Embed Size (px)
Citation preview



Breviář forenzní genetiky Forenzní DNA analýza v otázkách a odpovědích
Halina Šimková
Tribun EU
2012


Breviář forenzní genetiky Forenzní DNA analýza v otázkách a odpovědích
Halina Šimková
recenzovali:
doc. Jiří Drábek, Ph.D.
Ing. Aleš Hořínek
Mgr. Vlastimil Stenzl

Tato publikace je spolufinancována Evropským sociálním
fondem a státním rozpočtem České republiky z projektu
CZ.1.07/2.3.00/09.0080 – „4n6gen – vzdělávání v oblasti
forenzní genetiky“.
Řešitelem uvedeného projektu je Československá
společnost pro forenzní genetiku – www.cssfg.org.
Publikace obsahuje řadu ilustrativních příkladů užití forenzně
genetické expertízy v reálných situacích. Tyto příklady vycházejí ze
skutečných případů, na nichž autorka v průběhu své praxe pracovala,
či s nimiž se seznámila, jsou však oproti realitě zjednodušeny či
pozměněny tak, aby co nejlépe sloužily jako modelové; jsou vynechána
či zcela změněna všechna identifikační data týkající se jak osob, tak
míst.
© Halina Šimková, 2012
Illustrations © Halina Šimková, 2012
Cover illustrations © Halina Šimková, 2012
This edition © Tribun EU, 2012
ISBN

5
Obsah Seznam ilustrací ........................................................................................... 7
Předmluva .................................................................................................. 11
Forenzní genetika jako obor ................................................................ 13
Biologické stopy a biologický materiál ............................................ 18
Obecné pojmy .................................................................................................... 18
Tkáňový původ, druhový původ a původce biologického
materiálu ............................................................................................................. 19
Využitelnost biologického materiálu pro forenzně genetickou
analýzu ................................................................................................................. 25
Nakládání s biologickými stopami ............................................................ 29
Vlastnosti a chování biologického materiálu ve stopách................. 37
Důkazní potenciál stopy ................................................................................ 46
Seznámení s genetikou ........................................................................... 50
Základní fakta o DNA ...................................................................................... 50
Základní principy fungování dědičné informace ................................ 58
Forenzní potenciál genetické analýzy .............................................. 69
Individualita a identifikace .......................................................................... 69
Určení příbuznosti ........................................................................................... 80
Predikce biogeografického původu a fenotypových znaků osoby
................................................................................................................................. 95
Obecné principy forenzně genetické analýzy ................................ 99
Principy identifikace ....................................................................................... 99
Principy určení příbuznosti ...................................................................... 117
Principy predikce biogeografického původu a fenotypových
znaků osoby..................................................................................................... 122
Technologie forenzně genetické analýzy ..................................... 127
Základní kroky forenzně genetické analýzy ...................................... 127
Izolace (extrakce) DNA ............................................................................... 129
Kvantifikace (a případná charakterizace) DNA ................................ 140
Amplifikace DNA ........................................................................................... 146
Elektroforéza DNA ........................................................................................ 153

6
Analýza STR lokusů ...................................................................................... 158
Analýza HVR mt DNA ................................................................................... 161
Analýza SNP lokusů ...................................................................................... 164
Nakládání se stopami obsahujícími degradovanou DNA
a/nebo malá množství DNA ...................................................................... 166
Interpretace forenzně genetických dat ......................................... 175
Obecné kroky a pravidla interpretace dat .......................................... 175
Spolehlivost dat – technická kvalita dat ............................................... 176
Kvalita v laboratoři ....................................................................................... 179
Vypovídací hodnota dat .............................................................................. 182
Zdroje chyb ve forenzně genetické expertíze .................................... 185
Uchovávání zdrojů genetických informací ................................... 190
Banky (archivy, depozity) biologického materiálu ......................... 190
Banky izolátů DNA ........................................................................................ 191
DNA databáze .................................................................................................. 192
„Non–human“ forenzní analýza živočišné, rostlinné a
mikrobiální DNA .................................................................................... 196
Forenzní analýza živočišné DNA ............................................................. 196
Forenzní analýza rostlinné DNA ............................................................. 198
Forenzní analýza mikrobiální DNA ........................................................ 199
Doslov ........................................................................................................ 201
O autorce ................................................................................................... 202
Doporučená literatura a informační zdroje ................................. 203
Rejstřík ...................................................................................................... 204

7
Seznam ilustrací
Obrázek 1 – Charakteristiky biologické stopy .........................................................20
Obrázek 2 – Degradace DNA – příčiny .........................................................................32
Obrázek 3 – Odběr biologického materiálu ...............................................................34
Obrázek 4 – Kontaminace stopy .....................................................................................36
Obrázek 5 – Primární a sekundární přenos biologického materiálu .............44
Obrázek 6 – Struktura jednořetězcové DNA ............................................................51
Obrázek 7 – Struktura dvouřetězcové DNA .............................................................52
Obrázek 8 – Karyotyp ..........................................................................................................53
Obrázek 9 – Genetický kód ...............................................................................................55
Obrázek 10 – DNA v lidské buňce ..................................................................................57
Obrázek 11 – Variabilita lokusů v DNA ......................................................................60
Obrázek 12 – Meióza (redukční dělení) .....................................................................62
Obrázek 13 – Oplození .......................................................................................................63
Obrázek 14 – Dědičnost autozomálních lokusů .....................................................64
Obrázek 15 – Dědičnost Y–chromozomálních lokusů .........................................66
Obrázek 16 – Dědičnost X–chromozomálních lokusů . ........................................67
Obrázek 17 – Dědičnost mitochondriální DNA .......................................................68
Obrázek 18 – Genetický profil osoby ..........................................................................71
Obrázek 19 – Genetická informace jednovaječných dvojčat ............................74
Obrázek 20 – Genetická informace dvouvaječných dvojčat .............................76
Obrázek 21 – Genetická informace chimérického jedince .................................78
Obrázek 22 – Rodokmen – stupeň příbuznosti ......................................................87
Obrázek 23 – Rodokmen – dědičnost autozomálních lokusů ..........................88
Obrázek 24 – Rodokmen – dědičnost X–chromozomálních lokusů ...............89
Obrázek 25 – Rodokmen – dědičnost Y–chromozomálních lokusů ...............90
Obrázek 26 – Rodokmen – dědičnost mitochondriální DNA ............................91
Obrázek 27 – Rodokmen – konsangvinita a incest ...............................................93
Obrázek 28 – Rodokmen – mnohotný incest ...........................................................94
Obrázek 29 – Struktura krátké tandemové repetice (STR) . .......................... 101
Obrázek 30 – Alely STR lokusu – standardní a mikrovariantní. ................... 103
Obrázek 31 – Srovnání genotypu stopy a osoby .................................................. 106
Obrázek 32 – Struktura jednonukleotidového polymorfismu (SNP). ........ 109
Obrázek 33 – Struktura mitochondriální DNA (mtDNA) ................................ 113
Obrázek 34 – Určení pohlaví ........................................................................................ 123

8
Obrázek 35 – Základní kroky technologického zpracování stopy................ 128
Obrázek 36 – Princip organické extrakce DNA. .................................................... 133
Obrázek 37 – Princip vyvázání enzymatických kofaktorů .............................. 134
Obrázek 38 – FTA karty ................................................................................................. 135
Obrázek 39 – Vazba DNA na siliku ............................................................................ 137
Obrázek 40 – Diferenciální lýza .................................................................................. 138
Obrázek 41 – Laserová mikrodisekce (LCM) ......................................................... 139
Obrázek 42 – Spektrofotometrická kvantifikace ................................................. 142
Obrázek 43 – Fluorimetrická kvantifikace ............................................................. 143
Obrázek 44 – Specifická kvantifikace – princip .................................................... 144
Obrázek 45 – Specifická kvantifikace – slot blot hybridizace ........................ 145
Obrázek 46 – Složení PCR směsi a denaturační fáze PCR ............................... 148
Obrázek 47 – Anelační a elongační fáze PCR ........................................................ 149
Obrázek 48 – Princip elektroforézy DNA ............................................................... 154
Obrázek 49 – Princip kapilární elektroforézy ....................................................... 157
Obrázek 50 – Monoplexová a multiplexová analýza STR lokusů . ............... 159
Obrázek 51 – Sekvenace asymetrickou PCR ......................................................... 163
Obrázek 52 – Monoplexová a multiplexová analýza SNP lokusů ................ 165
Obrázek 53 – Mechanismy degradace DNA .......................................................... 166
Obrázek 54 – Klasická a miniSTR analýza ............................................................. 169
Obrázek 55 – Stochastické jevy ................................................................................... 173
Obrázek 56 – Základní kroky procesu práce s daty. ........................................... 175
Obrázek 57 – Možné formy uchovávání zdrojů genetických informací .... 190

9

10
Motto:
Cílem vědy není otvírat dveře nekonečné moudrosti, nýbrž
vytknout meze nekonečnému omylu.
Eugen Berthold Friedrich Brecht
Děkuji recenzentům za všechny cenné připomínky, výtky a postřehy, které
nemalou měrou přispěly ke zkvalitnění této publikace. Své rodině pak
děkuji za neutuchající podporu, vstřícnost a pochopení.
Věnováno mému gymnaziálnímu profesorovi biologie Petru Pačovskému,
jehož expresívní líčení tajného vnitřního života buněk, jakož i ostatních
mystérií živé přírody, bylo zcela neodolatelné.

11
Předmluva,
která by neměla zůstat nepřečtena
Forenzně genetický posudek bývá silným argumentem jak v oblasti
trestního práva, tak v rámci práva občanského. Jako každý takto
mocný nástroj může při správném použití velmi účinně pomáhat
k nastolení spravedlnosti a práva, při nesprávném použití však
může napáchat nesmírné škody.
Umět správně vnímat a posoudit výsledky jakékoliv genetické
analýzy znamená rozumět alespoň základům věci, principům, které
se v genetice uplatňují, a jejich logickým důsledkům.
Pro člověka bez přírodovědného vzdělání je ovšem velmi těžké, ba
až nemožné porozumět všem zákonitostem, postupům a závěrům
tohoto oboru na základě „samostudia“. Musí to tedy být znalec, kdo
se pokusí objasnit všechny aspekty forenzní genetiky řečí
srozumitelnou a přístupnou.
Tento breviář je proto psán jako populárně naučné pojednání,
primárně zaměřené na čtenáře, který má o genetice a forenzní
genetice jen základní představu, ale rád by do oboru pronikl
hlouběji; rozebírány jsou především univerzálně platné odborné
aspekty forenzní genetiky, přičemž publikace se současně záměrně
úzkostlivě vyhýbá právním aspektům forenzně genetických
zkoumání.
Mou snahou bylo podat někdy i poměrně obtížnou problematiku co
nejsrozumitelněji, což se místy neobešlo bez určitého vědomého
zjednodušování (proto v publikaci najdete řadu příslovcí
„zpravidla“, „často“, „obvykle“ atd., odkazujících, že zde popsané
není jediné možné). Věřím, že takový postup je ku prospěchu věci.
Podrobnější informace jsou dohledatelné v odborných publikacích,

12
přičemž tipy na některé z nich najde čtenář v závěrečné Doporučené
literatuře.
Mým přáním je, aby čtenář, ač sám zcela jiného odborného
zaměření, pročítal tento breviář s porozuměním a získal díky němu
vhled do forenzní genetiky jako oboru možná v detailech složitého,
avšak jako celku nesmírně krásného a logického.
autorka
ve východních Čechách, květen 2012

13
Forenzní genetika jako obor
Co je to genetika?
Termín genetika pochází z řeckého genetíkos, odvozeného od
genesis – původ, počátek. V nejširším pojetí je genetika nauka
o dědičnosti a proměnlivosti živých organismů a o buněčných
základech této dědičnosti, tj. o dědičné informaci nesené v DNA.
Co je to forenzní?
Termín forenzní pochází z lat. forensis, odvozeného od forum –
náměstí. Na náměstí v antickém Římě se konala veřejná soudní
jednání a slovo forenzní dnes používáme ve významu „týkající se
soudu či dokazování“.
Co je to forenzní genetika?
Forenzní genetika, též označovaná jako forenzní DNA analýza, je
v nejširším pojetí věda zabývající se genetickým zkoumáním pro
potřeby dokazování a objasňování v trestních i civilních řízeních
před státními orgány, ale též pro potřeby posuzování hypotéz mimo
tato řízení – např. v rámci jiných vědních oborů nebo v soukromých
záležitostech fyzických osob.
Jaká jsou hlavní aplikace forenzní genetiky?
Základními aplikacemi forenzní genetiky jsou: kriminalistika,
identifikace a posuzování biologické příbuznosti jedinců. Tyto
směry pochopitelně nejsou striktně oddělené a často se vzájemně
prolínají.
Kriminalistická genetika zabývá genetickým zkoumáním
biologických stop a srovnávacích vzorků osob v rámci vyšetřování a
dokazování trestných činů či jiných kriminalisticky relevantních

14
událostí. Hlavním účelem těchto zkoumání zpravidla bývá určení
původce příslušných biologických stop.
Identifikační genetika se zabývá genetickou identifikací osob
živých (imigrační řízení, ověřování totožnosti atd.) či mrtvých
(nálezy neznámých mrtvol, hromadná neštěstí, oběti válečných
konfliktů v hromadných hrobech atd.).
Posuzování biologické příbuznosti jedinců zahrnuje zejména
paternitní zkoumání v případech, kdy je třeba určit otce dítěte, ale
též posuzování jiných biologických příbuzenských vztahů pro
nejrůznější účely (dědická řízení, genealogická zkoumání atd.).
Jaké další aplikace forenzní genetika má?
Forenzně genetická analýza se uplatňuje dále v chovatelství,
zemědělství a potravinářství (zkoumání pravosti rostlinných a
živočišných produktů, identifikace zvířecích jedinců, rodokmenové
studie chovných zvířat, identifikace rostlinných odrůd a živočišných
plemen atd.).
V archeologii se uplatňuje forenzní genetice velmi blízká
archeogenetika neboli analýza starodávné DNA – aDNA (angl.
ancient DNA), užívaná ke zkoumáním archeologického biologického
materiálu, jako jsou lidské i zvířecí kosterní nálezy, ale též tkáně
mumií, rostlinný materiál atd.
Rovněž některé okruhy lékařské genetiky jsou s forenzní
genetikou metodicky téměř totožné; spadají sem například
identifikace buněčných linií, průkaz ztráty heterozygozity nebo
mikrosatelitové nestability (u pacientů s nádorovým
onemocněním), průkaz monozygozity dvojčat (v případech
závažného dědičného onemocnění u jednoho z dvojčat), průkaz či
vyloučení paternity (v případě neočekávaného výsledku
popírajícího základní pravidla genetiky při nepřímé DNA

15
diagnostice v rodině postižené dědičným onemocněním) nebo
určování chromozomálních odchylek plodu analýzou plodové vody,
choriových klků či fetální krve.
Kdy poprvé byla forenzně genetická analýza použita?
Poznatky genetiky jsou v rámci forenzních zkoumání uplatňovány
již několik desítek let. Typickým příkladem je posuzování
příbuznosti osob na základě znalosti principů dědičnosti krevních
skupin. Tyto postupy však nepracují přímo s rozborem DNA, pouze
se znalostmi dědičnosti určitých znaků.
Zásadním milníkem v historii forenzní genetiky se stal převratný
objev, který učinil v roce 1984 britský vědec Alec John Jeffreys,
genetik působící na univerzitě v Leicesteru. Při studiích věnovaných
zkoumání variability genových rodin „mimochodem“ nalezl metodu,
kterou bylo možno zobrazit genetickou jedinečnost každého
člověka. Tuto metodu nazval DNA fingerprinting, tedy „genetický
otisk prstu“.
První reálný případ, v němž byl užit DNA fingerprinting
k identifikaci pachatele, byly vraždy a znásilnění Lyndy Mann
(†1983) a Dawn Ashworth (†1986) v hrabství Leicestershire.
Biologické stopy, zajištěné v obou případech, byly analyzovány roku
1987 Alecem Jeffreysem jeho metodou DNA fingerprintingu a
posléze úspěšně ztotožněny s konkrétním mužem – místním
pekařem Colinem Pitchforkem.
V našem prostředí byla forenzně genetická analýza poprvé použita
v čerstvě porevolučním Československu; v červnu 1990 byla na
dámském WC Pedagogické fakulty Masarykovy univerzity v Brně
brutálně napadena a ubodána devatenáctiletá studentka Jana
Krkošková. Na místě činu byly mimo jiné zajištěny krevní kapky,
které mohly vzniknout otřepáním poraněné ruky. Krátce nato byl

16
zadržen podezřelý – již dříve pro sexuální napadení trestaný Milan
Lubas. Průlomový znalecký posudek, který jasně ztotožnil krevní
kapky z místa činu s tímto podezřelým a naopak – krevní stopy
z jeho oděvu se zavražděnou, zpracoval Doc. RNDr. Vladimír
Ferák, CSc., vedoucí katedry genetiky a molekulární chemie
Přírodovědecké fakulty Univerzity Komenského v Bratislavě. První
expert Kriminalistického ústavu Praha a rovněž velmi významný
průkopník oboru, RNDr. Jaroslav Brouček, CSc., začal na
kriminálních případech pracovat roku 1992, kdy byl také vyškolen
u FBI.
Jak se obor změnil od dob prvních analýz?
Přestože původní Jeffreysova metoda byla ve své době zcela
převratná, měla některá omezení: především k úspěšné analýze
bylo třeba relativně velké množství biologického materiálu,
obsahujícího nedegradovanou či jen málo degradovanou DNA, a
rovněž získaná data nebylo možno jednoduše databázovat a
vzájemně ve větším objemu porovnávat.
Další vývoj tedy směřoval k práci se vzorky degradovanými a
s malým množstvím využitelné DNA, přičemž původní Jeffreysova
metoda byla zcela opuštěna a nahrazena metodami založenými na
amplifikaci, tj. zmnožování DNA ve vzorku. Tyto nové metody navíc
poskytují data snadno převoditelná do digitální podoby, a tudíž
mnohem vhodnější pro elektronické databázové zpracování.
Kam nyní forenzní genetika směřuje?
Hlavními vytýčenými současnými směry rozvoje jsou zrychlení a
automatizace analýz včetně získání výsledků v reálném čase přímo
na místě činu, práce s vysoce problematickými vzorky (včetně
snížení hranice detekčního limitu pří zachování kvality analýz) a
rozšíření potenciálu oboru na prediktivní analýzy (tj. stanovování

17
určitých tělesných charakteristik osob ze stop na místě činu).
Pozornost oboru se též obrací k metodám umožňujícím určovat a
tkáňový původ stopy molekulárně genetickými metodami a též
určovat stáří stopy.

18
Biologické stopy a biologický materiál
Obecné pojmy
Co je to biologický materiál?
Jako biologický materiál obecně označujeme vše, co bylo či je
součástí nebo produktem živého organismu. Biologický materiál tak
může mít nesmírně mnoho podob: může jím být sušená bylinná
čajová směs stejně jako prázdná motýlí kukla, ohryzek jablka, moč
v toaletě, modřínové prkno, krev na náplasti v odpadkovém koši,
koňský trus, zkumavka s virem Eboly, cukrářské droždí, hovězí
steak, kožešinový límec atd.
Co je to biologická stopa?
Biologický materiál, který je zajištěn například na místě trestného
činu a stane se předmětem znaleckého zkoumání, je označován jako
biologická stopaPřestože z výše uvedeného příkladmého seznamu
se znalec zcela jistě nejpravděpodobněji setká s krvavou náplastí,
v určitých zvláštních situacích mohou být předmětem jeho
zkoumání i ostatní uvedené materiály. Obecně tedy lze jen velmi
obtížně vymezit, jaký typ biologického materiálu může být
forenzně zkoumán. Vždy to záleží na konkrétní situaci.
Kdy by kupříkladu připadalo v úvahu forenzně biologické a genetické
zkoumání hovězího steaku? Třeba tehdy, pokud by skot v daném státě
byl zasažen určitou infekční chorobou, například kravskou chřipkou
(taková nemoc sice zatím neexistuje, ale média ji jistě dříve nebo
později objeví...). Příslušná veterinární správa v této situaci často
nařídí likvidaci zasažených chovů. Pokud by se objevilo podezření, že
někdo z chovatelů nařízení nedodržel a zvířata z porážky neskončila
v kafilérii, nýbrž na stolech nic netušících strávníků, lze v principu

19
forenzně genetickou analýzou dohledat přesný původ takového
podezřelého masa.
V praxi se velmi často setkáme s tím, že stopa je jako „biologická“
označena už kriminalistickým technikem při zajištění. To ovšem
může být někdy zavádějící: pouze a jedině znalec totiž může
určit, zda stopa má skutečně povahu biologického materiálu či
nikoli. Zatímco někdy je tato biologická povaha stopy zjevná, a to i
laikovi (např. červenohnědá hmota na dámské hygienické vložce,
mléčně bělavá vazká hmota uvnitř použitého prezervativu atd.),
jindy je přítomnost biologického materiálu ve stopě pouze
suspektní (kupříkladu nedopalek cigarety sice biologický materiál
obsahuje často, ale ne vždy; světlejší červenohnědé skvrny na oděvu
mohou a nemusejí být krví, a to dokonce i tehdy, reagují–li pozitivně
na orientační testy, o nichž budeme mluvit později; atd.).
V případu obchodování s „bílým masem“ byly postupně na různých
místech činu zajišťovány nedopalky cigaret, a to vždy několika
značek; analýza byla bezproblémová, pouze u jedné konkrétní značky
cigaret se nikdy nepodařilo izolovat detekovatelné množství DNA a
určit genetický profil osoby, což bylo zvláštní. Po vyřešení případu se
příčina vyjasnila: jedna z pachatelek nosila s oblibou dámské
rukavičky a cigarety kouřila s použitím cigaretové špičky, takže
nedošlo ke kontaktu cigarety se rty.
Tkáňový původ, druhový původ a původce
biologického materiálu
U biologických stop nás obvykle nejvíce zajímá, jaký tkáňový
původ biologický materiál má, z jakého organismu pochází (tzv.
druhový původ) a kdo je jeho konkrétním původcem.

20
Obrázek 1 – Charakteristiky, které zpravidla chceme znát u biologické stopy: tkáňový původ, druhový původ a původce.
Co je to tkáňový původ (též označovaný jako druh)
biologického materiálu?
Tkáňovým původem (druhem) biologického materiálu
rozumíme typovou podstatu materiálu – tedy zda se jedná o krev,
sperma, sliny, moč, výkaly, poševní materiál, povrchové epitelie
kůže atd. Určení se provádí především mikroskopickými,
biochemickými a imunologickými zkouškami, v relativně blízké
budoucnosti pak lze v této oblasti očekávat rovněž nástup
molekulárně genetické diagnostiky.

21
Jaký je rozdíl mezi orientační a specifickou zkouškou
tkáňového původu biologického materiálu?
Orientační a specifické zkoušky se liší svou vypovídací hodnotou,
cenou a rychlostí provedení.
Pozitivní výsledek orientační zkoušky indikuje, že by se mohlo
(ale nemusí!) jednat o daný materiál (např. pozitivní výsledek
orientačního testu přítomnosti krve znamená, že testovaná skvrna
by mohla být krev; může se však jednat i např. o ovocnou šťávu).
Orientační zkoušky bývají zpravidla levné a rychlé, s nízkou
frekvencí falešně negativních výsledků a s vyšší frekvencí falešně
pozitivních výsledků. Bývají zaměřeny na detekci přítomnosti
určitých chemických látek či enzymů, nebo jsou to zkoušky fyzikální
povahy – užití alternativních světelných zdrojů.
Naproti tomu pozitivní výsledek specifické zkoušky prokazuje, že
se s určitostí jedná o daný materiál (např. pozitivní výsledek
specifického testu přítomnosti spermatu znamená, že testovaná
skvrna je s určitostí sperma). Specifické zkoušky bývají obvykle
zaměřeny na detekci přítomnosti specifických bílkovin, hormonů a
podobně. Nutno však mít na paměti, že i nejspecifičtější zkouška
může s určitou velmi malou pravděpodobností poskytovat falešně
pozitivní výsledek.
Kdy má smysl tkáňový původ biologického materiálu
stanovovat?
Stanovení tkáňového původu materiálu je důležité zejména
tehdy, kdy nám tato informace poslouží k potvrzení či
vyvrácení našich hypotéz.

22
Zatímco v některých případech je typ biologického materiálu
irelevantní, jindy může zásadně změnit pohled na hodnotu DNA
profilování a důkazu jako takového.
V případu znásilnění byly ke zkoumání předloženy dvě věcné stopy:
domácí telefon a parafínová svíce. Poškozená uváděla, že pachatel jí
do pochvy nejprve zasouval svíčku, poté na ní vykonal soulož
s ejakulací do pochvy a nakonec jí do pochvy zasouval sluchátko
domácího telefonu. Obviněný toto popíral. Biologickou a genetickou
analýzou byla na svíci zjištěna přítomnost většího množství poševních
epitelií poškozené, na sluchátku telefonu byla zjištěna přítomnost
směsi poševních epitelií poškozené a spermatu obviněného. Tato
zjištění významně podpořila pravdivost výpovědi poškozené. Bez
stanovení tkáňového původu biologického materiálu by v tomto
případě důkazní hodnota stop značně klesla (samotné genetické
zkoumání by se omezilo na konstatování, že na svíci se nachází
neurčený biologický materiál poškozené a na sluchátku směs
neurčených biologických materiálů poškozené a obviněného, kteří oba
mohli telefon používat k jeho původnímu účelu).
Kdy nemá smysl tkáňový původ biologického
materiálu stanovovat?
Zatímco ve výše uvedeném příkladu je určení tkáňového původu
biologického materiálu zcela zásadní, jindy je tato analýza
nadbytečná až zbytečná a někdy může být i ke škodě věci.
Především u stop s předpokládaným malým až velmi malým
obsahem biologického materiálu je vhodné vždy pečlivě uvážit,
jak moc je stanovení tkáňového původu materiálu přínosné a
potřebné, neboť toto stanovení téměř vždy znamená odebrání
určitého množství materiálu ze stopy.
Typickou situací je požadavek na průkaz slin a lidské bílkoviny
v nedopalku spojený s požadavkem na stanovení genetického

23
profilu osoby. Poměrně často totiž po provedení zkoušky na
přítomnost slin a lidské bílkoviny již pro genetickou analýzu zbývá
tak málo materiálu, že se nepodaří požadovaný genetický profil
stanovit v potřebné kvalitě. Přitom detekce slin a lidské bílkoviny je
zde ve své podstatě bezpředmětná, jelikož sama o sobě nemá
žádnou důkazní hodnotu: pokud se v nedopalku nachází biologický
materiál, pak to v drtivé většině případů jsou lidské sliny a epitelie
ústní sliznice či pokožky rtů, případně s malou příměsí krve (např.
při zánětu dásní, oparu atd.); s případem, kdy by se v nedopalku
nacházelo například prasečí sperma či kočičí moč, se – pokud je mi
známo – žádný znalec dosud nesetkal.
Dříve tyto testy mohly najít své opodstatnění jako jakési “síto“
k oddělení stop nepoužitelných pro genetické zkoumání (kupříkladu
u zmíněných nedopalků nepřítomnost slin a lidské bílkoviny
indikovala nemožnost stanovit genetický profil). V dnešní době je už
tento přístup ale překonán, neboť jednak forenzní genetika
disponuje mnohem přesnějšími metodami posouzení obsahu
využitelného biologického materiálu ve stopě, jednak i z nedopalků
s negativním výsledkem testu na přítomnost slin lze někdy stanovit
relevantní genetický profil.
Co je to určení druhového původu a průkaz lidského
původu?
Určení druhového původu materiálu, tedy zjištění, z jakého
rostlinného či živočišného druhu stopa pochází, hraje (na rozdíl od
určení druhu biologického materiálu) obvykle roli zásadní, přičemž
nejčastěji jde o průkaz lidského původu daného materiálu. Lze jej
provést samostatně imunologickými metodami (reakcí lidské
bílkoviny se specifickou protilátkou), častěji ale tento průkaz
lidského původu vyplyne „samovolně, jako vedlejší produkt“
z výsledku genetického zkoumání. Většina forenzně genetických
testů užívaných k analýzám lidského biologického materiálu totiž

24
poskytuje standardní pozitivní výsledek právě jen a pouze
u lidského materiálu – říkáme, že tyto testy jsou druhově
specifické. Tedy: poskytne–li analýza stopy pozitivní výsledek,
lze zpětně konstatovat, že stopa obsahovala s jistotou lidský
biologický materiál. Z tohoto důvodu se od imunologického
průkazu lidské bílkoviny většinou upouští, a to zejména u stop
s předpokládaným malým či velmi malým množstvím využitelného
biologického materiálu (důvody jsou obdobné jako u již
zmiňovaného určování druhu biologického materiálu).
Na policii se dostavila žena, živící se jako prostitutka, a vypověděla, že
předchozího dne měla konflikt se zákazníkem, který vyvrcholil tím, že
muž jí začal vyhrožovat zabitím. Jako důkaz, že svá slova míní vážně,
ji odvedl ke kufru auta a ukázal jí v něm ležící mrtvolu muže. Poté
odjel. Podle popisu byl tento zákazník v relativně krátké době nalezen,
přičemž v kufru jeho auta byly skutečně zjištěny krevní otěry.
Podezřelý se hájil tím, že se jedná o krev zvířecí, jelikož je členem
mysliveckého svazu. Jak imunologická, tak genetická analýza však
toto tvrzení jednoznačně vyvrátily – bylo potvrzeno, že se jedná o krev
lidskou, a genetickou analýzou byl stanoven genetický profil osoby
mužského pohlaví. Podezřelý k věci odmítl dále vypovídat. Pokud je mi
známo, mrtvola muže nebyla dosud nalezena a nebyla ani zjištěna
jeho možná totožnost.
Lze prokázat i jiný než lidský původ?
Ve zvláštních případech je třeba specificky prokázat i jiný než lidský
původ – například psí, králičí atd. Zde je nejčastěji využíváno
imunologické analýzy, založené na reakci s určitým specifickým
sérem. Genetické analýzy se v tomto směru v České republice
využívají spíše omezeně, nicméně v principu platí totéž, co bylo
řečeno u průkazu lidského původu: pozitivní výsledek určitého
druhově specifického genetického zkoumání je současně
potvrzením, že stopa byla tvořena biologickým materiálem daného

25
druhu. Genetickým analýzám jiného než lidského biologického
materiálu je věnována samostatná kapitola na konci tohoto
breviáře.
Co je to určení původce biologické stopy?
Pomyslnou nejvyšší metou, jíž lze forenzně genetickým zkoumáním
dosáhnout, je určení zdroje, původce stopy, tedy konkrétního
jedince, jehož biologickým materiálem je stopa tvořena – tedy např.
Karel Novák, r.č. 123456/7890, pes Haryk majitele Františka Flinty
atd. Jednoznačné určení původce stopy nazýváme individuální
identifikace.
Využitelnost biologického materiálu pro forenzně
genetickou analýzu
Jaký biologický materiál je pro forenzně genetickou
expertízu vhodný?
Obecně je forenzní analýza úspěšně aplikovatelná na jakýkoli
materiál, obsahující DNA v množství a kvalitě dostatečné k jejímu
provedení, což je, přiznejme si, definice kruhem (bohužel však
jediná vystihující podstatu věci). Úsloví, pocházející dle Oxford
Dictionary of Citations snad už z počátku 14. století, říká: „Zda je
puding k jídlu, to se pozná při jedení!“ („The proof of the pudding is
in the eating!“) Stejně tak my můžeme říci: „Zda je materiál
analyzovatelný, to se pozná při analýze!“
Je nějaký zásadní rozdíl ve využitelnosti biologických
materiálů analyzovaných v rámci jednotlivých
aplikací forenzní genetiky?
V praxi je velmi podstatný principiální rozdíl mezi analýzami
biologických stop a analýzami řízeně odebraných vzorků osob

26
(např. při určování příbuznosti). Kriminalistické analýzy
biologických stop jsou charakteristické tím, že znalec nemá žádnou
možnost volby, výběru biologického materiálu, musí pracovat s tím,
co je; navíc se jedná o vzorky často velice nestandardní co do
kvantity i kvality obsažené DNA. Stejně problematické mohou být i
materiály z těl vysokého stupně rozkladu či kosterních nálezů.
Naproti tomu řízené odběry srovnávacích vzorků umožňují obvykle
vybrat nejvhodnější materiál pro danou expertízu, zaručující
dostatečné množství kvalitní DNA pro její provedení.
Jaké materiály jsou zajišťovány jako srovnávací
vzorky osob?
V současnosti nejběžnějším srovnávacím vzorkem biologického
materiálu je bukální stěr, což je otěr sliznice vnitřní strany tváře
(tedy v ústech) na odběrový tampón či kartáček. Odběr je rychlý,
nebolestivý a neintimní, může ho provádět osoba sama či libovolný
k tomu pověřený pracovník, navíc odebraný materiál se následně
velmi dobře a nenáročně uchovává, a to i po dlouhou dobu.
Dříve běžně odebíraným, dnes již spíše opouštěným srovnávacím
vzorkem je periferní žilní krev, zajišťovaná nejčastěji klasicky
odběrem ze žíly v loketní jamce, případně u novorozenců a kojenců
ze žíly na hlavičce (stejně, jako při běžných zdravotnických
vyšetřeních). Odběr je rovněž rychlý, avšak spojený s jistou dávkou
diskomfortu (bolest, nepříjemné pocity) a za současného narušení
integrity těla včetně nenulového (byť nepatrného) rizika infekce,
musí ho provádět zdravotnický personál a vzorky se o něco
náročněji uchovávají. Pro forenzně genetickou analýzu mohou být
principiálně použity i krevní vzorky zajištěné pro jiný účel (např. ke
zdravotnickému vyšetření), záleží však na způsobu zajištění,
zejména na použitém protisrážlivém médiu, a též na způsobu
nakládání se vzorkem před předáním k forenzní analýze. Na tom lze

27
demonstrovat, jak neostré jsou hranice mezi zdravotnickými a
forenzními aplikacemi.
Jednoho vlahého večera zastavila policie mladého řidiče a po kontrole
dokladů jej podrobila dechové zkoušce na alkohol. Vzhledem k tomu,
že test byl pozitivní, byl řidič převezen do zdravotnického zařízení
na odběr krve, který původní podezření jednoznačně potvrdil. Kauza
se tak dostala do roviny trestní, ovšem majitel osobního dokladu pan
Karel před soudem argumentoval, že to nebyl on, kdo řídil, a že
na místě trestního činu v té době vůbec nebyl. Byl proto ustanoven
soudní znalec v oboru DNA analýzy, jehož úkolem bylo pouze
odpovědět na v podstatě jednoduchou otázku – byl pan Karel opravdu
oním řidičem, kterému byla odebrána krev na alkohol? Po porovnání
profilu pana Karla a profilu z testované krve bylo již na první pohled
jasné, že pan Karel oním řidičem nebyl, neboť dané profily se
jednoznačně lišily. Tím vše mohlo skončit a kauza mohla být
uzavřena. Pozorný znalec si však všimnul, že ve velké části
testovaných lokusů měl pan Karel s neznámým řidičem jednu alelu
shodnou, což napovídalo hypotéze o existenci biologického
příbuzenského vztahu. DNA obou byla proto navíc ještě přetestována
na haplotyp Y chromozómu, neboť pokud by muži byli příbuzní
po paternální linii, musel by být haplotyp Y chromozomu naprosto
shodný, což se následně opravdu potvrdilo. Neznámý však nemohl být
otec pana Karla, neboť ve třech autozomálních lokusech byl jako jeho
otec vyloučen (vzorky neměly společné alely). Jako
nejpravděpodobnější byla znalcem označena hypotéza, že se jedná
o bratry, čemuž by napovídal i podobný věk a podoba neznámého. Jak
jest patrno, vzorek původně odebraný pro toxikologické vyšetření byl
následně použit k identifikaci (tedy v oblasti forenzní analýzy) a ještě
později se zkoumání přesunulo do sféry určení biologické příbuznosti.
Ve zvláštních případech mohou být zajišťovány i jiné typy
srovnávacích vzorků živých osob, například odstřižené nehty či

28
kožní seškraby; děje se tak například při genetických výzkumech
lidských přírodních populací, u nichž by se odběr jiného typu
vzorku nesetkal s porozuměním.
Pro vyšetření dosud nenarozených dětí (embryí či plodů) může být
použita plodová voda odebraná amniocentézou (AMC), choriové
klky odebrané choriocentézou (CVS) či pupečníková krev plodu
odebraná kordocentézou (CC); některé genetické testy mohou být
provedeny i analýzou fetální DNA, která je přítomna v krvi matky.
Zde je třeba důrazně upozornit, že odběr plodové vody,
choriových klků i pupečníkové krve jsou invazivní metody
spojené s nezanedbatelným rizikem pro plod (udávané riziko
potratu po odběru je cca 0,5–1%) a mohou být provedeny pouze
ze zdravotních důvodů k potvrzení či vyloučení závažné genetické
odchylky plodu (např. Downův syndrom). Zbytková část materiálu
z takového odběru může být použita pro forenzní účely (např.
určení otcovství), pro čistě forenzní účel však nelze tento odběr
z etických důvodů nařídit, požadovat ani doporučovat (s
analýzou je potom třeba počkat až po narození dítěte).
Vyvraťme zde jeden hluboce zakořeněný mýtus, který stále přetrvává,
totiž tvrzení, že paternitní analýzu lze provést až v jednom roce věku
dítěte. Pro to neexistují žádné odborné ani právní důvody. Analýza je
technicky proveditelná již v okamžiku narození dítěte a – jak se lze
dočíst v předchozím odstavci – v případě, že je dostupný materiál
plodu, pak už v průběhu těhotenství. Komerční novinkou jsou dokonce
paternitní testy plodu z krve matky, které je možno provést již od cca
9. – 10. týdne těhotenství.
V případě potratu (spontánního nebo při umělém ukončení
těhotenství) může být k analýze použit plodový materiál zajištěný
z dutiny děložní; zde je třeba upozornit, že u velmi raných fází
těhotenství je velikost embrya velmi malá (v 5. týdnu těhotenství
cca 4–5 mm) a rozpoznání embryonální tkáně ve směsi s dalším

29
odsátým materiálem děložní sliznice může být dosti problematické.
V pozdějších stádiích již rozpoznání embrya problém nečiní.
U mrtvých osob je škála srovnávacích vzorků mnohem pestřejší:
dle stavu rozkladu těla mohou být odebrány krev, svalová tkáň,
kůže, nehty, zuby, tkáň některých orgánů nebo kost. Tyto srovnávací
vzorky mohou být zejména u těl ve vyšším stupni rozkladu kvůli
degradaci již poměrně náročné na získání analyzovatelné DNA, a
tedy se často svým charakterem blíží spíše stopám.
U pohřešovaných či již dříve zemřelých osob jsou za srovnávací
vzorky považovány materiály nejrůznější povahy, které lze s velkou
pravděpodobností osobě přisoudit, například materiál v jejím
zubním kartáčku, materiál z jejích osobních předmětů či materiál
zajištěný a deponovaný pro zdravotní účely (parafínem fixované
tkáně z biopsií, mikroskopické preparáty atd.). Všechny tyto vzorky
jsou co do kvality a kvantity DNA opět spíše povahy stop, a tak je
s nimi také nakládáno.
Nevhodnými srovnávacími vzorky jsou odstřižené či vypadlé vlasy a
chlupy, které obsahují jen velmi malá množství jaderné DNA.
Nakládání s biologickými stopami
Biologické stopy mohou být velmi cennými, často i rozhodujícími
důkazy o vině či nevině. Aby takto mohly posloužit, je ale nezbytné
s nimi správně nakládat, tedy je třeba dodržovat určité postupy a
zásady, popsané níže. Nevhodné zajištění, nesprávné uchovávání,
neopatrná manipulace – to vše může stopu částečně nebo úplně
znehodnotit nebo v horším případě dokonce změnit tak, že
genetická analýza poskytne zcela falešný výsledek.

