Embed Size (px)
Citation preview

Molecular and Cellular Pathobiology
Cancer Angiogenesis Induced by Kaposi Sarcoma–Associated Herpesvirus Is Mediated by EZH2
Meilan He1,4, Wei Zhang4, Thomas Bakken1, Melissa Schutten3, Zsolt Toth4, Jae U. Jung4,Parkash Gill5, Mark Cannon1,2, and Shou-Jiang Gao4
AbstractEZH2 is a component of the epigenetic regulator PRC2 that suppresses gene expression. Elevated expression of
EZH2 is common in human cancers and is associated with tumor progression and poor prognosis. In this study,we show that EZH2 elevation is associated with epigenetic modifications of Kaposi sarcoma–associatedherpesvirus (KSHV), an oncogenic virus that promotes the development of Kaposi sarcoma and othermalignancies that occur in patients with chronic HIV infections. KSHV induction of EZH2 expression wasessential for KSHV-induced angiogenesis. High expression of EZH2 was observed in Kaposi sarcoma tumors. Incell culture, latent KSHV infection upregulated the expression of EZH2 in human endothelial cells through theexpression of vFLIP and LANA, two KSHV-latent genes that activate the NF-kB pathway. KSHV-mediatedupregulation of EZH2 was required for the induction of Ephrin-B2, an essential proangiogenic factor that drivesendothelial cell tubule formation. Taken together, our findings indicate that KSHV regulates the host epigeneticmodifier EZH2 to promote angiogenesis. Cancer Res; 72(14); 3582–92. �2012 AACR.
IntroductionKaposi sarcoma–associated herpesvirus (KSHV) is the eti-
ologic agent of Kaposi's sarcoma, the most common cancer inHIV-infected subjects. Kaposi sarcoma is a multifocal neovas-cular tumor of proliferating spindle endothelial cells latentlyinfected by KSHV (1). In cell culture, KSHV infection convertshuman primary endothelial cells into spindle shapes withaltered expression of cell-surface markers and a distinct cel-lular gene expression program (2–6). Extensive studies haveidentified a number of cellular factors that mediate KSHV-induced malignant cellular proliferation (1).
Kaposi sarcoma is a highly vascularized and angiogenictumor. Indeed, KSHV infection promotes the angiogenic phe-notypes of endothelial cells, which include induction of tubuleformation on Matrigel, capillary morphogenesis in low growthfactor conditions, and enhanced migration, invasion, and
permeability (7–11). Several proangiogenic factors are inducedfollowing KSHV infection of human endothelial cells. Theseinclude VEGF-A and VEGF-C, angiopoietin-2, interleukin (IL)-6, IL-8, Ephrin-B, matrix metalloproteinase (MMP)-1, MMP-2,and MMP-9 (7, 8, 12–14). Upregulation of these proangiogeniccytokines is, in part, mediated by multiple mitogen-activatedprotein kinase pathways, Notch pathway, and NF-kB pathway(7, 8, 12, 13, 15).
Enhancer of Zeste Homolog 2 (EZH2) is a main componentof polycomb repressive complex 2 (PRC2), which regulateschromatin structure and gene expression through posttrans-lational modifications of histone (16). EZH2 has attractedintense interest because of its role inmaintaining cell stemnessand development of cancer (16). When the PRC2 complex isrecruited to chromatin, EZH2 catalyses the trimethylation ofthe lysine 27 of histone H3 (H3K27me3), which leads tosubsequent recruitment of the PRC1 complex that monoubi-quitylates the lysine 119 of histone H2A (H2AK119ub1) toprevent RNA polymerase II–dependent transcriptional elon-gation leading to transcriptional repression (16, 17). Severalreports suggest that EZH2 directly interacts with DNAmethyl-transferases (DNMT1, DNMT3A, andDNMT3B) and that EZH2is necessary for the maintenance of DNA methylation andrepression of specific genes, including tumor-suppressor genes(16, 17). EZH2 is highly expressed in numerous human cancers,including breast and prostate, with high EZH2 levels oftencorrelating with advanced stages of tumor progression andpoor prognosis (18, 19). EZH2 connects the estrogen and Wntsignaling pathways by directly interacting with estrogen recep-tor a and b-catenin, resulting in transcriptional upregulationof c-myc and cyclin D1 in breast cancer cells (20). Moreover,mutation of EZH2 Y641, which results in gain-of-function, hasbeen described in lymphomas (21). Recent studies have shownthat both VEGF and FGF2 induce angiogenesis through
Authors' Affiliations: Departments of 1Medicine and 2Microbiology, and3Veterinary Population Medicine, The University of Minnesota, Minneapo-lis, Minnesota; and Departments of 4Molecular Microbiology and Immu-nology, and 5Medicine, University of Southern California Keck School ofMedicine, Los Angeles, California
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Current address for M. Schutten: Genentech, Inc., 1 DNA Way, South SanFrancisco, CA 94114.
Corresponding Authors: Shou-Jiang Gao, Department of MolecularMicrobiology and Immunology, University of Southern California KeckSchool of Medicine, 1450 Biggy Street, Los Angeles, CA 90033. E-mail:[email protected]; Meilan He, Phone: 323-442-8058; Fax: 323-442-7868;E-mail: [email protected]; andMark Cannon, Department ofMedicine,The University of Minnesota, Minneapolis, MN. E-mail:[email protected]
doi: 10.1158/0008-5472.CAN-11-2876
�2012 American Association for Cancer Research.
CancerResearch
Cancer Res; 72(14) July 15, 20123582
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

upregulation of EZH2 expression, and inhibition of EZH2attenuated the cell proliferation, migration, and tubule forma-tion of glioblastoma cells (18, 22–24).Because of important biologic roles of EZH2 in tumorigen-
esis and angiogenesis and the results of recent studies thathave shown the connection of EZH2 with the epigeneticmodifications of the KSHV genome (25, 26), we sought toinvestigate how KSHV infection might regulate EZH2 andwhether such regulation might contribute to KSHV-inducedangiogenesis. In this work, we have found that EZH2 is highlyexpressed in Kaposi sarcoma tumors, and KSHV infection ofendothelial cells upregulates the expression of EZH2. Further-more, KSHV upregulation of EZH2 is partially mediated by theNF-kB pathway and the viral latent genes vFLIP (ORF71) andLANA (ORF73). Finally, we have shown that EZH2 mediatesKSHV-induced endothelial cell tubule formation on Matrigelby regulating the expression of proangiogenic gene Ephrin-B2.
Materials and MethodsCell lines and reagentsUninfected and KSHV-infected SLK cells, named SLKn and
SLKp, respectively, were provided by Dr. Adam Grundhoffat Heinrich Dr. Adam Grundhoff 's location is Heinrich-Pette-Institute for Experimental Virology and Immunology,Hamburg Germany Pette Institute, Hamburg, Germany, inSeptember, 2010, and cultured as described (27). The statusof KSHV infection was verified in this study (Fig. 2C). Severalbatches of primary human blood outgrowth endothelial cells(BOEC)were obtained fromDr. RobertHebbel at theUniversityof Minnesota, Minneapolis, MN, between 2009 and 2010 andmaintained in EGM-2 plus Bullet Kit (Lonza) on collagen-coated plates or dishes as previously described (28). BOECs canbe passaged in vitro for more than 30 passages while main-taining their phenotypes. We used cells at passage 7 to 8 for theexperiments.NF-kB super suppressor IkBa mutant (IkBaM) lentiviral
construct (p156RRLsinPPTCMV-IkBaM) was provided by Dr.Inder Verma at Salk Institute, San Diego, CA. vFLIP lentiviralconstruct was obtained from Dr. Chris Boshoff at UCL CancerInstitute, London, UK. EZH2 promoter firefly luciferase report-er construct, which contains a DNA fragment from �1,093 bptoþ48 bp of the transcriptional start site, was provided by Dr.Felix Hoppe-Seyler at German Cancer Research Center, Hei-delberg, Germany. The thymidine kinase promoter Renillaluciferase reporter plasmid (pRL-TK) was used as a controlfor transfection efficiency (Promega). Lentivirus cloneTRCN000040076 for one of the EZH2 short hairpin RNA(shRNA) constructs was obtained from Open Biosystems. Asecond lentiviral EZH2 shRNA construct was previouslydescribed (26). Lentivirus production and infection were car-ried out as previously described (29). Ephrin-B2 shRNA lenti-viral particles containing 3 different target-specific constructswere from Santa Cruz Biotechnology (sc-39438-V).
