Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 1988, p. 143-1490099-2240/88/010143-07$02.00/0Copyright C) 1988, American Society for Microbiology
Vol. 54, No. 1
Biodegradation of Alpha- and Beta-Hexachlorocyclohexane in a SoilSlurry under Different Redox Conditions
A. BACHMANN,t P. WALET, P. WIJNEN, W. DE BRUIN, J. L. M. HUNTJENS, W. ROELOFSEN,AND A. J. B. ZEHNDER*
Department of Microbiology, Agricultural University, 6703 CT Wageningen, The Netherlands
Received 3 September 1987/Accepted 20 October 1987
Aerobic conditions proved to be best for the microbial conversion of alpha-hexachlorocyclohexane(alpha-HCH) in a soil slurry. The dry soil contained 400 mg of alpha-HCH per kg. This xenobiotic compoundwas mineralized within about 18 days at an initial rate of 23 mg/kg of soil per day by the mixed native microbialpopulation of the soil. The only intermediate that was detected during breakdown was pentachlorocyclohexene,which was detected at very small concentrations. Alpha-HCH was also bioconverted under methanogenicconditions. However, a rather long acclimation period (about 30 days) was necessary before degradationstarted, at a rate of 13 mg/kg of soil per day. Mass balance calculations showed that about 85% of the initialalpha-HCH that was present was converted to monochlorobenzene, 3,5-dichlorophenol, and a trichlorophenolisomer, possibly 2,4,5-trichlorophenol. Under both denitrifying and sulfate-reducing conditions, no significantbioconversion of alpha-HCH was observed. The beta isomer of HCH was recalcitrant at all of the four redoxconditions studied. We propose that the specific spatial chloride arrangement of the beta isomer is responsiblefor its stability. The results reported here with complex soil slurry systems showed that alpha-HCH is, incontrast to the existing data in the literature, best degraded biologically in the presence of oxygen.
Close to 6,000 sites of soil contaminated with organicchemicals are known in The Netherlands, and their numberis still increasing (Dutch Ministry of Housing, PhysicalPlanning and Environment, 1985). Because the groundwaterlevel is generally very high in The Netherlands and becausegroundwater is the major source of drinking water, thesecontaminated soil sites constitute an obvious potential healthhazard. About 100 of these known contaminated soil sitesare mainly polluted with various isomers of hexachlorocy-clohexane (HCH). Much work on HCH bioconvertibility hasbeen done already. In most of the studies, however, theconcern was over lindane, the gamma isomer of HCH; andthe studies have been performed with generally low (<10mg/liter) HCH concentrations (3-9, 13-15, 18, 20, 24, 28, 31).According to the results presented in the literature, HCHcan be degraded more easily under anaerobic than aerobicconditions (3-6, 8, 17, 18, 31). Several investigators haveclaimed (9, 14, 15) that all four HCH isomers are bioconvert-ible, i.e., alpha-, beta-, gamma-, and delta-HCH. Bacteriathat are able to degrade gamma-HCH anaerobically formtetrachlorocyclohexene as the main intermediate (3, 6, 9, 12,17, 20). Small amounts of tri- and tetrachlorinated benzeneshave been found as well (9). Besides these compounds,monochlorobenzene (MCB) seems to be an important inter-mediate under certain experimental conditions (8, 16). Lin-dane was metabolized by several aerobic microorganisms,which were isolated from a loamy sand, to pentachlorocy-clohexene, tetrachlorocyclohexene, pentachlorobenzene,tetrachlorobenzenes, as well as trichlorobenzenes (28). Withregard to the relative biodegradation rates of the differentHCH isomers, it is generally accepted that the bioconversionrates are independent of the redox conditions (3, 20). Therelative biodegradation rates are as follows: gamma-HCH >alpha-HCH > delta-HCH - beta-HCH. Only aerobic micro-
* Corresponding author.t Present address: Electrowatt Engineering Services Ltd., CH-
8022 Zurich, Switzerland.
organisms show growth with lindane as the sole carbon andenergy source (28). Dechlorination of lindane to gamma-pentachlorocyclohexene was also shown to occur with fungiin aqueous suspensions (1, 12) and in algal cultures (26). Fewstudies have been done on the biodegradation ofHCH underdenitrifying conditions. Only one pure culture of a denitrify-ing organism (Paracoccus denitrificans) has been tested upto now (9). No HCH conversion was observed. HCHdegradation under sulfate-reducing conditions has not yetbeen studied.
