Embed Size (px)
Citation preview

Biological Conservation 170 (2014) 110–119
Contents lists available at ScienceDirect
Biological Conservation
journal homepage: www.elsevier .com/locate /b iocon
Capacity of older plantations to host bird assemblagesof naturally-regenerated conifer forests: A test at standand landscape levels
0006-3207/$ - see front matter � 2014 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.biocon.2013.12.023
⇑ Corresponding author. Tel.: +1 506 858 4292.E-mail addresses: [email protected] (A. MacKay), allardmax@icloud.
com (M. Allard), [email protected] (M.-A. Villard).1 Tel.: +1819 870 1917.
Allison MacKay a,1, Maxime Allard b, Marc-André Villard a,⇑a Département de biologie, Université de Moncton, Moncton, New Brunswick E1A 3E9, Canadab Département des sciences biologiques, Université du Québec à Montréal, Montréal, Québec H3C 3P8, Canada
a r t i c l e i n f o a b s t r a c t
Article history:Received 7 April 2013Received in revised form 10 December 2013Accepted 16 December 2013
Keywords:Bird communitiesForest managementNesting guildsCavity-nesting birdsBayesian hierarchical modelsSilviculture
Conifer plantations are thought to provide lower-quality habitat for many old forest specialists because oftheir simplified stand structure and relative paucity of habitat elements such as dead wood. However, atthe landscape scale, plantations may have a positive influence on biodiversity, e.g. through landscapesupplementation. Here, we test the quality of older spruce plantations as breeding habitat for forest birdsat local and landscape levels. We compared bird species assemblages of 40–50 year-old native spruceplantations with those of mature, naturally-regenerated conifer forests using a Bayesian hierarchicalmodel, which accounts for imperfect detection of species. Median species richness was higher in natu-rally-regenerated conifer stands than in spruce plantations (29.2 vs. 24.4; 95% probability inter-val = 3.9–5.7), which probably reflects the absence of primary cavity nesters from many stationswithin plantations. In contrast, more species of canopy nesters were detected in plantations. The propor-tion of naturally-regenerated conifer forest in the landscape was a predictor of species occurrence for fiveground nesters, four primary cavity nesters, and two weak excavators. Older spruce plantations did notprovide suitable habitat for some species, mainly cavity nesters. Snag densities in plantations were lowerthan the estimated threshold requirements for several cavity-nesting birds. Hence, mature, naturally-regenerated forest patches are essential components of intensively-managed forest landscapes tomaintain rich forest bird assemblages.
� 2014 Elsevier Ltd. All rights reserved.
1. Introduction
Plantation silviculture is extensively used to increase timberyields and their predictability. As of 2010, planted forests com-prised 7% of the world’s total forest area, increasing at a rate of5 million hectares per year (FAO, 2010). The potential implicationsof plantation silviculture for biodiversity conservation at local andlandscape scales have fuelled debate (Waldick et al., 1999; Bettset al., 2005) because planted stands often feature a single speciesof tree and a simplified structure compared to naturally-regener-ated forest stands. These characteristics, paired with generallyshort-rotation management, make plantations unsuitable to manyspecies associated with old forests (Barlow et al., 2007; du Bus deWarnaffe and Deconchat, 2008).
There may be a negative bias against the conservation valueof tree plantations as a result of unfair comparisons between
young, non-native plantations and mature native stands(Stephens and Wagner, 2007). A reduction in species richnessin non-native tree plantations has been particularly well docu-mented (Hartley, 2002; Haskell et al., 2006; Bremer and Farley,2010; Thinh et al., 2012). However, evidence regarding the con-tribution of native tree plantations to biodiversity is equivocal.Some studies (Barlow et al., 2007; Farwig et al., 2008; Pietrekand Branch, 2011) have reported neutral or positive effectswhereas others have reported lower biodiversity in nativetree plantations than in natural stands (Waldick et al., 1999;Brockerhoff et al., 2008). Furthermore, the scale at which biodi-versity is assessed can be misleading. The use of species richnessalone as an indicator can be problematic because it does notaccount for species composition (Canterbury et al., 2001). Onthe other hand, using one or even a few focal species (e.g. Gjerdeet al., 2005; Barrientos, 2010) as indicators of overall biodiversitymay fail to protect non-target species (Lindenmayer, 1999;Carignan and Villard, 2002; Roberge and Angelstam, 2006). Thus,we used both species occurrence in relation to habitat variables,and overall species richness as indicators of bird response toplantation silviculture.

A. MacKay et al. / Biological Conservation 170 (2014) 110–119 111
Even if they host lower stand-level biodiversity, commercialplantations may benefit landscape-level biodiversity by (1) supple-menting or complementing resources available in nearby habitat,(2) facilitating dispersal, or (3) creating a buffer zone around hab-itat fragments that reduces negative edge effects (Fischer and Lin-denmayer, 2006; Brockerhoff et al., 2008). If landscapesupplementation (sensu Dunning et al., 1992) takes place, planta-tions surrounded by high proportions of naturally-regenerated for-est would be expected to host a greater number of mature forestspecialists. However, plantations may also act as an ecological trapfor certain species (Remeš, 2003; Hollander et al., 2012) or theymay reduce landscape permeability for species using contrastinghabitat, e.g. spruce plantations from the perspective of a deciduousforest bird (Villard and Haché, 2012).
As a rule, the value of plantations for biodiversity conservationdepends on pre-plantation land use (Brockerhoff et al., 2008; Bre-mer and Farley, 2010). When established on degraded land, planta-tions, especially of native trees, can increase biodiversity (Stephensand Wagner, 2007), whereas those established after harvestingnaturally-regenerated forests tend to have negative effects. Nega-tive effects can often be attributed to a paucity of habitat compo-nents associated with tree senescence, such as dead wood, or tolow structural complexity (e.g. few canopy gaps, sparse under-story). Even-aged structure and intensive silvicultural treatmentstend to reduce dead wood volume (Kirby et al., 1998; Johnsonand Freedman, 2003; Paquette and Messier, 2011). Furthermore,plantations fragment continuous natural forest cover, are usuallycomposed of only one or a few tree species, and their understorycomposition tends to be simplified (Paritsis and Aizen, 2008),which has been shown to reduce species richness of forest birds(Estades and Temple, 1999).
Researchers have investigated the effects of plantation silvicul-ture on species associated with dead wood that belong to severaltaxa, e.g. bryophytes and lichens (Humphrey et al., 2002), fungi(Amaranthus et al., 1994), invertebrates (Brin et al., 2009, 2011),birds (Gjerde et al., 2005; Barrientos, 2010), and mammals (McCayand Komoroski, 2004). Most of these studies suggest increasingdead wood in plantations to increase their conservation value. Thismay be a viable option in plantations established for soil and waterconservation or for wind protection (Carnus et al., 2006), but whenwood production is the main objective, allowing substantial num-bers of planted trees to die in order to create deadwood may rep-resent significant economic losses. However, restoration efforts inintensively-managed forests of Fennoscandia have had some suc-cess. For example, Berglund et al. (2011) found that prescribedburning and artificial creation of dead wood (up to 60 m3 ha�1)positively influenced the diversity and occurrence of fungi.
This study was conducted in an intensively managed forestdistrict of New Brunswick, Canada, which features some of thefirst plantations established in the province. The proportion ofland devoted to plantations continues to increase in New Bruns-wick (New Brunswick Department of Natural Resources, 2009)although controversy exists regarding their ecological implica-tions (Waldick et al., 1999; Betts et al., 2005). A study conductedin southeastern New Brunswick indicated that bird assemblagesfrom spruce plantations tended to converge towards those of ma-ture, naturally-regenerated stands as they reached 18 years of age(Parker et al., 1994). In the same region, Johnson and Freedman(2003) documented avian use of 3–21 year old spruce plantationsand found that after 13 years, some bird species typical of mature(65–85 years old) natural conifer forests were present. However,the latter authors predicted that older plantations would not sup-port dead wood-dependent bird species. In addition, they pre-dicted that bird species requiring large-diameter trees wouldalso likely be absent. Studies are thus needed to examinebiodiversity in older plantations, but they must account for the
potential influence of landscape context (Betts et al., 2005;St-Laurent et al., 2007).