KSHV virus production and cell infectionRecombinant KSHV-GFP was isolated as previously
described (27). Isolated virus was purified by centrifugation
at 24,000 rpm for 2 hours with a 20/35% Nycodenz gradient(Thermo Fisher Scientific). The gradient junction band con-tainingKSHVwas collected. The purified virus preparationwasaliquoted and stored at �80�C until use. The purified KSHVwas diluted inOpti-MEM I Reduced SerumMediumbefore use.For KSHV infection, BOECswere plated on collagen-coated 12-well plate at 3� 104 cells per well overnight andmock infectedor infected with KSHV at more than 80% infection rates basedon the percentages GFP-positive cells at 2 days postinfection.The concentration of infectious viral particles was determinedbefore infection as published (3). To enhance infection effi-ciency, plates were centrifuged at 3,000 rpm for 1 hour at 25�Cimmediately after addition of KSHV to cells. At 3 hourspostinfection, cells were washed 3 times with PBS and culturedin full EGM-2 medium.
Immunofluorescence and immunohistochemistryImmunofluorescence was done as previously described (29).
An anti-LANA ratmonoclonal antibody (AdvancedBiotechnol-ogies Inc) and a Rhodamine Red-X–conjugated AffinipureDonkey Anti-Rat secondary antibody (Jackson ImmunoRe-search Laboratories, Inc.,) were used for LANA staining. Immu-nohistochemistry for EZH2was carried out on a formalin-fixedparaffin-embedded tissue microarray obtained from the AIDSand Cancer Specimen Resource (ACSR) of the National CancerInstitute. The tissue microarray slide was subjected to citrateantigen retrieval for 30 minutes and blocked for nonspecificprotein binding with Universal Protein Block (Dako). Slideswere incubated with an anti-EZH2 antibody (#36-6300; Invi-trogen) at 1:200 followed by an anti-rabbit IgG secondaryantibody and a DAB chromogen for color development (Envi-sion; Dako). LANA staining and hematoxylin and eosin (H&E)staining of tissue microarray sections from the same blockwere carried out by ACSR. Images were captured using a NikonE800M microscope equipped with a Nikon DXM1200 digitalcamera and the Nikon ACT-1 imaging software system (NikonInstruments Inc.).
Western blottingWesternblottingwas carriedout aspreviously described (29)
using antibodies to EZH2 (Cell Signaling Technology), Ephrin-B2 (Abcam), and IkBa (Santa Cruz Biotechnology). Glyceral-dehyde-3-phosphate dehydrogenase (GAPDH), detected by anantibody (Santa Cruz Biotechnology), was used as a loadingcontrol. Immunoreactive bands were visualized by autoradi-ography following development with an enhanced chemilumi-nescence system (Amersham).
RNA extraction, reverse transcription, and real-timequantitative PCR
Total RNAwas isolated using the TRIzol kit (Invitrogen) andtreated with DNase I (Ambion) following the manufacturer'sinstructions. RNA was reverse transcribed into cDNA usingSuperscript II reverse transcriptase as described in the proto-col (Invitrogen). Amplification reactions were carried out in a25 mL reaction volume containing 50 ng total RNA, specificprimers of EZH2, GAPDH, Ephrin-B2 or vFLIP, and SYBRAdvantage qPCR Premix (Clontech). The specificity of the
EZH2 Mediates KSHV-Induced Angiogenesis
www.aacrjournals.org Cancer Res; 72(14) July 15, 2012 3583
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

amplified products was controlled by postamplification dis-sociation curve analyses and agarose gel electrophoresis of theamplified products. The primer sequences are as follows:EZH2-F: 50-TTGTTGGCGGAAGCGTGTAAAATC-30, EZH2-R:50-TCCCTAGTCCCGCGCAATGAGC-30; Ephrin-B2-F: 50- GGA-AGAAGTTCGACAACAAGTCC-30, Ephrin-B2-R: 50- TTCAGCA-AGAGGACCACCAGCGT-30; GAPDH-F: 50-CGGAGTCAACGG-ATTTGGTCGTAT-30, GAPDH-R: 50-AGCCTTCTCCATGGTG-GTGAAGAC; vFLIP-F: 50-CGTCTACGTGGAGAACAGTGA-GCT-30, vFLIP-R: 50-CTGGGCACGGATGACAGGGAAGTG-30.Results were presented as mean � SD from 3 independentexperiments.
Tubule formation assayChilled growth factor-reduced Matrigel (BD Biosciences)
was added to the wells of prechilled 96-well plate at 70 mL perwell. The platewas placed in a 37�C incubator for 30minutes toallow the Matrigel to solidify. BOECs were serum starvedovernight, washed with PBS twice, detached with Accutase(Invitrogen), resuspended in full culturemedium, and plated at3,000 cells per well in the 96-well plates containing the solid-ified Matrigel. Cells were cultured in CO2 incubator at 37�C for7 hours, and images were taken with the Zeiss Axiovertmicroscope with a �10 objective (Carl Zeiss MicroimagingInc.). Tubule length was counted using the Axiovision software(Carl Zeiss Microimaging Inc.). Similar procedures were car-ried out with the SLK cells, except that the step of serumstarvationwas omitted, and cells were plated at 10,000 cells perwell in the Matrigel-coated plate.
Chromatin immunoprecipitation assayChromatin immunoprecipitation (ChIP) assay was carried
out to detect EZH2 binding to the Ephrin-B2 promoter locus,as previously described, using antibody to EZH2 and IgG as acontrol (26). The enrichment of EZH2 on Ephrin-B2 promot-er locus was calculated as the fold increase of the immu-noprecipitated DNA compared with IgG-precipitated DNA.Loci of LANA and RTA promoters were used as low and highbinding controls, as previously described (26). Each datapoints in ChIP figures were averages of at least 3 indepen-dent ChIPs using 3 independent chromatin preparations.The primers for LANA promoter locus are 50-GTTTATAAGT-CAGCCGGACCAA-30 (LANA-F) and 50-GATATAACTCCGC-CCTCCACTA-30 (LANA-R), which amplify a DNA fragment at0.5-kb upstream of transcriptional start site. The primers forRTA promoter locus are 50-CCCCAACACAAGGACCTTTA-30
(RTA-F) and 50-GCTTTTGGATACCCTGGTGA-30 (RTA-R),which amplify a DNA fragment at 1.0-kb upstream oftranscriptional start site. The primers for Ephrin-B2promoter locus are 50-TCGCCATGGCTGTGAGAAGGGA-30
(Ephrin-B2-F) and 50-GGATCGCGGACGCCACTTACT-30
(Ephrin-B2-R), which amplify a DNA fragment from �5 bpto þ142 bp of the transcriptional start site.
Reporter assaySLKp cells were cotransfected with both EZH2 promoter
firefly luciferase reporter construct and pRL-TK Renillaluciferase reporter construct using Lipofectamine 2000
(Invitrogen). Cells were split at 24 hours posttransfectionat 4 � 104 cells per well on 12-well plate. Cells were treatedwith Bay-11 at 38 hours posttransfection for 10 hours. Dualluciferase assay was then carried out following the instruc-tions of the manufacturer (Promega).
