Embed Size (px)
DESCRIPTION
BIOL2007 - INBREEDING AND NEUTRAL EVOLUTION Tutorial work due on Fri 25 Jan by 4:30 pm Put in “BIOL2007 Hand-in box” in Wolfson House office 306. Tutorial times to be announced. PREVIOUSLY Deterministic evolution, via natural selection. TODAY - PowerPoint PPT Presentation
Citation preview

BIOL2007 - INBREEDING AND NEUTRAL EVOLUTION
Tutorial work due on Fri 25 Jan by 4:30 pmPut in “BIOL2007 Hand-in box” in Wolfson House office 306. Tutorial times to be announced.
PREVIOUSLYDeterministic evolution, via natural selection. TODAYInbreeding: does not cause evolution on its own; -- affects arrangement of genes in populations; & has important fitness effects.
However, finite population size both inbreeding and random evolutionary change (or "genetic drift"). Stochastic.Mutation: also causes random genetic change;
but genetic drift is usually faster
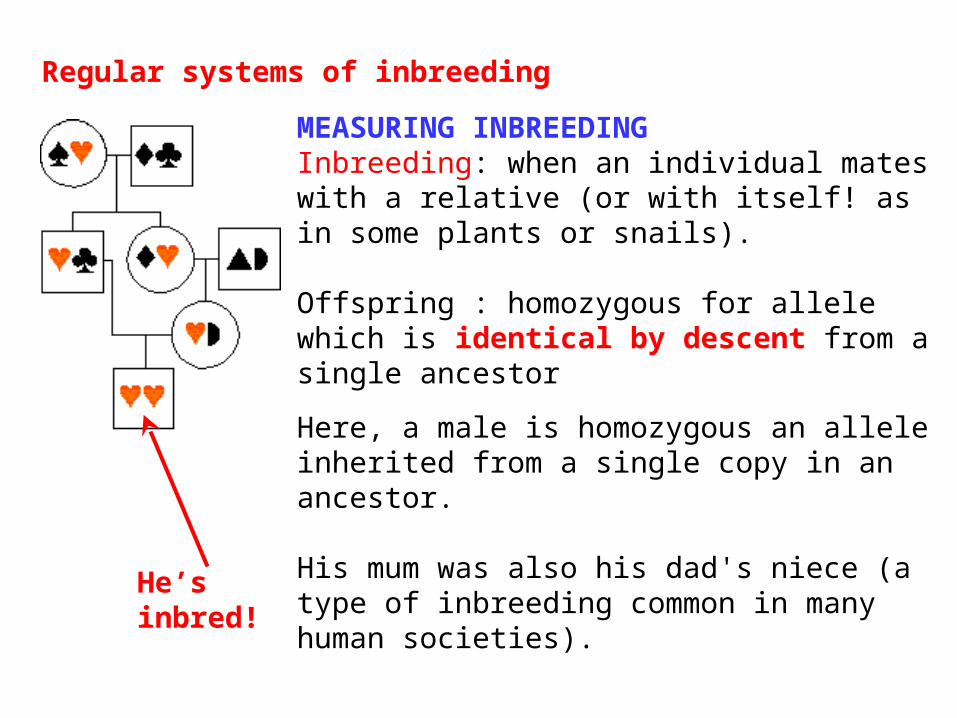
MEASURING INBREEDING Inbreeding: when an individual mates with a relative (or with itself! as in some plants or snails).
Offspring : homozygous for allele which is identical by descent from a single ancestor
Here, a male is homozygous an allele inherited from a single copy in an ancestor.
His mum was also his dad's niece (a type of inbreeding common in many human societies).
Regular systems of inbreeding
He’s inbred!

