Embed Size (px)
Citation preview

TIG ~ October 1988, Vol. 4, no. I0
The powerful genetic procedures that can be applied to the yeasts enable these organisms to be used for the investigation of a wide variety of cell and molecular biology prob- lems. Unfortunately, many of these procedures are either time consum- ing or impossible to apply to mamma- lian cells. Recessive mutants cannot be readily isolated in diploid cell lines and subsequent genetic analysis is hampered by long generation times, karyotype instability, and the limita- tions of somatic cell genetics. Man- ipulation of genes in marnw~li,n chro- mosomes by site-specific mutagen- esis has also proved to be very diflicu]t. However, the ability to transfer genes between yeasts and mammalian cells opens up new approaches that allow the powerful yeast genetic procedures to be applied to mamm~li~u genes. These approaches have been particularly useful for investigating ras gene function. In this article we describe some of the gene functions involved in cell cycle control of the fission yeast Schizosacclmromyces pombe and consider how these new approaches will be useful for investigating the cell cycle in nqh~zlxmli~n cells.
Two cell cycle controls Two major cell cycle controls operate during the
fission yeast cell cycle t'z. The first acts in late G1 just before S phase and is called start 3. Once past this point, cells become committed to the mitotic cell cycle, and the alternative developmental fate of conjugation cannot take place until the cell cycle in progress has been completed. Two gene functions encoded by cdc2 + and cdclO + are required for traverse of start Lz. Temperature-sensitive mutants in these two genes arrest in the cell cycle before start when incubated at the restrictive temperature (Fig. 1).
The second control acts in late (;2 and determines when mitosis is initiated during the cell cycle 4,s. Four gene functions encode components acting at this control: cdc2 +, wed +, cdc25 + and niml +. Mutations in these genes lead to cells being advanced into mitosis at a reduced siz~ 1'2'4-e (Fig. 1).
c d c 2 + encodes a 34 kDa protein kinase The cdc2 + gene plays a major role in cell cycle control,
being required at both the start and mitotic control points. The gene, which has four introns, has the potential to encode a 34kDa protein with significant homologies to protein kinases ~. Antibodies raised against the cdc2 + gene product detect a 34 kDa phosphoprote~n called p34 ~dcz which has protein kinase activity in vitro. In addition, specific mutants that disrupt regions of i)34 °~c2 thought to be important for this activity abolish cdc2 + gene function 9. These experiments suggest that protein phosphorylation plays a key role in cell cycle control.
Thec~c~otein kinase activity and phosphorylation state of p34 is correlated with the proliferative state of the cells s. When cells are arrested before start by nitrogen deprivation, p34 cdcz loses its protein kinase activity and also becomes dephosphorylated. When nitrogen is re-fed to the cells, p34 cdcz regains its protein kinase activity after a lag consistent with the notion that its activity is required for progress through the cell cycle. Therefore © 198& F.bev~r Pu~catio~ Cambridge 01~8- SS,~SSt$02.e0
review Cell cycle control genes in
fission yeast and mammalian cells Melanie Lee and Paul Nurse
The study of the cell cycle of Schizosaccharomyces pombe has identified many genes involoed in eulemyo~ cell cycle control. Recent ooservations suggest that there is a remarkable conservation of these genes and controlling elements in mammalian ceils, and have kigkl~hted the power of yeast genetics for investigation of mammalian
gene function.
regulation of the p34 ~d~ protein kinase could be part of the mechanism by which the cell controls exit from and entry into the cell cycle. The protein kinase activity could be determined by the phosphorylation state of the protein, but, alternatively, dephosphorylation may be a consequence rather than the caase of the fall in protein kinase activity.
Casein has been used as the in vitro substrate for the p34 c~2 protein ldnase assays but its in vivo substrates are unknown. Various proteins, such as histones and lamins, which are involved in chromatin and nucleoskeletai organization, undergo changes in phosphorylation during the cell cycle. Therefore the p34 ~ protein kinase could initiate entry into S phase and mitosis at the two control points by phosphorylating such proteins ~. It is clearly important to establish what the in vivo substrates are, and possible genetical approaches to this problem are indicated later.