ResultsEZH2 is overexpressed in KSHV tumors
EZH2 deregulation is observed in various malignanciesand associated with tumor cell proliferation and angiogen-esis (18, 19, 22). We examined the expression of EZH2 inKaposi sarcoma tumors in a tissue microarray consisting of38 normal biopsies and 173 Kaposi sarcoma biopsies fromdifferent organs. EZH2 was highly expressed in Kaposisarcoma specimens compared with normal controls(Table 1). Of 38 normal specimens, only 4 (11%) had weakEZH2 staining. In contrast, among the 173 Kaposi sarcomaspecimens, 157 (91%) had strong EZH2 staining. Figure 1shows the representative staining EZH2 patterns in skin andlymph node Kaposi sarcoma specimens. Of 25 lymph nodeKaposi sarcoma specimens, 24 (96%) had strong EZH2staining (Table 1). In contrast, only 1 of the 28 (4%) normalcontrol lymph node specimens had weak EZH2 staining. Of77 skin Kaposi sarcoma specimens, 75 (97%) were positivefor EZH2, whereas only 1 of 5 (20%) normal skin controlsshowed very light EZH2 staining (Table 1). The weak stain-ing of this normal skin sample was due to nonspecific uptakeby the macrophages. Insufficient clinical data on the Kaposisarcoma specimens was available for further analysis of thecorrelation of the EZH2 staining pattern and intensity withKaposi sarcoma disease status.
Table 1. Summary of EZH2 staining results
Tissue (n)EZH2-positivetissue, n (%)
Normal tissue (38) 4 (11)Lymph node (28) 1 (3.6)Skin (5) 1 (20)Spleen (2) 0 (0)Anus (2) 1 (50)Mouth (1) 1 (100)
KS tissue (173) 157 (91)Skin (77) 75 (97)Mouth (32) 27 (84)Lymph node (25) 24 (96)Tonsil/adenoids (10) 8 (80)Head/neck (8) 6 (75)Rectum (8) 7 (88)Lung (5) 3 (60)Spleen (2) 2 (100)Anus (2) 2 (100)Small bow (2) 1 (50)Stomach (1) 1 (100)Other (1) 1 (100)
He et al.
Cancer Res; 72(14) July 15, 2012 Cancer Research3584
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

KSHV infection upregulates EZH2 in endothelial cellsTo determine whether KSHV infection induced EZH2
expression in endothelial cells in culture, we infected BOECswith KSHV. BOECs are derived from blood circulating precur-sor cells and have typical endothelial cobblestone morphologyand features (28). BOECs harbor KSHV in Kaposi sarcomapatients and support KSHV infection (30). Almost all cells inthe KSHV-infected BOEC cultures were LANA positive 5 daysafter infection (Fig. 2A) without detectable expression of virallytic protein ORF65 (data not shown), indicating that thesecells were latently infected by KSHV. As expected, KSHVinfection converted the BOECs from cobblestone shape tospindle shape (Supplementary Fig. S1A), with an increase inthe expression of lymphaticmarkers FLT1, PROX1, LYVE1, andpodoplanin (Supplementary Fig. S1B). Thus, similar to vascularendothelial cells (4–6), KSHV infection reprograms BOECs toacquire markers of lymphatic endothelial cells, which arefeatures of Kaposi sarcoma tumor cells. As shown in Fig. 2B,KSHV infection of BOECs increased the expression of EZH2mRNA and protein. Histone H3 is a known substrate of thePRC2. One of the common marks targeted by PRC2 isH3K27me3. Consistent with the upregulation of EZH2, KSHVinfection also increased the level of H3K27me3 in BOECs (Fig.2B). To further confirm these observations, we examined theEZH2 expression in SLK cells with andwithoutKSHV infection.SLK cells are KSHV-negative endothelial cells isolated from a
Kaposi sarcoma tumor. Similar to BOECs, KSHV establishedlatent infection in SLK cells based on the detection of LANAprotein (Fig. 2C) but not lytic protein ORF65 (data not shown).Comparedwith uninfected cells (SLKn), KSHV infection of SLKcells (SLKp) led to the upregulation of EZH2 protein and anincrease in H3K27me3 (Fig. 2D). Together these data indicatedthat KSHV infection upregulated EZH2 and caused an increaseof its substrate H3K27me3 in endothelial cells.
KSHV upregulation of EZH2 is mediated by the NF-kBpathway and viral latent genes vFLIP and LANA
Activation of NF-kB pathway has been shown to upregulatethe expression of EZH2 (31). Because the NF-kB pathway isconstitutively activated in KSHV-infected cells (32), we deter-mined whether it might mediate KSHV induction of EZH2expression. Indeed, inhibition of the NF-kB pathway withspecific inhibitor Bay-11 reduced the expression of EZH2protein in a dose-dependent manner in SLKp cells (Fig. 3A).Similar results were also observed in BOECs. To confirm theseresults, we inhibited the NF-kB pathway with a super sup-pressor IkBaM. Expression of IkBaM reduced the expressionof EZH2 protein (Fig. 3A). Bay-11 also inhibited the expressionof EZH2 mRNA by 80% (Fig. 3B); however, it only inhibitedEZH2 promoter activity by 40% in a reporter assay, suggestingthat the NF-kB pathway might only partially regulate theexpression of EZH2 at the transcriptional level. Alternative
Figure 1. EZH2 is highly expressedin Kaposi sarcoma tumors.Immunohistochemical stainingof EZH2 protein in a paraffin-embedded tissue microarrayconsisting of Kaposi sarcoma andcontrol tissues. A, representativeimages of H&E, LANA, and EZH2staining of a skin Kaposi sarcomaand a normal skin control. B,representative images of H&E,LANA, and EZH2 staining of a lymphnode Kaposi sarcoma and a normallymph node control.
A H&E LANA EZH2 Enlarged
No
rma
l skin
S
kin
KS
B H&E LANA EZH2 Enlarged
No
rma
l ly
mp
h n
od
e
Lym
ph
no
de
KS
100 μm 100 μm 100 μm
100 μm100 μm 100 μm
100 μm 100 μm100 μm
100 μm100 μm100 μm
EZH2 Mediates KSHV-Induced Angiogenesis
www.aacrjournals.org Cancer Res; 72(14) July 15, 2012 3585
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

mechanism such as mRNA stability might also be involved inthe regulation of EZH2 mRNA level by this pathway.
Because KSHV established latent infection in both BOECsand SLK cells, we searched for latent genes that might activatethe NF-kB pathway and cause the upregulation of EZH2. Wefocused on vFLIP and LANA because both of them have beenshown to activate the NF-kB pathway (33, 34). Indeed, expres-sion of vFLIP in BOECs upregulated the expression of EZH2 atboth protein and mRNA levels, which was inhibited by Bay-11(Fig. 3C). Because we were not able to obtain a functionalantibody to vFLIP, we examined the expression of vFLIPmRNAby quantitative reverse transcription PCR (qRT-PCR). Wedetected robust expression of vFLIP mRNA in the vFLIP-transduced cells, which was not affected by Bay-11 (Fig. 3C).Similar to vFLIP, expression of LANA also increased theexpression of EZH2, however, to a less extent, by less than2-fold, in BOECs (Fig. 3C). In contrast to vFLIP and LANA, wedid not observe any increase in EZH2 expression following theexpression of KSHV-latent gene vCyclin (ORF72, data notshown). KSHV infection induces inflammatory cytokines,some of which can activate the NF-kB pathway. Conditionedmedium fromKSHV-infected BOECsonlymarginally increasedthe expression of EZH2 protein (Fig. 3D). Together, theseresults indicated that KSHV upregulation of EZH2 is mainlymediated by vFLIP and LANA through the NF-kB pathway inlatent KSHV-infected endothelial cells.
Induction of EZH2 is essential for KSHV-inducedangiogenesis
Overexpression of EZH2 is often associated with advancedstage of tumors (19, 35). EZH2 has been shown to regulatecancer angiogenesis (22, 24). We determined whether KSHV-induced upregulation of EZH2 contributed to KSHV-induced
angiogenesis. We used an in vitroMatrigel tubulogenesis assayto measure the angiogenic activity of endothelial cells. KSHVinfection of BOECs increased the expression of EZH2 as well astubule formation, whereas knockdownof EZH2 reduced tubuleformation (Fig. 4A). Similar results were also observed in SLKcells (Fig. 4B). These results indicated that EZH2 mediatedKSHV-induced angiogenesis.
