Embed Size (px)
Citation preview

Brca2-Null Embryonic Survival is Prolongedon the BALB/c Genetic Background
L. Michelle Bennett,1* Kimberly A. McAllister,1* Pamela E. Blackshear,2 Jason Malphurs,1
Gina Goulding,3 N. Keith Collins,1 Toni Ward,1 Donna O. Bunch,3 Edward M. Eddy,3
Barbara J. Davis,2 and Roger W. Wiseman1
1Laboratory of Molecular Carcinogenesis, National Institutes of Health, National Institute of Environmental Health Sciences,Research Triangle Park, North Carolina2Laboratory of Experimental Pathology, National Institutes of Health, National Institute of Environmental Health Sciences,Research Triangle Park, North Carolina3Laboratory of Reproductive Development and Toxicology, National Institutes of Health,National Institute of Environmental Health Sciences, Research Triangle Park, North Carolina
Women who inherit mutations in the BRCA2 cancer susceptibility gene have an 85% chance of developing breastcancer. The function of the BRCA2 gene remains elusive, but there is evidence to support its role in transcriptionaltransactivation, tumor suppression, and the maintenance of genomic integrity. Individuals with identical BRCA2mutations display a different distribution of cancers, suggesting that there are low-penetrance genes that can modifydisease outcome. We hypothesized that genetic background could influence embryonic survival of a Brca2 mutation inmice. Brca2-null embryos with a 129/SvEv genetic background (129B2ÿ/ÿ) died before embryonic day 8.5. Transfer ofthis Brca2 mutation onto the BALB/cJ genetic background (BALB/cB2ÿ/ÿ) extended survival to embryonic day 10.5.These results indicate that the BALB/c background harbors genetic modifiers that can prolong Brca2-null embryonicsurvival. The extended survival of BALB/cB2ÿ/ÿ embryos enabled us to ask whether transcriptional regulation of theBrca1 and Brca2 genes is interdependent. The interdependence of Brca1 and Brca2 was evaluated by studying Brca2gene expression in BALB/cB1ÿ/ÿ embryos and Brca1 gene expression in BALB/cB2ÿ/ÿ embryos. Nonisotopic in situhybridization demonstrated that Brca2 transcript levels were comparable in BALB/cB1ÿ/ÿ embryos and wild-typelittermates. Likewise, reverse transcriptase±polymerase chain reactions confirmed Brca1 mRNA expression inembryonic day 8.5 BALB/cB2ÿ/ÿ embryos that was comparable to Brca2-heterozygous littermates. Thus, the Brca1 andBrca2 transcripts are expressed independently of one another in Brca1- and Brca2-null embryos. Mol. Carcinog.28:174±183, 2000. ß 2000 Wiley-Liss, Inc.
Key words: Brca2, BALB/c, gene targeting, embryonic lethality
INTRODUCTION
Genetic inheritance represents a major risk factorfor breast-cancer development. Alterations in anumber of genes have been linked to hereditarybreast cancer including BRCA1, BRCA2, p53, Ataxiatelangiectasia mutated, PTEN, and Adenomatouspolyposis coli. Inherited mutations in the breastcancer susceptibility genes, BRCA1 and BRCA2, areimplicated in 40±50% of women with a familialhistory of breast cancer and have been reported toincrease a woman's lifetime breast-cancer risk to ashigh as 85% [1,2]. BRCA2 mutations have beenassociated with a wide spectrum of disease outcomesincluding male breast, ovarian, prostatic, colon,pancreatic, and stomach cancers [3]. Studies of theBRCA2 999del5 mutation in the Icelandic popula-tion suggest the presence of modi®er genes that canin¯uence tumor phenotype [4]. However, theinheritance of a BRCA2 mutation in combinationwith a germline mutation in the breast-cancer±susceptibility gene BRCA1 has not been associatedwith more severe phenotypes [5,6].
Although the functions of the BRCA1 and BRCA2genes have yet to be elucidated, there is evidencethat they play a role in responses to DNA damageand the maintenance of genomic stability [7±13].BRCA1 and BRCA2 interact directly in a complexwith RAD51, suggesting interdependence in at leastone DNA damage-repair pathway [13]. We andothers have demonstrated that Brca1 and Brca2expression patterns throughout embryonic and
MOLECULAR CARCINOGENESIS 28:174±183 (2000)
ß 2000 WILEY-LISS, INC.
L. Michelle Bennett and Kimberly A. McAllister contributedequally to the work presented in this report.
The current address of Pamela E. Blackshear is Schering-PloughResearch Institute, 144 Route 94, P.O. Box 32, Lafayette, NJ 07848-0032.
*Correspondence to: Laboratory of Molecular Carcinogenesis,NIH, National Institute of Environmental Health Sciences, ResearchTriangle Park, NC.
Received 21 December 1999; Revised 17 May 2000; Accepted 23May 2000
Abbreviations: E, embryonic day; 129, 129/SvEv; BALB/c, BALB/cJ;ES cell, embryonic stem cell; PCR, polymerase chain reaction; neo,neomycin resistance; RT, reverse transcription.

mammary-gland development and in adult tissuesare very similar, although not identical [14,15].These similar expression patterns may or may not bethe consequence of direct interactions betweenfactors that control the expression of these twogenes.
Most targeted genetic mutations described to datein the Brca1 and Brca2 loci cause embryoniclethality when present in the homozygous state[16]. The age of embryonic lethality in several Brca1-and Brca2-null embryos is dependent on whichportion of the gene is targeted for disruption [17].Targeted disruption of Brca1 in the 5 0 region of thegene causes embryonic lethality from embryonicday (E) 4.5 to E7.5 [18,19]. A Brca1 mutation causingan in-frame exon 11 deletion can extend survivalbeyond E10.5 [20]. Similar observations have beenmade with Brca2 mice that target the 5 0 end of exon11. Targeted disruption 5 0 of the well-conservedBRC repeats in exon 11 results in embryoniclethality when two mutant alleles are inherited[21±23]. However, retention of three or more of theBRC repeats, which bind Rad51, appears to besuf®cient for survival of a small subgroup ofhomozygous-null animals to at least several monthsof age and is in¯uenced by genetic background[24,25].