¢ d c 2 + is regulated by other genes encoding protein kinase homologues
Three other ger~.s, wed +, cdc25 '~ and niml +, have been ~@ficated in the control of mitosis initiation 6'1°'11. Over-production of cdc25 + transcripts leads to mitotic advancement, whereas deletion of the gene blocks mitosis s. This in,cares that cdc25 + encodes a positive element or inducer in the mitotic control. The opposite results are seen with wed +, indicating that this gene
Fig. 1. S. pombe cells showing various phenotypes: wild- type cell dividing at normal size (centre cell), cell advanced into mitosis dividing at reduced size (upper cell), cell arrested in cell cycle progress and unable to divide (lower cell). 287

r iews encodes a negative element or inhibitor in the mitotic control 1°. The third gene, niml +, also encodes a mitotic inducer, since over-production of its transcripts ad- vances cells into mitosis u. A further gene product, encoded by win1+, also interacts with the functions of weel + and cdc25 + (Ref. 12). Mutants of niml have no effect on the mitotic control in the absence of weel +, suggesting that the niml + gene product acts upstream as an inhibitor of the wee1 + gene product, and thus as an inducer in the overall mitotic control.
Mutations that result in specifically altered p34 °~ca proteins lead to an advancement of cells into mitosis. They fall into two classes, one producing cells largely unresponsive to changes in wee1 + gene activity, and a second producing cells largely unresponsive to changes in cdc.25 +gcne activity. These results suggest that the signals generated by the wee1 + and cdc25 + gene products act independently and upstream of the cdc2 + gene prod- uct. These data and other experiments discussed in Refs 10, 11 c~n be understood in terms of the scheme shown in Fig. 2.
Sequencing indicates that cdc.25 +, wee1 + and him1 + encode proteins of 67 kDa, 96 kDa and 42 kDa, respec- tively. In addition, the weel + and him1 + sequences have significant homologies to protein kinases, suggesting that tl~ee protein kinases act in a regulatory sequence in the control of mitosis. The him1 + protein kinase would inhibit the activity of the weel + protein kinase which in turn would inhibit the activity of the cdc2 + protein kinase. Hewever, it is not clear wilether the protein kinases act directly on each other or if there are other intermediate steps.
Other functions interacting with cdc2 + Two other gene functions, encoded by sucl + and
cdc13 +, have been shown to interact closely with that of cdc2 +. Mutations in sucl +, and over-production of sucl+gene transcripts, allow certain temperature- sensitive mutants of cdc2 + to grow at their restrictive temperaturet3,~4. Because this suppression is restricted
nim 1 PK I
el Iw. 1,,I [ I
el I
cdc25 I (win 1)
cdc2PK J ( s u c l , cdc 13)
G2 [ Mitotic Initiation
Fig. 2. Scheme of mitotic control in S. pombe . Proposed regulatory relationships of mitotic control elements, based on genetic and biochemical analysis. The n i m I + ~no cdc25 + genes encode activators and the wee1 + gene an inhibitor; n im 1 ÷ exerts its positive influence by inhibiting the negative element wee1 +" win1 + probably interacts with cdc25 + and suc l + and cdc13 + probably interact with cdc2 +.
T I C , - October 1988, to',. 4, no. 10
to only certain mutant alleles, it has been argued that the sunl + and cdd?, + gene products directly interact with each other. Evidence in favour of this possibility has been obtained using antibodies raised against the predicted 13kDa sun/+ gene product 14. An affinity column us~g the anti-p!3 suc~ ant]3mdy binds about 5% of the p34 °~-~ protein 14;ls. Thus there may be a direct physical associa- tion between p13 eucl and p34 ec~2 in vivo.
The c~13 + gene function has been shown to interact closely with that of cdc2 + (Ref. 16). A cold-sensitive ode2 ~ mutant allele that is defective solely at the second control point at mitosis can be suppressed by a cdcl3 mutant, and, in addition, a cdcl3 ~ lethal mutant can be rescued at the restrictive temperature by over-production of cdc2 + gene transcripts. The roles of sunl + and cdcl3 + are not known, but given their close interactions with the cdc2 ÷ gene product, they could encode regulators or substrates of the p34 °a~2 protein kinase.
