Embed Size (px)
Citation preview

Full Length Article
Characterization of new strains of Hortaeawerneckii isolated from salt marshes of Egypt
Ashraf Elsayed *, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil,Nada I. MagdyBotany Department, Faculty of Science, Mansoura University, Mansoura, Egypt
A R T I C L E I N F O
Article history:
Received 28 June 2016
Received in revised form 8
September 2016
Accepted 9 September 2016
Available online
A B S T R A C T
Three new black yeast strains (EGYNDA08, EGYNDA16 and EGYNDA90) of marine origin were
isolated from the Egyptian off shore salt marshes and molecularly identified by ITS1 and
ITS2 5.8S rRNA gene sequencing. The molecular identification showed a high sequence iden-
tity between the two of them and Hortaea werneckii Hw6 strain, while the other strain was
unique. The biochemical characterization using different nutritional media showed differ-
ent growth capabilities and the qualitative enzyme tests (such as catalase, urease, lipase,
proteases, amylases and cellulose) showed different activity levels. The morphological char-
acterization showed different developmental stages of hyphal maturation. The phylogenetic
analysis of the three strains indicated that, two isolates were evolutionary relevant to the
Hw6 strain isolated from Spain and one novel strain was isolated with rather different mo-
lecular and morphological characteristics.
© 2016 Production and hosting by Elsevier B.V. on behalf of Mansoura University. This is
an open access article under the CC BY-NC-ND license (http://creativecommons.org/
licenses/by-nc-nd/4.0/).
Keywords:
Black yeast
Hortaea and salt marshes
1. Introduction
From the taxonomic and phylogenetic point of view, ‘Black yeast’is a polyphyletic group [1,2] of dematiaceous, filamentous [3]fungi of which several representatives share morphological fea-tures [4] such as reproducing by unicellular growth [5]. Theythrive in extreme environments characterized by scarce nu-trients, low oxygen tension, high temperature, harmful UVradiation, osmotic stress as well as a mixture of these condi-tions [4].
Hortaea werneckii was identified as an extreme halotolerantfungus [6] that belongs to the black yeast group and was iso-lated from different marine habitats. H. wernrckii was reportedto inhibit the afore-mentioned extreme conditions. Hortaea sp.belongs to the Capnodiales order in the Ascomycota phylum[7]. Based on the state of the niche (static or dynamic) [5],H. werneckii switches between two main life phases yeast phaseand hyphal phase [8]. Both phases ended up with sclerotialbodies under sever environmental conditions [8].
Under light microscope, H. werneckii is a synanomorphic mi-croorganism which produces buds, conidia, arthrocondia and
* Corresponding author.E-mail address: [email protected] (A. Elsayed).
http://dx.doi.org/10.1016/j.ejbas.2016.09.0012314-808X/© 2016 Production and hosting by Elsevier B.V. on behalf of Mansoura University. This is an open access article under theCC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
Available online at www.sciencedirect.com
journal homepage: ht tp : / /ees .e lsevier.com/ejbas/defaul t .asp
Q1
Q2
Q3
1bs_bs_query
2bs_bs_query
3bs_bs_query
4bs_bs_query
5bs_bs_query
6bs_bs_query
7bs_bs_query
8bs_bs_query
9bs_bs_query
10bs_bs_query
11bs_bs_query
12bs_bs_query
13bs_bs_query
14bs_bs_query
15bs_bs_query
16bs_bs_query
17bs_bs_query
18bs_bs_query
19bs_bs_query20bs_bs_query
21bs_bs_query
22bs_bs_query23bs_bs_query
24bs_bs_query25bs_bs_query
26bs_bs_query27bs_bs_query
28bs_bs_query
29bs_bs_query
30bs_bs_query
31bs_bs_query
32bs_bs_query
33bs_bs_query
34bs_bs_query
35bs_bs_query
36bs_bs_query
37bs_bs_query
38bs_bs_query
39bs_bs_query
40bs_bs_query
41bs_bs_query
42bs_bs_query
43bs_bs_query
44bs_bs_query
45bs_bs_query
46bs_bs_query
47bs_bs_query
48bs_bs_query
49bs_bs_query
50bs_bs_query
51bs_bs_query
52bs_bs_query
53bs_bs_query
54bs_bs_query
55bs_bs_query
56bs_bs_query
57bs_bs_query
58bs_bs_query
59bs_bs_query
60bs_bs_query
61bs_bs_query
HOSTED BY
ScienceDirect

endoconidia. Buds are enteroblastic annellidic ones [8] and theyemerged as polar, lateral, and bipolar from yeast-like forms [9].Conidiogenesis and budding mode are the same as both budsand conidia are released by the complete separation of the de-veloped abscission line in a septum [3]. When cells aresuccessively generated from the same locus, the remnants ofthe outer layers of the cell wall are arranged consecutivelyaround the tip, an increase in neck (collars) of the mother cellresults in bottle-shaped cells marked by many rings directedtoward the pole and the number of rings is dependent on thenumber of buds or conidia released [8]. The conidia are pro-duced from the generative apex of annellidophores which areunbranched intercalary hyphal cells with tapered tips. Conidiaare mainly unicellular; however two-celled conidia separatedby conspicuous dark cross wall were found [3]. H. werneckii pro-duces hydrophobic dematiaceous septated mycelia [3] whichare formed of coherent thalli and extended from the mothercell wall [8] and lateral branches sometimes are expanded fromthem [9].
