Embed Size (px)
Citation preview

of July 10, 2018.This information is current as
SchistosomiasisTh17 Cell Responses in MurineCritical for the Development of Pathogenic CD209a Expression on Dendritic Cells Is
Nir Hacohen and Miguel J. StadeckerParnell, Tae J. Yun, Cheolho Cheong, Stephen C. Bunnell, Russo, D. Brenda Salantes, Chao-Qiang Lai, Laurence D.Raktima Raychowdhury, Bridget M. Larkin, Joanne M. Holly E. Ponichtera, Mara G. Shainheit, Beiyun C. Liu,
http://www.jimmunol.org/content/192/10/4655doi: 10.4049/jimmunol.1400121April 2014;
2014; 192:4655-4665; Prepublished online 11J Immunol
MaterialSupplementary
1.DCSupplementalhttp://www.jimmunol.org/content/suppl/2014/04/11/jimmunol.140012
Referenceshttp://www.jimmunol.org/content/192/10/4655.full#ref-list-1
, 20 of which you can access for free at: cites 68 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. All rights reserved.1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 10, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
CD209a Expression on Dendritic Cells Is Critical for theDevelopment of Pathogenic Th17 Cell Responses in MurineSchistosomiasis
Holly E. Ponichtera,*,† Mara G. Shainheit,‡ Beiyun C. Liu,*,‡ Raktima Raychowdhury,x
Bridget M. Larkin,*,† Joanne M. Russo,* D. Brenda Salantes,*,† Chao-Qiang Lai,{
Laurence D. Parnell,{ Tae J. Yun,‖ Cheolho Cheong,‖ Stephen C. Bunnell,*,†
Nir Hacohen,x and Miguel J. Stadecker*,†
In murine schistosomiasis, immunopathology and cytokine production in response to parasite eggs are uneven and strain dependent.
CBA/J (CBA) mice develop severe hepatic granulomatous inflammation associated with prominent Th17 cell responses driven by
dendritic cell (DC)-derived IL-1b and IL-23. Such Th17 cells fail to develop in low-pathology C57BL/6 (BL/6) mice, and the reasons
for these strain-specific differences in APC reactivity to eggs remain unclear. We show by gene profiling that CBA DCs display an 18-
fold higher expression of the C-type lectin receptor CD209a, a murine homolog of human DC-specific ICAM-3–grabbing nonintegrin,
compared with BL/6 DCs. Higher CD209a expression was observed in CBA splenic and granuloma APC subpopulations, but only
DCs induced Th17 cell differentiation in response to schistosome eggs. Gene silencing in CBA DCs and overexpression in BL/6 DCs
demonstrated that CD209a is essential for egg-elicited IL-1b and IL-23 production and subsequent Th17 cell development, which is
associated with SRC, RAF-1, and ERK1/2 activation. These findings reveal a novel mechanism controlling the development of Th17
cell–mediated severe immunopathology in helminthic disease. The Journal of Immunology, 2014, 192: 4655–4665.
Schistosoma mansoni is a trematode helminth that causesextensive disease in the developing world, accounting for.200 million infections and 200,000 deaths per year. The
principal cause of morbidity and mortality in S. mansoni infection isgranulomatous inflammation and subsequent fibrosis around para-site eggs deposited in the liver and intestines (1–5). Most infectedindividuals develop mild gastrointestinal disease, but 5–10% developlife-threatening hepatosplenic schistosomiasis, which is character-ized by severe liver fibrosis, splenomegaly, ascites, and portal hy-pertension (1–5).Similar to human disease, heterogeneity of disease severity also
is observed in an experimental murine model of schistosomiasis.
Infected CBA/J (CBA) mice develop severe hepatic pathology char-acterized by large, poorly circumscribed perioval granulomas (6–8).The severe pathology is mediated largely by T cell IL-17 productioninduced by egg Ag-stimulated dendritic cell (DC) secretion of IL-1band IL-23 (9–12). In contrast, infected C57BL/6 (BL/6) mice developmild pathology, with significantly smaller liver granulomas in a Th2-polarized environment (13). IL-17 is largely the product of Th17cells, a highly proinflammatory subset of CD4+ effector T cells thatalso produce IL-22, CSF, CXCL1, CXCL2, and TNF-a (14–17).Presently, the mechanisms underlying the variation in egg-inducedimmunopathology and selection of dominant CD4+ T cell phenotypeare incompletely understood; however, it is noteworthy that a recentstudy (18) of Schistosoma hematobium infection in humans similarlylinked the development of pathology to an increase in Th17 cells.We now demonstrate that genetic differences in pattern recog-
nition receptor (PRR) expression predispose CBA and BL/6 DCsto develop divergent cytokine responses following stimulation withlive schistosome eggs. PRRs are innate sensors used by APCs torecognize conserved pathogen-associated molecular patterns (19,20). C-type lectin receptors (CLRs) are a family of PRRs capableof binding carbohydrates (21, 22), such as the glycans Lewis X(LeX), GalNAcb1–4GlcNAc (LacdiNAc [LDN]), and fucosylatedLDN (LDN-F) typically expressed by schistosome eggs (23–26).We found overall CLR expression to be higher in CBA cells thanin BL/6 cells, and in CBA DCs, there was a striking overexpressionof the CLR CD209a, a murine homolog of human DC-specificICAM-3–grabbing nonintegrin (DC-SIGN; CD209). CD209a wasshown to facilitate the induction of egg-induced Th17 cells re-sponsible for causing severe immunopathology.
Materials and MethodsMice, parasites, and infection
Five- to six-week-old female CBA and BL/6 mice were obtained from TheJackson Laboratory. Swiss Webster mice were obtained from Charles River
*Department of Integrative Physiology and Pathobiology, Tufts University School ofMedicine, Boston, MA 02111; †Sackler School of Graduate Biomedical Sciences, TuftsUniversity, Boston, MA 02111; ‡Department of Molecular Biology and Microbiology,Tufts University School of Medicine, Boston, MA 02111; xBroad Institute of MIT andHarvard, Cambridge, MA 02142; {Nutritional Genomics Laboratory, Jean Mayer–U.S.Department of Agriculture Human Nutrition Research Center on Aging at Tufts Univer-sity, Boston, MA 02111; and ‖Institut de Recherches Cliniques de Montreal, Montreal,Quebec H2W 1R7, Canada
Received for publication January 15, 2014. Accepted for publication February 26,2014.
This work was supported by U.S. Public Health Service Grant R01 18919 (to M.J.S.),National Institutes of Health Grant R01 AI076575-01 (to S.C.B.), National Institutesof Health Director’s New Innovator award DP2OD002230 (to N.H.), and CanadianInstitutes of Health Research MOP 125933 and National Research Foundation ofKorea-Global Research Network 2013S1A2A2035348 (to C.C.).
Address correspondence and reprint requests to Dr. Miguel J. Stadecker, Tufts Uni-versity School of Medicine and Sackler School of Graduate Biomedical Sciences, 150Harrison Avenue, Boston, MA 02111. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: BL/6, C57BL/6; BMDC, bone marrow–deriveddendritic cell; CBA, CBA/J; CLR, C-type lectin receptor; DC, dendritic cell; DC-SIGN, DC-specific ICAM-3–grabbing nonintegrin; GC, granuloma cell; GO, geneontology; LDN, LacdiNAc (GalNAcb1–4GlcNAc); LDN-F, fucosylated LacdiNAc(GalNAcb1–4GlcNAc); LeX, Lewis X; PRR, pattern recognition receptor; qRT-PCR,quantitative RT-PCR; shRNA, short hairpin RNA; shCD209a, CD209a-targetedshRNA; shCTRL, GFP-targeted shRNA; Tg, transgenic.
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1400121
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Laboratories. A CBA mouse expressing a transgenic (Tg) TCR specific forthe Sm-p40 schistosome egg Ag was made in-house, as previously de-scribed (12). All mice were maintained at the Tufts University School ofMedicine Animal Facility in accordance with the Association for As-sessment and Accreditation of Laboratory Animal Care guidelines. Forsome experiments, CBA and BL/6 mice were infected with 85 S. mansonicercariae (Puerto Rico strain) by i.p. injection. Cercariae were shed frominfected Biomphalaria glabrata snails provided to us by BEI Resources(Manassas, VA). All Swiss Webster mice were infected in an identicalfashion for the purpose of isolating schistosome eggs. Eggs were isolatedfrom livers of 7–8-wk infected mice under sterile conditions by a series ofblending and straining techniques, as described previously (11).