INBREEDING COEFFICIENT, F, ... used to gauge the strength of inbreeding.
F = probability that two alleles in an individual are identical by descent (IBD).
Identical by descent vs. identical in state
Identity in state (homozygosity) does not necessarily imply recent identity by descent. (Conversely ...)
F for fixation index: homozygosity, or “fixation”, results from inbreeding.
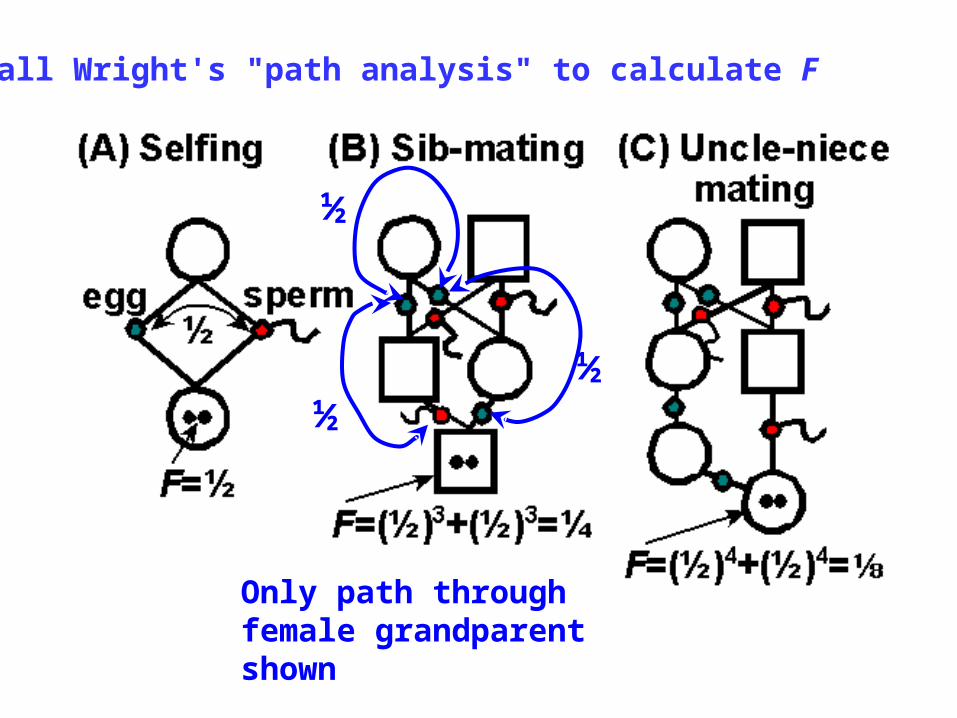
½
½
½
Only path through female grandparent shown
Sewall Wright's "path analysis" to calculate F