In this report we describe the biodegradation of alpha- andbeta-HCH in a heavily contaminated soil slurry. Dependingon the redox conditions applied, the soil was seeded withactivated sludge (aerobic and denitrification), anaerobic ma-rine sediment (sulfate reduction), and anaerobic sewagesludge (methanogenesis). The possibilities for the engineer-ing applications of the results for the cleanup of this andsimilarly contaminated soil environments are also discussed.
MATERIALS AND METHODS
Sampling. Large samples of contaminated soil were col-lected at a contaminated site in Hengelo, The Netherlands.The sampling depth was a maximum of 50 cm. The sampleswere sieved through an 8-mm-mesh sieve. Fifty kilograms ofthe sieved soil was made into a slurry (30% water; wt/wt),which was blended for several minutes until a smooth pastedeveloped in which only a few soil clumps remained visible.The soil slurry was kept in individual batches at -20°C.Physical-chemical soil parameters and xenobiotic soil con-taminants were analyzed from freshly thawed soil samples.Media and chemicals. The mineral salts medium used for
the experiments under methanogenic conditions containedthe following, per liter: K2HPO4, 60 mg; NaHCO3, 4 g;NH4NO3, 3.7 mg; FeCl2, 5 mg; CoCl2, 0.5 mg; NiCl2, 0.25mg; MgSO4 7H20, 150 mg; CaCl2, 25 mg; KCl, 25 mg. Forthe experiment under sulfate-reducing conditions, a modifiedmarine-type mnineral salts medium was used (34). It con-tained the following, per liter: NaCl, 19.89 g; MgCl2, 1.43 g;
143
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

144 BACHMANN ET AL.
KClI 298 mg; CaCl2, 111 mg; NH4Cl, 268 mg; KH2PO4, 204mg; Na2SO4, 2.84 g; NaHCO3, 2.52 g; NaBr, 82 mg; SrCl26H20, 24 mg; FeCl2, 2 mg; ZnCI2, 0.05 mg; MnCl2 4H20,0.05 mg; AlCl3, 0.05 mg; NiCl2, 0.05 mg; H3B03, 0.05 mg;CuC12, 0.03 mg; (NH4)6Mo7024 4H20, 0.05 mg; CoCl26H20, 0.05 mg; EDTA, 0.5 mg. For the experiment carriedout under denitrifying conditions, the mineral salts mediumcontained the following, per liter: NaNO3, 450 mg; K2HPO4,100 mg; MgSO4 7H20, 20 mg; FeSO4 7H20, 5 mg;
CaCl2, 2 mg; MnCl2 4H20, 0.2 mg; NaMoO4 2H20, 0.1mg. For the aerobic experimernts, the mineral salts mediumcontained the following, per liter: KH2PO4, 170 mg;K2HPO4, 570 mg; Na2HPO4, 668 mg; NaHCO3, 20 mg;
FeCl3 6H20, 2.03 mg; NH4NO3, 185.5 mg; MgSO4, 22.5mg; CaCl2, 27.5 mg. HCH isomers were obtained fromSchmidt B.V., Amsterdam, The Netherlands; [cx-14C]HCHwas obtained from Amersham International plc, Bucking-hamshire, England. All other chemicals were reagent gradeand were obtained from commercial sources.