The targeted disruption of mouse genes has madeit possible to study the modi®cation of speci®c genemutations in a controlled environment [17]. Severalstudies have demonstrated that combining a Brca1or Brca2 alteration with p53 or p21 mutationsextends embryonic survival. Brca1 and p53 or Brca1and p21 double-null embryos have survived to E9.5[19,23], whereas Brca2/p53 double nullizygotes, ona mixed 129/SvEv (129) and C57BL/6 background,appear in most cases to be developmentally similarto or slightly more advanced than Brca2 mutantsalone [23]. It is unclear how much of the phenotypicrescue described by introducing p53 or p21 in thesestudies is attributable to these speci®c targetedalterations as opposed to other genetic in¯uencesintroduced by inbred strain backgrounds.
We introduced a single Brca2 gene mutation onthe 129/SvEv (129) and BALB/cJ (BALB/c) geneticbackgrounds. Although embryonic survival inBrca1- and Brca2-null mice can be in¯uenced byan additional tumor suppressor gene alteration, theeffects of genetic background alone have not beencarefully investigated. We developed Brca2-de®-cient mice on a 129 background and establishedBALB/c inbred mice congenic for this alteration.The 129B2ÿ/ÿ embryos express an embryonic lethalphenotype similar to previously reported Brca2-de®cient mice with mutations introduced 5 0 ofthe BRC repeats [21±23]. However, we demonstratethat the BALB/c inbred genetic background canprolong survival of homozygous-null embryos. Theearly embryonic death of Brca2-null embryos limits
their use for studying loss of Brca1 and Brca2function during neoplastic transformation. Theextension of embryonic survivability enabled usto assess the interdependence of Brca1 and Brca2by examining the expression pattern of Brca2in BALB/cB1ÿ/ÿ embryos and Brca1 in BALB/cB2ÿ/ÿ
embryos. We report that the BALB/c genetic back-ground could enhance embryonic survivability inBrca2-null embryos and that Brca1 and Brca2transcripts were expressed independently of oneanother.
MMAATTEERRIIAALLSS AANNDD MMEETTHHOODDSS
DDeevveellooppmmeenntt ooff BBrrccaa22--DDee®®cciieenntt MMiiccee
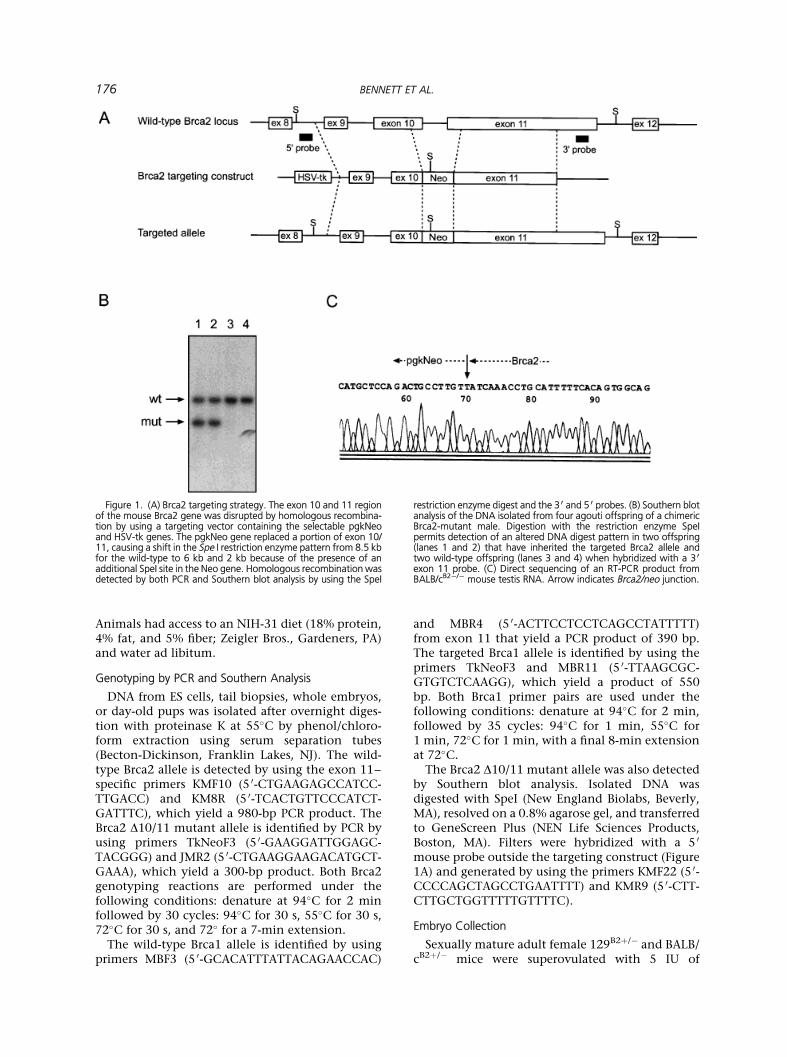
Brca2-de®cient mice were generated by disruptingexons 10 and 11 with a pgkNEO cassette (Figure 1A).The pgkNEO gene replaced a portion of exon 10,intron 10, and the 5 0 region of exon 11, extendingfrom nucleotide 1745 to 2033 (accession numberU89652) [26]. The targeting vector was linearizedwith SalI and electroporated (125 mFd, 0.36 kV;Bio-Rad gene pulser; Bio-Rad, Richmond, CA) into129/Ola-derived BK-4 embryonic stem (ES) cells[27]. After positive and negative selection withgeneticin (250 mg/mL; Gibco/BRL, Rockville, MD)and gancyclovir (2 mM; Roche, Hertfordshire, UK),three of 40 geneticin-resistant and gancyclovir-sensitive colonies were identi®ed as correctly tar-geted by polymerase chain reaction (PCR), Southernanalysis, and direct sequencing. One ES cell clone,2S2, was used to generate several chimeric males bywell-established techniques [28]. Initially, malechimeras were bred to wild-type C57BL/6 femalemice, and DNA isolated from agouti tail biopsies wasanalyzed by PCR to identify offspring that carry themutant Brca2 allele. The Brca2 mutation has beenestablished on the C57BL/6N (Charles River Labora-tory, Raleigh, NC) background by successive backcrossing. Chimeric male mice were also bred directlyto 129/SvEv (Taconic, Germantown, NY) andBALB/c (Jackson Laboratories, Bar Harbor, ME)females and back±crossed repeatedly to create inbredmouse strains congenic for the Brca2 mutation. TheBALB/cB2�/ÿmice used in this experiment were fromthe seventh back-cross generation (>99% contribu-tion from the BALB/c genetic background).