30
Jak se vyhledávají stopy?
Lakonicky řečeno: aby mohla stopa posloužit jako důkaz, je
třeba ji nejprve najít. To se zdá být samozřejmé, avšak ve
skutečnosti to někdy může představovat zdlouhavou a vyčerpávající
činnost. Stopy můžeme z tohoto hlediska rozdělit do několika
kategorií:
1. stopy markantní, viditelné okem i při zběžném pohledu
2. stopy viditelné okem při podrobném ohledání
3. stopy okem neviditelné, avšak detekovatelné zvláštními
testy
4. stopy zcela latentní
Do první kategorie spadají nepřehlédnutelné stopy, jako jsou
tratoliště krve a další velké krevní stopy, velké skvrny spermatu (na
oděvu, ložním prádle apod.), lejno atd. Vyhledání těchto stop nečiní
potíž ani naprostému laikovi a stopy lze obvykle snadno zajistit.
Do druhé kategorie lze zařadit drobné biologické stopy, jako jsou
kupříkladu krevní kapky a otěry, zaschlý nosní sekret, vlasy, stopy
slin na skleničkách atd. Kvůli malé velikosti může být jejich
vyhledání někdy obtížnější, avšak stále jsou zjistitelné pouhým
okem.
Třetí kategorii tvoří stopy „neviditelné“, které ale lze odhalit
s použitím speciálních pomůcek či postupů. Těmi mohou být
například monochromatické forenzní světelné zdroje (forenzní
„baterky“), které dle použité vlnové délky pomohou zviditelnit
určitý konkrétní druh biologického materiálu. Obecně známé pak je
vyhledávání velmi zředěných (a tudíž okem neviditelných) krevních
stop pomocí chemických činidel světélkujících při kontaktu s krví
(např. Luminol, FastBlue).

31
Latentní stopy, tvořící čtvrtou kategorii, nemohou být detekovány
ani okem, a prozatím ani pomocí screeningových testů. Typickým
příkladem takové stopy jsou povrchové kožní buňky ulpivší na
telefonním sluchátku. Tyto stopy se tudíž zajišťují „naslepo“: jsou
odebírány v místech, kde výskyt biologického materiálu
předpokládáme. Úspěšnost při jejich zajišťování tak závisí
především na dobrém odhadu a zkušenostech toho, kdo je zajišťuje.
Často ke zkoumání předkládanou věcnou stopou bývá pokrývka hlavy,
kterou pachatel ztratil na útěku. Zkušený znalec odebírá stěry najisto
u okraje v oblasti záuší, kde pokrývka nejtěsněji přiléhá k hlavě a při
nošení dochází k drobným pohybům, při nichž se odírají buňky
pokožky.
Velmi cenným vodítkem při vyhledávání stop mohou být i
výpovědi aktérů události.
Oběť znásilnění, ke kterému došlo v lednu v noci za opuštěnou
vlakovou zastávkou, vypověděla, že pachatel ejakuloval volně mimo
její pochvu; podle přesného popisu míst, kde se při tom oběť a
pachatel nacházeli, se kriminalistickému technikovi podařilo najít ve
sněhu malou proláklinu, která mohla být způsobena dopadem teplého
spermatu. Stopu proto po konzultaci se znalcem zajistil uválením do
tvaru sněhové koule o průměru cca 30 cm a takto byla
transportována do laboratoře. Ve stopě byla po rozpuštění prokázána
přítomnost spermatu a byl stanoven genetický profil, který byl posléze
ztotožněn s natipovaným podezřelým. Bez použití informací
z výpovědi poškozené by tato stopa zřejmě nebyla nikdy nalezena.
Jaká jsou pravidla zajišťování stop?
Zajišťování biologických stop pro forenzně genetické
zkoumání je nezbytné provádět tak, aby:

32
1. nedošlo k degradaci stopy (zničení biologického
materiálu)
2. bylo odebráno co možná největší množství biologického
materiálu (toto neplatí jen u rozsáhlých stop, jako jsou
například krevní tratoliště)
3. nedošlo ke kontaminaci stopy (vnesení cizorodého
biologického materiálu).
Jak zabránit degradaci stopy?
Poškození či úplné zničení biologického materiálu, spojené
s poškozením v něm přítomné DNA – tzv. degradací DNA, hrozí
tehdy, když stopu vystavíme nevhodným fyzikálním, chemickým či
biologickým vlivům.
Obrázek 2 – Řada přírodních činitelů může způsobit nevratné poškození molekul DNA ve stopě; tento proces označujeme jako degradaci DNA.

33
Blíže se o tom zmíníme později, zde pouze stručně: rozkladné
procesy jsou urychlovány vysokou teplotou a vlhkem, jež podporují
rychlý růst bakterií; nevratné chemické změny v dvoušroubovici
DNA způsobuje UV záření – tedy např. přímo dopadající sluneční
paprsky, stejně jako působení některých chemikálií. Degradaci DNA
lze zabránit pouze vytvořením správných předpisů týkajících se
zajišťování stop a následným dodržením v nich uvedených postupů.
Jak zajistit dostatek materiálu ze stopy?
Dostatečné množství zajištěného biologického materiálu zvyšuje
naději na pozitivní výsledek analýzy. Je důležité i proto, že někdy se
objeví potřeba stopu reanalyzovat (například při požadavku na
vypracování revizního posudku – superposudku). Nejvhodnější je
vždy dopravit stopu do laboratoře i s podkladem či přímo
předmětem, na němž se nachází. V některých případech to ovšem
není technicky možné (stopy na velmi rozměrných předmětech,
stopy na těle atd.) – potom musí odběr biologického materiálu
provést na místě kriminalistický technik nebo musí být znalec
povolán na místo činu. V praxi je většina latentních (dotykových
stop) zajišťována na místě trestného činu stěrem místa
s předpokládaným výskytem biologického materiálu.
Pro zajištění kriminalistických stop již existují specializované
pomůcky, které mimo jiné zaručují, že ze stopy bude odebráno velké
množství materiálu. Kupříkladu speciální forenzní nylonové
mikrokartáčky, jimiž se provádějí stěry, dokážou materiál zachytit
mnohonásobně efektivněji než běžné vatové tampóny. To má
význam zejména u latentních stop minimálních rozměrů. Nutno
ovšem říci, že pořizovací cena těchto pomůcek bývá zpravidla také
mnohonásobně vyšší, a jejich plošné používání tak v praxi naráží na
nedostatek financí ve státním sektoru. Jak bude na příkladu ukázáno
níže, někdy se šetření projeví jako nemístné škudlilství.
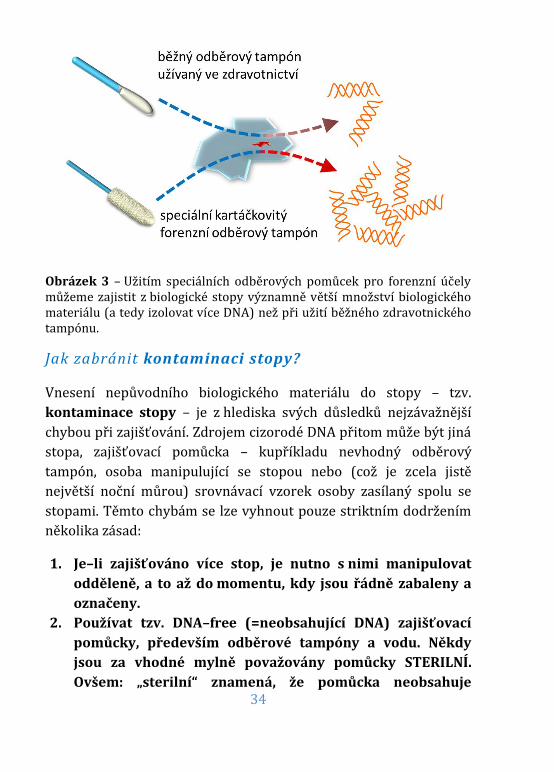
34
Obrázek 3 – Užitím speciálních odběrových pomůcek pro forenzní účely můžeme zajistit z biologické stopy významně větší množství biologického materiálu (a tedy izolovat více DNA) než při užití běžného zdravotnického tampónu.
Jak zabránit kontaminaci stopy?
Vnesení nepůvodního biologického materiálu do stopy – tzv.
kontaminace stopy – je z hlediska svých důsledků nejzávažnější
chybou při zajišťování. Zdrojem cizorodé DNA přitom může být jiná
stopa, zajišťovací pomůcka – kupříkladu nevhodný odběrový
tampón, osoba manipulující se stopou nebo (což je zcela jistě
největší noční můrou) srovnávací vzorek osoby zasílaný spolu se
stopami. Těmto chybám se lze vyhnout pouze striktním dodržením
několika zásad:
1. Je–li zajišťováno více stop, je nutno s nimi manipulovat
odděleně, a to až do momentu, kdy jsou řádně zabaleny a
označeny.
2. Používat tzv. DNA–free (=neobsahující DNA) zajišťovací
pomůcky, především odběrové tampóny a vodu. Někdy
jsou za vhodné mylně považovány pomůcky STERILNÍ.
Ovšem: „sterilní“ znamená, že pomůcka neobsahuje

35
infekční agens (živé bakterie, viry, plísně, kvasinky apod.).
Pro odběr stop je ale třeba, aby pomůcka neobsahovala
žádnou DNA, zejména ne lidskou.
Němečtí kriminalisté se jen pomalu a obtížně vzpamatovávají
z fatálního omylu, který proslul jako „Fantom z Heilbronnu“. Od roku
1993 zajistili v celkem 40 trestných činech včetně šesti vražd
biologické stopy vykazující jeden a týž genetický profil, náležející
neznámé ženě. Sekvenací mitochondriální DNA a analýzou evolučního
původu bylo určeno, že tato žena–monstrum může velmi
pravděpodobně pocházet z východní Evropy. Skutky páchala na území
celého Německa, dokonce i v Rakousku a Francii, nikdy však
v Bavorsku. Policie po mnoha letech neúspěšného pátrání dokonce
vypsala odměnu 300 tisíc euro za informace vedoucí k dopadení této
pachatelky. Obrat nastal, když byl její genetický profil zjištěn i na
spáleném těle prokazatelně mužského žadatele o azyl ve Francii.
Vzniklo podezření, že DNA tajemné ženy se do odběrových tampónů
dostala již během jejich výroby. Všechny tampóny použité v případech
připisovaných Fantomovi totiž pocházely od jediné firmy, rakouské
Greiner Bio–One International AG, a byly určeny pro užití
ve zdravotnictví. Proto byly po zabalení sterilizovány, což v nich sice
zahubilo všechny bakterie, ovšem neodstranilo DNA. Kvůli příznivé
ceně byly ve velkém nakupovány i policejními laboratořemi, které je
používaly k odběrům stop na místě činu. Pouze bavorská policie
užívala tampóny od jiného dodavatele, a tak se v Bavorsku Fantom
nikdy neobjevil. Greiner Bio–One zaměstnával ve výrobě i řadu dělnic
původem z východní Evropy. Tato rozsáhlá blamáž konečně hlasitě
upozornila kriminalisty v řadě zemí, že investování do speciálních
pomůcek určených přímo pro forenzní odběry má své opodstatnění.
3. Osoby zajišťující stopy musí používat dostatečné osobní
ochranné pomůcky – rukavice, roušky, případně celotělové

36
overaly. Kýchající, kouřící, pijící či dokonce stravující se
osoba nemá na místě činu co dělat.
Nebezpečí kontaminace stop technikem na místě činu je často
podceňováno. Přitom některé v zahraničí provedené studie jasně
ukazují, že i při používání ochranných pomůcek dochází v nemalém
procentu k nechtěné kontaminaci. Z technického hlediska
nejvhodnější cestou je proto jednak povinná výměna rukavic mezi
sběrem jednotlivých stop a vytvoření databáze genetických profilů
techniků a dalších osob manipulujících se stopami, aby takováto
kontaminace mohla být včas rozpoznána.
Obrázek 4 – Možnými zdroji kontaminace mohou být jak jiné stopy, tak i odběrové pomůcky či osoby manipulující se stopami. Proti těmto všem zdrojům je třeba se účinně bránit.

37
4. Srovnávací vzorky osob je naprosto nezbytné zajišťovat
odděleně od ohledání místa činu, nejlépe jinde a s časovým
odstupem.
Jak dlouho je možné stopy uchovávat?
Čím dříve jsou stopy předány ke zpracování do laboratoře, tím lépe.
I u té nejlépe zajištěné stopy není vyloučena pozvolná degradace,
proto je včasná analýza jedinou zárukou, že stopa nebude
znehodnocena ještě před získáním potřebných dat. Týká se to
především stop, u nichž degradace započala už před zajištěním. Na
druhou stranu, řada stop si uchovává biologický materiál
v analyzovatelném stavu i po mnoho desítek let. Takto stabilní
bývají obvykle například zaschlé materiály (krev, sliny, sperma).
Vlastnosti a chování biologického materiálu ve
stopách
Chceme–li si na základě výsledků genetické analýzy udělat obrázek
o dění na místě činu, je nezbytné mít alespoň základní představu
o vlastnostech a chování biologického materiálu jako takového.
V této oblasti panuje řada mylných domněnek, uveďme je tedy nyní
na pravou míru!
Kde se biologický materiál ve stopách bere?
Lidské tělo je dynamický systém – pohybujeme se, dýcháme, jíme a
vylučujeme, naše buňky se stále dělí, nové vznikají a staré umírají.
To vše je spojeno s produkcí velkého množství biologického
materiálu, z nějž část naše tělo opouští a dostává se do jeho bližšího
či vzdálenějšího okolí. Mohou to být:

38
1. přirozeně odcházející tělní tekutiny, sekrety a exkrementy
– menstruační krev, moč, stolice, pot, slzy, nosní sekret, sperma,
sliny, zvratky
2. přirozeně odcházející buňky kůže a kožní deriváty – vlasy,
chlupy, nehty, odumřelé kožní epitelie (povrchové buňky)
3. přirozeně odcházející buňky sliznice – poševní epitelie
(např. při pohlavním styku), povrchové epitelie pohlavního údu
(při pohlavním styku či jiné manipulaci s ním), epitelie dutiny
ústní a krku (při ústní hygieně, jídle, líbání, orálním styku
apod.), epitelie trávicího traktu (např. sliznice střeva
uvolňovaná do stolice), epitelie dýchacích cest (při kašli,
smrkání, kýchání, dýchání, řeči), epitelie močových cest (při
zánětu uvolňované do moči) atd.
4. materiály uvolňované nepřirozeně, a to v důsledku
nemoci, zranění, lékařského zákroku – krev, kůže jako celek,
svalová tkáň, tuková tkáň, při velmi hrubém poranění (včetně
chirurgického úkonu) i jakékoli další tkáně (mozková tkáň,
kost, tkáně orgánů dutiny břišní atd.).
Lze určit, jaký byl mechanismus vzniku stopy?
Rozhodnout o tom, jak stopa vznikla, bývá často velmi obtížné.
Forenzní věda se o takové zkoumání pokouší alespoň u krevních
stop; podle jejich tvaru a rozmístění lze někdy usuzovat, zda vznikly
otěrem, odkapáváním, stříkáním (např. po úderu) atd. Tyto analýzy
však nespadají do oboru forenzní genetiky, nýbrž do oboru
biomechaniky, a nebudeme je tedy blíže rozvádět. U ostatních druhů
biologického materiálu se „analýza“ mechanismu vzniku stopy
pohybuje obvykle už čistě v rovině spekulací (neexistují relevantní
studie, ze kterých by bylo možno vycházet).
V případu pohlavního zneužívání byly předloženy lůžkoviny z gauče,
na němž ke skutku mělo dojít, a byl mimo jiné vznesen tento
požadavek: „Na základě tvaru a rozmístění skvrn (spermatu a

39
poševního sekretu) určete, v jaké sexuální poloze k pohlavnímu styku
došlo.“ Relevantně, bez srovnávacích pokusů a jisté dávky spekulací na
takovou otázku prakticky nelze odpovědět. Samozřejmě každý
dospělý normálně sexuálně aktivní člověk si v principu umí představit,
kde přibližně se mohou uvedené biologické materiály při styku v té
které poloze ocitnout, nicméně taková představa může být i velmi
ošidná (třeba proto, že nepostihneme všechny aspekty, které mohly
mít v daný moment vliv), a nelze ji proto zaměňovat za odborně
relevantní znalecké posouzení.
Lze z množství biologického materiálu ve stopě
usuzovat na nějaké okolnosti skutku?
Souvislost mezi množstvím biologického materiálu ve stopě a
mechanismem jejího vzniku sice existuje, ale jen ve velmi hrubém
měřítku. Tedy: pokud je v bytě nalezeno tratoliště krve o metrovém
průměru a majitelka tvrdí, že se tam manžel řízl při holení, je její
tvrzení přinejmenším krajně podezřelé. Pokud ale jsou nalezeny
drobné krevní kapky a otěry, může se jednat jak o stopy po říznutí
při holení, tak o stopy po brutálním útoku řeznickým nožem. Jinak
řečeno, nález extrémně velkého množství biologického
materiálu vylučuje vznik stopy při drobné události, ale nález
malého množství biologického materiálu nevylučuje vznik
stopy při „velké“ události, například násilném útoku.
Často je tento problém diskutován při nálezu biologického
materiálu osoby A za nehty osoby B, neboť vždy vyvstává otázka,
zda je takový nález důkazem násilného kontaktu nebo ne. Obecně
lze říci, že u osob, které spolu nežijí, nepracují nebo nejsou jinak
v blízkém kontaktu, se zpravidla navzájem biologický materiál za
nehty nevyskytuje vůbec nebo jen v nepatrném měřítku (např. zcela
miniaturní množství přenesená v důsledku podání ruky, použití
stejného madla v tramvaji atd.). Naproti tomu u osob, které
v blízkém kontaktu jsou, může být množství biologického materiálu

40
za nehty toho druhého značné. V normálně fungující společně žijící
rodině tak běžně má žena za nehty materiál manžela i dětí, dítě
materiál sourozence i rodičů atd. Z toho vyplývá důležitý fakt:
u osob žijících společně nebo jsoucích v úzkém kontaktu nelze
nález biologického materiálu osoby A za nehty osoby B
považovat za důkaz násilného kontaktu. Stejně tak nelze velké
množství materiálu vyloučit tam, kde došlo (třeba i jednorázově)
k intenzivnímu fyzickému kontaktu, tedy např. mezi masérem a
masírovaným, prostitutkou a klientem, zdravotníkem a pacientem
apod.
Jak se stopa mění od okamžiku svého vzniku do
zajištění a analýzy?
Biologický materiál může být tvořen živými či mrtvými buňkami,
jejich zbytky a/nebo organickými, případně anorganickými látkami
produkovanými živými organismy. Velká část těchto komponentů
ihned po opuštění těla snadno podléhá nejrůznějším typům
rozkladných procesů, které souhrnně označujeme jako
biodegradace. (Jak takový proces ve své vrcholné fázi vypadá, to si
umí představit každý, kdo uprostřed července zapomněl v kufru
auta dva dny na parkovišti nákupní tašku s půlkilem vepřových
jater.) Rychlost biodegradace a její podoba závisejí především na
podmínkách, v nichž se stopa nachází.
Z hlediska genetického zkoumání je ovšem podstatné především to,
v jaké míře je ve stopě degradována DNA. Ta může být zasažena
několika typy vlivů:
1. fyzikálními činiteli – UV zářením, radioaktivním zářením,
extrémně vysokou teplotou
2. chemickými činiteli – oxidačními činidly, silnými kyselinami a
zásadami

41
3. biologickými činiteli – bakteriemi, plísněmi, ale i enzymatickou
sebedegradací (autolýzou)
a jejich kombinacemi. V praxi se zdaleka nejčastěji setkáváme
s bakteriální degradací stopy: vlhko a teplo (20–40°C) jsou
podmínkami optimálními pro růst bakterií a stopa může být v tomto
prostředí degradována velmi rychle.
V parku byla kolem desáté hodiny večerní cestou domů z jednání
brutálně napadena mladá žena: pachatel jí opakovaně udeřil tyčí do
zátylku a oblasti krku a zad, takže jí způsobil četná silně krvácející
poranění, poté jí odcizil kabelku a mobilní telefon a z místa činu utekl.
Celou noc drobně pršelo. Těžce zraněná žena byla nalezena pejskařem
až brzy ráno; muž zavolal záchrannou službu, ta po příjezdu při
poskytování pomoci ženu svlékla ze zakrváceného a deštěm
promočeného oděvu a tento uložila do igelitového pytle. Pytel byl
policii předán až o dva dny později: rozsáhlé krevní skvrny na oděvu
již byly hnědozelené barvy, silně zapáchající, a analýza neprokázala
v žádné ze skvrn přítomnost analyzovatelné DNA. Za daných okolností
tedy nejenže nebylo reálné najít na oděvu stopy DNA pachatele, ale
nebylo již možno – kvůli pokročilé bakteriální degradaci – stanovit
ani DNA oběti. (Pachatel byl přesto vypátrán a usvědčen díky stopám
na tyči, kterou odhodil poblíž místa činu; žena útok přežila, zůstává
však trvale ochrnutá).
Dalším z důležitých faktorů, které rozhodují o tom, zda bude stopa
později snadno či hůře analyzovatelná, případně zcela
neanalyzovatelná, je přítomnost tzv. inhibitorů. Souhrnně tak
označujeme látky, které negativně ovlivňují průběh dalších analýz,
zejména blokují namnožení vybraných úseků DNA metodou
označovanou jako PCR (polymerázová řetězová reakce), a bude
o nich pojednáno v části věnované technologii.

42
Moderní forenzně genetická laboratoř ovšem již disponuje jednak
metodami na zjištění přítomnosti inhibitorů ve vzorku, jednak
izolačními postupy schopnými ve většině případů tyto inhibitory
účinně odstranit.
Muž v době nepřítomnosti své manželky brutálně znásilnil a zavraždil
své dvě nevlastní dcery (14 a 16 let), poté jejich krví popsal zeď
v domě. Vzorky odebrané z této vápnem vybílené zdi obsahovaly
extrémně vysoká množství vápenatých iontů a analyzovat se je
podařilo až po dlouhé dekalcifikaci pomocí chelatačního činidla
(činidlo vyvazující dvojmocné ionty).
Existuje nějaká časová hranice , kdy už stopu s jistotou
nelze analyzovat?
Žádná pevně definovatelná hranice neexistuje. Za určitých
podmínek – jak už bylo řečeno – může stopa podlehnout zkáze
velmi rychle, pokud se ale dostane do stabilního prostředí, může zde
vydržet v dobrém stavu „celou věčnost“, což dokládají kupříkladu
pozitivní analýzy z některých historických či archeologických nálezů
včetně analýz mumií. Je zřejmé, že pro potřeby forenzní nám
postačuje mnohem kratší časový úsek – řádově maximálně desítky
let. Správně zajištěné a uchovávané stopy tak umožňují znovu
otevřít některé sporné případy a posoudit je na základě nových
analýz.
Lze určit, kdy biologická stopa vznikla?
Určení stáří biologické stopy není při současných znalostech
forenzní genetiky možné. Je sice známo, že u stopy dochází
k degradaci materiálu a ke změně v zastoupení a struktuře
některých složek, ovšem převést tyto vcelku obecné poznatky na
relevantní výpočet stáří stopy zatím nelze ani forenzně genetickými,

43
ani jinými metodami; je to však předmětem zájmu forenzně
genetického výzkumu.
Je možné, že osoba s předmětem manipuluje, a přitom
na něm její materiál neulpí?
Velmi zajímavou a často diskutovanou otázkou je, jak velká
množství biologického materiálu po sobě člověk zanechává.
Zejména se to týká povrchových epitelií kůže, které ulpívají na
předmětech přicházejících do kontaktu s nechráněnou pokožkou
(šperky, hodinky, brýle, psací potřeby, telefonní sluchátka a mobily,
volant a řadicí páka auta, kliky, zápisníky atd.). Studie ukázaly, že
toto množství je značně individuální: někdo je dobrý donor,
zanechávající svůj materiál i při letmém kontaktu, někdo je naopak
donor extrémně špatný, u něhož i po dlouhodobém kontaktu
zůstává na předmětech materiálu jen velmi málo. Tato vlastnost je
pravděpodobně dědičná a souvisí jednak s rychlostí obnovy
pokožky (tedy jak rychle se stará pokožka odlupuje a nová zespoda
dorůstá), jednak se soudržností jednotlivých buněk v pokožce.
Je možné, že materiál jedné osoby je ve stopě
detekovatelný, i když se stopou už manipulovala další
osoba/y?
Ano, a to zcela běžně.
Lze při nálezu biologického materiálu více osob na
témže předmětu určit, kdo s předmětem manipuloval
poslední?
Jednou z dosti rozšířených, leč mylných představ je, že na
předmětech dochází k „vrstvení“ biologického materiálu a že tak lze
říci, čí materiál na něm ulpěl jako poslední. Biologický materiál však
takto až na výjimečné situace nefunguje a určení „posledního

44
manipulujícího“ tudíž není možné – ve stopě se nachází směsný
biologický materiál, u něhož nelze analýzou DNA určit, kdy došlo
k přispění jednotlivých donorů.
Co je to primární přenos biologického materiálu?
Primárním přenosem rozumíme nanesení biologického materiálu
z těla původce na primární nosič (předmět či osobu). Typickými
příklady primárního přenosu mohou být zakrvácení čepele nože při
bodnutí, vstříknutí spermatu do prezervativu při ejakulaci, nanesení
kožních buněk na sluchátko při telefonování atd.
Co je to sekundární přenos biologického materiálu?
Sekundárním přenosem rozumíme nanesení biologického
materiálu z primárního nosiče na nosič sekundární. Typickými
příklady sekundárního přenosu mohou být otření zakrvácené
čepele nože o oděv, vytečení spermatu z prezervativu na koberec,
přenesení kožních buněk prvního telefonujícího na rukáv druhého
telefonujícího při užití téhož sluchátka atd.
Obrázek 5 – Primární přenos biologického materiálu (zde krev) může být následován přenosem sekundárním.

45
V hotelovém pokoji byla jako stopa zajištěna šmouha ze stropu.
V tomto vzorku byla zkoumáním prokázána přítomnost spermatu.
Později bylo objasněno, že šmouha vznikla tak, že hotelový host si po
souloži s partnerkou sejmul prezervativ, napnul ho a vystřelil proti
stropu.
Velmi specifickým jevem je sekundární přenos vektorem – např.
krvesajným hmyzem (komár, blecha atd.) Nasátá krev obsahuje po
určitou dobu (než je strávena) DNA v analyzovatelném stavu, a
tudíž kupříkladu zabitím komára klasickou metodou prudké rány
polštářem může na stěně či textilu vzniknout krevní stopa
identifikovatelná s konkrétní osobou. Je přitom prokázáno, že tento
hmyz má dosti velký akční rádius, a krev osoby tak může být
přenesena na poměrně vzdálené místo.
Existují ovšem i jiné, bizarnější mechanismy přenosu biologického
materiálu vektorem.
Ve státě New York byl řešen kuriózní případ paternity: dívka
otěhotněla a porodila dítě poté, co její partner použil při styku
prezervativ, který předtím vytáhnul použitý z odpadkového koše a
před nasazením na penis převrátil naruby, aby se neznečistil. Otcem
dítěte se tak stal mladík, který s dotyčnou dívkou nikdy neměl
pohlavní styk.
Sekundární přenos je z forenzního hlediska značně komplikujícím
jevem, neboť ve svém důsledku může vést k tomu, že:
na předmětu se vyskytuje biologický materiál osoby,
která s ním nemanipulovala (např. muž A si odplivne na
lavičku a odejde, nic netušící žena si do sekretu sedne,
později se stane obětí znásilnění a vraždy – na jejím oděvu
je zjištěna markantní biologická stopa a stanoven profil

46
muže A, ale ten s ní ve skutečnosti vůbec nepřišel do
kontaktu);
na místě činu se vyskytuje biologický materiál osoby,
která tam nikdy nebyla (v hotelu na pokoji č. 27 komár
nasaje krev spícího hosta A, poté vyletí oknem a vletí do
pokoje č. 59, kde je bdícím hostem B zabit, následně je
ovšem neznámým pachatelem zabit host B – na stěně je
nalezena krev a stanoven profil hosta A, ale ten na pokoji č.
59 nikdy nebyl).
Existence sekundárního přenosu biologického materiálu jako
taková ovšem rozhodně není důvodem zpochybnění důkazů
postavených na analýze DNA. Je ovšem vždy absolutně
nezbytné posuzovat DNA důkaz v kontextu ostatních zjištění a
zvážit i další možné hypotézy.
Co je to „nepravý“ sekundární přenos biologického
materiálu?
Za jakýsi „nepravý sekundární přenos“ lze považovat situaci, kdy je
přenesen biologický materiál s celým předmětem, a to jak
náhodně, tak úmyslně (muž A zapomene v restauraci zapalovač,
vezme si ho muž B, který později vykrade rekreační chatu, přičemž
tam zapalovač ztratí – zapalovač je zajištěn jako stopa a je v něm
zjištěn genetický profil muže A, který ale v chatě nikdy nebyl).
Důkazní potenciál stopy
Co je důkazní potenciál?
Podstatou každého dokazování je zvážení pravděpodobnosti
jednotlivých hypotéz o skutku ve světle faktů (svědectví,
znaleckých zkoumání atd.). Nejinak je tomu u důkazů
prostřednictvím analýzy DNA: po úspěšném provedení analýzy se

47
obvykle určitá hypotéza začne jevit pravděpodobnější, zatímco jiná
méně pravděpodobná až zcela nepravděpodobná.
Krátce po nočním vloupání do obchodu zadržela policie na nedaleké
tramvajové zastávce muže. Na rozbité výloze obchodu byla zajištěna
krev, na levé ruce zadrženého muže bylo zjištěno řezné poranění.
V tento moment existují dvě základní hypotézy: 1)původcem krve na
výloze je zadržený muž, nebo 2) původcem krve na výloze je někdo
jiný a to, že zadržený má pořezané zápěstí, je jen shoda okolností.
Analýza DNA následně razantně jednu z těchto hypotéz posílí a
druhou oslabí.
Čím více je jedna hypotéza analýzou stopy podpořena na úkor
ostatních, tím větší má stopa důkazní potenciál. Důkazní potenciál
stopy pramení ze dvou základních složek: z apriorní těsnosti
vazby stopy ke skutku a z očekávané rozdílnosti výsledku
příslušného znaleckého zkoumání za platnosti jednotlivých
zvažovaných hypotéz.
Co je apriorní těsnost vazby stopy ke skutku?
Tímto pojmem rozumíme míru naší jistoty, že stopa vznikla
v souvislosti se skutkem. Určité stopy mohou být již předem
považovány za velmi silně vázané ke skutku: například biologický
materiál plodu získaný při interrupci u čtrnáctileté dívky je
prakticky s naprostou jistotou vázán k pohlavnímu zneužívání této
poškozené. Naproti tomu u celé řady ve forenzní praxi běžně
analyzovaných stop je jejich vazba ke skutku nejistá: nedopalek
zajištěný na chodníku před vraty garáže, z níž bylo odcizeno auto,
může a nemusí mít vztah k tomuto skutku.

48
Co je očekávaná rozdílnost výsledku příslušného
znaleckého zkoumání za platnosti jednotlivých
zvažovaných hypotéz?
Aby zkoumání mělo smysl, musí existovat předpoklad, že analýza
poskytne jiný výsledek za platnosti hypotézy 1 než za platnosti
hypotézy 2, 3 atd. U výše uvedeného příkladu – případ nočního
vloupání – očekáváme, že při platnosti hypotézy 1 (původcem krve
na rozbité výloze je zadržený muž) analýza zjistí shodu profilu ze
stopy a srovnávacího vzorku muže, zatímco při platnosti hypotézy 2
(původcem krve na rozbité výloze je někdo jiný) budou tyto
genetické profily různé.
V jakých situacích má stopa z principu nízký či nulový
důkazní potenciál?
Za určitých okolností lze již předem (tj. bez provedení analýzy)
vyhodnotit, že daná biologická stopa jako důkaz posloužit nemůže,
nebo jen ve velmi omezené míře – říkáme, že má nízký či nulový
apriorní důkazní potenciál. Sem patří:
situace, kdy je malá těsnost vazby stopy ke skutku, neboli
vztah zajištěného biologického materiálu a skutku je velmi
sporný až vyloučitelný
Pachatel loupežného přepadení odhodil při útěku kuklu do křoví, ta
byla zajištěna jako stopa, z nepochopitelných důvodů byla zajištěna i
značně degradovaná použitá dámská menstruační vložka, která
ležela v křoví opodál; vztah této „stopy“ ke skutku je z logiky věci
nulový.
situace, kdy u zvažovaných hypotéz očekáváme totožný
výsledek analýzy

49
Byl oznámen případ znásilnění mezi manželi, kdy žena tvrdila, že
manžel ji znásilnil, ten tvrdil, že mezi nimi došlo k dobrovolnému
pohlavnímu styku. Ke zkoumání byl předložen poševní stěr poškozené.
Zkoumání je v tomto případě irelevantní, neboť u obou hypotéz lze
očekávat zjištění přítomnosti manželova spermatu v poševním stěru
manželky.
Znalci nepřísluší rozhodovat o právních otázkách ani o tom, jak má
být při vyšetřování postupováno. Nicméně, na tomto místě se
přimlouváme za to, aby kompetentní osoby vždy pečlivě zvažovaly,
jaký bude reálný přínos znaleckého zkoumání dané stopy pro
vyšetřování.

50
Seznámení s genetikou
Základní fakta o DNA
Co je to DNA?
DNA neboli deoxyribonukleová kyselina je jednou z funkčních
makromolekul, které se vyskytují v buňce. Označení „kyselina“
poukazuje na některé její chemické vlastnosti (ve vodném roztoku
odštěpuje protony a stává se záporně nabitou, což je jedna
z charakteristik kyselin), avšak rozhodně si ji nelze představovat
jako „klasické kyseliny“ – není to žádná leptající kapalina, nýbrž
vláknitá makromolekula.
Čistá DNA není ve vodném roztoku vidět – je rozpuštěná. Lze ji ovšem
určitými postupy vysrážet a tím „zviditelnit“: například v silně
koncentrovaném etanolu bude velké množství DNA patrné jako bělavé
vláknité chomáčky.
Jakou má DNA strukturu?
Základní struktura DNA je velmi jednoduchá: jednotlivé vlákno DNA
(říkáme mu jednořetězcová DNA) vzniká řetězením stavebních
kamenů – tzv. nukleotidů – lineárně za sebou (jako do řetízku
vzájemně slepené korálky).
Nukleotid sestává ze tří funkčních částí – trifosfátu, cukru
deoxyribózy a tzv. báze. Zatímco deoxyribóza i trifosfát jsou ve
všech nukleotidech stejné, báze se vyskytuje v DNA ve 4 různých
verzích – nazýváme je adenin, guanin, cytozin a tymin a
označujeme A, G, C a T. Existují tedy čtyři základní typy nukleotidů
podle toho, kterou bázi nesou (jako bychom měli čtyři různé „barvy
korálků“).

51
Sled nukleotidů ve vlákně – tzv. sekvence nukleotidů – vytváří
jednoduchý lineární kód o čtyřech znacích. (Obdobně například
naše písmo – latinka – je lineární kód, užívající ovšem podstatně
více znaků – v anglické abecedě 26, v té české dokonce 37 – a to
počítáme pouze písmena, nikoli interpunkční a jiná znaménka!).
Obrázek 6 – Základní struktura jednořetězcové DNA. DNA je lineárním polymerem nukleotidů, každý nukleotid sestává z trifosfátu, cukru deoxyribózy a báze.
V „genetickém světě“ zapisujeme sekvenci nukleotidů v DNA pomocí
zmíněných čtyř písmen, označujících jednotlivé báze, tedy například:
ACCAGTGAGATCCCAATATAGTACACACACTGCTGCTACTTCTAGAGG
CAAACTTAGT
Vlákno DNA může za určitých okolností existovat v popsané
jednořetězcové formě, běžněji se ale v buňce setkáváme
s dvouřetězcovou DNA. Ta je tvořena dvěma řetězci nukleotidů,

52
které jsou navzájem propojeny pomocí vazeb mezi bázemi. Tento
komplex je prostorově svinutý do tvaru známé dvoušroubovice.
Při vzniku dvouřetězcové DNA se uplatňuje zásadní pravidlo,
kterému říkáme princip (pravidlo) komplementarity: každý typ
báze páruje pouze s jediným jiným typem báze. A páruje vždy s T (a
logicky T s A) a C páruje vždy s G (a logicky G s C). Oba řetězce pak
označujeme jako vzájemně komplementární.
Obrázek 7 – Struktura dvouřetězcové DNA. Dvě vlákna spolu párují prostřednictvím bazí.
Vezměme si krátký úsek jednořetězcové DNA, jehož sekvence je
AGGTAC. Při vzniku druhého řetězce se uplatní princip
komplementarity a dvouřetězec tedy bude vypadat:
AGGTAC
| | | | | |
TCCATG
V běžné tělní lidské buňce je DNA o celkové délce cca 6,6 miliardy
nukleotidů, což de facto představuje dvoušroubovici o délce zhruba
2 metry. Velikost lidské buňky je však dramaticky menší – například

53
bílé krvinky mají průměr řádově pouhých 0,00001 metru. Jedinou
možností, jak vtěsnat něco tak dlouhého do něčeho tak malého, je
mnohonásobně to smotat.
DNA je proto v buňce velmi složitě uspořádána do vyšších a vyšších
stupňů: dvoušroubovicové vlákno se ovíjí kolem bílkovinných
„klubíček“ histonů, takto navinuté vytváří smyčky uchycené na
bílkovinné „lešení“ a celá tato struktura se spirálovitě stáčí a vytváří
tzv. chromatinové vlákno.
Co jsou chromozómy?
Vzhledem ke zmíněné značné délce DNA by i toto chromatinové
vlákno bylo neúměrně dlouhé a buňka by s ním mohla jen obtížně
pracovat. Proto je rozděleno na kusy, kterým říkáme chromozómy,
podobně jako třeba Ottův slovník naučný je rozdělen do svazků.
Soubor chromozómů v jádře nazýváme karyotyp.
Obrázek 8 – Karyotyp neboli soubor chromozómů v jádře.