Hyphal cells septa have an abscission line separatingbetween septum sheets. In addition to the simple central pore,triangular points in mature cells are laying at the anastomo-sis of inner and outer layer which marks the future points ofseparation and the adjacent outer layer remains intact withouta separation line until releasing enteroarthric thin-walled scar-free cells either in the form of an endogenous conidia fromthe yeast-like cells or arthroconidia from the fragmentationof hyphae by shedding off parts of the outer cell wall layers,which are easily ruptured [8,9].
Up to our knowledge, isolation and characterization ofH. werneckii from the Egyptian environment was not reportedbefore. In this survey, the isolated strains of H. werneckii fromEgypt were molecularly, biochemically and morphologicallycharacterized to focus on differences between them andbetween other previously published characterized H. werneckiistrains.
2. Materials and methods
2.1. Sample collection
Water samples from different off shore salt marshes locatedafter 10 kilometers from west of Gamasa on Damietta road,were collected at the beginning of December and three samplesin the end of March. Samples were stored in glass sealed jars.
2.2. Isolation and purification of the marine black yeast
One milliliter from the collected samples was inoculated in twosaline enriched liquid medium (0.5% yeast extract without/with 1% glucose) dissolved in sea water and 100 μg/ml ofstreptomycin was added after sterilization in order to inhibitundesired bacterial growth. The incubation was held at 15 °C,25 °C and 30 °C in an orbital incubator shaker at 150 rpm forthree weeks [10].
After one week, sub-culturing from the liquid media of tur-bidity was performed on the same saline enriched agar media.Temperatures 15 °C, 25 °C and 30 °C were repeated until the
filamentous community invading during the first week and thendeveloped to unicellular communities appeared on the thirdweek until the appearance of yeast-like melanized colonieswhich are purified by sub-culturing on media containing 10%NaCl. The pure cultures were preserved in 20% glycerol andstored at −20 °C.
2.3. Molecular identification of the isolates
Fungal DNA was extracted by employing the FastDNA® SpinKit according to the supplier’s instructions.The yield of genomicDNA was measured by a Nano Drop spectrophotometer by mea-suring the absorbance at 260 nm. The ITS1 region from DNAsample extracts was amplified in triplicate using primers withhigh specificity for ascomycete fungi (18FITS1 (CTTGGTCATTTAGAGGAAGTAA) and 18RITS4 (TCCTCCGCTTATTGATATGC). ThePCR reactions were performed in a thermo-cycler at a totalvolume of 50 μl using the temperature programs: 94 °C for 5 min,94 °C for 40 s, 55 °C for 45 s, 72 °C for 1.5 min, 72 °C for 7 min(35 cycles). The sizes of the PCR products were determined byelectrophoresis on 1.5% agarose gels.The desired products wereexcised and purified by the Qiagen II Agarose Gel ExtractionKit.
The sequencing reactions were performed in a thermocycler(Master cycler, Eppendorf, Hamburg, Germany) at a total volumeof 10 μl by using the temperature program: 96 °C for 1 min, 96 °Cfor 30 s, 60 °C for 10 s, 60 °C for 4 min, 72 °C for 5 min (25 cycles).The sequencing reaction products were purified by employ-ing the Dye Ex 2.0 Spin Kit. The purified sequencing reactionproducts were dried in vacuum centrifuge and then analyzedusing applied biosystems (ABI PRISM Big Dye Terminator v1.1),Ready Reaction Cycle Sequencing Kit and employing an ABI3130 XL Genetic Analyser (Applied Biosystems, Darmstadt) [11].The obtained sequences were annotated using theSequencher™ 4.8 Software. DNA similarity searches were per-formed using the BlastN program and the databases ofEuropean Molecular Biology Laboratory (EMBL) and GenBankfrom the National Center for Biotechnology Information website(NCBI).
Phylogenetic and molecular evolutionary analyses for 18SrRNA gene nucleotide sequences were conducted for se-quence alignments using the computer programs ClustalW andBioEdit 7.0.5.3 and implement in MEGA software version 5. Phy-logenetic trees were constructed using the Neighbor-Joining [12]algorithm method. Distances were generated using the KimuraMatrix, and the tree stability was supported through Boot-strap analysis (1000 replications).