C D C 2 8 i s functionally equivalent to cdc2 + in budding yeas t
The equivalent gene to cdc2 + in the budding yeast Saccharomyces cerevisiae is CDC28 (Ref. 17). This gene function is required at a start control in G1, and there is also a report that, like cdc~ +, it is required again later in the cell cycle for mitosis. The cdc2 + and CDC28 genes are functionally interchangeable, encode proteins that differ in length by only one amino acid, and share 62% identity in amino acid sequer~ce. The CDC28 gene product also has protein kinase activity ts. The activity was assayed in an immunoprecipitate using as substrate a 40 kDa protein co-precipitated from the cell extracts; it is possible that this protein is a CDC28 substrate in vivo.
On nutritional depletion leading to cell cycle arrest before start, p34 cv .zs loses its protein kinase activity, behaviour similar to that described for p34 cdcz. Analysis of changes in the p34 cDczs protein kinase activity during the cell cycle suggests that activity peaks during G1 at around start Is. If cells are withdrawn from the cell cycle or proceed to later stages in the cell cycle, the protein kinase activity is much reduced. Because the substrate used in the assays is also derived from the cells it is difficult to know ff the c ~h~, ges in ~ctivity are related to changes in the substrate, in activating or inhibitory factors, or in p34 cnc2s protein itself. However, clearly there are cyclical changes in overall !)34 cDc2s protein kinase activity correlated with traverse through the cell cycle. Some of the p34 cvc2s protein is found in an insoluble component, leading to the suggestion that the p34 cnc2s is a component of the. cytoskeleton is.
Humvn homologue of c d c 2 + Given the central role of the cdc2 +/CDC28 gene for
cell cycle control in yeasts, it was of interest to establish whether a similar gene function exists in other eukary- otes, including mammalian cells. The human homologue of cdc2 + was cloned by expression of a human cDNA library in a cdc2 ts mutant and selection for cDNA clones complementing the defective cdc2 ts function 2°. The cDNA library was made from actively proliferating cells by Okayarna and Berg, using a simian virus 40 (SV40) expression vector. The SV40 promoter works in S. pombe, and as a consequence the human cDNA library was well expressed in the yeast cells. Out of a million transfonnants screened, five could grow at the restric- tive temperature, and each of these contained a cDNA
T I G - Oc¢~ber 1988, Vol. 4, no. 10
GO G1 Mitotic cell cycle review
R/W points p ~ Pm/Ps MPF
_ . . . ..1_ Mitosis_.jL
7
Multistep signal pathways in response to growth factors etc.
CDC2 CDC2
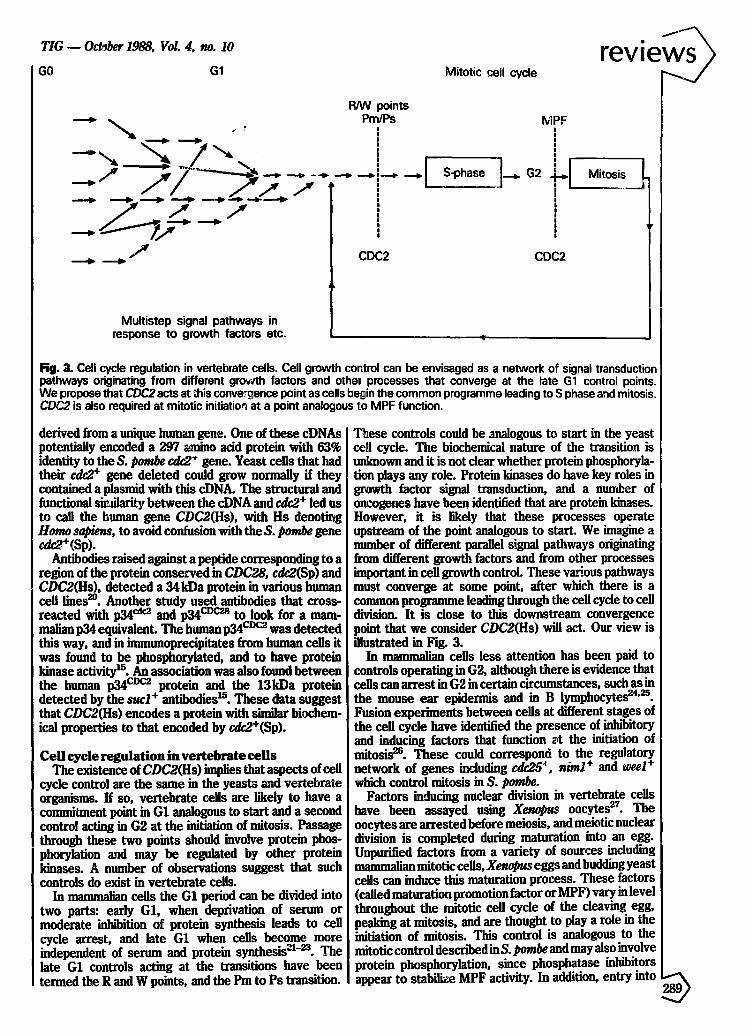
Fig. 3. Cell cycle regulation in vertebrate cells. Cell growth cor.tml can be envisaged as a network of signal transduction pathways originating from different growth factors and othe~ processes that converge at the late G1 control points. We propose that CDC2 acts at ;his conve~ence point as cells begin the common programme leading to S phase and mitosis, CDC2 is also required at mitotic initiation at a point analogous to MPF function.