Biodegradation studies. Biodegradation experiments were
carried out in the dark at 30°C in screw-cap glass septumvials (volume, 15 ml). They were continuously shaken on an
overhead rotary shaker (8 rpm; amplitude, 20 cm). Externalmass transfer limitations were found to be nonlimiting at theslurry concentrations used. A total glucose plus acetateconcentration, in a 1:1 ratio, of 630 mg/liter was added to theincubation vials, unless stated otherwise. Soil toxicity testswere done by using the anaerobic toxicity assay describedby Owen et al. (21). The soil slurry samples were seeded (0.7g [dry weight]/liter) with activated sludge (aerobic and deni-trification), anaerobic marine sediment (sulfate reduction),and anaerobic sewage sludge (methanogenesis). Contam-inated soil concentrations of 100 g (dry weight)/liter wereused in all experiments, unless stated otherwise. Sterilecontrols used for biodegradation experiments were treatedwith gamma-radiation at 2.5 M rad (Gammaster, B. V., Ede,The Netherlands), followed by pasteurization for 30 min at70°C. Pasteurization was necessary to inactivate free en-
zymes.Except for the mixtures used in the toxicity test, no HCH
was added to the incubation mixtures. The extent of 14CHdegradation reported relates to the HCH that was alreadypresent in the contaminated soil. The data are given inmilligram per kilogram of dry soil, unless specified other-wise.
Analytical methods. After 10 ml of acetonitrile was addedto 5 ml of soil slurry and the incubation mixture was shakenfor 16 h, 1 ,ul of centrifuged and filtered acetonitrile-waterextract was analyzed with a gas chromatograph (model 438;Packard Instrument Co., Inc., Rockville, Md.). The unit wasequipped with a splitter (1:100) and an electron capturedetector fitted to a WCOT fused capillary column (SIL5,Chrompack), which was kept at 160°C. Free chloride was
determined potentiometrically by titration with a microchlo-rocounter (Marius, Utrecht, The Netherlands).
Soil contaminants and metabolites produced during incu-bation were analyzed by high-pressure liquid chromatogra-phy, gas chromatography, or mass spectrometry. Identifica-tions by gas chromatography and high-pressure liquidchromatography were performed by comnparing the retentiontimes of the unknown compound with those of pure sub-stances. Most compounds were identified by at least twomethods. High-pressure liquid chromatography was donewith a pump (2150; LKB Instruments, Inc., Rockville, Md.)and a controller (2152; LKB); an Ultra Pac column (250 by 4mm) filled with Lichrosorb RP-8 and RP-18 (diameter 10
pm), was used; this was preceded by chromatography on auniversal column (75 by 2.1 mm; RP; Chrompack). Themobile phase was acetonitrile-water (ratio, by volume,60:40), the flow rate was 1 ml/min, and 20- and 50-,u sampleswere analyzed with a detector (SD; Uvicord) at 206 nm andquantified with a computing integrator (4270; Spectra Phys-ics). Identification by mass spectrometry was performedwith a mass spectrometer (VG 7070) that was operated in theelectron impact ionization mode (70 eV). The compoundswere directly introduced with a gas chromatograph (Pye 204)equipped with a fused silica capillary column (26 m by 0.25mm; CP SIL 19 CB). Gas analyses (02, N2, N20, CH4, C02)were performed on a gas chromatograph (model 406;Becker). The biochemical oxygen demand assay was done ina Sapromat (11). S2- was determined by the method de-scribed by Trueper and Schlegel (27), and denitrificationactivity was determined as outlined by Yoshinari et al. (32).Bacteria were counted in contaminated soil by the methoddescribed by Zvyagintsev et al. (34). Aerobic alpha-HCH-degrading bacteria were counted by the most probablenumber method (22). Before bacteria were counted, theywere removed from the soil particles by high-speed blending(2x/min), followed by sonification (two times, 10 s eachtime, at 150 W).
Analytical precision. All reported biodegradation data arebased on results of duplicate samples. Standard errors werewithin 15% of the values determined by gas chromatographicanalyses and within 7% of the values determined by freechloride analyses. The HCH detection limit was 5 mg/kg ofdry soil.
