Embed Size (px)
Citation preview

abundance of parasitic spiders of other species at high-resource sites.
This spider system bears many similarities to colonies of social insects. Group selection, which in these species translates to colony-level selection and in some cases involves direct warfare between colonies, is thought to shape social insect traits6. Like social spider colonies, social insect colonies are com-posed of close kin, which probably explains why group selection has seemingly had such strong evolutionary impacts. Although actu-ally quantifying selection on colony-level traits of social insects has been elusive7, the social mechanisms that regulate some such traits, referred to as social physiology, are well studied in social insects and are understood to be colony-level adaptations8 that are shaped by colony-level selection9.
This highlights a key remaining challenge in understanding the social spider system. The precise social and developmental mecha-nisms that enable spider colonies to adaptively adjust their colony composition must be char-acterized. Pruitt and Goodnight suggest that within-colony conflict or the collective ces-sation of reproduction are involved, which emphasizes another crucial issue: although
the authors present a compelling case for a strong evolutionary role of group selection, it is important to determine whether individual selection also operates within colonies. That is, does the survival and reproduction of indi-viduals within groups depend on whether the individual is aggressive or docile?
Another line of future research will be to study the genetic basis of group traits. Adapta-tion can occur only when selection acts on her-itable traits, so researchers need to understand the nature of the genetic architecture of specific traits together with patterns of selection on these traits. Some of the most persuasive evi-dence for the potential evolutionary strength of group selection comes from animal-breeding studies demonstrating that artificial selection at the group level can increase traits associated with productivity, even when these traits have long been the target of artificial individual-level selection10. The intuitive explanation for this is that an unintended side effect of individual-level selection for productivity is the evolution of highly aggressive individuals that monopolize resources to the detriment of group members. By contrast, group selection acts more efficiently on heritable traits under-lying positive social interactions, leading to
the evolution of groups of amiable animals with high overall reproductive success 11. The ability of group selection to more effectively act on heritable social traits may also underlie the seemingly strong evolutionary influence of group selection in Pruitt and Goodnight’s study and earlier studies. ■
Timothy Linksvayer is in the Department of Biology, University of Pennsylvania, Philadelphia, Pennsylvania 19104, USA.e-mail: [email protected]
1. Okasha, S. Evolution and the Levels of Selection (Oxford Univ. Press, 2006).
2. Pruitt, J. N. & Goodnight, C. J. Nature 514, 359–362 (2014).
3. Hamilton, W. D. J. Theoret. Biol. 7, 1–16 (1964).4. Wade, M. J. Science 210, 665–667 (1980).5. Lehmann, L. et al. Proc. Natl Acad. Sci. USA 104,
6736–6739 (2007).6. Hölldobler, B. & Wilson, E. O. The Superorganism:
The Beauty, Elegance, and Strangeness of Insect Societies (Norton, 2009).
7. Gordon, D. M. Nature 498, 91–93 (2013).8. Seeley, T. D. Am. Nat. 150, S22–S41 (1997).9. Linksvayer, T. A., Fondrk, M. K. & Page, R. E. Jr Am.
Nat. 173, E99–E107 (2009).10. Wade, M. J. et al. Evol. Appl. 3, 453–465 (2010).11. Bijma, P., Muir, W. A. & Van Arendonk, J. A. M.
Genetics 175, 277–288 (2007).
This article was published online on 1 October 2014.
A L E S S I A B O T T O S & N A N C Y E . H Y N E S
A key goal in cancer research is to under-stand the mechanisms underlying the metastatic process, by which cancer
cells spread from a primary tumour to other sites and form secondary tumours. Patients with breast cancer, for example, usually do not die from their primary tumours, which are surgically removed, but from metastatic tumours in organs such as the bone, liver or lung. Writing in Cell, Aceto et al.1 provide information about this complex pathway. They find that the cells that escape from primary tumours sometimes do so as clusters, and that these clusters have a higher ability to form lung metastases than single escaped cells, despite being present at much lower levels in the blood. The authors show that this difference stems from the expression of a cell-adhesion protein that allows the cells to stick together in clusters, thereby providing
a survival advantage in the lungs.Blood-borne circulating tumour cells
(CTCs) that have broken away from primary tumours were described more than 30 years ago, but technological limitations have until recently made it challenging to study them. With the advent of improved methods to detect, quantify and isolate CTCs2, we now know that these cells have tumour-forming ability3,4 and that CTC numbers have prognos-tic significance in many types of cancer (see ref. 5 for a review).
Considering that there are many more CTCs in the blood than there are metastatic tumours, it is of interest to discover the characteristics of a successful circulating cell. By mixing tumour cells labelled with different colours, Aceto and colleagues generated multi coloured primary breast tumours in mice, which allowed them to visualize multi coloured CTC clusters in the blood and lungs. Quantification revealed that even though cell clusters made up less than
3% of total CTC ‘events’ in the blood, more than 50% of the metastases were derived from clusters. The authors also found that CTC clusters represent aggregates of cells origina ting from the tumour and entering the vasculature, rather than tumour cells that clump together in the circulation to form clusters.
