Embed Size (px)
Citation preview

Gastroenterology 2014;146:995–1005
BASIC AND TRANSLATIONAL—ALIMENTARY TRACT
CD36- and GPR120-Mediated Ca2D Signaling in Human Taste BudCells Mediates Differential Responses to Fatty Acids and Is Alteredin Obese MiceMehmet Hakan Ozdener,1,* Selvakumar Subramaniam,2,* Sinju Sundaresan,3 Omar Sery,4
Toshihiro Hashimoto,5 Yoshinori Asakawa,5 Philippe Besnard,2 Nada A. Abumrad,3 andNaim Akhtar Khan2
1Monell Chemical Senses Center, Philadelphia, Pennsylvania; 2UMR U866 INSERM, Dijon, France; 3Center for Human Nutritionand Department of Cell Biology and Physiology, Washington University, St Louis, Missouri; 4Academy of Science, Brno, CzechRepublic; and 5Faculty of Pharmaceutical Sciences, Tokushima Bunri University, Tokushima, Japan
BASICAN
DTR
ANSLAT
IONA
LAT
BACKGROUND & AIMS: It is important to increase our un-derstanding of gustatory detection of dietary fat and itscontribution to fat preference. We studied the roles of the fattaste receptors CD36 and GPR120 and their interactions viaCa2þ signaling in fungiform taste bud cells (TBC). METHODS:We measured Ca2þ signaling in human TBC, transfected withsmall interfering RNAs against messenger RNAs encoding CD36and GPR120 (or control small interfering RNAs). We alsostudied Ca2þ signaling in TBC from CD36�/� mice and fromwild-type lean and obese mice. Additional studies were con-ducted with mouse enteroendocrine cell line STC-1 that expressGPR120 and stably transfected with human CD36. Wemeasured release of serotonin and glucagon-like peptide-1from human and mice TBC in response to CD36 and GPR120activation. RESULTS: High concentrations of linoleic acidinduced Ca2þ signaling via CD36 and GPR120 in human andmice TBC, as well as in STC-1 cells, and low concentrationsinduced Ca2þ signaling via only CD36. Incubation of human andmice fungiform TBC with lineoleic acid down-regulated CD36and up-regulated GPR120 in membrane lipid rafts. Obese micehad decreased spontaneous preference for fat. Fungiform TBCfrom obese mice had reduced Ca2þ and serotonin responses,but increased release of glucagon-like peptide-1, along withreduced levels of CD36 and increased levels of GPR120 in lipidrafts. CONCLUSIONS: CD36 and GPR120 have nonoverlappingroles in TBC signaling during orogustatory perception ofdietary lipids; these are differentially regulated by obesity.
Keywords: Serotonin; Linoleic Acid; GLP-1; Lipids.
ral perception of dietary fat was, until recently,
*Authors share co-first authorship.
Abbreviations used in this paper: [Ca2D]i, intracellular calcium; ER,endoplasmic reticulum; GA, grifolic acid; GLP-1, glucagon-like peptide-1;GPCRs, G protein-coupled receptors; HFD, high-fat diet; IP3, inositol-tris-phosphate; LA, linoleic acid; LCFA, long-chain fatty acid; siRNA, smallinterfering RNA; SOC, store-operated Ca2D; TBC, taste bud cells; WT, wildtype.
© 2014 by the AGA Institute0016-5085/$36.00
http://dx.doi.org/10.1053/j.gastro.2014.01.006
Othought to involve mainly texture and olfactorycues; however, accumulating evidence strongly suggests theexistence of a taste modality devoted to the detection oflong-chain fatty acids (LCFA).1 Mice can recognize dietaryfat and FA solutions in the oral cavity in the absence ofolfactory or textural cues.2 Humans can taste LCFA evenwhen textural properties are masked and olfaction is elim-inated using a nose clip.2
CD36 is highly expressed apically on lingual gustatoryepithelium in rats and mice3,4 and CD36 deletion completely
abolishes the spontaneous preference of wild-type mice forLCFA.4 In mouse circumvallate taste bud cells (TBC), LCFA-mediated activation of CD36 released Ca2þ from the endo-plasmic reticulum (ER) via a phospholipase-C (PLC)�dependent mechanism.5 In addition, LCFA via CD36 inducedthe phosphorylation of Src kinases, which regulate theopening of store-operated Ca2þ (SOC) channels in TBC. TheSOC channels in the mouse TBC are composed of orai1/3proteins, and are under the control of stromal interactionmolecule-1, which orchestrates ER Ca2þ sensing andrelease.6 Stromal interaction molecule-1 might play a keyrole in fat perception, as its deletion eliminates the mice’sspontaneous preference for fat.6
Similar to CD36, the GPR120 and GPR40 members ofG-protein�coupled receptors (GPCRs) were shown tomediate the taste responses to FAs.7 Both GPR120 andGPR40 knock-out mice showed a diminished preference forlinoleic and oleic acids with reduced taste nerve responses.GPR40 expression was undetectable in rat8 and humanTBC9 and only GPR120, is expressed in type II cells, the truetaste receptor cells of the lingual epithelium.7
The findings to date suggest that TBC CD36 andGPR120 are the potential candidates for lipid tasteperception.1,10 However, information on their relative rolesremains largely unavailable. Why are there 2 receptorcandidates for 1 taste?11 There is evidence in mice thatTBC CD36 and GPR120 might respond differently todietary fat. During a meal, lingual CD36 expression isdown-regulated, which is paralleled by reduction of fatpreference.12 In contrast, no changes are observed in
996 Ozdener et al Gastroenterology Vol. 146, No. 4
BASICAND
TRANSLATIONALAT
GPR120 expression in circumvallate papillae during themeal,12 suggesting that this receptor plays an alternativerole distinct from that of CD36. The current study wasundertaken to test the hypothesis that CD36, which hashigh affinity for LCFA, might function as the primary re-ceptor for fat taste detection, and GPR120 would operateto amplify signaling mainly under excess FA supply. Forthis we elucidated Ca2þ signaling pathways, triggered byLCFA interaction with CD36 or GPR120 vs a specificagonist of GPR120 in human fungiform TBC. Selective smallinterfering RNA (siRNA) silencing of CD36 and GPR120was used to dissect their respective roles. The data werefurther validated using TBC from CD36�/� mice and STC-1endocrine cells, endogenously expressing GPR120 andtransfected with human CD36. The effect of high-fat feedingon CD36 and GPR120 expression levels and membranedistribution were examined in fungiform TBC from leanand obese mice. Lipid gustatory preference, TBC Ca2þ
signaling, and the associated release of serotonin andglucagon-like peptide-1 (GLP-1) were determined.
Materials and MethodsMaterials
Human13–15 and C57BL/6J mice TBC5,6 were prepared andmaintained as reported previously. All studies adhered toprotocols approved by the Schulman Associates InstitutionalReview Board (Cincinnatti, OH) for human TBC and by theRegional Ethical Committee of Dijon (France) and the AnimalStudies Committee of Washington University (St Louis, MO) formice TBC. Cell culture media were from Lonza Verviers(Belgium) and Fura-2/AM from Invitrogen (Carlsbad, CA) (seeSupplementary Materials).
Measurement of Ca2þ SignalingIsolated TBC suspended in fresh Iscove’s modified Dulbec-
co’s medium containing 10% fetal bovine serum were seeded(2 � 105/well) onto Willico-Dish wells.6 Changes in intracel-lular Ca2þ ([Ca2þ]i) were monitored using a Nikon microscope(TiU) equipped with EM-CCD (Luca-S) camera for real-timerecording of digital images and an S-fluor 40� oil immersionobjective (Nikon, Tokyo, Japan). Planes were taken at Z in-tervals of 0.3 mm and the images analyzed using NIS-Elementssoftware (Nikon, Tokyo, Japan). Changes in [Ca2þ]i wereexpressed as DRatio, calculated as the difference between thepeak F340/F380. The data were averaged for 20�40 individualcells per run from 3�9 experiments with at least 3 cell prep-arations. For experiments in Ca2þ-free medium, CaCl2 wasreplaced by ethylene glycol-bis(b-aminoethyl ether)-N,N,N0,N0-tetraacetic acid (2 mM).
Statistical AnalysisData are presented as mean � SEM. Significance of differ-
ences between mean values was determined by one-wayanalysis of variance (Statistica, Statsoft, France), followed by aleast-significant difference test.
ResultsHuman Fungiform TBC Co-express GPR120and CD36
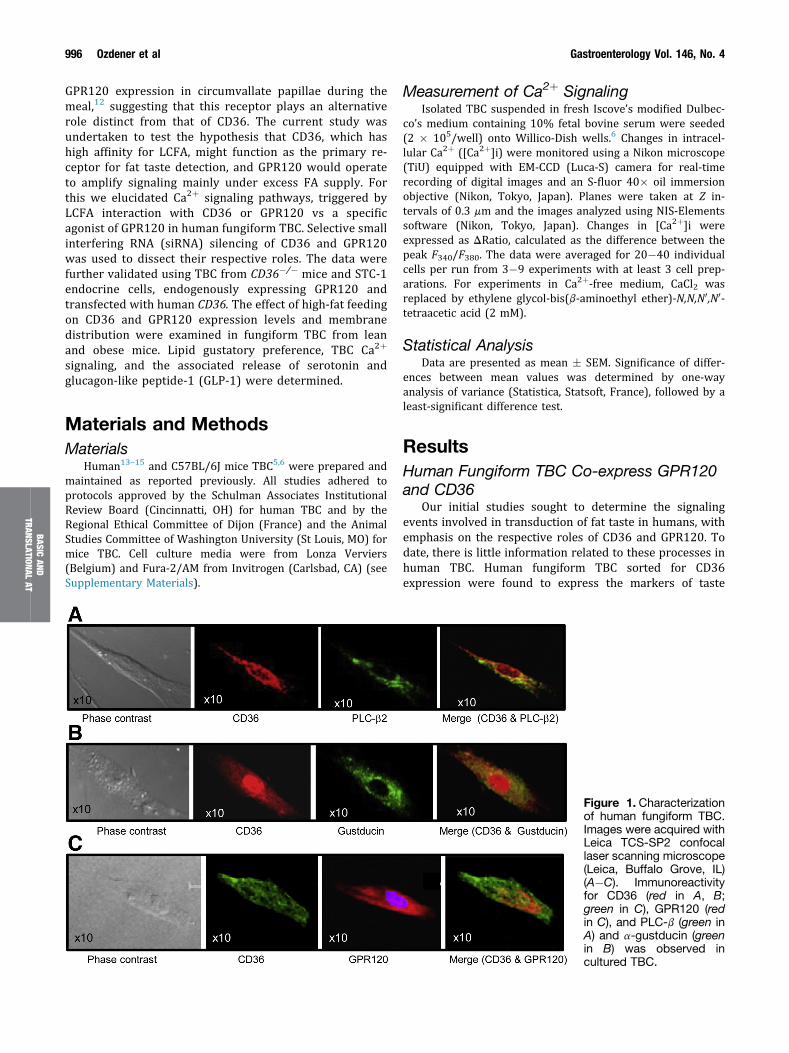
Our initial studies sought to determine the signalingevents involved in transduction of fat taste in humans, withemphasis on the respective roles of CD36 and GPR120. Todate, there is little information related to these processes inhuman TBC. Human fungiform TBC sorted for CD36expression were found to express the markers of taste
Figure 1. Characterizationof human fungiform TBC.Images were acquired withLeica TCS-SP2 confocallaser scanning microscope(Leica, Buffalo Grove, IL)(A�C). Immunoreactivityfor CD36 (red in A, B;green in C), GPR120 (redin C), and PLC-b (green inA) and a-gustducin (greenin B) was observed incultured TBC.