In this study, we aimed to determine whether point richness offorest birds in older (40–50 year-old) native spruce plantationsdoes, in fact, converge towards that of mature, naturally-regener-ated conifer forests. Bird occurrence, abundance, and fitnessparameters are considered to be valuable indicators of the effectsof human activities at a variety of spatial scales (i.e. from micro-habitat to landscape) (Carignan and Villard, 2002). In addition tobeing relatively easy to detect, forest birds include keystonespecies such as woodpeckers (Virkkala, 2006; Drever et al., 2008;Mikusinski et al., 2001), which provide cavities that are essentialfor the survival and reproduction of a number of other vertebrate,and even some invertebrate species (Bonar, 2000; Martin et al.,2004).
We applied a Bayesian hierarchical model inspired from that ofZipkin et al. (2010) to compare avian diversity between coniferplantations and naturally-regenerated conifer stands at three lev-els: overall species richness, richness of predefined nesting guilds,and occurrence of individual species. The model estimated speciesresponses to stand and landscape-level parameters a posteriori,thus avoiding the bias associated with the a priori selection of indi-cator species. We hypothesized that, even at commercial maturity,spruce plantations would provide habitat of lower quality thanthat of mature, naturally-regenerated conifer stands for manyold-forest specialists, owing to the paucity of critical structuressuch as dead wood and large-diameter trees. Hence, we predictedthat species richness would be higher in naturally-regeneratedconifer stands and that this difference would be most evident innesting guilds comprised of species dependent on dead wood orlarge-diameter trees (e.g., cavity nesters). We also predicted thatold-forest specialists would exhibit positive species-specific re-sponses to the proportion of naturally-regenerated conifer forestin the landscape. We further hypothesized that bird species rich-ness in older spruce plantations would increase with the propor-tion of naturally-regenerated forest in the surroundings becauseolder plantations can supplement resources.
2. Methodology
2.1. Study area and experimental design
The study was conducted in the Black Brook District (47�230N,67�400W), a privately-owned managed forest (ca. 200,000 ha) lo-cated in northwestern New Brunswick (Fig. 1). The District is lo-cated on an undulating plateau, historically covered by aheterogeneous mosaic of tolerant deciduous, mixed and coniferousforest (New Brunswick Department of Natural Resources, 2013).Commonly occurring native coniferous species are red, white,and black spruce (Picea rubens, Picea glauca; Picea mariana) and bal-sam fir (Abies balsamea). Since its purchase by J.D. Irving Ltd. (JDI)in 1944, the District has undergone important changes in land-scape composition (Etheridge et al., 2005) and it now comprisessome of the most intensively-managed forest land in Canada(Porter et al., 2004; Montigny and MacLean, 2005). Conifer planta-tions were established in 1957 and now represent ca. 37% of theDistrict’s area. The main species planted are white and blackspruce. Native red spruce, white pine (Pinus strobus) andnon-native Norway spruce (Picea abies) are also planted, but to alesser extent. Naturally-regenerated conifer forests represent only4% of the land base. These stands are mostly designated as bench-mark reserves and are protected from harvesting for conservationand scientific research (MacLean et al., 2010), although some haveundergone light salvage harvesting following Spruce Budworm(Choristoneura fumiferana) outbreaks (G. Adams, personal commu-

Fig. 1. Map of the study area in New Brunswick (inset) and location of point count stations (dots) within 40–50 year-old plantations and NRF. Streams and rivers (curvedlines) and roads (straight lines) are also shown.
112 A. MacKay et al. / Biological Conservation 170 (2014) 110–119
nication). In naturally regenerated stands, the dominant coniferspecies are white, black, and red spruces, balsam fir, as well asEastern white cedar (Thuja occidentalis) in poorly-drained areas.
We surveyed two different forest types to characterize andcompare their bird assemblages: naturally-regenerated, matureconifer stands (hereafter, NRF) (n = 42 point count stations) andcommercially mature (40–50 year old) spruce plantations (n = 60)(Fig. 1). NRF had a more important and diverse deciduous compo-nent than did conifer plantations. However, we only conducted
surveys in stands that were dominated by conifers and that werelarge enough to accommodate P 2 point count stations separatedby 250 m. Due to its scarcity in the District, most of the NRF thatmet our selection criteria was sampled. Plantations were selecteda priori from a forest inventory map and final selection was basedon accessibility, age (P40 years old) and area (P2 stations).Although silvicultural intensity varied among spruce plantations,most had received pre-commercial and commercial thinning treat-ments, and had been subjected to post-planting herbicide spraying

A. MacKay et al. / Biological Conservation 170 (2014) 110–119 113
to control competing deciduous undergrowth (G. Adams, personalcommunication). Besides planted spruces, balsam fir and tremblingaspen (Populus tremuloides) were generally present as scatteredtrees.
Within the stands that met our selection criteria, we positionedpoint count stations a priori on the georeferenced forest inventoryin order to maximize the number of stations while meeting the fol-lowing constraints (Fig. 1). Stations had to be separated by at least250 m in order to reduce the risk of double-counting individualbirds. They also had to be located at least 150 m from the nearestedge (i.e. road, recent cutblock, or distinct forest type) to reducepotential edge effects. The forest stands selected, henceforth re-ferred to as study sites, could fit 2–6 point count stations.
2.2. Bird surveys
We recorded species presence using a modified point countmethod, including two silent observation periods of 5 min sepa-rated by a 5 min playback of selected species. The second 5 min si-lent period was useful to detect individuals who were late inresponding to the playback. The species whose vocalizations ordrumming were broadcast are Black-backed and American Three-toed Woodpeckers (Picoides arcticus and Picoides dorsalis), BrownCreeper (Certhia americana), Red-breasted Nuthatch (Sitta canaden-sis), and Boreal Chickadee (Poecile hudsonicus), as well as BarredOwl (Strix varia) and Gray Jay (Perisoreus canadensis). The lattertwo species were included in an attempt to elicit mobbing re-sponses and, in turn, to increase the detectability of wide-rangingspecies when they did not respond to conspecific sounds. Recordedtracks were compiled from Dendroica (http://www.naturein-struct.org/dendroica). Point count surveys were conducted be-tween 25 May and 1 July in 2010 and 2011, from 0500 to 1000.Only bird detections estimated to be within a 100-m radius wereincluded in the analysis. Each station was visited three times toestimate species detectability. Birds flying over the canopy wereexcluded from the analyses. For species with large territories andwhose calls or drumming can be heard over long distances (i.e.woodpeckers), we estimated their distance to the point count byfollowing the call after the point count was completed. Also, ifone of these species was detected at two consecutive point countsand we could not be sure that they were different individuals (noresponse call, same sex), we attributed the occurrence to the firstpoint count station at which the individual was detected. Eachyear, the two observers sampled every station at least once to re-duce observer bias.
2.3. Local habitat and landscape metrics
We sampled habitat variables at and around (50 m in each maincardinal direction) each point count station, for a total of 5 habitatplots/station. Within a circular plot of 0.04 ha, we measured thedensity, species, status (alive or dead), and diameter at breast high(dbh) of all trees (dbh P 8).
We estimated the proportion of NRF within a 700 m radius ofeach point count station using a georeferenced forest inventorymap provided by JDI and Quantum GIS (version 1.8.0). This radiuswas chosen to be large enough to encompass the typical homerange of the most area-demanding species we detected, theBlack-backed Woodpecker (150 ha; Tremblay et al., 2010), yet re-stricted enough to allow for heterogeneity in landscape composi-tion around each sampling point and avoid pseudoreplication.NRF is sparsely distributed in the study area (ca. 4% of the District;Etheridge et al., 2005) and is mostly set aside as reserves. It is dif-ficult to assign a precise age to these reserves owing to post-bud-worm salvage harvesting.