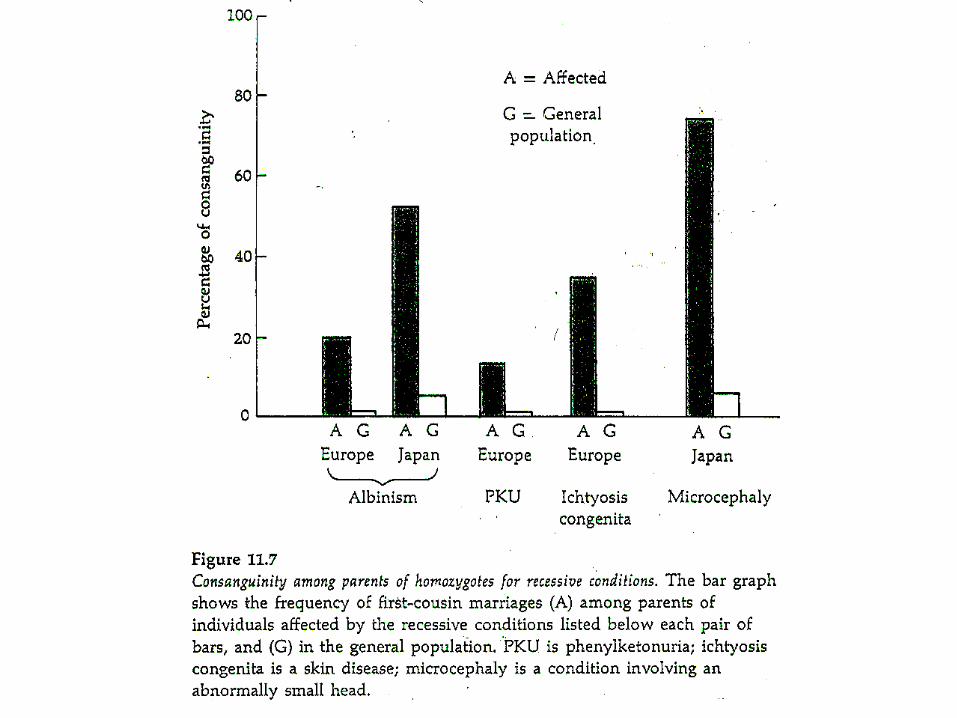
Problems of inbreeding:Deleterious recessive alleles in most populations.
Few deleterious recessives per gene (usually << 10-3)…but many deleterious alleles per genome.
You and I each carry about 1 strongly deleterious recessive mutation, or “lethal equivalent”.
When homozygous, these mutations cause problems,(inbreeding depression).
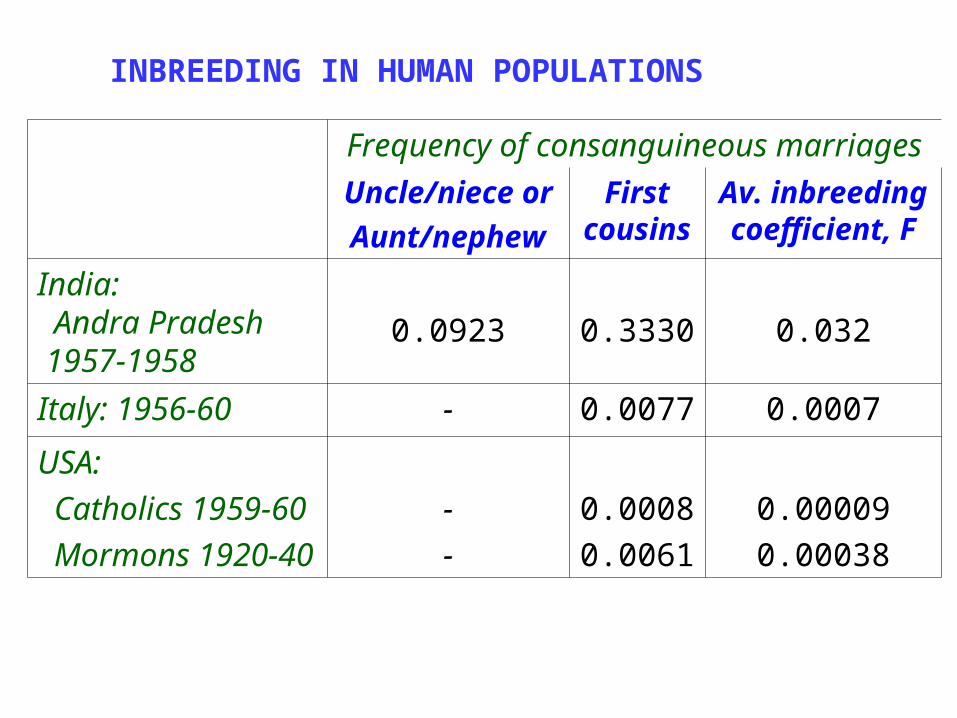
Frequency of consanguineous marriages
Uncle/niece or
Aunt/nephew
First cousin
s
Av. inbreeding coefficient,
F
India: Andra Pradesh 1957-1958
0.0923 0.3330 0.032
Italy: 1956-60 - 0.0077 0.0007
USA: Catholics 1959-60 Mormons 1920-40
--
0.00080.0061
0.000090.00038
INBREEDING IN HUMAN POPULATIONS