54
Biologické druhy mají různý počet chromozómů – např. člověk má
23 párů chromozómů, kočka 18 párů chromozómů, damašská růže
28 chromozómů atd. Vyšší počet chromozómů však neznamená, že
daný druh má delší DNA: pouze ukazuje, do kolika „svazků“ je tato
DNA rozdělena.
V lidských buňkách najdeme za normálních okolností 1 pár
chromozómů, které určují, jaké bude pohlaví jedince – říkáme jim
proto chromozómy pohlavní neboli gonozómy a označujeme je X
a Y. Zatímco muži mají jeden chromozóm X a jeden Y, ženy mají dva
chromozómy X.
Zbylých 22 párů chromozómů se na určení pohlaví nijak nepodílí –
říkáme jim proto chromozómy nepohlavní neboli autozómy a
označujeme je čísly 1–22.
Hovoříme zde o „párech“ chromozómů – to proto, že od každého
chromozómu najdeme v běžné tělní buňce dvě kopie, z nichž jedna
pochází od matky, druhá od otce. Celý princip bude popsán dále
v kapitole o podstatě dědičnosti.
Jak je v DNA zapsána informace?
Vlastní funkční uspořádání DNA je nesmírně složité, omezíme se
proto pouze na velmi zjednodušující popis.
V DNA lze nalézt úseky, které nesou informaci o struktuře
buněčných bílkovin. Těmto úsekům říkáme obecně geny. Ostatní
úseky, které takovou informaci nenesou, označujeme termínem
extragenová DNA.
V genech je využíván tzv. univerzální genetický kód. Tak jako je
DNA řetězcem nukleotidů, je bílkovina řetězcem aminokyselin,
kterých existuje 21 druhů. Každá trojice nukleotidů v genu – tzv.
triplet – představuje kód pro konkrétní aminokyselinu. Například

55
úsek DNA s kódem TCCAAAGGT znamená, že v bílkovině mají být za
sebou serin–alanin–glycin, neboť TCC je kód pro aminokyselinu
serin, AAA pro alanin a GGT pro glycin.
Obrázek 9 – Genetický kód: trojice nukleotidů (triplety) kódují konkrétní aminokyseliny zařazované do struktury bílkoviny.
Vlastní exprese (tj. vyjádření informace) genu probíhá tak, že podle
příslušného vlákna DNA je dle výše zmíněného pravidla
komplementarity syntetizováno vlákno RNA (ribonukleová
kyselina, velmi podobná DNA) – tomuto procesu říkáme
transkripce, tedy přepis. Vlákno RNA je po úpravách použito jako
předloha, podle níž se zařazují aminokyseliny do vznikající
bílkoviny – tento proces se nazývá translace, tedy překlad, nebo též
proteosyntéza neboli syntéza bílkoviny.
Na rozdíl od genů, extragenová DNA tímto způsobem překládána
není a informaci o struktuře bílkovin nenese. (Mezi námi, není to
úplně pravda – i v extragenové DNA se vyskytují kódující oblasti či
jejich zbytky; nicméně buňka je aktivně nevyužívá, spíše je jimi

56
sama někdy zneužívána, a pro potřeby této publikace zcela postačí
zjednodušující tvrzení, že v extragenové DNA kódující oblasti
nejsou.)
Co je to gen, lokus, marker?
Protože v užívání těchto pojmů nepanuje přílišná jednota ani mezi
samotnými odborníky, pokusme se je nyní blíže vyspecifikovat a
správně definovat.
Jako gen označujeme funkční úsek DNA, jehož určitá část je
překládána do struktury bílkovin a ostatní části se tohoto překladu
nějakým způsobem účastní. Pokud slovně specifikujeme, o jaký gen
se jedná, nejčastěji použijeme název příslušné bílkoviny, která podle
něj vzniká. Tedy: gen pro beta podjednotku alkoholdehydrogenázy
1, gen pro alfa podjednotku ATP syntázy atd.
Na tomto místě vyvraťme jednu oblíbenou novinářskou floskuli,
zaplevelující stránky novin i internetových portálů s pozoruhodnou
životaschopností: neexistuje nic jako „gen pro nemoc“. Titulek „Vědci
objevili gen rakoviny prostaty“ je nesmyslný. Každý gen v naší
buňce totiž má svou významnou pozitivní roli a přispívá k fungování
buňky jako celku. Sama přítomnost jakéhokoli genu nemá negativní
vliv na buňku. To, co vědci objevili v našem příkladu, je vadná forma
onoho genu; právě špatné fungování genu totiž má za následek
určitou nemoc organismu. (Nemluvě o tom, že asociativní vztah mezi
prostatou a formou genu je jen pravděpodobnostní.)
Řekněme to ještě jinak, obrazněji: to, že parašutista má ve své výbavě
padák, mu smrt nezpůsobí, naopak. Fatální pro něj bude, pokud má
nefungující padák.
Jako lokus označujeme libovolný adresný úsek DNA či místo v DNA.
Lokus DNA lze považovat za vůbec nejobecnější pojem – můžeme
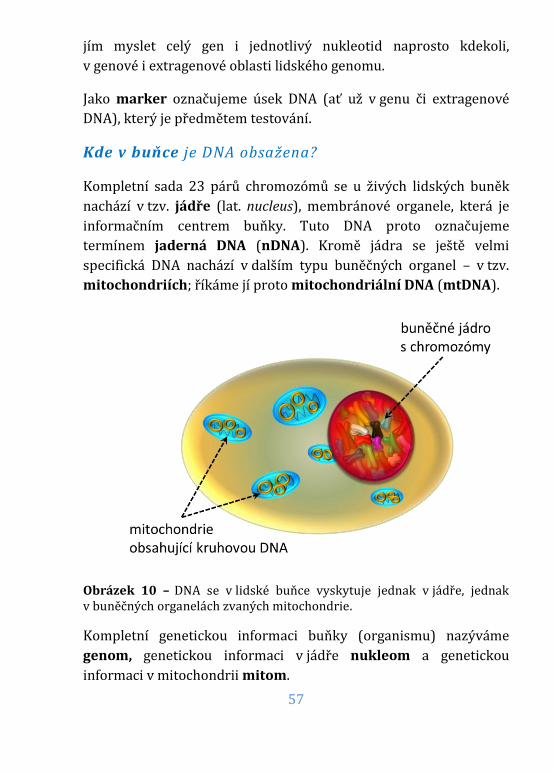
57
jím myslet celý gen i jednotlivý nukleotid naprosto kdekoli,
v genové i extragenové oblasti lidského genomu.
Jako marker označujeme úsek DNA (ať už v genu či extragenové
DNA), který je předmětem testování.
Kde v buňce je DNA obsažena?
Kompletní sada 23 párů chromozómů se u živých lidských buněk
nachází v tzv. jádře (lat. nucleus), membránové organele, která je
informačním centrem buňky. Tuto DNA proto označujeme
termínem jaderná DNA (nDNA). Kromě jádra se ještě velmi
specifická DNA nachází v dalším typu buněčných organel – v tzv.
mitochondriích; říkáme jí proto mitochondriální DNA (mtDNA).
Obrázek 10 – DNA se v lidské buňce vyskytuje jednak v jádře, jednak v buněčných organelách zvaných mitochondrie.
Kompletní genetickou informaci buňky (organismu) nazýváme
genom, genetickou informaci v jádře nukleom a genetickou
informaci v mitochondrii mitom.

58
U rostlin najdeme ještě další organely, obsahující DNA – plastidy,
z nichž laikům asi nejznámější jsou chloroplasty – organely, kde
rostlinné buňky zachycují sluneční záření do fotonových pastí –
fotosyntetizují. Souboru genetické informace v plastidech říkáme
plastidom.
Kde v těle je DNA obsažena?
Kompletní genetická informace, tedy příslušných 23 párů
chromozómů, se nachází téměř ve všech typech lidských tělních
buněk (říkáme, že genetická výbava všech tělních buněk je
uniformní). Výjimku tvoří červené krvinky (erytrocyty) a krevní
destičky (trombocyty), které neobsahují DNA vůbec, a buňky
pohlavní (gametocyty), tedy spermie a vajíčka (oocyty), které
obsahují pouze polovinu této dědičné informace – od každého
chromozómu pouze jednu kopii. Proč tomu tak je se dozvíte
v následující pasáži o dědičnosti.
Základní principy fungování dědičné informace
Jak vznikají viditelné či měřitelné rozdíly mezi lidmi?
Už při prostém pohledu kolem sebe vidíme, že lidé se navzájem liší
v řadě znaků – barvě očí, vlasů, tvaru obličeje, krevní skupině,
snášenlivosti alkoholu, temperamentu… Téměř neexistuje nic, v čem
by všichni lidé byli zcela uniformní.
Tomu, jak nějaký znak u dané osoby konkrétně vypadá, říkáme
fenotyp. Tato výsledná podoba znaku je dána obvykle společným
vlivem dvou faktorů: vlivu prostředí a dědičné informace. Existují
znaky, které jsou založeny téměř výhradně na dědičné informaci, a
žádný vliv prostředí je nemůže změnit; říkáme, že jsou silně
geneticky determinované. Takovým znakem je například krevní
skupina.

59
Naproti tomu existují znaky, kde hraje prostředí významnou roli a
daný znak jím může být silně ovlivněn; genetická informace se
u nich uplatňuje pouze jako jakýsi velmi tvárný základ, nebo
chceme–li, dispozice. Říkáme, že tyto znaky jsou slabě geneticky
determinované. Typickým příkladem budiž tělesná hmotnost.
Co je polymorfismus?
Všichni lidé mají stejné geny, přičemž odhadem je genů kolem 23
tisíc. Některé geny však existují ve více verzích – říkáme, že jsou
variabilní, polymorfní, a daný jev označujeme jako
polymorfismus. Jednotlivé verze genu nazýváme alely. Lidé mají
tedy stejné geny, ale nemají stejné alely. K tomuto základnímu
principu se přidává ještě další, neméně významný: i zcela stejná
varianta genu (alela) může být u dvou osob využívána buňkami
různě často a různě intenzivně; takovým rozdílům říkáme
epigenetické.
Představit si to lze kupříkladu takto: všechny restaurace mají ve své
obrovské spíži prakticky tytéž suroviny – mouku, česnek, med, mléko,
rajský protlak, red curry, vajíčka, jablka, olivový olej, atd. – celkem
několik tisíc přísad. Každá tato surovina se ale vyskytuje ve více
verzích, takže zatímco jedna restaurace nakupuje levný česnek z Číny,
jiná má k dispozici řízný česnek od farmáře z Vysočiny. Stejně tak
existují desítky různých odrůd jablek, rozličné druhy medu atd.
Přičteme–li k tomu ještě rozdíly v postupu přípravy (jeden kuchař
používá více, jiný méně pepře, jeden do salátu dává raději konopný
olej, zatímco druhý směs olivového a palmového), bude ve výsledku
totéž jídlo v každé restauraci trochu jiné (a někdy i hodně jiné).
Jak polymorfní jsou jednotlivé lokusy v DNA?
Velmi zhruba můžeme říci, že co do variability najdeme v lidské
DNA tři typy lokusů:

60
1. lokusy nepolymorfní, tedy zcela minimálně variabilní (říkáme
jim též vysoce konzervativní sekvence); obvykle jsou to nějaké
zásadní funkční úseky, bez nichž se buňka neobejde a které
musejí mít určitou přesnou sekvenci, jinak svou funkci ztratí;
2. lokusy nízce až středně polymorfní, vyskytující se v populaci
v několika variantách; sem spadají především geny;
3. lokusy vysoce polymorfní (multialelické), vyskytující se
typicky v mnoha variantách, jejichž počet může jít i do mnoha
desítek; najdeme je především v určitých oblastech nekódující
(extragenové) DNA;
Obrázek 11 – Variabilita lokusů v DNA může být velmi rozdílná.
Největší rozdíly mezi osobami budou z logiky věci právě u posledně
jmenovaných vysoce polymorfních lokusů. Především na ně se tudíž
bude zaměřovat identifikační analýza DNA.
Lidský genom má 2 x 3,3 miliardy nukleotidů, ale jen 2 x cca 3
milióny z nich jsou variantní. To znamená, že dvakrát 3 297 000 000
nukleotidů nás činí lidmi a složení zbývajícího jednoho promile
počtu nukleotidů nás činí originálním člověkem.

61
Mimochodem, když genetici porovnali kompletní sekvenci lidského a
šimpanzího genomu, zjistili fascinující věc: 98,4 % této sekvence je
shodné, jinak řečeno, od šimpanze se lišíme v pouhém 1,6 % své
dědičné informace. Je téměř neuvěřitelné, že tak málo postačuje
k tomu, abychom byli tak velmi odlišní.
Co jsou mutace?
Termínem mutace souhrnně označujeme změny v sekvenci DNA.
Pro účely této publikace se stran mutací omezme jen na pár
základních informací:
Co do svého rozsahu mohou být mutace zcela nepatrné –
jednobodové, spočívající v záměně jediného nukleotidu v sekvenci
(např. ATTACCGTT se změní na ATTTCCGTT), přes mutace
spočívající v přesunu, vymazání nebo naopak „připsání“ části
sekvence až po mutace obrovského rozsahu, spočívající
v přestavbách celých chromozómů či dokonce změnách počtu
chromozómů či jejich sad.
Sám rozsah mutace ovšem není úměrný účinku na organismus:
jednobodová mutace v kritickém místě nějakého genu může mít
zcela fatální důsledky, zatímco některé přestavby chromozómů
nemají žádný viditelný účinek a odhalíme je až náhodně při
genetickém vyšetření.
Svým původem mohou být mutace spontánní (laicky řečeno:
změnu sekvence způsobí chyba ve vnitřních buněčných pochodech)
nebo indukované (k mutaci dojde vlivem nějakého biologického,
fyzikálního či jiného činitele).
K mutacím dochází jak v tělních buňkách – pak mluvíme o mutacích
somatických, tak v zárodečných buňkách, z nichž vznikají buňky
pohlavní – takové mutace nazýváme gametické (nebo též mutace
v germinální linii).

62
Jak je genetická informace přenášena z rodičů na děti?
Potomci dědí vlastnosti svých rodičů – to bylo známo už antickým
myslitelům. Dnes víme, že je to proto, že dítě získá od obou rodičů
kopii části jejich dědičné informace v podobě DNA.
Jak jsou děděny lokusy na nepohlavních
chromozómech?
Lokusy na autozómech (nepohlavních chromozómech) jsou děděny
tzv. mendelovskou dědičností (princip objevil J. G. Mendel).
Člověk má v tělních buňkách dvě sady autozómů, a tedy dvě kopie
každého autozomálního lokusu. Tyto dvě kopie však nemusejí být
zcela identické – mohou to být dvě různé varianty daného lokusu,
tedy dvě různé alely.
Obrázek 12 – Meióza neboli redukční dělení je mechanismus tvorby pohlavních buněk, při němž se redukuje dědičná informace na polovinu.

63
Při tvorbě pohlavních buněk se dědičná informace složitým
mechanismem nazývaným meióza redukuje na polovinu a do zralé
pohlavní buňky se dostane pouze jedna kopie každého
autozomálního lokusu, tedy jedna alela daného lokusu. Záleží
víceméně na náhodě, která z obou alel to bude.
Spojením spermie otce a vajíčka matky vznikne znovu jediná
buňka (zygota, oplozené vajíčko) nesoucí dvě kompletní sady
chromozómů, a tedy dvě kopie každého autozomálního lokusu.
Obrázek 13 – Při oplození nese každá z pohlavních buněk genetickou informaci redukovanou na polovinu. Splynutím vajíčka a spermie tak vzniká prvotní buňka (zygota) s normálním množstvím genetické informace.
Co je to heterozygot a homozygot?
Již několikrát jsme zde zmínili, že každý autozomální lokus má
člověk ve výbavě svých tělních buněk dvakrát – jednu kopii od

64
matky, druhou od otce. Pokud jsou obě kopie identické (jsou to dvě
stejné alely), říkáme, že daný lokus je v homozygotním stavu a
daný člověk je v tomto lokusu homozygot (homozygotní). Naproti
tomu pokud je na každém párovém chromozómu v daném lokusu
jiná alela, říkáme, že daný lokus je v heterozygotním stavu a daný
člověk je v tomto lokusu heterozygot (heterozygotní).
Tedy: v případě homozygotního stavu zdědil v daném lokusu
jedinec od matky a otce dvě stejné alely. V případě heterozygotního
stavu zdědil od matky a otce dvě různé alely.
Obrázek 14 – Dědičnost autozomálních lokusů se řídí tzv. Mendelovými zákony. Dědičná informace dítěte je zkombinována z části dědičné informace matky a části dědičné informace otce.

65
Prosím povšimněte si, že homozygozita či heterozygozita je
vlastností konkrétního lokusu na nepohlavních chromozómech,
nikoli genetické informace jako celku! Tedy: každý z nás je v řadě
svých lokusů heterozygotem a v řadě homozygotem.
Sám termín má původ v řeckém „homo–/hetero–“ (= stejno–/různo–)
a „zygós“ (pohlavní buňka). U homozygotního lokusu totiž všechny
pohlavní buňky dané osoby ponesou stejnou alelu tohoto lokusu (jiná
není k dispozici), zatímco u heterozygotního lokusu nese zhruba
polovina pohlavních buněk jednu alelu původem z prvního párového
chromozómu a zbytek (tj. druhá polovina) druhou alelu původem
z druhého párového chromozómu.
Jak jsou děděny lokusy na pohlavních
chromozómech?
Dědičnost lokusů na gonozómech (pohlavních chromozómech) je
různorodější – existují zde oblasti, ve kterých je sekvence X i Y
chromozómu velmi podobná – nazýváme je oblasti
pseudoautozomální, a dále oblasti, v nichž se X a Y chromozóm
zásadně liší. Pseudoautozomální oblasti obsahují na chromozómu
X i Y tytéž lokusy, jež jsou děděny mendelovsky, tak jak bylo
popsáno výše. Ostatní oblasti pohlavních chromozómů X a Y se
chovají jinak.
Chromozóm Y je přítomen v jedné kopii u mužů a dědí se
paternálně (patrilineárně, po meči). Lokusy jsou předávány
v nezměněném stavu „jako balík“ – tzv. haplotyp – z otce na syny.
Jediný mechanismus, kterým se může dědičná informace v této části
Y–chromozómu za normálních okolností změnit, je mutace.

66
Obrázek 15 – Dědičnost Y–chromozomálních lokusů. Dědičná informace syna pochází pouze od otce.
Chromozóm X je přítomen ve dvou kopiích u žen a v jedné kopii
u mužů (tam je druhým pohlavním chromozómem Y). U žen se tedy
i X lokusy vyskytují ve dvou kopiích, zatímco u mužů jen v jedné.
Žena předává potomkovi vždy jeden ze svých X chromozómů. Muž
předá potomkovi buď svůj chromozóm X – pak to bude děvče, nebo
svůj chromozóm Y – pak to bude syn.
Na tomto místě nelze nepoznamenat, že ženy byly po dlouhá staletí
obviňovány z neschopnosti dát manželovi syna a někdy za to i
zaplatily životem (že, Jindřichu VII.?!) Objevy v oblasti genetiky ovšem
ukázaly, že je to spermie muže, která nese příslušný chromozóm (X
nebo Y), a tudíž je zodpovědná za to, zda dítě bude děvče či chlapec.
Načež další objevy v oblasti fyziologie početí ukazují, že je to vajíčko,
které důmyslnými mechanismy vybírá, kterou spermii „pustit dovnitř“.

67
Ergo, ač se mi to neříká lehce, původní nařčení byla minimálně
částečně opodstatněná…
Obrázek 16 – Dědičnost X–chromozomálních lokusů. Dědičná informace dcery je zkombinována z části dědičné informace matky a části dědičné informace otce, dědičná informace syna pochází pouze od matky.
Co je to hemizygot?
Pokud se určitý lokus nachází pouze v polovině pohlavních buněk
daného člověka, zatímco ve druhé polovině zcela chybí, říkáme, že
tento člověk je v daném lokusu hemizygot (hemizygotní).
To nastává u lokusů lokalizovaných na X chromozómu: zatímco
ženy mají standardně dva X chromozómy a jsou v X–lokusech buď
homozygotní, nebo heterozygotní, muži mají pouze jeden
chromozóm X (a jeden chromozóm Y) a v X–lokusech jsou
hemizygotní. Stejně tak za hemizygotní můžeme považovat u mužů
lokusy lokalizované na Y–chromozómu. (Obé konstatování se týká

68
lokusů mimo pseudoautozomální oblasti, jejichž dědičnost byla
zmíněna výše).
Jak je děděna mitochondriální DNA?
U člověka a celé řady dalších živočišných i rostlinných druhů byla
prakticky prokázána tzv. striktní matroklinita mitochondriální
DNA, tedy dědičnost „po přeslici“: potomek (dcera i syn) dědí
mitochondriální DNA výhradně po matce, a má s ní tudíž shodný tzv.
mitotyp.
Obrázek 17 – Dědičnost mitochondriální DNA. Dědičná informace dětí pochází pouze od matky.
Mimochodem – některé výzkumy ukazují, že striktní matroklinita
nemusí u člověka vždy platit. V přírodě navíc existují druhy, u nichž je
DNA děděna jinak než „po přeslici“, například oblíbené ovoce kiwi
(Actinidia) má paternální dědičnost mtDNA, tedy „po meči“. Inu,
příroda je ráda komplikovaná…

69
Forenzní potenciál genetické analýzy
Z forenzního hlediska lze genetickou analýzou získat informace,
které nám
pomohou identifikovat původce biologického materiálu
pomohou určit typ biologického vztahu mezi osobami
(biologickou příbuznost)
pomohou predikovat některé biologické charakteristiky
původce biologického materiálu
jinak pomohou posílit/oslabit naše hypotézy,
přičemž prvý bod dominuje stávající kriminalistické genetice, druhý
zase občansko–právním řízením o určení otcovství.
Individualita a identifikace
Co je to identifikace?
Identifikací ve forenzní genetice rozumíme určení shodného
původu dvou vzorků biologických materiálů – např. stopy a
srovnávacího vzorku. V praxi to znamená, že posuzujeme určité
variabilní znaky v DNA a zjišťujeme, zda oba materiály vykazují
v takto variabilních znacích shodu.
Některé z metod principiálně umožňují identifikaci až na úroveň
konkrétního jedince s vysokou mírou spolehlivosti (blížící se jistotě)
– potom mluvíme o individuální identifikaci.
Jiné metody už ze svého principu takovou rozlišovací schopnost
nemají – sem patří například analýzy mitochondriální DNA a
haplotypu Y–chromozómu; přesto mají tyto analýzy ve forenzní
praxi své nezastupitelné místo a bude o nich též pojednáno
v samostatných kapitolách.

70
Co je to genetický profil osoby?
Termínem genetický profil (syn. DNA profil) označujeme obecně
set genotypů několika polymorfních lokusů dané osoby; v praxi je
tak nejčastěji označován set genotypů autozomálních STR lokusů.
Genetický profil osobu částečně či zcela identifikuje.
Co je to genetické individuum?
Každý lidský jedinec vzniká mechanismem pohlavního
rozmnožování, tj. proniknutím otcovy spermie do matčina vajíčka;
tento děj nazýváme oplození. Po oplození dojde uvnitř vajíčka ke
splynutí jeho prvojádra s prvojádrem spermie do jediného celku,
nazývaného synkaryon, čímž je vytvořen zcela jedinečný soubor
genů a extragenové DNA – genetická informace nového, unikátního
jedince. Tato první buňka s kompletním jádrem se nazývá zygota.
Všechny buňky, vzniklé jejím dělením, nesou prakticky totožnou
genetickou informaci a tvoří genetické individuum, nazývané
odborně geneta. Jakékoli prohlášení o shodě či neshodě dvou
biologických materiálů se vždy a bezvýhradně týká pouze toho,
zda tyto materiály mohou či nemohou pocházet z téže genety.
To je zcela zásadní atribut forenzního testování a je třeba jej
mít vždy na paměti. Jak se dozvíme později, ne vždy je totiž jedinec
genetický totožný s jedincem fyzickým.
Co je to fyzické individuum?
Fyzickým individuem rozumíme konkrétního, prostorově
ohraničeného jedince; odborně jej v biologii nazýváme rameta.
U lidí je tedy rametou konkrétní člověk a v běžném životě je tak
rameta vnímána jako základní forma individua, která je jednotkou
sociálních interakcí ve společnosti i jednotkou v právním smyslu
slova (fyzickou osobou).

71
Obrázek 18 – Typický genetický profil osoby: set genotypů autozomálních STR lokusů a amelogeninu (bližší vysvětlení, o jaké lokusy se jedná a proč je testujeme, najdete v dalších částech knihy).

72
Jaký je vzájemný vztah genety a ramety?
Člověk patří spolu s dalšími obratlovci mezi tzv. unitární
organismy – ty se vyznačují tím, že z oplozeného vajíčka se
postupně vyvíjí jedinec s poměrně striktně daným uspořádáním
těla, prochází nejprve fází růstu, potom plodným obdobím a jeho
život je zakončen stářím a smrtí. Jedna geneta se tak stává jednou
rametou.
V přírodě se můžeme setkat ještě s jiným rozšířeným modelem –
s tzv. modulárními organismy, jakými jsou například rostliny. Ty
mají naopak velmi proměnlivou a přizpůsobivou podobu těla, které
sestává z opakujících se částí – modulů (např. listy, květy), jejich
vývoj není nikdy ukončený (jejich tělo stále roste) a jednotlivé
moduly mohou dát za příznivých podmínek vzniknout dalšímu
fyzickému jedinci. Jedna geneta se tak může stát mnoha
rametami.
Tedy, souhrnně řečeno: u člověka a dalších unitárních
organismů je rameta v drtivé většině případů totožná
s genetou; jeden jedinec genetický se vyvine v jediného jedince
fyzického. U rostlin a dalších modulárních organismů naopak
může jeden jedinec genetický existovat ve formě i mnoha
desítek jedinců fyzických.
Až půjdete kolem stánku s květinami, povšimněte si některé z váz
s kyticí nádherných růží, které jsou jedna jako druhá: můžete si být
téměř jisti, že ačkoli byly před pár hodinami nastříhány kdesi
v holandské pěstírně z mnoha desítek růžových keřů, jedná se
o jediného genetického jedince.

73
Může u člověka existovat i jiný vzájemný vztah genety a
ramety?
Ano. Existuje hned několik situací, kdy výše zmíněné pravidlo
jedinec genetický = jedinec fyzický neplatí. To je z hlediska
identifikace nesmírně závažné a je nutné mít při hodnocení případu
tyto potenciální situace vždy na paměti, neboť jak již bylo řečeno,
identifikace analýzou DNA se vždy týká genety.
Těmito výjimečnými situacemi jsou především: jednovaječná
dvojčata, vrozený tetragametický chimérismus, směna buněk mezi
dvojvaječnými dvojčaty, maternální a fetální mikrochimérismus a
trvalý posttransplantační chimérismus. (Prosím čtenáře, aby
nepropadal panice – všechny tyto pojmy si v následujících
odstavcích jeden po druhém objasníme).
Jak se vzájemně liší jednovaječná dvojčata?
První a nejčetnější výjimečnou situací jsou jednovaječná
(identická, monozygotní) dvojčata, případně vícerčata. Už sám
jejich název napovídá, že jsou původem z jednoho vajíčka a že jsou
v určitém ohledu stejná – identická. Vznikají tak, že dělící se
oplozené vajíčko se v určité velmi rané fázi rozvolní na dvě (nebo i
více) skupin buněk a každá tato skupina se zanoří do děložní
sliznice samostatně a dá pak vzniknout samostatnému jedinci –
dítěti. Jednovaječná dvojčata (vícerčata) jsou tedy jednou
genetou, ale více rametami.
Díky svému společnému původu se proto jednovaječná dvojčata
prakticky neliší svou dědičnou informací; jediný mechanismus,
který může dát vzniknout rozdílům v jejich DNA, jsou mutace.
Pozorovatelné či měřitelné rozdíly jsou u jednovaječných dvojčat
ovšem častěji způsobeny nestejným vlivem prostředí – např.
rozdílná tělesná hmotnost, rozdílná barva pokožky, jiné rozložení
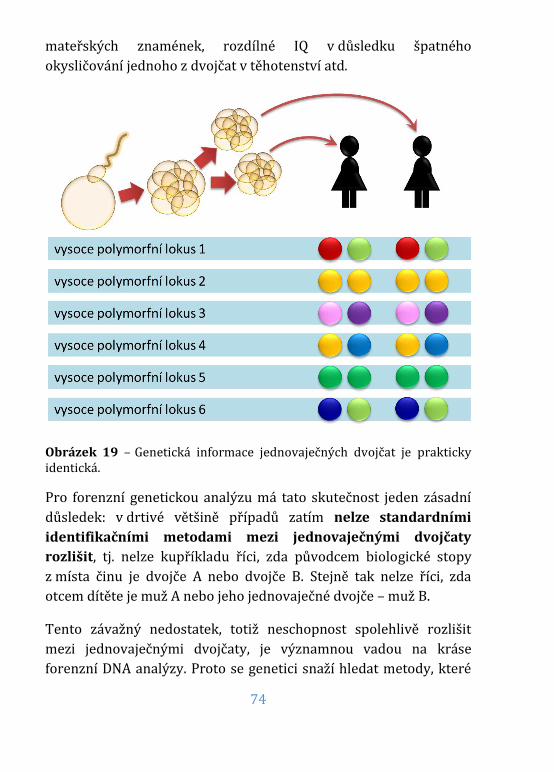
74
mateřských znamének, rozdílné IQ v důsledku špatného
okysličování jednoho z dvojčat v těhotenství atd.
Obrázek 19 – Genetická informace jednovaječných dvojčat je prakticky identická.
Pro forenzní genetickou analýzu má tato skutečnost jeden zásadní
důsledek: v drtivé většině případů zatím nelze standardními
identifikačními metodami mezi jednovaječnými dvojčaty
rozlišit, tj. nelze kupříkladu říci, zda původcem biologické stopy
z místa činu je dvojče A nebo dvojče B. Stejně tak nelze říci, zda
otcem dítěte je muž A nebo jeho jednovaječné dvojče – muž B.
Tento závažný nedostatek, totiž neschopnost spolehlivě rozlišit
mezi jednovaječnými dvojčaty, je významnou vadou na kráse
forenzní DNA analýzy. Proto se genetici snaží hledat metody, které

75
by dokázaly k rozlišení identických dvojčat využít těch změn v DNA,
k nimž dochází až po vzniku samostatných embryí v jejich tělních
buňkách. Přestože některé postupy se jeví jako nadějné, dosud
nebyla nalezena a v odborných časopisech publikována žádná
validní metoda, kterou by bylo možno s velkou spolehlivostí použít
jako důkazní nástroj.
Na rozdíl od analýzy DNA, jiné identifikační obory – např.
daktyloskopie, forenzní odontologie atd. – rozlišení mezi
jednovaječnými dvojčaty umožňují. Je to proto, že kupříkladu otisky
prstů vznikají sice na základě dědičné informace, ale na jejich
přesnou výslednou podobu má určitý vliv i prostředí, které není
u obou dvojčat zcela stejné (byť sdílejí tutéž dělohu). Otisky prstů
jednovaječných dvojčat se proto v malé míře liší.
Jak se vzájemně liší dvojvaječná dvojčata?
Původ dvojvaječných dvojčat (vícevaječných vícerčat) je zcela
odlišný: vznikají v důsledku vícečetného oplození, tj. při ovulaci se
uvolní více vajíček a každé toto vajíčko je samostatně oplozeno.
V některých případech nenastává ovulace současně, ale vajíčka se
uvolní a jsou oplozena s určitým časovým odstupem, někdy i
několikatýdenním.
Jednoduše lze říci, že dvojvaječná (fraternální, dizygotní)
dvojčata jsou normálními biologickými sourozenci, ovšem
počatými v témže okamžiku (či s velmi krátkým časovým
odstupem) a sdílejícími tudíž během prenatálního vývoje tutéž
dělohu. Díky tomu lze mezi nimi analýzou DNA rozlišit stejně
jako mezi biologickými sourozenci.
Protože dvojvaječná dvojčata vznikají dvěma nezávislými
oplozeními, vstupuje do hry ještě další možnost, totiž že
biologickým otcem každého z nich je jiný muž. Tento jev se odborně

76
nazývá heteropaternální superfekundace a, jak dokládá báje
o dvojčatech Héraklovi a Ífiklovi, jeho existence je lidstvu známa již
nejméně od dob starého Řecka.
Obrázek 20 – Genetická informace dvouvaječných dvojčat se liší stejně jako genetická informace běžných sourozenců.
Co je to chimérismus?
Jako chimérismus označujeme stav, kdy je jeden fyzický jedinec
tvořen buňkami původem z více zygot, neboli více genetami
(genetickými jedinci). Rozložení obou buněčných linií v organismu
může být nesmírně různorodé, závisí na způsobu vzniku
chimérismu i na řadě dalších faktorů včetně náhody. Principiálně
existuje více mechanismů, jak k chimérismu může dojít.

77
Co je to vrozený tetragametický chimérismus?
Tetragametický chimérismus je jakýmsi pomyslným opakem
jednovaječných dvojčat. Zatímco u jednovaječných dvojčat se jedna
geneta rozdělí na dvě ramety, u tetragametického chimérismu se
dvě genety spojí do jediné ramety. V těle ženy tedy dojde
k oplození dvou vajíček dvěma spermiemi, která by za normálních
okolností dala vzniknout dvojvaječným dvojčatům. Tato oplozená
dělící se vajíčka se však cestou vejcovodem přiblíží k sobě natolik,
že se navzájem slepí a do děložní sliznice zanoří jako jeden celek.
Následně dají vzniknout jedinému fyzickému jedinci, který ale je
tvořen buňkami dvojího, „sourozeneckého“ původu (celkem dvěma
vajíčky a dvěma spermiemi, tedy čtyřmi pohlavními buňkami –
odtud název tetragametický).
Tetragametický chimérismus je velmi často nepozorovatelný,
skrytý (kryptický). Na osobě často nejsou patrné žádné fyzické ani
jiné odlišnosti, které by nám napověděly, že se může jednat
o chiméru. Jinak řečeno, chimérou může být kdokoli z nás, aniž to
tuší. Nicméně nutno konstatovat, že je to jev velmi řídký – dosud
popsané případy se čítají řádově na desítky osob. Je to však jev
forenzně nesmírně zajímavý a se zásadními důsledky pro oblast
identifikace a určení biologické příbuznosti analýzou DNA.
U tetragametické chiméry totiž neplatí, že všechny tkáně daného
jedince vykazují identický DNA profil, neboť každá z obou
sourozeneckých buněčných linií nese profil jiný. Záleží tedy na tom,
jakým způsobem jsou tyto linie v těle rozmístěny: může například
nastat situace, kdy krev dotyčné osoby vykazuje jiný DNA profil než
její sperma.

78
Obrázek 21 – Genetická informace chimérického jedince vzniklého splynutím zárodků je dvojího typu; takový jedinec je vlastně spojením dvou sourozenců do jediného těla.
Co je to vzájemná směna buněk mezi dvojvaječnými
dvojčaty?
U dvojvaječných dvojčat může v některých případech dojít
k částečnému (byť třeba velmi malému) propojení jejich
krevních oběhů. Krevní buňky jednoho dítěte tak mohou putovat
krevním řečištěm druhého, a naopak. Mezi těmito buňkami jsou i
buňky krvetvorné, které se mohou „uhnízdit“ v kostní dřeni
dvojčete a dát tam natrvalo vzniknout celé skupině krvetvorných
buněk, z nichž vznikají krvinky a krevní destičky. Ve výsledku se
tedy v kostní dřeni jednoho z dvojčat nachází malá část kostní dřeně

79
druhého dvojčete, která produkuje bílé krvinky nesoucí DNA tohoto
dvojčete. Někdy je tento jev jednostranný (cizorodá linie buněk je
pouze v jednom dvojčeti), někdy vzájemný (cizorodou linii mají obě
dvojčata).
Z forenzního hlediska je to jev méně významný, protože množství
cizorodých krvinek je v poměru k množství vlastních krvinek
nepatrné a ve výsledku tato „příměs“ nijak zásadně neovlivňuje
identifikaci analýzou DNA.
Co je to maternální a fetální mikrochimérismus?
Pod tímto neprůhledným pojmem se skrývá další z mechanismů
vzniku chimérismu. Do embrya se během nitroděložního vývoje
mohou implementovat a trvale v něm přežívat buňky matky a
naopak, buňky plodu mohou v tomto období trvale kolonizovat
tělo matky. Odborné studie prokázaly přetrvávání fetálních buněk
v těle matky i po více než 50 letech od porodu. Uvedené
mechanismy tedy prokazatelně existují, avšak jejich přesné příčiny,
principy a též dopady na organismus jsou zatím předmětem
zkoumání. Z forenzně genetického hlediska má maternální/fetální
mikrochimérismus jen omezený význam. Experimentálně bylo
například potvrzeno, že v některých případech lze díky tomuto jevu
zjistit haplotyp chromozómu Y určitého muže rozborem krve jeho
matky. Ve standardní praxi se však tyto analýzy nepoužívají.
Může chimérismus vzniknout uměle?
Ano. Vznik chimérického jedince je důsledkem některých
moderních lékařských zákroků, při nichž jsou do organismu
pacienta vpraveny živé buňky jiného člověka. Trvale chiméričtí
jedinci vznikají v důsledku transplantací orgánů od dárce (ať už
biologického příbuzného, či osoby zcela nepříbuzné). Výběr
vhodného dárce se řídí tzv. tkáňovou kompatibilitou, tedy shodou

80
v tkáňových antigenech. Ty nemají nic společného s genetickým
profilem užívaným k forenzně genetické identifikaci, a dárce a
příjemce se tak genetickým profilem liší.
Z forenzního hlediska nejzávažnější důsledky má bezesporu
transplantace kostní dřeně, při níž jsou pacientovi vpraveny
krvetvorné buňky kostní dřeně od dárce. Taková osoba potom má
jiný genetický profil v krvi a prokrvených orgánech než kupříkladu
v bukálním stěru či spermatu. To může např. vést k falešnému
vyloučení osoby jako původce krevní stopy, je–li porovnávána se
vzorkem bukálního stěru. A naopak: taková stopa může být
nesprávně ztotožněna s osobou, která byla dárcem kostní dřeně.
Určení příbuznosti
Co je to biologická příbuznost?
Biologickou příbuzností dvou osob ve forenzní genetice rozumíme
jednak stav, kdy jedna z těchto osob je přímým předkem druhé,
nebo stav, kdy tyto dvě osoby mají blízkého společného předka.
V prvém případě se typicky jedná o dvojice rodič–dítě, prarodič–
vnouče, případně praprarodič–pravnouče. Ve druhém případě jsou
to nejčastěji dvojice sourozenecké a polovičně sourozenecké, a dále
dvojice strýc/teta – synovec/neteř, dvojice bratranecké, sestřenické
a dvojice bratranec–sestřenice.
Co je to stupeň příbuznosti?
Stupněm příbuznosti dvou osob vyjadřujeme, jak velké procento
autozomální genetické informace těchto osob je shodného původu.
Absolutní příbuznost představuje stav, kdy 100 % autozomální
genetické informace je shodného původu. V absolutní příbuznosti
jsou teoreticky pouze identická (jednovaječná dvojčata).

81
První stupeň příbuznosti představuje stav, kdy polovina, tj. 50 %
autozomální genetické informace je shodného původu. V prvním
stupni příbuznosti jsou dvojice rodič–dítě a úplné sourozenecké
dvojice.
Druhý stupeň příbuznosti představuje stav, kdy čtvrtina, tj. 25 %
autozomální genetické informace je shodného původu. Ve druhém
stupni příbuznosti jsou dvojice prarodič–vnouče, poloviční
sourozenecké dvojice a dvojice biologický strýc/biologická teta –
synovec/neteř.
Třetí stupeň příbuznosti představuje stav, kdy osmina, tj. 12,5 %
autozomální genetické informace je shodného původu. Ve třetím
stupni příbuznosti jsou dvojice praprarodič–pravnouče, dvojice
poloviční biologický strýc/ poloviční biologická teta –
synovec/neteř a dvojice bratranecké, sestřenické a dvojice
bratranec–sestřenice.
Jaké genetické znaky jsou pro určení příbuznosti
používány?
K posouzení příbuznosti osob užíváme v zásadě čtyř skupin DNA
lokusů:
1. lokusy autozomální, děděné dle mendelovských pravidel, tj.
z matky i z otce na všechny děti
2. X–chromozomální haplotyp, děděný z matky na všechny děti
a z otce pouze na dcery
3. Y–chromozomální haplotyp, děděný výhradně z otce na syny
4. mitotyp (lokusy mtDNA), děděný z matky na všechny děti.
Jak je vidět, každá z těchto čtyř skupin lokusů se dědí podle jiných
pravidel. Vhodné lokusy pro testování vybíráme tedy na základě
toho, jaký příbuzenský vztah chceme ověřit či vyvrátit.

82
Jak lze předpokládaný biologický vztah ověřit?
Máme–li určitou konkrétní hypotézu, tj. předpokládáme–li mezi
dvěma osobami určitý konkrétní biologický vztah (např. otec–syn),
víme, jakou měrou by se u něj měly shodovat jednotlivé výše
zmíněné typy DNA lokusů. Provedeme analýzu a posoudíme, jak
dobře zjištěná míra shody odpovídá míře očekávané.
Někdy nemáme o příbuzenství dvou osob jen jednu, ale hned dvě či
více alternativních hypotéz. Například: paní Novákové se narodil
syn Adam, jehož otcem může být manžel paní Novákové Jiří Novák
nebo jeho bratr Jindřich Novák nebo soused Novákových pan Josef
Svoboda. O dvojici Adam Novák – Jiří Novák tedy můžeme říci, že
mohou být a) otec a syn, b) strýc a synovec nebo c) nepříbuzné
osoby. V takovém případě opět posoudíme, jak zjištěná míra shody
odpovídá míře očekávané u jednotlivých hypotéz.
V extrémních případech nemáme o povaze biologického vztahu
mezi dvěma osobami žádnou informaci, pouze nás zajímá, zda jsou
tyto osoby biologicky příbuzné či nikoli. Zde je třeba upozornit, že
na jisté úrovni jsou samozřejmě navzájem biologicky příbuzní
všichni žijící lidé, a dotaz po vzájemné příbuznosti je tedy nutné
omezit jen na určitý okruh možných příbuzenství – tedy např. na
příbuzenství do třetího stupně příbuznosti. V takovém případě tedy
postupujeme tak, že testováním ověříme všechny biologické vztahy,
které připadají u dané dvojice v úvahu.
Jaká je očekávaná shoda u vybraných příbuzenských
dvojic?
Následující odstavce nejsou plným výčtem, ale představují
nejběžnější posuzované biologické vztahy a příslušnou shodu znaků
DNA.