Cells
Bone marrow–derived DCs. Bone marrow was flushed from femurs and tibiasof normal CBA and BL/6 mice. RBCs were lysed with Tris ammoniumchloride buffer, and cells were cultured in complete RPMI 1640 medium(Lonza) containing 10% FBS (Aleken Biologicals) and rGM-CSF at 15ng/ml (PeproTech; AF-315-03) or GM-CSF–containing supernatant fromthe J558L transfectant B cell hybridoma. The medium was changed ondays 3 and 5, and cells were harvested on day 7. CD11c+ DC purity was.85% by flow cytometric analysis.
CD4+ T cells. Single-cell suspensions were prepared from the spleens ofnormal CBA and BL/6mice, RBCs were lysed, and CD4+ T cells were purifiedby negative selection using a CD4+ T Cell Isolation Kit II for mouse (MiltenyiBiotec). CD4+ T cell purity was .95% by flow cytometric analysis.
Gene expression profiling
CBA and BL/6 bone marrow–derived DCs (BMDCs) prepared from in-dividual mice were plated in replicate at 1 3 106 cells/ml in 48-well tissueculture plates (BD Falcon). Replicates were pooled after 4 h, and totalRNAwas obtained by TRIzol reagent (Invitrogen) extraction, according tothe manufacturer’s instructions. Amplified and labeled cRNAwas assessedwith a Mouse Gene 1.0 ST array (Affymetrix, Santa Clara, CA), followingthe manufacturer’s instructions. Affymetrix Expression Console 1.1 soft-ware was used to generate annotated NetAffx CSV files for analysis.Microarray data were deposited in the National Center for BiotechnologyInformation Gene Expression Omnibus database under accession numberGSE55307 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE55307).Data from a single chip is representative of gene expression of unstimulatedDCs from an individual mouse, and two chips were run per mouse strain.
Gene ontology analysis
Genes with$ 2-fold difference in expression between CBA and BL/6 DCsand known biological function based on Ensembl release 54 gene archive(http://may2009.archive.ensembl.org/index.html) were defined as differ-entially expressed. Differentially expressed genes characterized to haveputative immunological functions were further selected for gene ontology(GO) and pathway analysis using the public Web server, g:Profiler withdefault settings (http://biit.cs.ut.ee/gprofiler/; Institute of Computer Sci-ence, University of Tartu, Tartu, Estonia) (27).
Cocultures
Purified naive CD4+ T cells from normal spleen and syngeneic BMDCswere cultured at an 8:1 ratio in complete RPMI 1640 medium togetherwith anti-CD3/CD28–coated beads (2 3 105 beads/ml, Dynabeads Invi-trogen) and freshly isolated live schistosome eggs for 96 h.
Cytokine analysis
ELISA. Supernatants from 96 h cocultures were assessed for IL-1b, IL-23,and IL-17A using ELISA kits from R&D Systems.
Quantitative RT-PCR. RNA from cocultures was obtained with TRIzolreagent or a TurboCapture 96 mRNA Kit (QIAGEN), and cDNA wassynthesized with a High Capacity Reverse Transcription Kit (Invitrogen) ora Sensiscript RT kit (QIAGEN). TaqMan probes for Il17a (Mm00439618)and Gapdh (4352339E) were used in combination with TaqMan GeneExpression Master Mix (all from Applied Biosystems).
CD209a analysis
Flow cytometry and cell sorting. Spleen cells from individual normal andinfected CBA and BL/6 spleens were isolated, and RBCs were lysed. Cellswere blocked with rat IgG and stained with fluorescently labeled Absspecific for CD11c (BD Pharmingen; 553802), CD19 (BD Pharmingen;553786), Gr-1 (BD Pharmingen; 553127), or F4/80 (AbD Serotec;MCA497APC) in combination with biotin-conjugated anti-CD209a (BD
Pharmingen; 558073). Subsequently, cells were stained with Alexa Fluor647–conjugated streptavidin (Invitrogen; S-21374). Data were acquiredwith a FACSCalibur flow cytometer and CellQuest software version 3.2.1(Becton Dickinson) and analyzed with Summit Software. Spleen andgranuloma cells (GCs) were gated for viability based on forward scatterand side scatter parameters, as well as propidium iodide exclusion. NormalCBA splenocytes were sorted at the Tufts Flow Cytometry Core Facilityusing the Abs listed above.
Quantitative RT-PCR. Normal and 7-wk infected CBA and BL/6 spleenor liver tissue was homogenized in TRIzol reagent, and cDNA was syn-thesized, as described above. A TaqMan probe for Cd209a (AppliedBiosystems; Mm00460067) was used in combination with TaqMan GeneExpression Master Mix.
Immunohistochemistry. Ten-micrometer OCT-embedded liver and spleencryostat sections were fixed in acetone. Liver sections were stained withanti-CD209a Ab (BMD10) for 18 h at 4˚C, followed by mouse anti-ratIgG2a-HRP for 30 min and Tyramide signal amplification (Invitrogen), orwith anti-CD209a plus anti-CD11c (N418), followed by streptavidin-HRPand Tyramide signal amplification. Spleen sections also were stained withanti-B220–Alexa Fluor 647. Fluorescent staining was observed using anLSM 710 confocal microscope (Zeiss).
RNA interference
Knockdown. CBA BMDCs derived with rGM-CSF were infected withlentivirus containing CD209a- or GFP-targeted short hairpin RNA (shRNA)(RNAi Platform of the Broad Institute of MITand Harvard). Puromycin wasused to select for infected cells, and BMDCs were harvested for cocultureexperiments on day 10.
Overexpression. The open reading frame sequence for CD209a (28, 29)was inserted into a lentiviral plasmid and packaged into viral particlesin HEK293T cells with X-tremeGENE 9 DNA Transfection Reagent(Roche). Concentrated virus was used to infect BMDCs, as describedabove.
Signaling protein analysis
Intracellular staining. BMDCs were washed, fixed with 3% paraformal-dehyde, blocked with rat IgG, and stained with a fluorescently labeled Abspecific for CD11c. Subsequently, cells were washed, permeabilized withmethanol, and stained with a primary Ab specific for phospho-ERK (CellSignaling; 4695), followed by a fluorescently labeled secondary anti-rabbitIgG Ab (Invitrogen; A11034). Data were acquired with the FACSCaliburflow cytometer and CellQuest software version 3.2.1 (Becton Dickinson)and analyzed with Summit Software.
Western blot. BMDCs were washed, lysed, and prepared with Laemmli(SDS-Sample) Buffer (Boston BioProducts). Samples were run on an SDS-PAGE gel and transferred to an Immobilon-P polyvinylidene difluoride(Millipore) membrane, which was blocked in 5% BSA. The activation ofMAPKs was detected with Abs specific for phospho-Src Tyr416 (2101),phospho–RAF-1 Ser338 (9427), phospho-ERK1/2 Thr202/Tyr204 (4695P),phospho-JNK Thr183/Tyr185 (9251S), and phospho-p38 Thr180/Tyr182(9215S; all from Cell Signaling). Total kinase expression was detectedwith Abs specific for Src (32G6), RAF-1 (9422P), ERK1/2 (4695), JNK(9252), p38 (9212), and GAPDH (2118S; all from Cell Signaling).
Statistical analysis
ANOVA and the Student t test were used to statistically analyze differencesbetween groups. The p values , 0.05 were considered significant.
ResultsGene profiling reveals elevated CLR expression by CBA versusBL/6 DCs
CBA and BL/6 DCs were previously shown to induce divergentschistosome egg-specific cytokine profiles (9–12). To identifygenetic differences intrinsic to CBA and BL/6 DCs that may in-fluence the cytokine response to schistosome Ags, we usedAffymetrix microarray technology to assess baseline expression of.22,000 genes in CBA and BL/6 BMDCs (Fig. 1A). Genes ofknown biological function with a$2-fold difference in expressionbetween CBA and BL/6 BMDCs were functionally categorized byGO analysis using the public Web server, g:Profiler (27) (Fig. 1B,Supplemental Table I). Of 180 biologically characterized genes thatwere elevated $2-fold in CBA versus BL/6 BMDCs, 35 had known
4656 DC CD209a INDUCES Th17 RESPONSE IN SCHISTOSOMIASIS
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

immunological functions (Supplemental Table II). GO analysisrevealed elevated expression of genes with predicted roles in cellmembrane–intrinsic immune defense response to external stimuli,carbohydrate binding, and molecular-transducer activity (Fig. 1B,Supplemental Tables I, II). Of 157 genes that were elevated $2-fold in BL/6 BMDCs versus CBA BMDCs, 17 had known im-munological function (Supplemental Table III). GO analysis re-vealed that BL/6 BMDCs exhibited a markedly different expressionprofile, characterized by genes involved in the regulation of im-mune effector processes and various metabolic processes (Fig. 1B,Supplemental Table I).Among all of the genes assessed by microarray technology,
the most apparent difference between CBA and BL/6 BMDCswas in PRR expression. In CBA DCs, the majority of overex-pressed PRR genes belonged to the CLR family, among whichthere was a striking 18-fold increase in expression of the CLRCd209a (Fig. 1C). CD209a, also known as mouse DC-SIGN andSIGNR5, is one of eight murine homologs of human DC-SIGN,a CLR known to bind to schistosome egg glycans (25, 28, 30).Elevated expression of CD209a by CBA DCs was confirmed inBMDCs from individual CBA and BL/6 mice by quantitative RT-PCR (qRT-PCR) (Fig. 1D). These results denote a significant dif-ference in baseline CLR gene expression between CBA and BL/6BMDCs.