April 2014 Ca2+ Signaling in Human Taste Bud Cells 997
receptor cells PLC-b2 and a-gustducin (Figure 1A and B)and all CD36 expressing TBC also co-expressed GPR120(Figure 1C).
Linoleic Acid and Grifolic Acid InduceCapacitative Ca2þ Signaling in HumanTBC Through Protein Tyrosine Kinaseand G-Protein�Dependent Mechanisms
Two taste receptor ligands, linoleic acid (LA) and grifolicacid (GA), were used for the fat taste studies to help deter-mine specificity of agonist responses and sensitivity tovarious inhibitors. LA, used by us and others,1,2 is primarily aCD36 ligand that is abundantly present in vegetable oil and istherefore a representative fatty component of the Westerndiet. GA, purified by Hara et al16 from the edible mushroomAlbatrellus dispansus, is a selective GPR120 ligand based onstudies of Ca2þ signaling in enteroendocrine STC-1 cells.16
The addition of LA to human fungiform TBC at a con-centration of 20 mM induced a rise in [Ca2þ]i (Figure 2A
Figure 2. Effects of LA and GA on Ca2þ signaling in human fuloaded with Fura-2/AM, and the changes in intracellular Ca2þ (FCa2þ-containing (A�F) and in Ca2þ-free media (B, D). In panels[Ca2þ]i evoked, respectively by LA and GA. In panels E and Fincubated (20 minutes) with SU6656 (5 mM), PP2 (10 mM),2-aminoethoxydiphenyl borate (2-APB; 30 mM), pertussis toxinS; 300 mM), or methyl-b-cyclodextrin (M-bCD; 2.5 mM). Data ar
and B), and a similar effect was observed with 20 mM GA(Figure 2C and D). LA- and GA-induced increases in [Ca2þ]iwere higher in Ca2þ-containing, as compared withCa2þ-free, medium (Figure 2B and D). In the presence ofCa2þ, blockers of SOC channels (ie, SKF96365, 2-amino-ethoxydiphenyl borate, and econazole) significantly dimin-ished LA-induced increases in [Ca2þ]i (Figure 2E; P < .001)and, to a lesser extent, that induced by GA (Figure 2F;P < .001). Different inhibitors of protein tyrosine kinases(ie, PP2, genistein, and SU6656) markedly (60%�90%)curtailed the rise of [Ca2þ]i after LA (Figure 2E; P < .001),and significant but less pronounced (20%�40%) inhibitionwas observed with GA (Figure 2F; P < .001).
Pertussis toxin, a G-protein inhibitor, and guanosine-50-O-2-thiodiphosphate, a nonhydrolyzable guanosine diphosphateanalogue, significantly diminished both LA- (30% and 60%)and GA-evoked (40% and 30%) increases of [Ca2þ]i in humanTBC (Figure 2E and F). Disruption of lipid rafts by methyl-b-cyclodextrin also decreased Ca2þ signaling in the presenceof either LA (30%) (Figure 2E) or GA (50%) (Figure 2F).The sulfo-N-succinimidyl derivative of oleate, which binds
ngiform TBC. The cultured TBC (2 � 105 cells/assay) were340/F380) were monitored. The experiments were performed inA and C, the colored time-lapse images show the changes in, TBC before exposure to 20 mM LA (E) or GA (F) were pre-genistein (30 mM), SKF96365 (15 mM), econazole (30 mM),(PTX; 10 ng/mL), guanosine-50-O-2-thiodiphosphate (GDP-b-e mean � SEM (n ¼ 7). *P < .001 as compared with control.
BASICAN
DTR
ANSLAT
IONA
LAT