2.4. Biochemical characterization
2.4.1. Nutritional characteristicsDifferent media were tested for supporting the growth of theisolates such as Malt Extract Agar media (MEA) (2% malt extractand 1.5% agar) [2], Potato Dextrose Agar media (PDA) (2% dex-trose 20 g, sliced potato 1.5% agar) and modified Yeast ExtractPeptone Dextrose media (YPD) (2% glucose-1% pepton-1% yeastextract-1.5% agar) [13]. All media were dissolved in sea waterand cultures were incubated at 25 °C.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
2 e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
Q4
62bs_bs_query
63bs_bs_query
64bs_bs_query
65bs_bs_query
66bs_bs_query
67bs_bs_query
68bs_bs_query
69bs_bs_query
70bs_bs_query
71bs_bs_query
72bs_bs_query
73bs_bs_query
74bs_bs_query
75bs_bs_query
76bs_bs_query
77bs_bs_query
78bs_bs_query
79bs_bs_query
80bs_bs_query
81bs_bs_query
82bs_bs_query
83bs_bs_query
84bs_bs_query
85bs_bs_query
86bs_bs_query
87bs_bs_query
88bs_bs_query
89bs_bs_query
90bs_bs_query
91bs_bs_query
92bs_bs_query
93bs_bs_query
94bs_bs_query
95bs_bs_query
96bs_bs_query
97bs_bs_query
98bs_bs_query
99bs_bs_query
100bs_bs_query
101bs_bs_query
102bs_bs_query
103bs_bs_query
104bs_bs_query
105bs_bs_query
106bs_bs_query
107bs_bs_query
108bs_bs_query
109bs_bs_query
110bs_bs_query
111bs_bs_query
112bs_bs_query
113bs_bs_query
114bs_bs_query
115bs_bs_query
116bs_bs_query
117bs_bs_query
118bs_bs_query
119bs_bs_query
120bs_bs_query
121bs_bs_query
122bs_bs_query
123bs_bs_query
124bs_bs_query
125bs_bs_query
126bs_bs_query
127bs_bs_query
128bs_bs_query
129bs_bs_query
130bs_bs_query
131bs_bs_query
132bs_bs_query
133bs_bs_query
134bs_bs_query
135bs_bs_query
136bs_bs_query
137bs_bs_query
138bs_bs_query
139bs_bs_query
140bs_bs_query
141bs_bs_query
142bs_bs_query
143bs_bs_query
144bs_bs_query
145bs_bs_query
146bs_bs_query
147bs_bs_query
148bs_bs_query
149bs_bs_query
150bs_bs_query
151bs_bs_query
152bs_bs_query
153bs_bs_query
154bs_bs_query
155bs_bs_query
156bs_bs_query
157bs_bs_query
158bs_bs_query
159bs_bs_query
160bs_bs_query
161bs_bs_query
162bs_bs_query
163bs_bs_query
164bs_bs_query
165bs_bs_query
166bs_bs_query
167bs_bs_query
168bs_bs_query
169bs_bs_query
170bs_bs_query
171bs_bs_query
172bs_bs_query
173bs_bs_query
174bs_bs_query
175bs_bs_query
176bs_bs_query
177bs_bs_query
178bs_bs_query

2.4.2. Enzyme tests
2.4.2.1. Catalase test. In room temperature, a drop of diluted30% Hydrogen Peroxide was placed onto a slide containing adrop of liquid modified YPD containing tested organisms with6 days old. Effervescence reaction could be observed in-stantly as a positive result [14].
2.4.2.2. Urease test. Rustigian and Stuart’s urea broth media[15] containing urea 20 g, monopotassium phosphate 9.1 g, di-potassium phosphate 9.5 g and Phenol Red 0.01 g were dissolvedin one liter of distilled water with final pH 6.8, were sterilizedusing Millipore filter papers, poured into sterilized tubes, in-oculated with culture and incubated at 25 °C for five days. Thechange in media color from yellow to pink was regarded as apositive result.
2.4.2.3. Lipase test. Sierra lipase test protocol was done [16]using Tween-80 (10.0 g) and agar 20.0 g in one liter of sea waterand the final pH was 6. Agar was dissolved in sea water andautoclaved and Tween was sterilized separately. After inocu-lation, the plates were incubated at 25 °C for three days. Theobservation of white precipitation around the growinginoculums was considered as a positive result.
2.4.2.4. Proteases test. A modified protocol of 30% skim milk-agar medium [17] was done by adding 30 ml packed liquid skimmilk (the used skim milk nutrition label on the pack: Fats .44 g,Carbohydrates: 10.6 g, Protein: 6.9 g, Vitamin: B1 .1 mg andVitamin: B2 .14 mg). The formation of transparent zone aroundthe growing isolates was considered as a positive result.
2.4.2.5. Amylases test. Vedder starch media composed ofsoluble starch 10 g, agar 12 g and sodium nitrate 6.5 g were dis-solved in one liter sea water and adjusted to pH 6. Amylasesactivity was detected by flooding the surface of inoculatedmedia with Grams Iodine [18].
2.4.2.6. Cellulase test. The Carboxymethylcellulose (CMC) media[19,20], which were composed of: CMC 10 g, Sodium Nitrate 6.5 g,Potassium Hydrogen Phosphate 6.5 g, Potassium chloride 6.5 g,Magnesium sulphate heptahydrate 3.0 g and Agar 17.5 g in oneliter with final pH of 6, were used for cellulose activity. The ac-tivity was visualized by staining the media with Congo red dyefollowed by de-staining with Sodium Chloride solution. De-
staining process was repeated continuously until a clear zoneappeared around the growing hyphae.
2.5. Microscopic characterization
Specimens of the three strains of the five days old light greencolonies for yeast phase formation and one month old blackcolonies for hyphal phase formation were examined under lightmicroscope (40×).