MMiiccee
BALB/cB1�/ÿ mutant mice were maintained byback crossing to wild-type BALB/c mice and havebeen described previously as Brca1D223±763 mice[20]. The neomycin-resistance (neo) insertion inthe BALB/cB1�/ÿ mutant mice results in an alter-natively spliced transcript that encodes an in-framedeleted Brca1 protein lacking amino acids 223±763from exon 11. DBA/2J mice were purchased fromJackson Laboratories. All mice were group housedin plastic cages with pressed wood-chip bedding.
BBRRCCAA22--NNUULLLL EEMMBBRRYYOONNIICC SSUURRVVIIVVAALL OONN TTHHEE BBAALLBB//CC BBAACCKKGGRROOUUNNDD 117755
Animals had access to an NIH-31 diet (18% protein,4% fat, and 5% ®ber; Zeigler Bros., Gardeners, PA)and water ad libitum.
GGeennoottyyppiinngg bbyy PPCCRR aanndd SSoouutthheerrnn AAnnaallyyssiiss
DNA from ES cells, tail biopsies, whole embryos,or day-old pups was isolated after overnight diges-tion with proteinase K at 55�C by phenol/chloro-form extraction using serum separation tubes(Becton-Dickinson, Franklin Lakes, NJ). The wild-type Brca2 allele is detected by using the exon 11±speci®c primers KMF10 (5 0-CTGAAGAGCCATCC-TTGACC) and KM8R (5 0-TCACTGTTCCCATCT-GATTTC), which yield a 980-bp PCR product. TheBrca2 D10/11 mutant allele is identi®ed by PCR byusing primers TkNeoF3 (5 0-GAAGGATTGGAGC-TACGGG) and JMR2 (5 0-CTGAAGGAAGACATGCT-GAAA), which yield a 300-bp product. Both Brca2genotyping reactions are performed under thefollowing conditions: denature at 94�C for 2 minfollowed by 30 cycles: 94�C for 30 s, 55�C for 30 s,72�C for 30 s, and 72� for a 7-min extension.
The wild-type Brca1 allele is identi®ed by usingprimers MBF3 (5 0-GCACATTTATTACAGAACCAC)
and MBR4 (5 0-ACTTCCTCCTCAGCCTATTTTT)from exon 11 that yield a PCR product of 390 bp.The targeted Brca1 allele is identi®ed by using theprimers TkNeoF3 and MBR11 (5 0-TTAAGCGC-GTGTCTCAAGG), which yield a product of 550bp. Both Brca1 primer pairs are used under thefollowing conditions: denature at 94�C for 2 min,followed by 35 cycles: 94�C for 1 min, 55�C for1 min, 72�C for 1 min, with a ®nal 8-min extensionat 72�C.
The Brca2 D10/11 mutant allele was also detectedby Southern blot analysis. Isolated DNA wasdigested with SpeI (New England Biolabs, Beverly,MA), resolved on a 0.8% agarose gel, and transferredto GeneScreen Plus (NEN Life Sciences Products,Boston, MA). Filters were hybridized with a 5 0
mouse probe outside the targeting construct (Figure1A) and generated by using the primers KMF22 (5 0-CCCCAGCTAGCCTGAATTTT) and KMR9 (5 0-CTT-CTTGCTGGTTTTTGTTTTC).
EEmmbbrryyoo CCoolllleeccttiioonn
Sexually mature adult female 129B2�/ÿ and BALB/cB2�/ÿ mice were superovulated with 5 IU of
FFiigguurree 11.. ((AA)) BBrrccaa22 ttaarrggeettiinngg ssttrraatteeggyy.. TThhee eexxoonn 1100 aanndd 1111 rreeggiioonnooff tthhee mmoouussee BBrrccaa22 ggeennee wwaass ddiissrruupptteedd bbyy hhoommoollooggoouuss rreeccoommbbiinnaa--ttiioonn bbyy uussiinngg aa ttaarrggeettiinngg vveeccttoorr ccoonnttaaiinniinngg tthhee sseelleeccttaabbllee ppggkkNNeeooaanndd HHSSVV--ttkk ggeenneess.. TThhee ppggkkNNeeoo ggeennee rreeppllaacceedd aa ppoorrttiioonn ooff eexxoonn 1100//1111,, ccaauussiinngg aa sshhiifftt iinn tthhee SSppee II rreessttrriiccttiioonn eennzzyymmee ppaatttteerrnn ffrroomm 88..55 kkbbffoorr tthhee wwiilldd--ttyyppee ttoo 66 kkbb aanndd 22 kkbb bbeeccaauussee ooff tthhee pprreesseennccee ooff aannaaddddiittiioonnaall SSppeeII ssiittee iinn tthhee NNeeoo ggeennee.. HHoommoollooggoouuss rreeccoommbbiinnaattiioonn wwaassddeetteecctteedd bbyy bbootthh PPCCRR aanndd SSoouutthheerrnn bblloott aannaallyyssiiss bbyy uussiinngg tthhee SSppeeII
rreessttrriiccttiioonn eennzzyymmee ddiiggeesstt aanndd tthhee 33 00 aanndd 55 00 pprroobbeess.. ((BB)) SSoouutthheerrnn bblloottaannaallyyssiiss ooff tthhee DDNNAA iissoollaatteedd ffrroomm ffoouurr aaggoouuttii ooffffsspprriinngg ooff aa cchhiimmeerriiccBBrrccaa22--mmuuttaanntt mmaallee.. DDiiggeessttiioonn wwiitthh tthhee rreessttrriiccttiioonn eennzzyymmee SSppeeIIppeerrmmiittss ddeetteeccttiioonn ooff aann aalltteerreedd DDNNAA ddiiggeesstt ppaatttteerrnn iinn ttwwoo ooffffsspprriinngg((llaanneess 11 aanndd 22)) tthhaatt hhaavvee iinnhheerriitteedd tthhee ttaarrggeetteedd BBrrccaa22 aalllleellee aannddttwwoo wwiilldd--ttyyppee ooffffsspprriinngg ((llaanneess 33 aanndd 44)) wwhheenn hhyybbrriiddiizzeedd wwiitthh aa 33 00eexxoonn 1111 pprroobbee.. ((CC)) DDiirreecctt sseeqquueenncciinngg ooff aann RRTT--PPCCRR pprroodduucctt ffrroommBBAALLBB//ccBB22ÿÿ//ÿÿ mmoouussee tteessttiiss RRNNAA.. AArrrrooww iinnddiiccaatteess BBrrccaa22//nneeoo jjuunnccttiioonn..