Inbreeding not all bad Many organisms habitually inbreed! e.g. fig wasps, parasites, weeds.
Advantages to inbreeding? Ecological: a single female can colonize May also usefully prevent recombinationDeleterious recessives in inbreeding species purged by selection.
Inbreeding in humans Ancient Egyptians, modern European royals, Indian subcontinent.Mild inbreeding, such as mating between first cousins, or uncle-niece isn't so dangerous. Example: Charles Darwin: married first cousin, 10 kids.
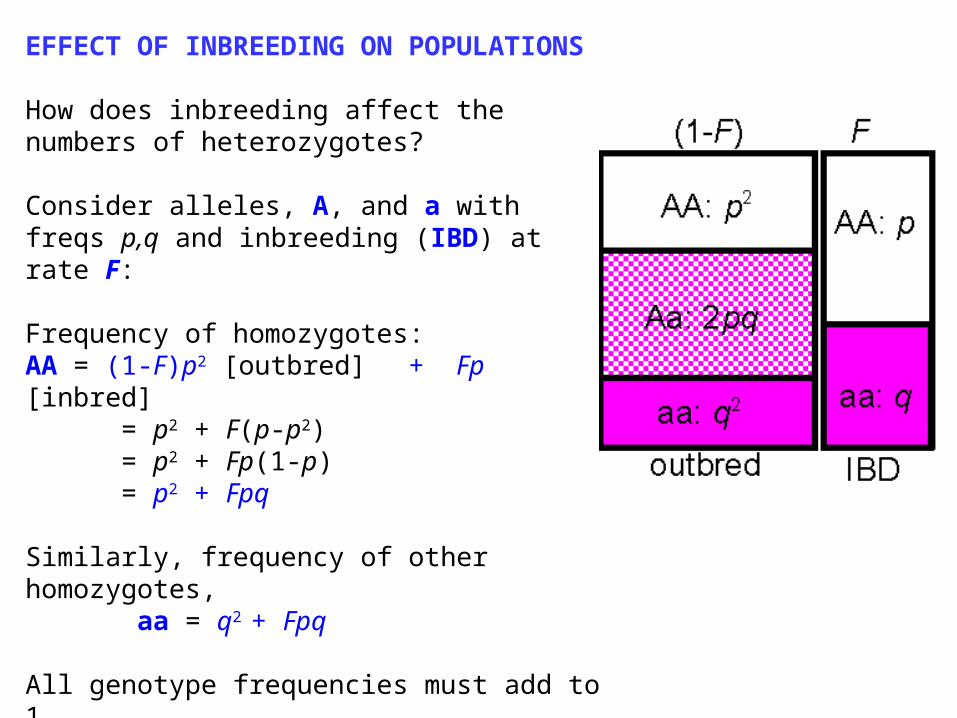
EFFECT OF INBREEDING ON POPULATIONS
How does inbreeding affect the numbers of heterozygotes?
Consider alleles, A, and a with freqs p,q and inbreeding (IBD) at rate F:
Frequency of homozygotes: AA = (1-F)p2 [outbred] + Fp [inbred]
= p2 + F(p-p2) = p2 + Fp(1-p) = p2 + Fpq
Similarly, frequency of other homozygotes, aa = q2 + Fpq
All genotype frequencies must add to 1so the extra 2Fpq AA and aa homozygotes must have come from the heterozygotes

Inbreeding leads to a reduction in heterozygosity. Heterozygosity (Het, i.e. fraction that are heterozygotes under inbreeding) is reduced by a fraction F compared with the outbred (Hardy-Weinberg) expectation HetHW = 2pq: Het = HetHW (1 - F)
F measures reduction of heterozygosity, or heterozygote deficit compared to Hardy-Weinberg, as well as probability of identity by descent!
genotype
AA Aa aa (Sum)
frequency
p2+Fpq 2pq(1-F)
q2+Fpq p2+Fpq+2pq-2Fpq+Fpq+q2
=1
EFFECT OF INBREEDING ON POPULATIONS