2.4. Statistical analyses
We adapted the hierarchical community model developed byZipkin et al. (2010) to incorporate multiple site-level replicates.This Bayesian modeling approach is an extension of the single spe-cies occurrence model (MacKenzie et al., 2002), hence it providesestimates of species richness that are corrected for imperfectdetection of individual species as well as direct species-level re-sponses to habitat variables. Species-level responses are modeledas a function of habitat (spruce snags and large-diameter trees)and landscape (proportion of NRF) variables.
Even though rare species are often of particular conservationconcern and may exhibit unique responses to landscape alteration,it is often difficult to include them in occurrence models becausethey are detected too infrequently (MacKenzie et al., 2005). Thehierarchical model (Zipkin et al., 2010) uses collective trends inthe species assemblage derived from commonly co-occurring spe-cies in the data set to adjust detection probabilities of uncommonspecies. Thus, there is no minimum threshold occurrence value forinclusion in the model. This modeling approach also allowed us toincorporate habitat and landscape variables and to analyze speciesoccurrence in response to these variables. We used JAGS, a pro-gram which preforms Markov Chain Monte Carlo (MCMC) simula-tion (Plummer, 2003), to run the model. Model convergence wasverified through the investigation of trace plots and the R-hat sta-tistic (Brooks and Gelman, 1998).
Species were grouped into five nesting guilds: ground nesters,understory nesters, canopy nesters, primary cavity nesters [wood-peckers and Brown Creeper (C. americana)], and weak excavators[chickadees and the Red-breasted Nuthatch (S. canadensis)](Appendix A, Table A1). Nesting guilds have been found to be moreclosely associated with habitat structure than guilds determinedaccording to foraging ecology (Canterbury et al., 2001).
Using estimated species richness values obtained from theBayesian hierarchical model, we related median species richnessto the aforementioned habitat variables using generalized linearmodels (glm) (Hoffmann, 2003). A separate model was createdfor total species richness as well as for each nesting guild. We ad-dressed model selection uncertainty by averaging the parametersof the top-fit models (DAIC < 2) using the MuMIn package in R(Barton, 2013). We considered two-way interactions, but nothigher-level interactions, and only those holding biologically-meaningful interpretations.
2.5. Modeling framework
In our models, species occurrence z(i, j) is defined as a binaryvariable in which z(i, j) = 1 if species i occurs within 100 m of sta-tion j, and z(i, j) = 0 otherwise. The occurrence state is assumed tobe the outcome of a Bernoulli random variable (z(i, j) � Bern(wi,j)),where wi,j is the probability that species i occurs at station j. Imper-fect observation of true occurrence, and therefore the estimate ofwi,j, is corrected through a detection model of the observed datawhich allows for distinction between true absence and non-detec-tion (MacKenzie et al., 2002). Temporal replicates (k = 3) for eachstation (j) were performed within the same breeding season, forwhich we applied the closure assumption. Spot mapping was con-ducted repeatedly in a few study sites prior to the beginning of thesampling period to ensure that migratory species had begun set-tling on their territories prior to the beginning of point count sur-veys. We define detection as x(i, j,k) � Bern(pi,j,k z(i,j)), where pi,j,k isthe detection probability of species i at station j during the tempo-ral replicate k.
Occurrence (wi,j) probabilities at the species level may reflectstand characteristics (density of spruce snags P 20 cm dbh andlive large-diameter trees P 30 cm dbh), landscape composition

114 A. MacKay et al. / Biological Conservation 170 (2014) 110–119
(proportion of NRF within 700 m), or simply forest type (plantationor NRF). The environmental variables were standardized and incor-porated into the model using the logit link function as fixed vari-ables, whereas forest type was incorporated as a two-way factor.Similarly, study site was added as a random variable to accountfor spatial clustering of stations. The occurrence model was similarto that of Zipkin et al. (2010):
logitðw; i; jÞ ¼ Occði; j; Þ þ a1iðlarge trees jÞþ a2iðdeadwood jÞ þ a3iðlandscape jÞ; ð1Þ
where Occ(i, j) is the probability of occurrence, on a logit scale, forbird species i at station j. The coefficients pertaining to the threeai terms are the linear effects of the fixed variables on species i. Con-trary to Zipkin et al. (2010), we did not incorporate an augmentedmatrix and, therefore, did not estimate potentially unobserved spe-cies. The detection model was:
logitðpði; j; kÞÞ ¼ pi þ b1iðdatej;kÞ þ b2iðdate2j;kÞ ð2Þ
This model estimates the detection probability pi (also on a logitscale) as a function of the linear and squared effects of the samplingdate (coefficients for the two b terms). These covariates were stan-dardized to have a mean of zero and a standard deviation of one.
3. Results
3.1. Habitat structure
Average density of spruce snags P 8 cm dbh was higher in NRF(30.4 ± 3.8 SE; range = 0–110) stems ha�1 than in spruce plantation(4.4 ± 1.2 SE; range = 0–50) (Mann–Whitney U test, p < 0.001).Average density of spruce snags P 20 cm dbh was also higher inNRF (6.07 ± 0.75 SE) stems ha�1 (range = 0–22) than in spruceplantations (0.88 ± 0.23; range = 0–10). Finally, density oftrees P 30 cm dbh was higher in NRF than in plantations(17.8 ± 1.7 SE; range = 3–58 vs. 5.1 ± 0.61 SE; range = 0–25)(Mann–Whitney U test, p < 0.001).
3.2. Bird species assemblages
A total of 46 bird species were detected during our surveys,including 43 in plantations and 44 in NRF. Mean detection proba-bility varied widely among species (0.026–0.70). Estimated medianspecies richness (corrected for imperfect detection) was signifi-cantly higher in NRF than in plantations (29.2 vs. 24.4; t = 10.5;95% PI = 3.9–5.7). Variability in bird species richness was notice-ably higher at low values of either stand or landscape variablesin either forest type (Fig. 2).
3.3. Nesting guilds
Species richness of canopy nesters was higher in plantationsthan in NRF (10.2 vs. 9.1; 95% PI = �1.61 to �0.61; t = �4,4) andnegatively related to the proportion of NRF in the landscape(Fig. 2e). For each of the remaining guilds, species richness inNRF was higher than in plantations (Appendix A, Table A2). Thiswas most obvious for primary cavity nesters, whose mean speciesrichness in NRF was twice as high as in plantations (6.7 vs. 3.4; 95%PI = 3.02–3.78; t = 17.6). The response of primary cavity-nestingbirds to the proportion of NRF in the landscape and the local den-sity of spruce snags (Fig. 2c and d) reflected most closely the pat-tern observed for total species richness (Fig. 2a and b). In fact,the proportion of NRF in the adjacent landscape explained 76% ofthe variability in the species richness of primary cavity nesters(Fig. 2c).
3.4. Species-specific responses
None of the 46 species detected responded to the density ofspruce snags. Two ground-nesting species responded to the den-sity of large-diameter trees, whereas 12 species (a mixture of weakexcavators, primary cavity and ground-nesting species) respondedpositively to the proportion of NRF in the landscape (Table 1). Onlythe Bay-breasted Warbler (Setophaga castanea), a canopy nester,exhibited a negative response to the proportion of NRF in the land-scape. No other species from the canopy or understory nestingguilds responded significantly to any of the three habitat variables.