EZH2 regulation of KSHV-induced angiogenesis ismediated by Ephrin-B2
To delineate the mechanism involved in EZH2-mediatedKSHV-induced tubule formation, we screened a panel ofproangiogenic genes that could be regulated by EZH2 inKSHV-infected endothelial cells. We found that Ephrin ligandEphrin-B2 protein was upregulated in KSHV-infected BOECsand constitutively expressed in SLKp cells (Fig. 5A). Knock-down of EZH2 reduced the expression of Ephrin-B2 protein(Fig. 5A). Upregulation of Ephrin-B2 by KSHV was alsoobserved at the mRNA level (Fig. 5B). Knockdown of EZH2reduced the expression of Ephrin-B2 mRNA in KSHV-infectedBOECs (Fig. 5B). To further determine whether EZH2 directlybinds to the Ephrin-B2 promoter to regulate its expression, wecarried out ChIP assay in KSHV-infected SLKp cells. A previousstudy has shown that EZH2binds strongly to theRTApromoterchromatin but weakly to the LANA promoter chromatin inKSHV-infected cells. We used these KSHV loci as controls forour experiments. Indeed, comparedwith IgG control, an EZH2-specific antibody enriched 18-fold of the RTA promoter chro-matin but only 3.1-fold of the LANA promoter chromatin (Fig.5C). Knockdown of EZH2 reduced its binding to the RTApromoter chromatin by 82% but had marginal effect onthe LANA promoter chromatin (Fig. 5C). Similar to the RTApromoter, EZH2 enriched Ephrin-B2 promoter chromatin by
A
DA
PI
LA
NA
C
D SLKn SLKpMock KSHV
EZH2
GAPDH
B Mock KSHV
DA
PI
LA
NA
Mock KSHV SLKn SLKp
EZH2
H3K27me3
GAPDH
EZH2
H3K27me3
GAPDHmRNA
Protein Protein
Figure 2. KSHV infectionupregulates EZH2 expression inhuman endothelial cells. A,LANA staining of mock- andKSHV-infected BOECs.Immunofluorescence staining wascarried out at 5 days postinfection.B, KSHV infection upregulated theexpression of EZH2 mRNA andprotein, as well as the level ofH3K27me3 in BOECs. EZH2mRNA was detected bysemiquantitativeRT-PCR (left), andWestern blotting was carried outfor EZH2 andH3K27me3 (right) at 5days postinfection. C, LANAstaining of uninfected SLK cells(SLKn) and KSHV-infected SLKcells (SLKp). D, KSHV infectionupregulated the expression ofEZH2 protein and the level ofH3K27me3 in SLK cells. DAPI,40, 6-diamidino-2-phenylindole.
He et al.
Cancer Res; 72(14) July 15, 2012 Cancer Research3586
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876
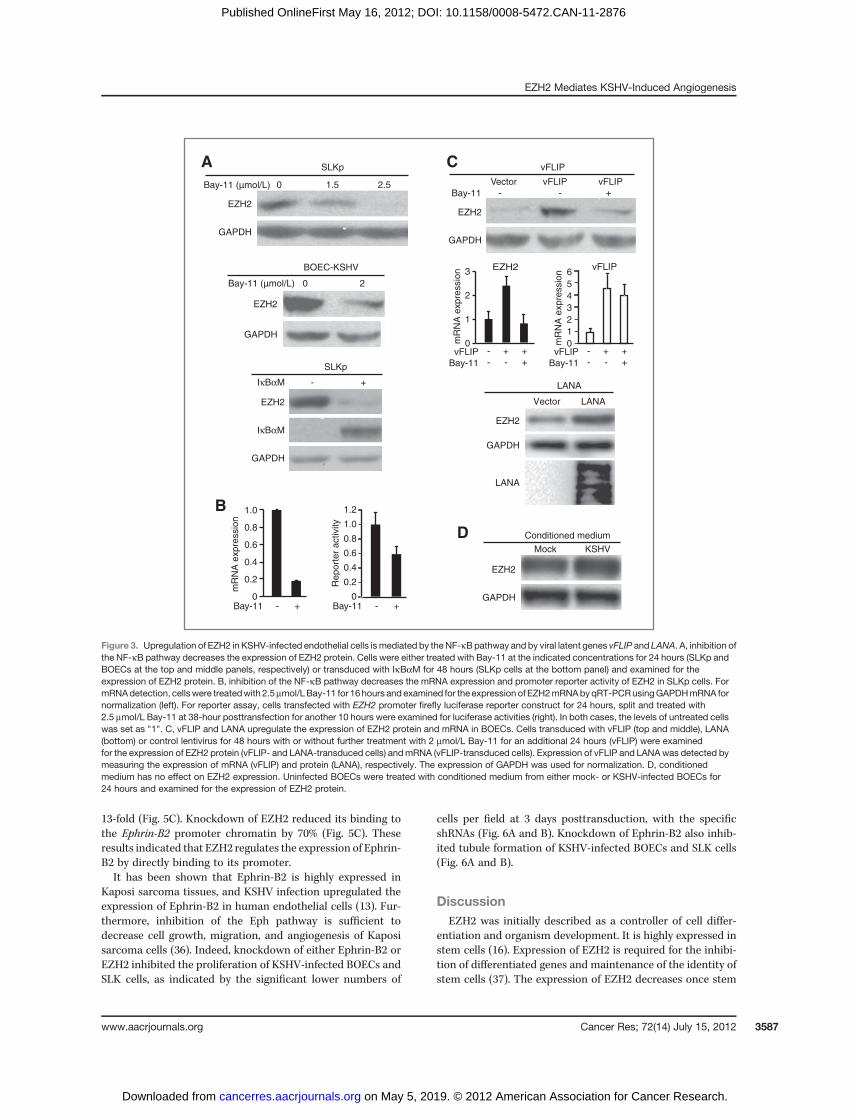
13-fold (Fig. 5C). Knockdown of EZH2 reduced its binding tothe Ephrin-B2 promoter chromatin by 70% (Fig. 5C). Theseresults indicated that EZH2 regulates the expression of Ephrin-B2 by directly binding to its promoter.It has been shown that Ephrin-B2 is highly expressed in
Kaposi sarcoma tissues, and KSHV infection upregulated theexpression of Ephrin-B2 in human endothelial cells (13). Fur-thermore, inhibition of the Eph pathway is sufficient todecrease cell growth, migration, and angiogenesis of Kaposisarcoma cells (36). Indeed, knockdown of either Ephrin-B2 orEZH2 inhibited the proliferation of KSHV-infected BOECs andSLK cells, as indicated by the significant lower numbers of
cells per field at 3 days posttransduction, with the specificshRNAs (Fig. 6A and B). Knockdown of Ephrin-B2 also inhib-ited tubule formation of KSHV-infected BOECs and SLK cells(Fig. 6A and B).