3. Results
3.1. Black yeast isolation
The incubational temperature 25 °C and 10% NaCl enhancedthe growth of the black yeast isolates. Two isolates were re-covered from water samples, EGYNDA08 at the beginning ofDecember and EGYNDA16 at the end of March, while anotherisolate EGYNDA90 was recovered from NaCl crystals of saltmarshes at the beginning of May.
3.2. Molecular identification
From the phylogenetic tree (Fig. 1), three new black yeast strains(EGYNDA08, EGYNDA16 and EGYNDA90) were visualized. Oneof them (EGYNDA90, accession number KU341734) lay in a sepa-rate branch while the others, EGYNDA08 (accession numberKU341732) and EGYNDA16 (accession number KU341733), werevery close to each other and to Hw6-JN997370.1 which was iso-lated from Spain. The similarity between EGYNDA16 and theother ecotypes was 97.4% maximum using Pairwise SequenceAlignment (Table 1).
3.3. Biochemical characterization
3.3.1. Nutritional characteristicsEGYNDA08 grew on MEA as black small colonies with hyphalgrowth stimulation; also it grew on the modified YPD media,meanwhile, EGYNDA16 and EGYNDA90 grew on YPD media andthey hardly grew on MEA and the media contained 0.5% yeastextract.
3.3.2. Enzymes tests
3.3.2.1. Catalase test. The effervescence that resulted on a slidewas very weak in case of EGYNDA08, very strong in case of
Fig. 1 – Phylogenetic tree constructed by MEGA using Neighbor-Joining tree, with number of Bootstrap replications 500,between different Hortaea werneckii strains including the three isolates and Phaeotheca triangularis. T = ex-type strain.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
3e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
Q5
Q6
179bs_bs_query
180bs_bs_query
181bs_bs_query
182bs_bs_query
183bs_bs_query
184bs_bs_query
185bs_bs_query
186bs_bs_query
187bs_bs_query
188bs_bs_query
189bs_bs_query
190bs_bs_query
191bs_bs_query
192bs_bs_query
193bs_bs_query
194bs_bs_query
195bs_bs_query
196bs_bs_query
197bs_bs_query
198bs_bs_query
199bs_bs_query
200bs_bs_query
201bs_bs_query
202bs_bs_query
203bs_bs_query
204bs_bs_query
205bs_bs_query
206bs_bs_query
207bs_bs_query
208bs_bs_query
209bs_bs_query
210bs_bs_query
211bs_bs_query
212bs_bs_query
213bs_bs_query
214bs_bs_query
215bs_bs_query
216bs_bs_query
217bs_bs_query
218bs_bs_query
219bs_bs_query
220bs_bs_query
221bs_bs_query
222bs_bs_query
223bs_bs_query
224bs_bs_query
225bs_bs_query
226bs_bs_query
227bs_bs_query
228bs_bs_query
229bs_bs_query
230bs_bs_query
231bs_bs_query
232bs_bs_query
233bs_bs_query
234bs_bs_query
235bs_bs_query
236bs_bs_query
237bs_bs_query
238bs_bs_query
239bs_bs_query
240bs_bs_query
241bs_bs_query
242bs_bs_query
243bs_bs_query
244bs_bs_query
245bs_bs_query
246bs_bs_query
247bs_bs_query
248bs_bs_query
249bs_bs_query
250bs_bs_query
251bs_bs_query
252bs_bs_query
253bs_bs_query
254bs_bs_query
255bs_bs_query
256bs_bs_query
257bs_bs_query
258bs_bs_query
259bs_bs_query
260bs_bs_query
261bs_bs_query
262bs_bs_query
263bs_bs_query
264bs_bs_query
265bs_bs_query
266bs_bs_query
267bs_bs_query
268bs_bs_query
269bs_bs_query
270bs_bs_query
271bs_bs_query
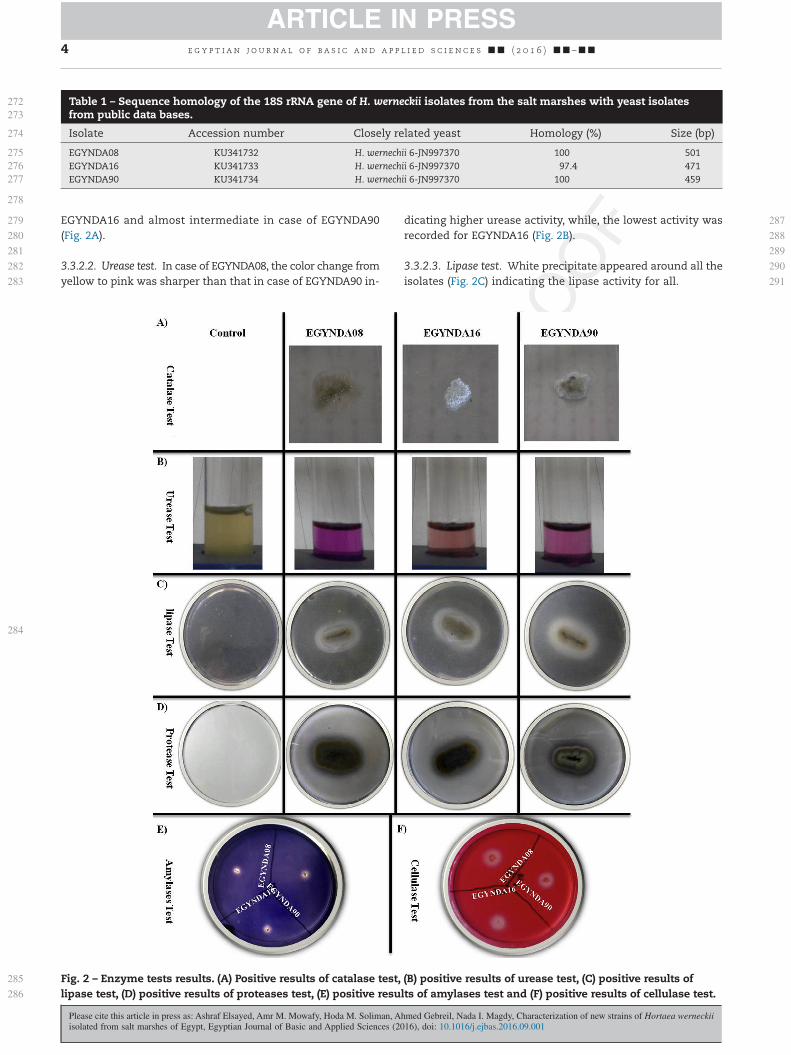
EGYNDA16 and almost intermediate in case of EGYNDA90(Fig. 2A).
