H. (1994). Molecular mechanisms of clostridialneurotoxins. Ann. NY Acad. Sci. 733, 245-255.
Jantti, J., Keranen, S., Toikkanen, J., Kuismanen, E., Ehnholm, C.,Soderlund, H. and Olkkonen, V. M. (1994). Membrane insertion andintracellular transport of yeast syntaxin Sso2p in mammalian cells. J. CellSci. 107, 3623-3633.
Jin, P., Hardy, S. and Morgan, D. O. (1998). Nuclear localization of cyclinB1 controls mitotic entry after DNA damage. J. Cell Biol. 141, 875-885.
Kee, Y., Lin, R. C., Hsu, S. C. and Scheller, R. H. (1995). Distinct domainsof syntaxin are required for synaptic vesicle fusion complex formation anddissociation. Neuron 14, 991-998.
Klausner, R. D., Donaldson, J. G. and Lippincott-Schwartz, J. (1992).Brefeldin A: insights into the control of membrane traffic and organellestructure. J. Cell Biol. 116, 1071-1080.
Kundu, A., Avalos, R. T., Sanderson, C. M. and Nayak, D. P. (1996).Transmembrane domain of influenza virus neuraminidase, a type II protein,possesses an apical sorting signal in polarized MDCK cells. J. Virol. 70,6508-6515.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assemblyof the head of bacteriophage T4. Nature 227, 680-685.
B. Quiñones and others

4303Characterization of syntaxin 2 variants
Lafont, F., Verkade, P., Galli, T., Wimmer, C., Louvard, D. and Simons,K. (1999). Raft association of SNAP receptors acting in apical traffickingin madin-darby canine kidney cells [In Process Citation]. Proc. Natl. Acad.Sci. USA 96, 3734-3738.
Lippincott-Schwartz, J. (1993). Membrane cycling between the ER andGolgi apparatus and its role in biosynthetic transport. Subcell. Biochem. 21,95-119.
Littleton, J. T., Chapman, E. R., Kreber, R., Garment, M. B., Carlson, S.D. and Ganetzky, B. (1998). Temperature-sensitive paralytic mutationsdemonstrate that synaptic exocytosis requires SNARE complex assemblyand disassembly. Neuron 21, 401-413.
Low, S. H., Chapin, S. J., Weimbs, T., Komuves, L. G., Bennett, M. K.and Mostov, K. E. (1996). Differential localization of syntaxin isoformsin polarized Madin-Darby canine kidney cells. Mol. Biol. Cell 7, 2007-2018.
Low, S. H., Roche, P. A., Anderson, H. A., van Ijzendoorn, S. C., Zhang,M., Mostov, K. E. and Weimbs, T. (1998). Targeting of SNAP-23 andSNAP-25 in polarized epithelial cells. J. Biol. Chem. 273, 3422-3430.
McMahon, H. T., Ushkaryov, Y. A., Edelmann, L., Link, E., Binz, T.,Niemann, H., Jahn, R. and Sudhof, T. C. (1993). Cellubrevin is aubiquitous tetanus-toxin substrate homologous to a putative synaptic vesiclefusion protein [see comments]. Nature 364, 346-349.
Melikyan, G. B., White, J. M. and Cohen, F. S. (1995). GPI-anchoredinfluenza hemagglutinin induces hemifusion to both red blood cell andplanar bilayer membranes. J. Cell Biol. 131, 679-691.
Montecucco, C. (1998). Protein toxins and membrane transport. Curr. Opin.Cell Biol. 10, 530-536.
Munro, S. (1995). An investigation of the role of transmembrane domains inGolgi protein retention. EMBO J. 14, 4695-4704.
Nakamura, N., Rabouille, C., Watson, R., Nilsson, T., Hui, N., Slusarewicz,P., Kreis, T. E. and Warren, G. (1995). Characterization of a cis-Golgimatrix protein, GM130. J. Cell Biol. 131, 1715-1726.
Novick, P. and Zerial, M. (1997). The diversity of Rab proteins in vesicletransport. Curr. Opin. Cell Biol. 9, 496-504.
Ogawa, H., Harada, S., Sassa, T., Yamamoto, H. and Hosono, R. (1998).Functional properties of the unc-64 gene encoding a Caenorhabditis eleganssyntaxin. J. Biol. Chem. 273, 2192-2198.
Otto, H., Hanson, P. I. and Jahn, R. (1997). Assembly and disassembly ofa ternary complex of synaptobrevin, syntaxin, and SNAP-25 in themembrane of synaptic vesicles. Proc. Natl. Acad. Sci. USA 94, 6197-6201.
Oyler, G. A., Higgins, G. A., Hart, R. A., Battenberg, E., Billingsley, M.,Bloom, F. E. and Wilson, M. C. (1989). The identification of a novelsynaptosomal-associated protein, SNAP-25, differentially expressed byneuronal subpopulations. J. Cell Biol. 109, 3039-3052.