DiscussionEZH2 was initially described as a controller of cell differ-
entiation and organism development. It is highly expressed instem cells (16). Expression of EZH2 is required for the inhibi-tion of differentiated genes and maintenance of the identity ofstem cells (37). The expression of EZH2 decreases once stem
ABay-11 (µmol/L) 0 1.5 2.5
EZH2
GAPDH
IκBαM - +
B 1.0
0.8
0.6
0.4
0.2
0
mR
NA
expre
ssio
n
Bay-11 - +
Report
er
activity
CVector vFLIP vFLIP
- - +Bay-11
3
2
1
0
6
5
4
3
2
1
0mR
NA
expre
ssio
n
vFLIP
Bay-11
EZH2 vFLIP
Vector LANA
Mock KSHV
Bay-11 (µmol/L) 0 2
EZH2
GAPDH
EZH2
IκBαM
GAPDH
1.2
1.0
0.8
0.6
0.4
0.2
0Bay-11 - +
EZH2
GAPDH
EZH2
GAPDH
LANA
mR
NA
expre
ssio
n
- + +
- - +vFLIP
Bay-11
- + +
- - +
EZH2
GAPDH
BOEC-KSHV
vFLIP
LANA
Conditioned mediumD
SLKp
SLKp
Figure 3. Upregulation of EZH2 in KSHV-infected endothelial cells ismediated by theNF-kBpathway and by viral latent genes vFLIP and LANA. A, inhibition ofthe NF-kB pathway decreases the expression of EZH2 protein. Cells were either treated with Bay-11 at the indicated concentrations for 24 hours (SLKp andBOECs at the top and middle panels, respectively) or transduced with IkBaM for 48 hours (SLKp cells at the bottom panel) and examined for theexpression of EZH2 protein. B, inhibition of the NF-kB pathway decreases the mRNA expression and promoter reporter activity of EZH2 in SLKp cells. FormRNAdetection, cellswere treatedwith 2.5mmol/LBay-11 for 16hours and examined for the expression of EZH2mRNAbyqRT-PCRusingGAPDHmRNA fornormalization (left). For reporter assay, cells transfected with EZH2 promoter firefly luciferase reporter construct for 24 hours, split and treated with2.5 mmol/L Bay-11 at 38-hour posttransfection for another 10 hours were examined for luciferase activities (right). In both cases, the levels of untreated cellswas set as "1". C, vFLIP and LANA upregulate the expression of EZH2 protein and mRNA in BOECs. Cells transduced with vFLIP (top and middle), LANA(bottom) or control lentivirus for 48 hours with or without further treatment with 2 mmol/L Bay-11 for an additional 24 hours (vFLIP) were examinedfor the expression of EZH2 protein (vFLIP- and LANA-transduced cells) andmRNA (vFLIP-transduced cells). Expression of vFLIP and LANAwas detected bymeasuring the expression of mRNA (vFLIP) and protein (LANA), respectively. The expression of GAPDH was used for normalization. D, conditionedmedium has no effect on EZH2 expression. Uninfected BOECs were treated with conditioned medium from either mock- or KSHV-infected BOECs for24 hours and examined for the expression of EZH2 protein.
EZH2 Mediates KSHV-Induced Angiogenesis
www.aacrjournals.org Cancer Res; 72(14) July 15, 2012 3587
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

cells differentiated into specific type of cells (38). Subsequentstudies revealed roles of EZH2 in normal cell proliferation andcell-cycle control as well as progression of cancer. In cellculture, overexpression of EZH2 induces cell invasion, growthin soft agar, andmotility, whereas knockdown of EZH2 inhibitscell proliferation and cell invasion (18, 35). Dysregulation ofEZH2 has been associated with tumor aggressiveness, thera-peutic refractory, and tumor angiogenesis in prostate, breast,and ovarian cancer cells (18, 19, 22). EZH2 is regulated byseveral pathways including retinoblastoma-E2F (39), c-Myc(40), NF-kB (31), and p53 (41) pathways, as well as severalmiRNAs (23, 40). In this study, we have found that EZH2 ishighly expressed in Kaposi sarcoma tumors and highly upre-gulated in KSHV-infected endothelial cells. Furthermore, wehave shown that KSHV-induced upregulation of EZH2 ismediated by the NF-kB pathway and KSHV-latent genes vFLIPand LANA.
Like all herpesvirus, KSHVhas both latent and lytic phases inits life cycle. Most tumor cells in Kaposi sarcoma lesionsdisplay KSHV-latent replication program, expressing only ahandful of latent gene products that are involved in host cellsurvival, proliferation, and manipulation of the tumor micro-environment. AmongKSHV-latent genes, vFLIP andLANAhave
been shown to activate the NF-kB pathway (33, 34). Our resultshave shown that KSHV upregulation of EZH2 is mainly medi-ated by vFLIP, and to a less extent, by LANA.Our previous studyhas shown that vFLIP inhibits KSHV lytic replication programto promote viral latency through activation of the NF-kBpathway and inhibition of the AP-1 pathway (42). A recentstudy has shown that EZH2mediates KSHV latency in primaryeffusion lymphomas and in KSHV-infected endothelial cells byregulating latency-specific histone-repressive modificationssuch as H3K27me3 (26). Thus, in addition to the AP-1 pathway,this study has identified EZH2 as a new cellular target thatmediates vFLIP regulation of KSHV latency through the NF-kBpathway.
Because Kaposi sarcoma is a highly angiogenic tumor andKSHV infection of endothelial cells enhances the angiogenicphenotype of these cells, we examined the effect of upregula-tion of EZH2 on KSHV-induced tubule formation on Matrigel.EZH2 is an epigenetic repressor and inhibited the expression ofseveral tumor suppressor genes, including p16, E-cadherin,DAB2IP, and RUNX3 among others (35). Upregulation of EZH2byVEGFpromotes angiogenesis by inhibiting the expression ofangiogenesis inhibitor VASH1 through epigenetic silencing(22). Maspin, an angiogenesis inhibitor, is also downregulated
AControl shRNA Control shRNA
EZH2 shRNA1 EZH2 shRNA2
B
4
3
2
1
0KSHV
shRNA
- + + +
Control Control shRNA1 shRNA2
Fo
ld in
cre
ase
of
tub
ule
le
ng
th
EZH2
GAPDH
Mock KSHV
KSHV KSHV
Tubule formation
Quantification of tubule length
No shRNA Control shRNA
EZH2 shRNA1 EZH2 shRNA2
1.25
1.00
0.75
0.50
0.25
0
shRNA - Control shRNA1 shRNA2
Fo
ld in
cre
ase
of
tub
ule
le
ng
th
Quantification of tubule length
Tubule formation
KSHV
shRNA - + + +
Control Control shRNA1 shRNA2
Protein expression
EZH2
GAPDH
Protein expression
shRNA - Control shRNA1 shRNA2
Figure 4. EZH2 mediates KSHV-induced tubule formation inendothelial cells. A and B,knockdown of EZH2 inhibits tubuleformation in KSHV-infectedBOECs (A) and SLKp (B) cells.Mock- or KSHV-infected BOECswere transduced with lentivirus ofEZH2 shRNA or control shRNA for4 days, serum starved for 12 hoursand examined for the tubuleformation on Matrigel in fullmedium. Representative images oftubule formation (top) were taken at7 hours postplating and quantifiedfor tubule length (middle). Cellswere also examined for theexpressionof EZH2protein at day4lentivirus posttransduction(bottom). Experiments with SLKpcells (B) were carried out as theBOECs, except that cells were notsubjected to serum starvationbefore the tubulogenesis assay.
He et al.
Cancer Res; 72(14) July 15, 2012 Cancer Research3588
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

by EZH2 in a microarray analysis (38). Examination of KSHV-infected BOECs showed that the expression of both genes wereat the lower detection limit of qRT-PCR, albeit VASH1 seemedto be downregulated. Interestingly, we did not observe anysignificant increases in the expression of VASH1 followingEZH2 knockdown (data not shown). In addition to angiogen-esis inhibitors, we have also examined proangiogenenic factorsincluding Eph ligand Ephrin-B2, Notch ligands Jag1 and DLL4,VEGFR3, and MMP-2 and MMP-9. Among these factors,Ephrin-B2 is the only one consistently upregulated followingKSHV infection. Knockdown of EZH2 downregulated theexpression of Ephrin-B2 in KSHV-infected BOECs and SLKcells.Ephrin-B2 is a transmembrane ligand and regulates
angiogenic growth of the blood vessel and lymphatic endo-thelium (43). Eph-ephrin binding leads to bidirectionalsignaling in cells expressing the receptor (forward signaling)or ligand (reverse signaling). Ephrin-B2 is upregulated
during physiologic and pathologic angiogenesis in the adult,and loss of Ephrin-B2 leads to vascular defects (44). Intrigu-ingly, Ephrin-B2–mediated reverse signaling controlsVEGFR internalization. Endothelial cells lacking Ephrin-B2reverse signaling are unable to internalize VEGFR2 andVEGFR3 and cannot properly transmit VEGF signals, result-ing in impairment of angiogenesis (45, 46). Previous studieshave shown that Ephrin-B2 is highly expressed in Kaposisarcoma tissues, KSHV infection upregulates Ephrin-B2 inendothelial cells (13), and the inhibition of Ephrin-B2blocked tumor growth and reduced tumor vessel densityand perfusion in a Kaposi sarcoma tumor xenograft model(36). Our results have confirmed that Ephrin-B2 expressionis increased following KSHV infection. Furthermore, we haveshown that knockdown of EZH2 inhibits Ephrin-B2 expres-sion at both protein and mRNA levels, and knockdown ofeither Ephrin-B2 or EZH2 inhibits cell growth and tubuleformation in KSHV-infected endothelial cells.