998 Ozdener et al Gastroenterology Vol. 146, No. 4
BASICAND
TRANSLATIONALAT
lysine-164 in CD36 and blunts LCFA-induced Ca2þ signaling,17
curtailed (75%) LA-induced increase in [Ca2þ]i (F340/F380ratio, 0.21 � 0.016 vs 0.06 � 0.003; P < .001).
LA and GA Induce Serotonin and GLP-1Release in Human TBC
We previously showed that mice TBC respond to LCFAby releasing serotonin (5-hydroxytryptamine), and thisrelease is dependent on the rise of [Ca2þ]i.5 In humanfungiform TBC, LA (20 mM) and GA (20 mM) both triggeredrelease of serotonin, with GA (61.3% � 2.48% above con-trol; P < .001) being more potent than LA (36.4% � 1.39%above control; P < .001).
GLP-1 and its receptor, GLP-1R, have been identified inmouse TBC, suggesting their involvement in taste percep-tion.18 The addition of 20 mM LA or GA to human TBCinduced a small and equivalent release of GLP-1 (LA: 103.06� 1.61 pmol per 1 � 106 cells vs GA: 102.93 � 1.45 pmolper 1 � 106 cells) that significantly (P < .001) exceeded theGLP-1 released by control TBC (plus 0.1% v/v ethanolvehicle) (90.29 � 1.20 pmol of GLP-1 per 1 � 106 cells).
LA and GA Induce Additive Effects on Ca2þ
Signaling and IP3 Production in Human TBCLA and GA exerted an additive response on Ca2þ
signaling (Supplementary Figure 1A�C). GA-induced rise of
[Ca2þ]i exceeded that triggered by LA both at 20 mM or100 mM (Supplementary Figure 1B and C). Consistent withthis, production of IP3, which triggers release of ER Ca2þ
and subsequent SOC Ca2þ influx, was higher in GA-treatedTBC as compared with LA-treated cells (SupplementaryFigure 2). In addition, IP3 release in response to LA andGA combined exceeded that observed with each alone(Supplementary Figure 2).
Knockdown of CD36 and GPR120 ExertAdditive Suppression of Ca2þ Signaling inHuman TBC
To dissect the CD36- vs GPR120-specific signaling re-sponses, we used siRNA knockdown to selectively diminishthe expression of these proteins in human fungiform TBC(Figure 3A). CD36 siRNA-transfected TBC had a reduced(approximately 45%) Ca2þ response to 20 mM LA, with noinfluence on that induced by 20 mM GA (Figure 3B). TBCtransfected with GPR120 siRNA had a small but significantdecrease (approximately 18%) in the LA-induced Ca2þ rise,with a much stronger reduction (approximately 75%) in theGA-induced response (Figure 3B). Combined transfectionwith CD36 and GPR120 siRNA suppressed LA-induced Ca2þ
signaling by about 80%, but caused no additional reductionsin the response to GA as compared with cells transfectedwith GPR120 siRNA alone (Figure 3B).
Figure 3. Effects of siRNAtargeting CD36 or GPR120on LA- and GA-evokedCa2þ signaling in humanfungiform TBC. (A) Westernblots showing selectivedown-regulation of CD36and GPR120 by siRNAtargeting CD36 and/orGPR120. Mock: non-targeting siRNA. b-actin isthe loading control. (B�E)Increases in [Ca2þ]iinduced by either LA or GAin TBC transfected by thevarious siRNA. Data aremeans � SEM (n ¼ 4)conducted in triplicates.*P < .001 as comparedwith control.

April 2014 Ca2+ Signaling in Human Taste Bud Cells 999
LA at Low Concentrations Induces Ca2þ
Signaling Only via CD36 in Human andMouse TBC
LA at 5 mM failed to raise [Ca2þ]i in human TBC, trans-fected with CD36 siRNA (Figure 3C and D). In TBC trans-fected with GPR120 siRNA, LA (5 mM), but not GA (5 mM),induced a significant rise of [Ca2þ]i (Figure 3C and E).
To strengthen these observations, we conducted exper-iments on mice fungiform TBC, isolated from CD36 null mice(CD36�/�). LA at 5 mM increased [Ca2þ]i in wild-type (WT)mice TBC, and no rise was observed in TBC from CD36�/�
mice (Figure 4A). LA at 20 mM evoked in CD36�/� TBC asmall rise in [Ca2þ]i, which was substantially less (70%)than that observed in TBC from WT mice (Figure 4B). Therewas no difference in GA signaling to [Ca2þ]i between TBCfrom WT or CD36�/� mice (Figure 4C). Addition of 20 mMGA after that of 20 mM LA failed to induce an additive rise of[Ca2þ]i in TBC from CD36�/� mice (Figure 4D), indicatingthat LA is an effective competitor of GA for GPR120 acti-vation of Ca2þ signaling.16 In TBC from WT mice, the addi-tion of LA and then GA triggered an additive response(Figure 4E) in line with LA primarily interacting with CD36(Figure 4B), while GA only activates GPR120 (Figure 4C).
BASICAN
DAN
SLAT
IONA
LAT
LA and GA Evoke Differential Ca2þ Signalingin Enteroendocrine STC-1 Cells
To confirm these observations, we analyzed the changesin [Ca2þ]i in STC-1 cells. STC-1 cells express GPR120, butnot CD36, and a line stably expressing human CD36 wasgenerated19 (Supplementary Figure 3A) to dissect the rela-tive roles of the 2 receptors.
Figure 4. Effects of CD36 deficiency on LA- and GA-evoked Cwere performed on fungiform TBC from WT or CD36�/� mice,Figure 2. The changes in [Ca2þ]i shown in response to (A) LA 5 m20 mM.The data are representative of 4�5 experiments conduc
In cells lacking CD36, LA at 5 mM did not increase [Ca2þ]i(Figure 5A), and a response was observed in CD36-expressing cells (Figure 5B), which was not altered byGPR120 knock-down (Figure 5C). LA at 20 mM evoked asmall rise in [Ca2þ]i in STC-1 cells expressing GPR120, butlacking CD36 (Figure 5D). A strong rise in [Ca2þ]i wasobserved in cells expressing CD36 (Figure 5E), which wasnot affected by GPR120 knock-down (Figure 5F). Pretreat-ment with the CD36 inhibitor sulfo-N-succinimidyl deriva-tive of oleate curtailed LA-induced increases in [Ca2þ]i inSTC-1 cells expressing CD36 (Supplementary Figure 3B).
LA Alters Distribution of CD36 andGPR120 in Membrane Rafts of Humanand Mouse TBC
CD36 is localized in membrane microdomains known asrafts,20 although less is known about membrane distributionof GPR120. Because localization in lipid rafts can impactCa2þ signaling by influencing G-protein activation and IP3production,21,22 the effect of LA on CD36 and GPR120membrane distribution was explored. Treatment of humanor mice fungiform TBC for 20 minutes with LA did not alterprotein levels of CD36 and GPR120 (not shown), but didcause changes in membrane distribution. In human fungi-form TBC, CD36 and GPR120 were present in both raft(fractions 3�6) and nonraft (fractions 7�10) triton-solublemembranes (Figure 6A�C). LA treatment of human fungi-form TBC decreased CD36 level in the rafts without affectinglevel in the soluble fractions (Figure 6A�C). In contrast, LAexposure increased GPR120 levels in the raft fractions(Figure 6A and B).
a2þ signaling in mouse fungiform TBC. Ca2þ imaging studiesand the changes in [Ca2þ]i (F340/F380) were monitored as forM; (B) LA 20 mM; (C) GA 20 mM; (D&E) LA 20 mM; and then GAted in triplicate.
TR