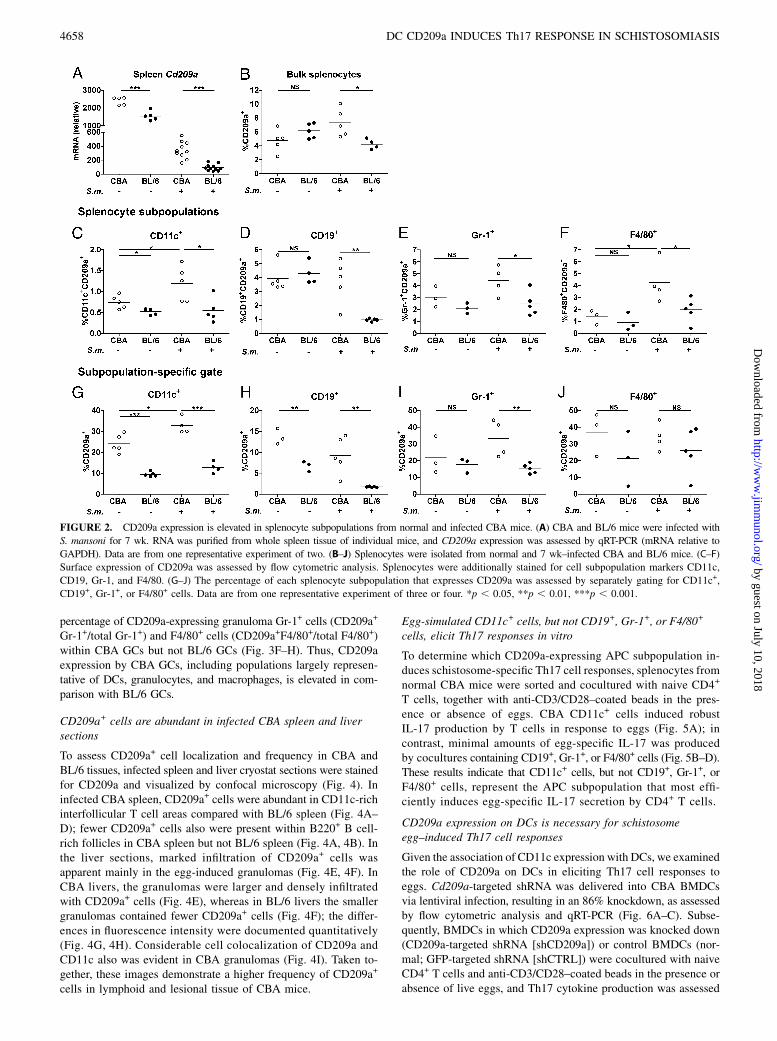
CD209a expression is elevated in splenocyte subpopulationsfrom normal and schistosome-infected CBA mice
Cd209a expression was significantly higher in the spleens of bothnormal and infected CBA mice versus BL/6 mice (Fig. 2A). Theoverall relative expression of Cd209a decreased in infected spleenbecause of the development of pronounced splenomegaly after7 wk of infection, primarily caused by clonal T and B cell ex-pansion; however, elevation of Cd209a in CBA spleen comparedwith BL/6 spleen remained intact (Fig. 2A, 2B).Several studies (22, 31–33) demonstrated that human DC-SIGN
is primarily expressed on DCs, but little is known about CD209aexpression by APC subpopulations in mice. CD11c+, CD19+,Gr-1+, and F4/80+ splenocytes from normal and infected CBA and
BL/6 mice, largely representative of DCs, B cells, granulocytes,and macrophages, respectively, were assessed for CD209a ex-pression by flow cytometric analysis. The percentage of CD209a-expressing CD11c+ cells within normal and infected splenocytes(CD209a+CD11c+/bulk spleen and subpopulation-specific–gatedCD209a+CD11c+/total CD11c+) was higher in CBA mice. More-over, CD209a-expressing CD11c+ cells increased after 7 wk ofinfection in CBA spleen but remained unchanged in BL/6 spleen(Fig. 2C, 2G). Similarly elevated CD209a expression was seen onCD19+, Gr-1+, and F4/80+ cells in infected CBA spleen versusBL/6 spleen (Fig. 2D–F). Subpopulation-specific gating revealedthat a higher percentage of CD19+ cells (CD209a+CD19+/totalCD19+) and Gr-1+ cells (CD209a+Gr-1+/total Gr-1+) expressedCD209a in normal and infected CBA spleen versus BL/6 spleen(Fig. 2H, 2I). There was no difference in the percentage ofCD209a-expressing F4/80+ cells by subpopulation-specific gating(Fig. 2J). These data indicate that CD209a is expressed by variousAPC subpopulations, and elevated CD209a expression is apparentin normal and schistosome-infected CBA APCs versus BL/6APCs.
CD209a expression is higher in GCs from infected CBA miceversus BL/6 mice
As the liver is the principal site of egg-induced granulomatouspathology in S. mansoni infection, we investigated Cd209a ex-pression in livers from normal and infected CBA and BL/6 mice.Although there was no difference in CD209a expression in normallivers from these strains when assessed by qRT-PCR, after 7 wk ofinfection we detected a significant increase in CD209a expressionin CBA liver (Fig. 3A). Additionally, elevated CD209a expressionwas confirmed in bulk CBA liver GCs by flow cytometric analysis(Fig. 3B).Within GCs, the percentage of CD209a-expressing CD11c+
cells (CD209a+CD11c+/bulk GCs and subpopulation-specific–gatedCD209a+CD11c+/total CD11c+) was elevated in infected CBAversus BL/6 mice (Fig. 3C, 3F). Similarly, CD209a expressionalso was higher in Gr-1+ and F4/80+ bulk CBA GCs (Fig. 3D,3E). Subpopulation-specific gating revealed an increase in the
FIGURE 1. CLR expression is elevated in CBA
DCs versus BL/6 DCs. (A) BL/6 and CBA BMDCs
were differentiated in medium containing GM-CSF
over 7 d. RNA was purified, and gene expression
was assessed by Affymetrix Mouse 1.0 ST Gene
Array technology. Scatter plot shows log2-trans-
formed gene expression. Arrow points to CD209a.
(B) Genes with $2-fold difference in expression
between mouse strains with characterized biologi-
cal function were selected for GO analysis. A
functional profile for differentially expressed genes
was obtained using the Web server, g:Profiler. (C)
PRRs with $2-fold difference in expression by
CBA DCs versus BL/6 DCs. Bars represent mean6SD of two independent microarray chips for each
strain. (D) CD209a expression by BMDCs from
four individual CBA and BL/6 mice was deter-
mined by qRT-PCR (mRNA relative to GAPDH).
The Journal of Immunology 4657
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
percentage of CD209a-expressing granuloma Gr-1+ cells (CD209a+
Gr-1+/total Gr-1+) and F4/80+ cells (CD209a+F4/80+/total F4/80+)within CBA GCs but not BL/6 GCs (Fig. 3F–H). Thus, CD209aexpression by CBA GCs, including populations largely represen-tative of DCs, granulocytes, and macrophages, is elevated in com-parison with BL/6 GCs.