3.3.2.2. Urease test. In case of EGYNDA08, the color change fromyellow to pink was sharper than that in case of EGYNDA90 in-
dicating higher urease activity, while, the lowest activity wasrecorded for EGYNDA16 (Fig. 2B).
3.3.2.3. Lipase test. White precipitate appeared around all theisolates (Fig. 2C) indicating the lipase activity for all.
Table 1 – Sequence homology of the 18S rRNA gene of H. werneckii isolates from the salt marshes with yeast isolatesfrom public data bases.
Isolate Accession number Closely related yeast Homology (%) Size (bp)
EGYNDA08 KU341732 H. wernechii 6-JN997370 100 501EGYNDA16 KU341733 H. wernechii 6-JN997370 97.4 471EGYNDA90 KU341734 H. wernechii 6-JN997370 100 459
Fig. 2 – Enzyme tests results. (A) Positive results of catalase test, (B) positive results of urease test, (C) positive results oflipase test, (D) positive results of proteases test, (E) positive results of amylases test and (F) positive results of cellulase test.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
4 e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
272bs_bs_query
273bs_bs_query
274bs_bs_query
275bs_bs_query
276bs_bs_query
277bs_bs_query
278bs_bs_query
279bs_bs_query
280bs_bs_query
281bs_bs_query
282bs_bs_query
283bs_bs_query
284bs_bs_query
285bs_bs_query
286bs_bs_query
287bs_bs_query
288bs_bs_query
289bs_bs_query
290bs_bs_query
291bs_bs_query

3.3.2.4. Protease test. Hydrolysis of proteins around colonieswas achieved by the three isolates (Fig. 2D) indicating prote-ase activity for all the isolates.
3.3.2.5. Amylase test. Clear zone around colonies was achievedby the three isolates (Fig. 2F) indicating the ability for all of themto degrade starch.
3.3.3. Microscopic characteristicsGenerally, the isolated strains have all morphological fea-tures recorded previously to other H. werneckii strains andmentioned briefly in the introduction. Table 2 summarizes themorphological differences among the isolated black yeast
strains. EGYNDA08 was studied as a model for the develop-mental stages of hyphal maturation. Newly developedhyphae appeared as non-septated coenocytic thin hyphae(Fig. 3D1).
Polar septum formation started from the older parts of thefilaments at the colony base and continued to the tip of thefilaments and the intercalary cells become wider by age andfollowed by conidiogenesis (Fig. 3D2). Two weeks old fila-ments terminated conidiogenesis and intercalary cells startedto divide leading to the irregular size and shape of cells. Frag-mentation has also been noticed in old filaments (Fig. 3D3) thensclerotial elements production was the final step in the cycle(Fig. 3D4).
Table 2 – Morphological characteristics of H. werneckii strains isolated from salt marshes.
Characteristic Strain
EGYNDA08 EGYNDA16 EGYNDA90
Cell length 5.2–7.8 μm 5.2–46.8 μm 5.2–20.8 μmCell width 2.6–5 μm ~2.6 μm 2.6–5.2 μmSpherical cells Not present 10.4 μm 6.5–7.8 μmCell description Two-celled pattern, conspicuous and heavily
melanized thick septum and cell wall. (Fig. 3A).Melanosomes were low in numbers and large insize, they were located inside and outside thecell (Fig. 3B1), anastomosis triangles in septumare pigmented (Fig. 3B2).
Two shapes of cells: rod andspherical (Fig. 4A). Cell wall,septum and melanization wereweak (Fig. 4A). Many finemelanosomes.
Cell wall and septum weremedium in thickness andmelanizationMelanosomes were many andfine (Fig. 5A).
Hyphal development Short mucilage Rare, if present, zigzag non-septated (Fig. 4C).
Zigzag coenocytic (Fig. 5B). Long,albino aerial mycelium (Fig. 2D).