A B
20
15
10
5
0
Fold
of enrichm
ent
lgG
EZH2
LANA RTA Ephrin-B2
Enrichm
ent (%
)Control shRNA
EZH2 shRNA
4
3
2
1
0
mR
NA
expre
ssio
n
C
EZH2
Ephrin-B2
KSHV
shRNA - + + +
Control Control shRNA1 shRNA2
shRNA - Control shRNA1 shRNA2
EZH2
Ephrin-B2
GAPDH
EZH2
Ephrin-B2
GAPDH
KSHV
shRNA - + + +
Control Control shRNA1 shRNA2
4
3
2
1
0
mR
NA
expre
ssio
n
KSHV
shRNA - + + +
Control Control shRNA1 shRNA2
LANA RTA Ephrin-B2
150
120
90
60
30
0
BOEC
SLKp
Figure 5. EZH2 is required for the expression of Ephrin-B2 in KSHV-infected endothelial cells. A, EZH2 is required for the expression of Ephrin-B2 protein inBOECs and SLK cells. Mock- and KSHV-infected BOECs (top) and SLKp cells (bottom) were transduced with lentivirus of EZH2 shRNA or control shRNA for4 days and examined for the expression of EZH2 and Ephrin-B2 proteins. B, EZH2 is required for the expression of Ephrin-B2 mRNA in BOECs.Mock- and KSHV-infected BOECs described in A were also examined for the expression of EZH2 and Ephrin-B2 mRNAs. C, EZH2 directly bindsto the Ephrin-B2 promoter. Chromatin from SLKp cells with or without transduction with lentivirus of EZH2 shRNA or control shRNA for 4 days wereimmunoprecipitated with an EZH2 antibody or IgG control. Immunoprecipitated DNA was extracted and examined for the presence of Ephrin-B2promoter DNA by qPCR. RTA and LANA promoters known to have strong and weak EZH2 binding, respectively, were used as controls. Chromatinenrichment by the EZH2 antibody was calculated as the percentage of the immunoprecipitated DNA and compared with IgG precipitated DNA(1 in the left panel). Binding of EZH2 protein to the promoters in cells with shRNA knockdown was compared with those transduced with control shRNA(100% in the right panel).
EZH2 Mediates KSHV-Induced Angiogenesis
www.aacrjournals.org Cancer Res; 72(14) July 15, 2012 3589
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

EZH2 and its associated PRC2 complex typically repressor maintain the repression status of their target genes(37, 47, 48). EZH2 is associated with the promoters of anumber of tumor suppressor genes and therefore inhibitstheir expression (35, 47). However, EZH2 also upregulatessome positive regulators, including CCND1, CCNE1, CCNA2,and c-Myb among others (20, 39). A recent study has shownthat EZH2, through interacting with RelA/RelB, is requiredfor the establishment of constitutive activation of a subset ofNF-kB–targeted genes (49). Our results have shown that
EZH2 binds to the promoter of angiogenesis enhancerEphrin-B2 and upregulated its expression, indicating EZH2might also be required for either the activation or themaintenance of the activated state of this positive angio-genesis regulator. Further studies are required to delineatethe mechanism mediating this positive regulation.
In summary, we have shown that KSHV promotes angio-genesis by upregulating a proangiogenic Ephrin-B2 throughEZH2. These findings suggest that both EZH2 and Ephrin-B2are promising novel targets for KSHV-induced angiogenesis.
A B
EZH2
EphrinB2
GAPDH
4
3
2
1
0 - + + +
Control Control Ephrin-B2 EZH2
Fo
ld in
cre
ase
of
tub
ule
le
ng
th
Mock KSHV
KSHV KSHV
No shRNA Control shRNA
Ephrin-B2 shRNA EZH2 shRNA
Morphology Morphology
Control shRNA Control shRNA
Ephrin-B2 shRNA EZH2 shRNA
Mock KSHV
KSHV KSHV
No shRNA Control shRNA
Ephrin B2-shRNA EZH2 shRNA
Tubule formation Tubule formation
KSHV
shRNA
Quantification of tubule length Quantification of tubule length
shRNA - Control Ephrin-B2 EZH2
1.6
1.2
0.8
0.4
0Fo
ld in
cre
ase
of
tub
ule
le
ng
th
- + + +
Control Control Ephrin-B2 EZH2
KSHV
shRNA
EZH2
EphrinB2
GAPDH
Protein expression Protein expression
shRNA - Control Ephrin-B2 EZH2
Control shRNA Control shRNA
Ephrin-B2 shRNA EZH2 shRNA
Figure 6. Ephrin-B2 mediates EZH2enhancement of KSHV-inducedtubule formation in endothelialcells. A and B, knockdown of eitherEphrin-B2 or EZH2 inhibits cellgrowth and tubule formation inKSHV-infected BOECs (A) andSLKp (B) cells. Mock or KSHV-infected BOECs were transducedwith lentivirus of EZH2 shRNA, 3pooled Ephrin-B2 shRNAs, orcontrol shRNA for 4 days andexamined for cell growthbydirectlycounting of cell number on thephase contrast images (top).Transduced cells were also serumstarved for 12 hours and examinedfor the tubule formation onMatrigelin full medium. Representativeimagesof tubule formation (secondpanel) were taken at 7-hourpostplating and quantified fortubule length (third panel). Cellswere examined for the expressionof EZH2 and Ephrin-B2 proteins atday 4 lentivirus posttransduction(bottom). Experiments with SLKpcells were carried out as theBOECs, except cells were notsubjected to serum starvationbefore the tubulogenesis assay.
He et al.
Cancer Res; 72(14) July 15, 2012 Cancer Research3590
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: M. He, M. Cannon, S.-J. GaoDevelopment of methodology: M. He, M. Schutten, P. Gill, M. CannonAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.): T. BakkenAnalysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): M. He, M. Schutten, Z. Toth, M. Cannon, S.-J. GaoWriting, review, and/or revision of the manuscript:M. He, M. Schutten, Z.Toth, P. Gill, M. Cannon, S.-J. GaoAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases):M. Schutten, Z. Toth, J.U. Jung, S.-J. GaoStudy supervision: M. Cannon, S.-J. GaoPerformed RT-qPCR for the Bay-11 study: W. Zhang
AcknowledgmentsThe authors thank Drs. Adam Grundhoff, Robert Hebbel, Inder Verma, Chris
Boshoff, and Felix Hoppe-Seyler for providing us critical reagents, Paula Overnin the Comparative Pathology Shared Resource, Masonic Cancer Center atthe University of Minnesota for her technical assistance for the IHC experiments,and members of Dr. Gao's laboratory for technical assistance and helpfuldiscussions.
Grant SupportThis work was funded by an American Cancer Society Research Scholar grant
(RSG-10-060-01-MPC) to M. Cannon and NIH grants to S.-J. Gao (R01CA132637,R01CA124332, and R01CA096512).
Received August 29, 2011; revised May 3, 2012; accepted May 5, 2012;published OnlineFirst May 16, 2012.