RESULTS
Characterization of the contaminated soil. Some of thecharacteristics of the soil slurry are as follows: soil watercontent, 30.0 ± 0.2% (standard deviation [SD] based on sixsamples); soil pH, 7.7 ± 0.1 (SD based on three determina-tions); soil organic content, 0.9 ± 0.06% (SD based on sixsamples); soil Kjeldahl nitrogen content, 0.052 ± 0.003%(SD based on five determinations); soil biochemical oxygendemand, below the detection limit; CaCO3, 38 + 2% (SDbased on three determinations); specific weight, 1,870 ± 40g/liter (SD based on three determinations); total soil bacte-ria, >108 cells per ml; aerobic alpha-HCH degraders, >103cells per ml. The soil that was used had a rather high pH(about 7.7) and a low soil organic and Kjeldahl nitrogencontent of about 0.9 and 0.05% (wt/wt), respectively. About60% of the constituents of the soil was sandy material. Theoriginal sandy soil was treated with 38% (wt/wt) lime by thechemical company as they disposed their HCH wastes.The soil was found to be contaminated with various
chlorinated benzenes, as well as two dichlorophenol isomers(Table 1). The contamination level of these aromatic com-pounds was relatively low, ranging from 0.32 mg/kg of drysoil for 2,3-dichlorophenol (2,3-DCP) to 4.34 mg/kg of drysoil for 1,2,4-trichlorobenzene. The major pollutants werealpha-HCH (400 mg/kg of dry soil) and beta-HCH (250 mg/kgof dry soil). Two other 14CH isomers, gamnma and delta,were also detected but at much lower levels (22 and 13 mg/kgof dry soil, respectively).To differentiate between the inability of native popula-
tions, added microbial populations, or both to degrade HCHand the inhibition of HCH breakdown due to the chemicals(HCH and others) that were present in the contaminatedsoil, a toxicity test was performed. Depending on the redoxconditions applied, oxygen consumption, N20 production,
APPL. ENVIRON. MICROBIOL.
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

HCH BIOCONVERSION IN SOIL SLURRY 145
TABLE 1. Xenobiotic contaminants found in the originalhomogenized soil slurry
Compound Concn (mg/kg of drysoil)"
MCB ................................. 2.23 0.18"1,2-DCB ................................. 1.30 ± 0.151,4-DCB ............... ................... 1.53 ± 0.111,2,4-Trichlorobenzene ............................ 4.34 ± 0.061,3,5-Trichlorobenzene ............................ 0.83 ± 0.211,2,3,4-Tetrachlorobenzene ....................... 2.08 t 0.251,2,3,5-Tetrachlorobenzene ....................... 1.61 ± 0.452,3-DCP .................................. 0.32 ± 0.453,5-DCP ................................. 1.25 ± 0.54alpha-HCH ................................. 400 ± 36beta-HCH ................. ................ 230 ± 15gamma-HCH ................................. 22 ± 0.1delta-HCH .................................. 13 ± 0.3
a SDs are based on eight and four samples for aromatic and HCH values,respectively.
S2- formation, or methane release was used as the parame-ter to be tested. Toxicity was expressed by the maximumrate ratio. The maximum rate ratio was calculated by divid-ing the initial production or consumption rate in samplescontaining various amounts of contaminated soil by theinitial rate observed in a sample without soil. According toOwen et al. (21), a maximum rate ratio of less than 0.90 isalready indicative of significant toxicity in the case ofmethane production. Measurements were done for 6 daysafter the start of the experiment.
Toxicity data are presented in Fig. 1. From these data itcan be concluded that the contaminated soil was severelytoxic for the mixed methanogenic and the marine sulfate-reducing populations that were used but did not inhibitaerobic respiration or denitrification. To investigate whetherthe HCH that was present in the soil caused the observedinhibition, its effects on oxygen consumption and methaneformation were tested as well. In these experiments, no soilwas added and only the seed microorganisms (0.7 g [dryweight]/liter) were exposed to various alpha-HCH concen-trations. No adverse effect was found under aerobic condi-tions (Fig. 1, dashed line). Methanogenesis was slightlyreduced, but was reduced much less than at comparableHCH levels in soil. Therefore, it is probable that, in additionto HCH, other compounds (or combinations thereof) that arealso present in the contaminated soil are responsible for theinhibition of methanogenesis.
In all later soil experiments a contaminated soil concen-tration of 100 g (dry weight)/liter was chosen as a conse-quence of these results. This amount of soil was judged to besufficiently small as to prevent excess toxicity and wassufficiently large to study the biodegradation of the xenobio-tic compounds that are present in a soil slurry environment.External mass transfer limitations were found to be nonlim-iting at the slurry concentrations used.