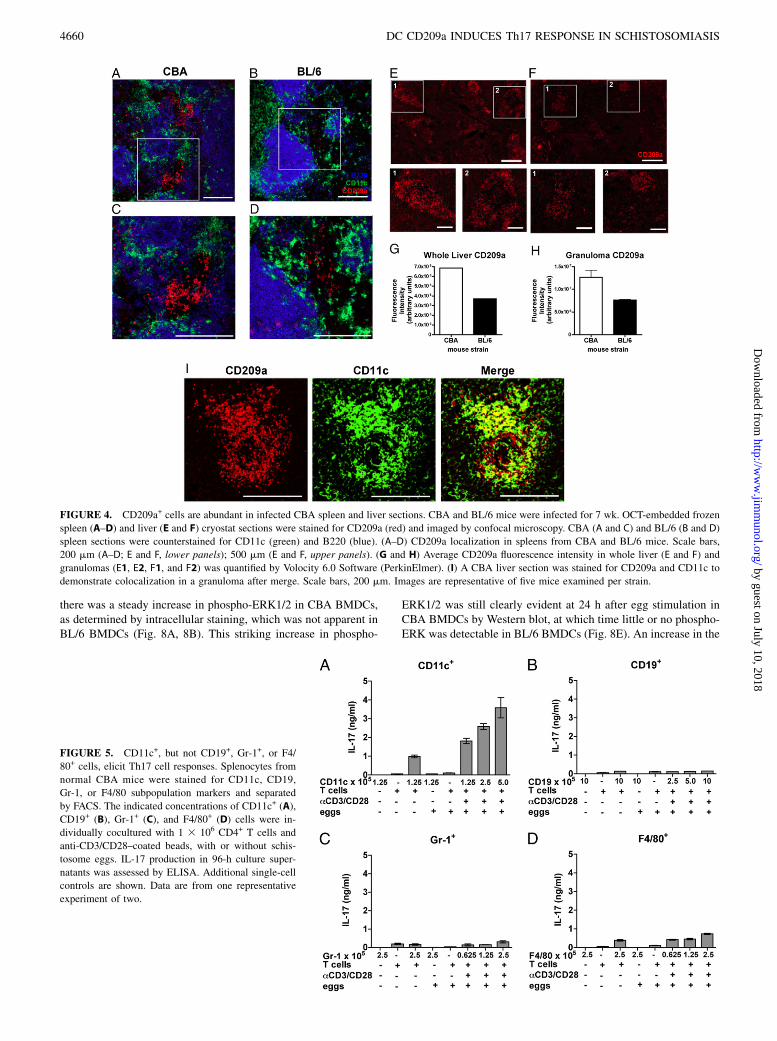
CD209a+ cells are abundant in infected CBA spleen and liversections
To assess CD209a+ cell localization and frequency in CBA andBL/6 tissues, infected spleen and liver cryostat sections were stainedfor CD209a and visualized by confocal microscopy (Fig. 4). Ininfected CBA spleen, CD209a+ cells were abundant in CD11c-richinterfollicular T cell areas compared with BL/6 spleen (Fig. 4A–D); fewer CD209a+ cells also were present within B220+ B cell-rich follicles in CBA spleen but not BL/6 spleen (Fig. 4A, 4B). Inthe liver sections, marked infiltration of CD209a+ cells wasapparent mainly in the egg-induced granulomas (Fig. 4E, 4F). InCBA livers, the granulomas were larger and densely infiltratedwith CD209a+ cells (Fig. 4E), whereas in BL/6 livers the smallergranulomas contained fewer CD209a+ cells (Fig. 4F); the differ-ences in fluorescence intensity were documented quantitatively(Fig. 4G, 4H). Considerable cell colocalization of CD209a andCD11c also was evident in CBA granulomas (Fig. 4I). Taken to-gether, these images demonstrate a higher frequency of CD209a+
cells in lymphoid and lesional tissue of CBA mice.
Egg-simulated CD11c+ cells, but not CD19+, Gr-1+, or F4/80+
cells, elicit Th17 responses in vitro
To determine which CD209a-expressing APC subpopulation in-duces schistosome-specific Th17 cell responses, splenocytes fromnormal CBA mice were sorted and cocultured with naive CD4+
T cells, together with anti-CD3/CD28–coated beads in the pres-ence or absence of eggs. CBA CD11c+ cells induced robustIL-17 production by T cells in response to eggs (Fig. 5A); incontrast, minimal amounts of egg-specific IL-17 was producedby cocultures containing CD19+, Gr-1+, or F4/80+ cells (Fig. 5B–D).These results indicate that CD11c+ cells, but not CD19+, Gr-1+, orF4/80+ cells, represent the APC subpopulation that most effi-ciently induces egg-specific IL-17 secretion by CD4+ T cells.
CD209a expression on DCs is necessary for schistosomeegg–induced Th17 cell responses
Given the association of CD11c expression with DCs, we examinedthe role of CD209a on DCs in eliciting Th17 cell responses toeggs. Cd209a-targeted shRNA was delivered into CBA BMDCsvia lentiviral infection, resulting in an 86% knockdown, as assessedby flow cytometric analysis and qRT-PCR (Fig. 6A–C). Subse-quently, BMDCs in which CD209a expression was knocked down(CD209a-targeted shRNA [shCD209a]) or control BMDCs (nor-mal; GFP-targeted shRNA [shCTRL]) were cocultured with naiveCD4+ T cells and anti-CD3/CD28–coated beads in the presence orabsence of live eggs, and Th17 cytokine production was assessed
FIGURE 2. CD209a expression is elevated in splenocyte subpopulations from normal and infected CBA mice. (A) CBA and BL/6 mice were infected with
S. mansoni for 7 wk. RNA was purified from whole spleen tissue of individual mice, and CD209a expression was assessed by qRT-PCR (mRNA relative to
GAPDH). Data are from one representative experiment of two. (B–J) Splenocytes were isolated from normal and 7 wk–infected CBA and BL/6 mice. (C–F)
Surface expression of CD209a was assessed by flow cytometric analysis. Splenocytes were additionally stained for cell subpopulation markers CD11c,
CD19, Gr-1, and F4/80. (G–J) The percentage of each splenocyte subpopulation that expresses CD209a was assessed by separately gating for CD11c+,
CD19+, Gr-1+, or F4/80+ cells. Data are from one representative experiment of three or four. *p , 0.05, **p , 0.01, ***p , 0.001.
4658 DC CD209a INDUCES Th17 RESPONSE IN SCHISTOSOMIASIS
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

by ELISA and qRT-PCR. In response to eggs, cocultures con-taining shCD209a DCs produced significantly less IL-1b andIL-23 compared with normal or shCTRL DCs (Fig. 6D, 6E). Ac-cordingly, there was a substantial decrease in egg-stimulated T cellIL-17 secretion and mRNA expression in cocultures containingshCD209a DCs compared with cocultures containing control DCs(Fig. 6F, 6G). Significantly, shCD209a DCs also elicited lowerIL-17 production in egg-stimulated TCR-Tg CD4+ T cells specificfor the immunodominant peptide of the Sm-p40 major schistosomeegg Ag (Fig. 6H) (12).Further assessment of transcription factors required for line-
age commitment of CD4+ T cells revealed that egg-stimulatedcocultures containing T cells and shCD209a DCs exhibited sig-nificantly lower expression of the Th17-associated transcriptionfactor Rorc. In contrast, the Th1- and Th2-associated transcriptionfactors, Tbx21 and Gata3, were unchanged, regardless of the DCpopulation present in the cultures (Fig. 6I–K). Collectively, thesedata indicate that CD209a expression on CBA DCs is necessaryfor egg-stimulated IL-23 and IL-1b production, which inducesTh17 cell differentiation.
CD209a overexpression confers on BL/6 DCs the capacity toinduce Th17 cell responses
CD209a expression is significantly lower in BL/6 DCs than in CBADCs (Figs. 1–3), and BL/6 DCs fail to induce Th17 cell responses toeggs in vitro (9–12). Thus, we investigated whether CD209a might
confer on BL/6 DCs the capacity to induce egg-specific Th17 cellresponses. A lentiviral vector was used to overexpress CD209a inBL/6 BMDCs, which was accomplished successfully, as determinedby flow cytometric analysis and qRT-PCR (Fig. 7A–C).BL/6 CD209a-expressing BMDCs were cocultured with naive
BL/6 CD4+ T cells and anti-CD3/CD28–coated beads in thepresence or absence of live eggs, and Th17-associated cytokineproduction was assessed by ELISA and qRT-PCR. Egg-stimulatedCD209a-expressing BMDCs produced higher levels of IL-1b andIL-23 in comparison with control BMDCs (Fig. 7D, 7E); IL-17secretion and mRNA expression also were enhanced in CD209aBMDC cocultures compared with cocultures containing controlBMDCs (Fig. 7F, 7G). Additionally, Rorc expression was elevatedin CD209a BMDC cocultures compared with control BMDCs(Fig. 7H), but there was no significant change in Tbx21 or Gata3expression (Fig. 7I, 7J). Overall, these results confirm that CD209aexpression enables egg-stimulated DCs to induce IL-1b and IL-23secretion, leading to Th17 cell development.