117766 BBEENNNNEETTTT EETT AALL..

pregnant mare serum gonadotropin (Sigma, St.Louis, MO) followed by 5 IU of human chorionicgonadotropin (Sigma) 48 h later and were thenmated with 129B2�/ÿ and BALB/cB2�/ÿ males,respectively. E8.5, E10.5, and E11.5 embryos ordeciduas were collected from timed-pregnantmice and ®xed in 10% neutral buffered formalinfor 2±24 h or frozen atÿ70�C for genotypic analysis.Suf®cient tissue was not always available for theextraction of DNA and RNA. Female BALB/cB1�/ÿ
mice were also superovulated and mated withBALB/cB1�/ÿ males to generate Brca1 embryos ofall genotypes. E10.5 embryos or deciduas weresubsequently collected from these timed-pregnantBALB/cB1�/ÿ females and ®xed as described earlier.
RReevveerrssee TTrraannssccrriippttiioonn ((RRTT))±±PPCCRR
RNA was extracted from mouse testes or wholeembryos with Tri-Reagent (Molecular ResearchCenter, Inc., Cincinnati, OH) as directed. Frozentissue samples were ®nely ground with mortar andpestle in liquid nitrogen. One milliliter of Tri-Reagent was added per 50±100 mg of tissue, thesamples were homogenized, and RNA was isolated.The cDNA was generated by RT from the mRNAusing Superscript II-RT (Boerhinger Mannheim,Indianapolis, IN). The Brca2 cDNA was ampli®edby an exon 10±speci®c forward primer KCF1 (5 0-AGGACAGCATTTTGAATCACT) and a neo-speci®creverse primer KCR1 (5 0-GGTGGATGTGGAATG-TGT). This RT-PCR product was gel isolated andsequenced directly with an internal reverse neoprimer (5 0-AGACGTGCTACTTCCATTTGT). Brca1cDNA from Brca2-intercross progeny was ampli®edby RT-PCR with the following primer pair thatspanned from exon 6 to exon 7 of Brca1: MBF29(5 0-GGGAAGCACAAGGTTTAGTCA) and MBR10(5 0-GGTGGCATTTCCAGGTTC).
IInn SSiittuu HHyybbrriiddiizzaattiioonn
Brca1 and Brca2 digoxigenin-labeled probes weregenerated as described by Blackshear et al. [14]except that the transcription reactions were per-formed with the Maxiscript T3/T7 kit (Ambion,Austin, TX). Paraf®n-embedded sections were hybri-dized and washed at 60�C. Whole-embryo in situhybridization was performed essentially asdescribed by Wilkinson [29]. After ®xation, theembryos were prehybridized for 1 h at 63�C andthen hybridized overnight to either a Brca2-anti-sense or a Brca2-sense digoxigenin-labeled probe.After hybridization, the embryos were takenthrough a series of stringent 63�C washes (50%formamide, 0.75 M sodium chloride, and 0.075 Msodium citrate). The embryos were then treated withribonuclease A to remove unbound single-strandedprobe. The embryos were incubated with 10% sheepserum for 60±90 min and then incubated overnightat 4�C with antidigoxigenin antibody conjugated to
alkaline phosphatase. The embryos were incubatedwith nitroblue tertrazolium/5-bromo-4-chromo-3-indolyl phosphate (Boehringer Mannheim) in thedark to develop the alkaline phosphatase histo-chemical reaction.
RREESSUULLTTSS
GGeennoottyyppee aanndd TTrraannssccrriipptt AAnnaallyyssiiss iinnBBrrccaa22--DDee®®cciieenntt MMiiccee
The genotypes of agouti offspring resulting fromcrosses between chimeric males and C57BL/6females were determined by Southern analysis(Figure 1B) and PCR (data not shown). RT-PCRanalysis was performed on BALB/cB2�/ÿ testis RNAbecause Brca2 RNA is abundantly expressed in thistissue [14,15,26]. A 550-bp fragment was generatedfrom the BALB/cB2�/ÿ samples with primers speci®cfor Brca2 exon 10 and the neo gene. Directsequencing of this PCR product indicated that theBrca2/neo fusion transcript was expressed (Figure1C). The putative Brca2 protein is truncated atcodon 569, with 75 additional amino acids con-tributed by the pgkNeo cassette. Repeated attemptsto visualize a truncated transcript by northern blotanalysis were unsuccessful, presumably due tounstable mRNA.
PPhheennoottyyppiicc EEvvaalluuaattiioonn ooff BBrrccaa22--DDee®®cciieennttaanndd BBrrccaa22--NNuullll MMiiccee
Mendelian inheritance of the Brca2 mutation wasstudied by intercrossing male and female Brca2heterozygous mice, with a mixed 129�C57BL/6background, to generate Brca2-null mice. Genotypicanalyses by Southern blotting and PCR ampli®ca-tion failed to identify a single Brca2-null animalfrom 142 intercross offspring examined at weaning(Table 1). Mice heterozygous for a germline Brca2mutation did not display any overt phenotypicabnormalities compared with their wild-type litter-mates at birth, weaning, or as adults. Female andmale Brca2-heterozygous mice were able to breednormally. Females carried their pups to term and didnot exhibit dif®culty with lactation. We did notobserve an increased incidence of spontaneoustumors in 129B2�/ÿ virgin females versus wild-typelittermates up to 2 yr of age.
GGeenneettiicc MMooddii®®ccaattiioonn ooff EEmmbbrryyoonniicc SSuurrvviivvaabbiilliittyy
Genetic background has been determined toin¯uence phenotype and survival of mutants inseveral gene knockout mouse models [30]. Brca1-null offspring on a mixed 129�DBA/2�C57BL/6genetic background survive up to E13.5 [20], whichis several days beyond that described for otherBrca1-null mice. This altered phenotype may be theconsequence of this speci®c Brca1 mutation thatyields an in-frame deletion of exon 11 (D223±763)or, alternatively, may be enhanced by the presence
BBRRCCAA22--NNUULLLL EEMMBBRRYYOONNIICC SSUURRVVIIVVAALL OONN TTHHEE BBAALLBB//CC BBAACCKKGGRROOUUNNDD 117777

of the DBA/2 genetic background. Thus, we eval-uated survival to birth of Brca2-null mice by usingseveral combinations of the DBA/2 with 129 and/orC57BL/6 genetic backgrounds. Seventy-one off-spring from the following crosses were genotypedat 1 d of age: (DBA/2�129 B2�/ÿ)F2, (DBA/2�C57BL/6B2�/ÿ)F2, and (DBA/2�C57BL/6B2�/ÿ)F1�129B2�/ÿ. As shown in Table 1, the incorporation ofthe DBA/2 genetic background did not prolong thesurvival of Brca2-null offspring to birth.