Figure 5. Effects of LA andGA on Ca2þ signaling inSTC-1 cells. The studieswere performed on STC-1cells that endogenouslyexpress GPR120 but lackCD36 (GPR120þ/CD36�),that stably express humanCD36 (GPR120þ/CD36þ),or that express CD36and are transfected withGPR120 siRNA (GPR120�/CD36þ). The changes inintracellular Ca2þ (F340/F380) are shown in re-sponse to 5 mM LA (A-C)or 20 mM LA (D-F). Thedata are representative of4–5 experiments conduct-ed in triplicate.
1000 Ozdener et al Gastroenterology Vol. 146, No. 4
BASICAND
TRANSLATIONALAT
Comparable results were obtained with mouse fungi-form TBC (Figure 6D�F), as LA treatment was associatedwith less CD36 and more GPR120 (Figure 6D and E) in theraft fractions.
High-Fat�Fed Obese Mice Have ReducedFat Preference With Abnormalities of TBCCa2þ Signaling, Release of Serotonin, andGLP-1 and Altered CD36 and GPR120Membrane Raft Distribution
To gain insight into the physiological role of FA-inducedCa2þ signaling, we explored whether it is altered duringobesity. Studies were conducted with mice fed a high-fatdiet (HFD) for 2 months. This regimen resulted in 20%more body weight gain as compared with controls fedstandard chow and in hyperglycemia (glucose levels: 1.1 �0.8 g/L in control mice vs 1.75 � 0.05 g/L in HFD mice). GAand LA increased [Ca2þ]i in the TBC of mice fed the standarddiet as expected (Figure 7A). The HFD reduced the Ca2þ
response to LA, and it increased that induced by GA(Figure 7A). Expression of messenger RNA encoding CD36,GPR120, and a-gustducin was not altered by the HFD (notshown). However, CD36 protein level in both raft andnonraft fractions decreased (Figure 7C�F), and moreGPR120 distributed to lipid rafts in the TBC of HFD fed mice(Figure 7C and E).
Control mice fed the standard diet exhibited a strongpreference for 0.1% canola oil as compared with a control
solution (Figure 7B), and this preference was diminishedsignificantly in HFD-fed obese mice (Figure 7B).
LA-induced serotonin release was reduced in TBC fromHFD as compared with standard diet�fed mice(Supplementary Figure 4A). However, GA-activated releaseof serotonin was not altered (Supplementary Figure 4A).Interestingly, GLP-1 release triggered by GA was enhancedin TBC from HFD as compared with standard diet�fed mice(Supplementary Figure 4B), and LA-stimulated GLP-1release was unchanged (Supplementary Figure 4B). Thissuggested that GLP-1 release is mediated by GPR120 and itis facilitated by recruitment of GPR120 to lipid rafts.
DiscussionThe current study showed that human fungiform TBC
sorted for CD36 expression also express GPR120 togetherwith the markers of taste receptor cells a-gustducin andPLC-b. Both CD36 and GPR120 are implicated in orogusta-tory perception of dietary lipids.1,4,6,7 Expression,messenger RNA and protein, of CD363,4 and GPR1208,12 hasbeen reported in circumvallate, foliate, and fungiformpapillae of rodents. However, this is the first documentationof their coexpression in individual human TBC.
Our study evaluated Ca2þ signaling, an early eventdownstream of taste receptor activation, in response to anLCFA, LA, a ligand of CD36 and presumably of GPR120, ascompared with GA, a selective GPR120 agonist.16 Both LAand GA induced increases in IP3 production and [Ca2þ]i,

Figure 6. Effects of LA exposure on the distribution of CD36 and GPR120 in raft fractions in human (A�C) and mouse (D� F)fungiform TBC. The TBC were treated or not (control) with 20 mM LA for 20 minutes. (A, D) Cells were homogenized in 1%Triton X-100 at 4�C, and the lysates were subjected to discontinuous 5%�40% sucrose gradient centrifugation. Differentfractions were collected from the top of the gradient and equal volumes of aliquots from each fraction were subjected toWestern blot. Histograms show the relative band intensity (arbitrary units) measured by densitometry of protein content in raft(3�6) and soluble fractions (7�10). The data were normalized with respect to band intensity of caveolin-1 (Cav-1) for raftfractions and b-actin for soluble fractions, measured under similar conditions. Panels B and C are derived from panel A, andpanels E and F are derived from panel D. Data are means � SEM conducted in triplicate.
April 2014 Ca2+ Signaling in Human Taste Bud Cells 1001
BASICAN
DTR
ANSLAT
IONA
LAT
suggesting they acted to mobilize Ca2þ from the ER via aPLC-dependent mechanism, possibly via PLC-b present inhuman fungiform TBC. LA-evoked Ca2þ signaling wassignificantly curtailed in the presence of sulfo-N-succini-midyl derivative of oleate, indicating that this FA mobilizedCa2þ, in large part, by binding to CD36.5,23 Like in mouseTBC,5 CD36 in human fungiform TBC appears coupled toprotein tyrosine kinase and Ca2þ signaling.
As reported in the mouse,5,6 the current study confirmsinvolvement of SOC channels in the Ca2þ response to LCFAby human fungiform TBC. Disruption of G-proteins alsodiminished the Ca2þ signaling responses to both LA and GA.GPR120 has been documented to elevate [Ca2þ]i via inter-acting with the Gaq/11 family of G proteins,24 but the link
between CD36 and G-proteins is less clear.24 CD36 signalingmight induce IP3 generation directly via phosphorylation ofPLC by the CD36 interacting src kinase, which has beenreported to phosphorylate the PLCg isoforms.25
Treatment of human or mice fungiform TBC for 20 mi-nutes with LA did not significantly down-regulate totalCD36 protein, which has been shown to become ubiquiti-nated and degraded after FA treatments.19 Chevrot et al26
reported lipid-mediated down-regulation of CD36 inmouse TBC after a 60-minute exposure and Tran et al27
reported its down-regulation in the small intestine 2hours after lipid intake. CD36 down-regulation wouldrepresent a feedback mechanism aiming to reduce FA acti-vation of CD36 signaling. Although this down-regulation