CD209a expression is associated with schistosomeegg–stimulated MAPK activation in DCs
Although signaling pathways associated with CD209a are un-known, recent studies demonstrated that the MAPKs RAF-1 andERK1/2 are activated in DCs following ligation of human DC-SIGN (34–37). Therefore, we investigated MAPK activation inCBA and BL/6 BMDCs. Following stimulation with live eggs,
FIGURE 3. CD209a expression is higher in GCs from infected CBA mice versus BL/6 mice. (A) CBA and BL/6 mice were infected with S. mansoni for
7 wk. RNAwas purified from whole liver tissue of individual mice, and CD209a expression was assessed by qRT-PCR (mRNA relative to GAPDH). Data
are from one representative experiment of two. (B–H) Liver GCs were isolated from livers by collagenase digestion, followed by Lympholyte cell sep-
aration. Surface expression of CD209a was assessed by flow cytometric analysis. (C–E) GCs also were stained for cell subpopulation markers CD11c, Gr-1,
and F4/80. (F–H) The percentage of each granuloma subpopulation that expresses CD209a was assessed by separately gating for CD11c+, Gr-1+, or F4/80+
cells. Data are from one representative experiment of three. *p , 0.05, **p , 0.01, ***p , 0.001.
The Journal of Immunology 4659
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
there was a steady increase in phospho-ERK1/2 in CBA BMDCs,as determined by intracellular staining, which was not apparent inBL/6 BMDCs (Fig. 8A, 8B). This striking increase in phospho-
ERK1/2 was still clearly evident at 24 h after egg stimulation inCBA BMDCs by Western blot, at which time little or no phospho-ERK was detectable in BL/6 BMDCs (Fig. 8E). An increase in the
FIGURE 4. CD209a+ cells are abundant in infected CBA spleen and liver sections. CBA and BL/6 mice were infected for 7 wk. OCT-embedded frozen
spleen (A–D) and liver (E and F) cryostat sections were stained for CD209a (red) and imaged by confocal microscopy. CBA (A and C) and BL/6 (B and D)
spleen sections were counterstained for CD11c (green) and B220 (blue). (A–D) CD209a localization in spleens from CBA and BL/6 mice. Scale bars,
200 mm (A–D; E and F, lower panels); 500 mm (E and F, upper panels). (G and H) Average CD209a fluorescence intensity in whole liver (E and F) and
granulomas (E1, E2, F1, and F2) was quantified by Volocity 6.0 Software (PerkinElmer). (I) A CBA liver section was stained for CD209a and CD11c to
demonstrate colocalization in a granuloma after merge. Scale bars, 200 mm. Images are representative of five mice examined per strain.
FIGURE 5. CD11c+, but not CD19+, Gr-1+, or F4/
80+ cells, elicit Th17 cell responses. Splenocytes from
normal CBA mice were stained for CD11c, CD19,
Gr-1, or F4/80 subpopulation markers and separated
by FACS. The indicated concentrations of CD11c+ (A),
CD19+ (B), Gr-1+ (C), and F4/80+ (D) cells were in-
dividually cocultured with 1 3 106 CD4+ T cells and
anti-CD3/CD28–coated beads, with or without schis-
tosome eggs. IL-17 production in 96-h culture super-
natants was assessed by ELISA. Additional single-cell
controls are shown. Data are from one representative
experiment of two.
4660 DC CD209a INDUCES Th17 RESPONSE IN SCHISTOSOMIASIS
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

phosphorylation of the upstream MAPK RAF-1, as well as SRCkinase, also was detected in CBA BMDCs compared with BL/6BMDCs (Fig. 8C, 8D). In contrast, there was no significant acti-vation of the MAPKs p38 or JNK (Fig. 8F, 8G). These results areconsistent with the notion that egg-stimulated CD209a expressionon CBA DCs is associated with SRC, RAF-1, and ERK1/2 MAPKactivation.
DiscussionDCs are potent APCs that sense foreign Ag through PRRs andinduce the differentiation of naive CD4+ T cells into various ef-fector T cell populations. Diverse cellular, environmental, andgenetic factors may influence DCs to bias Th cell differentiationtoward Th1, Th2, Th17, or regulatory T cell lineages (4, 20, 22,31, 38). In murine S. mansoni infection, the development of aTh17 cell response is detrimental to the host, rather than protec-tive, and leads to the development of severe pathology (5, 9–12).In the current study, comprehensive gene profiling revealed
major differences in the expression of PRRs between CBA andBL/6 mice. We found a strikingly higher expression of the CLRCD209a on CBA DCs compared with BL/6 DCs. CD209a ex-pression on DCs, but not on B cells, granulocytes, or macrophages,was required for the induction of pathogenic CD4+ Th17 cellresponses to live eggs. The absence of CD209a expression, eithernaturally on normal BL/6 DCs or silenced in CBA DCs, resultedin a marked reduction in IL-1b and IL-23 following egg stimulationand consequent Th17 cell development. Conversely, overexpression
of CD209a in BL/6 DCs enabled egg-stimulated IL-1b and IL-23production and subsequent Th17 cell development. Finally, CD209aexpression on egg-stimulated CBA DCs was associated with MAPKactivation.Proinflammatory cytokine responses to various bacterial, viral,
fungal, and parasitic Ag have been characterized downstream ofPRR–Ag recognition by APCs (19–22, 39, 40). Of the variousfamilies of PRRs, CLRs are the best characterized in the contextof schistosome Ag recognition (25, 26, 41–44). CLRs are a largefamily of calcium-dependent receptors that bind glycans on bothpathogen and host cell surfaces, facilitating recognition of a widerange of glycosylation patterns (21, 22). Schistosome-specific gly-cans, both O- and N-linked, are rich in fucose and include LeX,polyLeX, pseudo–Lewis Y, LDN, LDN-F, CAA, F-LDN, F-LDN-F,and HexNAc-DF (23, 24, 26). Such glycans are heavily expressedon the surface of S. mansoni eggs and are actively secreted in theform of soluble glycoproteins (45). Human CLRs, such as DC-SIGN, DC-SIGNR, macrophage galactose-type lectin, and themannose receptor, bind to schistosome egg glycans, such asLeX, LDN, and LDN-F (25, 41–44, 46, 47). Additionally, Ritteret al. (48) demonstrated that components of schistosome solu-ble egg Ag stimulate murine Dectin-2, leading to inflammasomeactivation and IL-1b production. Although two murine homo-logs of DC-SIGN, CD209b and CD209d, were shown to bindthe schistosome-expressed glycan LeX (29, 49), no in vivo functionhas been attributed to these CLRs, and their possible role in pa-thology has not been described in murine schistosomiasis. Our
FIGURE 6. CD209a expression on CBA DCs is necessary for the induction of Th17 cell responses. (A and B) CBA BMDCs were differentiated in rGM-
CSF–containing medium over 10 d. shRNA delivered by a lentiviral vector was used to knock down CD209a expression (shCD209a). Infected DCs were
enriched by puromycin selection, and CD209a knockdown efficiency was assessed by flow cytometric analysis relative to CBA BMDCs receiving a GFP-
targeted control shRNA (shCTRL) or normal BMDCs. (C) CD209a expression knockdown also was assessed by qRT-PCR (mRNA relative to GAPDH).
(D–F) Normal CBA, shCTRL, or shCD209a DCs were cocultured with naive CBA CD4+ T cells and anti-CD3/CD28–coated beads, with or without
schistosome eggs, for 96 h. IL-1b, IL-23, and IL-17 in supernatants were assessed by ELISA. (G) Il17a was assessed by qRT-PCR. (H) Normal CBA,
shCTRL, or shCD209a DCs were cocultured with S. mansoni major egg Ag Sm-p40–specific Tg T cells, with or without schistosome eggs. IL-17 was
assessed by ELISA. (I–K) Transcription factors Rorc, Tbx21, and Gata3 were assessed by qRT-PCR in egg-stimulated cocultures. Bars represent the
mean 6 SD of three biological replicates of one representative experiment of five. *p , 0.05, **p , 0.01, ***p , 0.001.
The Journal of Immunology 4661
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

novel findings suggest that CD209a plays a critical role in pre-cipitating Th17 cell–mediated inflammation following stimulationwith egg glycans.CD209a is one of eight murine homologs of human DC-SIGN,
a CLR capable of binding various mannose- and fucose-containingbacterial and viral glycans, as well as facilitating endocytosis andcytokine production (22, 28, 29, 50). The specific glycan ligand forCD209a has not been identified (29), but some studies (29, 44)suggested that it may play a role in Ag internalization and pre-sentation. Peptide sequence analysis showed that CD209a retainsmany structural properties of human DC-SIGN, including onecarbohydrate- and calcium-binding domain in the extracellularpeptide sequence, one conserved 23-aa sequence in the neck re-gion, and triacidic cluster and dileucine motifs in the cytoplasmictail (28, 29, 51). Such cytoplasmic tail motifs promote receptorinternalization and may function in receptor–Ag uptake (51). Assuch, DC-SIGN expressed on human DCs was shown to inter-nalize and target soluble egg Ag to lysosomal compartmentscontaining MHC class II molecules (44). Along these same lines,CD209a+ monocyte-derived DCs were shown to possess powerfulAg-presenting capability (29). A more recent study (52) on bac-terial sepsis was the first to reveal that leukocyte cell–derivedchemotaxin-2 interacts with CD209a on murine macrophages topromote endocytosis, bacterial killing, and cytokine production.