Budding style Polar (Fig. 3C1), sided polar (Fig. 3C2), bipolar(Fig. 3C3) and sympodial (Fig. 3C4)
Mainly polar and other styleswere rare
Mainly polar and other styleswere rare
Annellidic ring Few rings max. 3 (Fig. 3D2) Many rings (Fig. 4B1, B2 and B3). Medium number of rings
Fig. 3 – EGYNDA08 isolate under light microscope (40×). (A) The most frequent morphological characteristics (two-celledpattern, conspicuous septa and melanized cell wall. (B1) Bold arrows showing the cytoplasmic melanosomes aggregatedinside and outside the cell. (B2 and B3) Light arrows referring to the accumulation of melanin position. (C1–C4) Differentstyles of budding cells (C1) polar, (C2) sided polar, (C3), bipolar (C4) sympodial. (D1–D4) Filaments at different ages. (D1) Fivedays old hyphae, (D2) ten days old hyphae, arrows are pointing to annellidophores, (D3) twenty days old petri-dish boldarrow pointing to fragmented filament and double-headed arrow is pointing to the same number of vesicles inside twocells and inside fragmented hyphae, (D4) one month old culture with irregular sclerotial elements.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
5e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
292bs_bs_query
293bs_bs_query
294bs_bs_query
295bs_bs_query
296bs_bs_query
297bs_bs_query
298bs_bs_query
299bs_bs_query
300bs_bs_query
301bs_bs_query
302bs_bs_query
303bs_bs_query
304bs_bs_query
305bs_bs_query
306bs_bs_query
307bs_bs_query
308bs_bs_query
309bs_bs_query
310bs_bs_query
311bs_bs_query
312bs_bs_query
313bs_bs_query
314bs_bs_query
315bs_bs_query
316bs_bs_query
317bs_bs_query
318bs_bs_query
319bs_bs_query
320bs_bs_query
321bs_bs_query
322bs_bs_query
323bs_bs_query
324bs_bs_query
325bs_bs_query
326bs_bs_query
327bs_bs_query
328bs_bs_query
329bs_bs_query
330bs_bs_query
331bs_bs_query
332bs_bs_query
333bs_bs_query
334bs_bs_query
335bs_bs_query
336bs_bs_query
337bs_bs_query
338bs_bs_query
339bs_bs_query
340bs_bs_query
341bs_bs_query
342bs_bs_query
343bs_bs_query

4. Discussion
The biggest obstacle to the halophilic black yeasts study is notlaying in extreme conditions but in their inability to prohibitthe invasion of other competing microorganisms that is similarto the behavior of the acidophilic fungi [21]. Additionally, iso-lating H. werneckii was a big challenge [4] because a successfulpurification depends on choosing the unsuitable isolation con-ditions to other fungi contaminating the sample and in thesame time tolerable by the organism of interest such as highsalt concentration in media [4], high pH >9 and low tempera-ture <20 °C. Media type, temperature and media state affectedthe examined shape and the stages of Hortaea cells under mi-croscope, even the temperature in which they are growingaffected the relativity between hyphal and yeast phase. Yeast
phase was more frequently observed and could be describedas the dominant phase at low temperatures (15–20 °C), however,hyphal phase was the dominant phase at high temperatures(25–30 °C). When a four-day-old colony was picked and exam-ined by light microscope, it showed the different growth phasesexcept the sclerotial bodies. On the other hand, the isolatesexamined from the liquid media were much more homog-enous cells with almost the same shape and size.
Morphological differences were observed and recorded bycareful comparison between the three isolates cultivated underthe same laboratory conditions to avoid the gradual changeand loss in the unnecessary morphological characteristics ofthe tested lab organisms over the research study period.
The general environmental or controlled conditions sur-rounding black yeasts are majorly responsible for sculpturingtheir morphological identity so culturing the organisms re-peatedly in controlled conditions probably leads to differentphenotype. As an example, EGYNDA16 was rarely observed toproduce filaments until it was cultured repeatedly on solidmedia at which wide spaces between spots were consideredindicating that the laboratory conditions and the cultivationmethod forced the cultured organism to produce hyphae whichmight be necessary for the organism to obtain nutrients.
The yeast-like pattern gave a wide variety in budding styles:polar and sided polar budding (Fig. 3C1 and C2) which was trig-gered by growing on solid media may be when cells on thesurface of colony were budding vertically downward then hori-zontally outward. Bipolar budding (Fig. 3C3) maybe due tonuclear division occurrence in one mother cell and each nucleiordered its half with budding from each pole before a sepa-rating septum is formed. Sympodial budding (Fig. 3C4) wastriggered in old cultures. Hyphal branches’ irregularity has beenmentioned as a cause to prolonged culturing of the organismin liquid culture [22]. The characteristic shape of zigzag hyphaehas been noticed in Ashbya gossypii and the dimorphic patho-genic fungus Histoplasma capsulatum as the studies proved thatthe hyphae shape might be affected with calcium availabilityin surrounding media [23] or calcium binding protein gene mu-tation [24].
Fig. 4 – EGYNDA16 isolate under light microscope (40×). (A) Black bold arrows are pointing to big cells while thin arrows arepointing to long cells. (B1–B3) Annellidic budding and brackets are referring to annellidic conidiogenesis, arrows arepointing to the original cell (B3) which was potential for shedding off endoconidia. (C) Isolated hyphaea from broth media.