Figure 7. HFD-inducedobesity modulates Ca2þ
signaling, gustatory fatpreference, and CD36 andGPR120 raft distribution inmouse fungiform TBC. Thefungiform TBC were iso-lated from mice fed astandard chow diet (Std.Diet) or a HFD for 2months. (A) Changes in[Ca2þ]i. (B) Spontaneousfat preference of mice fedstandard diet or HFD.Values are means � SEM(n ¼ 6). *P < .001; oil ascompared with controlsolution. (C, D) Distributionof CD36 and GPR120 inraft (R, 3�6) and soluble(S, 7�10) fractions frommice fed standard diet orHFD. Caveolin-1 (cav-1) isa raft marker. (E, F) Rela-tive densitometry quanti-fying data (arbitrary units)derived from panels C andD after normalization forband intensity of cav-1 forraft fractions and b-actinfor soluble fractions. Dataare means � SEM con-ducted in triplicate.
1002 Ozdener et al Gastroenterology Vol. 146, No. 4
BASICAND
TRANSLATIONALAT
was not measurable after a short 20-minute FA treatment, itis a likely contributor to the reduction of TBC CD36 levelsthat we observed in HFD-fed obese mice.
Cholesterol depletion of plasma membrane rafts cur-tailed LA- and GA-evoked increases in [Ca2þ]i in humanfungiform TBC, suggesting that raft integrity plays animportant role in the regulation of Ca2þ signaling. Mem-brane rafts are domains, rich in cholesterol and sphingoli-pids, that provide a platform for the assembly of signalingcomplexes.28 In mouse TBC, clustering of sweet taste re-ceptors into the rafts is thought to enhance sweetener actionby facilitating tastant-triggered G-protein�coupled receptorsignaling.29 Studies in which lipid rafts were disrupted orreconstituted showed these domains to be indispensable forsweet receptor responsiveness to sucralose.29 Similarly, the
presence of CD36 in lipid rafts has been reported to beimportant for CD36-mediated downstream signaling, asreviewed recently.30
Our findings support the interpretation that both CD36and GPR120 are functional taste receptors in humanfungiform TBC. LA and GA both induced the release of se-rotonin, as was previously shown for LA with mice TBC.5,6
GA-induced serotonin release by human TBC exceededthat triggered by LA and it paralleled the higher GA-inducedrise in [Ca2þ]i, consistent with the direct coupling of Ca2þ
signaling to serotonin secretion.31 In this context, the higherrelative potency of 20 mM GA vs 20 mM LA in human TBC,and not in mouse TBC, might reflect a dietary effect on TBCCD36 in humans. Human intake of dietary fat often exceeds40% as compared with <6% for mice fed standard diets,

April 2014 Ca2+ Signaling in Human Taste Bud Cells 1003
BASICAN
DTR
ANSLAT
IONA
LAT
which would be expected to down-regulate CD36 level andfunction in human TBC. Future studies will explore thispossibility.
CD36 and GPR120 are structurally different glycopro-teins predicted to have 2 and 7 transmembrane segments,respectively. Although both recognize LCFA, they markedlydiffer in affinity for the FA, with CD36 exhibiting muchhigher affinity28 than GPR120.32 Our data indicate thatCD36 is the primary LCFA receptor in TBC. Selective knock-down of either CD36 or GPR120 in human fungiform TBCshowed that LA at low concentration induces Ca2þ signalingvia CD36 and not through GPR120. LA at a low concentra-tion also failed to increase [Ca2þ]i in TBC obtained fromCD36�/� mice. Although a high concentration of LA trig-gered a rise in [Ca2þ]i, the response was much smaller thanthat obtained in WT mice. In contrast to CD36, which rec-ognizes low FA levels, GPR120 only responds to high FAconcentrations and the response obtained is modest. Thisdifferential sensitivity was also observed in STC-1 cellsexpressing GPR120, but not CD36,33 which were unre-sponsive to 5 mM LA (or oleic acid or docosahexaenoic acid,data not shown). STC-1 responsiveness to LCFA was ac-quired with CD36 expression. High concentrations of LA(20 mM) evoked only a slight rise of [Ca2þ]i in CD36-negative STC-1 cells and, although the LA response wasincreased 4-fold by CD36 expression, it was minimallyreduced by knock-down of GPR120. These data support theinterpretation that CD36 is necessary for fat taste detectionat physiological FA concentrations. This would be consistentwith the finding that CD36�/� mice lose their fat preference4
and that down-regulation of CD36 in circumvallate papillaeeliminates fat preference in the absence of changes inGPR120.34 Together our observations suggest that GPR120,being poorly activated by LCFA, might act downstream ofLCFA receptors, such as CD36 (and possibly others), toamplify the response to high concentrations of tastants,including dietary FA. This interpretation would be consis-tent with GPR120 expression in a variety of taste cellsresponsive to sweet, bitter, and umami stimuli.35 Interest-ingly, a recent study in humans showed that non-FA ago-nists of GPR120 (such as GA in our study) do not elicit ataste response similar to that of LA, despite the finding thatthey activate the glossopharyngeal nerve in mice.36 There-fore, although CD36 would associate with oral LCFArecognition and “taste detection,” GPR120 might function insignal amplification for the “sustained sensing” of food.
Our data with both human and mice fungiform TBCshow that acute exposure to LCFA decreases membranelipid raft localization of CD36 while it increases that ofGPR120. This effect is greatly exaggerated by consumptionof a HFD, which also decreases total CD36, probablyreflecting the effect of chronic exposure to excess FA.Localization in membrane rafts as compared with nonraftdetergent-soluble domains would modulate IP3 release andCa2þ signaling,21,22 events integral to TBC CD36 andGPR120 function. In addition to modulating CD36 mem-brane localization, FA exposure would regulate CD36internalization and ubiquitination,30 possibly involving theactivation of CD36-associated src kinases.37
Our data indicate that HFD, by resulting in low CD36expression and distribution to lipid rafts, diminishes LCFA-induced Ca2þ signaling and serotonin release. The releasedserotonin in mice TBC has been shown to act downstream ofCa2þ signaling to transmit the fat taste response to afferentnerve fibers.5,6 Diminished release of this neurotransmittertoward synaptic clefts would contribute to the reduced oralsensitivity to FA in obesity. Consistent with this, subjectswith attenuated expression of CD36, as a result of geneticvariants, exhibit decreased sensitivity for orogustatoryperception of dietary lipids.38
How the HFD-induced changes in CD36 and GPR120level and signaling might influence the etiology or pro-gression of obesity remains to be determined. Stewartet al39 have reported that the ability to detect oleic acid bothorally and within the gastrointestinal tract is compromisedin obese men, suggesting that the decreased sensitivitymight promote more fat intake to reach the same tasteperception. However, direct proof for this is lacking andawaits additional investigation.
In contrast to its effect on CD36, the HFD increased raftlocalization of GPR120, which might constitute a compen-satory event serving to enhance signal amplification, asshown for other GPCR proteins.40 This amplification couldinvolve, at least in part, the enhanced release of GLP-1shown in obese mice. GLP-1 receptor knockout mice havereduced taste responses to sweeteners18 and dietary fat34 inbehavioral tests. GLP-1 signaling is involved in normaldiurnal down-regulation of CD36 by dietary fat in the mousecircumvallate papillae,34 contributing to progressivereduction of fat taste sensitivity during a meal and possiblymodifying feeding behavior.34
In summary, our data provided new information on therespective signaling functions of CD36 and GPR120 in oro-gustatory fat perception. It would be of interest to extendthe studies to the highly innervated circumvallate papillae.Our data documented the mechanistic changes underlyingthe hyposensitivity to lipid taste observed in obesity. Theyshould guide future studies into the mechanisms respon-sible for the hedonic preference for fat that associates withhigh-fat consumption that has been reported in obesesubjects.41
Supplementary MaterialNote: To access the supplementary material accompanyingthis article, visit the online version of Gastroenterology atwww.gastrojournal.org, and at http://dx.doi.org/10.1053/j.gastro.2014.01.006.
References
1. Khan NA, Besnard P. Oro-sensory perception of dietarylipids: new insights into the fat taste transduction. Bio-chim Biophys Acta 2009;1791:149–155.
2. Mattes RD. Is there a fatty acid taste? Annu Rev Nutr2009;29:305–327.
3. Fukuwatari T, Kawada T, Tsuruta M, et al. Expression ofthe putative membrane fatty acid transporter (FAT) in