Signaling events proximal to ligand binding by CD209a havenot been characterized; however, various human studies (34–37)documented MAPK activation downstream of DC-SIGN. MAPKsare evolutionarily conserved signaling molecules essential formediating rapid communication of extracellular signals to thenucleus during diverse cellular processes (53, 54). Gringhuis et al.(35, 36) showed that, in the presence of TLR4 stimulation, DC-SIGN ligation by ManLam stimulates the recruitment of a sig-nalosome of scaffolding and signaling proteins that leads toMAPK activation, modification of NF-kB, and IL-12p40, IL-10,IL-12p35, and IL-6 production. Caparros et al. (34) revealed thatERK1/2 and PI3K, but not p38, are activated following engage-ment of DC-SIGN, which also coprecipitates with tyrosine kinasesLyn and Syk in human DCs. A similar study (37) documentinginteractions between DC-SIGN and syncytial virus glycoprotein Galso demonstrated ERK/1/2 stimulation post–DC-SIGN ligation.Although MAPK activation was reported following cross-linkingof CD209b (SIGNR1) (55), as well as CD209d (SIGNR3) (56),the signaling mechanisms critical for Th17 cell–associated cyto-kine production downstream of murine DC-SIGN homologs, par-ticularly in response to schistosome products, are largely unknown.Our findings demonstrate that CD209a expression on DCs is
essential for the induction of egg-stimulated Th17 responses as-sociated with DC ERK1/2 activation. Although little is known
FIGURE 7. CD209a overexpression confers on BL/6 DCs the capacity to induce Th17 cell responses. (A and B) BL/6 BMDCs were differentiated in
rGM-CSF–containing medium over 10 d. CD209a overexpression was achieved using a lentiviral vector and confirmed by flow cytometric analysis relative
to BL/6 control BMDCs that overexpressed RFP (CTRL) or normal BMDCs. (C) CD209a overexpression also was assessed by qRT-PCR (mRNA relative to
GAPDH). (D–F) Normal BL/6, CTRL, or CD209a DCs were cocultured with naive BL/6 CD4+ T cells and anti-CD3/CD28–coated beads, with or without
schistosome eggs, for 96 h. IL-1b, IL-23, and IL-17 in supernatants were assessed by ELISA. (G) Il17a was assessed by qRT-PCR. (H–J) Transcription
factors Rorc, Tbx21, and Gata3 were assessed by qRT-PCR in egg-stimulated cocultures. Bars represent the mean6 SD of three biological replicates of one
experiment representative of three. *p , 0.05, ***p , 0.001.
4662 DC CD209a INDUCES Th17 RESPONSE IN SCHISTOSOMIASIS
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

of the relationship between ERK1/2 activation and Th17 cellresponses, signaling through ERK1/2 was shown to promote IL-23p19 and IL-1b production, leading to Th17 cell differentiationin a study of primary human fibroblasts (57). More relevant toimmunity in schistosome infection, ERK1/2 activation, in theabsence of significant JNK or p38 activation, also was reported inresponse to lacto-N-fucopentaose III and ES62 from Acan-thocheilonema viteae; however, these studies (58–62) focusedon the anti-inflammatory properties of helminth-related mole-cules and the induction of Th2 cell responses. Indeed, numer-ous studies (61–66) established the immunomodulatory Th2-promoting properties of helminth products that may signalthrough receptors, such as human DC-SIGN and the mannosereceptor. Prior to this study, the only CLRs known to facilitateTh17 cell responses were Dectin-1 and Dectin-2, which signalvia Syk kinase and CARD9 in response to fungal Ags, resultingin IL-23 production (21, 67, 68). Our finding that a novel CLR,CD209a, mediates proinflammatory Th17 cell responses indi-cates that the eight murine homologs of DC-SIGN, each ofwhich retains slightly different structural properties of humanDC-SIGN, may have evolved to mediate divergent functions.As reflected in the nomenclature of DC-SIGN, its natural ligand
is ICAM-3, one of several transmembrane glycoproteins andICAMs that facilitate DC–T cell interactions (32). However, it isunknown whether murine DC-SIGN homologs also bind adhesionmolecules to promote cell–cell interaction. Immune synapse for-mation between DCs and T cells is required for efficient Agpresentation and DC–T cell intercellular feedback that leads tocytokine production. Future assessment of the ICAM-binding
capacity of CD209a will clarify whether murine DC-SIGN homo-logs also have the ability to promote cell adhesion and interaction.In sum, severe hepatic granulomatous pathology in murine
schistosomiasis, as seen in infected CBA mice, is dependent onCD4+ Th17 cell responses. In this article, we show that pathogenicTh17 cell cytokine responses to live S. mansoni eggs are largelydependent on the expression of CD209a by DCs. Future workinvestigating the outcome of pathology in S. mansoni–infectedCD209a-deficient CBA mice will clarify the function of CD209ain murine schistosomiasis in vivo.
AcknowledgmentsWe thank Allen Parmelee and Stephen Kwok at the Tufts Flow Cytometry
Core Facility. The U.S. Department of Agriculture is an equal opportunity
provider and employer.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bica, I., D. H. Hamer, and M. J. Stadecker. 2000. Hepatic schistosomiasis. Infect.
Dis. Clin. North Am. 14: 583–604, viii.2. Pearce, E. J., and A. S. MacDonald. 2002. The immunobiology of schistoso-
miasis. Nat. Rev. Immunol. 2: 499–511.3. Fallon, P. G. 2000. Immunopathology of schistosomiasis: a cautionary tale of
mice and men. Immunol. Today 21: 29–35.4. Wilson, M. S., M. M. Mentink-Kane, J. T. Pesce, T. R. Ramalingam,
R. Thompson, and T. A. Wynn. 2007. Immunopathology of schistosomiasis.Immunol. Cell Biol. 85: 148–154.
5. Larkin, B. M., P. M. Smith, H. E. Ponichtera, M. G. Shainheit, L. I. Rutitzky, andM. J. Stadecker. 2012. Induction and regulation of pathogenic Th17 cellresponses in schistosomiasis. Semin. Immunopathol. 34: 873–888.
FIGURE 8. Egg stimulation of CBA DCs is associated with greater MAPK activation. (A and B) CBA and BL/6 BMDCs were differentiated in GM-
CSF–containing medium over 7 d and cultured with schistosome eggs for the indicated time periods. Cells were fixed, permeabilized, and analyzed by
intracellular staining for phospho-ERK1/2. Flow cytometry plots were gated for viability and CD11c+ cells, which were .80% for both strains. Data are
from one representative experiment of three. (C–G) CBA and BL/6 BMDCs were cultured, with or without schistosome eggs, for 24 h; phosphorylated
versus total SRC, RAF-1, ERK1/2, JNK, and p38 expression was assessed by Western blot, and relative band density was quantified. Data are from one
representative experiment of four.
The Journal of Immunology 4663
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

6. Fanning, M. M., P. A. Peters, R. S. Davis, J. W. Kazura, and A. A. Mahmoud.1981. Immunopathology of murine infection with Schistosoma mansoni: relation-ship of genetic background to hepatosplenic disease and modulation. J. Infect. Dis.144: 148–153.
7. Cheever, A. W., R. H. Duvall, T. A. Hallack, Jr., R. G. Minker, J. D. Malley, andK. G. Malley. 1987. Variation of hepatic fibrosis and granuloma size among mousestrains infected with Schistosoma mansoni. Am. J. Trop. Med. Hyg. 37: 85–97.
8. Hernandez, H. J., W. C. Trzyna, J. S. Cordingley, P. H. Brodeur, andM. J. Stadecker. 1997. Differential antigen recognition by T cell populationsfrom strains of mice developing polar forms of granulomatous inflammation inresponse to eggs of Schistosoma mansoni. Eur. J. Immunol. 27: 666–670.
9. Rutitzky, L. I., J. R. Lopes da Rosa, and M. J. Stadecker. 2005. Severe CD4T cell-mediated immunopathology in murine schistosomiasis is dependent onIL-12p40 and correlates with high levels of IL-17. J. Immunol. 175: 3920–3926.