To strengthen their conclusions, the authors compared the ability of single CTCs and CTC clusters to generate lung metastases directly. Following injection into the tail vein of mice, cells from both populations efficiently reached the lungs, but the single CTCs underwent high levels of apoptotic cell death, whereas the CTC clusters survived much better and thus formed metastases more often.
The results from these experimental models are exciting, but do they hold up in patients with cancer? To look at the prognostic signifi-cance of CTC clusters, the researchers moni-tored the blood of patients with advanced metastatic cancer. CTCs were found in 68% of the patients; among these, clusters were con-tinuously detected in 5.6% of the cases. Despite the rarity of the clusters, their continued pres-ence was correlated with a significantly shorter period without disease progression in patients with breast cancer and with reduced overall survival in patients with prostate cancer.
To identify the molecular mechanisms governing CTC-cluster formation, Aceto and colleagues used devices called negCTC-iChips6 to isolate single CTCs and CTC clusters from the blood of patients with breast cancer, and then sequenced RNA transcripts from the cells. Although there were no large gene-expression
C A N C E R
Staying together on the road to metastasisMost deaths from breast cancer occur when the primary tumour spreads to secondary sites. It now emerges that clusters of tumour cells that enter the bloodstream form metastases more often than single circulating tumour cells.
1 6 O C T O B E R 2 0 1 4 | V O L 5 1 4 | N A T U R E | 3 0 9
NEWS & VIEWS RESEARCH
© 2014 Macmillan Publishers Limited. All rights reserved
differences in the cells originating from the two populations, expression of some cluster-associated genes, including that encoding the protein plakoglobin, was increased in cells from clusters.
Plakoglobin was discovered more than 30 years ago as an adhesion protein associ-ated with the cell membrane. Although it has previously been implicated in cancer, its role is controversial (see ref. 7 for a review). For example, one study proposed plakoglobin as a tumour-suppressor protein, whose expres-sion is downregulated by epigenetic (non-DNA-sequence-changing) modulation in some tumours8, whereas another suggested that it has a tumour-inducing effect9. It is possible that plakoglobin has different roles in different cancer types, but from Aceto and col-leagues’ results it seems clear that the protein acts as a metastasis promoter in breast cancer. Indeed, in an in silico analysis of publicly available data sets from almost 2,000 patients, the authors correlated high plako globin levels with a significant reduction in metastasis-free survival time.
Returning to mouse studies, the researchers found that reducing plakoglobin expression in breast-cancer cells resulted in the destruction of CTC clusters and reduced their metastatic ability. These data suggest that plakoglobin expression in tumour cells is responsible for the formation of CTC clusters and that plakoglobin-mediated clustering provides these cells with a survival advantage when they reach the lungs (Fig. 1).
But how does plakoglobin work? Is its effect mediated by adhesion activity and/or by controlling signalling pathways in the cell? Plakoglobin is a homologue of the protein β-catenin, and competes with it for binding
to E-cadherin (another adhesion protein) and transcription factors of the TCF family. These proteins are involved in the Wnt signalling pathway, which is abnormally active in several types of breast cancer. Perhaps plakoglobin influences signalling by antago-nizing β-catenin and thereby affects Wnt signalling in breast-cancer cells. This and other hypotheses will need to be investigated.
It will also be of interest to assess whether plakoglobin is a drug-targetable molecule in metastases. Reduction of plakoglobin expres-sion did not affect the dissemination of single CTCs to the lungs in Aceto and colleagues’ mouse studies, and these cells, although they have less metastatic power than CTC
clusters, can still colonize distant organs. Thus, identifying molecular targets that are common to all CTC populations remains a goal for future studies.
Although several questions about CTCs are still open, the efforts currently being expended on characterizing these cells are motivated by their clear clinical relevance10. Thanks to the feasibility of blood sampling and the fact that CTCs can be detected even in the early stages of primary cancer, these cells could be excel-lent biomarkers for early diagnosis. CTCs and CTC clusters could also be quantified to moni-tor treatment responses and the progression of metastatic disease. Moreover, considering the genetic differences reported between primary tumours and metastases11, choosing effective therapies to treat metastases might be better informed by analysing CTCs, the cells capable of disseminating to distant organs, rather than by analysing primary tumour cells. ■
Alessia Bottos and Nancy E. Hynes are at the Friedrich Miescher Institute for Biomedical Research, 4058 Basel, Switzerland.e-mails: [email protected]; [email protected]
1. Aceto, N. et al. Cell 158, 1110–1122 (2014).2. Yu, M., Stott, S., Toner, M., Maheswaran, S. &
Haber, D. A. J. Cell Biol. 192, 373–382 (2011).3. Baccelli, I. et al. Nature Biotechnol. 31, 539–544
(2013). 4. Hodgkinson, C. L. et al. Nature Med. 20, 897–903
(2014).5. Alix-Panabières, C. & Pantel, K. Nature Rev. Cancer
14, 623–631 (2014).6. Ozkumur, E. et al. Sci. Transl. Med. 5, 179ra47 (2013).7. Aktary, Z. & Pasdar, M. Int. J. Cell Biol. 2012,
189521 (2012).8. Shiina, H. et al. Cancer Res. 65, 2130–2138 (2005). 9. Hakimelahi, S. et al. J. Biol. Chem. 275,
10905–10911 (2000).10. Krebs, M. G. et al. Nature Rev. Clin. Oncol. 11,
129–144 (2014).11. Shah, S. P. et al. Nature 461, 809–813 (2009).