The embryonic lethal phenotype was studied inBrca2-null mice on the 129 genetic background.Intercrosses between male and female 129B2�/ÿ
mice generated 22 E8.5 embryos, but genotypicanalysis of the embryos was not completed becauseof the large number of reabsorbed fetuses observedat this time point. The 129B2�/ÿ intercrosses gener-ated 35 E10.5 embryos, 11 of which were dissectedout of the decidua or the yolk sacs and genotyped.Only heterozygotes and wild types were present at
E10.5. Thus, 129B2ÿ/ÿ embryos die and are largelyreabsorbed before E8.5, which is consistent withprevious reports of Brca2-de®cient mice on a mixedC57BL/6 and 129 genetic background [21±23].
In contrast, when the Brca2 mutation was placedon a BALB/c genetic background, the day ofembryonic lethality in the Brca2-null mice wasextended to approximately E10.5. Ten E8.5 embryoswere generated from BALB/cB2�/ÿ intercross mat-ings. Seven of 10 E8.5 embryos were genotyped, andthe remaining three were either partly or completelyreabsorbed. PCR analysis conclusively identi®edthree BALB/cB2ÿ/ÿ embryos and four BALB/cB2�/ÿ
embryos (Figure 2). Genotyping was performed onone sample that was partly reabsorbed, but the PCRproduct was probably due to maternal contamina-tion (Figure 2, lane 4). BALB/cB2ÿ/ÿ E8.5 embryoswere grossly smaller, developmentally delayed, andresembled E6±7 embryos (Figure 3). Twenty-six yolksacs and/or embryos were isolated from intercrosses
FFiigguurree 22.. GGeennoottyyppiicc aannaallyyssiiss ooff DDNNAA eexxttrraacctteedd ffrroomm eeiigghhtt iinntteerr--ccrroossss EE88..55 BBAALLBB//ccBB22 eemmbbrryyooss.. TThhee uuppppeerr ggeell ccoonnttaaiinnss PPCCRR pprroodduuccttssuussiinngg pprriimmeerrss tthhaatt aammpplliiffyy tthhee mmuuttaanntt aalllleellee.. TThhee lloowweerr ggeell ccoonnttaaiinnssPPCCRR pprroodduuccttss ggeenneerraatteedd wwiitthh wwiilldd--ttyyppee±±ssppeeccii®®cc pprriimmeerrss.. EEaacchh llaannee
rreepprreesseennttss aann iinnddiivviidduuaall eemmbbrryyoo.. NNoo ssaammpplleess wweerree llooaaddeedd iinn llaanneess 33,,66,, aanndd 88.. LLaannee 11:: DDNNAA MMWW mmaarrkkeerr.. LLaanneess 22,, 55 aanndd 77:: BBAALLBB//ccBB22ÿÿ//ÿÿ
eemmbbrryyoo.. LLaannee 44:: iinnccoonncclluussiivvee rreessuulltt.. LLaanneess 99±±1122:: BBAALLBB//ccBB22��//ÿÿ
eemmbbrryyooss.. LLaannee 1133:: ccoonnttrrooll aadduulltt BBAALLBB//ccBB22��//ÿÿ DDNNAA..
TTaabbllee 11.. GGeennoottyyppeess ooff BBrrccaa22--IInntteerrccrroossss PPrrooggeennyy oonn DDiiffffeerreenntt IInnbbrreedd GGeenneettiicc BBaacckkggrroouunnddss
Brca2 genotype
Strain background Time point �/� �/ÿ ÿ/ÿ(C57B2�/ÿ� 129B2�/ÿ) F2 3 wk 65 77 0(DBA/2�C57B2�/ÿ) F2 1 d 6 19 0(DBA/2� 129B2�/ÿ) F2 1 d 4 17 0(DBA/2�C57B2�/ÿ) F1� 129B2�/ÿ 1 d 13 12 0
117788 BBEENNNNEETTTT EETT AALL..

at E10.5. The yolk sacs and/or embryos weregenotyped by PCR and con®rmed that all morpho-logically normal embryos were of the heterozygousor wild-type genotypes. The one con®rmed BALB/cB2ÿ/ÿ embryo that survived until E10.5 was devel-opmentally delayed and smaller than its hetero-zygous littermates. The cranial neural tuberemained open, characteristic of an E8.5 embryo.However, the turning process was completed andthe second brachial arch and the heart wereprominent, indicating some development beyondthe E8.5 time point. Three additional embryosdisplayed similar developmental delays, but con-®rmation of genotype was hindered by maternaltissue contamination. No embryonic tissue wasfound in ®ve reabsorption sites. At the E11.5 timepoint, 10 morphologically normal and two devel-opmentally delayed yolk sacs were dissected. Therewere three completely reabsorbed sites that couldnot be dissected. The 10 morphologically normalembryos were genotyped as wild types or hetero-zygotes. The con®rmation of genotype for the two
developmentally delayed embryos was not possiblebecause of contaminating maternal tissue. Devel-opmentally, these presumed mutant embryos werecharacteristic of normal E8.5 embryos, similar tothat observed at the E10.5 time point.
The extension of Brca2-null embryo survival onthe BALB/c genetic background enabled us toaddress whether transcriptional regulation of theBrca1 and Brca2 genes was interdependent. Brca1expression was evaluated by RT-PCR analysis ofmRNA isolated from E8.5 BALB/cB2ÿ/ÿ embryos.These studies demonstrated that Brca1 expression inBALB/cB2ÿ/ÿ E8.5 embryos was comparable to age-matched wild-type BALB/c B2�/ÿ embryos (Figure 4).