The model converged for all species, with the exception of theRed-breasted Nuthatch (S. canadensis) (R-hat value > 1.2). The nut-hatch was detected twice as often in 2010 (occurrence = 0.83) thanin 2011 (occurrence = 0.41), a significant difference (G = 6.638;d.f. = 1; p < 0.01). Furthermore, in 2011, its occurrence in planta-tions was 0.34 compared to 0.72 in 2010 (Appendix A, Fig. A1), a38% reduction (G = 29.03; d.f. = 1; p < 0.001). In contrast, therewas only a 17% reduction (1.00–0.83) in the occurrence of this spe-cies in NRF (G = 6.08; d.f. = 1; p = 0.01).
4. Discussion
This study aimed to determine the conservation value of com-mercially mature plantations of native spruce from the perspectiveof breeding forest birds. Bird species richness was higher in natu-rally-regenerated, mature conifer stands, probably as a result ofthe paucity of large-diameter snags in plantations. Indeed, the pro-portion of NRF in the landscape and the density of spruce snagswere important predictors of total species richness. Although med-ian species richness was significantly higher in NRF, the fact that astation was located within NRF (i.e. forest type) was not a predictorof high species richness. The rate at which species richness in-creased in response to the proportion of NRF and the density oflarge snags (which was also a significant predictor) was similarin plantations and NRF. Small or narrow patches of NRF tendedto be characterized by a lower species richness than larger NRFstands. The density of large-diameter trees in plantations was alsolow compared to NRF, but as a predictor of total species richness,this variable was less important than expected (see Table 2).
Guild-level results point out the importance of investigatingpatterns beyond those revealed by total species richness. Re-sponses to structural and landscape factors differed in both inten-sity and direction among the four nesting guilds modeled. Therewere too few species in the weak excavator guild (n = 3) to obtainguild-level trends. Of the four remaining nesting guilds, primarycavity nesters showed the greatest contrast in species richness be-tween plantations and NRF, the number of species being lower inplantations. None of the plantations hosted the maximum numberof cavity-nesting species encountered in NRF. Of the three covari-ates, the proportion of NRF in the landscape had the greatest influ-ence on species richness of primary cavity nesters, explaining 76%of the variance, but snag density was also a predictor of cavity ne-ster occurrence (see Table 2). Once again, the paucity of large-diameter snags in plantations may explain why the proportion ofNRF in the landscape was so important for the occurrence of cavitynesters.
Ground and understory nesters also occurred less frequently inplantations than in NRF (Appendix A, Table A2). The proportion ofNRF within 700 m contributed to explain the occurrence of groundnesters, whereas forest type was a significant predictor for under-story nesters. This may reflect the lower floristic diversity in theplantations, although we did not quantify this variable.
Conversely, species occurrence of canopy nesters was higher inplantations than in NRF and was negatively related to the propor-

Fig. 2. Variation in species richness of bird assemblages (a and b), and of cavity- (c and d) and canopy- (e) nesting guilds as a function of habitat and landscape variables asdetermined using generalized linear models. Open dots represent point count stations within 40–50 year-old plantations, whereas closed dots correspond to NRF. Dottedlines represent 95% confidence intervals.
A. MacKay et al. / Biological Conservation 170 (2014) 110–119 115
tion of NRF in the landscape. This is likely driven by species such asthe Bay-breasted Warbler (S. castanea). Plantations apparently of-fered plenty of good foraging space for this species: the basal areaof live spruce trees was almost twice as high in plantations(14.4 ± 6.0 m2 ha�1) as in NRF (7.7 ± 6.3 m2 ha�1).
Species-specific responses were consistent with trends found atthe nesting guild level. Species responding significantly were al-most exclusively associated with the proportion of NRF in the land-scape, and positive associations were limited to species from theground (4 species), primary cavity (5) and weak excavator (2) nest-ing guilds. Among these were old-forest specialists such as Black-backed and Pileated Woodpeckers (Dryocopus pileatus) (Settering-ton et al., 2000) and Brown Creeper (Poulin et al., 2008, 2013), aswell as both chickadee species, who require well-decayed snags
for nesting (New Brunswick Department of Natural Resources,2005). The significant influence of the proportion of NRF on thesespecies suggests that NRF plays an essential role in maintainingbiodiversity within intensively-managed landscapes.
With respect to landscape supplementation, results were incon-sistent. For isolated patches of equivalent size, NRF stands likelycontained more old forest habitat components (e.g. large snags)and hosted more old-forest specialists than plantations. However,occurrence of old forest specialists would have been expected toincrease in plantations located near NRF if those specialists useplantations to supplement NRF resources. Indeed, we expected thatspecies richness would increase at a faster rate in plantations thanin NRF as a function of the proportion of NRF in the landscape. If so,forest type or an interaction term including proportion of NRF

Table 1Species-specific responses to stand and landscape-level variables as estimated by a Bayesian hierarchical model. Linear responses are either positively (+) or negatively (�)correlated with the variable and are significant for 90% or 95% probability intervals.
Explanatory variable Probability interval (%) Species Direction of relationship Nesting guild
Density of snags (P20 cm dbh) 95 None na na90 None na na
Density of large trees (P30 cm dbh) 95 Ovenbird (Seiurus aurocapilla) + Ground90 White-throated Sparrow (Zonotrichia albicollis) + Ground
Proportion of NRF (700 m radius) 95 Black-backed Woodpecker (Picoides arcticus) + Primary cavityPileated Woodpecker (Dryocopus pileatus) + Primary cavityBlack-capped Chickadee (Poecile atricapillus) + Weak excavatorsCanada Warbler (Cardellina canadensis) + Ground
90 Northern Flicker (Colaptes auratus) + Primary cavityYellow-bellied Flycatcher (Empidonax flaviventris) + GroundBoreal Chickadee (Poecile hudsonicus) + Weak excavatorsBrown Creeper, (Certhia americana) + Primary cavityVeery (Catharus fuscescens) + GroundBay-breasted Warbler (Setophaga castanea) � CanopyMourning Warbler (Geothlypis philadelphia) + GroundNorthern Waterthrush (Parkesia noveboracensis) + Ground
Table 2Influence of site and landscape variables on species richness of the complete bird assemblage, and for each nesting guild separately, according to model averaging fromgeneralized linear models. Models with a delta AIC < 2 were averaged.
Variable Estimate Adj. SE z p
Total species richnessModel averaging included 2 models
Intercept 3.130 0.033 93.061 <0.001*
NRF 700 m 0.208 0.102 2.028 0.043*
Snags 0.072 0.034 2.088 0.037*
NRF 700 m: Snags �0.086 0.070 1.211 0.226
Species richness of ground-nesting speciesModel averaging included 4 models
Intercept 1.652 0.091 18.07 <0.001*
NRF 700 m 0.450 0.143 3.142 0.002*
Large trees 0.046 0.048 0.943 0.346Snags 0.026 0.052 0.495 0.621Forest type (plantations) 0.049 0.141 0.350 0.727
Species richness of canopy-nesting speciesModel averaging included 3 models
Intercept 2.344 0.053 44.01 <0.001*
NRF 700 m �0.232 0.120 2.115 0.035*
Snags 0.027 0.042 0.646 0.518Large trees �0.023 0.037 0.630 0.529
Species richness of understory-nesting speciesModel averaging included 8 models
Intercept 1.356 0.206 6.589 <0.001*
Forest type (plantation) �0.256 0.136 1.877 0.061NRF 700 m 0.297 0.200 1.484 0.138Large trees 0.091 0.066 1.370 0.171Snags 0.054 0.070 0.778 0.437
Species richness of cavity-nesting speciesModel averaging included 6 models
Intercept 1.214 0.206 5.904 <0.001*
NRF 700 m 0.958 0.348 2.749 0.006*
Snags 0.235 0.113 2.084 0.037*
NRF 700 m: Snags �0.333 0.172 1.932 0.053Forest type (plantations) �0.263 0.195 1.351 0.178Large trees �0.087 0.083 1.049 0.294Snags: Large trees 0.083 0.062 1.345 0.179
* p < 0.05.