derived from a unique human gene. One of these cDNAs potentially encoded a 297 ~Lmino acid protein with 63% identity to the S. pombe c.dc2 + gene. Yeast cells that had their cdc2 + gene deleted could grow normally if they contained a plasmid with this cDNA. The structural and functional siv, tilarity between the cDNA and cdc2 + led us to call the human gene CDC2(Hs), with Hs denoting Homo sapiens, to avoid com~usion with the S. pombe gene cdc2+(Sp).
Antibodies raised against a peptide corresponding to a region of the protein conserved in CDC28, cdc2(Sp) and CDC2(Hs), detected a 34kDa protein in various human cell lines z°. Another study used antibodies that cross- reacted with p34 cs~ and p34 c~c~s to look for a mam- malian p34 equivalent. The hmnan p34 c°c~ was detected this way, and in immunoprecipitates from human cells it was found to be phosphorylated, and to have protein kinase activity Is. An association was also found between the human p34 cvc2 protein and the 13kDa protein
+ 15 detected by the sue1 antibodies . These data suggest that CDC2(Hs) encodes a protein with s~rnilnr biochem- ical properties to that encoded by cdc2+(Sp).
Cell cycle regulation in vertebrate cells The existence of CDC2(Hs) implies that aspects of cell
cycle control are the same in the yeasts and vertebrate organisms. If so, vertebrate cells are likely to have a commiUnent point in G1 analogous to start and a second control acting in (32 at the initiation of mitosis. Passage through these two points should involve protein phos- phorylation and may be regulated by other protein kinases. A number of observations suggest that such controls do exist in vertebrate cells.
In mammalian cells the G1 period can be divided into two parts: early G1, when deprivation of serum or moderate inhibition of protein synthesis leads to cell cycle arrest, and late G1 when cells become more independent of serum and protein synthesis "~l-m. The late G1 controls acting at the transitions have been termed the R and W points, and the Pin to Ps transition.
T~ese controls could be analogous to start in the yeast ce~l cycle. The biochemical nature of the transition is unlmown and it is not clear whether protein phosphoryla- tion plays any role. Protein kinases do have key roles in 8t~wth factor signal transduction, and a number of oncogenes have been identified that are protein kinases. However, it is likely that these processes operate upstream of the point analogous to starL We imagine a number of different parallel signal pathways originating from different growth factors and from other processes important in cell growth control These various pathways must converge at some point, after which there is a common programme leading through the cell cycle to cell division. It is close to this downstream convergence point that we consider CDC2(Hs) will act. Our view is illustrated in Fig. 3.