83
matka – dcera
50 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, z toho jeden je shodný
původem
mitotyp je shodný původem
matka – syn
50 % autozomální DNA je shodné původem
matka má dva X chromozómy, syn jeden, a to shodný původem
s jedním z matčiných X chromozómů
mitotyp je shodný původem
otec – dcera
50 % autozomální DNA je shodné původem
dcera má dva X chromozómy, otec jeden, a to shodný původem
s jedním z dceřiných X chromozómů
mitotyp není shodný původem
otec – syn
50 % autozomální DNA je shodné původem
otec i syn mají po jednom X chromozómu, tyto nejsou shodné
původem
otec i syn mají po jednom Y chromozómu, tyto jsou shodné
původem
mitotyp není shodný původem
sestra – sestra
v průměru 50 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, jeden nebo oba jsou
shodné původem
mitotyp je shodný původem

84
sestra – bratr
v průměru 50 % autozomální DNA je shodné původem
sestra má dva X chromozómy, bratr jeden, tento může a nemusí
být shodný původem s jedním ze sestřiných X chromozómů
mitotyp je shodný původem
bratr – bratr
50 % autozomální DNA je shodné původem
oba mají po jednom X chromozómu, tyto mohou a nemusí být
shodné původem
oba mají po jednom Y chromozómu, tyto jsou shodné původem
mitotyp je shodný původem
babička z matčiny strany – vnučka
v průměru 25 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, z toho jeden může a
nemusí být shodný původem
mitotyp je shodný původem
babička z matčiny strany – vnuk
v průměru 25 % autozomální DNA je shodné původem
babička má dva X chromozómy, vnuk jeden, tento může a
nemusí být shodný původem s jedním z babiččiných X
chromozómů
mitotyp je shodný původem
dědeček z matčiny strany – vnučka
v průměru 25 % autozomální DNA je shodné původem
vnučka má dva X chromozómy, dědeček jeden, tento může a
nemusí být shodný původem s jedním z vnuččiných X
chromozómů
mitotyp není shodný původem
dědeček z matčiny strany – vnuk
v průměru 25 % autozomální DNA je shodné původem

85
oba mají po jednom X chromozómu, tyto mohou a nemusí být
shodné původem
oba mají po jednom Y chromozómu, tyto nejsou shodné
původem
mitotyp není shodný původem
babička z otcovy strany – vnučka
v průměru 25 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, z toho jeden je shodný
původem
mitotyp není shodný původem
babička z otcovy strany – vnuk
v průměru 25 % autozomální DNA je shodné původem
babička má dva X chromozómy, vnuk jeden, tento není shodný
původem ani s jedním z babiččiných X chromozómů
mitotyp není shodný původem
dědeček z otcovy strany – vnučka
v průměru 25 % autozomální DNA je shodné původem
vnučka má dva X chromozómy, dědeček jeden, tento není
shodný původem ani s jedním z vnuččiných X chromozómů
mitotyp není shodný původem
dědeček z otcovy strany – vnuk
25 % autozomální DNA je shodné původem
oba mají po jednom X chromozómu, tyto nejsou shodné
původem
oba mají po jednom Y chromozómu, tyto jsou shodné původem
mitotyp není shodný původem
nevlastní sestry (společná matka)
v průměru 25 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, z toho jeden nebo žádný je
shodný původem

86
mitotyp je shodný původem
nevlastní sestry (společný otec)
v průměru 25 % autozomální DNA je shodné původem
obě mají po dvou X chromozómech, z toho jeden je shodný
původem
mitotyp není shodný původem
nevlastní sestra a bratr (společná matka)
v průměru 25 % autozomální DNA je shodné původem
sestra má dva X chromozómy, bratr jeden, tento může a nemusí
být shodný původem s jedním ze sestřiných X chromozómů
mitotyp je shodný původem
nevlastní sestra a bratr (společný otec)
v průměru 25 % autozomální DNA je shodné původem
sestra má dva X chromozómy, bratr jeden, tyto nejsou shodné
původem
mitotyp není shodný původem
nevlastní bratři (společná matka)
v průměru 25 % autozomální DNA je shodné původem
oba mají po jednom X chromozómu, tyto mohou a nemusí být
shodné původem
oba mají po jednom Y chromozómu, tyto nejsou shodné
původem
mitotyp je shodný původem
nevlastní bratři (společný otec)
v průměru 25 % autozomální DNA je shodné původem
oba mají po jednom X chromozómu, tyto nejsou shodné
původem
oba mají po jednom Y chromozómu, tyto jsou shodné původem
mitotyp není shodný původem

87
Obrázek 22 – Rodokmen ilustrující stupeň příbuznosti běžných biologických příbuzenských vztahů. Osobu, k níž se porovnání vztahuje, označujeme běžně termínem proband (tak použito i v dalších obrázcích).
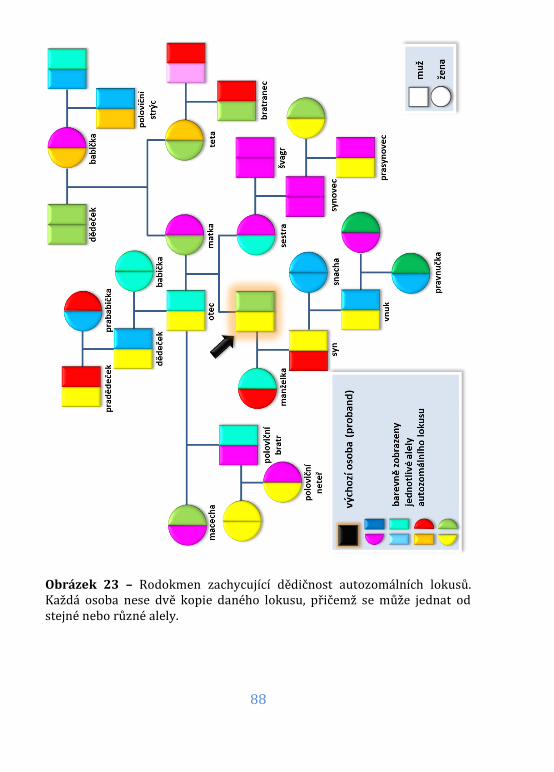
88
Obrázek 23 – Rodokmen zachycující dědičnost autozomálních lokusů. Každá osoba nese dvě kopie daného lokusu, přičemž se může jednat od stejné nebo různé alely.

89
Obrázek 24 – Rodokmen zachycující dědičnost X–chromozomálních lokusů. Osoby ženského pohlaví nesou vždy dvě kopie téhož lokusu, osoby mužského pohlaví pouze jednu.

90
Obrázek 25 – Rodokmen zachycující dědičnost Y–chromozomálních lokusů. Osoby ženského pohlaví nenesou žádnou kopii téhož lokusu, osoby mužského pohlaví nesou kopii jednu.

91
Obrázek 26 – Rodokmen zachycující dědičnost mitochondriální DNA. Každá osoba nese určitý mitotyp (vyjma osob s tzv. heteroplazmíí, o nichž bude pojednáno později).

92
Existují nějaké situace, kde výše uváděné míry shody
neplatí?
Ano. Námi popsané míry shody jsou platné jen tehdy, když daná
dvojice osob je právě a pouze v uvedeném vztahu a tyto osoby
nejsou zároveň příbuzné ještě jinak. Co máme na mysli?
V praxi se poměrně často setkáme s takzvaným příbuzenským
křížením neboli inbreedingem, který má významný vliv na
očekávané míry shody genetických znaků. Inbreeding zahrnuje
celou škálu jevů: jeho nejtvrdší formou je incest, tj. příbuzenské
křížení mezi otcem a dcerou, matkou a synem, dědem a vnučkou,
babičkou a vnukem nebo mezi sourozenci. O něco mírnější formou
inbreedingu je konsangvinita neboli pokrevní příbuznost –
typickým příkladem budiž manželství (a společné rodičovství)
bratrance a sestřenice. Za nejmírnější stupeň inbreedingu pak
můžeme považovat křížení osob vzdáleně příbuzných, tak jak
k němu běžně docházelo po dlouhá staletí i na našem území, a to
zejména v izolovanějších subpopulacích. Partner pro uzavření
manželství byl obvykle vybírán v té samé vesnici nebo v okruhu
několika málo vesnic, což při dlouhodobém systematickém
uplatňování této strategie vedlo k významnému zvýšení koeficientu
příbuznosti obyvatel daného regionu, zvlášť přičteme–li k tomu
ještě „posilující efekt“ skrytě konsangvinních svazků (manželé
mohli být často nevlastní sourozenci, aniž to tušili, v důsledku
nevěry svých rodičů).
Při konstruování možných hypotéz biologické příbuznosti a
určování očekávané míry shody genetických znaků je tedy třeba
vždy zohlednit případnou předpokládanou či přímo
potvrzenou konsangvinitu, či dokonce incest. Správné určení
očekávané míry shody může být u složitějších či vícestupňově
konsangvinních hypotéz opravdu nelehkou úlohou.

93
Obrázek 27 – Rodokmen ilustrující zvýšený stupeň příbuznosti osob v rodině, kde došlo jak ke konsangvinnímu sňatku bratrance a sestřenice (značen modrou úsečkou), tak k nechtěně incestnímu sňatku probanda s nevlastní sestrou (značen červenou úsečkou).

94
Byl posuzován případ možného incestního vztahu, kdy ze zjištěných
údajů vyplývalo, že muž (68) měl se svojí sestřenicí dceru, s touto
dcerou měl další dceru (tedy zároveň vnučku) a s touto dcerou–
vnučkou zplodil syna (tedy zároveň vnuka a pravnuka). Pomineme–li
trestně právní a sociální rozměr případu, je tato kauza z hlediska
stanovení správných očekávání genetické shody velmi zajímavá.
Obrázek 28 – Rodokmen ilustrující extrémní stupeň příbuznosti osob u mnohotně incestního případu (popis případu viz žlutý box výše).

95
Predikce biogeografického původu a
fenotypových znaků osoby
Co je etnický původ?
Pojem etnický původ používáme v souvislosti s pojmenováním
příslušnosti konkrétního jedince k určitému lidskému subtypu,
subpopulaci, skupině. Zcela zásadní skutečností, kterou je třeba si
v této souvislosti uvědomit a ujasnit, je, že skupiny označované
jako rasa, národ, etnikum jsou definovány primárně socio–
kulturně, nikoli antropologicky či biologicky.
Zůstává ovšem zjevné, že mezi skupinami lidí na Zemi existují
markantní biologické rozdíly – osoba původem z Afriky má jiné
biologické vlastnosti než osoba původem z Asie. V rámci biologie a
antropologie proto používáme termín biogeografický původ
osoby. Ten zahrnuje a vystihuje jak biologické (a tedy i genetické)
rozdíly mezi jednotlivými populacemi, tak geografické rozmístění
těchto populací.
Co je predikce biogeografického nebo etnického
původu analýzou DNA?
Predikcí biogeografického původu molekulárně genetickými
metodami rozumíme určení, do jaké lidské subpopulace osoba patří,
na základě analýzy DNA. Vzhledem k tomu, že – jak bylo řečeno
o odstavec výše – lidské subpopulace jsou obvykle definovány a
tedy i označovány především na základě sociokulturních souvislostí,
je praktický potenciál analýzy DNA v této oblasti limitovaný.
Zřejmé je, že vysoce spolehlivých výsledků dosahujeme tehdy,
pokud se snažíme o rozlišení „v nejhrubším měřítku“, tj. o určení,
zda osoba je evropského, asijského, afrického původu. Čím jemnější
rozlišení požadujeme, tím větší statistickou chybou je analýza

96
zatížena a tím méně se lze na její výsledek spoléhat. Kupříkladu
rozlišení mezi jednotlivými blízkými národy (Čech vs. Slovák) je
prakticky nemožné, přestože statisticky mezi nimi jisté drobné
rozdíly existují.
Současně je potenciál této analýzy významně ovlivněn dvěma
dalšími faktory: zaprvé mírou mobility obyvatelstva (např. pohyb
obyvatelstva v Evropě je významně menší než v USA) a zadruhé
podílem smíšených partnerství, tedy partnerství osob z původně
geneticky výrazně vzdálených a odlišných populací – typicky např.
žena Nativní Američanka (my bychom ji označili jako Indiánku) a
muž Afroameričan (my bychom jej označili jako černocha) – opět je
podíl těchto partnerství v Evropě významně nižší než v USA.
Národ je typickou socio–kulturně a geopoliticky definovanou
skupinou – příslušníci národa sdílejí především jazyk, kulturu,
území a historii; pochopitelně řada z nich sdílí i biogeografický
původ, ovšem toto kritérium není pro národ primární a určující.
Zatímco u národů obývajících dlouhodobě významně izolovaná
teritoria (např. ostrovy) je společný biogeografický původ silný,
u jiných tomu tak není; kupříkladu my Češi jsme typickým národem
dlouhodobě obývajícím „průchozí území“, což se odráží v našem
genofondu: lakonicky řečeno, v různých obdobích naší historie do
něj přispěl významně kde kdo (Slované, Hunové, Kelti, Turci,
Švédové, Italové, Francouzi, Němci) a hledat tak dnes jakési
„genetické češství“ je beznadějné.
Co je fenotypový znak?
Fenotypovým znakem rozumíme libovolnou vlastnost organismu
– v tomto případě člověka, která je zjevná nebo zjistitelná (např.
biochemickou analýzou). Můžeme sem zahrnout znaky tělesné
(výška, váha, tvar lebky, barva vlasů, barva očí atd.), znaky
biochemické a fyziologické (krevní skupina, rychlost „spalování“

97
glukózy atd.) i třeba znaky neurologické a psychické (svalové
napětí, motorika chůze, temperament, inteligence atd.).
Co je predikce fenotypu analýzou DNA?
Predikcí fenotypu molekulárně genetickými metodami rozumíme
určení, jak daný znak u osoby vypadá, na základě analýzy DNA. To je
pochopitelně nesmírně zajímavé především z investigativního
hlediska – moci z biologické stopy získat popis osoby, která ji
zanechala, je snem všech kriminalistů.
Jaké znaky jsou pro takovouto predikci nejvhodnější?
Především ty, které
1. jsou stálé v čase, tj. relativně neměnné (např. tělesná výška
je u dospělé osoby mnohem stálejší znak než tělesná hmotnost)
2. jsou zjevné i při běžném kontaktu s osobou, a lze je tedy
zjistit bez složitých vyšetření, a často dokonce bez vědomí
dotyčné osoby (např. barvu očí zjistíme pouhým pohledem,
zatímco krevní skupinu osoby musíme vyšetřit sérologicky či
geneticky)
3. jsou dostatečně variabilní, díky čemuž můžeme na základě
daného znaku osoby rozlišit na ty, které se se stopou shodují, a
ty, které ne (např. barva očí je mnohem variabilnější znak než
počet očí)
4. jsou silně geneticky determinované a mohou být analýzou
DNA predikovány s vysokou spolehlivostí (např. již zmíněná
barva očí je mnohem snáze predikovatelný znak než tvar a
rozměry úst).

98
Co tedy lze v současnosti z biologické stopy s relativně
velkou spolehlivostí predikovat?
1. Pohlaví (prakticky s jistotou);
2. Biogeografický původ v hrubém měřítku;
3. Přirozenou barvu očí (v hrubém rozlišení – např. modrá/
hnědá);
4. Přirozenou barvu vlasů (v hrubém rozlišení – např.
blond/rezavá/tmavá);
5. Přirozenou barvu pokožky (v hrubém rozlišení – např. velmi
světlá/velmi tmavá);
Vyjma určení pohlaví – což je rutinní záležitost – byly dosud ve
světě tyto predikce provedeny přímo z forenzních vzorků jen
v několika málo kriminálních případech a příslušné metody jsou
zatím spíše vyvíjeny a ověřovány. Problematice se v minulých letech
intenzivně věnovala americká společnost DNAprint Genomics,
sídlící na Floridě, která vytvořila první komerčně dostupné testy
predikce etnicity a fenotypových znaků; po určité době však tyto své
původně nadějné projekty revidovala, nabízené produkty stáhla a
v roce 2009 ukončila svou činnost.
Které forenzně zajímavé fenotypové znaky dosud nelze
ze stopy věrohodně predikovat?
Dosud zůstávají zcela nepredikovatelné či velmi nespolehlivě
predikovatelné například stáří osoby, tělesná výška a rysy
obličeje.

99
Obecné principy forenzně genetické
analýzy
Principy identifikace
Na základě analýzy jakých lokusů v DNA lze osobu
identifikovat?
Pro identifikace jsou užívány lokusy polymorfní až vysoce
polymorfní, tedy ty, u nichž existuje v populaci několik (často i
desítky) možných variant – alel. Právě díky existenci mnoha variant
se osoby v populaci navzájem velmi často liší tím, kterou konkrétní
alelu či alely ve své DNA nesou.
Co jsou repetitivní sekvence DNA?
Jedněmi z nejpolymorfnějších lokusů lidské (a nejen lidské) DNA
jsou tzv. repetitivní sekvence. Jsou to takové sekvence, které se
v genomu nevyskytují pouze jednou, ale opakovaně. Tedy – určitý
motiv (určitá přesná sekvence) se v genomu objevuje vícekrát.
Pokud jsou jednotlivé motivy v DNA zařazeny těsně za sebou –
v tandemu, nazýváme takový úsek DNA tandemová repetice.
Pokud se jednotlivá opakování motivu vyskytují rozptýleně na
různých místech genomu, a to třeba i na různých chromozómech,
mluvíme o repetici rozptýlené.
Které repetitivní sekvence jsou forenzně využívány pro
identifikaci?
V počátcích forenzní genetiky byly k identifikačním analýzám
užívány tzv. dlouhé tandemové repetice s variabilním počtem
opakování, označované jako VNTR (angl. Variable Number of
Tandem Repeats) neboli minisatelity. V současnosti jsou pro

100
identifikaci zdaleka nejužívanějšími lokusy tzv. krátké tandemové
repetice, označované jako STR (angl. Short Tandem Repeats) neboli
mikrosatelity.
Co jsou minisatelity neboli VNTR?
Minisatelity neboli VNTR (angl. Variable Number of Tandem
Repeats) čili dlouhé tandemové repetice s variabilním počtem
opakování jsou repetitivní sekvence s typickou délkou základního
motivu 10–100 nukleotidů; tento motiv se vyskytuje na určitém
místě DNA v jednotkách až stovkách opakování těsně za sebou.
Jednotlivé alely minisatelitu se liší počtem opakování základního
motivu, a tedy délkou.
Dejme tomu, že určitá VNTR repetice lidského genomu má délku
základního motivu 17 nukleotidů, přičemž motiv se může opakovat
70–450krát. Nejkratší alela této repetice je tedy dlouhá 1190
nukleotidů a nejdelší alela 7650 nukleotidů.
VNTR lokusy se vyskytují na mnoha místech genomu a byly prvními,
které byly pro účely identifikace využity. Dnes už jsou postupy
založené na jejich analýze překonány a pro identifikace se od
poloviny 90. let 20. století prakticky neužívají, a proto se jim
nebudeme již podrobněji věnovat.
Co jsou mikrosatelity neboli STR?
Mikrosatelity neboli STR (angl. Short Tandem Repeats) čili krátké
tandemové repetice (označované někdy též SSR – angl. Simple
Sequence Repeat) jsou repetitivní sekvence s typickou délkou
základního motivu 2–9 nukleotidů; tento motiv se vyskytuje na
určitém místě DNA v jednotkách až desítkách opakování těsně za
sebou. Jednotlivé alely mikrosatelitu se tak stejně jako
u minisatelitů liší počtem opakování základního motivu, a tedy

101
délkou, nicméně celková délka repetice je u STR výrazně kratší než
u VNTR.
Dejme tomu, že určitá STR repetice lidského genomu má délku
základního motivu 4 nukleotidy, přičemž motiv se může opakovat 8–
15krát. Nejkratší alela této repetice je tedy dlouhá 32 nukleotidů a
nejdelší alela 60 nukleotidů. (Prosím srovnejte s délkami nejkratší a
nejdelší alely typické VNTR, uvedenými v textu o něco výše).
Obrázek 29 – Struktura krátké tandemové repetice (STR) neboli mikrosatelitu.
STR lokusy se vyskytují na všech lidských chromozómech včetně
pohlavních chromozómů X a Y, nenajdeme je pouze
v mitochondriální DNA. Jsou jich známy tisíce a tvoří cca 3 %

102
lidského genomu, nicméně zdaleka ne všechny tyto mikrosatelity
jsou variabilní.
Jak mikrosatelity (STR lokusy) a jejich alely
klasifikujeme?
Jednotlivé mikrosatelity mohou mít systematická i vžitá (triviální)
označení. Obecně mikrosatelity objevené a k analýzám užívané již
dávno mají spíše označení triviální, vycházející obvykle z názvu
blízké genové sekvence – např. TPOX, FGA, vWA apod. Lokusy
„novější“ jsou označovány systematickým názvem – např. D21S11,
D19S433 či D5S818, ne však vždy (např. relativně nově užívané
pětinukleotidové lokusy jsou známy pod názvem Penta E a Penta D).
Jak již bylo řečeno výše, mikrosatelity mohou mít délku základního
motivu 2–9 nukleotidů, a právě podle ní klasifikujeme jednotlivé
mikrosatelity na di–, tri–, tetra–, penta–, hexa–, hepta–, okta– a
nonanukleotidové. Pro forenzní analýzy lidí jsou využívány
prakticky výhradně STR lokusy tetra– nebo pentanukleotidové,
tedy ty se základním motivem dlouhým 4 nebo 5 nukleotidů.
U repetic s kratším motivem (2 či 3 nukleotidy) může při analýze
docházet ve velké míře k těžko odstranitelnému nežádoucímu jevu,
zvanému stuttering (prokluz polymerázy, jev je podrobněji popsán
v kapitolách pojednávajících o technologii forenzně genetické
analýzy), repetice s delším motivem (6–9 nukleotidů) jsou již zase
celkově dosti dlouhé (čtyřnukleotidová repetice s 20 opakováními
motivu měří 80 nukleotidů, zatímco osminukleotidová repetice se
stejným počtem opakování měří 160 nukleotidů), což může vést
k problémům s jejich analýzou u vzorků s degradovanou DNA.
Jednotlivé alely STR lokusů označujeme číslem vyjadřujícím
počet opakování základního motivu u této konkrétní alely. Tedy –
alela 7 představuje takovou variantu daného STR lokusu, v níž se
základní motiv opakuje 7krát.

103
Některé alely však nejsou tvořeny celým počtem opakování
základního motivu: lokus je tvořen například 7 celými opakováními
čtyřnukleotidového motivu, za nimiž následují ještě dva nukleotidy
z osmého opakování (tj. osmé opakování motivu není celé, ale
v repetici jsou z něj přítomny jen dva nukleotidy). Takové alely
označujeme jako mikrovarianty a značíme je počtem úplných
opakování základního motivu, za nímž následuje tečka a počet
„nadbytečných“ nukleotidů – v našem příkladu tedy 7.2.
Obrázek 30 – Alely STR lokusu – standardní a mikrovariantní.
Jak byly vybrány konkrétní STR lokusy (mikrosatelity)
pro forenzní testování?
Pro potřeby forenzního testování byly zvoleny takové lokusy, které
mají kromě již výše zmíněné vhodné délky základního motivu (4

104
nebo 5 nukleotidů) také velkou variabilitu, tj. co největší počet
existujících alel, a pokud možno vyvážené frekvence těchto alel.
Nutno říci, že ne všechny z testovaných lokusů tuto podmínku
splňují a v základním setu testovaných lokusů zůstávají zejména
z historických důvodů.
Další důležitou podmínkou je, aby jednotlivé testované lokusy
nebyly v tzv. genetické vazbě – laicky řečeno, aby v DNA nebyly
příliš blízko sebe. Ideálně jsou vybírány do testování STR lokusy
lokalizované na různých chromozómech, a pokud už jsou na témže
chromozómu, pak dostatečně daleko od sebe. (Tento princip neplatí
u STR lokusů na pohlavních chromozómech X a Y, o kterých bude
později pojednáno samostatně).
Principiálně je důležité, aby po celém světě byly pro forenzní
analýzy používány pokud možno stejné lokusy – jedině tak lze
data získaná v různých laboratořích vzájemně srovnávat (u dvou
vzorků – např. stopy a srovnávacího vzorku – lze porovnat vždy
pouze výsledky analýzy stejných lokusů).
V současné době existuje pro identifikaci několik oficiálně
doporučených setů autozomálních STR lokusů, které se v mnoha
zařazených lokusech shodují. Jsou to:
The CODIS STR Loci – 13 STR lokusů jako základní standardní
set pro oblast USA;
The Expanded U.S. Core Loci – 18 STR lokusů (13 původních
+ 5 nových) jako rozšířený standardní set pro oblast USA;
The European Standard Set (ESS) – 7 STR lokusů jako
základní standardní set pro Evropu; je totožný s The Interpol
Standard Set of Loci (ISSOL);
The Extended European Standard Set (extended ESS) –
12 STR lokusů (7 původních + 5 nových) jako rozšířený
standardní set pro Evropu.

105
Pro analýzu Y chromozomálních STR lokusů rovněž existují
doporučené sety těchto markerů – hovoříme o tzv. minimálním a
rozšířeném Y–STR haplotypu. Minimální haplotyp označovaný
minHt zahrnuje 8 Y–STR lokusů, rozšířený pak dalších 8 Y–STR
lokusů.
Pro analýzu X chromozomálních STR lokusů tak významná
doporučení neexistují, v rámci forenzních analýz je otestováno
celkem cca 33 různých X–STR a výběr závisí především na
rozhodnutí dané laboratoře.
Jak vypadá genotyp STR lokusu konkrétního jedince?
U STR lokusu, který se nachází na nepohlavních chromozómech
(autozómech), má konkrétní jedinec dvě kopie daného lokusu, a
může tedy být buď heterozygotní (např. 11/12 – tj. má jednu alelu
11 a jednu 12), nebo homozygotní (např. 13/13 – tj. má dvě kopie
alely 13).
U STR lokusu, který se nachází na X chromozómu, je muž vždy
hemizygotní – má pouze jeden X chromozóm, a tedy pouze jednu
kopii daného lokusu (např. alelu 12), zatímco žena má dva
X chromozómy, a může tedy být heterozygotní (např. 12/14) nebo
homozygotní (např. 11/11).
U STR lokusu, který se nachází na Y chromozómu, je daný lokus
přítomen pouze u muže, a to v jedné kopii (má např. alelu 12). Toto
konstatování však neplatí u některých Y–STR lokusů, jež jsou na
Y chromozómu duplikovány (zdvojeny), přičemž každá ze dvou
kopií může obsahovat jinou alelu a muž pak má genotyp např.
11/14 (nejedná se však o heterozygozitu).

106
Obrázek 31 – Možné varianty srovnání genotypu stopy a osoby: neshoda, náhodná shoda nebo zákonitá shoda genotypu.
Jak probíhá identifikace (určení shody) pomocí
analýzy autozomálních STR lokusů?
Pokud mají dva biologické vzorky (např. krevní stopa a
srovnávací vzorek) téhož původce neboli stejný zdroj, pak bude
forenzně genetickou analýzou určitého STR lokusu v obou
vzorcích stanoven identický genotyp tohoto lokusu; tedy
kupříkladu je–li ve stopě zjištěn genotyp 11/15, pak i v bukálním
stěru jejího původce bude stanoven genotyp 11/15. (Bohužel, toto
tvrzení nemusí být vždy pravdivé a vzácně se objeví i situace, kdy se
genotyp dvou vzorků nemusí shodovat, přestože mají stejného
původce; jednou z nich je již dříve popsaný chimérismus, dále se
může jednat o tzv. mozaicismus – mutaci přítomnou pouze
v některých buňkách dané osoby.)

107
Naproti tomu, pokud mají dva biologické vzorky různého
původce, pak forenzně genetickou analýzou bude v poměrně
velkém procentu případů zjištěn rozdílný genotyp (např. stopa
11/15 a srovnávací vzorek 12/15). Může nicméně nastat situace,
kdy i u takovýchto vzorků s různým původcem bude zjištěn stejný
genotyp – tedy např. u obou 13/14), a to proto, že oba původci se
koincidencí, náhodou shodují v genotypu testovaného lokusu.
Je tedy zřejmé, že testování jediného STR lokusu nám nepomůže
otázku na společný respektive rozdílný původ obou vzorků
spolehlivě zodpovědět. Proto je při forenzně genetické analýze
současně testován větší počet STR lokusů (v ČR obvykle typicky
15 autozomálních STR lokusů), čímž se náhodná shoda dvou osob
stává extrémně nepravděpodobnou a testování má vysokou
vypovídací hodnotu.
Analýza setu autozomálních STR lokusů (a současně s ní
prováděná analýza pohlaví) je nejfrekventovanějším postupem
současné kriminalistické a identifikační forenzní genetiky. Své
uplatnění nachází v nejrůznějších typech kriminálních případů či
identifikačních úkonů a může v principu vést až k individuální
identifikaci zůstavitele biologického materiálu.
Jak probíhá identifikace (určení shody) pomocí
analýzy X–chromozomálních STR lokusů?
Pokud mají dva biologické vzorky téhož původce neboli stejný
zdroj, pak se u nich bude shodovat i genotyp X–
chromozomálních STR lokusů; pro přímou identifikaci se ovšem
analýza X–chromozomálních markerů prakticky nevyužívá, neboť
nepřináší žádný benefit. Je však velmi zajímavým nástrojem při
určování některých typů biologických vztahů – viz část věnovaná
principům určení příbuznosti.

108
Jak probíhá identifikace (určení shody) pomocí
analýzy Y–chromozomálních STR lokusů?
Pokud mají dva biologické vzorky téhož původce neboli stejný
zdroj, pak se u nich bude shodovat i haplotyp Y–
chromozomálních STR lokusů; opět ovšem nelze vyloučit možnost
existence drobné odchylky, způsobené mutací.
Y–chromozóm je předáván z otce na všechny syny, a to
v nezměněné podobě, proto tuto analýzu samu o sobě nelze
použít k individuální identifikaci osoby. Při identifikacích tedy
tuto analýzu užíváme v některých specifických situacích, kdy je
analýza autozomálních STR neinformativní: typicky je to analýza
stopy tvořené ženským biologickým materiálem s malou příměsí
materiálu mužského, kdy analýza autozomálních STR zpravidla není
schopna detekovat alely minoritní složky směsi, tj. alely muže.
Takovými typickými vzorky jsou
1. poševní stěr po znásilnění, při němž nedošlo k ejakulaci
pachatele do pochvy poškozené (anebo je pachatel
azoospermický, tj. nemá v ejakulátu spermie),
2. vzorek materiálu zpoza nehtů ženy fyzicky napadené mužem.
Analýzou Y–STR v těchto případech sice nedosáhneme individuální
identifikace pachatele, ale v případě neshody haplotypů můžeme
podezřelou osobu prakticky s jistotou vyloučit.
Jaké další lokusy kromě repetic jsou forenzně
využívány pro identifikaci?
Dalšími lokusy, které lze využít pro identifikaci, jsou tzv.
jednonukleotidové polymorfismy, označované jako SNP (angl.
Single Nucleotide Polymorphisms). Konečně v určitých specifických
případech je pro skupinovou identifikaci osob užívána analýza tzv.

109
hypervariabilních oblastí mitochondriální DNA, označovaných
jako HVR mtDNA (angl. Hypervariable Region of mitochondrial
DNA).
Co jsou jednonukleotidové polymorfismy neboli SNP?
Jednonukleotidové (bodové) polymorfismy jsou ve své podstatě
nejjednodušším možným polymorfismem, jaký může v DNA
existovat. V určitém konkrétním místě DNA se u jedné osoby
vyskytuje jiný nukleotid než u druhé. Kupříkladu: zatímco osoba A
má v určitém místě sekvenci AATGGATA, osoba B má v tomtéž místě
sekvenci AATGGGTA; vidíme, že šestý nukleotid této sekvence je u
obou osob různý, a dané místo je tedy polymorfní. Protože tento
polymorfismus spočívá ve změně jediného nukleotidu sekvence,
hovoříme o jednonukleotidovém polymorfismu.
Obrázek 32 – Struktura jednonukleotidového polymorfismu (SNP).

110
SNP se vyskytují po celém lidském genomu s četností cca 1 na 1000
nukleotidů – u každého z nás lze tedy nalézt miliony takovýchto
polymorfismů.
Jak SNP lokusy a jejich alely klasifikujeme?
Jednotlivé SNP lokusy jsou zpravidla specifikovány
prostřednictvím svého referenčního čísla v databázi dbSNP, vedené
NCBI (National Center for Biotechnology Information). Označení
konkrétního SNP pak je kupříkladu rs1493232.
Jednotlivé alely SNP lokusů označujeme písmenem označujícím
příslušný nukleotid – tj. A, C, G nebo T.
Principiálně mohou u každého SNP lokusu existovat nanejvýše čtyři
možné alely: A, C, T a G (více základních typů nukleotidů v DNA
není). Valná část SNP lokusů je však tzv. bialelických – to znamená,
že v populaci se vyskytují pouze dvě varianty (alely) tohoto SNP;
například větší procento populace má v daném místě nukleotid C,
menší procento pak nukleotid A. Při popisu příslušného SNP
užíváme zápis rs1493232 C>A nebo rs1493232 C/A, kterým
vyjadřujeme, jaké alely se v tomto SNP vyskytují a která z nich je
frekventovanější. Nutno však říci, že nomenklatura SNP není
v tomto směru zcela jednotná a existují i jiné způsoby zápisu.
Jak vypadá genotyp SNP lokusu konkrétního jedince?
U SNP lokusu, který se nachází na nepohlavních chromozómech
(autozómech), má konkrétní jedinec dvě kopie daného lokusu,
a může tedy být buď heterozygotní (např. AT – tj. má jednu alelu A a
jednu T), nebo homozygotní (např. TT – tj. má dvě kopie alely T).
U SNP lokusu, který se nachází na X chromozómu, je muž vždy
hemizygotní – má pouze jeden X chromozóm, a tedy pouze jednu
kopii daného lokusu (např. alelu C), zatímco žena má dva X

111
chromozómy, a může tedy být heterozygotní (např. CG) nebo
homozygotní (např. GG).
U SNP lokusu, který se nachází na Y chromozómu, je daný lokus
přítomen pouze u muže, a to v jedné kopii (má např. alelu T).
Jak probíhá identifikace (určení shody) pomocí analýzy
SNP lokusů?
Princip identifikace pomocí SNP je prakticky shodný s principem
identifikace u STR: pokud předpokládáme, že dva vzorky mají
téhož původce, měl by v nich být stanoven i stejný genotyp SNP
lokusu. Mají–li různého původce, může a nemusí být v nich
stanovený genotyp shodný.
Poměrně podstatným rozdílem oproti STR, který si musíme
uvědomit, je malá alelická škála SNP lokusu: zatímco u typického
STR jsme hovořili o desítkách možných alel, u SNP mohou
z podstaty věci existovat nanejvýše čtyři alely, ale v praxi jsou
dokonce zpravidla užívány již zmíněné lokusy bialelické, tedy
s pouhými dvěma alelami. Důsledkem je podstatně větší
pravděpodobnost náhodné shody osoby se vzorkem, jehož
původcem je někdo jiný. Abychom tento efekt malé alelické škály
vyvážili, je pro dosažení potřebné míry spolehlivosti do
testování třeba zařadit mnohem více SNP lokusů, než tomu
bylo u STR, a to až desítky.
Přestože analýza SNP má prakticky stejný identifikační potenciál
jako analýza STR, v praxi je mnohem méně využívána. Hlavními
příčinami jsou zejména
1. Nutnost převést laboratoř (včetně celé metodiky a systému
kvality) na analýzu SNP, někdy i částečně převybavit laboratoř
technologicky (analýza základních forenzních SNP je

112
proveditelná na zařízeních používaných pro STR analýzu, celá
řada SNP aplikací je však založena na odlišných technologiích)
2. Nekompatibilita dat (pokud jsou v databázích uloženy STR
profily, pak SNP profil není s nimi porovnatelný, jedná se o jiný
identifikační údaj)
3. Problematičtější použitelnost pro analýzu směsí biologických
materiálů
4. Souvislost mnohých SNP s dědičnými chorobami.
Pro svůj velmi zajímavý potenciál v oblasti predikce
biogeografického původu a fenotypových znaků (viz dále) však
pravděpodobně budou v budoucnu SNP lokusy stále
vyhledávanějším cílem forenzně genetické analýzy.
Co jsou hypervariabilní oblasti mitochondriální DNA
neboli HVR mtDNA?
Mitochondriální DNA (mtDNA) je velmi specifickou částí dědičné
informace v buňce. Je to jednoduchá kruhová molekula DNA (nemá
tedy podobu chromozómu) a typově je stejná jako DNA bakterií.
Současné teorie předpokládají, že mitochondrie mají skutečně svůj
původ v bakteriích, které se postupně začlenily do pokročilých
eukaryotních buněk a staly se jejich nedílnou součástí.
MtDNA je z velké části tvořena kódujícími sekvencemi (geny),
extragenové DNA zde najdeme relativně velmi málo. Výjimkou je
oblast tzv. D–kličky (smyčky) (angl. D–loop), zvaná též kontrolní
(řídící) oblast (angl. control region). Zhruba uprostřed této oblasti
leží tzv. ori H – místo, odkud se mtDNA kopíruje, a po jeho obou
stranách se nacházejí nekódující úseky, v nichž najdeme velký počet
variabilních míst; proto tyto úseky označujeme jako
hypervariabilní a značíme HVR (angl. Hypervariable Region).
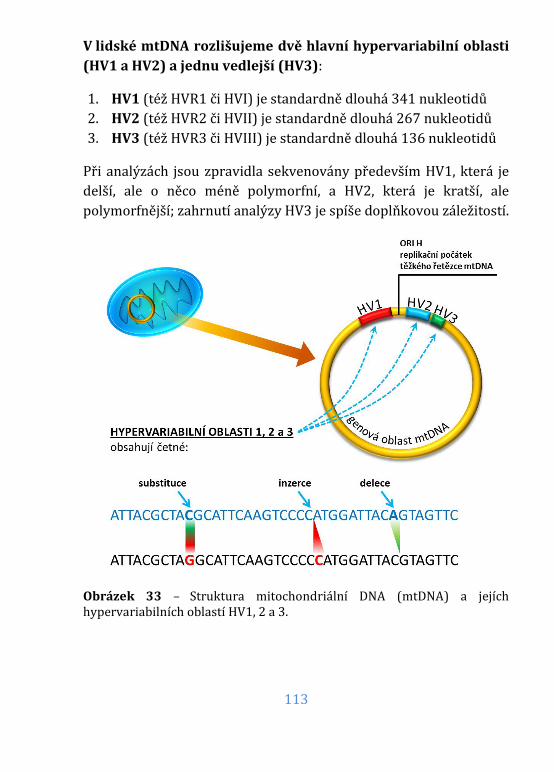
113
V lidské mtDNA rozlišujeme dvě hlavní hypervariabilní oblasti
(HV1 a HV2) a jednu vedlejší (HV3):
1. HV1 (též HVR1 či HVI) je standardně dlouhá 341 nukleotidů
2. HV2 (též HVR2 či HVII) je standardně dlouhá 267 nukleotidů
3. HV3 (též HVR3 či HVIII) je standardně dlouhá 136 nukleotidů
Při analýzách jsou zpravidla sekvenovány především HV1, která je
delší, ale o něco méně polymorfní, a HV2, která je kratší, ale
polymorfnější; zahrnutí analýzy HV3 je spíše doplňkovou záležitostí.
Obrázek 33 – Struktura mitochondriální DNA (mtDNA) a jejích hypervariabilních oblastí HV1, 2 a 3.

114
Jak vypadá HVR mtDNA konkrétního jedince , jak
popisujeme jednotlivé alely HVR mtDNA?
Pokud porovnáme sekvenci hypervariabilní oblasti mtDNA dvou
osob, najdeme zde zpravidla několik drobných rozdílů, kterými
mohou být:
1. Drobné záměny jednotlivých nukleotidů (tedy vlastně SNP)
– tzv. nukleotidové substituce: např. osoba A má v pozici 16126
nukleotid T, zatímco osoba B má v téže pozici nukleotid C.
2. Drobné délkové změny – inzerce (vřazení) či naopak delece
(vypadnutí) jednoho, dvou či tří nukleotidů: například osoba A
má v určitém místě HVR sekvenci AAACCCCTCCAT, osoba B
sekvenci AAACCCCTTCCAT a osoba C naopak jen
AAACCCTCCAT
Popsat stav hypervariabilní oblasti konkrétní osoby bychom
samozřejmě mohli tím, že bychom uvedli její kompletní sekvenci;
takový zápis by však byl pro svou délku a nepřehlednost značně
nepraktický. Postupujeme tedy tak, že zjištěnou sekvenci
konkrétní osoby srovnáme s referenční sekvencí, kterou
označujeme jako rCRS (angl. revised Cambridge Reference Sequence),
a zapíšeme odchylky od této sekvence, a to takto:
1. U substitucí uvedeme nukleotid, který je na dané pozici
přítomen místo standardního nukleotidu referenční sekvence,
tedy kupříkladu 16126C (= „na pozici 16126 je u vyšetřované
osoby nukleotid C“).
2. U delecí uvedeme pozici a „d“, „del“ nebo „–“, tedy kupříkladu
315d respektive 315del či 315– (= „u vyšetřované osoby
nukleotid na pozici 315 chybí“).