GENETIC DRIFT Deterministic vs. stochastic evolution Hardy-Weinberg: no gene frequency change.
True in an infinitely large population; evolution deterministic.
Only approximately true in populations of finite size.
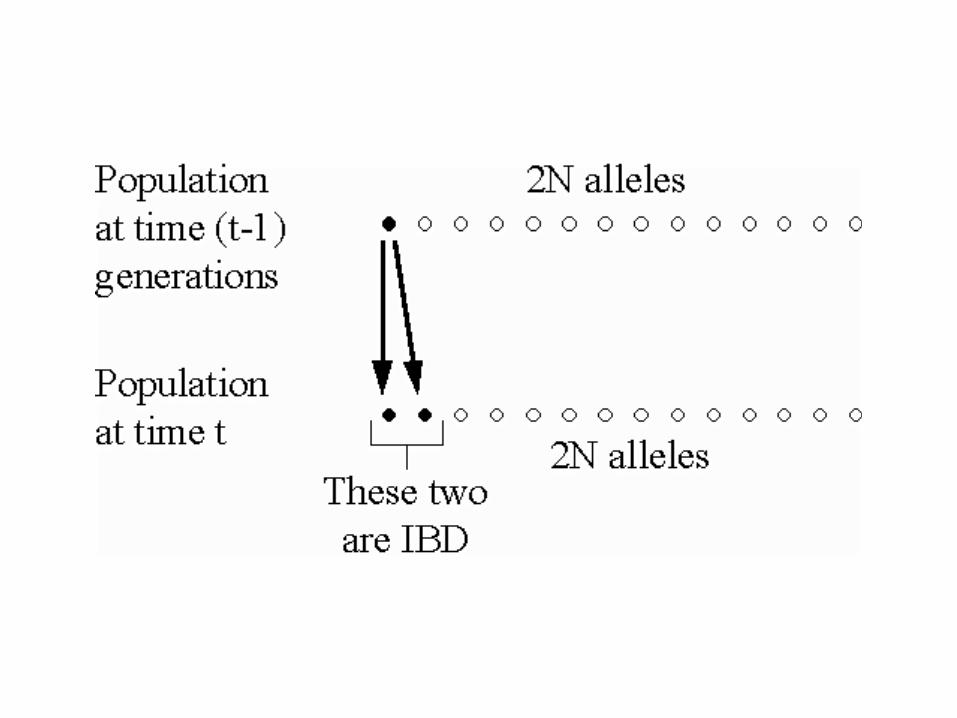
Assume a diploid population of constant size N. Each of 2N alleles are copied into gametes.
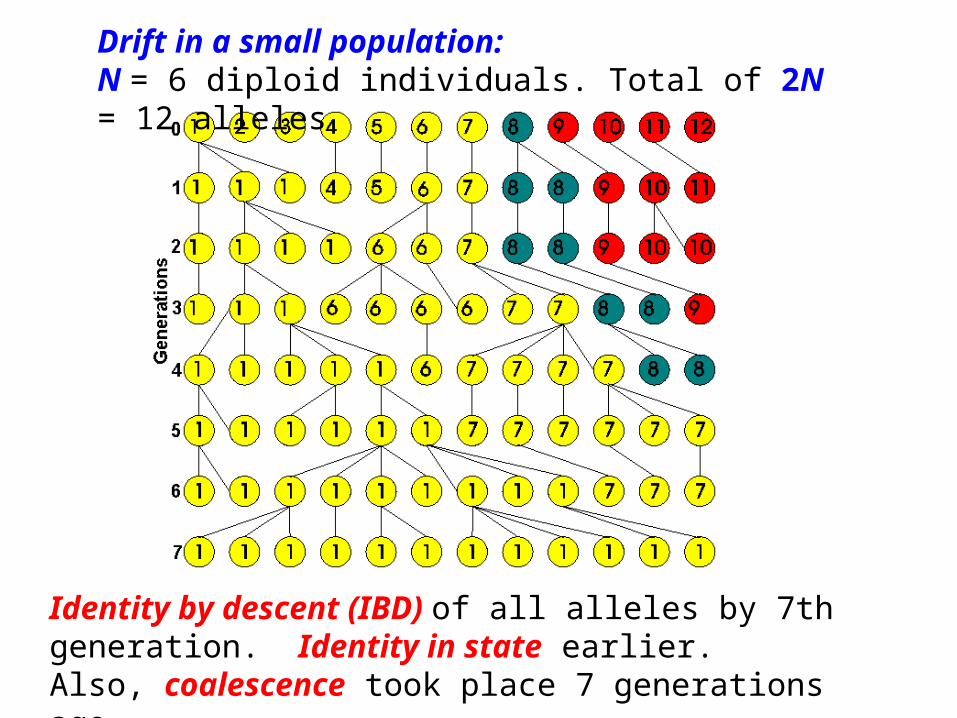
Drift in a small population:N = 6 diploid individuals. Total of 2N = 12 alleles