References1. Mesri EA, Cesarman E, Boshoff C. Kaposi's sarcoma and its associ-
ated herpesvirus. Nat Rev Cancer 2010;10:707–19.2. Ciufo DM, Cannon JS, Poole LJ, Wu FY, Murray P, Ambinder RF, et al.
Spindle cell conversion by Kaposi's sarcoma-associated herpesvirus:formation of colonies and plaques with mixed lytic and latent geneexpression in infected primary dermal microvascular endothelial cellcultures. J Virol 2001;75:5614–26.
3. Gao SJ, Deng JH, Zhou FC. Productive lytic replication of a recom-binant Kaposi's sarcoma-associated herpesvirus in efficient primaryinfection of primary human endothelial cells. J Virol 2003;77:9738–49.
4. Hong YK, Foreman K, Shin JW, Hirakawa S, Curry CL, Sage DR, et al.Lymphatic reprogramming of blood vascular endothelium by Kaposi'ssarcoma-associated herpesvirus. Nat Genet 2004;36:683–5.
5. WangHW, TrotterMW, Lagos D, Bourboulia D, Henderson S,MakinenT, et al. Kaposi's sarcoma herpesvirus-induced cellular reprogram-ming contributes to the lymphatic endothelial gene expression inKaposi sarcoma. Nat Genet 2004;36:687–93.
6. Carroll PA, Brazeau E, Lagunoff M. Kaposi's sarcoma-associatedherpesvirus infection of blood endothelial cells induces lymphaticdifferentiation. Virology 2004;328:7–18.
7. Ye FC, Blackbourn DJ, Mengel M, Xie JP, Qian LW, Greene W, et al.Kaposi's sarcoma-associated herpesvirus promotes angiogenesis byinducing angiopoietin-2 expression via AP-1 and Ets1. J Virol2007;81:3980–91.
8. Qian LW, Xie J, Ye F, Gao SJ. Kaposi's sarcoma-associated herpes-virus infection promotes invasion of primary human umbilical veinendothelial cells by inducing matrix metalloproteinases. J Virol2007;81:7001–10.
9. Qian LW, Greene W, Ye F, Gao SJ. Kaposi's sarcoma-associatedherpesvirus disrupts adherens junctions and increases endothelialpermeability by inducing degradation of VE-cadherin. J Virol 2008;82:11902–12.
10. Shin YC, Joo CH, Gack MU, Lee HR, Jung JU. Kaposi's sarcoma-associated herpesvirus viral IFN regulatory factor 3 stabilizes hypoxia-inducible factor-1 alpha to induce vascular endothelial growth factorexpression. Cancer Res 2008;68:1751–9.
11. Wang L, Damania B. Kaposi's sarcoma-associated herpesvirus con-fers a survival advantage to endothelial cells. Cancer Res 2008;68:4640–8.
12. Xie J, Pan H, Yoo S, Gao SJ. Kaposi's sarcoma-associated herpes-virus induction of AP-1 and interleukin 6 during primary infectionmediated by multiple mitogen-activated protein kinase pathways. JVirol 2005;79:15027–37.
13. MasoodR, XiaG, SmithDL, Scalia P, Still JG, Tulpule A, et al. Ephrin B2expression in Kaposi's sarcoma is induced by human herpesvirus type8: phenotype switch from venous to arterial endothelium. Blood2005;105:1310–8.
14. Sivakumar R, Sharma-Walia N, Raghu H, Veettil MV, Sadagopan S,Bottero V, et al. Kaposi's sarcoma-associated herpesvirus inducessustained levels of vascular endothelial growth factors A and C early
during in vitro infection of human microvascular dermal endothelialcells: biological implications. J Virol 2008;82:1759–76.
15. Iso T, Maeno T, Oike Y, Yamazaki M, Doi H, Arai M, et al. Dll4-selectiveNotch signaling induces ephrinB2 gene expression in endothelial cells.Biochem Biophys Res Commun 2006;341:708–14.
16. Sauvageau M, Sauvageau G. Polycomb group proteins: multi-facetedregulators of somatic stem cells and cancer. Cell Stem Cell 2010;7:299–313.
17. Vire E, Brenner C, Deplus R, Blanchon L, Fraga M, Didelot C, et al. ThePolycomb group protein EZH2 directly controls DNA methylation.Nature 2006;439:871–4.
18. Chang CJ, Yang JY, Xia W, Chen CT, Xie X, Chao CH, et al. EZH2promotes expansion of breast tumor initiating cells through activationof RAF1-beta-catenin signaling. Cancer Cell 2011;19:86–100.
19. Varambally S, Dhanasekaran SM, Zhou M, Barrette TR, Kumar-Sinha C, Sanda MG, et al. The polycomb group protein EZH2is involved in progression of prostate cancer. Nature 2002;419:624–9.
20. Shi B, Liang J, Yang X, Wang Y, Zhao Y, Wu H, et al. Integrationof estrogen and Wnt signaling circuits by the polycomb groupprotein EZH2 in breast cancer cells. Mol Cell Biol 2007;27:5105–19.
21. McCabe MT, Graves AP, Ganji G, Diaz E, Halsey WS, Jiang Y, et al.Mutation of A677 in histone methyltransferase EZH2 in humanB-cell lymphoma promotes hypertrimethylation of histone H3on lysine 27 (H3K27). Proc Natl Acad Sci U S A 2012;109:2989–94.
22. Lu C, Han HD, Mangala LS, Ali-Fehmi R, Newton CS, Ozbun L, et al.Regulation of tumor angiogenesis by EZH2. Cancer Cell 2010;18:185–97.
23. Kottakis F, Polytarchou C, Foltopoulou P, Sanidas I, Kampranis SC,Tsichlis PN. FGF-2 regulates cell proliferation, migration, and angio-genesis through an NDY1/KDM2B-miR-101-EZH2 pathway. Mol Cell2011;43:285–98.
24. Smits M, Nilsson J, Mir SE, van der Stoop PM, Hulleman E, Niers JM,et al. miR-101 is down-regulated in glioblastoma resulting in EZH2-induced proliferation, migration, and angiogenesis. Oncotarget 2010;1:710–20.
25. Gunther T, Grundhoff A. The epigenetic landscape of latent Kaposi'ssarcoma-associated herpesvirus genomes. PLoS Pathog 2010;6:e1000935.
26. Toth Z, Maglinte DT, Lee SH, Lee HR, Wong LY, Brulois KF, et al.Epigenetic analysis of KSHV latent and lytic genomes. PLoS Pathog2010;6:e1001013.
27. Grundhoff A, Ganem D. Inefficient establishment of KSHV latencysuggests an additional role for continued lytic replication in Kaposi'ssarcoma pathogenesis. J Clin Invest 2004;113:124–36.
28. Lin Y, Weisdorf DJ, Solovey A, Hebbel RP. Origins of circulatingendothelial cells and endothelial outgrowth from blood. J Clin Invest2000;105:71–7.
EZH2 Mediates KSHV-Induced Angiogenesis
www.aacrjournals.org Cancer Res; 72(14) July 15, 2012 3591
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

29. He M, Bakken T, Kassimova A, Boshoff C, Philpott N, Cannon ML.Focal adhesion kinase is required for KSHV vGPCR signaling. MolCarcinog 2011 [Epub ahead of print].
30. DellaBella S, TaddeoA,CalabroML,Brambilla L, BellinviaM,BergamoE, et al. Peripheral blood endothelial progenitors as potential reservoirsof Kaposi's sarcoma-associated herpesvirus. PLoS One 2008;3:e1520.
31. Trenkmann M, Brock M, Gay RE, Kolling C, Speich R, Michel BA, et al.Expression and function of EZH2 in synovial fibroblasts: epigeneticrepression of the Wnt inhibitor SFRP1 in rheumatoid arthritis. AnnRheum Dis 2011;70:1482–8.