In addition, we examined the timing of embryo-nic lethality and the expression of Brca2 in Brca1-null embryos on the BALB/c genetic background.Forty-two embryos were generated from BALB/cB1�/ÿ intercrosses, and 19 viable progeny wereexamined. Genotypic analysis showed two BALB/cB1ÿ/ÿ, seven BALB/cB1�/�, and 10 BALB/cB1�/ÿ
offspring. The BALB/cB1ÿ/ÿ embryos were grossly
FFiigguurree 33.. GGrroowwtthh iinnhhiibbiittiioonn ooff BBAALLBB//ccBB22ÿÿ//ÿÿ eemmbbrryyooss aatt EE88..55 aannddEE1100..55.. TThhee EE88..55 ggrroowwtthh--rreettaarrddeedd BBAALLBB//ccBB22ÿÿ//ÿÿ eemmbbrryyoo iiss ccoommppaarreeddwwiitthh aa nnoorrmmaall BBAALLBB//ccBB22��//ÿÿ lliitttteerrmmaattee.. TThhee BBrrccaa22 ((ÿÿ//ÿÿ)) eemmbbrryyoo iiss ssttiilllliinn aann eegggg--ccyylliinnddeerr ssttaaggee.. TThhee BBrrccaa22 ((��//ÿÿ)) eemmbbrryyoo hhaass aapppprrooxxiimmaatteellyyffoouurr ttoo ®®vvee ssoommiitteess aanndd wweellll--ddeevveellooppeedd eeccttooppllaacceennttaall ccoonnee aanndd yyoollkk
ssaacc.. TThhee EE1100..55 ggrroowwtthh--rreettaarrddeedd BBAALLBB//ccBB22ÿÿ//ÿÿ eemmbbrryyoo iiss ccoommppaarreeddwwiitthh aa mmoorrpphhoollooggiiccaallllyy nnoorrmmaall wwiilldd--ttyyppee lliitttteerrmmaattee.. TThhee BBrrccaa22 ((ÿÿ//ÿÿ))eemmbbrryyoo iiss ddeevveellooppmmeennttaallllyy ddeellaayyeedd,, wwiitthh aann ooppeenn ccrraanniiaall nneeuurraall ttuubbeecchhaarraacctteerriissttiicc ooff aa nnoorrmmaall EE88..55 eemmbbrryyoo..
BBRRCCAA22--NNUULLLL EEMMBBRRYYOONNIICC SSUURRVVIIVVAALL OONN TTHHEE BBAALLBB//CC BBAACCKKGGRROOUUNNDD 117799

smaller and developmentally delayed. The E10.5Brca1-null embryos had completed the turningprocess characteristic at this stage of development.However, the open cranial neural tube was char-acteristic of E8±8.5 embryos. These observations arecomparable to results reported by Gowen et al. [20]with Brca1-null mice on a mixed 129, DBA/2, andC57BL/6 genetic background. Thus, we did notobserve a dramatic alteration in the timing ofembryonic lethality in BALB/cB1ÿ/ÿ embryos.
Brca2 expression was examined in E10.5 BALB/cB1ÿ/ÿ embryos. In situ hybridization analysisdetected widespread Brca2 expression in Brca1-nullembryos that was qualitatively comparable to thelevels of Brca2 expression in age-matched BALB/cB1�/� embryos (Figure 5). The most intense locali-zation of Brca2 mRNA was in the neuroepithelialcells lining the neural tube and was similar betweenwild-type and null embryos (data not shown).Neuroepithelial cells comprised the most activeproliferating population, correlating with rapidbrain development, in E8±10 embryos.
DDIISSCCUUSSSSIIOONN
This study demonstrates that the BALB/c geneticbackground can modify survival of Brca2-nullmouse embryos. Previous reports have associatedembryonic survival with the location of mutationswithin the gene but have not shown an effect due tovariation of genetic background alone. The 129B2ÿ/ÿ
embryos were presumed to have died before E8.5because 129B2�/ÿ intercrosses had few viable pro-geny, as indicated by the large percentage ofreabsorbed fetuses. This observation is consistentwith other reports of Brca2 mutant mice in whichthe targeted alteration was 5 0 of the BRC repeatregion [21±23]. In contrast, matings between BALB/cB2�/ÿ mice generated BALB/cB2ÿ/ÿ embryos thatsurvived to E10.5. Thus, the BALB/c genetic back-ground extends survival of Brca2-null embryos thatinherit an alteration in exons 10 and 11 in Brca2.
Genetic background has been shown to in¯uencephenotype and survival of mutants in a variety ofgene knockout animal models [30]. For example,the BALB/c genetic background has a dramatic effecton the normal development of p130 knockout mice[31]. Likewise, the timing of embryonic lethality ofPTEN knockout embryos is modi®ed by the CD-1genetic background [32,33]. Epidermal growthfactor receptor knockout mice with a 129 inbredbackground die during mid-gestation but survive tobirth or longer with mixed inbred or outbredgenetic backgrounds [34,35]. Thus, embryoniclethality for a variety of gene defects can bemodi®ed by allelic differences contributed byvarious inbred strain backgrounds.
We have demonstrated that modi®ers in theBALB/c genetic background can prolong the survivalof Brca2-null embryos. Variation of disease onsetand severity has been reported for women with
FFiigguurree 44.. AAnnaallyyssiiss ooff BBrrccaa11 eexxpprreessssiioonn iinn tthhee EE88..55 BBAALLBB//ccBB22
eemmbbrryyooss ffrroomm FFiigguurree 33 bbyy RRTT--PPCCRR.. TThhee ggeennoottyyppeess aarree iinnddiiccaatteedd..TThhee uuppppeerr ggeell sshhoowwss BBrrccaa11 ccDDNNAA ((221177 bbpp)) aammppllii®®eedd wwiitthh aa pprriimmeerrppaaiirr tthhaatt ssppaannss eexxoonnss 66 aanndd 77.. GG33PPDDHH,, uusseedd aass aann iinntteerrnnaall ccoonnttrrooll,, iisssshhoowwnn oonn tthhee lloowweerr ggeell.. LLaanneess 11 aanndd 1122:: DDNNAA MMWW mmaarrkkeerrss.. LLaanneess
22±±44,, 66,, 88,, aanndd 99:: aammppllii®®ccaattiioonn ooff BBrrccaa11 ccDDNNAA.. LLaannee 1111:: ppoossiittiivveeccoonnttrrooll.. LLaanneess 55 aanndd 77:: nnoo BBrrccaa11 ccDDNNAA wwaass aammppllii®®eedd bbeeccaauussee ooff tthheemmiinniimmaall aammoouunntt ooff eemmbbrryyoo ttiissssuuee aavvaaiillaabbllee.. LLaannee 1100:: nneeggaattiivveeccoonnttrrooll..
118800 BBEENNNNEETTTT EETT AALL..

various mutations in BRCA1 and BRCA2 [4,36].Penetrance variations in different populations sug-gest that genetic modi®er loci or environmentalfactors must modulate the effect of BRCA1 orBRCA2 inactivation [1]. The inheritance of muta-tions in both BRCA1 and BRCA2 does not appear toaffect severity or latency of disease outcome inhumans [5,6], as might be expected if the genesdirectly interacted. However, modi®er genes arestrongly implicated by the various disease pheno-types that have been observed in individuals withidentical BRCA2 mutations [4]. Certainly, there maybe highly prevalent low-penetrance susceptibilitygenes that have a more profound effect on breast-cancer development in the general population thanBRCA1 or BRCA2 mutations do [37,38]. Such genesmay also modify breast-cancer risk in BRCA1 andBRCA2 mutation carriers, but the limitations ofhuman linkage analysis makes identifying thosegenes very challenging.