116 A. MacKay et al. / Biological Conservation 170 (2014) 110–119
should have been significant predictors of total species richness.Instead, we observed a positive trend for species richness in re-sponse to the proportion of NRF, but neither forest type nor anyinteraction term was significant. Hence, there was no indicationof landscape supplementation with respect to total speciesrichness.
At the nesting guild level, canopy nesters responded negativelyto the proportion of NRF within a 700 m radius. Hence, they
showed no evidence for landscape supplementation. However, inthe case of understory nesters, landscape supplementation couldbe invoked, as forest type may have had an effect on species rich-ness (p = 0.061). Finally, for primary cavity nesters, which tend tohave large home ranges and highly-specialized foraging require-ments, landscape supplementation was likely taking place: theinteraction between the proportion of NRF and the density ofspruce snags (p = 0.053) suggests that plantations may supplement

A. MacKay et al. / Biological Conservation 170 (2014) 110–119 117
resources to NRF, but only when snag density in plantations ishigh. Therefore, allowing plantations adjacent to NRF patches toreach the ‘‘overmature’’ stage could contribute positively to thespecies richness of cavity nesters in the landscape through land-scape supplementation.
The increased occurrence of certain species with the proportionof NRF in the landscape might also reflect source-sink dynamics(Pulliam, 1988). Because we did not follow individuals or monitorreproductive success, we could not test these alternative explana-tions. Nonetheless, inter-annual variations in the occurrence ofRed-breasted Nuthatch (S. canadensis) suggest the possibility ofsuch regional dynamics. In 2010, point count stations within NRFwere saturated (100% occupancy) by this species, whereas planta-tions were occupied at 72%. In 2011, there was a major drop in theregional abundance of this species, but the decline was signifi-cantly stronger in plantations than in NRF, suggesting that theRed-breasted Nuthatch preferred NRF during the breeding season(Appendix A, Table A1). The 2010 season may have represented apopulation eruption following a mast-seeding event (Koenig andKnops, 2000), during which individuals (possibly floaters) spilledover into plantations. Not surprisingly, the model did not convergefor this species. The reproductive success of the Red-breasted Nut-hatch should be quantified in both plantations and NRF to test thisexplanation.
Interpreting the effects of the landscape variable (proportion ofNRF) is complicated by its correlation with the random variable(forest type). Forest type was nonetheless included in the analysisbecause it was used alone or in interaction terms to investigatelandscape supplementation. For stations within plantations, theproportion of NRF in the landscape may increase the likelihoodof finding species that can use plantations but require resourcesfound in NRF. For stations located in NRF, the proportion of NRFwithin 700 m may reflect patch size or patch shape effects. For spe-cies restricted to NRF, elongated patches (lower proportion of NRFin the landscape) require individuals to move more extensively tomeet their foraging requirements and local species richness maybe reduced as a result. Indeed, species richness was consistentlyhigher in stations where the proportion of NRF in the landscapewas greater than 0.6, whereas those located in areas where thisproportion was lower than 0.6 exhibited higher variability andgenerally lower species richness (Fig. 2a and c).
The importance of maintaining naturally-regenerated forestpatches in intensively-managed forests has frequently been con-sidered. In Australia, patches of native Eucalyptus increased avianbiodiversity in adjoining plantations of non-native pine (Linden-
2010
2011
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
NRF Plantations
Fig. A.1. Occurrence of the Red-breasted Nuthatch at stations located in NRF(n = 42) and in 40–50 year-old spruce plantations (n = 60). Occurrence differedbetween years in both NRF and plantations (likelihood ratio test, NRF: G = 6.08,d.f. = 1, p = 0.01 and plantations: G = 29.03, d.f. = 1, p < 0.001).
mayer and Hobbs, 2004). In Spain, patches of naturally-regener-ated forest had a similar importance for the Great SpottedWoodpecker (Dendrocopos major) in a landscape dominated bynon-native pine plantations (Barrientos, 2010). Native forest prob-ably provided its preferred nesting substrates. Here, naturally-regenerated forest had a similar importance for several primarycavity-nesting birds even though our study was conducted in alandscape featuring native planted trees. One of those species,the Black-backed Woodpecker, has been shown to require35 m3 ha�1 of dead wood in the boreal forest (Tremblay et al.,2009). A closely–related species, the Eurasian Three-toed Wood-pecker (Picoides tridactylus), requires at least 15 m3 ha�1
(1.3 m2 ha�1 basal area) (Bütler et al., 2004), and possibly evenmore (Roberge et al., 2008). In the case of the Black-backed Wood-pecker, not only the quantity but also the quality of dead woodmatters because it nests preferentially in large-diameter snags(Setterington et al., 2000) and mainly uses recently-dead treesfor foraging (Tremblay et al., 2010). Hence, the low basal area(average = 0.13 ± 0.26 m2 ha�1) of relatively small-diameter snags(<24 cm dbh) in these plantations does not appear to be sufficientto meet both its nesting and foraging requirements, which may ex-plain the importance of nearby NRF, which offers both a higherquantity (average basal area = 0.9 ± 1.4 m2 ha�1), and higher qual-ity of spruce snags (up to 56 cm dbh). For species such as theBrown Creeper, the positive relationship with NRF in the surround-ings may in fact reflect the higher snag diversity within NRF andnot simply higher snag abundance. The Brown Creeper nests pref-erentially on balsam fir snags (Poulin et al., 2008) which, althoughthey occur at low density within spruce plantations, are notplanted and are much sparser than in NRF (0.2 ± 0.5 m2 ha�1 vs.1.3 ± 1.6 m2 ha�1 respectively).
In this study, 40–50 year-old spruce plantations did not providesuitable habitat to meet the foraging or nesting requirements ofseveral old forest specialists, most likely as a result of a paucityof large-diameter snags. This is cause for concern given the increas-ing rarity of mature and old, naturally-regenerated forest in man-aged landscapes of New Brunswick, Canada (Betts et al., 2007;Erdle and Ward, 2008), or elsewhere in the world. The ecologicalconsequences of the current expansion of plantation silvicultureshould be monitored to ensure the persistence of species such asthe Black-backed Woodpecker and others that are strongly associ-ated with mature, naturally-regenerated conifer forests.
Acknowledgements
We thank Christian Roy for his consultation on R code and sta-tistical analyses. G. Adams, A. Chiasson, M.-H. St-Laurent, and G.Moreau made insightful comments on a previous version, and N.Tran and A. Bulot contributed intensively to data collection. Weare grateful to J.D. Irving Ltd., especially P. Poitras and G. Pelletier,for logistical support, and C. Michaud for her assistance with GIS.This study was funded by a scholarship from the Natural Sciencesand Engineering Research Council of Canada (NSERC) and J.D. Ir-ving Ltd. to AM, and by grants from NSERC and the New BrunswickInnovation Foundation to MAV and D. MacLean.
Appendix A
See Fig. A1 and Tables A1–A3.
Appendix B. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.biocon.2013.12.023. These data include Google maps of the most important areasdescribed in this article.