In mammalian cells less attention has been paid to controls operating in (;2, although there is evidence that cells can arrest in (;2 in certain circumstances, such as in the mouse ear epidermis and in B lymphocytes u'2s. Fusion experiments between cells at different stages of the cell cycle have identified the presence of inhibitory and inducing factors that function ~t the initiation of mitosis 26. These could correspond to the regulatory network of genes including cdc25 +, niml + and weel + which control mitosis in S. pombe.
Factors inducing nuclear division in vertebrate cells have been assayed using X~opus oocytes ~7. The oocytes are arrested before meiosis, and meiotic nuclear division is completed during maturation into an egg. Unpurified factors from a variety of sources including malnm~linrl mitotic cells, Xenopus eggs and budding yeast cells can induce this maturation process. These factors (called maturation promotion factor or MP F) vary in level throughout the mitotic cell cycle of the cleaving egg, peaking at mitosis, and are thought to play a role in the initiation of mitosis. This control is analogous to the mitotic control described in S. pombe and may also involve protein phosphorylation, since phosphatase inhibitors appear to stabilize MPF activity. In addition, entry into ®

r iews mitosis is accompanied by changes in the phosphorylation of a number of proteins L,t animal cells.
Recently, MPF has been highly purified fromXenopus and found to contain two major protein components, one of approximately 32 kDa and a second of 45 kDa 2s. The smaller component is immunoblotted and immunoprecipi- tater by antibodies raised against the fission yeast cdc2* gene product (see Monitor by Peter Fantes in this issue). This result strongly suggests that p34 cdcz is a component of MPF and that cdc2 homologues will piay an tmportant role in mitotic control of vertebrate cells.
Mammalian genes in simple eukaryotes The ability to identify h~.:~mologous genes from mamma-
lian cells by function, using simple eukaryotes such as S. pombe, opens up new approaches for the analysis of the cell cycle in mammals. Once a functional homologue has been isolated from mammalian cells the simple eukaryote can be used as a surrogate organism for genetical investigation of the mammalian gene. Of particular interest are dominant mutations, which can be isolated using a further copy of the mammalian gone introduced into the simple eukaryote on a plasmid ~. Dominant mutations can block the proper function of the wild-type gone product and it may be possible to use such mutations to knock out gone functions in mammalian cells. Suppres- sors of dominant mutations can also be isolated, allowing other mammalian gene functions that interact with the original gene to be identified. These procedures should be useful for identifying in rive substxates of p34 cdcz and of the other protein kinases involved in its regulation.
A further advantage of these approaches will be in the functional analysis of gene sequences. Because of the evolutionary distance between simple eukaryotes and mammafian cells, considerable divergence can be expected between homologous gone sequences, and therefore conserved regions are likely to represer, t protein domains that are essential for the gone function. Evolution has carried out the mutagenesis experiments necessary to establish the important regions for function. For example, cdc2 + encodes a protein kinase and con- tains consensus sequences that are found in all protein kinases. Clearly, these cannot be important in estab- lishing the specific function of the p34 cecz protein kinase with respect to regulators and substrates. However, a comparison of the cdc2 + homologues from the yeasts and mammalian sources can be used to identify those regions which are common to all the homologues but differ from other protein kinases. These regions act as the 'finger
, cdc2 prints that define the p34 specificity. An example is 2o the perfectly conserved peptide EGVPSTAIREISLLKE ,
antibodies against which have detected a 32--36 kDa protein in a wide range of eukaryotic cells.
The approaches outlined above can be used for the investigation of gone functions required in any cellular process, and should be useful for bypassing some of the problems associated with genetical analysis in mammalian cells.