10. Rutitzky, L. I., L. Bazzone, M. G. Shainheit, B. Joyce-Shaikh, D. J. Cua, andM. J. Stadecker. 2008. IL-23 is required for the development of severe egg-induced immunopathology in schistosomiasis and for lesional expression ofIL-17. J. Immunol. 180: 2486–2495.
11. Shainheit, M. G., P. M. Smith, L. E. Bazzone, A. C. Wang, L. I. Rutitzky, andM. J. Stadecker. 2008. Dendritic cell IL-23 and IL-1 production in response toschistosome eggs induces Th17 cells in a mouse strain prone to severe immu-nopathology. J. Immunol. 181: 8559–8567.
12. Shainheit, M. G., K. W. Lasocki, E. Finger, B. M. Larkin, P. M. Smith,A. H. Sharpe, C. A. Dinarello, L. I. Rutitzky, and M. J. Stadecker. 2011. Thepathogenic Th17 cell response to major schistosome egg antigen is sequentiallydependent on IL-23 and IL-1b. J. Immunol. 187: 5328–5335.
13. Pearce, E. J., P. Caspar, J. M. Grzych, F. A. Lewis, and A. Sher. 1991. Down-regulation of Th1 cytokine production accompanies induction of Th2 responsesby a parasitic helminth, Schistosoma mansoni. J. Exp. Med. 173: 159–166.
14. Dong, C. 2006. Diversification of T-helper-cell lineages: finding the family rootof IL-17-producing cells. Nat. Rev. Immunol. 6: 329–333.
15. Bettelli, E., M. Oukka, and V. K. Kuchroo. 2007. T(H)-17 cells in the circle ofimmunity and autoimmunity. Nat. Immunol. 8: 345–350.
16. Stockinger, B., and M. Veldhoen. 2007. Differentiation and function of Th17T cells. Curr. Opin. Immunol. 19: 281–286.
17. Iwakura, Y., S. Nakae, S. Saijo, and H. Ishigame. 2008. The roles of IL-17A ininflammatory immune responses and host defense against pathogens. Immunol.Rev. 226: 57–79.
18. Mbow, M., B. M. Larkin, L. Meurs, L. J. Wammes, S. E. de Jong, L. A. Labuda,M. Camara, H. H. Smits, K. Polman, T. N. Dieye, et al. 2013. T-helper 17 cellsare associated with pathology in human schistosomiasis. J. Infect. Dis. 207:186–195.
19. Medzhitov, R. 2007. TLR-mediated innate immune recognition. Semin. Immu-nol. 19: 1–2.
20. Takeuchi, O., and S. Akira. 2010. Pattern recognition receptors and inflamma-tion. Cell 140: 805–820.
21. Robinson, M. J., D. Sancho, E. C. Slack, S. LeibundGut-Landmann, andC. Reis e Sousa. 2006. Myeloid C-type lectins in innate immunity. Nat. Immunol.7: 1258–1265.
22. Geijtenbeek, T. B., and S. I. Gringhuis. 2009. Signalling through C-type lectinreceptors: shaping immune responses. Nat. Rev. Immunol. 9: 465–479.
23. Cummings, R. D., and A. K. Nyame. 1999. Schistosome glysoconjugates. Bio-chim. Biophys. Acta 1455: 363–374.
24. Hokke, C. H., and A. M. Deelder. 2001. Schistosome glycoconjugates in host-parasite interplay. Glycoconj. J. 18: 573–587.
25. van Die, I., S. J. van Vliet, A. K. Nyame, R. D. Cummings, C. M. Bank,B. Appelmelk, T. B. Geijtenbeek, and Y. van Kooyk. 2003. The dendritic cell-specific C-type lectin DC-SIGN is a receptor for Schistosoma mansoni eggantigens and recognizes the glycan antigen Lewis x. Glycobiology 13: 471–478.
26. Hokke, C. H., and M. Yazdanbakhsh. 2005. Schistosome glycans and innateimmunity. Parasite Immunol. 27: 257–264.
27. Reimand, J., T. Arak, and J. Vilo. 2011. g:Profiler–a web server for functionalinterpretation of gene lists (2011 update). Nucleic Acids Res. 39(Web Serverissue): W307–315.
28. Park, C. G., K. Takahara, E. Umemoto, Y. Yashima, K. Matsubara, Y. Matsuda,B. E. Clausen, K. Inaba, and R. M. Steinman. 2001. Five mouse homologues ofthe human dendritic cell C-type lectin, DC-SIGN. Int. Immunol. 13: 1283–1290.
29. Cheong, C., I. Matos, J. H. Choi, D. B. Dandamudi, E. Shrestha, M. P. Longhi,K. L. Jeffrey, R. M. Anthony, C. Kluger, G. Nchinda, et al. 2010. Microbialstimulation fully differentiates monocytes to DC-SIGN/CD209(+) dendritic cellsfor immune T cell areas. Cell 143: 416–429.
30. Powlesland, A. S., E. M. Ward, S. K. Sadhu, Y. Guo, M. E. Taylor, andK. Drickamer. 2006. Widely divergent biochemical properties of the completeset of mouse DC-SIGN-related proteins. J. Biol. Chem. 281: 20440–20449.
31. van Vliet, S. J., J. J. Garcıa-Vallejo, and Y. van Kooyk. 2008. Dendritic cells andC-type lectin receptors: coupling innate to adaptive immune responses. Immunol.Cell Biol. 86: 580–587.
32. Geijtenbeek, T. B., R. Torensma, S. J. van Vliet, G. C. van Duijnhoven,G. J. Adema, Y. van Kooyk, and C. G. Figdor. 2000. Identification of DC-SIGN,a novel dendritic cell-specific ICAM-3 receptor that supports primary immuneresponses. Cell 100: 575–585.
33. Geijtenbeek, T. B., D. S. Kwon, R. Torensma, S. J. van Vliet, G. C. vanDuijnhoven, J. Middel, I. L. Cornelissen, H. S. Nottet, V. N. KewalRamani,D. R. Littman, et al. 2000. DC-SIGN, a dendritic cell-specific HIV-1-bindingprotein that enhances trans-infection of T cells. Cell 100: 587–597.
34. Caparros, E., P. Munoz, E. Sierra-Filardi, D. Serrano-Gomez, A. Puig-Kroger,J. L. Rodrıguez-Fernandez, M. Mellado, J. Sancho, M. Zubiaur, and A. L. Corbı.
2006. DC-SIGN ligation on dendritic cells results in ERK and PI3K activationand modulates cytokine production. Blood 107: 3950–3958.
35. Gringhuis, S. I., J. den Dunnen, M. Litjens, B. van Het Hof, Y. van Kooyk, andT. B. Geijtenbeek. 2007. C-type lectin DC-SIGN modulates Toll-like receptorsignaling via Raf-1 kinase-dependent acetylation of transcription factor NF-kappaB. Immunity 26: 605–616.
36. Gringhuis, S. I., J. den Dunnen, M. Litjens, M. van der Vlist, andT. B. Geijtenbeek. 2009. Carbohydrate-specific signaling through the DC-SIGNsignalosome tailors immunity to Mycobacterium tuberculosis, HIV-1 and Heli-cobacter pylori. Nat. Immunol. 10: 1081–1088.
37. Johnson, T. R., J. S. McLellan, and B. S. Graham. 2012. Respiratory syncytialvirus glycoprotein G interacts with DC-SIGN and L-SIGN to activate ERK1 andERK2. J. Virol. 86: 1339–1347.
38. Bettelli, E., Y. Carrier, W. Gao, T. Korn, T. B. Strom, M. Oukka, H. L. Weiner,and V. K. Kuchroo. 2006. Reciprocal developmental pathways for the generationof pathogenic effector TH17 and regulatory T cells. Nature 441: 235–238.
39. Sansonetti, P. J. 2006. The innate signaling of dangers and the dangers of innatesignaling. Nat. Immunol. 7: 1237–1242.
40. Kanneganti, T. D., M. Lamkanfi, and G. Nunez. 2007. Intracellular NOD-likereceptors in host defense and disease. Immunity 27: 549–559.
41. Van Liempt, E., A. Imberty, C. M. Bank, S. J. Van Vliet, Y. Van Kooyk,T. B. Geijtenbeek, and I. Van Die. 2004. Molecular basis of the differences inbinding properties of the highly related C-type lectins DC-SIGN and L-SIGN toLewis X trisaccharide and Schistosoma mansoni egg antigens. J. Biol. Chem.279: 33161–33167.