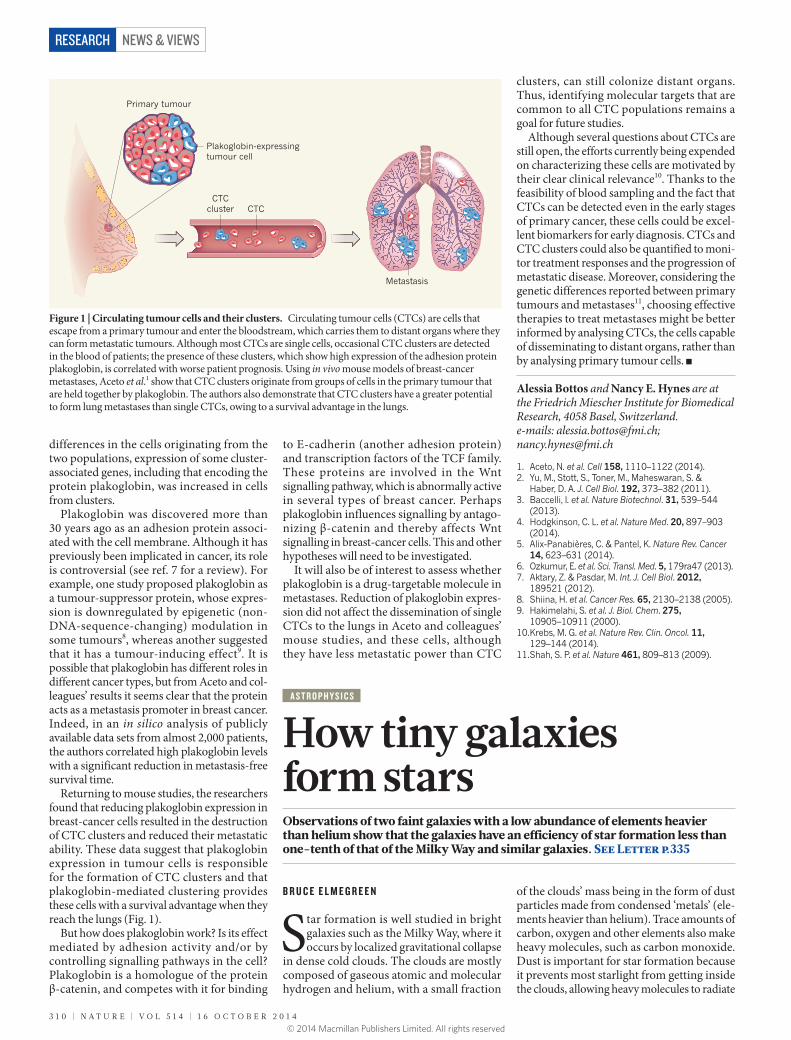
Figure 1 | Circulating tumour cells and their clusters. Circulating tumour cells (CTCs) are cells that escape from a primary tumour and enter the bloodstream, which carries them to distant organs where they can form metastatic tumours. Although most CTCs are single cells, occasional CTC clusters are detected in the blood of patients; the presence of these clusters, which show high expression of the adhesion protein plakoglobin, is correlated with worse patient prognosis. Using in vivo mouse models of breast-cancer metastases, Aceto et al.1 show that CTC clusters originate from groups of cells in the primary tumour that are held together by plakoglobin. The authors also demonstrate that CTC clusters have a greater potential to form lung metastases than single CTCs, owing to a survival advantage in the lungs.
Primary tumour
Plakoglobin-expressingtumour cell
Metastasis
CTCcluster CTC
A S T R O P H Y S I C S
How tiny galaxies form starsObservations of two faint galaxies with a low abundance of elements heavier than helium show that the galaxies have an efficiency of star formation less than one-tenth of that of the Milky Way and similar galaxies. See Letter p.335
B R U C E E L M E G R E E N
Star formation is well studied in bright galaxies such as the Milky Way, where it occurs by localized gravitational collapse
in dense cold clouds. The clouds are mostly composed of gaseous atomic and molecular hydrogen and helium, with a small fraction
of the clouds’ mass being in the form of dust particles made from condensed ‘metals’ (ele-ments heavier than helium). Trace amounts of carbon, oxygen and other elements also make heavy molecules, such as carbon monoxide. Dust is important for star formation because it prevents most starlight from getting inside the clouds, allowing heavy molecules to radiate
3 1 0 | N A T U R E | V O L 5 1 4 | 1 6 O C T O B E R 2 0 1 4
NEWS & VIEWSRESEARCH
© 2014 Macmillan Publishers Limited. All rights reserved