1004 Ozdener et al Gastroenterology Vol. 146, No. 4
BASICAND
TRANSLATIONALAT
taste buds of the circumvallate papillae in rats. FEBS Lett1997;414:461–464.
4. Laugerette F, Passilly-Degrace P, Patris B, et al. CD36involvement in orosensory detection of dietary lipids,spontaneous fat preference, and digestive secretions.J Clin Invest 2005;115:3177–3184.
5. El-Yassimi A, Hichami A, Besnard P, et al. Linoleic acidinduces calcium signaling, SRC-kinase phosphorylationand neurotransmitters release in mouse CD36-positivegustatory cells. J Biol Chem 2008;283:12949–12959.
6. Dramane G, Abdoul-Azize S, Hichami A, et al. STIM1regulates calcium signaling in taste bud cells and pref-erence for fat in mice. J Clin Invest 2012;122:2267–2282.
7. Cartoni C, Yasumatsu K, Ohkuri T, et al. Taste prefer-ence for fatty acids is mediated by GPR40 and GPR120.J Neurosci 2010;30:8376–8382.
8. Matsumura S, Mizushige T, Yoneda T, et al. GPRexpression in the rat taste bud relating to fatty acidsensing. Biomed Res 2007;28:49–55.
9. Galindo MM, Voigt N, Stein J, et al. G protein-coupledreceptors in human fat taste perception. Chem Senses2012;37:123–139.
10. Abumrad NA. CD36 may determine our desire for dietaryfats. J Clin Invest 2005;115:2965–2967.
11. Abdoul-Azize S, Selvakumar S, Sadou H, et al. Ca2þ
signaling in taste bud cells and spontaneous preferencefor fat: unresolved roles of CD36 and GPR120. Biochimie2014;96:8–13.
12. Martin C, Passilly-Degrace P, Gaillard D, et al. The lipid-sensor candidates CD36 and GPR120 are differentiallyregulated by dietary lipids in mouse taste buds: impacton spontaneous fat preference. PLoS One 2011;6:e24014.
13. Ozdener MH, Brand JG, Spielman AI, et al. Character-ization of human fungiform papillae cells in culture. ChemSenses 2011;36:601–612.
14. Ozdener MH, Rawson NE. Primary culture of mammaliantaste epithelium. Methods Mol Biol 2013;945:95–107.
15. Ozdener H, Spielman AI, Rawson NE. Isolation and cul-ture of human fungiform taste papillae cells. J Vis Exp2012;63:e3730.
16. Hara T, Hirasawa A, Sun Q, et al. Novel selective ligandsfor free fatty acid receptors GPR120 and GPR40. NaunynSchmiedebergs Arch Pharmacol 2009;380:247–255.
17. Kuda O, Pietka TA, Demianova Z, et al. Sulfo-N-succi-nimidyl oleate (SSO) inhibits fatty acid uptake andsignaling for intracellular calcium via binding CD36 lysine164. SSO also inhibits oxLDL uptake by macrophages.J Biol Chem 2013;288:15547–15555.
18. Shin YK, Martin B, Golden E, et al. Modulation of tastesensitivity by GLP-1 signaling. J Neurochem 2008;106:455–463.
19. Smith J, Su X, El-Maghrabi R, et al. Opposite regulationof CD36 ubiquitination by fatty acids and insulin: effectson fatty acid uptake. J Biol Chem 2008;283:13578–13585.
20. Pohl J, Ring A, Korkmaz U, et al. FAT/CD36-mediatedlong-chain fatty acid uptake in adipocytes requiresplasma membrane rafts. Mol Biol Cell 2005;16:24–31.
21. Tovey SC, Taylor CW. Cyclic AMP directs inositol (1,4,5)-trisphosphate evoked Ca2þ signalling to differentintracellular Ca2þ stores. J Cell Sci 2013;126:2305–2313.
22. Liao Y, Plummer NW, George MD, et al. A role for Orai inTRPC-mediated Ca2þ entry suggests that a TRPC: Oraicomplex may mediate store and receptor operatedCa2þ entry. Proc Natl Acad Sci U S A 2009;106:3202–3206.
23. Gaillard D, Laugerette F, Darcel N, et al. The gustatorypathway is involved in CD36-mediated orosensory per-ception of long-chain fatty acids in the mouse. FASEB J2008;22:1458–1468.
24. Park YM, Drazba JA, Vasanji A, et al. Oxidized LDL/CD36interaction induces loss of cell polarity and inhibitsmacrophage locomotion. Mol Biol Cell 2012;23:3057–3068.
25. Liao F, Shin HS, Rhee SG. In vitro tyrosine phosphory-lation of PLC-gamma 1 and PLC-gamma 2 by src-familyprotein tyrosine kinases. Biochem Biophys Res Commun1993;191:1028–1033.
26. Chevrot M, Bernard A, Ancel D, et al. Obesity alters thegustatory perception of lipids in the mouse: plausibleinvolvement of lingual CD36. J Lipid Res 2013;54:2485–2494.
27. Tran TT, Poirier H, Clément L, et al. Luminal lipidregulates CD36 levels and downstream signaling tostimulate chylomicron synthesis. J Biol Chem 2011;286:25201–25210.
28. Barnett-Norris J, Lynch D, Reggio PH. Lipids, lipid raftsand caveolae: their importance for GPCR signaling andtheir centrality to the endocannabinoid system. Life Sci2005;77:1625–1639.
29. Llegems E, Iwatsuki K, Kokrashvili Z, et al. REEP2 en-hances sweet receptor function by recruitment to lipidrafts. J Neurosci 2010;30:13774–13783.
30. Su X, Abumrad NA. Cellular fatty acid uptake: a pathwayunder construction. Trends Endocrinol Metab 2009;20:72–77.
31. Ehrlich D, Humpel C. Effects of ethanol on aggregation,serotonin release, and amyloid precursor protein pro-cessing in rat and human platelets. Platelets 2014;25:16–22.
32. Smith NJ. Low Affinity GPCRs for metabolic in-termediates: challenges for pharmacologists. FrontEndocrinol 2012;3:1.
33. Sundaresan S, Shahid R, Riehl TE, et al. CD36-dependent signaling mediates fatty acid-induced gutrelease of secretin and cholecystokinin. FASEB J 2013;27:1191–1202.
34. Martin C, Passilly-Degrace P, Chevrot M, et al. Lipid-mediated release of GLP-1 by mouse taste buds fromcircumvallate papillae: putative involvement of GPR120and impact on taste sensitivity. J Lipid Res 2012;53:2256–2265.
35. Damak S, Le-Coutre J, Bezencon C, et al. Fat taste re-ceptors and their methods of use. International applica-tion published under the patent cooperation treaty.2007;WO2007/014824 A1.