115
3. U inzercí uvedeme poslední standardní pozici, tečku, pořadí
inzertovaného nukleotidu a typ nukleotidu, tedy kupříkladu
211.1A (=“u vyšetřované osoby je za standardní pozicí 211
navíc začleněn nukleotid A“).
4. Za určitých okolností může být některý z nukleotidů analýzou
opakovaně „nečitelný“ (nelze zjistit, který nukleotid v dané
pozici je) – potom užijeme označení „N“ – například 16143N
(=“u vyšetřované osoby nebylo možno určit typ nukleotidu
v pozici 16143“).
Ve valné většině případů mají všechny tkáně i jednotlivé buňky
osoby stejnou sekvenci hypervariabilních oblastí mtDNA. Nicméně,
neplatí to vždy; relativně vzácně se můžeme setkat s jevem,
označovaným jako heteroplazmie. Je to jednak stav, kdy některé
skupiny buněk v rámci téže tkáně mají jinou sekvenci HVR než
jiné, a jednak stav, kdy se mitochondrie s rozdílnou sekvencí
HVR nacházejí společně přímo v rámci jednotlivých buněk.
Zpravidla se obě linie liší jediným zaměněným nukleotidem – danou
pozici pak zapíšeme např. 16126C/T, někdy se lze setkat i s
označením 16126Y (Y se použije pro heteroplazmii C/T, R se
použije pro heteroplazmii A/G).
Jak probíhá identifikace (určení shody) pomocí
analýzy HVR mtDNA?
Při porovnávání dvou vzorků vycházíme opět z předpokladu, že
mají–li tyto vzorky společného původce, měly by mít i stejnou
sekvenci hypervariabilních oblastí. Porovnáme tedy námi zjištěné
odchylky od standardní sekvence obou vzorků.
Zde je třeba znovu konstatovat a zdůraznit, že sekvence HVR
mtDNA nejsou z logiky věci markery individuálně identifikující:
jsou děděny v nezměněné podobě v maternální linii (po přeslici) a

116
všechny osoby v této linii mají – nedojde–li k mutaci – sekvence
mtDNA identické. Analýzou tedy nelze rozlišit např. mezi sourozenci
(pokud mají společnou matku), mezi matkou a jejími dětmi, mezi
vnukem a babičkou z matčiny strany atd.
V případě zjištění shody sekvencí obou vzorků proto pouze
společného původce obou vzorků nevyloučíme, ale rozhodně
nepotvrdíme.
Na druhou stranu, vzhledem k mnohonásobně vyšší mutační
rychlosti DNA mitochondrií než má jaderná DNA, a též vzhledem
k již zmíněné možnosti heteroplazmie existuje statisticky nenulová
možnost, že i dva vzorky stejného původce se mohou lišit
jedním nukleotidem. Proto teprve zjištění většího počtu rozdílů
(2 a více) v HVR sekvenci obou vzorků nás opravňuje hypotézu
společného původce vzorků vyloučit.
Analýza mtDNA, podobně jako analýza Y–STR, není schopna osobu
jednoznačně identifikovat. Své uplatnění v kriminalistické a
identifikační forenzní genetice však nachází tam, kde je analýza
jaderné DNA (autozomálních, X a Y–chromozomálních lokusů)
nemožná či velmi obtížná; těmito situacemi jsou zejména:
1. Analýza kosterního materiálu s vysoce degradovanou DNA
2. Analýza fragmentů vlasů a chlupů „bez kořínku“, tj. v katagenní
či telogenní fázi vývoje, či pouhých vlasových fragmentů.
Analýzou mtDNA opět nedosáhneme individuální identifikace
pachatele, ale v případě neshody HVR můžeme podezřelou osobu –
stejně jako u haplotypu Y–chromozómu – prakticky s jistotou
vyloučit. Analýza mtDNA je rovněž cenným nástrojem při
posuzování příbuzenských vztahů (viz dále).

117
Principy určení příbuznosti
Na základě analýzy jakých lokusů v DNA lze
posuzovat biologickou příbuznost osob?
Pro posouzení příbuznosti jsou stejně jako pro identifikace užívány
lokusy polymorfní až vysoce polymorfní. V části Forenzní
potenciál genetické analýzy jsme si již řekli, že k posouzení
příbuznosti využíváme v zásadě čtyř typů DNA lokusů, a to:
1. lokusů autozomálních, děděných z matky i z otce na všechny
děti; zde se zaměřujeme (stejně jako u identifikací) zejména na
STR lokusy,
2. lokusů X chromozomálních, děděných z matky na všechny
děti a z otce pouze na dcery; ve forenzní praxi jsou využívány
téměř výhradně X–STR lokusy (principiálně by bylo možno
použít i X–SNP lokusy, které se však užívají spíše v rámci
evolučně migračních studií),
3. lokusů Y chromozomálních, děděných výhradně z otce na
syny; ve forenzní praxi jsou využívány téměř výhradně Y–STR
lokusy (principiálně by bylo možno použít i Y–SNP lokusy,
které se však užívají spíše v rámci evolučně migračních studií),
4. lokusů mitochondriálních, děděných z matky na všechny děti;
stejně jako u identifikací se zde analýza soustředí na
hypervariabilní oblasti neboli HVR.
Jak lze posoudit příbuznost osob na základě
autozomálních STR lokusů?
Základní pravidla, kterými se primárně řídí dědičnost
autozomálních lokusů, jsme zde již zmínili, a lze je shrnout takto:
1. Rodič svému dítěti předá vždy jednu ze svých dvou alel.

118
2. O tom, která z obou rodičových alel bude předána, rozhoduje
náhoda.
3. S určitou velmi malou pravděpodobností může „v době
předávání“ (tj. při vzniku vajíčka či spermie, z něhož posléze
dítě vznikne) dojít k mutaci, která předávanou alelu pozmění
(např. z alely 11 se stane alela 12).
Dvě osoby (bez ohledu na to, zda jsou příbuzné či nikoliv) mohou
nést tutéž alelu určitého lokusu – např. jedna osoba má genotyp
11/12, druhá osoba je 12/14 – shodně mají tedy obě alelu 12. Této
alele pak říkáme alela shodná stavem.
Osoby biologicky příbuzné mají některé své alely shodné
původem; to znamená, že příslušná alela pochází od společného
předka obou těchto osob, nebo dokonce že jedna z těchto osob je
přímým předkem druhé osoby.
Alely shodné původem mohou, ale nemusejí být shodné stavem
(shodné nejsou, nastala–li mezigenerační mutace). Alely shodné
stavem mohou, ale nemusejí být shodné původem (lidé se
náhodně shodují, ačkoli nejsou příbuzní). Pro lepší pochopení
uveďme konkrétní případ:
Pavel a Marie jsou manželé a – pokud je jim známo – nejsou nijak
biologicky příbuzní. Pavel má v lokusu A genotyp 8/12, Marie genotyp
8/14. Alela 8 je u nich alelou shodnou stavem, nikoli však
původem.
Pavlovi a Marii se narodí syn David, který má v lokusu A genotyp
8/14. Je zřejmé, že David alelu 14 získal od matky Marie (je to tedy
jeho maternální alela) a můžeme konstatovat, že alela 14 je
u Davida a Marie shodná stavem i původem. Alelu 8 musel proto
David získat od otce Pavla (je to tedy jeho paternální alela) a můžeme
konstatovat, že alela 8 je u Davida a Pavla shodná stavem

119
i původem. Marie má také alelu 8, ovšem tu s jistotou Davidovi
nepředala (neboť mu předala alelu 14), a můžeme tedy konstatovat,
že alela 8 je u Davida a Marie shodná stavem, nikoli však
původem.
Po čase se Pavlovi a Marii narodí syn Lukáš, který má v lokusu A
genotyp 11/14. Pavel se s Lukášem tedy neshoduje ani v jedné z alel.
Analýza řady dalších lokusů prokáže, že příčinou není Mariina nevěra,
ale mezigenerační mutace Pavlovy alely 12 na Lukášovu alelu 11.
Můžeme tedy konstatovat, že Pavlova alela 12 a Lukášova alela 11
jsou shodné původem, nikoli však stavem.
Jak již jsme konstatovali v části věnované obecným informacím
o posouzení příbuznosti, každý typ biologického příbuzenského
vztahu je charakterizován určitým podílem alel shodných
původem. Při analýze ovšem tento podíl nedokážeme nikdy určit
přímo, ale pouze zprostředkovaně – pomocí vyhodnocení
přítomnosti alel shodných stavem.
Lze posoudit otcovství muže k dítěti i bez vzorku
matky dítěte?
Ano. Znalost DNA profilu matky umožňuje zpravidla identifikovat,
která alela dítěte je paternálního původu, což zlepšuje statistické
parametry zkoumání, není však v žádném případě nezbytnou
podmínkou posouzení otcovství. K tomu stačí znalost profilu dítěte
a znalost profilu potenciálního otce.
Lze posoudit otcovství muže k dítěti i bez jeho
vzorku?
Za určitých okolností ano. Není–li k dispozici přímo vzorek
nařčeného (potenciálního otce), je možné použít ke srovnání vzorky
jeho blízkých biologických příbuzných – rodičů (tj. potenciální

120
babičky a dědečka dítěte), dalších biologických dětí nařčeného (tj.
potenciálních sourozenců dítěte) apod. Vypovídací hodnota
zkoumání pak výrazně závisí na příbuznosti a počtu takto
dostupných „nepřímých“ vzorků. Existuje zde ovšem riziko, že
dostupné osoby nejsou ve skutečnosti v předpokládaném
biologickém vztahu k nařčenému – tj. například že jeho otec (tedy
potenciální dědeček dítěte) není ve skutečnosti jeho biologickým
otcem.
Jak lze posoudit příbuznost osob na základě X–
chromozomálních DNA lokusů?
Jak již bylo popsáno dříve, ženy mají ženy dva chromozómy X,
zatímco muži mají jeden chromozóm X a jeden Y.
U ženy se při tvorbě pohlavních buněk (meióze) dostává do vajíčka
vždy jeden z jejích chromozómů X. (Mezi jejími dvěma chromozómy
X může rovněž během meiózy dojít k vzájemné výměně částí DNA –
tzv. rekombinaci; do vajíčka se pak dostane jeden rekombinovaný
chromozóm X, který je vlastně kombinací obou původních ženiných
X chromozómů). U muže tento jev nenastává – chromozóm X a Y
spolu (vyjma pseudoautozomálních oblastí) nerekombinují; do
poloviny spermií se dostane chromozóm X v nezměněné podobě,
zatímco do druhé poloviny chromozóm Y, a to rovněž v nezměněné
podobě (samozřejmě za předpokladu, že nenastane mutace).
Výše uvedené má některé důležité důsledky pro posouzení
příbuznosti. Obecně užíváme analýzu X chromozomálních
lokusů spíše výjimečně, a to v případech, kdy analýza
autozomálních lokusů nemá sama o sobě dostatečnou
vypovídací hodnotu. Typickými situacemi jsou:
1. průkaz otcovství u páru otec–dcera, kdy není k dispozici vzorek
matky

121
2. průkaz otcovství dcery, kdy není k dispozici vzorek otce, ale
pouze jeho matky (tj. analýza páru babička z otcovy strany–
vnučka)
3. průkaz mateřství u páru matka–syn či matka–dcera, kdy není
k dispozici vzorek otce
4. průkaz polovičního sourozenectví sester, které mají společného
otce
5. některé incestní situace; analýza X–STR například může pomoci
rozhodnout, zda dítě zplodil otec nebo bratr jeho matky.
Jak lze posoudit příbuznost osob na základě Y–
chromozomálních DNA lokusů?
Posouzení příbuznosti osob na základě analýzy Y–chromozomálních
lokusů je v principu velmi jednoduché: Y chromozóm se dědí
v nezměněné podobě z otce na syny, tj. po meči. Jediným
mechanismem změny genotypu Y–chromozomálních lokusů jsou
mutace.
Při hodnocení možné příbuznosti dvou osob v paternální linii proto
porovnáváme zjištěné haplotypy Y–chromozómu – u osob takto
příbuzných by měly být identické, nanejvýše s nepatrným rozdílem
způsobeným mezigenerační mutací. V případě zjištění více
odchylek, které již nelze přičíst na vrub mutacím, příbuzenství
v paternální linii vyloučíme. Tohoto jevu je hojně využíváno
v geneticko – genealogickém výzkumu.
Jak lze posoudit příbuznost osob na základě lokusů
mitochondriální DNA?
Posuzování příbuznosti pomocí analýzy HVR mtDNA je obdobou
určení příbuznosti na základě analýzy Y–chromozomálních
markerů. MtDNA se dědí v nezměněné podobě z matky na všechny

122
její děti (dcery i syny), tj. po přeslici. Jediným mechanismem změny
mtDNA lokusů jsou mutace.
Při hodnocení možné příbuznosti dvou osob v maternální linii proto
porovnáváme zjištěné sekvence HVR mtDNA – u osob takto
příbuzných by měly být identické, nanejvýše s nepatrným rozdílem
způsobeným mezigenerační mutací. V případě zjištění více
odchylek, které již nelze přičíst na vrub mutacím, příbuzenství
v maternální linii vyloučíme.
Principy predikce biogeografického původu a
fenotypových znaků osoby
Jak se pomocí analýzy DNA určuje pohlaví osoby?
Určení pohlaví je založeno na skutečnosti, že ženské tělní buňky
obsahují dva chromozómy X, zatímco mužské obsahují jeden
chromozóm X a jeden Y.
Historicky je pro DNA analýzu pohlaví nejdéle a nejčastěji
využíván gen pro amelogenin, který se vyskytuje jak na
chromozómu X, kde je plně funkční (dává vzniknout bílkovině zubní
skloviny), tak na chromozómu Y, kde se nachází jeho nefunkční
verze – tzv. pseudogen. Mezi genem na X a pseudogenem na Y
existuje několik drobných konstantních rozdílů; například určitý
úsek tohoto genu je na X chromozómu o 6 nukleotidů kratší než
tentýž úsek na Y chromozómu. Při vlastní analýze tedy tento úsek
„změříme“ (pochopitelně molekulárně genetickými metodami,
nikoliv pravítkem): u žen zjistíme jedinou délku (tzn. jedinou alelu X
o délce např. 104 bp), u mužů délky dvě (tzn. jednu alelu X o délce
např. 104 bp a jednu alelu Y o délce 110 bp).

123
Obrázek 34 – Určení pohlaví pomocí analýzy krátkého úseku DNA v genu pro amelogenin, který se nachází na X i na Y chromozómu. U muže detekujeme dva rozdílně dlouhé fragmenty, u ženy fragment jediný.
Jinou možností určení pohlaví je analýza specificky mužských
lokusů – tedy těch, které leží na Y chromozómu (např. SRY).
Pozitivní výsledek analýzy je potvrzením, že materiál obsahoval
buňky mužského původu, u srovnávacího vzorku tedy, že patří
muži. U ženy dostaneme negativní výsledek analýzy. V tom ovšem
tkví i úskalí tohoto způsobu určení pohlaví – při nesprávném
provedení analýzy či při analýze degradovaného a/nebo
inhibovaného materiálu můžeme dostat falešně negativní výsledek
(tj. vzorek je mužský, ale analýza nedala pozitivní výsledek). Bránit
tomu lze současnou analýzou nějakého kontrolního autozomálního
lokusu, který by pozitivní výsledek měl poskytnout jak u mužského,
tak u ženského vzorku.

124
Jak se pomocí analýzy DNA určuje biogeografický
původ osoby?
Jak bylo deklarováno již výše, je určování biogeografického původu
osoby analýzou DNA zatím ve svých počátcích a není možné při
těchto predikcích dosáhnout vysoké míry spolehlivosti (například
srovnatelné se spolehlivostí identifikace pomocí STR). Principiálně
při hledání biogeografického původu osoby můžeme užít analýzu
mtDNA, analýzu Y–chromozómu a analýzu některých
autozomálních lokusů, které mohou informovat o původu osoby.
Mitochondriální DNA je, jak již zde bylo vícekrát konstatováno –
děděna po přeslici neboli maternálně. Nové linie mtDNA vznikají
s velmi malou frekvencí a náhodně – mutací. Tedy – matka předá
svým dětem svou mtDNA v nezměněné podobě, při vzniku jednoho
jejího vajíčka však dojde k mutaci a jedno z jejích dětí obdrží
mutovanou verzi mtDNA; nová linie je založena. K další mutaci
dojde v průměru za několik desítek generací (současné odhady
rychlosti těchto mutací se však výrazně liší, proto neuvádíme žádné
konkrétní číslo).
Popsaný mechanismus spolu se způsobem, jakým moderní člověk
postupně osidloval jednotlivé kontinenty, má velmi zajímavý
důsledek: různé linie mtDNA se vyskytují na různých
kontinentech, nebo mají alespoň rozdílné frekvence výskytu.
Kupříkladu některé mtDNA linie nenajdeme jinde než v Asii, jiné
zase téměř výhradně v Evropě a na Blízkém východě.
Stanovením sekvence HVR mtDNA linie tak můžeme odhadovat,
odkud daná osoba pochází. Tento na první pohled půvabný princip
je však problematizován dalšími faktory. Uveďme alespoň dva:
Za prvé – analýza umožňuje odhad jen v hrubším měřítku; dle
mitotypu můžeme např. relativně spolehlivě určit, že osoba pochází

125
s velkou pravděpodobností z Evropy, avšak už nedokážeme určit, ze
kterého státu.
Za druhé – informace z mtDNA se týká výhradně již zmíněné linie
„po přeslici“, ostatní předkové nejsou při této analýze zohledněni.
Představme si například fiktivní čínskou dívku Fu Li, která se v době
Marca Pola zamiluje do některého z evropských obchodníků,
cestujících pro zboží do Asie, a odejde s ním do jeho slunné Itálie.
Jejich společná dcera, která se po čase narodí, nese plně asijský
mitotyp, který se bude dále předávat přes dcery do dalších generací.
Jednoho dne roku 2011 si na univerzitě v Bologni studentka Fulvia
udělá v rámci praktik forenzní genetiky sekvenaci HVR mtDNA.
S překvapením zjistí, že přísluší k tzv. haploskupině G, typické pro
oblast severovýchodní a centrální Asie. Je to pro ni veliké překvapení –
ona je přece každým coulem Italka, nic asijského na sobě nikdy
nepozorovala. Inu, není divu – mezi ní a její maternální prapředkyní
Fu Li, která pro lásku neváhala opustit navždy domov, leží přibližně
30 generací.
Určení biogeografického původu osoby prostřednictvím
analýzy haplotypu Y–chromozómu je založeno na stejném
principu jako prostřednictvím analýzy mtDNA. Také různé Y–
chromozomální linie jsou geograficky více či méně specifické. Stejně
jako u mtDNA, i zde je metoda limitována malou rozlišovací
schopností a tím, že nezohledňuje jinou než čistě paternální část
dědičné informace.
Velmi nadějnou metodou určení biogeografického původu je
užití autozomálních SNP lokusů, označovaných jako AIMs (angl.
Ancestry Informative Markers) – tedy markery informující o
původu. Princip této analýzy je opět jednoduchý: již jsme si uvedli,
že u typického SNP existují zpravidla dvě alely, které mají v populaci
určitou frekvenci. U AIMs je tato frekvence v různých populacích

126
různá. Kupříkladu: SNP rs729172 má alely A a C; v rámci USA se
alela A vyskytuje u Afroameričanů s frekvencí 23 %, u Asiatů
s frekvencí 6 % a u europoidního typu s frekvencí 38 %. Jinak
řečeno, alelu A najdeme čtyřikrát častěji u Afroameričanů než
u Asiatů a více než šestkrát častěji u europoidního typu než u Asiatů.
Analýza jednotlivého AIM není sama o sobě dostatečně
informativní; pokud ale analyzujeme celý set takovýchto lokusů
(zhruba kolem padesáti) a výsledky statisticky vyhodnotíme, již
můžeme dostat informaci s relativně solidní vypovídací hodnotou.
Navíc tato informace – na rozdíl od mtDNA a Y–chromozomálního
haplotypu – zohledňuje genetický příspěvek všech předků testované
osoby.
Jak se pomocí analýzy DNA určují fenotypové znaky
osoby?
Opět je třeba zdůraznit a připomenout, co již bylo řečeno výše:
predikce fenotypových znaků je v tuto chvíli ve svých počátcích a
není zatím dostatečně spolehlivá. Dílčí úspěch však již přináší, a
proto se o ní zde rovněž zmiňujeme.
Analýza fenotypových znaků je založena na stanovení genotypu
SNP, které se nacházejí v těsné blízkosti genů podmiňujících
příslušný znak, případně přímo v těchto genech. V roce 2010 byl
kupříkladu validován IrisPlex, test šesti SNP vázaných s geny
ovlivňujícími pigmentaci oční duhovky. Tento test umí s relativně
velkou spolehlivostí odlišit světlé (modré) oči od tmavých
(hnědých). V roce 2011 pak byl validován další systém – HIrisPlex,
umožňující současnou predikci barvy vlasů i oční duhovky.

127
Technologie forenzně genetické analýzy
Základní kroky forenzně genetické analýzy
Jaké technologické postupy jsou využívány při
forenzní analýze DNA?
Spektrum užívaných metod je velmi široké a nelze ho nijak striktně
omezit. Vzhledem k šíři záběru forenzní genetiky se zde v různých
situacích uplatňuje celá řada specifických postupů, a jak se obor
rozvíjí, stále další přibývají (a zastaralé jsou opouštěny). Žádná
z těchto metod není uplatňována jen a pouze ve forenzní genetice,
ale jsou užívány i jinými obory molekulární genetiky a DNA
diagnostiky – setkáme se s nimi v lékařské diagnostice,
šlechtitelství, veterinární genetice a podobně.
V následujících odstavcích budou popsány základní kroky a
postupy, běžně uplatňované při forenzně genetickém
zkoumání. Nejedná se o plný výčet, ten by ostatně ani nebyl
vzhledem k výše uvedenému možný, ale o stručný přehled, který by
měl čtenáři umožnit zorientovat se v základních otázkách technické
stránky forenzní genetiky. S ohledem na zaměření této publikace
jsou jednotlivé metody popisovány co nejjednodušeji, bez příliš
technicistních detailů – důraz je kladen na princip daného postupu.
Jaké kroky zahrnuje forenzně genetická analýza?
Po zajištění stopy a jejím uchování až do doby analýzy, kteréžto
kroky byly popsány v první části, pojednávající o stopách, následuje
– již v laboratoři – izolace neboli extrakce DNA (tyto pojmy jsou
běžně používány jako synonyma – „izolace“ znamená oddělení,
rozuměj DNA od ostatních látek, „extrakce“ znamená vytažení,
rozuměj DNA ze směsi látek).
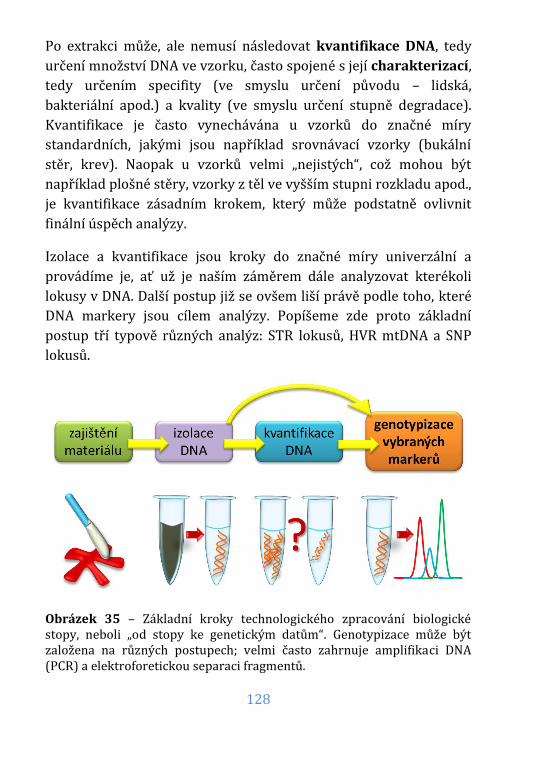
128
Po extrakci může, ale nemusí následovat kvantifikace DNA, tedy
určení množství DNA ve vzorku, často spojené s její charakterizací,
tedy určením specifity (ve smyslu určení původu – lidská,
bakteriální apod.) a kvality (ve smyslu určení stupně degradace).
Kvantifikace je často vynechávána u vzorků do značné míry
standardních, jakými jsou například srovnávací vzorky (bukální
stěr, krev). Naopak u vzorků velmi „nejistých“, což mohou být
například plošné stěry, vzorky z těl ve vyšším stupni rozkladu apod.,
je kvantifikace zásadním krokem, který může podstatně ovlivnit
finální úspěch analýzy.
Izolace a kvantifikace jsou kroky do značné míry univerzální a
provádíme je, ať už je naším záměrem dále analyzovat kterékoli
lokusy v DNA. Další postup již se ovšem liší právě podle toho, které
DNA markery jsou cílem analýzy. Popíšeme zde proto základní
postup tří typově různých analýz: STR lokusů, HVR mtDNA a SNP
lokusů.
Obrázek 35 – Základní kroky technologického zpracování biologické stopy, neboli „od stopy ke genetickým datům“. Genotypizace může být založena na různých postupech; velmi často zahrnuje amplifikaci DNA (PCR) a elektroforetickou separaci fragmentů.

129
Jak dlouho trvá provedení forenzně genetické analýzy?
Délka celé analýzy závisí na tom, které kroky jsou do ní zařazeny a
jaká je jejich časová náročnost. Hlavním faktorem je bezesporu
délka izolace DNA, která může být u různých typů izolovaného
materiálu i řádově rozdílná – kupříkladu izolace z kostí může být
dle zvoleného postupu otázkou až dnů, naproti tomu izolace
z bukálních stěrů může být redukována na pouhé minuty. Druhým
rozhodujícím faktorem je doba potřebná k amplifikaci
zájmových lokusů, která se může dle zvoleného postupu lišit
o desítky minut. Zkrácení izolace i amplifikace je prioritou výzkumu
a na trhu již se objevují komerční kity se zkrácenou délkou těchto
kroků – u amplifikace je to např. Identifiler® Plus Kit, či PowerPlex
18D Kit.
Izolace (extrakce) DNA
Co je smyslem izolace DNA?
V biologickém materiálu (stop i srovnávacích vzorků) je kromě
DNA přítomno velké množství nejrůznějších dalších látek, např.
bílkoviny, lipidy, polysacharidy atd. Řada z těchto látek v principu
znemožňuje některé další kroky analýzy (patří mezi ně např.
takzvané inhibitory, o nichž je pojednáno dále), a proto je třeba je ze
vzorku odstranit. Navíc – DNA v biologickém materiálu je často
nestabilní a může být snadno poškozena ve stopě přítomnými
látkami (např. enzymy původem z buněk vzorku či enzymy
produkovanými mikroorganismy), je třeba ji tedy z tohoto
„ohrožení“ dostat. Ideální izolační postup je ten, který:
má co nejvyšší separační účinnost, tj. v maximální možné
míře odstraní ze vzorku všechny látky vyjma nukleových
kyselin (tj. DNA a RNA)

130
má co nejvyšší výtěžnost, tj. získá z materiálu pokud možno
veškerou přítomnou DNA; tato podmínka je důležitá především
u vzorků s malým množstvím využitelného materiálu. Naopak,
u vzorků bohatých na DNA často dáváme přednost takovému
izolačnímu postupu, který nám izoluje DNA v určitém
standardním množství (tj. víme, že na konci izolace budeme mít
k dispozici roztok DNA o určité standardní koncentraci).
převede DNA do stabilního stavu – do roztoku či na pevnou
fázi (nosič), které jsou bez problémů použitelné pro následující
analýzy.
Musí být izolace provedena vždy?
Existují některé speciální postupy či přípravky, které u určitých
typů vzorků umožňují celý izolační proces redukovat na minimum –
řadí se sem např. Prep–n–GoTM Buffer firmy Life Technologies či
SwabSolution Kit firmy Promega. Jejich užití v rámci forenzní
analýzy je možné, ale spíše jen při zpracování srovnávacích vzorků.
Co je izolát?
Izolátem DNA rozumíme finální produkt izolace DNA; obvykle je
to vodný roztok DNA (tj. ředidlem je voda, v izolátu mohou být
přítomny i další látky, např. soli).
Pozor! Zatímco pojmy „analýza DNA“ a „genetická analýza“ jsou
prakticky synonyma, pojem „izolát DNA“ a „genetický/genový izolát“
rozhodně ne: jako genetické/genové izoláty jsou označována
teritoria, kde určitá subpopulace dlouhodobě žije do značné míry
izolovaně – např. některé odlehlé horské osady.
Co jsou inhibitory?
Jako inhibitory označujeme látky nejrůznější povahy, které mají
negativní účinek na průběh jednoho ze zásadních kroků analýzy,

131
tzv. polymerázové řetězové reakce (PCR) – viz dále. Jejich
přítomnost ve vzorku může vést ke zhoršené kvalitě výsledků
analýzy či k úplnému selhání analýzy.
Inhibitory mohou mít svůj původ jednak přímo ve vzorku (tj.
jsou přítomny už v původním biologickém materiálu, který je dodán
do laboratoře ke zkoumání), nebo se mohou do vzorku dostat až
v průběhu izolace z reagencií pro izolaci užívaných.
K hlavním inhibitorům původem ze vzorku řadíme krevní barvivo
hemoglobin, jeho rezidua heminy, indigový pigment z jeansoviny,
huminové kyseliny z půdy, vlasový a kožní pigment melanin a
mnoho jiných.
Zanesení inhibitorů z reagencií je vždy důsledkem nesprávně
zvoleného nebo nesprávně provedeného izolačního postupu.
Typickými inhibitory tohoto původu jsou alkoholy, fenol či některé
typy detergentů.
Jak lze inhibitory odstranit?
Některé inhibitory lze odstranit už během izolace, některé jsou
odstranitelné specifickou purifikací izolátu (viz dále) a některé jsou
odstranitelné jen velmi obtížně. U inhibitorů původem z krevních
vzorků lze do jisté míry potlačit jejich negativní účinek až v průběhu
samotné PCR, a to přidáním tzv. bovinního sérového albuminu.
Co je purifikace DNA?
Termínem purifikace DNA označujeme přečištění izolátu DNA, a to
právě kvůli účinnému odstranění inhibitorů, případně dalších
nežádoucích látek, které zůstaly ve vzorku přítomny i po izolaci.

132
Co je zkoncentrování izolátu DNA?
Zkoncentrováním izolátu rozumíme snížení objemu izolátu při
zachování veškeré DNA, čímž se roztok „zahustí“; tento postup
používáme zejména u vzorků s malým obsahem DNA.
Zkoncentrování lze provést buď vyvázáním DNA z roztoku na
pevnou fázi (např. oxid křemičitý) a její následné uvolnění do
roztoku o menším objemu, nebo centrifugací pomocí tzv.
mikrokoncentrátorů: roztok DNA je v mikrokoncentrátoru při
centrifugaci „hnán“ přes velmi husté sítko, které propustí pouze
rozpouštědlo a drobné molekuly, makromolekuly DNA však ne – ty
v něm zůstanou zachyceny a opět mohou být uvolněny do roztoku
o menším objemu, než měl původní izolát.
Jaké jsou základní izolační mechanismy?
Prvním krokem izolace je prakticky vždy lýza buněk, tedy jejich
rozrušení a uvolnění buněčného obsahu do roztoku. Lýza je
nejčastěji založena na osmóze: buňky v destilované vodě (případně
lyzačním pufru) zvyšují svůj objem (molekuly vody pronikají přes
buněčnou membránu dovnitř), až popraskají. Často je tento
jednoduchý mechanismus podpořen zvýšenou teplotou,
protřepáváním vzorku a přidáním některých enzymů, např.
proteinázy, štěpící bílkoviny. Výsledkem je tzv. buněčný lyzát.
Po lýze buněk může následovat několik základních typů izolačních
postupů:
Postupy založené na oddělení polárních a nepolárních látek
Postupy založené na inaktivaci enzymatických kofaktorů
Postupy založené na vazbě DNA na papírový (tj. celulózový)
absorbent
Postupy založené na vazbě DNA na pevnou fázi
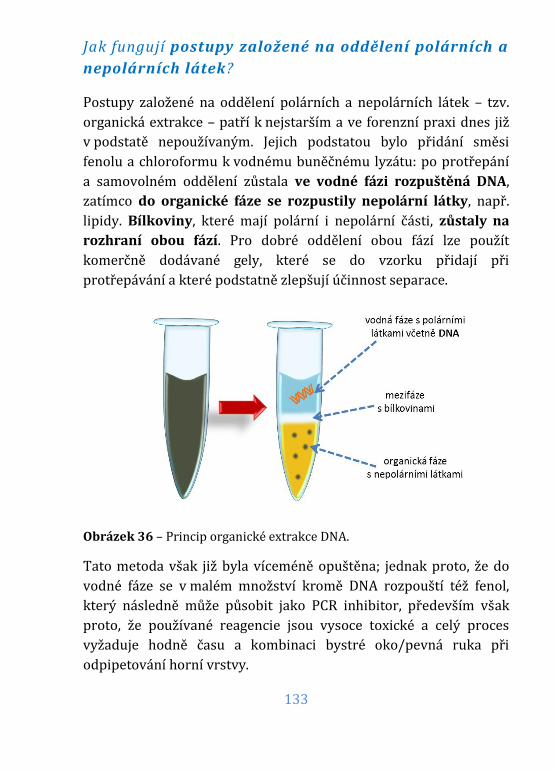
133
Jak fungují postupy založené na oddělení polárních a
nepolárních látek?
Postupy založené na oddělení polárních a nepolárních látek – tzv.
organická extrakce – patří k nejstarším a ve forenzní praxi dnes již
v podstatě nepoužívaným. Jejich podstatou bylo přidání směsi
fenolu a chloroformu k vodnému buněčnému lyzátu: po protřepání
a samovolném oddělení zůstala ve vodné fázi rozpuštěná DNA,
zatímco do organické fáze se rozpustily nepolární látky, např.
lipidy. Bílkoviny, které mají polární i nepolární části, zůstaly na
rozhraní obou fází. Pro dobré oddělení obou fází lze použít
komerčně dodávané gely, které se do vzorku přidají při
protřepávání a které podstatně zlepšují účinnost separace.
Obrázek 36 – Princip organické extrakce DNA.
Tato metoda však již byla víceméně opuštěna; jednak proto, že do
vodné fáze se v malém množství kromě DNA rozpouští též fenol,
který následně může působit jako PCR inhibitor, především však
proto, že používané reagencie jsou vysoce toxické a celý proces
vyžaduje hodně času a kombinaci bystré oko/pevná ruka při
odpipetování horní vrstvy.

134
Jak fungují postupy založené na inaktivaci
enzymatických kofaktorů?
Postupy založené na inaktivaci enzymatických kofaktorů jsou
založeny na jednoduché lýze a následné separaci lyzovaných
buněk pomocí centrifugace (případně s přidáním proteináz či
proteáz k rozrušení bílkovin), po nichž následuje přidání tzv.
chelatačních činidel – látek, které jsou schopny vyvazovat na sebe
některé typy iontů; v praxi je nejčastěji používán Chelex–100,
iontoměničová pryskyřice, která vyvazuje dvojmocné ionty kovů.
Při lýze buněk se do roztoku mimo jiné uvolní endonukleázy –
enzymy, které mohou napadat a rozrušovat molekuly DNA.
Endonukleázy potřebují ke svému fungování kofaktory, kterými
jsou hořečnaté ionty. Přidáním Chelexu–100 k lyzátu však dojde
k vyvázání hořečnatých iontů z roztoku, takže tyto nemohou plnit
funkci kofaktorů, a tudíž endonukleázy nemohou napadat DNA; tím
je izolát stabilizován.
Obrázek 37 – Princip vyvázání enzymatických kofaktorů při extrakci DNA.
Tento izolační postup je velmi jednoduchý, rychlý a velmi levný,
nejsou však při něm z roztoku účinně odstraňovány inhibitory, a své
uplatnění tak nachází především u vzorků s malým množstvím

135
inhibitorů – plošné stěry, vzorky slin (např. cigaretové nedopalky),
malé krevní vzorky apod. Je též velmi často používán pro izolace ze
srovnávacích vzorků bukálních stěrů.
Jak fungují postupy založené na vazbě DNA na
papírový absorbent?
Postupy založené na vazbě DNA na celulózový absorbent jsou
reprezentovány především tzv. FTA papírem (FTA kartami).
Podstatou tohoto absorbentu je papír, obsahující látky
inaktivující nukleázy a látky bránící růstu bakterií. Na FTA
papír může být nakápnut či natřen tekutý nebo rozpuštěný
biologický materiál (krev, sliny apod.). U modernějších typů FTA
karet je nanesení biologického materiálu okamžitě indikováno
změnou barvy papírku v daném místě. Materiál nasákne do papíru
a vyschne, čímž je DNA imobilizována a pevně v papíru
ukotvena. V takovém stavu může být i neizolovaný vzorek při
pokojové teplotě uchováván řadu měsíců.
Obrázek 38 – FTA karty slouží k imobilizaci a trvalému uchování DNA ve stabilním stavu.

136
Následná izolace se provádí tak, že z karty je mechanicky vyražen
malý terčík, ten se vloží do zkumavky a jsou k němu přidány izolační
reagencie; dojde k odstranění nežádoucích látek (např. vymytí
inhibitorů), přičemž DNA stále zůstává fixována na papírovém
terčíku.
Jak fungují postupy založené na vazbě DNA na
pevnou fázi?
Postupy založené na vazbě DNA na pevnou fázi využívají toho, že
DNA je molekula s parciálním (tj. částečným) záporným
nábojem. Jako zmíněná pevná fáze je zpravidla užívána silika
(polymerní oxid křemičitý), která je na povrchu rovněž záporně
nabita. V čisté vodě se tak DNA a silika navzájem odpuzují. Pokud
však do roztoku přidáme tzv. chaotropní soli, zafungují jako
zprostředkovatel vazby mezi DNA a silikou. Touto vazbou je pak
DNA pevně vázána k silice a může být spolu s ní
transportována. Po změně složení roztoku (snížení koncentrace
solí, změně pH z kyselého na zásadité) dojde ke zrušení vazby DNA
na silikát a jejímu uvolnění do roztoku.
V praxi jsou užívána dvě základní uspořádání tohoto izolačního
postupu: za prvé tzv. silikátové kolonky, kde je DNA zachycena na
kolonce během centrifugace (případně vakuovým odsátím) lyzátu,
pak přečištěna a následně uvolněna do čistého elučního roztoku; za
druhé tzv. magnetosilikové partikule, kde se DNA naváže na tyto
paramagnetické kuličky, které mohou být manipulovány pomocí
magnetu – např. vyjmuty z roztoku, přeneseny do přečišťovacích
roztoků a z nich nakonec do elučního roztoku, kde je DNA uvolněna.