Fig. 5 – EGYNDA90 isolate under light microscope. (A) Blackbold arrow is pointing to a relatively big cell while thinarrow is pointing to long cells. (B) Zigzag-shaped branchedhyphae growing on the figure after few days; filament isstraightened once it gets septated.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
6 e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
Q7 Q8
344bs_bs_query
345bs_bs_query
346bs_bs_query
347bs_bs_query
348bs_bs_query
349bs_bs_query
350bs_bs_query
351bs_bs_query
352bs_bs_query
353bs_bs_query
354bs_bs_query
355bs_bs_query
356bs_bs_query
357bs_bs_query
358bs_bs_query
359bs_bs_query
360bs_bs_query
361bs_bs_query
362bs_bs_query
363bs_bs_query
364bs_bs_query
365bs_bs_query
366bs_bs_query
367bs_bs_query
368bs_bs_query
369bs_bs_query
370bs_bs_query
371bs_bs_query
372bs_bs_query
373bs_bs_query
374bs_bs_query
375bs_bs_query
376bs_bs_query
377bs_bs_query
378bs_bs_query
379bs_bs_query
380bs_bs_query
381bs_bs_query
382bs_bs_query
383bs_bs_query
384bs_bs_query
385bs_bs_query
386bs_bs_query
387bs_bs_query
388bs_bs_query
389bs_bs_query
390bs_bs_query
391bs_bs_query
392bs_bs_query
393bs_bs_query
394bs_bs_query
395bs_bs_query
396bs_bs_query
397bs_bs_query
398bs_bs_query
399bs_bs_query
400bs_bs_query
401bs_bs_query
402bs_bs_query
403bs_bs_query
404bs_bs_query
405bs_bs_query
406bs_bs_query
407bs_bs_query
408bs_bs_query

It could be concluded that new black yeast strains have beenisolated and characterized in this study.They all have been iden-tified as H. werneckii, two of them EGYNDA08 and EGYNDA16were ecotypes of each other and EGYNDA90 lay in a separatebranch. Up to our knowledge, this is the first report describ-ing the occurrence of this organism in Egypt.The isolates showdifferences in morphological characteristics as described in theresults and the enzymatic assays showed qualitative differ-ences. Further studies are ongoing to verify the pleomorphiccharacterization and the adaptation mechanism of these iso-lates to the saline habitat they could survive.
Acknowledgments
Many thanks to Prof. Samia Ali Haroun, the former head ofbotany department, Faculty of Science, Mansoura University,Egypt and many thanks to Prof. Heshmat Soliman Aldesuquythe current head of the department for their valuable help andencouragement. We thank Dr. Eladl Gala from the same de-partment for his continuous support and valuable discussions.
R E F E R E N C E S
[1] De Hoog G, Zalar P, Urzi C, De Leo F, Yurlova N, Sterflinger K.Relationships of dothideaceous black yeasts andmeristematic fungi based on 5.8 S and ITS2 rDNA sequencecomparison. Stud Mycol 1999;31–7.
[2] Sterflinger K, De Hoog G, Haase G. Phylogeny and ecology ofmeristematic ascomycetes. Stud Mycol 1999;5–22.
[3] Ajello L. The black yeasts as disease agents: historicalperspective. Pan Am Health Organ Sci Pub 1978;356:9–19.
[4] Sterflinger K. Black yeasts and meristematic fungi: ecology,diversity and identification Biodiversity and ecophysiologyof yeasts. Springer; 2006. p. 501–14.
[5] de Hoog GS. Evolution of black yeasts: possible adaptation tothe human host. Antonie Van Leeuwenhoek 1993;63(2):105–9.
[6] Vaupotic T, Gunde-Cimerman N, Plemenitas A. Novel 3’-phosphoadenosine-5’-phosphatases from extremelyhalotolerant Hortaea werneckii reveal insight into moleculardeterminants of salt tolerance of black yeasts. Fungal GenetBiol 2007;44(11):1109–22.
[7] Revankar SG, Sutton DA. Melanized fungi in human disease.Clin Microbiol Rev 2010;23(4):884–928.
[8] de Hoog GS, Gerrits van den Ende AH. Nutritional patternand eco-physiology of Hortaea werneckii, agent of humantinea nigra. Antonie Van Leeuwenhoek 1992;62(4):321–9.
[9] Mittag H. The fine structure of Hortaea werneckii. Mycoses1993;36(11–12):343–50.
[10] Gunde-Cimermana N, Zalarb P, de Hoogc S, Plemenitasd A.Hypersaline waters in salterns – natural ecological nichesfor halophilic black yeasts. FEMS Microbiol Ecol2000;32(3):235–40.
[11] Mullis KB. The unusual origin of the polymerase chainreaction. Sci Am 1990;262(4):56–61.
[12] Muzyka D, Pantin-Jackwood M, Stegniy B, Rula O, Bolotin V,Stegniy A. Wild bird surveillance for avian paramyxovirusesin the Azov-black sea region of Ukraine (2006 to 2011)reveals epidemiological connections with Europe and Africa.Appl Environ Microbiol 2014;80(17):5427–38.