32. Keller SA, Hernandez-Hopkins D, Vider J, Ponomarev V, Hyjek E,Schattner EJ, et al. NF-kappaB is essential for the progression ofKSHV- and EBV-infected lymphomas in vivo. Blood 2006;107:3295–302.
33. MattaH,Chaudhary PM.Activation of alternativeNF-kappaBpathwayby human herpes virus 8-encoded Fas-associated death domain-likeIL-1 beta-converting enzyme inhibitory protein (vFLIP). Proc Natl AcadSci U S A 2004;101:9399–404.
34. Konrad A, Wies E, Thurau M, Marquardt G, Naschberger E, HentschelS, et al. A systemsbiology approach to identify the combination effectsof human herpesvirus 8 genes on NF-kappaB activation. J Virol2009;83:2563–74.
35. Chang CJ, Hung MC. The role of EZH2 in tumour progression. Br JCancer 2012;106:243–7.
36. Scehnet JS, Ley EJ, Krasnoperov V, Liu R, Manchanda PK, Sjoberg E,et al. The role of Ephs, Ephrins, andgrowth factors inKaposi's sarcomaand implications of EphrinB2 blockade. Blood 2009;113:254–63.
37. Delgado-Olguin P, Huang Y, Li X, Christodoulou D, Seidman CE,Seidman JG, et al. Epigenetic repression of cardiac progenitor geneexpression by Ezh2 is required for postnatal cardiac homeostasis. NatGenet 2012;44:343–7.
38. Velichutina I, Shaknovich R, Geng H, Johnson NA, Gascoyne RD,Melnick AM, et al. EZH2-mediated epigenetic silencing in germinalcenter B cells contributes to proliferation and lymphomagenesis.Blood 2010;116:5247–55.
39. Bracken AP, Pasini D, Capra M, Prosperini E, Colli E, Helin K. EZH2 isdownstream of the pRB-E2F pathway, essential for proliferation andamplified in cancer. EMBO J 2003;22:5323–35.
40. SanderS,Bullinger L,KlapprothK, Fiedler K,KestlerHA,Barth TF, et al.MYC stimulates EZH2 expression by repression of its negative reg-ulator miR-26a. Blood 2008;112:4202–12.
41. Tang X, MilyavskyM, Shats I, Erez N, Goldfinger N, Rotter V. Activatedp53 suppresses the histone methyltransferase EZH2 gene. Oncogene2004;23:5759–69.
42. Ye FC, Zhou FC, Xie JP, Kang T, Greene W, Kuhne K, et al. Kaposi'ssarcoma-associated herpesvirus latent gene vFLIP inhibits viral lyticreplication through NF-kappaB-mediated suppression of the AP-1pathway: a novel mechanism of virus control of latency. J Virol2008;82:4235–49.
43. Kullander K, Klein R. Mechanisms and functions of Eph and ephrinsignalling. Nat Rev Mol Cell Biol 2002;3:475–86.
44. Salvucci O, Maric D, Economopoulou M, Sakakibara S, Merlin S,Follenzi A, et al. EphrinB reverse signaling contributes to endothelialand mural cell assembly into vascular structures. Blood 2009;114:1707–16.
45. Sawamiphak S, Seidel S, Essmann CL, Wilkinson GA, Pitulescu ME,Acker T, et al. Ephrin-B2 regulates VEGFR2 function in developmentaland tumour angiogenesis. Nature 2010;465:487–91.
46. Wang Y, Nakayama M, Pitulescu ME, Schmidt TS, Bochenek ML,Sakakibara A, et al. Ephrin-B2 controls VEGF-induced angiogenesisand lymphangiogenesis. Nature 2010;465:483–6.
47. Cheng AS, Lau SS, Chen Y, Kondo Y, Li MS, Feng H, et al. EZH2-mediated concordant repression of Wnt antagonists promotesbeta-catenin-dependent hepatocarcinogenesis. Cancer Res 2011;71:4028–39.
48. Paul TA, Bies J, Small D, Wolff L. Signatures of polycomb repressionand reduced H3K4 trimethylation are associated with p15INK4b DNAmethylation in AML. Blood 2010;115:3098–108.
49. Lee ST, Li Z, Wu Z, AauM, Guan P, Karuturi RK, et al. Context-specificregulation of NF-kappaB target gene expression by EZH2 in breastcancers. Mol Cell 2011;43:798–810.
Cancer Res; 72(14) July 15, 2012 Cancer Research3592
He et al.
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876

Correction
Correction: Cancer Angiogenesis Induced byKaposi's Sarcoma-Associated Herpesvirus IsMediated by EZH2
In this article (Cancer Res 2012;72:3582–92), which was published in the July 15,2012 issue of Cancer Research (1), an incorrect version of Figure 2B right panel,Figure 3A middle panel, and Figure 3D was published. The correct version of thefigure panels is provided below. The conclusions of the article remainunchanged. The authors regret this error.
Published panel
Mock KSHV
EZH2
H3K27me3
GAPDH
Protein
Corrected panel
Mock KSHV
EZH2
H3K27me3
GAPDH
Protein
Mock KSHV
Published original Corrected original
Mock KSHV
GAPDH GAPDH
EZH2 EZH2
Repeat
GAPDH
Mock KSHV
EZH2
Figure 2B.
CancerResearch
www.aacrjournals.org 5603

Bay-11 (µmol/L) 0 2
EZH2
GAPDH
BOEC-KSHV
Bay-11 (µmol/L) 0 2
EZH2
GAPDH
BOEC-KSHV
Published panel Corrected panel
Bay-11 (µmol/L) 0 2
Published original Corrected original
Bay-11 0 2
(µmol/L)
Bay-11 (µmol/L) 0 2
Bay-11 0 2
(µmol/L)
GAPDHGAPDH
EZH2EZH2
Repeat
Bay-11 0 2
(µmol/L)
Bay-11 0 2
(µmol/L)
GAPDH
EZH2
Figure 3A.
Correction
Cancer Res; 73(17) September 1, 2013 Cancer Research5604

Reference1. He ML, Zhang W, Bakken T, Schutten M, Toth Z, Jung JU, et al. Cancer angiogenesis
induced by Kaposi's sarcoma-associated herpesvirus is mediated by EZH2. Cancer Res2012;72:3582–92.
Published OnlineFirst August 15, 2013.doi: 10.1158/0008-5472.CAN-13-2115�2013 American Association for Cancer Research.
Mock KSHV
EZH2
GAPDH
Conditioned medium Conditioned medium
Published panel Corrected panel
Mock KSHV
EZH2
GAPDH
Published original Corrected original
Mock KSHV Mock KSHV
GAPDH GAPDH
EZH2
GAPDH
Mock KSHV
Repeat
Mock KSHV
Figure 3D.
Correction
www.aacrjournals.org Cancer Res; 73(17) September 1, 2013 5605

2012;72:3582-3592. Published OnlineFirst May 16, 2012.Cancer Res Meilan He, Wei Zhang, Thomas Bakken, et al. Herpesvirus Is Mediated by EZH2
Associated−Cancer Angiogenesis Induced by Kaposi Sarcoma
Updated version
10.1158/0008-5472.CAN-11-2876doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2012/05/15/0008-5472.CAN-11-2876.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/72/14/3582.full#ref-list-1
This article cites 48 articles, 24 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/72/14/3582.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/72/14/3582To request permission to re-use all or part of this article, use this link
on May 5, 2019. © 2012 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst May 16, 2012; DOI: 10.1158/0008-5472.CAN-11-2876



![Aquaporins as diagnostic and therapeutic targets in cancer ...cancer, Laryngeal cancer, Lung cancer [43], Nasopharyngeal cancer, Ovarian cancer [37] Tumor grade, prognosis, tumor angiogenesis,](https://img.dokumen.tips/doc/110x75/5ffa8fafa51a2a21db58011f/aquaporins-as-diagnostic-and-therapeutic-targets-in-cancer-cancer-laryngeal.jpg)

