137
Obrázek 39 – Vazba DNA na siliku v magnetosilikových partikulích umožňuje manipulovat DNA v roztoku pomocí magnetického pole.
Tyto postupy jsou vysoce efektivní co do odstranění nežádoucích
látek z roztoku, mají velkou výtěžnost a mohou být automatizovány
(izolace je pak prováděna roboticky), což z nich činí ideální izolační
metody pro forenzní analýzu; nevýhodou je jejich relativně vysoká
cena.
Lze nějakým mechanismem oddělit biologický
materiál jednotlivých přispěvatelů vzorku?
Za určitých okolností ano. V této souvislosti je třeba zmínit dva
postupy, a to diferenciální lýzu a laserovou mikrodisekci.
Diferenciální buněčná lýza je velmi specifickým postupem
užívaným u vzorků tvořených směsí ženských epiteliálních buněk a
spermií – typicky u vzorků poševních stěrů po znásilnění (název je
zavedeným, leč poněkud otrockým překladem původního
anglického termínu differential cell lysis – slovo „differential“ zde
ovšem funguje ve významu „odstupňovaný“, „založený na rozdílu“, a
asi vhodnější by tedy bylo užití termínu odstupňovaná buněčná
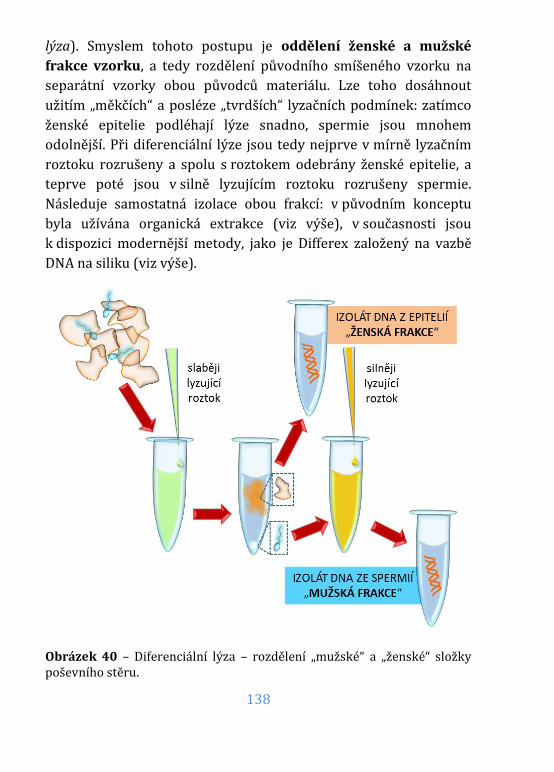
138
lýza). Smyslem tohoto postupu je oddělení ženské a mužské
frakce vzorku, a tedy rozdělení původního smíšeného vzorku na
separátní vzorky obou původců materiálu. Lze toho dosáhnout
užitím „měkčích“ a posléze „tvrdších“ lyzačních podmínek: zatímco
ženské epitelie podléhají lýze snadno, spermie jsou mnohem
odolnější. Při diferenciální lýze jsou tedy nejprve v mírně lyzačním
roztoku rozrušeny a spolu s roztokem odebrány ženské epitelie, a
teprve poté jsou v silně lyzujícím roztoku rozrušeny spermie.
Následuje samostatná izolace obou frakcí: v původním konceptu
byla užívána organická extrakce (viz výše), v současnosti jsou
k dispozici modernější metody, jako je Differex založený na vazbě
DNA na siliku (viz výše).
Obrázek 40 – Diferenciální lýza – rozdělení „mužské“ a „ženské“ složky poševního stěru.
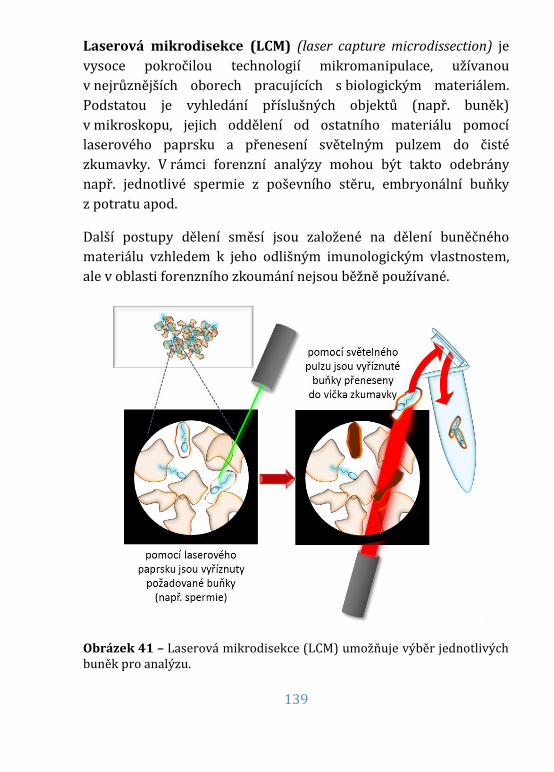
139
Laserová mikrodisekce (LCM) (laser capture microdissection) je
vysoce pokročilou technologií mikromanipulace, užívanou
v nejrůznějších oborech pracujících s biologickým materiálem.
Podstatou je vyhledání příslušných objektů (např. buněk)
v mikroskopu, jejich oddělení od ostatního materiálu pomocí
laserového paprsku a přenesení světelným pulzem do čisté
zkumavky. V rámci forenzní analýzy mohou být takto odebrány
např. jednotlivé spermie z poševního stěru, embryonální buňky
z potratu apod.
Další postupy dělení směsí jsou založené na dělení buněčného
materiálu vzhledem k jeho odlišným imunologickým vlastnostem,
ale v oblasti forenzního zkoumání nejsou běžně používané.
Obrázek 41 – Laserová mikrodisekce (LCM) umožňuje výběr jednotlivých buněk pro analýzu.

140
Kvantifikace (a případná charakterizace) DNA
Co je to kvantifikace DNA a jak vyjadřujeme
množství DNA?
Kvantifikací rozumíme stanovení množství DNA ve vzorku. Toto
množství může být vyjádřeno několika způsoby; nejčastěji je to tzv.
hmotnostní koncentrace DNA ve vzorku, jejíž základní jednotka je
kg/m3 („kilogram na metr krychlový“). Ve forenzní analýze, která
pracuje s velmi malými objemy vzorků, však používáme odvozené
jednotky ng/μl („nanogram na mikrolitr“) a pg/μl („pikogram na
mikrolitr“). Platí, že 1 kg/m3 = 1 000 ng/μl = 1 000 000 pg/μl.
Jedna lidská tělní buňka obsahuje zhruba 6,6 pg DNA. Pokud
bychom tedy měli čistý izolát ze 100 lidských buněk o objemu 1 μl,
bude mít tento izolát hmotnostní koncentraci DNA 660 pg/μl neboli
0,66 ng/μl. A obráceně: mikrolitrový izolát o koncentraci 1,00 ng/μl
obsahuje DNA ze zhruba 152 buněk. Množství DNA ve spermiích je
poloviční – 3,3 pg. Mikrolitrový izolát o koncentraci 1,00 ng/μl tedy
obsahuje DNA ze zhruba 304 spermií.
Jaký smysl má kvantifikace DNA?
Stanovení množství DNA ve vzorku je důležité ze dvou důvodů: za
prvé, na základě výsledků můžeme vzájemně porovnat účinnost
několika izolačních postupů. Pokud například odebereme vždy ze
stopy dva stejné vzorky, tyto izolujeme dvěma různými postupy A a
B a izolát kvantifikujeme, můžeme zjistit, že izolační postup A nám
systematicky poskytuje vyšší koncentrace DNA v izolátu než postup
B, a je tedy zjevně účinnější izolační metodou.
Za druhé, stanovení koncentrace DNA ve vzorku je důležité pro
vyladění následně prováděné polymerázové řetězové reakce
(PCR – viz dále). Tato metoda je citlivá na správná vstupní množství

141
DNA – příliš malá, ale především příliš velká vstupní množství DNA
mohou vést ke zhoršenému průběhu reakce nebo k její úplné
inhibici. Kupříkladu je–li optimální vstupní množství DNA do určité
PCR reakce 1 ng, pak vstupní množství 10 ng DNA způsobí
částečnou inhibici reakce a PCR proběhne velmi problematicky.
Kvantifikace je tak nesmírně důležitá zejména u vzorků s těžko
odhadnutelným obsahem DNA (plošné stěry, tkáně v rozkladu
apod.)
Co je to nespecifická a specifická kvantifikace DNA?
Izolační postup zpravidla do výsledného izolátu převede veškerou
DNA, přítomnou původně ve vzorku. U biologických stop tak může
výsledný izolát obsahovat kromě lidské DNA též DNA zvířecí,
rostlinnou, bakteriální apod. Mimochodem – u srovnávacího vzorku
to může platit též – např. u vzorků bukálního stěru izolujeme kromě
vlastní DNA buněk bukální sliznice též DNA bakterií přítomných
v ústech, tvořících zubní plak.
Kvantifikační postupy, které určí množství veškeré DNA ve vzorku
bez ohledu na to, z jakého zdroje tato DNA pochází, označujeme jako
nespecifické. Naproti tomu postupy, zaměřené na stanovení
množství pouze určitého specifického druhu DNA (nejčastěji DNA
lidské), označujeme jako specifické.
Jaké jsou základní principy nespecifické kvantifikace
DNA?
Nejjednodušším a pro forenzní praxi nejméně vhodným postupem
je spektrofotometrická kvantifikace DNA. Je založena na tom, že
molekuly DNA silně absorbují elektromagnetické záření vlnové
délky 260 nm (tj. ultrafialové záření); paprsek této vlnové délky se
nechá procházet roztokem DNA (např. izolátem) a změří se pokles
intenzity záření – ten je tím větší, čím je roztok DNA

142
koncentrovanější. Metoda není vhodná pro forenzní účely
(užívala se pouze v naprostých počátcích), neboť není příliš citlivá,
tj. nezachytí malá množství DNA, spotřebovává se při ní velké
množství vzorku a výsledek měření může být velmi silně ovlivněn
přítomností některých dalších chemických látek ve vzorku, jako je
fenol, bílkoviny apod. Výjimku tvoří mikrospektrofotometrické
metody použitelné ve forenzní genetice především pro získání
rychlé orientační informace o množství DNA ve vzorku.
Obrázek 42 – Spektrofotometrická kvantifikace je založena na měření poklesu intenzity světla procházejícího roztokem DNA.
Pro forenzní účely vhodnější metodou je detekce nespecificky
inkorporovaného fluorescenčního barviva. Pro tyto účely se
užívají chemické sloučeniny – barviva (např. ethidiumbromid,
PicoGreen, OligoGreen), která se navážou na/do DNA, tím se změní
jejich prostorové uspořádání a začnou být při dopadu světla určité
vlnové délky silně fluorescenčně aktivní, tj. vyzařují světlo, jehož
množství měříme fluorimetrem. Množství vyzářeného světla je
tedy přímo úměrné množství navázaného barviva, a to je zase přímo
úměrné množství DNA přítomné ve vzorku. Výhodou metody je

143
relativně vysoká citlivost a přesnost měření, malé nároky na
spotřebu vzorku, možnost měření automatizovat a malá cena
analýzy; nevýhodou zůstává nespecifita měření.
Obrázek 43 – Nespecifická kvantifikace založená na navázání fluorescenčního barviva do DNA. Měření se provádí pomocí fluorimetru.
Jaké jsou základní principy specifické kvantifikace
DNA?
Při specifické kvantifikaci musí být dosaženo toho, že metoda
zobrazí výlučně příslušnou specifickou DNA – nejčastěji DNA
lidskou. Toho dosahujeme použitím tzv. druhově specifických sond
či druhově specifických primerů, což obojí jsou krátké
jednořetězcové fragmenty DNA, které jsou komplementární (laicky
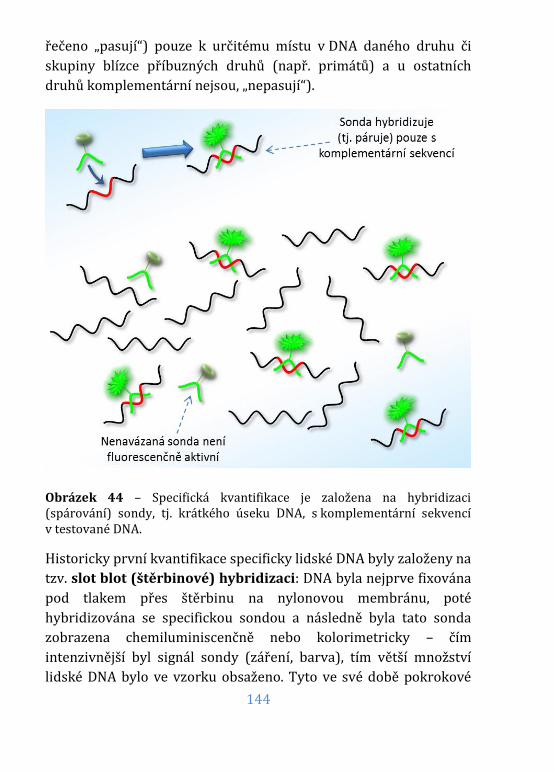
144
řečeno „pasují“) pouze k určitému místu v DNA daného druhu či
skupiny blízce příbuzných druhů (např. primátů) a u ostatních
druhů komplementární nejsou, „nepasují“).
Obrázek 44 – Specifická kvantifikace je založena na hybridizaci (spárování) sondy, tj. krátkého úseku DNA, s komplementární sekvencí v testované DNA.
Historicky první kvantifikace specificky lidské DNA byly založeny na
tzv. slot blot (štěrbinové) hybridizaci: DNA byla nejprve fixována
pod tlakem přes štěrbinu na nylonovou membránu, poté
hybridizována se specifickou sondou a následně byla tato sonda
zobrazena chemiluminiscenčně nebo kolorimetricky – čím
intenzivnější byl signál sondy (záření, barva), tím větší množství
lidské DNA bylo ve vzorku obsaženo. Tyto ve své době pokrokové

145
metody (nejznámější z nich byl QuantiBlot Human DNA
Quantitation Kit) se dnes již prakticky neužívají.
Obrázek 45 – Takto vypadal výsledek kvantifikace specificky lidské DNA pomocí slot blot hybridizace: kvantita DNA v testovaných vzorcích se odhadovala porovnáním intenzity vybarvení s kalibrační řadou (tj. se vzorky známé koncentrace – v obr. uvedena v ng).
Dalším principem kvantifikace specificky lidské DNA byla
luminometrická kvantifikace AluQuant Human DNA
Quantitation System. Při ní byla DNA hybridizována se specifickou
sondou, což v roztoku vedlo k sérii enzymatických reakcí až po
finální oxidaci luciferinu enzymem luciferázou za vzniku viditelného
světla, jehož množství bylo změřeno luminometrem. (Mimochodem,
je to identická enzymatická reakce, kterou používají samečci
světlušek). Čím více lidské DNA bylo ve vzorku přítomno, tím více
sondy hybridizovalo a tím intenzivnější světlo v konečné fázi
z roztoku zazářilo. Přestože tato metoda byla po mnoha stránkách
pro forenzní účely vynikající, dnes už se prakticky neužívá, neboť
byla vytlačena moderní metodou qPCR.
Nejmodernějšími a v současnosti zdaleka nejužívanějšími metodami
specifické kvantifikace jsou metody založené na polymerázové

146
řetězové reakci (viz níže) – tzv. kvantitativní PCR (qPCR).
Technicky může mít qPCR mnoho konceptů, jejichž popis a
vysvětlení by šly nad rámec této knihy. Koncepty užívané ve
forenzní analýze jsou nejčastěji založeny na v zásadě jednoduchém
principu: je prováděna PCR, tj. cyklické kopírování vybraného
malého úseku DNA, a současně je průběžně měřeno množství těchto
nakopírovaných DNA fragmentů ve vzorku. Z toho, jakým způsobem
během PCR toto množství rostlo, lze zpětně dovodit původní
vstupní množství DNA ve vzorku (opět se kalibruje pomocí vzorků
známé koncentrace). Metoda kvantitativní PCR nám ze všech
kvantifikačních metod přináší nejvíce informací: je vysoce přesná,
vysoce citlivá (zobrazí i velmi malá množství DNA), přísně
specifická (poskytuje pozitivní výsledek prakticky pouze u daného
druhu respektive velmi blízce příbuzných druhů) a detekuje i
případnou přítomnost PCR inhibitorů a rozsah DNA
fragmentace.
Amplifikace DNA
Co je to amplifikace DNA?
Termín amplifikace označuje obecně zesílení nebo též rozmnožení
(z lat. amplificare – zvětšovat). V případě DNA tak označujeme
řízené laboratorní mnohočetné kopírování DNA, jehož smyslem
je získat ve větším počtu kopií DNA identickou s DNA původní.
Amplifikace se téměř vždy zaměřuje na určitý konkrétní úsek DNA,
který nás zajímá, a kopírování se týká právě pouze tohoto úseku
(lokusu).
Na jakém principu je amplifikace DNA prováděna?
Obecně v molekulární genetice najdeme celou řadu postupů,
kterými může být DNA namnožena. Zcela vůdčí metodou však je
PCR – polymerázová řetězová reakce (angl. Polymerase Chain

147
Reaction). PCR je prakticky jedinou amplifikační metodou, užívanou
ve forenzní genetice, a to v řadě postupů; proto si ji zde blíže
popíšeme.
Co je PCR a jak probíhá?
PCR neboli polymerázová řetězová reakce je cyklická enzymatická
reakce, prováděná v laboratorních podmínkách (in vitro), která
je ve své podstatě napodobením přirozeného kopírování DNA
v buňkách (in vivo).
PCR se provádí v zařízení zvaném thermocycler (termocyklér),
což je de facto termoblok, do něhož se vloží zkumavky s reakční
směsí a který je schopný v rychlém sledu měnit teplotu v těchto
zkumavkách (proces je naprogramován a přesně řízen počítačovým
čipem).
Vstupní reakční směs obsahuje následující komponenty:
1. PCR pufr, tj. vodu obsahující ionty a další látky potřebné pro
správný průběh reakce (Mg2+, případně K+, NH4+ apod.)
2. volné nukleotidy, označované jako dNTPs, což jsou stavební
kameny DNA, z nichž budou tvořeny nově vznikající řetězce
DNA; až na výjimky je pro PCR užívána vyvážená směs 4
základních dNTP – dATP, dCTP, dGTP a dTTP.
3. primery – krátké úseky DNA o délce cca 20–30 nukleotidů,
které slouží k nalezení specifického místa v DNA, jež má být
kopírováno, a od nichž toto kopírování začíná; primery jsou
navrženy tak, aby bezchybně párovaly právě pouze na začátek a
konec zájmového úseku na každém z obou DNA vláken;
(primery v páru pak označujeme jako F – forward a R– reverse.
4. DNA polymerázu – enzym, který provádí vlastní kopírování;
jsou užívány tzv. termostabilní polymerázy, které dobře snášejí
vysoké teploty, kterým jsou během PCR vystaveny.

148
5. templátovou DNA neboli matriční DNA, což je vlastní
předloha, podle níž má kopírování probíhat; nejčastěji se jedná
o DNA izolovanou z nějakého biologického materiálu (stopy
nebo srovnávacího vzorku).
PCR probíhá formou cyklů, které se opakují 20–40krát (počet se liší
dle aplikace). Jeden cyklus zahrnuje tři po sobě jdoucí fáze:
1. fázi denaturační, při níž vlivem vysoké teploty (cca 95°C)
dojde k oddělení obou řetězců DNA (dvoušroubovice se
„rozplete“ na dvě samostatná vlákna); fáze trvá zpravidla
několik desítek vteřin.
Obrázek 46 – Složení PCR směsi a první – denaturační fáze PCR.

149
2. fázi hybridizační (anelační), při níž se teplota sníží
(konkrétní teplota závisí na sekvenci a délce užitých primerů,
pohybuje se nejčastěji kolem 50–65°C) a dojde k nasednutí
primerů na oba volné řetězce DNA v místech, kam bezchybně
párují (tj. na začátek a konec úseku, který chceme kopírovat);
tato fáze rovněž trvá několik desítek vteřin.
Obrázek 47 – Druhá – anelační – a třetí – elongační fáze PCR.
3. fázi prodlužovací (extenzní, elongační), při níž se teplota
mírně zvýší (na cca 72°C) a polymeráza začne od místa
navázaného primeru připojovat na základě sekvence v matriční
DNA komplementární nukleotidy (od primeru dále tedy vzniká
podél jednořetězcové DNA druhé komplementární vlákno, a
vytváří se tak dvoušroubovice; současně probíhá stejný proces

150
na komplementárním templátovém vláknu). Délka fáze závisí
na rychlosti polymerázy a na tom, jak dlouhý úsek je třeba
okopírovat, zpravidla se pohybuje v řádu desítek vteřin až
několika minut.
Po ukončení prodlužovací fáze ihned následuje denaturační fáze
následujícího cyklu. Nově vzniklá vlákna se při ní oddělí od matriční
DNA a v následující anelační fázi mohou sama posloužit jako
matriční DNA. Tímto mechanismem narůstá exponenciálně počet
nově nasyntetizovaných vláken DNA v roztoku; po několika
desítkách cyklů tak PCR směs obsahuje miliardy kopií příslušného
úseku DNA. Nutno ovšem upozornit, že PCR nemůže probíhat
donekonečna: po určitém počtu opakování dojde reakce do tzv. fáze
plató, kdy již nové řetězce nevznikají, a zvýšení počtu cyklů tak už
nevede ke zvýšení množství amplifikované DNA.
Čím může být PCR negativně ovlivněna?
Negativní vliv ve smyslu snížení účinnosti amplifikace (někdy až
k nule – amplifikace neprobíhá) mají látky zvané PCR inhibitory,
zmíněné již v kapitole týkající se izolací DNA.
Účinnost amplifikace též může být snížena nesprávným složením
PCR reakční směsi – nedostatečnou koncentrací dNTPs, Mg2+,
primerů či DNA polymerázy. Naopak – i příliš vysoká koncentrace
Mg2+ snižuje účinnost reakce, neboť brání úplné denaturaci
(rozpletení dvoušroubovice). V praxi však je master mix většinou
namíchán předem jako součást komerčního kitu. Účinnost snižuje i
vyšší než optimální hybridizační teplota (ani specifické primery
se „neudrží“).
Zásadní vliv na účinnost amplifikace má též vstupní množství
templátové DNA, jak již jsme zmínili v kapitole o kvantifikaci DNA:
příliš nízké množství templátové DNA vede k pomalejšímu „náběhu“

151
reakce a mohou se při něm snáze uplatnit některé náhodné
(stochastické) efekty, jako je drop–out, o nichž bude pojednáno
v kapitole o interpretaci dat (nicméně – PCR může zdárně
proběhnout i pokud je ve vstupní reakci přítomna jedna jediná
nepoškozená kopie cílového úseku DNA). Zcela fatální důsledky pro
PCR však má příliš vysoké vstupní množství DNA, které může vést
až k úplné inhibici reakce.
Negativní vliv ve smyslu nesprávného kopírování (vznikající
řetězec není zcela identický s předlohou nebo není identický vůbec)
může být dán jednak nesprávným nasednutím primerů (jinam,
než kam by měly), pak hovoříme o snížení specifity reakce, jednak
chybami při činnosti polymerázy.
Snížení specifity může být vyvoláno nesprávným složením PCR
reakční směsi – zejména příliš vysokou koncentrací Mg2+, a též
příliš nízkou anelační teplotou (v obou případech se primery
mohou „udržet“ i na místech, kam zcela nepárují).
Základní chyby, k nimž dochází během činnosti polymerázy, je
zařazení nesprávného nukleotidu a prokluz polymerázy.
Frekvence zařazení nesprávného nukleotidu je dána přirozenou
chybovostí té které polymerázy (nikdo nejsme dokonalý). Obecně
menší chybovost mají tzv. polymerázy s korektorskou funkcí
(angl. proofreading, někdy označované též jako HiFi polymerázy),
které po sobě zařazený nukleotid zkontrolují. Prokluz polymerázy je
specifickou chybou, k níž dochází při amplifikaci STR lokusů a jejímž
důsledkem je vznik tzv. koktavého píku (angl. stutter peak), a bude
blíže popsáno v kapitole o analýze LCN a degradované DNA.
Co je monoplexová a co multiplexová PCR?
Amplifikujeme–li v rámci jedné reakční směsi (tj. v jedné zkumavce)
s použitím jednoho páru primerů pouze jediný DNA lokus, hovoříme

152
o monoplexové PCR. Amplifikujeme–li lokusů více, pak zpravidla
používáme termín vystihující počet amplifikovaných lokusů –
duplexová PCR (= 2 lokusy), triplexová PCR (= 3 lokusy) atd.,
u většího počtu lokusů můžeme použít též obecný pojem
multiplexová PCR (= mnoho lokusů).
Co je symetrická a co asymetrická PCR?
Pokud jsou oba primery v rámci primerového páru přidány do
reakce ve stejném množství, hovoříme o symetrické PCR.
U některých aplikací je však vhodné, aby bylo preferováno
kopírování v jednom směru, a do reakce proto přidáváme příslušný
primer ve větším množství – pak hovoříme o asymetrické PCR. Její
extrémní formou je PCR s použitím jediného primeru, s níž se
setkáme např. při sekvenaci (popsána bude u analýzy HVR mtDNA).
Při použití jediného primeru neroste množství okopírovaných DNA
fragmentů v roztoku exponenciálně, ale lineárně.
Kde v rámci forenzně genetických technologií se
amplifikace DNA pomocí PCR uplatňuje?
Užití PCR ve forenzní genetice je široké a aplikací najdeme opravdu
mnoho. Rámcově lze říci, že:
1. Účelem PCR může být pouhé namnožení zájmových lokusů
před provedením dalších analýz (u SNP lokusů, HVR mtDNA
apod.).
2. PCR může být jedním z více kroků nějaké analýzy (mono– či
multiplexová PCR jako součást analýzy STR lokusů, asymetrická
PCR jako součást sekvenace HVR mtDNA apod.)
3. Na samotné PCR může být přímo založena určitá analýza
(qPCR jako metoda specifické kvantifikace).

153
Elektroforéza DNA
Co je elektroforéza?
Elektroforézou nazýváme souhrnně separační metody založené
na pohybu nabitých částic ve stejnosměrném elektrickém poli,
přičemž různé částice mají různou elektroforetickou mobilitu (tj.
pohybují se v poli s rozdílnou rychlostí). Na základě této rozdílné
pohyblivosti tak dojde elektroforézou k separaci jednotlivých typů
částic.
Obecně je mobilita dána především velikostí elektrického napětí a
fyzikálně chemickými vlastnostmi média, v němž separace probíhá.
Existuje celá řada možných uspořádání elektroforézy – např.
zónová, kapilární, elektroforéza v plynné fázi atd.
Jak funguje elektroforéza DNA?
DNA je za běžných podmínek díky fosfátu ve své struktuře záporně
nabitá molekula, a ve stejnosměrném elektrickém poli se proto
pohybuje směrem ke kladné elektrodě (anodě). Pokud je tento
pohyb prováděn v médiu, které ho částečně brzdí – např. v gelu,
dojde k postupnému rozdělení DNA fragmentů podle jejich
elektroforetické mobility.
Relativní elektroforetická mobilita je primárně dána délkou
fragmentu DNA (s počtem bazí mobilita klesá, tj. čím delší fragment
je, tím pomaleji se pohybuje) a případnou přítomností na DNA
navázaných dalších molekul – např. fluorescenčních značek
(navázaná molekula zvyšuje molekulární hmotnost celého
fragmentu, a tedy snižuje jeho mobilitu). Mobilitu ovšem ovlivňují
též další faktory, například podíl C a G nukleotidů ve fragmentu (C a
G nukleotidy jsou těžší než A a T nukleotidy) a též konformace
molekul DNA. Nejčastěji jsou proto elektroforeticky separovány

154
lineární fragmenty DNA, kdy jejich mobilita není rozdílnou
konformací ovlivněna. Elektroforéza nám v takovém případě
pomůže přesně určit délku fragmentů DNA ve vzorku.
Obrázek 48 – Princip elektroforézy DNA.
S jakými typy elektroforézy DNA se lze setkat při
forenzně genetických zkoumáních?
V rámci metod forenzní genetiky se užívají především dvě možná
uspořádání elektroforézy – klasická gelová elektroforéza (GE) a
kapilární (gelová) elektroforéza (CE).
Co je klasická gelová elektroforéza DNA?
Při klasické gelové elektroforéze (GE) je vzorek DNA nanesen do
malé jamky v gelovém plátu ponořeném v pufru mezi elektrodami

155
(plát může být umístěn vodorovně či svisle) a poté je na elektrody
vloženo napětí. DNA ze vzorku se začne pohybovat gelem směrem
k anodě, přičemž – jak již bylo popsáno – kratší fragmenty putují
rychleji než delší. V praxi se nejčastěji lze setkat s užitím
agarózového (AGE) či polyakrylamidového gelu (PAGE).
Po určité době je elektroforéza ukončena a DNA v gelu je zobrazena
(zobrazovacích metod je nepřeberné množství – může to být např.
pomocí fluorescenční barvy, radioaktivně atd.). Na gelu se objeví
místa, v nichž se nachází DNA; často jsou to tzv. bandy – proužky
obsahující fragmenty stejné délky. Vzdálenost bandu od nanášecí
jamky je tím větší, čím kratší fragmenty obsahuje.
Aby bylo možno určit přesnou délku fragmentů v bandu, užívá se
tzv. velikostní standard (žebříček), což je uměle připravená směs
fragmentů několika známých délek (žebříček tedy např. obsahuje
fragmenty o délce 50, 100, 150, 200, 300 a 400 bazí). Žebříček je
nanesen vedle vzorku do sousední jamky a během elektroforézy se
separuje za stejných podmínek a stejným způsobem jako vzorek. Po
skončení elektroforézy a zobrazení DNA lze délku fragmentů
v jednotlivých bandech vzorku zjistit srovnáním s polohou bandů
žebříčku.
V jakém přístroji je klasická gelová elektroforéza
DNA prováděna?
Klasická gelová elektroforéza nevyžaduje žádné vysoce
specializované vybavení. Postačuje elektroforetická vana
s lůžkem pro gel, a to v horizontální (AGE) či vertikální (PAGE)
orientaci, a spolehlivý programovatelný zdroj napětí.
Existuje i vysoce sofistikovaná a automatizovaná forma gelové
elektroforézy – automatický analyzátor, v němž je celý proces
řízený počítačem a detekce fragmentů v gelu probíhá pomocí

156
fluorescenčního scanneru; v rámci molekulárně genetických technik
je však tento přístroj vytěsňován genetickými analyzátory
založenými na principu kapilární elektroforézy.
Co je kapilární elektroforéza DNA?
Při kapilární elektroforéze (CE) je vzorek elektrickým pulsem
nabrán na začátek tenké skleněné kapiláry naplněné polymerem
lineárního akrylamidu nebo polydimethylamidu. Na kapiláru je
pomocí elektrod zanořených do pufru přivedeno napětí a vzorek
začne putovat kapilárou, přičemž obdobně jako u klasické
gelové elektroforézy dochází k separaci fragmentů na základě
rozdílné mobility.
V případě kapilární elektroforézy je detekce fragmentů prováděna
v průběhu času: každý fragment je již před analýzou označen
fluorescenční barvou (zpravidla začleněním označeného primeru
v průběhu PCR). V koncové části kapiláry se nachází okénko, kterým
prosvěcuje laserový paprsek; ten v okamžiku průchodu
fluorescenčně značených fragmentů okénkem vybudí silnou
fluorescenci (světelné záření), která je zaznamenána CCD
detektorem (stejným, jako je v digitálních fotoaparátech).
Ani kapilární elektroforéza se neobejde bez standardizace pomocí
velikostního žebříčku. V tomto případě jej přidáváme přímo do
analyzovaného vzorku, proto jej nazýváme vnitřní žebříček.
Čím kratší fragment je, tím dříve doputuje k okénku v kapiláře:
zatímco u zonální elektroforézy tedy určujeme délku neznámého
fragmentu podle relativní vzdálenosti od nanášecí jamky, u
kapilární elektroforézy ji určujeme podle relativní doby, kterou
fragment potřebuje k dosažení okénka kapiláry (v obou případech
na základě porovnání s velikostním standardem – žebříčkem).

157
Obrázek 49 – Princip kapilární elektroforézy fluorescenčně značených fragmentů DNA.
V jakém přístroji je kapilární elektroforéza DNA
prováděna?
Kapilární elektroforéza je proces technicky velmi náročný, a proto je
prováděn téměř výhradně pomocí automatizovaných genetických
analyzátorů (často je pro ně užíván nepřesný leč vžitý slangový
název sekvenátor). Nejrozšířenějšími jsou bezesporu analyzátory
firmy Applied Biosystems (součást Life Technologies). Dnes již jsou
laboratoře běžně vybaveny 4, 16 i 96kapilárovými verzemi tohoto
přístroje, umožňujícími současnou analýzu příslušného počtu
vzorků.

158
Analýza STR lokusů
Jakými metodami jsou analyzovány STR lokusy?
Přestože technicky existuje několik možností, jak analyzovat STR
lokusy, forenzní praxi v současnosti dominuje jediná z nich: analýza
založená na PCR amplifikaci a následném stanovení délky
amplifikovaného fragmentu pomocí kapilární elektroforézy.
Jak tato analýza teoreticky probíhá?
Pro srozumitelnost popíšeme celý proces nejprve na analýze
jediného STR lokusu. V prvém kroku je připraven PCR mastermix
(viz popis PCR), který obsahuje jediný pár primerů; jeden z těchto
primerů nasedá na známé místo v DNA těsně před a druhý těsně za
náš zájmový STR lokus. Protože jsme předvídaví, je jeden z těchto
primerů označen fluorescenční barvou – tak, aby nám posléze
umožnil detekci fragmentu.
Poté je v thermocycleru provedena klasická PCR, přičemž vznikají
fragmenty dlouhé přesně tolik párů bazí, kolik měří celý úsek
ohraničený oběma primery. Protože polymorfismus STR lokusů –
jak již bylo vysvětleno dříve – je polymorfismem délkovým, bude
u různých alel našeho zájmového lokusu výsledný fragment
různě dlouhý.
Po skončení PCR proto potřebujeme stanovit délku vzniklých
fragmentů; k tomu použijeme výše zmíněnou kapilární
elektroforézu prováděnou v genetickém analyzátoru. Část
amplifikátu, tj. produktu PCR, spolu s vnitřním žebříčkem
napipetujeme do zkumavky s formamidem, který má silně
denaturující účinek, a tedy zajišťuje, že fragmenty zůstanou
v jednořetězcovém stavu. Poté již proces probíhá automaticky podle
předem nastaveného programu – vzorek je elektrokineticky

159
injikován do kapiláry, na kterou je následně vloženo napětí. Při
průchodu DNA fragmentu okénkem kapiláry excituje laser
fluorescenční barvičku, která je na fragmentu vázána, a ta následně
emituje silné záření v určité oblasti spektra, což se projeví
v elektroforetogramu (časovém záznamu elektroforézy)
charakteristickým píkem určité barvy.
Obrázek 50 – Výsledek monoplexové a multiplexové analýzy STR lokusů.
Srovnáním s píky vnitřního žebříčku software vypočte s vysokou
přesností délku námi amplifikovaného fragmentu. Protože víme,
kam přesně nasedaly primery a jak dlouhé jsou tedy konstantní
úseky po obou stranách STR, odečteme jejich délku od stanovené
délky fragmentu, čímž získáme vlastní délku repetice. Vydělením
této hodnoty délkou základního motivu repetice zjistíme, kolikrát se
v našem fragmentu tento motiv opakuje; toto číslo současně
označuje příslušnou alelu.

160
Př.: Kapilární elektroforézou jsme změřili, že námi amplifikovaný
fragment má délku 255 bp. Víme, že ohraničením pomocí primerů
byly do amplifikace kromě vlastní STR též zahrnuty konstantní úseky
po obou jejích stranách v celkové délce 167 bp. Délka samotné
repetice je tedy 255 – 167 = 88 bp. Víme, že tato repetice je
tetranukleotidová, neboli že základní motiv má délku 4 bp. Jednoduše
tudíž zjistíme, že v našem fragmentu je 88/4=22 opakování, a jedná se
tedy o alelu 22.
Jak je tato analýza prováděna v praxi?
Oproti popisu v předchozích odstavcích se reálná analýza STR
poněkud liší, a to především v následujícím:
vyjma velmi specifických situací není prováděna analýza
jediného lokusu, ale analyzuje se celý set lokusů najednou;
k tomu slouží již dříve zmíněná multiplexová PCR, při níž je
použito více párů primerů – vždy jeden pár pro jeden lokus;
veškeré komponenty potřebné pro analýzu jsou již
předpřipraveny a ověřeny výrobcem a do laboratoře jsou
dodávány ve formě tzv. kitu neboli soupravy. V českém
prostředí hojně užívanými kity jsou např. PowerPlex 16,
Identifiler, Identifiler Plus, MiniFiler, PowerPlex Y či Y–filer;
zejména pro analýzy příbuznosti jsou užívány kity s velkým
počtem testovaných lokusů – např. PowerPlex 21 a PowerPlex
Y23;
kromě primerů pro STR lokusy tyto kity zpravidla obsahují pár
primerů pro amplifikaci specifického úseku v genu pro
amelogenin, který je užíván ke zjištění pohlaví nositele
analyzované DNA;
celý proces na genetickém analyzátoru je již přednastaven,
odečet délek fragmentů a jejich přepočet na číslo alely je
prováděn automaticky pomocí software; znalec se do

161
procesu zapojuje jednak jako kontrolní entita (zda vše probíhá
podle plánu), jednak jako ten, kdo vyhodnocuje nejednoznačné
nebo komplikované výsledky – lze ho tedy přirovnat k pilotovi
dopravního letounu, který sice ve standardních situacích hojně
využívá autopilota, avšak pro kontrolu průběhu letu a řešení
situací nestandardních je jeho přítomnost absolutně klíčová.
Dlužno podotknout, že stopy jsou zřídkakdy povahy
standardního vzorku.
Analýza HVR mt DNA
Jakými metodami jsou analyzovány HVR
v mitochondriální DNA?
Polymorfismus hypervariabilních oblastí mtDNA je sekvenční;
k jeho zjišťování jsou proto užívány buď metody sekvenační, nebo
metody založené na hybridizaci s tzv. alelově–specifickou
sondou (SSO) (angl. Sequence–Specific Oligonucleotides).
Jak je prováděna sekvenace mtDNA?
Sekvenací rozumíme stanovení pořadí nukleotidů
v analyzovaném úseku DNA. Existuje mnoho konceptů a aplikací
této metody, popíšeme zde velmi stručně pouze postup v rámci
forenzní praxe v současnosti nejužívanější – sekvenaci pomocí
asymetrické PCR s tzv. terminátory.
Při tomto postupu je HVR oblast nejprve namnožena pomocí
jednoho páru primerů klasickou PCR. Poté následuje přečištění
amplifikátu a též zpravidla kontrola úspěšnosti (detekce
namnoženého fragmentu). Ten pak dále poslouží jako templát do
následující, tentokráte asymetrické PCR reakce.