Pelham, H. R. (1991). Multiple targets for brefeldin A. Cell 67, 449-451.Pelham, H. R. (1996). The dynamic organisation of the secretory pathway.
Cell Struct. Funct. 21, 413-419.Pelham, H. R. (1999). SNAREs and the secretory pathway-lessons from yeast.
Exp. Cell Res. 247, 1-8.Peng, X. R., Yao, X., Chow, D. C., Forte, J. G. and Bennett, M. K. (1997).
Association of syntaxin 3 and vesicle-associated membrane protein (VAMP)with H+/K(+)-ATPase-containing tubulovesicles in gastric parietal cells.Mol. Biol. Cell 8, 399-407.
Pevsner, J. (1996). The role of Sec1p-related proteins in vesicle trafficking inthe nerve terminal. J. Neurosci. Res. 45, 89-95.
Pevsner, J., Hsu, S. C., Braun, J. E., Calakos, N., Ting, A. E., Bennett, M.K. and Scheller, R. H. (1994a). Specificity and regulation of a synapticvesicle docking complex. Neuron 13, 353-361.
Pevsner, J., Hsu, S. C. and Scheller, R. H. (1994b). n-Sec1: a neural-specificsyntaxin-binding protein. Proc. Natl. Acad. Sci. USA 91, 1445-1449.
Poirier, M. A., Hao, J. C., Malkus, P. N., Chan, C., Moore, M. F., King, D.S. and Bennett, M. K. (1998a). Protease resistance of syntaxin.SNAP-25.VAMP complexes. Implications for assembly and structure. J. Biol.Chem. 273, 11370-11377.
Poirier, M. A., Xiao, W., Macosko, J. C., Chan, C., Shin, Y. K. and Bennett,M. K. (1998b). The synaptic SNARE complex is a parallel four-strandedhelical bundle. Nat. Struct. Biol. 5, 765-769.
Prekeris, R., Klumperman, J., Chen, Y. A. and Scheller, R. H. (1998).Syntaxin 13 mediates cycling of plasma membrane proteins viatubulovesicular recycling endosomes. J. Cell Biol. 143, 957-971.
Pryer, N. K., Wuestehube, L. J. and Schekman, R. (1992). Vesicle-mediatedprotein sorting. Annu. Rev. Biochem. 61, 471-516.
Riento, K., Galli, T., Jansson, S., Ehnholm, C., Lehtonen, E. andOlkkonen, V. M. (1998). Interaction of munc-18-2 with syntaxin 3 controls
the association of apical SNAREs in epithelial cells [In Process Citation].J. Cell Sci. 111, 2681-2688.
Risinger, C. and Bennett, M. K. (1999). Differential phosphorylation ofsyntaxin and synaptosome-associated protein of 25 kDa (SNAP-25)isoforms [In Process Citation]. J. Neurochem. 72, 614-624.
Roth, J., Taatjes, D. J., Lucocq, J. M., Weinstein, J. and Paulson, J. C.(1985). Demonstration of an extensive trans-tubular network continuouswith the Golgi apparatus stack that may function in glycosylation. Cell 43,287-295.
Rothman, J. E. (1994). Mechanisms of intracellular protein transport. Nature372, 55-63.
Saifee, O., Wei, L. and Nonet, M. L. (1998). The Caenorhabditis elegansunc-64 locus encodes a syntaxin that interacts genetically withsynaptobrevin. Mol. Biol. Cell 9, 1235-1252.
Sambrook, J. M., Fritsch, E.F., and Maniatis, T. (1989). Molecular Cloning:A Laboratory Manual. New York: Cold Spring Harbor Laboratory Press.
Scheiffele, P., Roth, M. G. and Simons, K. (1997). Interaction of influenzavirus haemagglutinin with sphingolipid- cholesterol membrane domains viaits transmembrane domain. EMBO J. 16, 5501-5508.
Schlesinger, M. J., Veit, M. and Schmidt, M. F. G. (1993). LipidModifications of Proteins. Boca Raton, FL: CRC Press.
Schulze, K. L., Broadie, K., Perin, M. S. and Bellen, H. J. (1995). Geneticand electrophysiological studies of Drosophila syntaxin-1A demonstrate itsrole in nonneuronal secretion and neurotransmission. Cell 80, 311-320.
Simons, K. and Ikonen, E. (1997). Functional rafts in cell membranes. Nature387, 569-572.
Simonsen, A., Bremnes, B., Ronning, E., Aasland, R. and Stenmark, H.(1998). Syntaxin-16, a putative Golgi t-SNARE. Eur. J. Cell Biol. 75, 223-231.