Identity by descent (IBD) of all alleles by 7th generation. Identity in state earlier. Also, coalescence took place 7 generations ago.
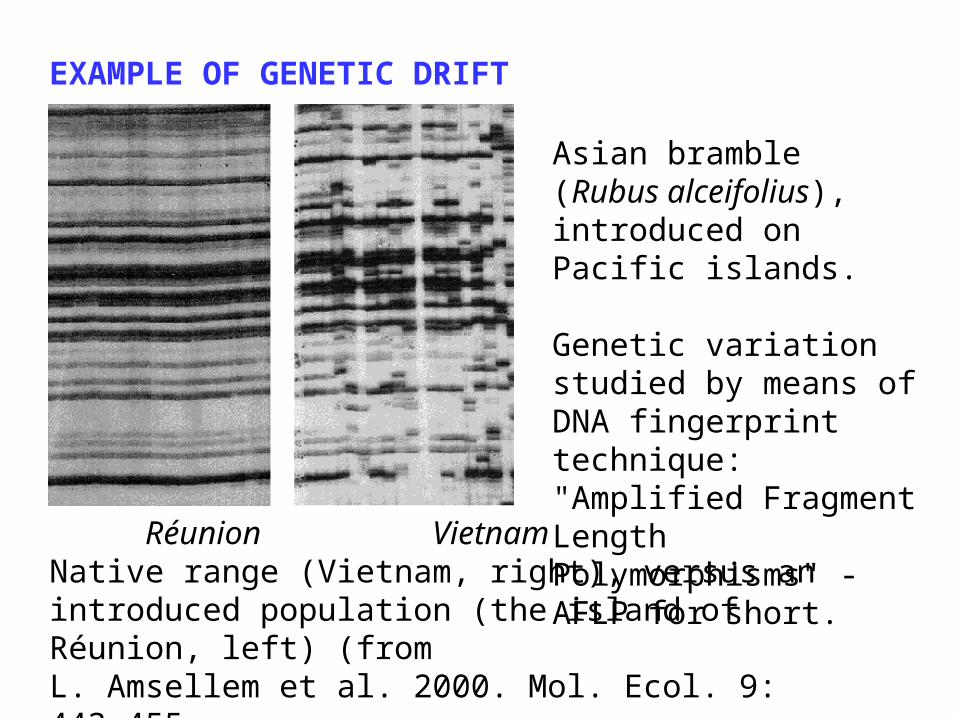
EXAMPLE OF GENETIC DRIFT
Réunion VietnamNative range (Vietnam, right), versus an introduced population (the island of Réunion, left) (from L. Amsellem et al. 2000. Mol. Ecol. 9: 443-455.
Asian bramble (Rubus alceifolius), introduced on Pacific islands.
Genetic variation studied by means of DNA fingerprint technique: "Amplified Fragment Length Polymorphisms" - AFLP for short.