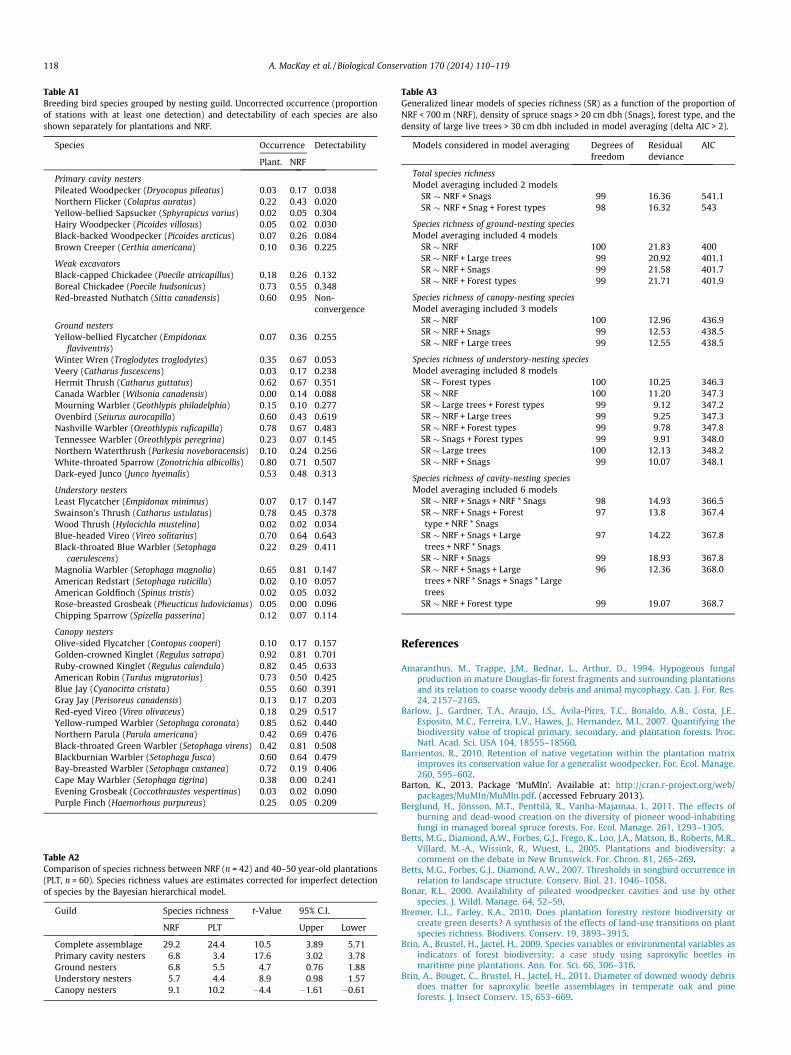
Table A1Breeding bird species grouped by nesting guild. Uncorrected occurrence (proportionof stations with at least one detection) and detectability of each species are alsoshown separately for plantations and NRF.
Species Occurrence Detectability
Plant. NRF
Primary cavity nestersPileated Woodpecker (Dryocopus pileatus) 0.03 0.17 0.038Northern Flicker (Colaptus auratus) 0.22 0.43 0.020Yellow-bellied Sapsucker (Sphyrapicus varius) 0.02 0.05 0.304Hairy Woodpecker (Picoides villosus) 0.05 0.02 0.030Black-backed Woodpecker (Picoides arcticus) 0.07 0.26 0.084Brown Creeper (Certhia americana) 0.10 0.36 0.225
Weak excavatorsBlack-capped Chickadee (Poecile atricapillus) 0.18 0.26 0.132Boreal Chickadee (Poecile hudsonicus) 0.73 0.55 0.348Red-breasted Nuthatch (Sitta canadensis) 0.60 0.95 Non-
convergence
Ground nestersYellow-bellied Flycatcher (Empidonax
flaviventris)0.07 0.36 0.255
Winter Wren (Troglodytes troglodytes) 0.35 0.67 0.053Veery (Catharus fuscescens) 0.03 0.17 0.238Hermit Thrush (Catharus guttatus) 0.62 0.67 0.351Canada Warbler (Wilsonia canadensis) 0.00 0.14 0.088Mourning Warbler (Geothlypis philadelphia) 0.15 0.10 0.277Ovenbird (Seiurus aurocapilla) 0.60 0.43 0.619Nashville Warbler (Oreothlypis ruficapilla) 0.78 0.67 0.483Tennessee Warbler (Oreothlypis peregrina) 0.23 0.07 0.145Northern Waterthrush (Parkesia noveboracensis) 0.10 0.24 0.256White-throated Sparrow (Zonotrichia albicollis) 0.80 0.71 0.507Dark-eyed Junco (Junco hyemalis) 0.53 0.48 0.313
Understory nestersLeast Flycatcher (Empidonax minimus) 0.07 0.17 0.147Swainson’s Thrush (Catharus ustulatus) 0.78 0.45 0.378Wood Thrush (Hylocichla mustelina) 0.02 0.02 0.034Blue-headed Vireo (Vireo solitarius) 0.70 0.64 0.643Black-throated Blue Warbler (Setophaga
caerulescens)0.22 0.29 0.411
Magnolia Warbler (Setophaga magnolia) 0.65 0.81 0.147American Redstart (Setophaga ruticilla) 0.02 0.10 0.057American Goldfinch (Spinus tristis) 0.02 0.05 0.032Rose-breasted Grosbeak (Pheucticus ludovicianus) 0.05 0.00 0.096Chipping Sparrow (Spizella passerina) 0.12 0.07 0.114
Canopy nestersOlive-sided Flycatcher (Contopus cooperi) 0.10 0.17 0.157Golden-crowned Kinglet (Regulus satrapa) 0.92 0.81 0.701Ruby-crowned Kinglet (Regulus calendula) 0.82 0.45 0.633American Robin (Turdus migratorius) 0.73 0.50 0.425Blue Jay (Cyanocitta cristata) 0.55 0.60 0.391Gray Jay (Perisoreus canadensis) 0.13 0.17 0.203Red-eyed Vireo (Vireo olivaceus) 0.18 0.29 0.517Yellow-rumped Warbler (Setophaga coronata) 0.85 0.62 0.440Northern Parula (Parula americana) 0.42 0.69 0.476Black-throated Green Warbler (Setophaga virens) 0.42 0.81 0.508Blackburnian Warbler (Setophaga fusca) 0.60 0.64 0.479Bay-breasted Warbler (Setophaga castanea) 0.72 0.19 0.406Cape May Warbler (Setophaga tigrina) 0.38 0.00 0.241Evening Grosbeak (Coccothraustes vespertinus) 0.03 0.02 0.090Purple Finch (Haemorhous purpureus) 0.25 0.05 0.209
Table A2Comparison of species richness between NRF (n = 42) and 40–50 year-old plantations(PLT, n = 60). Species richness values are estimates corrected for imperfect detectionof species by the Bayesian hierarchical model.
Guild Species richness t-Value 95% C.I.
NRF PLT Upper Lower
Complete assemblage 29.2 24.4 10.5 3.89 5.71Primary cavity nesters 6.8 3.4 17.6 3.02 3.78Ground nesters 6.8 5.5 4.7 0.76 1.88Understory nesters 5.7 4.4 8.9 0.98 1.57Canopy nesters 9.1 10.2 �4.4 �1.61 �0.61
Table A3Generalized linear models of species richness (SR) as a function of the proportion ofNRF < 700 m (NRF), density of spruce snags > 20 cm dbh (Snags), forest type, and thedensity of large live trees > 30 cm dbh included in model averaging (delta AIC > 2).
Models considered in model averaging Degrees offreedom
Residualdeviance
AIC
Total species richnessModel averaging included 2 models
SR � NRF + Snags 99 16.36 541.1SR � NRF + Snag + Forest types 98 16.32 543
Species richness of ground-nesting speciesModel averaging included 4 models
SR � NRF 100 21.83 400SR � NRF + Large trees 99 20.92 401.1SR � NRF + Snags 99 21.58 401.7SR � NRF + Forest types 99 21.71 401.9
Species richness of canopy-nesting speciesModel averaging included 3 models
SR � NRF 100 12.96 436.9SR � NRF + Snags 99 12.53 438.5SR � NRF + Large trees 99 12.55 438.5
Species richness of understory-nesting speciesModel averaging included 8 models
SR � Forest types 100 10.25 346.3SR � NRF 100 11.20 347.3SR � Large trees + Forest types 99 9.12 347.2SR � NRF + Large trees 99 9.25 347.3SR � NRF + Forest types 99 9.78 347.8SR � Snags + Forest types 99 9.91 348.0SR � Large trees 100 12.13 348.2SR � NRF + Snags 99 10.07 348.1
Species richness of cavity-nesting speciesModel averaging included 6 models
SR � NRF + Snags + NRF * Snags 98 14.93 366.5SR � NRF + Snags + Foresttype + NRF * Snags
97 13.8 367.4
SR � NRF + Snags + Largetrees + NRF * Snags
97 14.22 367.8
SR � NRF + Snags 99 18.93 367.8SR � NRF + Snags + Largetrees + NRF * Snags + Snags * Largetrees
96 12.36 368.0
SR � NRF + Forest type 99 19.07 368.7
118 A. MacKay et al. / Biological Conservation 170 (2014) 110–119
References
Amaranthus, M., Trappe, J.M., Bednar, L., Arthur, D., 1994. Hypogeous fungalproduction in mature Douglas-fir forest fragments and surrounding plantationsand its relation to coarse woody debris and animal mycophagy. Can. J. For. Res.24, 2157–2165.