April 2014 Ca2+ Signaling in Human Taste Bud Cells 1005
36. Godinot N, Yasumatsu K, Barcos ME, et al. Activationof tongue-expressed GPR40 and GPR120 by non caloricagonists is not sufficient to drive preference in mice.Neuroscience 2013;250:20–30.
37. Heit B, Kim H, Cosío G, et al. Multimolecular signalingcomplexes enable Syk-mediated signaling of CD36internalization. Dev Cell 2013;24:372–383.
38. Pepino MY, Love-Gregory L, Klein S, et al. The fatty acidtranslocase gene CD36 and lingual lipase influence oralsensitivity to fat in obese subjects. J Lipid Res 2012;53:561–566.
39. Stewart JE, Seimon RV, Otto B, et al. Marked differencesin gustatory and gastrointestinal sensitivity to oleic acidbetween lean and obese men. Am J Clin Nutr 2011;93:703–711.
40. Ostrom RS, Insel PA. The evolving role of lipid rafts andcaveolae in G protein-coupled receptor signaling: impli-cations for molecular pharmacology. Br J Pharmacol2004;143:235–245.
41. Stewart JE, Feinle-Bisset C, Golding M, et al. Oralsensitivity to fatty acids, food consumption and BMI inhuman subjects. Br J Nutr 2010;104:145–152.
Author names in bold designate shared co-first authorship.
Received July 26, 2013. Accepted January 4, 2014.
Reprint requestsAddress requests for reprints to: Naim A. Khan, PhD, Physiologie de laNutrition and Toxicologie, UMR U866 INSERM/Université de Bourgogne/AgroSup, 6 Boulevard Gabriel, F-21000 Dijon, France. e-mail: [email protected]; fax: (33) 3 80 39 63 30.
Conflicts of interestThe authors disclose no conflicts.
FundingThis work was supported by grants from the FrenchMinistry of Higher Educationand Research, Région Bourgogne (Post-Doc to SSub) and ANR SensoFat-2(ANR-12-BSV1-0027-01), and by grants (DK033301 and DK060022) from theNational Institutes of Health (Abumrad).
BASICAN
DTR
ANSLAT
IONA
LAT

Supplementary Materialsand MethodsMaterials
All other chemicals including MCDB153 medium werefrom Sigma (St Louis, MO). GA was purified from Albatrellusdispansus.1 The antibodies for CD36 were from R&D (Abing-don, UK) (AF 2519) and Santa Cruz Biotechnology (Santa Cruz,CA) (sc-52645), those for GPR120 were from Santa CruzBiotechnology (sc-48203), LifeSpan Biosciences, Inc (Seattle,WA) (LS-A2003), and Abcam (Cambridge, MA; ab75313),those for a-gustducin (sc-395) and PLC-b2 (sc-206) werefrom Santa Cruz Biotechnology and those for caveolin 1 werefrom BD Biosciences (Le Pont de Claix, France).
Cultures and Immunocytochemistry ofHuman Fungiform TBC
The CD36-positive TBC, by using a positive selectiontechnique, were isolated as described for mice TBC.2 TheTBC were fixed in 4% paraformaldehyde in phosphate-buffered saline (pH 7.2) for 1 hour at room temperature.For immunocytochemistry, primary antibodies were used at1:500 (anti�a-gustducin, anti�PLC-b2), 1:250 (GPR120) or1:50 (CD36) and secondary antibodies were anti-rabbit IgGAlexa 635 or anti-mouse IgG Alexa 488 (Molecular Probes,Grand Island, NY). Images were acquired (Leica TCS SP2Spectral Confocal Microscope) at 405 nm for 40,6-diamidino-2-phenylindole, 488 nm for Alexa Fluor 488, and 633nm forAlexa Fluor 633.
Culture of Mouse Fungiform TBCThe CD36-positive TBC, by using a positive selection
technique, from mouse lingual fungiform papillae wereisolated as described elsewhere.2 The TBC were seeded ontoa Biocoat poly-d-lysine�coated dishes and cultured for 24hours before being used for experiments.
Culture of STC-1 CellsThe STC-1 cells were cultured in Dulbecco’s modified
Eagle medium with 20% fetal bovine serum. STC-1 cellsexpressing human CD36 were generated by electroporation(Nucleofector Kit V; Lonza, Cologne, Germany) as describedpreviously.3
Measurement of IP3 ProductionThe human fungiform TBC cells (1 � 106) were treated
with LA (20 mM) or GA (20 mM) for 20 minutes. Combinedtreatment was carried out by adding consecutively LA andGA (or vice versa) in 10-minute intervals. The controlsreceived 0.1 % ethanol (v/v) for 20 minutes. After stimu-lation, the cells were washed twice with ice-cold phosphate-buffered saline and lysed with 20% ice-cold perchloric acid(0.2:1; v/v). After centrifugation, the supernatant wasassayed for IP3 production using a kit (NEK064) from Per-kin Elmer (Courtaboeuf, France) according to manufac-turer’s instructions.
Messenger RNA Detection by Real-TimeQuantitative Polymerase Chain Reaction
Total RNA, extracted from TBC by Trizol (Invitrogen),treated with DNAse (RNAse-free; Qiagen, Valencia, CA) andreverse transcribed (1 mg) with Super Script II RNAseH-reverse transcriptase (Invitrogen). Real-time quantitativepolymerase chain reaction was performed with the iCycleriQ Real-Time Detection System (Bio-Rad, Hercules, CA)using SYBR Green I. Primers used (forward; reverse) were:b-actin: 50-AGAGGGAAATCGTGCGTGA-30; 50-CAATAGTGATGACCTGGCCGT-30, GPR120: 50-CTGGGGCTCATCTTTGTCGT-30; 50-ACGACGAGCACTAGAGGGAT-30, CD36: 50-GGCCAAGCTATTGCGACATG-30; 50-CCGAACACAGCGTAGATAGAC-3’,a-gustducin: 50-ACACATTGCAGTCCATCCTAGC-30; 50-ATCACCATCTTCTAGTGTATTTGCC-30.
siRNA Knock-Down of CD36 and GPR120Human fungiform TBC were transfected with siRNA ON-
Target plus Smart pool (25 nM) or nontargeting siRNA as acontrol (Dharmacon, Pittsburgh, PA) using the DharmaFECTprotocol (Dharmacon). Target sequences for CD36 siRNAand GPR120 siRNA were, respectively: 50-GGAAAGUCA-CUGCGACAUG-30 and 50-GGAAGAGGCUCACGGUAAG-30. Twenty-four hours before siRNAtreatment, the cells were placed in the culture mediumwithout antibiotics. After transfection (24 hours), the me-dium was replaced with fresh medium without siRNA for48 hours before performing experiments.
Lipid Raft IsolationLipid rafts were isolated by sucrose density gradient
centrifugation.4 Briefly, control and LA-treated (20 minutes)fungiform TBC (2.5 � 107) were washed with phosphatebuffered saline, lysed on ice in 2 mL 1% Triton X-100 in 25mM morpholineethanesulfonic acid, 150 mM NaCl (pH 6.5),buffer with a protease inhibitor cocktail, and homogenized(Dounce homogenizer). Homogenates equalized based onprotein concentrations, mixed with 2 mL 80% sucrose in1% Triton X-100, 25 mM morpholineethanesulfonic acid,150 mM NaCl (pH 6.5), buffer, were placed in a centrifugetube, overlaid with 4 mL 30% sucrose and 4 mL 5% sucrose,and centrifuged at 175,000g for 20 hours at 4�C (SW41rotor, Beckman Ultracentrifuge; Beckman Coulter, Brea, CA).Ten fractions (1.2 mL each) were collected from the top ofthe gradient and fractions 3�6 were precipitated with 2volumes of cold acetone for immunoblotting. Caveolin wasthe positive control for lipid raft detection and b-actin wasused for soluble fractions.
Western BlotsThe TBC were lysed in 50 mL buffer (20 mM HEPES [pH
7.3], 1 mM EDTA, 1 mM ethylene glycol-bis(b-aminoethylether)-N,N,N0,N0-tetraacetic acid, 0.15 mM NaCl, 1% TritonX-100; 10% glycerol; 1 mM phenylmethylsulfonyl fluoride;2 mM sodium orthovanadate, and 2 mL/mL anti-proteasecocktail) and centrifuged (13,000g � 10 minutes). Dena-tured proteins (30 mg) were separated by sodium dodecyl
1005.e1 Ozdener et al Gastroenterology Vol. 146, No. 4