GENETIC DRIFT AS A CAUSE OF INBREEDING
As we have seen, inbreeding results from drift because alleles become identical by descent (IBD). We can therefore measure drift in terms of our inbreeding coefficient, F, and hence how the fraction of heterozygosity, Het, declines with time. We can show:
t
NHettHet
211
0
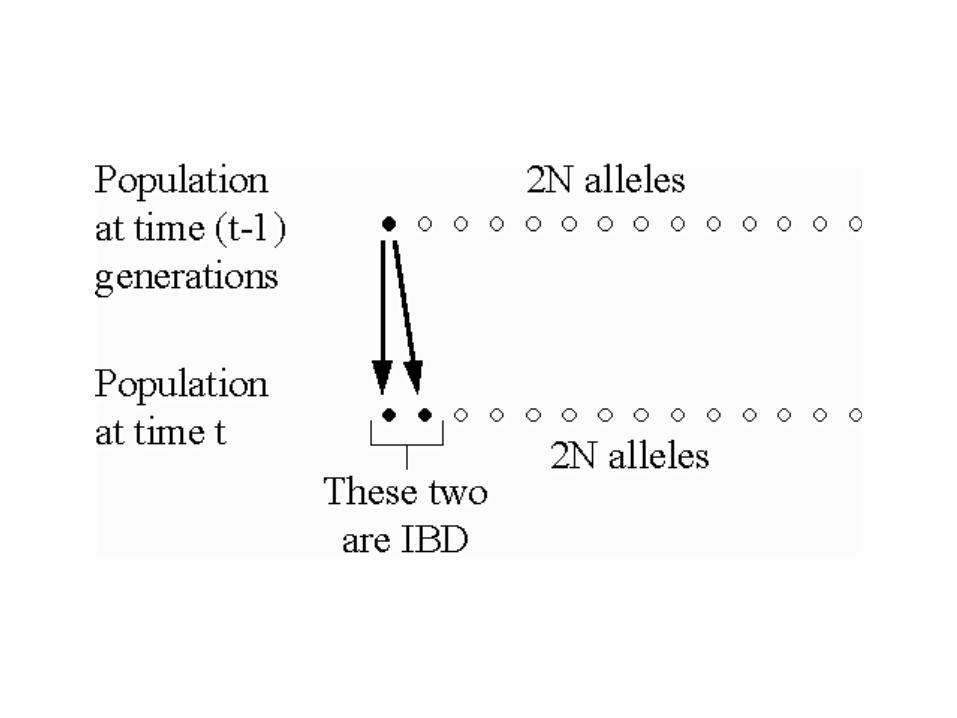