Barlow, J., Gardner, T.A., Araujo, I.S., Ávila-Pires, T.C., Bonaldo, A.B., Costa, J.E.,Esposito, M.C., Ferreira, L.V., Hawes, J., Hernandez, M.I., 2007. Quantifying thebiodiversity value of tropical primary, secondary, and plantation forests. Proc.Natl. Acad. Sci. USA 104, 18555–18560.
Barrientos, R., 2010. Retention of native vegetation within the plantation matriximproves its conservation value for a generalist woodpecker. For. Ecol. Manage.260, 595–602.
Barton, K., 2013. Package ‘MuMIn’. Available at: http://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf. (accessed February 2013).
Berglund, H., Jönsson, M.T., Penttilä, R., Vanha-Majamaa, I., 2011. The effects ofburning and dead-wood creation on the diversity of pioneer wood-inhabitingfungi in managed boreal spruce forests. For. Ecol. Manage. 261, 1293–1305.
Betts, M.G., Diamond, A.W., Forbes, G.J., Frego, K., Loo, J.A., Matson, B., Roberts, M.R.,Villard, M.-A., Wissink, R., Wuest, L., 2005. Plantations and biodiversity: acomment on the debate in New Brunswick. For. Chron. 81, 265–269.
Betts, M.G., Forbes, G.J., Diamond, A.W., 2007. Thresholds in songbird occurrence inrelation to landscape structure. Conserv. Biol. 21, 1046–1058.
Bonar, R.L., 2000. Availability of pileated woodpecker cavities and use by otherspecies. J. Wildl. Manage. 64, 52–59.
Bremer, L.L., Farley, K.A., 2010. Does plantation forestry restore biodiversity orcreate green deserts? A synthesis of the effects of land-use transitions on plantspecies richness. Biodivers. Conserv. 19, 3893–3915.
Brin, A., Brustel, H., Jactel, H., 2009. Species variables or environmental variables asindicators of forest biodiversity: a case study using saproxylic beetles inmaritime pine plantations. Ann. For. Sci. 66, 306–316.
Brin, A., Bouget, C., Brustel, H., Jactel, H., 2011. Diameter of downed woody debrisdoes matter for saproxylic beetle assemblages in temperate oak and pineforests. J. Insect Conserv. 15, 653–669.

A. MacKay et al. / Biological Conservation 170 (2014) 110–119 119
Brockerhoff, E.G., Jactel, H., Parrotta, J.A., Quine, C.P., Sayer, J., 2008. Plantationforests and biodiversity: oxymoron or opportunity? Biodivers. Conserv. 17,925–951.
Brooks, S.P., Gelman, A., 1998. General methods for monitoring convergence ofiterative simulations. J. Comp. Graph. Stat. 7, 434–455.
Bütler, R., Angelstam, P., Ekelund, P., Schlaepfer, R., 2004. Dead wood thresholdvalues for the three-toed woodpecker presence in boreal and sub-Alpine forest.Biol. Conserv. 119, 305–318.
Canterbury, G.E., Martin, T.E., Petit, D.R., Petit, L.J., Bradford, D.F., 2001. Birdcommunities and habitat as ecological indicators of forest condition in regionalmonitoring. Conserv. Biol. 14, 544–558.
Carignan, V., Villard, M.-A., 2002. Selecting indicator species to monitor ecologicalintegrity: a review. Environ. Monit. Assess. 78, 45–61.
Carnus, J.-M., Parrotta, J., Brockerhoff, E., Arbez, M., Jactel, H., Kremer, A., Lamb, D.,OHara, K., Walters, B., 2006. Planted forests and biodiversity. J. Forest. 104, 65–77.
Drever, M.C., Aitken, K.E., Norris, A.R., Martin, K., 2008. Woodpeckers as reliableindicators of bird richness, forest health and harvest. Biol. Conserv. 141, 624–634.
Du Bus de Warnaffe, G., Deconchat, M., 2008. Impact of four silvicultural systems onbirds in the Belgian Ardenne: implications for plantation management.Biodivers. Conserv. 17, 1041–1055.
Dunning, J.B., Danielson, B.J., Pulliam, H.R., 1992. Ecological processes that affectpopulations in complex landscapes. Oikos 65, 169–175.
Erdle, T., Ward, C., 2008. Management Alternatives for New Brunswick’sPublic Forest: Report of the New Brunswick Task Force on Forest Diversityand Wood Supply. New Brunswick Department of Natural Resources,Fredericton.
Estades, C.F., Temple, S.A., 1999. Deciduous-forest bird communities in afragmented landscape dominated by exotic pine plantations. Ecol. Appl. 9,573–585.
Etheridge, D.A., MacLean, D.A., Wagner, R.G., Wilson, J.S., 2005. Changes inlandscape composition and stand structure from 1945 2002 on an industrialforest in New Brunswick, Canada. Can. J. For. Res. 35, 1965–1977.
FAO, 2010. Global Forest Resources Assessment 2010: Main Report. Organization ofthe United Nations Forestry Department. Food and Agriculture Organization ofthe United Nations, Rome.
Farwig, N., Sajita, N., Böhning-Gaese, K., 2008. Conservation value of forestplantations for bird communities in western Kenya. For. Ecol. Manage. 255,3885–3892.
Fischer, J., Lindenmayer, D.B., 2006. Beyond fragmentation: the continuum modelfor fauna research and conservation in human-modified landscapes. Oikos 112,473–480.
Gjerde, I., Sætersdal, M., Nilsen, T., 2005. Abundance of two threatened woodpeckerspecies in relation to the proportion of spruce plantations in native pine forestsof western Norway. Biodivers. Conserv. 14, 377–393.
Hartley, M.J., 2002. Rationale and methods for conserving biodiversity in plantationforests. For. Ecol. Manage. 155, 81–95.
Haskell, D.G., Evans, J.P., Pelkey, N.W., 2006. Depauperate avifauna in plantationscompared to forests and exurban areas. PLoS One 1 (1), e63. http://dx.doi.org/10.1371/journal.pone.0000063.
Hoffmann, J.P., 2003. Generalized Linear Models. Allyn & Bacon, Boston.Hollander, F.A., Titeux, N., Van Dyck, H., 2012. Territorial resource valuation
deviates from habitat quality in an ecologically trapped, long-distancemigratory bird. Behav. Ecol. Sociobiol. 66, 777–783.
Humphrey, J.W., Davey, S., Peace, A.J., Ferris, R., Harding, K., 2002. Lichens andbryophyte communities of planted and semi-natural forests in Britain: theinfluence of site type, stand structure and deadwood. Biol. Conserv. 107, 165–180.
Johnson, G.A., Freedman, B., 2003. Breeding birds in forestry plantations and naturalforest in the vicinity of Fundy National Park, New Brunswick. Can. Field Nat.116, 475–487.
Kirby, K.J., Reid, C.M., Thomas, R.C., Goldsmith, F.B., 1998. Preliminary estimates offallen dead wood and standing dead trees in managed and unmanaged forestsin Britain. J. Appl. Ecol. 35, 148–155.