Radiation-induced DNA damage is repaired lessef®ciently in cells from BALB/c than in cells fromC57BL/6 mice. Chromosomal aberrations persist forup to 28 population doublings in irradiated BALB/cmammary epithelial cells, whereas similarly treatedC57BL/6 cells repair damage within six populationdoublings [39]. Perhaps this increased tolerance forDNA damage in BALB/c mice is related to the
prolonged embryonic survival of BALB/cB2ÿ/ÿ micecompared with other strains. In addition, a Brca2mutation on the BALB/c genetic background mayresult in more dramatic phenotypic outcomes in theadult mammary gland as a result of inef®cient DNAdamage repair and a predicted tissue speci®city.
This study also demonstrated that Brca1 mRNAwas expressed in Brca2-null embryos and Brca2mRNA was expressed in Brca1-null embryos. Expres-sion levels of the null embryos were qualitativelycomparable to those of wild-type and heterozygouslittermates, although small quantitative differencesin expression cannot be excluded. Thus, Brca1 andBrca2 transcripts are expressed independently ofeach other. If the regulation of either of these geneswere dependent on the other, dysregulated expres-sion would be expected in the gene-de®cientembryos. Our results support those of a report bySuzuki et al. [22] who used Brca1-null mice with atargeted disruption of exons 5 and 6 and Brca2-nullmice with disruption of exons 10 and 11 to studyBrca2 and Brca1 gene expression, respectively. Inthe mouse embryo, adequate proliferation to sustainnormal development appears to require the expres-sion of both Brca1 and Brca2. Mice that inheritmutations in both Brca1 and Brca2 genes do notdisplay more severe embryonic lethality, whichmight be expected if the effect of the losses were
FFiigguurree 55.. IInn ssiittuu hhyybbrriiddiizzaattiioonn aannaallyyssiiss ooff BBrrccaa22 eexxpprreessssiioonn iinn EE1100..55BBAALLBB//ccBB11��//�� aanndd BBAALLBB//ccBB11ÿÿ//ÿÿ eemmbbrryyooss:: BBAALLBB//ccBB11��//�� wwhhoollee eemmbbrryyoosswweerree hhyybbrriiddiizzeedd wwiitthh ccRRNNAA ddiiggooxxiiggeenniinn--llaabbeelleedd BBrrccaa22 aannttiisseennssee ((AA))
aanndd sseennssee ((BB)) pprroobbeess.. ((CC)) BBAALLBB//ccBB11ÿÿ//ÿÿ eemmbbrryyoo hhyybbrriiddiizzeedd wwiitthh ccRRNNAAddiiggooxxiiggeenniinn--llaabbeelleedd aannttiisseennssee pprroobbee.. BBaarr�� 11 mmmm..
BBRRCCAA22--NNUULLLL EEMMBBRRYYOONNIICC SSUURRVVIIVVAALL OONN TTHHEE BBAALLBB//CC BBAACCKKGGRROOUUNNDD 118811

additive [23]. Our investigation was limited tomRNA expression in embryos and did not addresspotential interactions between the Brca1 and Brca2proteins. Because these gene products form acomplex in vitro [13], it is possible that thefunctions of these proteins require a mutual inter-action that permits normal cellular proliferationsuch that dysregulation of either Brca1 or Brca2 cancontribute to neoplastic development.
We have described a substantial extension in thesurvival time of Brca2-null embryos on the BALB/cgenetic background. These results suggest the pre-sence of modi®er loci that could be mapped in thesemice. Future studies will evaluate the in¯uence ofthe BALB/c genetic background on mammary-glandmorphogenesis and neoplastic development inBrca2-de®cient mice. Identi®cation of such geneticmodi®ers in mice may permit the identi®cation ofhuman orthologues that could be evaluated for theirrole in breast-cancer susceptibility.
AACCKKNNOOWWLLEEDDGGMMEENNTTSS
We thank Yugi Mishina and Arti Patel for thecritical review of this article and for their helpfulsuggestions. We thank Beverly Koller for thegenerous gift of the Brca1-de®cient mice. The experttechnical assistance of Clyde Rogers, HeatherBrownlee, Charles Cochran, Astrid Haugen-Strano,and Sarah Hagevik with the development of theBrca2-de®cient mice is greatly appreciated. LMB andK.A.M. were funded during a part of this study byDepartment of Defense grants DAMD17-97-1-7027and DAMD1-98-1-8131, respectively. The workcontributed by PEB was in partial satisfaction ofthe requirements for the degree of Doctor ofPhilosophy at North Carolina State University.
RREEFFEERREENNCCEESS
1. Rahman N, Stratton M. The genetics of breast cancersusceptibility. Annu Rev Genet 1998;32:95±120.
2. Ford D, Easton D, Stratton M, et al. Genetic heterogeneityand penetrance analysis of the BRCA1 and BRCA2 genes inbreast cancer families. Am J Hum Genet 1998;62:676±689.
3. The Breast Cancer Linkage Consortium. Cancer risks inBRCA2 mutation carriers. J Natl Cancer Inst 1999;91:1310±1316.
4. Thorlacius, S Olafsdottir, G Tryggvadottir L, et al. A singleBRCA2 mutation in male and female breast cancer familiesfrom Iceland with varied cancer phenotypes. Nat Genet1996;13:117±119.
5. Ramus SJ, Friedman LS, Gayther SA, et al. A breast/ovariancancer patient with germline mutations in both BRCA1 andBRCA2. Nat Genet 1997;15:14±15.
6. Randall TC, Bell KA, Rebane BA, Rubin SC, Boyd J. Germlinemutations of the BRCA1 and BRCA2 genes in a breast andovarian cancer patient. Gynecol Oncol 1998;70:432±434.
7. Scully R, Chen J, Plug A, et al. Association of BRCA1 withRad51 in mitotic and meiotic cells. Cell 1997;88:265±275.
8. Patel KJ, Yu VPC, Lee HS, et al. Involvement of Brca2 in DNArepair. Mol Cell 1998;1:347±357.
9. Gowen LC, Avrutskaya AV, Latour AM, Koller BH, LeadonSA. BRCA1 required for transcription-coupled repair ofoxidative DNA damage. Science 1998;281:1009±1012.