In a population of size N, the probability that two alleles picked at random mating in generation t are IBD due to copying from generation t-1 is (on average).
(inbred in generation t)
This is the rate of inbreeding due to drift per generation.(it measures the strength of drift).
BUT the 2N alleles in the previous generation may be IBD themselves from inbreeding in previous generations. The fraction of alleles in generation t that are IBD because of inbreeding before generation t-1 is:
(outbred ingeneration t)
NtF21
1211
tF
NtF

take 1–(both sides):
111
21
1211
21
tF
tF
NtF
NNtF
11.
211
11
21
111
tF
NtF
NtF
tF
Nt
HettHet211
1
t
NHetHett
211
0
By definition, the heterozygosity after a single generation of inbreeding is reduced by a fraction F, Het = 2pq (1 - F) = HetHW (1 - F), so Het/HetHW = (1 - F).
From the above equation relating Ft to Ft-1 and cancelling HetHW’s:
So, after t generations of drift:
Summing the inbreeding in the current generation with inbreeding from previous generations, we have at time t:

So heterozygosity declines approximately by a factor per generation. However, ...
(a) Only true on average. (And if assumptions met)
(b) F can also measure inbreeding – and drift – as a result of subdivision into finite populations.
It is the heterozygote deficit or identity by descent produced by subdivision. Usually written FST, inbreeding (F) due to subdivision into Subpopulations relative to the Total population.
N21
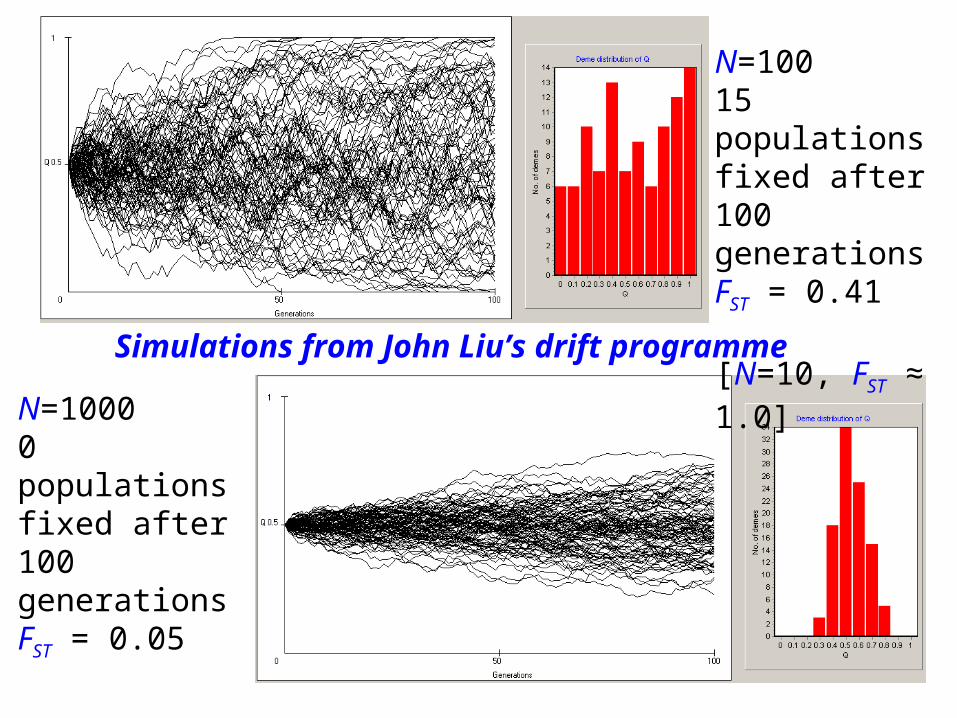
N=10015 populationsfixed after 100 generationsFST = 0.41
[N=10, FST ≈ 1.0]N=1000
0 populationsfixed after 100 generationsFST = 0.05
Simulations from John Liu’s drift programme

THE FLORIDA PANTHER
Genus PumaSpecies concolor Subspecies coryi
... may have a few problems of this nature.
Kink at the end of its tail; cowlick on back; 65% males cryptorchid; abnormal sperm – due to inbreeding?
Est. population in the wild: 50-70. Up from 30-50 in 1995.

EFFECTIVE POPULATION SIZE
Alleles usually do not have identical probability of being passed on, as required in simple models.
Population geneticists get around this by calculating effective population size, Ne that produces the same rate of genetic drift in their simple models with population size N.
Ne may differ from actual population size. Examples:
1) Separate sexes2) Unequal sex ratio3) Some males mate more than others

INBREEDING -- Conclusions Inbreeding coefficient, F – the measure of inbreeding.
Regular systems of inbreedingF is also a measure of heterozygote deficit.
Inbreeding due to genetic drift in finite populations.The extent of drift can also be measured by F … or Het = 2pq (1–F).
All evolution is somewhat stochastic: a mix of random genetic drift with deterministic – selection.
– an important equation in conservation, deduced from the effect of drift on inbreeding in population of size N.
t
NHetHett
211
0

FURTHER READING
FUTUYMA, DJ 2005. Evolution. Chapter 9: 197-199, 201-202, Chapter 10.
FREEMAN & HERRON 2004. Evolutionary Analysis.Chapter 6. pp. 204-252.














![On the expected relationship between inbreeding, …...extinction is correlated to neutral genetic variation. The model of Morton et al. [34] is still widely used to evaluate the e](https://img.dokumen.tips/doc/110x75/5f3b7a83e1708e18705a7d94/on-the-expected-relationship-between-inbreeding-extinction-is-correlated-to.jpg)