Koenig, W.D., Knops, J.M., 2000. Patterns of annual seed production by northernhemisphere trees: a global perspective. Am. Nat. 155, 59–69.
Lindenmayer, D.B., 1999. Future directions for biodiversity conservation inmanaged forests: indicator species, impact studies and monitoring programs.For. Ecol. Manage. 115, 277–287.
Lindenmayer, D.B., Hobbs, R.J., 2004. Fauna conservation in Australian plantationforests – a review. Biol. Conserv. 119, 151–168.
MacKenzie, D.I., Nichols, J.D., Lachman, G.B., Droege, S., Royle, J.A., Langtimm, C.A.,2002. Estimating site occupancy rates when detection probabilities are lessthan one. Ecology 83, 2248–2255.
MacKenzie, D.I., Nichols, J.D., Sutton, N., Kawanishi, K., Bailey, L.L., 2005. Improvinginferences in population studies of rare species that are detected imperfectly.Ecology 86, 1101–1113.
MacLean, D.A., Amos-Binks, L., Adams, G., Pelletier, G., Villard, M.-A., 2010.Legacy of the Sustainable Forest Management Network: Outcomes ofResearch Collaborations among J.D. Irving, Limited, University of NewBrunswick, and Université de Moncton. Sustainable Forest ManagementNetwork, Edmonton.
Martin, K., Aitken, K.E., Wiebe, K.L., 2004. Nest sites and nest webs for cavity-nestingcommunities in interior British Columbia, Canada: nest characteristics andniche partitioning. Condor 106, 5–19.
McCay, T.S., Komoroski, M.J., 2004. Demographic responses of shrews to removal ofcoarse woody debris in a managed pine forest. For. Ecol. Manage. 189, 387–395.
Mikusinski, G., Gromadzki, M., Chylarecki, P., 2001. Woodpeckers as indicators offorest bird diversity. Conserv. Biol. 15, 208–217.
Montigny, M.K., MacLean, D.A., 2005. Using heterogeneity and representation ofecosite criteria to select forest reserves in an intensively managed industrialforest. Biol. Conserv. 125, 237–248.
New Brunswick Department of Natural Resources, 2005. Habitat Definitions forOld-forest Vertebrates in New Brunswick. Government of New Brunswick,Fredericton.
New Brunswick Department of Natural Resources, 2009. Be... Sustainable in thisPlace. A Balanced Approach for New Brunswick’s Crown Forest. Government ofNew Brunswick, Fredericton.
New Brunswick Department of Natural Resources, 2013. Our Landscape Heritage.Central Uplands. Government of New Brunswick, Fredericton, 359 pp. <http://www2.gnb.ca/content/dam/gnb/Departments/nr-rn/pdf/en/CrownLandsForests/ProtectedNaturalAreas/OurLandscapeHeritage/Chapter09-e.pdf> (Chapter 9).
Paquette, A., Messier, C., 2011. The effect of biodiversity on tree productivity: fromtemperate to boreal forests. Global Ecol. Biogeogr. 20, 170–180.
Paritsis, J., Aizen, M.A., 2008. Effects of exotic conifer plantations on the biodiversityof understory plants, epigeal beetles and birds in Nothofagus dombeyi forests.For. Ecol. Manage. 255, 1575–1583.
Parker, G.R., Kimball, D.G., Dalzell, B., 1994. Bird communities breeding in selectedspruce and pine plantations in New Brunswick. Can. Field Nat. 108, 1–9.
Pietrek, A.G., Branch, L.Y.N.C., 2011. Native plantations as an important element forbiodiversity in vanishing forested landscapes: a study of the near threatenedaraucaria tit spinetail (Leptasthenura setaria, Furnariidae). Austral Ecol. 36, 109–116.
Plummer, M., 2003. JAGS: A Program for Analysis of Bayesian Graphical Modelsusing Gibbs Sampling. <http://citeseer.ist.psu.edu/plummer03jags.htm>.
Porter, K.B., Hemens, B., MacLean, D.A., 2004. Using insect-caused disturbancepatterns in northern New Brunswick to inform forest management. In: Perera,A.H., Buse, L.J., Weber, M.G. (Eds.), Emulating Natural Forest LandscapeDisturbances. Colombia University Press, New York, pp. 135–145.
Poulin, J.-F., Villard, M.-A., Edman, M., Goulet, P.J., Eriksson, A.-M., 2008. Thresholdsin nesting habitat requirements of an old forest specialist, the Brown Creeper(Certhia americana), as conservation targets. Biol. Conserv. 141, 1129–1137.
Poulin, J.-F., D’Astous, E., Villard, M.-A., Hejl, S.J., Newlon, K.R., McFadzen, M.E.,Young, J.S., Ghalambor, C.K., 2013. Brown creeper (Certhia americana). In: Poole,A. (Ed.), The Birds of North America Online. Cornell Lab of Ornithology, Ithaca.<http://bna.birds.cornell.edu/bna/species/669doi:10.2173/bna.669>.
Pulliam, H.R., 1988. Sources, sinks, and population regulation. Am. Nat. 132, 652–661.
Remeš, V., 2003. Effects of exotic habitat on nesting success, territory density, andsettlement patterns in the Blackcap (Sylvia atricapilla). Conserv. Biol. 17, 1127–1133.
Roberge, J.-M., Angelstam, P., 2006. Indicator species among resident forest birds – across-regional evaluation in northern Europe. Biol. Conserv. 130, 134–147.
Roberge, J.-M., Angelstam, P., Villard, M.-A., 2008. Specialized woodpeckers andnaturalness in hemiboreal forests – deriving quantitative targets forconservation planning. Biol. Conserv. 141, 997–1012.
Setterington, M.A., Thompson, I.D., Montevecchi, W.A., 2000. Woodpeckerabundance and habitat use in mature balsam fir forests in Newfoundland. J.Wildl. Manage. 64, 335–345.
Stephens, S., Wagner, M.R., 2007. Forest plantations and biodiversity: a freshperspective. J. Forest. 105, 307–313.
St-Laurent, M.-H., Ferron, J., Hins, C., Gagnon, R., 2007. Effects of stand structure andlandscape characteristics on habitat use by birds and small mammals inmanaged boreal forest of eastern Canada. Can. J. For. Res. 37, 1298–1309.
Thinh, V.U.T., Doherty, P.F., Huyvaert, K.P., 2012. Avian conservation value of pineplantation forests in northern Vietnam. Bird Conserv. Int. 22, 193–204.
Tremblay, J.A., Ibarzabal, J., Dussault, C., Savard, J.-P.L., 2009. Habitat requirementsof breeding black-backed Woodpeckers (Picoides arcticus) in managed,unburned boreal forest. Avian Conserv. Ecol. 4 (1), 2, <http://www.ace-eco.org/vol4/iss1/art2/>.
Tremblay, J.A., Ibarzabal, J., Savard, J.-P.L., 2010. Foraging ecology of black-backedwoodpeckers (Picoides arcticus) in unburned eastern boreal forest stands. Can. J.For. Res. 40, 991–999.
Villard, M.-A., Haché, S., 2012. Conifer plantations consistently act as barriers tomovement in a deciduous forest songbird: a translocation experiment. Biol.Conserv. 155, 33–37.
Virkkala, R., 2006. Why study woodpeckers? The significance of woodpeckers inforest ecosystems. Ann. Zool. Fenn. 43, 82–85.
Waldick, R.C., Freedman, B., Wassersug, R.J., 1999. The consequences for amphibiansof the conversion of natural, mixed-species forests to conifer plantations insouthern New Brunswick. Can. Field Nat. 113, 408–418.
Zipkin, E.F., Royle, J.A., Dawson, D.K., Bates, S., 2010. Multi-species occurrencemodels to evaluate the effects of conservation and management actions. Biol.Conserv. 143, 479–484.





