[13] Kejzar A, Gobec S, Plemenitas A, Lenassi M. Melanin iscrucial for growth of the black yeast Hortaea werneckii in itsnatural hypersaline environment. Fungal Biol2013;117(5):368–79.
[14] Taylor WI, Achanzar D. Catalase test as an aid to theidentification of Enterobacteriaceae. Appl Microbiol1972;24(1):58–61.
[15] Rustigian R, Stuart CA. Decomposition of urea by Proteus.Proc Soc Exp Biol Med 1941;47(1):108–12.
[16] Sierra G. A simple method for the detection of lipolyticactivity of micro-organisms and some observations on theinfluence of the contact between cells and fatty substrates.Antonie Van Leeuwenhoek 1957;23(1):15–22.
[17] Kazanas N. Proteolytic activity of microorganisms isolatedfrom freshwater fish. Appl Microbiol 1968;16(1):128–32.
[18] Vedder EB. Starch agar, a useful culture medium. J Infect Dis1915;16(3):385–8.
[19] Stewart JC, Stewart CS, Heptinstall J. The use of tritiatedcellulose in screening for cellulolytic microorganisms.Biotechnol Lett 1982;4(7):459–64.
[20] Shahriarinour M, Mohd Noor A, Ariff A, Mohamad R.Screening, isolation and selection of cellulolytic fungi fromoil palm empty fruit bunch fibre. Biotechnology2011;10(1):108–13.
[21] Selbmann L, de Hoog GS, Zucconi L, Isola D, Ruisi S, van denEnde AH. Drought meets acid: three new genera in adothidealean clade of extremotolerant fungi. Stud Mycol2008;61:1–20.
[22] Takeo K, De Hoog G. Karyology and hyphal characters astaxonomic criteria in ascomycetous black yeasts andrelated fungi. Antonie Van Leeuwenhoek 1991;60(1):35–42.
[23] Bauer Y, Knechtle P, Wendland J, Helfer H, Philippsen P. ARas-like GTPase is involved in hyphal growth guidance inthe filamentous fungus Ashbya gossypii. Mol Biol Cell2004;15(10):4622–32.
[24] Tian XB, Shearer G. The mold-specific MS8 gene is requiredfor normal hypha formation in the dimorphic pathogenicfungus Histoplasma capsulatum. Eukaryot Cell2002;1(2):249–56.
ARTICLE IN PRESS
Please cite this article in press as: Ashraf Elsayed, Amr M. Mowafy, Hoda M. Soliman, Ahmed Gebreil, Nada I. Magdy, Characterization of new strains of Hortaea werneckiiisolated from salt marshes of Egypt, Egyptian Journal of Basic and Applied Sciences (2016), doi: 10.1016/j.ejbas.2016.09.001
7e g y p t i an j o u rna l o f b a s i c and a p p l i e d s c i e n c e s ■■ ( 2 0 1 6 ) ■■ –■■
409bs_bs_query
410bs_bs_query
411bs_bs_query
412bs_bs_query
413bs_bs_query
414bs_bs_query
415bs_bs_query
416bs_bs_query
417bs_bs_query
418bs_bs_query
419bs_bs_query
420bs_bs_query
421bs_bs_query
422bs_bs_query
423bs_bs_query
424bs_bs_query
425bs_bs_query
426bs_bs_query
427bs_bs_query
428bs_bs_query
429bs_bs_query
430bs_bs_query
431bs_bs_query
432bs_bs_query
433bs_bs_query
434bs_bs_query
435bs_bs_query
436bs_bs_query
437bs_bs_query
438bs_bs_query
439bs_bs_query
440bs_bs_query
441bs_bs_query
442bs_bs_query
443bs_bs_query
444bs_bs_query
445bs_bs_query
446bs_bs_query
447bs_bs_query
448bs_bs_query
449bs_bs_query
450bs_bs_query
451bs_bs_query
452bs_bs_query
453bs_bs_query
454bs_bs_query
455bs_bs_query
456bs_bs_query
457bs_bs_query
458bs_bs_query
459bs_bs_query
460bs_bs_query
461bs_bs_query
462bs_bs_query
463bs_bs_query
464bs_bs_query
465bs_bs_query
466bs_bs_query
467bs_bs_query
468bs_bs_query
469bs_bs_query
470bs_bs_query
471bs_bs_query
472bs_bs_query
473bs_bs_query
474bs_bs_query
475bs_bs_query
476bs_bs_query
477bs_bs_query
478bs_bs_query
479bs_bs_query
480bs_bs_query
481bs_bs_query
482bs_bs_query
483bs_bs_query
484bs_bs_query
485bs_bs_query
486bs_bs_query
487bs_bs_query
488bs_bs_query
489bs_bs_query
490bs_bs_query
491bs_bs_query
492bs_bs_query
493bs_bs_query
494bs_bs_query
495bs_bs_query
496bs_bs_query
497bs_bs_query
498bs_bs_query
499bs_bs_query
500bs_bs_query
501bs_bs_query
502bs_bs_query
503bs_bs_query
504bs_bs_query
505bs_bs_query
506bs_bs_query
507bs_bs_query
508bs_bs_query
509bs_bs_query
510bs_bs_query
511bs_bs_query





