Sogaard, M., Tani, K., Ye, R. R., Geromanos, S., Tempst, P., Kirchhausen,T., Rothman, J. E. and Sollner, T. (1994). A rab protein is required for theassembly of SNARE complexes in the docking of transport vesicles. Cell78, 937-948.
Sollner, T., Bennett, M. K., Whiteheart, S. W., Scheller, R. H. andRothman, J. E. (1993a). A protein assembly-disassembly pathway in vitrothat may correspond to sequential steps of synaptic vesicle docking,activation, and fusion. Cell 75, 409-418.
Sollner, T., Whiteheart, S. W., Brunner, M., Erdjument-Bromage, H.,Geromanos, S., Tempst, P. and Rothman, J. E. (1993b). SNAP receptorsimplicated in vesicle targeting and fusion [see comments]. Nature 362, 318-324.
Steegmaier, M., Yang, B., Yoo, J. S., Huang, B., Shen, M., Yu, S., Luo, Y.and Scheller, R. H. (1998). Three novel proteins of the syntaxin/SNAP-25family. J. Biol. Chem. 273, 34171-34179.
Stevenson, B. R., Siliciano, J. D., Mooseker, M. S. and Goodenough, D. A.(1986). Identification of ZO-1: a high molecular weight polypeptideassociated with the tight junction (zonula occludens) in a variety of epithelia.J. Cell Biol. 103, 755-766.
Sutton, R. B., Fasshauer, D., Jahn, R. and Brunger, A. T. (1998). Crystalstructure of a SNARE complex involved in synaptic exocytosis at 2.4 Aresolution [In Process Citation]. Nature 395, 347-353.
Tang, B. L., Low, D. Y. and Hong, W. (1998a). Syntaxin 11: a member ofthe syntaxin family without a carboxyl terminal transmembrane domain.Biochem. Biophys. Res. Commun. 245, 627-632.
Tang, B. L., Low, D. Y., Lee, S. S., Tan, A. E. and Hong, W. (1998b).Molecular cloning and localization of human syntaxin 16, a member of thesyntaxin family of SNARE proteins. Biochem. Biophys. Res. Commun. 242,673-679.
Tang, B. L., Low, D. Y., Tan, A. E. and Hong, W. (1998c). Syntaxin 10: amember of the syntaxin family localized to the trans-Golgi network.Biochem. Biophys. Res. Commun. 242, 345-350.
Tang, B. L., Tan, A. E., Lim, L. K., Lee, S. S., Low, D. Y. and Hong, W.(1998d). Syntaxin 12, a member of the syntaxin family localized to theendosome. J. Biol. Chem. 273, 6944-6950.
Tang, B. L., Wong, S. H., Qi, X., Subramaniam, V. N. and Hong, W. (1992).Golgi-localized beta-galactoside alpha 2,6-sialyltransferase in transfectedCHO cells is redistributed into the endoplasmic reticulum by brefeldin A.Eur. J. Cell Biol. 59, 228-231.
Teter, K., Chandy, G., Quinones, B., Pereyra, K., Machen, T. and Moore,H. P. (1998). Cellubrevin-targeted fluorescence uncovers heterogeneity inthe recycling endosomes. J. Biol. Chem. 273, 19625-19633.
Tooze, J. and Hollinshead, M. (1991). Tubular early endosomal networks inAtT20 and other cells. J. Cell Biol. 115, 635-653.
Trimble, W. S., Cowan, D. M. and Scheller, R. H. (1988). VAMP-1: a

4304
synaptic vesicle-associated integral membrane protein. Proc. Natl. Acad.Sci. USA 85, 4538-4542.
Walch-Solimena, C., Blasi, J., Edelmann, L., Chapman, E. R., vonMollard, G. F. and Jahn, R. (1995). The t-SNAREs syntaxin 1 and SNAP-25 are present on organelles that participate in synaptic vesicle recycling. J.Cell Biol. 128, 637-645.
Weber, T., Zemelman, B. V., McNew, J. A., Westermann, B., Gmachl, M.,Parlati, F., Sollner, T. H. and Rothman, J. E. (1998). SNAREpins:minimal machinery for membrane fusion. Cell 92, 759-772.
Wong, S. H., Xu, Y., Zhang, T. and Hong, W. (1998). Syntaxin 7, a novelsyntaxin member associated with the early endosomal compartment. J. Biol.Chem. 273, 375-380.
Yang, B., Gonzalez, L., Jr., Prekeris, R., Steegmaier, M., Advani, R. J. andScheller, R. H. (1999). SNARE interactions are not selective. Implicationsfor membrane fusion specificity. J. Biol. Chem. 274, 5649-5653.
Yeaman, C., Grindstaff, K. K. and Nelson, W. J. (1999). New perspectiveson mechanisms involved in generating epithelial cell polarity. Physiol. Rev.79, 73-98.
B. Quiñones and others





