162
Při sekvenační reakci je v reakční směsi přítomen pouze jeden
z páru primerů (F nebo R), a nové fragmenty tak vznikají pouze
podle jednoho z obou komplementárních řetězců sekvenovaného
fragmentu. Reakční směs dále obsahuje kromě „obyčejných“
deoxyribonukleotidů (dNTP), jež slouží jako stavební kameny nově
vznikajících řetězců, též fluorescenčně značené
dideoxyribonukleotidy (ddNTP), nazývané pro svou úlohu v reakci
terminátory: pokud je totiž do nově vznikajícího řetězce zařazen
ddNTP namísto standardního dNTP, nemůže polymeráza na něj již
připojit další stavební kámen, a syntéza řetězce tak končí. Je–li
poměr dNTP a ddNTP cca 100:1, dosáhneme stavu, kdy výsledný
produkt této asymetrické PCR sestává z fragmentů různých délek,
přičemž každý fragment má na konci fluorescenčně značený
terminátor. Stejně jako dNTP, jsou i ddNTP v reakci přítomny 4
druhy (ddATP, ddCTP, ddGTP a ddTTP), každý značený jinou
fluorescenční barvou.
Produkt asymetrické PCR je následně elektroforeticky rozdělen
(nejběžněji pomocí kapilární elektroforézy); na elektroforetogramu
se délkově odlišné fragmenty zobrazí jako jednotlivé píky, přičemž
barva píku odpovídá typu ddNTP, který byl do fragmentu zařazen.
Celý elektroforetogram pak tedy má charakter „pík–za–píkem“
(každý následující pík představuje fragment o 1 bp delší) a
převedením barvy píků na typ ddNTP můžeme odečíst kompletní
sekvenci.
Velmi často se při této sekvenaci provádějí dvě paralelní
asymetrické PCR, přičemž do každé je použit jiný z původního páru
primerů (do jedné F, do druhé R); získané sekvence z obou reakcí
musejí být komplementární. Postup se užívá především tehdy,
pokud je z nějakého důvodu obtížné bezchybně „přečíst“ celou
délku zájmového úseku DNA s pomocí jednoho primeru; tímto
důvodem může být např. velká délka úseku, malá koncentrace
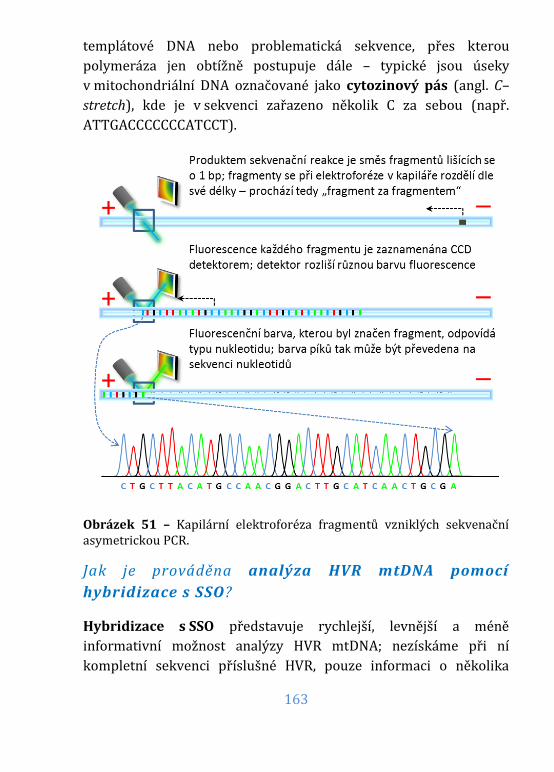
163
templátové DNA nebo problematická sekvence, přes kterou
polymeráza jen obtížně postupuje dále – typické jsou úseky
v mitochondriální DNA označované jako cytozinový pás (angl. C–
stretch), kde je v sekvenci zařazeno několik C za sebou (např.
ATTGACCCCCCCATCCT).
Obrázek 51 – Kapilární elektroforéza fragmentů vzniklých sekvenační asymetrickou PCR.
Jak je prováděna analýza HVR mtDNA pomocí
hybridizace s SSO?
Hybridizace s SSO představuje rychlejší, levnější a méně
informativní možnost analýzy HVR mtDNA; nezískáme při ní
kompletní sekvenci příslušné HVR, pouze informaci o několika

164
úsecích této HVR jako celku. Vybrány pro testování jsou úseky
zahrnující nejpolymorfnější nukleotidy. Analýza pomocí SSO se
uplatňuje především jako screeningová metoda při vyšetřování
většího množství stop či osob.
Podstata metody je velmi jednoduchá: pokud použitá SSO sonda je
zcela komplementární k příslušnému úseku HVR mtDNA ve
vyšetřovaném vzorku, dojde k jejímu navázání (hybridizaci) na
tento úsek, což je posléze detekováno nějakou vhodnou zobrazovací
metodou. Pokud se však sekvence sondy a příslušného úseku liší
byť v jediném nukleotidu, k hybridizaci nedojde. Získáme tak
informaci, zda je tento úsek jako celek komplementární se sondou či
nikoli. Pro analýzu lze užít i více různých sond pro stejný úsek, čímž
se analýza stane informativnější.
Analýza SNP lokusů
Jakými metodami jsou analyzovány SNP lokusy?
Existuje celá řada principů, na nichž může být zjištění genotypu
jednonukleotidových polymorfismů založeno. Ve forenzní praxi se
uplatňují především ty, které umožňují současnou genotypizaci
velkého množství SNPs, které je třeba k dosažení potřebné
vypovídací hodnoty analýzy. K základním patří minisekvenování
například pomocí soupravy SNaPshot; jeho obliba tkví i v tom, že jej
lze provádět se stejným laboratorním vybavením, jaké je užíváno
pro analýzu STR lokusů.
Jak je prováděna analýza SNPs minisekvenováním?
Při minisekvenování je nejprve pomocí klasické PCR reakce
namnožen úsek obsahující testovaný SNP. Poté je reakční směs
vystavena působení enzymů, které rozloží zbytky
nespotřebovaných primerů a zbylé volné dNTP.

165
Po úvodní amplifikaci následuje krok označovaný jako prodloužení
primeru: do reakce je přidán tzv. extenzní primer nasedající zcela
těsně před testovaný SNP, a směs čtyř různou barvou fluorescenčně
značených ddNTP neboli terminátorů (týchž, jaké byly popsány
v části věnované technologii sekvenování mtDNA). Polymeráza poté
na základě komplementarity s naším SNP připojí k primeru jediný
ddNTP (odtud název minisekvenování). Vzniklý fluorescenčně
značený produkt je poté analyzován pomocí kapilární elektroforézy:
barva detekovaného píku odpovídá typu terminátoru, který byl na
základě komplementarity s testovaným SNP do fragmentu zařazen.
Obrázek 52 – Výsledek monoplexové a multiplexové analýzy SNP lokusů.
Tato jednoduchá metoda může být úspěšně multiplexována, tj. je
množeno a analyzováno více zájmových lokusů najednou; pro
rozlišení příslušnosti jednotlivých píků k jednotlivým SNP je

166
používáno nejčastěji připojení různě dlouhých poly–T řetězců (úsek
tvořený pouze nukleotidy T) k extenzním primerům, což změní
výslednou pozici jednotlivých píků v elektroforetogramu.
Nakládání se stopami obsahujícími
degradovanou DNA a/nebo malá množství DNA
Co je degradovaná DNA?
V části věnované biologickým stopám jsme se již podrobněji zmínili
o jevu zvaném biodegradace včetně degradace DNA. Popsali jsme
základní činitele, podílející se na degradaci, a možné důsledky pro
analýzu.
Obrázek 53 – Dva základní mechanismy degradace DNA – fragmentace a narušení struktury.
Jako degradovanou DNA souhrnně označujeme DNA, která již není
v původním nativním stavu (tj. taková, jaká byla v živé buňce), ale
byla poškozena fyzikálními, chemickými či biologickými činiteli.

167
Toto poškození může mít charakter fragmentace, tj. rozštěpení na
kratší úseky, a/nebo narušení struktury, tj. změny jednotlivých
bází (např. v důsledku oxidace), výpadky bází apod. Uvedené změny
mohou nastat v různé míře, proto mluvíme o různém stupni
degradace DNA.
Můžeme určit stupeň degradace DNA ve vzorku?
Posoudit, do jaké míry je DNA ve vzorku poškozena a zda bude
vhodná k námi zamýšlené analýze, není nemožné, stále však platí, že
nejlepším testem, zda bude analýza fungovat, je analýza sama.
Obvykle u vzorku možnou degradaci již předvídáme na základě
nám dostupných informací (ať už získaných ze spisu, ohledáním
biologického materiálu či prostou zkušeností s daným typem
materiálu). Máme–li analyzovat čerstvou krev, odebranou osobě
den předem a uchovanou v ledničce, prakticky s jistotou očekáváme,
že DNA nebude ani mírně degradovaná. U vzorku kosti z 18. století
si naopak můžeme být vyšším stupněm degradace téměř jisti.
Lze provést některé orientační analýzy stupně degradace –
například rozdělit izolovanou DNA elektroforeticky na agarózovém
gelu a obarvit sloučeninou ethidiumbromid, která v UV světle září:
pokud se na gelu objeví dlouhá „rozplizlá“ šmouha v oblasti
krátkých fragmentů, máme co do činění s velmi fragmentovanou
DNA; pokud naopak intenzivně září pouze malá ohraničená skvrna
v oblasti, kde očekáváme velmi dlouhé fragmenty, je naše DNA
pravděpodobně velmi málo poškozena.
Míra degradace může být hodnocena i dalšími postupy, jako je
qPCR. Provedení takových testů sice nestojí příliš mnoho času,
avšak spotřebovává se při něm část vzorku; u nepatrných stop
s minimálním množstvím využitelné DNA si takové plýtvání dovolit
nemůžeme, a musíme přistoupit přímo k pokusu o analýzu.

168
Může být degradovaná DNA před analýzou nějak
reparována?
Zmínili jsme dva možné charaktery změn v degradované DNA –
fragmentaci a narušení struktury. Fragmentace je při současných
znalostech a schopnostech molekulární genetiky poškození
laboratorně nereparovatelné – neexistuje laboratorní postup,
kterým by bylo možno vyhledat a seřadit fragmenty tak, jak
původně byly v DNA za sebou, a znovu je spojit (v jiných odvětvích
molekulární genetiky se používají některé postupy „skládání
sekvence až v počítači“, ve forenzní oblasti se však zatím
neuplatňují.) Naproti tomu existují některé dílčím způsobem
úspěšné postupy opravy narušené struktury DNA pomocí
enzymatických koktejlů. Jejich užití ve forenzní genetice je však
zatím spíše otázkou budoucnosti.
Existují speciální postupy analýzy degradované
DNA?
Ano. Základním přístupem je maximalizace výtěžnosti izolačních
technik (tj. nelze si dovolit ztráty) a minimalizace délky
fragmentu potřebného k provedení příslušné analýzy (neboť je
mnohem větší pravděpodobnost, že ve vzorku se bude nacházet
několik nepoškozených zájmových fragmentů o délce 100 bp než
o délce 300 bp). V rámci analýzy STR lokusů se v tomto směru
uplatňuje především užití tzv. miniSTRs.
Co jsou miniSTRs a jak se analyzují?
Jako miniSTRs označujeme analýzy STR lokusů s pomocí primerů
nasedajících těsně před a za danou repetici; tímto blízkým
umístěním primerů se minimalizuje délka potřebného
neporušeného fragmentu, a tedy i zvyšuje pravděpodobnost, že
analýza bude úspěšná. Testování se zaměřuje na:
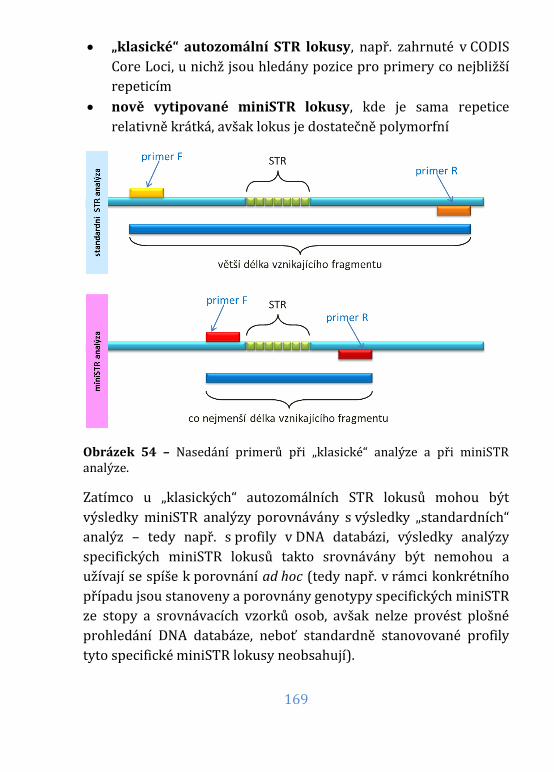
169
„klasické“ autozomální STR lokusy, např. zahrnuté v CODIS
Core Loci, u nichž jsou hledány pozice pro primery co nejbližší
repeticím
nově vytipované miniSTR lokusy, kde je sama repetice
relativně krátká, avšak lokus je dostatečně polymorfní
Obrázek 54 – Nasedání primerů při „klasické“ analýze a při miniSTR analýze.
Zatímco u „klasických“ autozomálních STR lokusů mohou být
výsledky miniSTR analýzy porovnávány s výsledky „standardních“
analýz – tedy např. s profily v DNA databázi, výsledky analýzy
specifických miniSTR lokusů takto srovnávány být nemohou a
užívají se spíše k porovnání ad hoc (tedy např. v rámci konkrétního
případu jsou stanoveny a porovnány genotypy specifických miniSTR
ze stopy a srovnávacích vzorků osob, avšak nelze provést plošné
prohledání DNA databáze, neboť standardně stanovované profily
tyto specifické miniSTR lokusy neobsahují).

170
Co je LCN?
Jako LCN (angl. Low–Copy Number, „malý počet kopií“) neboli LT–
DNA (angl. Low–Template DNA) jsou souhrnně označovány vzorky
s malým až velmi malým obsahem DNA využitelné pro analýzu.
Existují speciální postupy analýzy LCN (LT–DNA)?
Pro analýzu LCN vzorků bylo až dosud vyvinuto a je užíváno několik
specifických strategií, jejichž cílem je zvýšení citlivosti testování.
Základními jsou
zvýšení počtu PCR cyklů při amplifikaci, které může vést ke
zvýšení množství výsledného produktu, avšak zvyšuje i riziko
nespecifické amplifikace (tj. jevů typu drop–in);
užití tzv. nested PCR, kdy je nejprve lokus amplifikován
s pomocí páru primerů nasedajících dále od sebe (tzv. vnější
pár primerů) a poté s pomocí páru primerů nasedajících uvnitř
prvního amplikonu blíže k sobě (tzv. vnitřní pár primerů), což
může vést ke zvýšení množství výsledného produktu, avšak
vzhledem k manipulaci se vzorkem se zvyšuje i riziko
kontaminace;
snížení objemu PCR reakce, tedy jakési „zakoncentrování“
celé reakce; negativní efekt ovšem může mít současné
zakoncentrování PCR inhibitorů;
další snahy o vylepšení PCR, např. přidáním většího množství
polymerázy apod.
zvýšení objemu vzorku injektovaného do kapilární
elektroforézy
přečištění PCR produktu před injekcí do kapilární
elektroforézy, což rovněž zvýší množství vzorku, který se
nabere do kapiláry
užití „nadějnějších lokusů“, např. mtDNA, která je ve vzorku
přítomna v několikařádově více kopiích než autozomální lokusy

171
Jak se degradace DNA a/nebo malé množství DNA
projeví na analýze?
K tomu, aby analýza poskytla pozitivní výsledek, je teoreticky třeba,
aby ve vzorku byl přítomen minimálně jeden nepoškozený
fragment DNA zahrnující celý testovaný úsek DNA – v případě
PCR technik je to celý úsek mezi F a R primerem včetně obou konců,
kam tyto primery nasedají. V praxi je však pozitivní analýza vzorku
s jedinou kopií zájmového lokusu značně nejistá, závisí významně
na použité technologii a spolehlivých výsledků lze zpravidla
dosáhnout až při řádově vyšším počtu kopií, např. 10.
S postupující degradací DNA se počet nepoškozených fragmentů
zahrnujících testovaný úsek ve vzorku snižuje, ve vzorcích typu LCN
je rovněž počet nepoškozených fragmentů zahrnujících testovaný
úsek velmi malý; tím se mohou během PCR více a více projevit tzv.
stochastické efekty – laicky řečeno vlivy náhody, které snižují
spolehlivost získaných dat. Se snížením počtu nepoškozených
fragmentů zahrnujících testovaný úsek rovněž dramaticky roste
riziko, že se při analýze projeví přítomnost třeba i malého množství
kontaminující DNA (neboli cizorodé DNA zanesené do vzorku).
K jakým stochastickým efektům může docházet při
analýze degradované DNA či LT–DNA?
Nejčastějšími stochastickými efekty jsou
nerovnováha píků u heterozygota
alelický drop–out
lokusový drop–out
zvýšení stutteringu (tj. tvorby koktavých píků)
alelický drop–in

172
Jako nerovnováhu píků u heterozygota označujeme stav, kdy oba
píky reprezentující dvě alely heterozygotního lokusu nejsou stejně
vysoké (tak, jak by u vzorku jediné osoby měly být), ale svou výškou
se významně liší; pokud se výška jednoho z obou píků sníží až k nule
(tj. pík není de facto přítomen), mluvíme o ztrátě alely neboli
alelickém drop–outu. Dojde–li ke ztrátě obou alel lokusu (tj. žádný
pík daného lokusu není de facto přítomen), mluvíme o ztrátě lokusu
neboli lokusovém drop–outu.
Při amplifikaci lokusů typu STR dochází u čtyř– a
méněnukleotidových repetic k občasnému prokluzu polymerázy:
polymeráza během kopírování repetice „odpadne a znovu nasedne“,
přičemž v určitém procentu případů nasedne na templátovou DNA
až o motiv dále než odpadla, takže výsledný produkt amplifikace je
o motiv kratší. Tím vznikne určité množství fragmentů, které se
v elektroforetogramu objeví jako menší pík předsazený před vlastní
alelou o délku motivu, tedy kupříkladu o 4 bp. Tento jev nazýváme
zadrhávání, koktání (stuttering) a zmíněný „předpík“ označujeme
jako koktavý pík (stutter). U standardních vzorků s dostatečným
množstvím nepoškozené DNA se při klasické analýze autozomálních
tetranukleotidových STR pohybuje míra stutteringu kolem 10–15
%, u degradované DNA či LCN vzorků však může významně
narůstat, což spolu s ostatními stochastickými jevy vede k nejistotě,
zda daný pík představuje pouze zvýšený stutter, nebo se jedná
o relevantní alelu.
Vzhledem k malému obsahu templátové DNA v degradovaných či
LCN vzorcích se při amplifikaci může projevit i sporadická
kontaminace (kontaminace malými DNA rezidui), a to nejčastěji
v podobě tzv alelických drop–inů, tj. píků „navíc“.
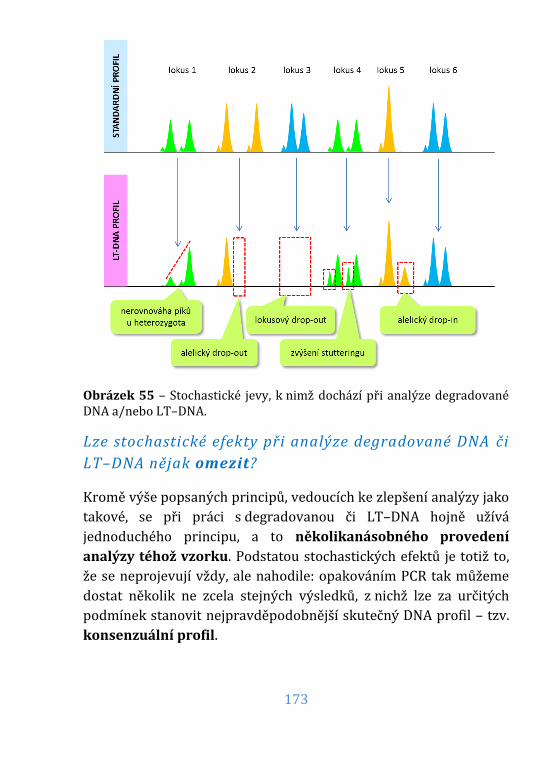
173
Obrázek 55 – Stochastické jevy, k nimž dochází při analýze degradované DNA a/nebo LT–DNA.
Lze stochastické efekty při analýze degradované DNA či
LT–DNA nějak omezit?
Kromě výše popsaných principů, vedoucích ke zlepšení analýzy jako
takové, se při práci s degradovanou či LT–DNA hojně užívá
jednoduchého principu, a to několikanásobného provedení
analýzy téhož vzorku. Podstatou stochastických efektů je totiž to,
že se neprojevují vždy, ale nahodile: opakováním PCR tak můžeme
dostat několik ne zcela stejných výsledků, z nichž lze za určitých
podmínek stanovit nejpravděpodobnější skutečný DNA profil – tzv.
konsenzuální profil.

174
Jaká hlavní rizika přináší užití vzorků s degradovanou
DNA a/nebo LCN vzorků ve forenzní praxi?
Jak již bylo naznačeno výše, analýza tohoto typu vzorků s sebou
vždy přináší snížení spolehlivosti získaných dat: uplatňují se snáze
stochastické efekty a roste riziko, že se projeví kontaminující DNA.
Přes tato negativa mohou být degradované a LCN vzorky
principiálně pro forenzní analýzu užity, pokud je s nimi správným
způsobem nakládáno a z nich získaná data jsou správně a obezřetně
interpretována. Na druhou stranu – data velmi sporné kvality a
původu již by neměla být referována jako relevantní a neměla by
být pro jakékoli forenzní interpretace užívána.

175
Interpretace forenzně genetických dat
Obecné kroky a pravidla interpretace dat
Co si představit pod pojmem interpretace dat?
Po vlastní analyzační fázi, tj. genetické analýze v laboratoři,
nastupuje neméně důležitá fáze interpretační; jejím smyslem je
zhodnotit získaná data, stanovit jejich spolehlivost a následně
určit jejich vypovídací hodnotu, tj. převést získanou informaci
z roviny úzce odborné do roviny reálného světa a života – neboli
říci, co vlastně výsledky analýzy v daném případě „znamenají“.
Obrázek 56 – Základní kroky procesu práce s daty neboli „od dat k posudku“.
Z čeho vycházíme při hodnocení spolehlivosti dat?
Abychom mohli odhadnout míru spolehlivosti dat (či naopak
chcete–li míru nejistoty), musíme především posoudit:

176
jak jsou získaná data technicky kvalitní, a jaká je tedy
spolehlivost vlastního odečtu dat
jak v principu spolehlivé jsou námi použité metody
analýzy a jak je z hlediska možného ovlivnění výsledků
bezpečný a vůči chybě odolný celý laboratorní proces
jak logicky konzistentní jsou jednotlivá dílčí zjištění
v procesu forenzně genetické analýzy navzájem (tj. zda si
např. dva dílčí výsledky zkoumání téhož vzorku neodporují)
jak dobře data odpovídají empirii (tj. zda jsou data
v souladu se zkušeností znalce a uživatele dat – soudce, či
naopak zda z ní vybočují).
Jakým způsobem posuzujeme vypovídací hodnotu dat?
Další posouzení dat by mělo vždy být založeno na inferenci
pravděpodobnostních jevů, tj. na vyjádření pravděpodobnosti
daného výsledku za platnosti dvou či více alternativních hypotéz.
Všechny ostatní způsoby posouzení dat (kategorické soudy,
frekventistický přístup aj.), které jsou užívány v některých jiných
forenzních oborech, jsou v rámci forenzně genetické analýzy
nedostatečné či dokonce hrubě zkreslující a znalec by se jich měl
vyvarovat.
Spolehlivost dat – technická kvalita dat
Jakou podobu mají výstupní laboratorní data?
Laboratorní data, která jsou výsledkem forenzně genetické analýzy,
mohou v principu mít nejrůznější podobu; v praxi se ovšem
nejčastěji setkáme s časovým záznamem změn fluorescence
v průběhu kapilární elektroforézy (tzv. elektroforetogramem), a to
jak u klasické identifikační STR analýzy, tak například při
sekvenování mtDNA či při minisekvenování SNP lokusů.

177
Co rozumíme pod pojmem technická kvalita
laboratorních dat?
Technickou kvalitou dat rozumíme to, jak zřetelně odečitatelná tato
data jsou, tj. do jaké míry jednoznačně z nich lze určit konkrétní
podobu studovaných polymorfismů DNA. Naše data mohou být
od vysoce kvalitních, úplných a bez nejasností, přes středně kvalitní,
neúplná a s určitými nejasnostmi až po zcela neinterpretovatelná.
Jak hodnotíme technickou kvalitu laboratorních dat?
Posouzení technické kvality dat spočívá v hodnocení celé řady
parametrů, jako je výška a tvar píků, vzájemné poměry výšek píků,
míra stutteringu, výška a charakter tzv. pozadí a podobně. Při
hodnocení přitom vycházíme ze standardu, který představují
data z validace (o validaci viz dále). Posouzení může být primárně
svěřeno kvalitnímu vyhodnocovacímu softwaru, stále ovšem platí,
že finální posouzení dat včetně jejich technické kvality je na
expertovi a jeho „cvičeném oku“.
Jak pracujeme s technicky středně a nízce kvalitními
daty?
Prvním krokem by vždy měla být snaha kvalitu dat zvýšit; v praxi
tak často po první analýze vzorku následuje i několik dalších
opakování s různými modifikacemi (změna vstupního množství
DNA do PCR, změna parametrů PCR, změna parametrů kapilární
elektroforézy atd.) Cílem je optimalizovat průběh celé analýzy
tak, aby výsledná data byla co nejčitelnější. Na tomto místě
zdůrazňuji, že se v žádném případě nejedná o snahu tak dlouho
zkoušet analýzu, až konečně dostanu výsledek, jaký potřebuji; jde
o snahu „vyladit“ celou analýzu tak, aby poskytla co nejúplnější a co
nejspolehlivější data.

178
V případě práce se silně degradovanou DNA a/nebo LCN vzorky,
kde se silně uplatňují již dříve popsané stochastické jevy (alelický
drop–out, nevyváženost píků atd.), se můžeme setkat se
sestavováním tzv. konsenzuálních profilů. Podstatou je
opakovaná analýza téhož vzorku s pomocí několika nezávislých PCR
a následné pomyslné spojení odečtených dat ze všech těchto analýz;
tím lze do značné míry eliminovat právě stochastické jevy, které se
objevují náhodně a pouze v některých PCR téhož vzorku.
V případě, že ani opakované analýzy a modifikované postupy
nevedou ke zkvalitnění dat, je třeba míru technické nekvality
zohlednit při stanovení vypovídací hodnoty dat.
V praxi tak můžeme například dospět k závěru, že:
u určitého autozomálního lokusu, v němž byla stanovena pouze
jediná alela (označme ji A), připouštíme možný drop–out druhé
alely; skutečný genotyp vzorku tudíž může být nejen AA, ale i AX,
kde X je libovolná jiná alela;
u určitého autozomálního lokusu byla jednoznačně stanovena
alela A a dále alela B cca poloviční výšky na pozici odpovídající
stutteru alely A, u níž vzhledem k technické kvalitě vzorku nelze
jednoznačně rozhodnout, zda se jedná o stutter či o druhou
relevantní alelu; skutečný genotyp vzorku tudíž může být jak AA,
tak AB;
u určitého autozomálního lokusu nebyla stanovena ani jedna
alela; skutečný genotyp vzorku tudíž může být jakýkoli.
U dat nízce kvalitních je často zdrojů nejistoty tak mnoho, že je již
prakticky nelze jako důkaz použít. Správný postup v tomto případě
neznamená snažit se i vysoce nekvalitní data interpretovat, ale
naopak: nekvalitní data rozpoznat a označit je jako
nehodnotitelná (nepoužitelná).

179
Kvalita v laboratoři
Proč laboratoře zavádějí systémy řízení kvality?
Řízení kvality ve forenzně genetických laboratořích sleduje několik
základních cílů:
prevence chyb ze známých zdrojů a prevence snížení kvality
(např. prevence záměny vzorků, prevence poškození vzorků
nevhodnou teplotou atd.)
odhalování nových, neznámých zdrojů chybovosti a
neznámých zdrojů snížení kvality (např. zjištění negativního
vlivu určité látky na průběh analýz apod.)
systémový přístup a standardizace analýz (se stejnými
vzorky je nakládáno stejně)
optimalizace laboratorních procesů z časového i
ekonomického hlediska (snaha dosáhnout rychlého zpracování
vzorků, snaha maximalizovat finanční efektivitu analýzy – tj.
množství informace získané ze vzorku za určitou jednotkovou
částku)
průběžné zkvalitňování analýz s ohledem na vývoj v oboru.
Co jsou to certifikace a akreditace laboratoří? Jaký je
mezi nimi rozdíl?
Certifikací rozumíme dobrozdání (certifikát) certifikačního
orgánu, že daný výrobek, postup nebo služba je ve shodě
s předepsanou normou nebo jiným normativním dokumentem.
V souvislosti s forenzně genetickou laboratorní praxí se nejčastěji
setkáme s certifikací o shodě s normou ISO 9001:2008 (týká se
systému managementu kvality).

180
Akreditací rozumíme dobrozdání akreditačního orgánu, že buď
a) daný certifikační orgán je po stránce odborné i jiné (nezávislost,
nestrannost atd.) způsobilý k certifikacím, nebo že b) daný subjekt
je odborně i jinak způsobilý k provádění určité činnosti.
V souvislosti s forenzně genetickou laboratorní praxí se nejčastěji
setkáme s akreditací laboratoří dle normy ISO 17025:2005 (týká
se všeobecných požadavků na způsobilost zkušebních a
kalibračních laboratoří, pro akreditaci forenzních laboratoří bylo
vydáno doporučení ILAC G19), případně okrajově s akreditací dle
normy ISO 15189:1998 (týká se zvláštních požadavků na kvalitu a
odbornou způsobilost zdravotnických laboratoří).
V souvislosti s ohledáním místa činu se můžeme setkat s akreditací
dle normy ISO 17020:1998 (týká se inspekce a inspektorů obecně,
pro akreditaci ohledání místa činu bylo vydáno zvláštní doporučení
EA–5/03).
Souhrnně lze konstatovat, že mezi certifikací forenzní laboratoře
dle ISO 9001:2008 a akreditací forenzní laboratoře dle ISO
17025:2005 je zásadní rozdíl: certifikace dle ISO 9001:2008
garantuje, že laboratoř má zaveden a uplatňuje systém řízení
kvality, ovšem již nijak negarantuje odbornou způsobilost dané
laboratoře k provádění příslušných zkoušek; naproti tomu
akreditace dle ISO 17025:2005 garantuje jak zavedení a
uplatňování systému managementu kvality, tak odbornou
způsobilost laboratoře k provádění příslušných zkoušek.
Znamená to tedy, že forenzně genetická laboratoř,
která není akreditována dle ISO 17025:2005 , není
odborně způsobilá a produkuje „špatná“ dat a?
Nikoliv. I laboratoře neakreditované mohou být velmi kvalitní, mít
vlastní systémy řízení kvality a kontroly správnosti dat a

181
produkovat vysoce validní data; tato jejich schopnost však není
garantována žádnou akreditační autoritou a je pouze na uvážení
„zákazníka“ (v tomto případě např. orgánu činného v trestním
řízení), jakou důvěryhodnost datům přikládá. V řadě oborů se již
akreditace laboratoře stala dokonce právně zakotvenou nezbytnou
podmínkou výkonu dané činnosti. Naproti tomu sama akreditace
není absolutní zárukou validity dat, pokud nejsou laboratoří
všechna pravidla důsledně uplatňována a naplňována.
Jak laboratoř vybírá vhodné metody pro danou
analýzu? Jak pozná, že metoda je v jejích podmínkách
funkční?
Při výběru metod pro analýzu se laboratoř řídí vědeckými poznatky
ve svém oboru; relevantní metody jsou vždy popsány v odborné
literatuře jako výstup vědeckých studií a zpravidla jsou
mnohonásobně ověřeny dalšími nezávislými studiemi jiných
odborných pracovišť. Řada metod je dnes dostupná komerční
cestou v podobě tzv. analyzačních kitů; firmy produkující tyto
kity garantují jejich funkčnost při dodržení stanoveného pracovního
postupu – kity jsou validované. Validací obecně rozumíme
„potvrzení prostřednictvím poskytnutí objektivních důkazů, že
požadavky na specifické zamýšlené použití nebo na specifickou
aplikaci byly splněny“. I validovaný kit však musí být verifikován
(tj. přezkoušen) přímo laboratoří, čímž je zaručeno, že funguje
správně i v podmínkách tohoto konkrétního pracoviště (tzn. na
jeho přístrojích atd.).
Jak laboratoř průběžně ověřuje, že produkuje
relevantní data?
Jednou z nejspolehlivějších metod ověření kvality a správnosti dat
produkovaných laboratoří je účast v tzv. okružních
(proficienčních) testech. Ty spočívají v tom, že zadavatel testů

182
připraví testovací vzorky, rozešle je testovaným laboratořím, ty je
analyzují a odešlou výsledky analýzy zpět zadavateli, který posoudí
správnost výsledků. Úspěšné laboratoře získají certifikát; testy se
opakují v určitých intervalech, kupříkladu jednou ročně. Tento
systém mezilaboratorní kontroly kvality může být organizován jak
na mezinárodní, tak i na národní úrovni.
Vypovídací hodnota dat
Jak funguje inferenční logika hodnocení dat?
Inference neboli usuzování, odvozování jedné skutečnosti z druhé,
je ve forenzní analýze založena na jednoduchém principu: některé
hypotézy jsou při daném výsledku pravděpodobnější a jiné
méně pravděpodobné; platí to tehdy, pokud jsou předpoklady
hypotézy a výsledky analýzy jevy závislé. Pro porozumění si
uveďme nejprve jednoduchý příklad zcela mimo forenzní obor:
Na konzultaci k panu Petrovi má přijít žena, říkejme jí Marie, o níž
Petr ví pouze to, že je jí právě 30 let. Klade si v duchu otázku, zda je
svobodná či nikoli. V tuto chvíli mu vodítkem může být pouze nějaká
obecná, předběžná (neboli apriorní) pravděpodobnost: dejme tomu,
že celorepubliková statistika říká, že v ročníku třicetiletých je dosud
svobodných zhruba 25 % žen. Z toho vyplývá, že i Marie bude
svobodná s pravděpodobností 25 %. Poté však kolega Petrovi během
řeči sdělí, že pokud je mu známo, vystudovala Marie práva. S novou
informací se původní (apriorní) pravděpodobnost změní: v ročníku
třicetiletých vysokoškolaček je totiž podle statistiky dosud svobodných
plných 40 %. O několik minut později zaklepe Marie na Petrovu
kancelář, a zatímco ji Petr vítá, všimne si, že na levém prsteníčku má
Marie zlatý prsten bez kamenu: pravděpodobnost, že je svobodná, v tu
chvíli dramaticky klesne.

183
Jak vidíme v našem příkladu, inferenční logika je založena na
úpravě našich očekávání na základě získání nových dat
(informací).
Stejným způsobem funguje hodnocení síly důkazu ve forenzní
oblasti – opět si uveďme příklad:
Manželé Novákovi ohlásili na policii, že jejich soused, pan Svoboda,
pravděpodobně zneužívá svou nezletilou dceru Lucii. Je známo, že cca
50 % takovýchto podezření se posléze potvrdí, zatímco ve zbylých 50
% případů se obvinění ukáže jako účelové nebo nesprávné. Prvotní
pravděpodobnost zneužívání Lucie (tj. ještě předtím, než kdokoli
cokoli začne zjišťovat), je tedy cca 50 %. Při následném pohovoru
s policejním psychologem si tento všimne, že Lucie si kouše nehty. Ze
studií přitom vyplývá, že kousání nehtů lze zaznamenat u cca 80 %
zneužívaných dětí a pouze u 10 % dětí nezneužívaných. Jinak řečeno:
kousání nehtů je 8x pravděpodobnější u zneužívaného dítěte než
u nezneužívaného. Na základě zjištění, že Lucie si kouše nehty,
stoupne i pravděpodobnost, že je zneužívána.
Jak funguje inferenční logika ve forenzní genetice?
Inferenční logika se ve forenzní genetice vztahuje především ke
třem typům hypotéz, a to:
hypotéz o shodném/rozdílném původci dvou vzorků
hypotéz o přítomnosti/nepřítomnosti biologického
materiálu určité osoby ve smíšené stopě
hypotéz o existenci/neexistenci určitého biologického
příbuzenského vztahu mezi dvěma osobami
V prvém případě nás zajímá především, jak významným důkazem
o stejném původci je shoda genotypů (genetických profilů). Ve
druhém případě nás zajímá, jak významně svědčí výsledek analýzy

184
smíšené stopy pro přítomnost biologického materiálu konkrétní
osoby. Ve třetím případě nás zajímá, jak silně pro hypotézu či proti
hypotéze o určitém příbuzenském vztahu svědčí zjištěné genotypy
(genetické profily) obou osob.
Jak konkrétně při určování vypovídací hodnoty
forenzně genetického důkazu znalec postupuje?
Vlastní interpretace dat je problematika nesmírně rozsáhlá a je
záměrem realizačního týmu Československé společnosti pro
forenzní genetiku vydat k tomuto tématu samostatnou podrobnou a
názornou publikaci o rozsahu minimálně takovém, jako má tento
breviář. Pro jeho potřeby se proto omezme na některá základní
konstatování:
znalec zvažuje výsledky analýzy za platnosti definovaných
alternativních hypotéz a s ohledem na specifika případu,
přičemž užívá tzv. bayesiánský přístup, založený na inferenční
logice
vychází z četnosti jednotlivých znaků v populaci a ze
způsobu, jakým jsou děděny
zohledňuje míru mutací, tj. změn v DNA, které mohou
zasáhnout i testované lokusy
hodnotí nejistotu měření neboli spolehlivost dat
bere v potaz velikost a strukturu populace, počet možných
pachatelů, míru příbuznosti osob v populaci atd.
Co by mělo být zásadním výstupem forenzně
genetické analýzy?
Výstupem práce znalce je tzv. věrohodnostní poměr (LR) (angl.
Likelihood Ratio) pro každou dvojici alternativních hypotéz; tento
věrohodnostní poměr může být kombinován s věrohodnostním
poměrem vzešlým jiných zkoumání (např. daktyloskopie) ke

185
stejné dvojici hypotéz nebo s dalšími – i nevědeckými – důkazy,
např. očitým svědectvím. Vynásobením věrohodnostního poměru a
apriorního podílu šancí získáváme aposteriorní podíl šancí
(převeditelný na aposteriorní pravděpodobnost).
Nutno říci, že praxe v tomto směru ještě stále zaostává za teorií:
věrohodnostní poměr není dosud v rámci našeho soudnictví příliš
vyžadován (soud obvykle volí jiný, mnohem intuitivnější přístup
k hodnocení důkazů jako celku), a není tudíž ani bezvýhradně
poskytován. Forenzně genetické posudky často obsahují jiné
statistické parametry – u identifikací to je pravděpodobnost náhodné
shody (RMP), u paternitních analýz například pravděpodobnost
otcovství (P) či síla vyloučení (PE). Tyto hodnoty nejsou zásadně
nesprávné, pro budoucnost je však žádoucí, aby se systém
hodnocení forenzních důkazů všech typů sjednotil a kodifikoval.
Blíže o tom bude pojednáno v samostatné publikaci o interpretaci
forenzních dat.
Zdroje chyb ve forenzně genetické expertíze
Je možné, aby při forenzně genetickém zkoumání došlo
k chybě?
Samozřejmě. Chybovost je přirozenou vlastností veškerých
procesů, ať už prováděných člověkem, strojem nebo přírodou, a
nevyhýbá se ani forenzně genetickému zkoumání (přestože bychom
rádi byli neomylní, nejsme). Pro věc je podstatné umět poznat a
popsat příčiny a mechanismy vzniku chyb a jejich důsledky, neboť
jedině tak je můžeme eliminovat či alespoň snížit pravděpodobnost,
že k nim dojde.