References I Nurse, P. (1985) Trends Genet. 1, 51-55 2 Hayles, J. and Nurse, P. (1986) ]. Cell Sci. 4 (Suppl.), 155-170 3 Nurse, P. and Bissett, Y. (1981) Nature 292, 558-.~0 4 Nurse, P. 0975)Nature 256, 547-,558 5 Fantes, P. A. (1983)Nature 302, 153-155 6 Russell, P. and Nurse, P. (1986) Cell 45, 145-153 7 Hindley, J. and Phear, G. A. (1984) Gene 31, 129-134
T I C , - October 1988, Vol. 4, no. I0
8 Simanis, V. and Nurse, P. (1986) Cell 45, 261-268 9 Booher, Ih and Beach, D. (1986) Moi. Cell. B/o/. 6, 3523--3530
10 Russell, P. and Nurse, P. (1987) Cell 49, 559-567 11 Russell, P. and Nurse, P. (1987) Cell 49, 569-578 12 Ogden, J. E. and Fantes, P. A. (1986) Curt. Genet. 10, 509-514 13 Hayles, J., Ayes, S. and Nurse, P. (1986)EMBOJ. 5, 3373-3379 14 Brianela, L., Draetta, G. and Beach, D. (1987) E M B O ]. 6,
3507--3514 15 Drsetta, G., Bfianela, L., Potashkin, J. and Beach, D. (1987)Cell
50, 319-325 16 Booher, R. and Beach, D. (1987) E M B O ] . 6, 3441-3447 17 Beach, D., Durkacz, B. andNurse, P. (1982)Nature300, 706-709 18 Reed, S. I., Madwiger, J. A. and Larincz, A. (1985) Prof. Natl
Aead. Sd. USA 82, 405,5-4059 19 Mendenhall, M., Jones, C. and Reed, S. (1987) Cell 50, 927--935 20 Lee, IV[. 6. and Nurse, P. (1987)Nature 327, 31--35 21 Pardee, A. B., Danbrow, R., Hamlin, J. L. and Kletzien, IL F.
(1978) Annuo Rev. Biofhem. 47, 715--750 22 Zetterberg, A~ and L~_sson, O. (1985) Prof. NatlAraut Sd . USA
82, 5365-,5369 23 Pledger, W., Stiles, C., Antoiades, IL and Scher, C. (1978) Prof.
Nati Acad. 8c£ USA 82, 7681-7685 24 Gelfant, S. (1962) Exp. Cell Res. 26, 395-403 25 Melchers, F. and Lemlmrdt, W. (1985) Prof. NatlAcad. 5¢i. USA
82, 7681-7685 26 Rao, P. and Jolmson, R. (1970) Nature 225, 159--164 27 Kirsdmer, M., Newport, J. and Gerbart, J. (1985) Trenda Goner.
I, 41-47 28 Lehka, M. J., Hayes, M. K. and M~ller, J. L. (1988) Prec. Natl
Acad. Sci. USA 85, 3009-3013 29 Mendeniml], M. D., Richardson, H. E. and Reed, S. [. (1988)
Prof. Natl Acad~ Sd. ~ ' t 1 5 , 4426-4430
M. Lee and P. Nurse are in the ICRF Cell Cycle Control Laboratory, Microbiology Unit, Department of Biodwmistry, University of Oxford, South Parks Road, Oxford OXI 3QU, UK; M. Lee is currently in the DOarlment of Genetics, Glaxo Group Research Ltd, Greenford UB6 0HE, UK.
SUBSCRIPTION NEWS!!! Trends Journals Subscriptions are now handled
from the Elsevier, Barking, UK office.
Al l new subscriptions, renewals and subscription
queries should now be addressed to:
Trends Subscriptions Department. A Elsevier Science Publishers Ltd ~F~ ~ ' ~ Crown House, c ~ o J Linton Road, ~ ~ " Barking, Essex, IGI 1 8JU, ~ ~-~ ,~ / UK J
t t ~ L a
Tel: +44 (1) 594 7272 ~
From USA and Canada all Trends subscriptions ~ r " ' 7 will still be forwarded in the usual way / ~ _ _ ~ . ~ , . f by writing to: Trends Subscriptions, JIC. L ? f ~ ~ - Elsevier Science Publishing Co Inc. ,,~ v -- 52 Vanderbilt Avenue, New York.





