sulfate polyacrylamide gel electrophoresis (10%), trans-ferred to polyvinylidine difluoride membranes, and probedfor CD36 (anti-CD36 antibody, 1:500 dilution) and GPR120(anti-GPR120, 1:500 dilution). Signal quantification wasdone by ChemiDoc XRS System (Bio-Rad).
HFDThe standard diet contained 4% fat5 and had the
following FA composition (percent of total FA): C14:0(0.73%), C16:0 (6.60%); 18:0 (2.36%); C18:1 (60.09%);C18:2 n-6 (19.41%), and C18:3 n-3 (8.88%). The HFD wasthe same as the standard diet except it contained 40% fat(palm oil) with the following FA composition (percent oftotal FA): C14:0 (1.25%), C16:0 (43.49%); 18:0 (4.46%);C18:1 (40.32%); C18:2 n-6 (9.93%), and C18:3 n-3 (0%).Diets were prepared every day and the mice consumedthem ad libitum.
Two-Bottle Preference Test of Lipid SolutionThe experiments on the spontaneous preference for
lipid-enriched solutions were performed by 2-bottle pref-erence test.6 The individually caged mice, fed either astandard diet or HFD, were allowed to choose between 0.1%canola oil emulsified in 0.3% xanthan gum (w/v) in water orwater with 0.3% xanthan gum (w/v) over a period of 12hours. The canola oil contained, as percent of fatty acids:61% oleic acid (18:1 n-9), 25% linoleic acid (18:2 n-6), and10% rumelenic acid (18:3 n-3).7 The intake was determinedby weighing the feeders.
Release of [3H]SerotoninThe release of [3H]serotonin was determined according
to our previously published procedure.6 Chlorgyline (100mL) and pargyline (100 mM) were used in order to preventserotonin metabolism. [3H] serotonin release was calculatedas percent of unstimulated cells.
Detection of GLP-1 ReleaseHuman fungiform TBC or freshly isolated TBC from
standard diet or HFD-fed mice were cultured in the
References1. Hara T, Hirasawa A, Sun Q, et al. Novel selective
ligands for free fatty acid receptors GPR120 andGPR40. Naunyn Schmiedebergs Arch Pharmacol 2009;380:247–255.
2. El-Yassimi A, Hichami A, Besnard P, et al. Linoleic acidinduces calcium signaling, SRC-kinase phosphorylationand neurotransmitters release in mouse CD36-positivegustatory cells. J Biol Chem 2008;283:12949–12959.
3. Sundaresan S, Shahid R, Riehl TE, et al. CD36-depen-dent signaling mediates fatty acid-induced gut releaseof secretin and cholecystokinin. FASEB J 2013;27:1191–1202.
4. Leon-Bollotte L, Subramaniam S, Cauvard O, et al.S-nitrosylation of the death receptor Fas promotes Fasligand–mediated apoptosis in cancer cells. Gastroenter-ology 2011;140:2009–2018.
5. Triboulot C, Hichami A, Denys A, et al. Dietary (n-3)polyunsaturated fatty acids exert antihypertensive effectsby modulating calcium signaling in T cells of rats. J Nutr2001;131:2364–2369.
6. Dramane G, Abdoul-Azize S, Hichami A, et al. STIM1regulates calcium signaling in taste bud cells andpreference for fat in mice. J Clin Invest 2012;122:2267–2282.
7. Ayorinde FO, Karen Garvin K, Saeed K. Determination ofthe fatty acid composition of saponified vegetable oilsusing matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun Mass Spec-trom 2000;14:608–615.
8. Martin C, Passilly-Degrace P, Chevrot M, et al. Lipid-mediated release of GLP-1 by mouse taste buds fromcircumvallate papillae: putative involvement of GPR120and impact on taste sensitivity. J Lipid Res 2012;53:2256–2265.
presence of LA (20 mM) or GA (20 mM) for 2 hours. TheGLP-1 concentrations in the supernatants were determinedby enzyme-linked immunosorbent assay (Millipore, Bill-erica, MA) according to the protocol furnished with the kitand mentioned elsewhere.8
April 2014 Ca2+ Signaling in Human Taste Bud Cells 1005.e2

Supplementary Figure 1. Effects of LA and GA on Ca2þ signaling in human fungiform TBC. The cultured TBC (2 � 105/assay)were loaded with Fura-2/AM and the changes in intracellular Ca2þ (F340/F380) monitored. (A, B) Increase in [Ca2+]i whenLA or GA were added at 20 mM individually or combined. (C) LA or GA added at 100 mM. Data in panels B and C are means �SEM (n ¼ 7).
Supplementary Figure 2. LA and GA induce IP3 productionin human fungiform TBC. The cultured TBC were exposed to20 mM LA or GA for 20 minutes and IP3 production wasdetermined. Data are means � SEM (n ¼ 5).
1005.e3 Ozdener et al Gastroenterology Vol. 146, No. 4

Supplementary Figure 3. CD36 and GPR120 expression incontrol STC-1 (Ctrl) and in STC-1 stably expressing humanCD36 (þCD36). (A) Western blots showing CD36 andGPR120 proteins with actin as the loading control. (B) Effectsof SSO on Ca2þ signaling in CD36 positive STC-1 cells. Cells(2 � 105/assay) were pretreated with SSO (20 mM, 15 mi-nutes) before addition of LA and [Ca2þ]i imaging. Data shownare representative of n ¼ 3 experiments.
April 2014 Ca2+ Signaling in Human Taste Bud Cells 1005.e4

Supplementary Figure 4. High-fat feeding reduces LA-induced serotonin release and enhances GA-inducedGLP-1 secretion by mice fungiform TBC. (A) Releaseof 3H-5-HT by TBC from mice fed a standard diet (Std)or a high-fat diet (HFD) for 2 months (n ¼ 7). Serotoninlevels were determined as described in the Supple-mentary Materials and Methods. (B) Effects of LA andGA on TBC release of GLP-1 (determined as describedin Supplementary Materials and Methods). Values aremeans � SEM (n ¼ 4).
1005.e5 Ozdener et al Gastroenterology Vol. 146, No. 4





