186
Kde v procesu forenzně genetické analýzy se chyby
objevují?
Prakticky kdekoli, ze statistik forenzních i jiných – například
klinických – laboratoří však vyplývá, že chyby mnohem častěji
vznikají mimo vlastní vědecké zkoumání a z jiných příčin než je
neodbornost. Spadají sem chyby jako nesprávné označení vzorku,
špatné přečtení označení vzorku (např. pro nečitelnost písma),
umístění vzorku na nesprávné místo, záměna dvou podobných stop
atd.
Jaké jsou zdroje chyb při zkoumání?
Četné příčiny chyb v laboratoři můžeme alespoň zhruba rozdělit
takto:
člověk vedený přímým úmyslem zkreslit či zcela změnit
výsledek zkoumání (např. osoba k tomu finančně motivovaná
pachatelem)
člověk vedený nepřímým úmyslem, kterému nejde
primárně o změnu výsledku, ale ví, že svým jednáním může
takovou změnu způsobit, a je s tím srozuměn (např. osoba,
která manipuluje s daty, aby zakryla svou chybu v pracovním
postupu)
člověk vědomě nedbalý, který ví, že svým jednáním může
způsobit chybu, ale doufá, že se tak nestane (dříve
všeobecně známo jako hřích „opovážlivé spoléhání na
milosrdenství Boží“; například osoba, která vědomě porušuje
pracovní postup, aby si ulehčila práci, přičemž tuší, že to může
způsobit chybu, ale doufá, že nezpůsobí)
člověk nevědomě nedbalý, který neví, že může způsobit
chybu, ale vědět by to měl (např. osoba manipulující
s přístrojem bez řádného proškolení)

187
běžný lidský omyl (přestože to mnozí neradi slyší, je
chybovost bez znaků nedbalosti integrální vlastností lidského
mozku – např. přepsání se v čísle, napipetování vzorku do
nesprávné zkumavky, překouknutí atd.)
technická chyba z očekávatelných příčin (počítačový virus,
závada na přístroji, nekvalitní chemikálie pro analýzu atd.)
technická chyba z neočekávatelných příčin (neodhalený
negativně působící fyzikální, chemický či biologický faktor –
v minulosti byly takto dodatečně „odhaleny“ např. pudrová
tvářenka laborantky či pylová zrna jasanu ve dvoře objektu,
které oboje velmi účinně blokovaly PCR).
Jaké se chybám uvedených typů bránit?
Pro jednotlivé zdroje chyb pochopitelně existují výrazně odlišné
mechanismy prevence:
v přímém či nepřímém úmyslu zkreslit či zcela změnit
výsledek zkoumání lze pracovníku laboratoře zabránit
poměrně obtížně, zejména je–li to osoba důvěrně znalá
prostředí a všech v něm fungujících procesů. Primárním
opatřením je samozřejmě prověření osob předtím, než se
pracovníky stanou; znesnadnit úmyslnou manipulaci lze
primárně výběrem pracovníků s morálním kreditem,
zavedením kontrolních bodů, jako jsou kontrolovaný pohyb
osob (např. čipové karty, přístup jen do určitých prostor),
kontrolovaný pohyb vzorku (uzamykatelné chladničky,
mrazáky apod.), kontrolovaný přístup do počítačů, řídících
jednotek přístrojů, databází (logování pomocí hesla, otisku
prstu apod.). I tak ovšem může zručný „insider“ najít slabá
místa systému.
v přímém či nepřímém úmyslu zkreslit či zcela změnit
výsledek zkoumání lze osobě zvenčí zabránit obdobnými

188
mechanismy jako v předchozím případě; účinnost opatření
bude u cizí osoby zcela jistě mnohem větší (osoba se nemůže
v laboratoři pohybovat „nenápadně“, není tak dobře
obeznámena s chodem laboratoře atd). Obecně je tendence
zcela minimalizovat přístup cizích osob do laboratorních
prostor (i proto, že tyto osoby mohou být zdrojem
kontaminující DNA).
nedbalostní jednání (ať už vědomé, či nevědomé) lze
nejlépe eliminovat zavedením přesných pravidel práce
v laboratoři a průběžnou kontrolou jejich dodržování
jednotlivými pracovníky; dobrým pomocníkem v tomto směru
může být software monitorující procesy v laboratoři, který umí
indikovat problematický faktor mnohem dříve, než by si jej
všiml člověk (např. identifikuje provázanost mezi častějšími
špatnými výsledky a určitým konkrétním pracovníkem).
je někdy snaha eliminovat běžný lidský omyl hrozbou trestu;
to je ovšem opatření zcela neúčinné, ba co víc –
kontraproduktivní. Člověk nemůže přestat přirozeně chybovat
jen proto, že mu hrozí trest; naopak – často hrozba trestu
způsobí, že osoby, která svůj omyl zjistí, se snaží na něj
neupozorňovat, případně jej i zastřít. Jedinými účinnými
opatřeními proti běžným chybám jsou zavedení kontrolních
mechanismů (např. data odečítají nezávisle na sobě dva experti
a musejí se shodnout, při pipetování jedna osoba kontroluje
druhou atd.) a především automatizace procesů, která vyloučí
lidský faktor ze hry (např. automatická izolace a amplifikace,
kdy vše provádí robot a vzorky jsou v procesu značeny a
identifikovány pomocí čárového či QR kódu)
technické chyby jsou do značné míry eliminovatelné
zavedením managementu kvality (zmíněné ISO17025), jehož
součástí jsou standardní operační postupy, které zahrnují i
pravidelné údržby, kalibrace a kontroly funkčnosti přístrojů,

189
kontroly kvality používaných chemikálií a laboratorního
materiálu atd.
Zásadním mechanismem, který nám pomáhá zvyšovat efektivitu
preventivních opatření, je zjištění příčiny a mechanismu každé
chyby, kterou v laboratoři odhalíme. Jedině tak můžeme
identifikovat a průběžně odstraňovat slabá místa celého procesu. I
tak ovšem zůstává pravděpodobnost chyby nenulová, a to i v té
nejspolehlivější laboratoři. Pokud takové důvodné podezření
existuje, je samozřejmě namístě – pokud je to možné – zpracování
revizního posudku v jiné laboratoři.

190
Uchovávání zdrojů genetických informací
Obrázek 57 – Možné formy uchovávání zdrojů genetických informací.
Banky (archivy, depozity) biologického materiálu
Co je banka biologického materiálu?
Základní možností, jak uchovat zdroj forenzně genetických
informací, je deponovat samotnou stopu či její část (např. setřený
biologický materiál). Bankou tedy rozumíme takovýto depozit;
jednotlivé stopy (biologické materiály) jsou evidovány pod
identifikačním kódem a k němu jsou vázána data – údaje o typu
stopy či materiálu, původu atd.
Jaké výhody a nevýhody má banka biologického
materiálu?
Provozní banka biologického materiálu funguje v každé forenzně
genetické laboratoři. Materiál je v ní uchováván po dobu provádění

191
expertíz a může být případně opakovaně testován či odebrán pro
novou izolaci DNA. Materiál v původním stavu navíc obsahuje i další
informace kromě vlastní DNA – lze z něj např. určit krevní skupinu,
mikroskopicky jej zkoumat a podobně. Nevýhodou uchovávání stop
a biologických materiálů však je, že ne vždy je při dlouhodobém
skladování možné zcela zabránit postupné degradaci
biologického materiálu ve stopě. Z provozního hlediska pak
zjevnou nevýhodu představují značné nároky na prostory
depozitáře.
Jak se biologický materiál deponuje?
Způsob depozice závisí na charakteru biologického materiálu.
Materiály, které jsou v suchém stavu nebo které je možné do něj
převést, se zpravidla uchovávají při pokojové teplotě v suchu a
temnu v prodyšných (např. papírových) obalech. Materiály jako je
tekutá krev lze krátkodobě skladovat v chladničce při 4°C, trvale
uchovány však tímto způsobem být nemohou. Materiály ze své
podstaty vlhké (např. svalová tkáň) se uchovávají hluboce zmražené
při teplotách kolem –80°C. Pravidla pro deponování biologického
materiálu jsou samozřejmě významně podrobnější, zde se
omezujeme jen na hlavní principy.
Banky izolátů DNA
Co je banka izolátů DNA?
Jak bylo popsáno v části věnované technologii forenzně genetické
analýzy, je z každého biologického materiálu nejprve izolována
DNA. Většina metod izolace, které jsou v rámci forenzní praxe
užívány, izoluje totálně, tj. izolát obsahuje veškerou DNA ze vzorku
(samozřejmě s určitými ztrátami danými účinností příslušné
metody; některé izolační metody také nepřevedou do izolátu velmi
krátké fragmenty DNA). Lze tedy říci, že z izolátu lze provádět

192
analýzu libovolných DNA lokusů. Stejně jako banka biologického
materiálu, i banka izolátů má evidenci vzorků založenou na
identifikačním kódu, k němuž jsou navázána data o izolátu (z jaké je
stopy, kdy a jakou metodou byl vytvořen atd.)
Jaké výhody a nevýhody má banka izolátů DNA?
Provozní banka izolátů funguje – stejně jako banka biologických
materiálů – v každé forenzně genetické laboratoři. Izoláty jsou v ní
uchovávány po dobu provádění expertíz a mohou být opakovaně
použity pro testování (kolik opakování lze provést, to závisí na
celkovém množství izolátu). Pokud laboratoř uchovává trvale
materiály, z nichž byla expertíza prováděna, pak se nejčastěji
jedná právě o izoláty DNA. Jejich skladování je mnohem snazší a
prostorově podstatně méně náročné než uchovávání biologického
materiálu jako takového.
Jak se izoláty DNA deponují?
V praxi jsou užívány dva hlavní způsoby depozice izolátů DNA:
zamražení (doporučovány teploty pod –50°C, ale prakticky lze i
kolem –20°C) nebo deponování v suchém stavu na FTA kartě.
Vzhledem k historicky relativně krátké době, po kterou je DNA
analýza prováděna, nelze přesně říci, jak dlouho zůstává
v deponovaném izolátu analyzovatelná DNA, rozhodně se však
jedná řádově o roky či spíše desítky let.
DNA databáze
Co jsou DNA databáze?
DNA databáze jsou organizované soubory DNA dat, tedy informací
vzniklých genotypováním (analýzou) určitých markerů v DNA.
V současnosti jsou provozovány již výhradně v elektronické podobě.

193
Setkáme se s nimi ve všech oblastech, v nichž se uplatňuje genetická
analýza – v klinické diagnostice, výzkumu i forenzní oblasti.
Jakou podobu mají forenzní (kriminalistické) DNA
databáze?
Ve forenzních DNA databázích jsou uchovávány výsledky
forenzních analýz jednotlivých vzorků v elektronické podobě –
mohou to být „klasické“ identifikační genetické profily STR lokusů,
výsledky Y–STR a Y–SNP genotypování, výsledky sekvenace HVR
mtDNA, ale i další markery užívané ve forenzním identifikačním
testování (vždy závisí na legislativě příslušného státu).
Data o jakých vzorcích jsou ve forenzních DNA
databázích vedeny?
Primárním úkolem forenzních DNA databází je objasňování
trestné činnosti; proto největší podíl v těchto databázích tvoří
jednak profily z biologických stop nalezených na místě
trestných činů, jednak profily osob – pachatelů trestné činnosti.
Dále se v databázích vyskytují profily osob neznámé totožnosti,
profily pohřešovaných či jejich příbuzných, profily obětí a též tzv.
eliminační profily – například profily pracovníků laboratoře.
Každý stát, provozující národní DNA databázi, má svá pravidla.
K čemu forenzní (kriminalistické) DNA databáze
slouží?
Strukturování dat ve forenzních DNA databázích a softwarová
nadstavba, která je zpravidla nedílnou součástí databáze, umožňují:
uchovávat forenzní DNA data
provádět srovnání DNA profilů jednotlivých vzorků
získávat z vložených dat statistické analýzy – populační data

194
Jaká porovnávání umožňuje DNA databáze?
Databázový software je obvykle postaven tak, aby umožňoval velmi
komplexně nastavovat parametry prohledávání. To umožňuje
přizpůsobit „odpověď“, kterou nám databáze dá, našim
požadavkům.
Máme například úplný genetický profil osoby, který jsme stanovili
z analyzačních dat vysoké technické kvality; chceme zjistit, zda se
v databázi nenachází profil téže osoby. V takovém případě nastavíme
parametry tak, aby prohledávání jako potenciálně shodný profil
označilo pouze ten, který se liší maximálně v jedné alele jednoho
markeru.
Naproti tomu pokud máme profil z dat nižší technické kvality, kde je
nejistota ohledně přesného stanovení genotypů větší, lze nastavit vyšší
toleranci odchylek – např. maximálně 6 alel.
Při vyhledávání možných biologických dětí genotypovaných osob
nastavujeme parametry tak, aby prohledávání označilo jako
potenciálně vyhovující takové profily, které se s naším shodují
v minimálně jedné alele v každém lokusu. Opět jiné bude nastavení
prohledávání při posuzování smíšených profilů.
Z výše uvedeného vyplývá jedna zásadní skutečnost: výsledek
každého porovnávání je pouze návrh, že označené profily by
mohly být potenciálně shodné či příbuzné. V případě, že se
nejedná o úplnou shodu ve všech alelách všech srovnávaných
lokusů, je namístě porovnat následně analyzační data obou vzorků a
teprve poté provádět další hodnocení a interpretaci včetně evaluace
míry shody.

195
Na jakém software stojí česká kriminalistická DNA
databáze?
Česká kriminalistická DNA databáze vznikla v roce 2002, kdy byl
České republice poskytnut databázový software CODIS, vyvinutý a
provozovaný americkým FBI. Na bázi tohoto software je
provozována řada národních databází v Evropě i ve světě.
Lze srovnávat profily i mimo Národní DNA databázi ,
tedy v zahraničí?
Možnosti srovnávání závisí jinak na technických, jednak na
legislativních podmínkách.
Po technické stránce lze srovnávat profily tehdy, pokud jsou
k testování užívány stejné markery (lokusy). Blíže se o tomto
tématu zmiňuje část věnovaná STR lokusům v kapitole Principy
identifikace.
Po legislativní stránce závisí možnost porovnání na konkrétních
podmínkách daného případu, typu požadavku, právních předpisech
partnerského státu atd.
Kromě individuálních požadavků na srovnávání jsou v současnosti
intenzivně budovány systémy mezinárodní výměny dat; na
celosvětové úrovni je vůdčím projektem Interpol DNA Gateway. Na
evropské úrovni je systém výměny genetických dat budován na
základě Prümské úmluvy.

196
„Non–human“ forenzní analýza živočišné,
rostlinné a mikrobiální DNA
Forenzní analýza živočišné DNA
Jaké forenzní analýzy lze u živočichů provádět?
Z biologického pohledu je člověk de facto pouze jedním z živočichů.
Proto zjevně nepřekvapí, že potenciál genetické analýzy je u
živočichů prakticky týž jako u člověka. Živočichy lze tedy stejně
jako osoby individuálně identifikovat a provádět u nich analýzu
biologické příbuznosti.
Kromě toho jsou v rámci živočišné forenzní genetiky prováděny též
specifické analýzy, jejichž cílem je určit živočišný druh, ze kterého
pochází daný biologický materiál.
Které živočišné druhy jsou nejčastěji zkoumány?
Identifikační a příbuzenské analýzy jsou prováděny zejména
u domestikovaných druhů. Jednak jsou to ty, které souhrnně
označujeme jako „domácí miláčky“ – tj. především kočka a pes, a
dále hospodářská zvířata, jako je skot, koně, ovce, prasata atd.
Účelem je zkoumání živočišného biologického materiálu v rámci
kriminálních případů, napadení člověka zvířetem, ale i např. pro
potřeby chovatelství.
Druhové identifikace jsou prováděny naopak zejména u živočichů
volně žijících; přitom dominují analýzy vzácných a chráněných
druhů v rámci aktivit na jejich ochranu (např. boj s ilegálním
obchodem s těmito druhy, s pytláctvím v národních parcích).

197
Jak je prováděna identifikační analýza u koček a psů?
U koček i psů jsou k identifikaci používány vybrané specificky psí či
kočičí tetranukleotidové krátké tandemové repetice (STR).
Metodika analýzy je tudíž prakticky identická s metodikou užívanou
v identifikačních analýzách člověka. DNA je dle charakteru případu
izolována ze srovnávacích vzorků (tekutá krev, bukální stěr) nebo
z biologických stop (chlupy, krev, sliny). Výraznější rozdíl zde
nacházíme pouze v interpretaci dat: některá plemena jsou vnitřně
mnohonásobně příbuzná oproti jiným, což je třeba vždy zohlednit
při výpočtu statistických parametrů míry identifikace.
U koček i psů lze též zkoumat patrilineárně či matrilineárně děděné
lokusy, jako jsou hypervariabilní oblasti mtDNA či Y–STR a Y–SNP
markery.
Jak je prováděna identifikační analýza
u hospodářských zvířat?
U hospodářských zvířat jsou k testování rovněž využívány nejčastěji
krátké tandemové repetice (STR), často se však jedná o repetice
s kratším, dinukleotidovým motivem.
Jak je prováděna identifikace živočišných druhů?
Druhová identifikace je založena na zkoumání takových lokusů
DNA, jejichž sekvence jsou identické nebo velmi podobné uvnitř
druhu (tj. u všech příslušníků daného druhu) a zároveň významněji
odlišné mezi jednotlivými druhy. Optimální v tomto směru se jeví
některé specifické lokusy v mitochondriální DNA: cytochrom B
gen, 12S rRNA gen a COI gen.
Analýza všech těchto lokusů je založena na sekvenaci a následném
porovnávání získané sekvence s databází sekvencí jednotlivých
druhů.

198
Hovězí steak může být např. zkoumán, pokud někdo
prodává/servíruje maso chráněných živočichů a vydává je za hovězí
dobytek nebo pokud je potřeba potvrdit, že steak je opravdu
připraven z býků plemene wagyu. Pomocí DNA analýzy nejde ovšem
potvrdit, že býk wagyu byl opravdu napájen saké...
Forenzní analýza rostlinné DNA
Jaké forenzní analýzy lze u rostlin provádět?
Forenzní analýza rostlinné DNA se zaměřuje na dvě základní
oblasti: prvou je identifikace rostlinného jedince v rámci
kriminalistických zkoumání, kdy smyslem je zpravidla určit
konkrétní místo původu nějakého rostlinného materiálu.
Druhou oblastí zkoumání je identifikace rostlinných drog,
především marihuany.
Jaké metody zkoumání se ve forenzní analýze
rostlinné DNA užívají?
Identifikační analýza rostlin je opět založena nejčastěji na
genotypizaci vysoce variabilních krátkých tandemových repetic
(STR). Významným faktorem, který je u rostlin třeba brát v potaz, je
to, že se jedná o modulární organismy, jak již bylo zmíněno v části
věnované individualitě a identifikaci. Jednoduše řečeno: tentýž
genetický jedinec může existovat jako mnoho jedinců fyzických.
Zatímco nový genetický jedinec vznikne pohlavním rozmnožováním
(tvorbou semen), noví fyzičtí jedinci mohou vznikat i nepohlavním
rozmnožováním (dělením cibulí, oddenků, řízkováním atd.).
Identifikace analýzou DNA se vždy týká jedince genetického.
I pro rostlinnou říši jsou užívány metody identifikace druhů na
základě sekvenace genů lokalizovaných v mitochondriální DNA;

199
v některých případech však neposkytují tak spolehlivé výsledky
jako u živočichů.
Forenzní analýza mikrobiální DNA
V jakých forenzních aplikacích je užívána analýza
mikrobiální DNA?
Analýza mikrobiální DNA je oborem ležícím na hranici forenzní
genetiky a forenzní mikrobiologie. Mikroorganismy jsou velmi
různorodá skupina organismů, vyskytující se v nejrůznějších
prostředích, proto i škála situací, kdy jsou mikroorganismy
předmětem forenzního zkoumání, je velmi pestrá.
Pomocí identifikace bakteriálních a houbových druhů lze
porovnávat půdní vzorky, a to nejlépe v kombinaci s fyzikálními
zkoumáními struktury, barvy atd. Každá půda je totiž osídlena
charakteristickým společenstvem mikroorganismů, tzv.
mikrobiomem, který je sice v čase ne zcela stálý, přesto však může
být k okamžitému porovnávání použit.
Identifikace konkrétních kmenů virulentních organismů, jako
jsou bakterie a viry, může být použita též k vysledování šíření
nákazy, typicky například u osob, které vědí, že jsou přenašeči
závažné infekční choroby (např. HIV, hepatitidy atd.), přesto však
nebrání nakažení dalších osob nebo je dokonce nakazí záměrně.
Určité mikroorganismy a jejich společenstva – tzv. mikroflóra – jsou
též charakteristické pro určitá tělní prostředí a tělní tekutiny.
Toho může být využito při identifikaci těchto materiálů – například
identifikace vaginálního sekretu je založena na stanovení
přítomnosti některých druhů laktobacilů.

200
Jaké metody zkoumání se ve forenzní analýze
mikrobiální DNA užívají?
Mikrobiální, a zejména bakteriální genom se výrazně odlišují od
genomu mnohobuněčných organismů, jakými jsou rostliny,
živočichové i člověk. Lokusy, které jsou u nich studovány, jsou proto
dosti odlišné a k jejich genotypizaci se používá celá řada
molekulárně genetických metod, s nimiž se v „klasické“ forenzní
genetice nesetkáme. Výčet a popis těchto metod už jde nicméně nad
rámec této publikace.

201
Doslov
Forenzně genetická analýza je pilířem kriminalistické práce při
vyšetřování závažných trestních činů, ale podílí se i na vyšetřování
méně závažných přestupků proti trestnímu i občanskému právu.
Z laického pohledu televizního diváka ovlivněného sledováním
seriálů typu CSI se jedná o bezchybnou a nezpochybnitelnou
metodu usvědčování zločinců. Z pohledu jiného genetického laika,
bojovníka za občanské soukromí, se jedná o potenciální hrozbu
úniku zneužitelných osobních údajů z kriminalistické DNA databáze.
Oba pohledy jsou pokřivené neznalostí. Pro pochopení výhod a
nevýhod použití DNA profilování a databázování kriminálníků je
nutné nejprve pochopit biologii genetických markerů, technologii
sběru stop, amplifikace DNA a detekce amplikonů a zásady
inferenční logiky.
V anglosaských zemích byla forenzní DNA analýza ve svých
počátcích podrobena důkladné kritice a široké diskusi, což umožnilo
obecné pochopení a přijetí jejich zákonitostí, postupů a omezení
všemi profesionálními účastníky soudního řízení a velké části laické
veřejnosti. Naproti tomu v české společnosti diskuse o DNA probíhá
kampaňovitě, vedena dílčími komerčními zájmy, a je ovlivněna
neznalostí základů forenzní genetiky. Je to zčásti proto, že v české
literatuře dosud chyběla kniha, která by uvedla do problematiky
soudce, státní zástupce, vyšetřovatele, obhájce, budoucí forenzně
genetické znalce nebo zainteresované laiky.
Autorce se podařilo čtivou formou otázek a odpovědí, doplněnou o
šťavnaté příklady z praxe, nastínit všechny důležité pojmy a
procesy, týkající se forenzní DNA analýzy předtím, než dojde k
završení analýzy – k vyhodnocení síly důkazů pomocí
věrohodnostního poměru. S obezřetností jsou přeskočeny všechny
vzorce a právní floskule, které by mohly čtenáře odradit.

202
Kniha může sloužit jako odrazové prkno pro všechny právníky, kteří
se chystají skočit do bazénu forenzně genetických analýz, i pro laiky,
kteří se cítí ohroženi existencí kriminalistické DNA databáze.
Ilustrativní obrázky svou přehledností a výpovědní hodnotou
nemají konkurenci na trhu českých genetických učebnic a
monografií.
Čtenáři, vychutnej si toto počtení!
doc. Mgr. Jiří Drábek, Ph.D.,
předseda Československé společnosti
pro forenzní genetiku
v Olomouci, červen 2012
O autorce
Mgr. Halina Šimková vystudovala odbornou biologii se specializací
na genetiku člověka a antropologii na Přírodovědecké fakultě
Univerzity Karlovy v Praze. Už v době studia působila dva roky jako
civilní stážistka na Kriminalistickém ústavu Praha Policie ČR, od
roku 2002 zde pracuje jako DNA expertka na oddělení genetických
expertíz. V roce 2005 se jako DNA specialistka účastnila mise
Interpolu při identifikacích obětí tsunami v Thajsku. Po ukončení
studií v r. 2002 spoluzaložila a přednáší předmět Forenzní genetika
na PřF UK. Velkou prioritou pro ni vždy byla a je didaktika oboru –
kromě odborných přednášek na univerzitě se věnuje přípravě
budoucích expertů, specializačnímu dalšímu vzdělávání stávajících
znalců a expertů, odbornému vzdělávání kriminalistů, přednáší pro
soudce a státní zástupce, ale nesmírně ráda také přibližuje obor
naprostým laikům při přednáškách pro veřejnost. Je zakládající
členkou Československé společnosti pro forenzní genetiku, od roku
2007 je její místopředsedkyní.

203
Doporučená literatura a informační zdroje
V českém jazyce pohříchu existuje jen velmi málo aktuální literatury
k tématu forenzně genetické analýzy (což byl ostatně jeden
z hlavních důvodů, proč vznikl tento breviář). Odborné informace
k oboru existují (jak už tomu u přírodních věd bývá) především
v jazyce anglickém. Zde odkazuji zejména na novější publikace, jako
například:
John M. Butler: Fundamentals of Forensic DNA Typing
John M. Butler: Advanced Topics in Forensic DNA Typing: Methodology
A. Jamie Cuticchia: Genetics: A Handbook for Lawyers
J. Thomas McClintock: Forensic DNA Analysis: A Laboratory Manual
Richard Li: Forensic Biology: Identification and DNA Analysis of
Biological Evidence
a další.
Z odborných periodik se tématu forenzní genetiky věnuje
především žurnál Forensic Science International: Genetics, dále
Journal of Forensic Sciences, Science & Justice a další.
Mezinárodní odbornou společností, která sdružuje odborníky
věnující se forenzní genetice, je ISFG – International Society for
Forensic Genetics (www.isfg.org). Na české národní úrovni je
takovouto odbornou společností ČSSFG – Československá
společnost pro forenzní genetiku (www.cssfg.org).
Existuje nepřeberné množství internetových zdrojů (ať už
otevřených, či uzavřených); zde však velmi důrazně varuji, že
mnoho z těchto zdrojů přináší nepřesné, zavádějící či zcela
nepravdivé informace, přičemž pro laika je velmi obtížné kvalitu
posoudit.

204
Rejstřík
1
12S rRNA gen ........................................ 197
A
adenin .......................................................... 50
AIMs ........................................................... 125
akreditace ............................................... 179
akreditační orgán ................................ 180
alela ....................................................... 59, 62
mikrovariantní . viz mikrovarianta
shodná původem ........................... 118
shodná stavem ............................... 118
alelově–specifická sonda ................. 161
AluQuant Human DNA Quantitation
System ................................................ 145
AMC ............................... viz amniocentéza
amelogenin ...................................122, 160
amniocentéza ........................................... 28
amplifikace DNA ........................146, 152
analyzátor genetický ...............156, 157
Ancestry Informative Markers ....... viz
AIMs
archeogenetika ........................................ 14
autozómy...................... viz chromozómy
nepohlavní
B
banka
biologického materiálu .............. 190
izolátů DNA ...................................... 191
barvivo
fluorescenční ................................... 142
biodegradace ................................. 40, 166
biologická stopa ...................................... 18
autolýza................................................. 41
degradace ............................................. 32
důkazní potenciál ............................. 46
důkazní potenciál nízký, nulový48
kontaminace ....................................... 34
latentní .................................................. 31
mechanismus vzniku ...................... 38
původce................................................. 25
uchovávání .......................................... 37
určení doby vzniku .......................... 42
vyhledávání ......................................... 30
biologický materiál ................................ 18
druhový původ .................................. 23
odběr ...................................................... 33
primární přenos ................................ 44
průkaz lidského původu ............... 23
sekundární přenos .......................... 44
sekundární přenos nepravý ........ 46
sekundární přenos vektorem ..... 45
tkáňový původ ................................... 20
určení množství ................................ 39
za nehty ................................................. 39
zdroje ..................................................... 37
bovinní sérový albumin ................... 131
BSA ........... viz bovinní sérový albumin
bukální stěr ............................................... 26
buňky
pohlavní ..................... viz gametocyty
C
Cambridge Reference Sequence ... 114
CE viz elektroforéza DNA kapilární

205
certifikace ............................................... 179
CODIS STR Loci ..................................... 104
COI gen ..................................................... 197
CVS................................. viz choriocentéza
cytochrom B gen .................................. 197
cytozin .........................................................50
cytozinový pás ...................................... 163
Č
červené krvinky ............. viz erytrocyty
činidlo
chelatační .......................................... 134
D
ddNTP .................. viz dideoxynukleotid
dědičnost
mendelovská ......................................62
mitochondriální DNA .....................68
na nepohlavních chromozómech
.............................................................62
na pohlavních chromozómech ...65
degradace DNA ................... 32, 166, 171
stupeň ................................................. 167
degradovaná DNA ............................... 173
dělení redukční...................... viz meióza
deoxyribonukleová kyselina . viz DNA
deoxyribóza ..............................................50
detekční činidlo
FastBlue ................................................30
Luminol .................................................30
dideoxyribonukleotid ........................ 162
diferenciální lýza ......................... viz lýza
diferenciální
Differex ..................................................... 138
D–klička ................................................... 112
DNA ...............................................................50
báze ........................................................ 50
degradovaná ......................... 166, 168
dvouřetězcová .................................. 51
extragenová........................................ 54
jaderná .................................................. 57
jednořetězcová ................................. 50
mikrobiální ............................ 196, 199
mitochondriální ............................... 57
rostlinná .................................. 196, 198
struktura .............................................. 50
templátová ............................. 148, 150
živočišná ............................................196
DNA databáze ........................................192
obsah ...................................................193
porovnávání .......................... 194, 195
software .............................................195
DNA fingerprinting ............................... 15
DNA polymeráza ..................................147
DNA profil .............. viz genetický profil
DNA profil osoby. viz genetický profil
osoby
drop–in
alelický ................................................172
drop–out
alelický ................................................171
lokusový .............................................171
druh
rostlinný - identifikace ................198
živočišný ............................................196
živočišný - identifikace ...............197
důkaz
vypovídací hodnota ......................184
dvojčata
dizygotní viz dvojčata dvojvaječná
dvojvaječná ................................. 75, 77
fraternální ....................... viz dvojčata
dvojvaječná
identická .......................... viz dvojčata
jednovaječná
jednovaječná ...................................... 73

206
monozygotní .................. viz dvojčata
jednovaječná
E
elektroforetická vana ........................ 155
elektroforetogram ....................159, 162
elektroforéza DNA .............................. 153
gelová ........................................154, 155
kapilární ............................................ 156
endonukleáza ........................................ 134
epigenetické rozdíly ............................. 59
erytrocyty .................................................. 58
ethidiumbromid................................... 142
etnicita ......................... viz původ etnický
etnikum ....................................................... 95
European Standard Set ..................... 104
extrakce DNA ............... viz izolace DNA
F
Fantom z Heilbronnu ........................... 35
fenotyp ........................................................ 58
fenotypový znak viz znak fenotypový
fluorimetr................................................ 142
forenzní světelné zdroje ..................... 30
fragmentace DNA ................................ 167
FTA card .............................. viz FTA papír
FTA papír ................................................ 135
G
GE
ciz elektroforéza DNA gelová .. 154
gel
agarózový ......................................... 155
polyakrylamidový ......................... 155
gen ................................................................. 56
geneta.................................... 70, 72, 73, 77
genetická data
spolehlivost ...................................... 175
technická kvalita............................ 176
uchovávání ....................................... 190
vypovídací hodnota ........... 175, 176
genetický kód
univerzální .......................................... 54
genetický profil ............................... 70, 71
genetický profil konsenzuální ...... 173,
178
genetický profil osoby.......................... 70
genetika....................................................... 13
forenzní ................................................. 13
identifikační ........................................ 14
kriminalistická................................... 13
lékařská................................................. 14
genom .......................................................... 57
geny ............................................................... 54
gonozómy . viz chromozómy pohlavní
guanin .......................................................... 50
H
haplotyp
Y-chromozómu ............. 79, 108, 125
Y-STR ................................................... 105
Y-STR minimální ............................ 105
Y-STR rozšířený ............................. 105
heminy ...................................................... 131
hemizygot ....................................... 67, 105
hemoglobin ............................................ 131
heteropaternální superfekundace viz
superfekundace heteropaternální
heteroplazmie ....................................... 115
heterozygot .................................... 63, 105
heterozygozita ......................................... 65
homozygot ...................................... 63, 105

207
homozygozita ...........................................65
huminové kyseliny.............................. 131
HV1 ............................................................. 113
HV2 ............................................................. 113
HV3 ............................................................. 113
HVR mtDNA................................. 109, 124
alely ...................................................... 114
analýza ............................................... 161
Ch
charakterizace DNA ........................... 128
Chelex–100 ............................................. 134
chimérismus .............................................76
posttransplantační .................. 73, 79
tetragametický .......................... 73, 77
chloroplasty ..............................................58
choriocentéza ...........................................28
chromatinové vlákno ............................53
chromozóm
X 54, 105, 120
Y 54, 65, 66, 105, 120, 121
Y - určení biogeografického
původu ......................................... 125
chromozómy .............................................53
nepohlavní ...........................................54
pohlavní ........................................ 54, 65
chybovost ................................................ 185
zdroje chyb ....................................... 186
I
identifikace
individuální .........................................25
identifikace ................................................69
identifikace
individuální .........................................69
identifikace ................................................70
identifikace
principy ................................................ 99
identifikace
individuální ......................................108
identifikace
individuální ......................................116
identifikace
rostlinných drog ............................198
Identifiler .................................................160
Identifiler Plus ......................................160
Identifiler® Plus Kit ...........................129
inbreeding ................................................. 92
incest ........................................................... 92
individuální identifikace .................... viz
identifikace individuální
individuum
fyzické .................................. viz rameta
genetické ............................. viz geneta
inference ..................................................182
pravděpodobnostních jevů .......176
inferenční logika hodnocení dat ...182
inhibitory .............................. 41, 130, 146
Interpol DNA Gateway ......................195
Interpretace forenzně genetických
dat .........................................................175
izolace DNA ................................. 127, 129
fenol-chloroformová ....................133
chelexová ...........................................134
na FTA papíru..................................135
vazbou na pevnou fázi ................136
izolát
DNA ......................................................130
genový (genetický) .......................130
J
jádro
buněčné ................................................ 57
jedinec

208
fyzický .................................. viz rameta
genetický .............................. viz geneta
Jeffreys, Alec John .................................. 15
K
karyotyp ..................................................... 53
kolonka
silikátová ........................................... 136
koncentrace DNA
hmotnostní ....................................... 140
konsangvinita........................................... 92
kontrolní oblast... viz oblast kontrolní
krevní destičky ............. viz trombocyty
křížení
příbuzenské ............... viz inbreeding
kvalita v laboratoři ............................. 179
kvantifikace DNA .......................128, 140
luminometrická ............................. 145
nespecifická ..................................... 141
specifická ................................141, 143
spektrofotometrická .................... 141
L
laserová mikrodisekce ............. viz LCM
LCM ..................................................137, 139
LCN ............................................................. 170
Likelihood Ratio..... viz věrohodnostní
poměr
lokus ............................................................. 56
mitochondriální ............................. 121
X-chromozomální ......................... 120
Y-chromozomální ......................... 121
lokusy
multialelické ....... viz lokusy vysoce
polymorfní
nepolymorfní ..................................... 60
nízce a středně polymorfní .......... 60
vysoce polymorfní ........ 60, 99, 117
Low–Copy Number ..................... viz LCN
Low–Template DNA ........... viz LT-DNA
LR viz věrohodnostní poměr
LT–DNA .................................................... 170
luminometr ............................................ 145
lýza
buněk .................................................. 132
diferenciální ..................................... 137
lyzát
buněčný .................................. 132, 134
M
marker ......................................................... 57
matroklinita .............................................. 68
meióza ......................................................... 63
melanin .................................................... 131
mikrodisekce
laserová ..................................... viz LCM
mikrochimérismus
fetální ............................................. 73, 79
maternální ................................... 73, 79
mikrokoncentrátor ............................. 132
mikrosatelity ................................. viz STR
mikrovarianta ....................................... 103
mikrovariantní alela ............................ viz
mikrovarianta
minHt . viz haplotyp Y-STR minimální
MiniFiler .................................................. 160
minisatelity ................................ viz VNTR
minisekvenování ................................. 164
miniSTRs ................................................. 168
mitochondriální DNA ...... 57, 112, 124
sekvenace .......................................... 161
určení biogeografického původu
.......................................................... 124
mitom ........................................................... 57

209
mitotyp ........................................................68
mobilita
elektroforetická ............................. 153
motiv
sekvenční .............................................99
mtDNA
viz mitochondriální DNA ..............57
mutace .........................................................61
gametické .............................................61
indukované..........................................61
somatické .............................................61
spontánní .............................................61
N
národ .................................................... 95, 96
narušení struktury DNA ................... 167
násilný kontakt
důkaz ......................................................40
nDNA ............................... viz jaderná DNA
nehty ..................................................... 27, 39
nerovnováha píků ............................... 171
non–human analýza ........................... 196
norma
ISO 17020
1998 ............................................... 180
ISO 17025
2005 ............................................... 180
nucleus ....................... viz jádro buněčné
nukleom ......................................................57
nukleotid ....................................................50
nukleotidová sekvence .. viz sekvence
nukleotidů
nukleotidy
volné .................................................... 147
O
oblast
hypervariabilní .... viz HVR mtDNA,
viz HVR mtDNA
kontrolní ............................................112
pseudoautozomální ........................ 65
okružní testy ..........................................181
OligoGreen ..............................................142
oocyty .......................................... viz vajíčka
oplození ...................................................... 70
organismy
modulární............................................ 72
unitární ................................................. 72
ori H ...........................................................112
otcovství ...................................................119
P
partikule
magnetosilikové .............................136
PCR ........................................ 131, 140, 146
asymetrická ......................................152
fáze denaturační ............................148
fáze hybridizační (anelační) .....149
fáze plató ...........................................150
fáze prodlužovací (elongační,
extenzní) ......................................149
kvantitativní.......................... viz qPCR
monoplex ...........................................151
multiplex ............................................151
negativní vlivy .................................150
nested ..................................................170
reakční směs ....................................147
sekvenační ........................................162
symetrická ........................................152
PicoGreen ................................................142
pigment
indigový .............................................131

210
vlasový a kožní ............................... 131
pík
koktavý ......................... 151, 171, 172
plastidom ................................................... 58
plastidy,....................................................... 58
plodová voda ............................................ 28
plodový materiál .................................... 28
pohlaví osoby
určení ........................................122, 160
polymeráza
prokluz .....................................151, 172
s korektorskou funkcí ................. 151
polymerázová řetězová reakce ...... viz
PCR
polymorfismus ........................................ 59
bodový ........................................ viz SNP
jednonukleotidový ............... viz SNP
poly–T řetězec ...................................... 166
pomůcky
DNA-free ............................................... 34
ochranné .............................................. 35
sterilní ................................................... 34
PowerPlex 16 ........................................ 160
PowerPlex 18D Kit ............................. 129
PowerPlex 21 ........................................ 160
PowerPlex Y ........................................... 160
PowerPlex Y23 ..................................... 160
pravidlo komplementarityviz princip
komplementarity
predikce
biogeografického původu ............ 95
fenotypu ............................................... 97
Prep–n–GoTM Buffer ........................... 130
primer .............................................147, 152
prodloužení ...................................... 165
princip komplementarity ................... 52
proband ...................................................... 87
proficienční testy ................................ 181
proteosyntéza .......................................... 55
Prümská úmluva.................................. 195
pryskyřice
iontoměničová ................................ 134
příbuznost
biologická ..................................... 14, 80
pokrevní ............... viz konsangvinita
principy určení ............................... 117
stupeň .................................................... 80
půdní vzorky.......................................... 199
purifikace DNA ..................................... 131
původ
biogeografický ........................ 95, 124
etnický ................................................... 95
Q
qPCR ............................................... 146, 167
QuantiBlot Human DNA Quantitation
Kit ......................................................... 145
R
rameta................................... 70, 72, 73, 77
rasa ................................................................ 95
rCRS .............. viz Cambridge Reference
Sequence
reparace DNA ........................................ 168
repetice ..........viz sekvence repetitivní
rozptýlená ............................................ 99
tandemová ........................................... 99
tandemové dlouhé ............ viz VNTR
tandemové krátké ................. viz STR
ribonukleová kyselina.............. viz RNA
RNA ............................................................... 55
rozptýlená repetice ........... viz repetice
rozptýlená

211
Ř
řídící oblast............ viz oblast kontrolní
S
sekvenace ................................................ 197
sekvenátor .............................................. 157
sekvence
nukleotidů ...........................................51
repetitivní ............................................99
silika........................................................... 136
slot blot hybridizace .......................... 144
SNaPshot ................................................. 164
SNP .................................................. 108, 109
alely ...................................................... 110
analýza ............................................... 164
autozomální ..................................... 110
identifikace ....................................... 111
určení biogeografického původu
.......................................................... 125
X-chromozomální .......................... 110
Y-chromozomální .......................... 111
spermie ............................................... 58, 70
srovnávací vzorky
mrtvá osoba ........................................29
pohřešovaná osoba .........................29
srovnávací vzorky
typy .........................................................26
SRY ............................................................. 123
SSO ................................................... 161, 163
SSR ...................................................... viz STR
starodávná DNA ......................................14
stochastické efekty ............................. 171
STR .................................................. 100, 197
alely ...................................................... 102
analýza ............................................... 168
autozomální ................ 105, 106, 117
genotyp .............................................. 105
klasifikace .........................................102
lokusy doporučené sety .............104
X-chromozomální ............... 105, 107
Y-chromozomální ............... 105, 108
základní motiv ..................... 100, 102
STR lokus
analýza ................................................158
stutter peak .................... viz pík koktavý
superfekundace
heteropaternální .............................. 76
SwabSolution Kit .................................130
syntéza bílkoviny ... viz proteosyntéza
T
tandemová repetice ........... viz repetice
tandemová
terminátor .................................... 162, 165
termocyklér ................ viz thermocycler
thermocycler .........................................147
transkripce ............................................... 55
transplantace ........................................... 79
kostní dřeně ....................................... 80
trifosfát ....................................................... 50
trombocyty ............................................... 58
tymin............................................................ 50
U
U.S. Core Loci .........................................104
účinnost
separační ...........................................129
V
vajíčka ......................................................... 58
vajíčko ......................................................... 70

212
validace .................................................... 181
verifikace ................................................. 181
věrohodnostní poměr ....................... 184
vícerčata
jednovaječná .................. viz dvojčata
jednovaječná
vícevaječná ...................... viz dvojčata
dvojvaječná
VNTR ................................................. 99, 100
výtěžnost
izolace ................................................. 130
Y
Y–filer........................................................ 160
Z
zkoncentrování izolátu DNA .......... 132
zkouška
orientační ............................................. 21
specifická.............................................. 21
znak
fenotypový ................................ 96, 126
ztráta alely ............................ viz drop-out
zygota ................................................... 63, 70





