10. Chen JJ, Silver D, Cantor S, Livingston DM, Scully R. BRCA1,BRCA2, and Rad51 operate in a common DNA damageresponse pathway. Cancer Res 1999;59:1752s±1756s.
11. Morimatsu M, Donoho G, Hasty P. Cells deleted for Brca2COOH terminus exhibit hypersensitivity to g-radiation andpremature senescence. Cancer Res 1998;58:3441±3447.
12. Zhong Q, Chen CF, Li S, et al. Association of BRCA1 with thehRad50±hMre11±p95 complex and the DNA damageresponse. Science 1999;285:747±750.
13. Chen J, Silver DP, Walpita D, et al. Stable interactionbetween the products of the BRCA1 and BRCA2 tumorsuppressor genes in mitotic and meiotic cells. Mol Cell1998;2:317±328.
14. Blackshear PE, Goldsworthy SM, Foley JF, et al. Brca1 andBrca2 expression patterns in mitotic and meiotic cells ofmice. Oncogene 1998;16:61±68.
15. Rajan JV, Marquis ST, Garder HP, Chodosh LA. Develop-mental expression of Brca2 colocalizes with Brca1 and isassociated with proliferation and differentiation in multipletissues. Dev Biol 1997;184:385±401.
16. Hakem R, de la Pompa JL, Mak TW. Developmental studiesof Brca1 and Brca2 knock-out mice. J Mammary Gland BiolNeoplasia 1998;3:431±445.
17. Bronson SK, Smithies O. Altering mice by homologousrecombination using embryonic stem cells. J Biol Chem1994;269:27155±27158.
18. Liu C-Y, Flesken-Nikitin A, Li S, Zeng Y, Lee W-H. Inactiva-tion of the mouse Brca1 gene leads to failure in themorphogenesis of the egg cylinder in early postimplantationdevelopment. Genes Dev 1996;10:1835±1843.
19. Hakem R, de la Pompa JL, Sirard C, et al. The tumorsuppressor gene Brca1 is required for embryonic cellularproliferation in the mouse. Cell 1996;85:1009±1023.
20. Gowen LC, Johnson BL, Latour AM, Sulik KK, Koller BH.Brca1 de®ciency results in early embryonic lethality char-acterized by neuroepithelial abnormalities. Nat Genet1996;12:191±194.
21. Sharan SK, Morimatsu M, Albrecht U, et al. Embryoniclethality and radiation hypersensitivity mediated by Rad51 inmice lacking Brca2. Nature 1997;386:804±810.
22. Suzuki A, de la Pompa JL, Hakem R, et al. Brca2 is requiredfor embryonic cellular proliferation in the mouse. Genes Dev1997;11:1242±1252.
23. Ludwig T, Chapman DL, Papaiouannou VE, Efstratiadis A.Targeted mutations of breast cancer susceptibility genehomologs in mice: Lethal phenotypes of Brca1, Brca2,Brca1/Brca2, Brca1/p53 and Brca2/p53 nullizygousembryos. Genes Dev 1997;11:1226±1241.
24. Connor F, Bertwistle D, Mee PJ, et al. Tumorigenesis and aDNA repair defect in mice with a truncating Brca2 mutation.Nat Genet 1997;17:423±430.
25. Friedman LS, Thislethwaite FC, Patel KJ, et al. Thymiclymphomas in mice with a truncating mutation in Brca2.Cancer Res 1998;58:1338±1343.
26. McAllister KA, Haugen-Strano A, Hagevik S, et al. Char-acterization of the rat and mouse homologues of theBRCA2 breast cancer susceptibility gene. Cancer Res1997;57:3121±3125.
27. Dix DJ, Allen JW, Collins BW, et al. Targeted gene disruptionof Hsp70-2 results in failed meiosis, germ cell apoptosis,and male infertility. Proc Natl Acad Sci USA 1996;93:3264±3268.
28. Hogan B, Beddington R, Costantini F, Lacy E. Manipulatingthe mouse embryo: A laboratory manual. Cold SpringHarbor: Cold Spring Harbor Press; 1994.
29. Wilkinson DG. Whole in situ hybridization of vertebrateembryos. In: Wilkinson DG, editor. In situ hybridization: Apractical approach. New York: Oxford University Press;1992. p 75±83.
30. Doetschman, T. Interpretation of phenotype in geneticallyengineered mice. Lab Animal Sci 1999;49:137±143.
118822 BBEENNNNEETTTT EETT AALL..

31. LeCouter JE, Kablar B, Whyte PFM, Ying C, Rudnicki MA.Strain-related embryonic lethality in mice lacking theretinoblastoma-related p130 gene. Development 1998;125:4669±4679.
32. Suzuki A, Luis de la Pompa J, Stambolic V, et al. High cancersusceptibility and embryonic lethality associated withmutation of the PTEN tumor suppressor gene in mice. CurrBiol 1998;8:1169±1178.
33. Di Cristofano A, Pesce B, Cordon-Cardo C, Pandol®PP. Pten is essential for embryonic developmentand tumor suppression. Nat Genet 1998;19:348±355.
34. Sibilia M, Wagner E. Strain-dependent epithelial defectsin mice lacking the EGF receptor. Science 1995;269:234±238.
35. Threadgill D, Dlugosz A, Hansen L, et al. Targeted disruptionof mouse EGF receptor: Effect of genetic background onmutant phenotype. Science 1995;269:230±234.
36. Gayther, SA, Pharoah PDP, Ponder BAJ. The genetics ofinherited breast cancer. J Mammary Gland Biol Neoplasia1998;3:365±376.
37. Rebbeck TR, Couch FJ, Kant, J, et al. Genetic heterogeneityin hereditary breast cancer: Role of BRCA1 and BRCA2. AmJ Hum Genet 1996;59:547±553.
38. Easton, DF. How many more breast cancer predispositiongenes are there? Breast Cancer Res 1999;1:1±4.
39. Ponnaiya B, Cornforth MN, Ullrich RL. Radiation-inducedchromosomal instability in BALB/c and C57BL/6 mice: Thedifference is as clear as black and white. Radiat Res 1997;147:121±125.
BBRRCCAA22--NNUULLLL EEMMBBRRYYOONNIICC SSUURRVVIIVVAALL OONN TTHHEE BBAALLBB//CC BBAACCKKGGRROOUUNNDD 118833










![Slamf1 -/- [ BALB/c.129]](https://img.dokumen.tips/doc/110x75/56815051550346895dbe5296/slamf1-balbc129.jpg)


