42. Meyer, S., E. van Liempt, A. Imberty, Y. van Kooyk, H. Geyer, R. Geyer, andI. van Die. 2005. DC-SIGN mediates binding of dendritic cells to authenticpseudo-LewisY glycolipids of Schistosoma mansoni cercariae, the first parasite-specific ligand of DC-SIGN. J. Biol. Chem. 280: 37349–37359.
43. van Vliet, S. J., E. van Liempt, E. Saeland, C. A. Aarnoudse, B. Appelmelk,T. Irimura, T. B. Geijtenbeek, O. Blixt, R. Alvarez, I. van Die, and Y. van Kooyk.2005. Carbohydrate profiling reveals a distinctive role for the C-type lectin MGLin the recognition of helminth parasites and tumor antigens by dendritic cells.Int. Immunol. 17: 661–669.
44. van Liempt, E., S. J. van Vliet, A. Engering, J. J. Garcıa Vallejo, C. M. Bank,M. Sanchez-Hernandez, Y. van Kooyk, and I. van Die. 2007. Schistosomamansoni soluble egg antigens are internalized by human dendritic cells throughmultiple C-type lectins and suppress TLR-induced dendritic cell activation. Mol.Immunol. 44: 2605–2615.
45. Cass, C. L., J. R. Johnson, L. L. Califf, T. Xu, H. J. Hernandez, M. J. Stadecker,J. R. Yates, III, and D. L. Williams. 2007. Proteomic analysis of Schistosomamansoni egg secretions. Mol. Biochem. Parasitol. 155: 84–93.
46. van Stijn, C. M., S. Meyer, M. van den Broek, S. C. Bruijns, Y. van Kooyk,R. Geyer, and I. van Die. 2010. Schistosoma mansoni worm glycolipids inducean inflammatory phenotype in human dendritic cells by cooperation of TLR4 andDC-SIGN. Mol. Immunol. 47: 1544–1552.
47. Meevissen, M. H., M. Yazdanbakhsh, and C. H. Hokke. 2012. Schistosomamansoni egg glycoproteins and C-type lectins of host immune cells: molecularpartners that shape immune responses. Exp. Parasitol. 132: 14–21.
48. Ritter, M., O. Gross, S. Kays, J. Ruland, F. Nimmerjahn, S. Saijo, J. Tschopp,L. E. Layland, and C. Prazeres da Costa. 2010. Schistosoma mansoni triggersDectin-2, which activates the Nlrp3 inflammasome and alters adaptive immuneresponses. Proc. Natl. Acad. Sci. USA 107: 20459–20464.
49. Saunders, S. P., C. M. Walsh, J. L. Barlow, N. E. Mangan, P. R. Taylor,A. N. McKenzie, P. Smith, and P. G. Fallon. 2009. The C-type lectin SIGNR1binds Schistosoma mansoni antigens in vitro, but SIGNR1-deficient micehave normal responses during schistosome infection. Infect. Immun. 77: 399–404.
50. Feinberg, H., D. A. Mitchell, K. Drickamer, and W. I. Weis. 2001. Structuralbasis for selective recognition of oligosaccharides by DC-SIGN and DC-SIGNR.Science 294: 2163–2166.
51. Figdor, C. G., Y. van Kooyk, and G. J. Adema. 2002. C-type lectin receptors ondendritic cells and Langerhans cells. Nat. Rev. Immunol. 2: 77–84.
52. Lu, X. J., J. Chen, C. H. Yu, Y. H. Shi, Y. Q. He, R. C. Zhang, Z. A. Huang,J. N. Lv, S. Zhang, and L. Xu. 2013. LECT2 protects mice against bacterialsepsis by activating macrophages via the CD209a receptor. J. Exp. Med.210: 5–13.
53. Wellbrock, C., M. Karasarides, and R. Marais. 2004. The RAF proteins takecentre stage. Nat. Rev. Mol. Cell Biol. 5: 875–885.
54. Kolch, W. 2005. Coordinating ERK/MAPK signalling through scaffolds andinhibitors. Nat. Rev. Mol. Cell Biol. 6: 827–837.
55. Numazaki, M., C. Kato, Y. Kawauchi, T. Kajiwara, M. Ishii, and N. Kojima.2009. Cross-linking of SIGNR1 activates JNK and induces TNF-alpha produc-tion in RAW264.7 cells that express SIGNR1. Biochem. Biophys. Res. Commun.386: 202–206.
56. Tanne, A., B. Ma, F. Boudou, L. Tailleux, H. Botella, E. Badell, F. Levillain,M. E. Taylor, K. Drickamer, J. Nigou, et al. 2009. A murine DC-SIGN homo-logue contributes to early host defense against Mycobacterium tuberculosis. J.Exp. Med. 206: 2205–2220.
57. Zhu, L., Y. Wu, H. Wei, S. Yang, N. Zhan, X. Xing, and B. Peng. 2012. Up-regulation of IL-23 p19 expression in human periodontal ligament fibroblasts byIL-1b via concurrent activation of the NF-kB and MAPKs/AP-1 pathways.Cytokine 60: 171–178.
58. Thomas, P. G., M. R. Carter, O. Atochina, A. A. Da’Dara, D. Piskorska,E. McGuire, and D. A. Harn. 2003. Maturation of dendritic cell 2 phenotype bya helminth glycan uses a Toll-like receptor 4-dependent mechanism. J. Immunol.171: 5837–5841.
4664 DC CD209a INDUCES Th17 RESPONSE IN SCHISTOSOMIASIS
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

59. Kane, C. M., L. Cervi, J. Sun, A. S. McKee, K. S. Masek, S. Shapira,C. A. Hunter, and E. J. Pearce. 2004. Helminth antigens modulate TLR-initiateddendritic cell activation. J. Immunol. 173: 7454–7461.
60. Goodridge, H. S., M. R. Deehan, W. Harnett, and M. M. Harnett. 2005. Sub-version of immunological signalling by a filarial nematode phosphorylcholine-containing secreted product. Cell. Signal. 17: 11–16.
61. Carvalho, L., J. Sun, C. Kane, F. Marshall, C. Krawczyk, and E. J. Pearce. 2009.Review series on helminths, immune modulation and the hygiene hypothesis:mechanisms underlying helminth modulation of dendritic cell function. Immu-nology 126: 28–34.
62. Harnett, M. M., A. J. Melendez, and W. Harnett. 2010. The therapeutic potentialof the filarial nematode-derived immunodulator, ES-62 in inflammatory disease.Clin. Exp. Immunol. 159: 256–267.
63. van Riet, E., F. C. Hartgers, and M. Yazdanbakhsh. 2007. Chronic helminthinfections induce immunomodulation: consequences and mechanisms. Immu-nobiology 212: 475–490.
64. Everts, B., G. Perona-Wright, H. H. Smits, C. H. Hokke, A. J. van der Ham,C. M. Fitzsimmons, M. J. Doenhoff, J. van der Bosch, K. Mohrs, H. Haas, et al.
2009. Omega-1, a glycoprotein secreted by Schistosoma mansoni eggs, drivesTh2 responses. J. Exp. Med. 206: 1673–1680.
65. Steinfelder, S., J. F. Andersen, J. L. Cannons, C. G. Feng, M. Joshi, D. Dwyer,P. Caspar, P. L. Schwartzberg, A. Sher, and D. Jankovic. 2009. The majorcomponent in schistosome eggs responsible for conditioning dendritic cells forTh2 polarization is a T2 ribonuclease (omega-1). J. Exp. Med. 206: 1681–1690.
66. Klaver, E. J., L. M. Kuijk, L. C. Laan, H. Kringel, S. J. van Vliet, G. Bouma,R. D. Cummings, G. Kraal, and I. van Die. 2013. Trichuris suis-induced modulationof human dendritic cell function is glycan-mediated. Int. J. Parasitol. 43: 191–200.
67. LeibundGut-Landmann, S., O. Gross, M. J. Robinson, F. Osorio, E. C. Slack,S. V. Tsoni, E. Schweighoffer, V. Tybulewicz, G. D. Brown, J. Ruland, andC. Reis e Sousa. 2007. Syk- and CARD9-dependent coupling of innate immunityto the induction of T helper cells that produce interleukin 17. Nat. Immunol. 8:630–638.
68. Gringhuis, S. I., T. M. Kaptein, B. A. Wevers, B. Theelen, M. van der Vlist,T. Boekhout, and T. B. Geijtenbeek. 2012. Dectin-1 is an extracellular pathogensensor for the induction and processing of IL-1b via a noncanonical caspase-8inflammasome. Nat. Immunol. 13: 246–254.
The Journal of Immunology 4665
by guest on July 10, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