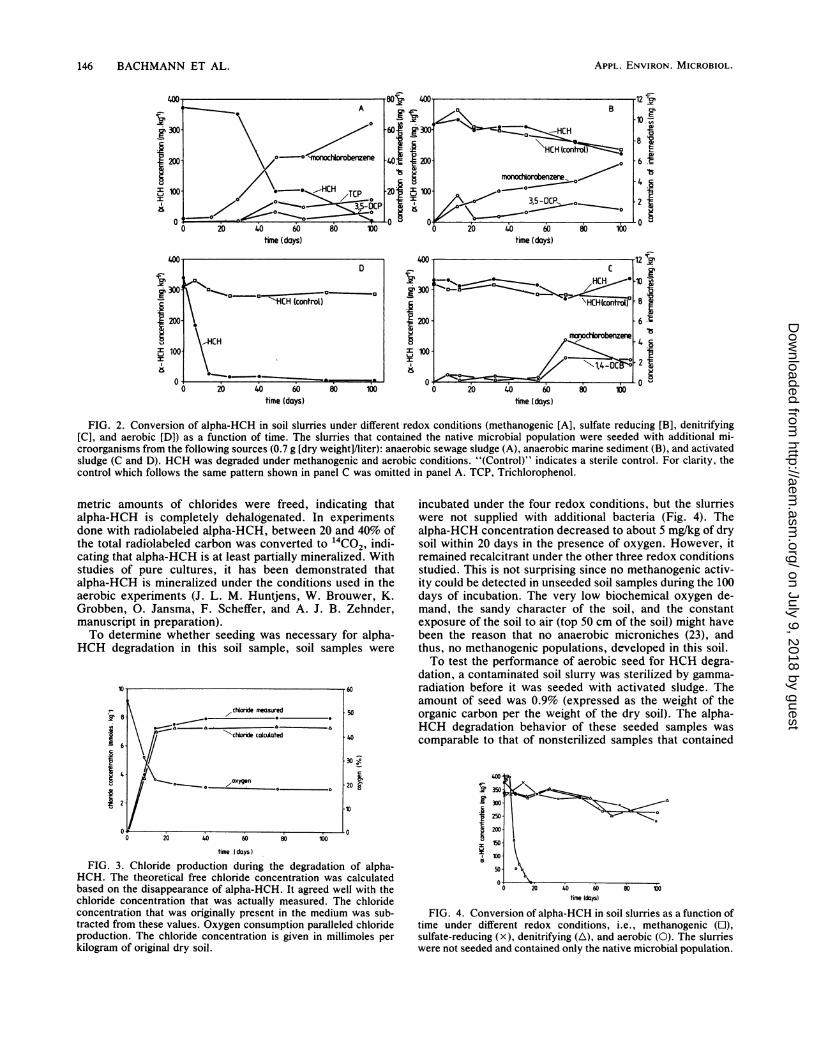
Microbial HCH degradation under different redox condi-tions. After a long lag time (about 25 days; Fig. 2A),alpha-HCH was bioconverted at an initial rate of 13 mg/kg ofdry soil per day under methanogenic conditions. The de-crease of alpha-HCH was paralleled by an increase in MCB.Within 100 days, approximately 65% of the alpha-HCH thatwas degraded was converted into MCB. In addition, 3,5-DCP and a trichlorophenol isomer, possibly 2,4,5-trichloro-phenol, also built up with time. A mass balance at day 100showed that about 85% of the initial alpha-HCH that waspresent in the soil was converted to these aromatic interme-
diates. Also, 1,4-dichlorobenzene (1,4-DCB) increased fromits background contamination level of 1.5 mg/kg to about 3.5mg/kg at day 100 (data not shown). This increase was sosmall, however, that it may have resulted from the biocon-version of other chlorinated benzenes that were alreadypresent in the soil (Table 1) and not directly from alpha-HCH. The rest of the soil contaminants showed only small,if any, changes in concentrations. Intraparticle chemicaladsorption might have caused this recalcitrance.Over the 100 days of incubation, no significant decrease in
alpha-HCH was obvious when sulfate served as the mainelectron acceptor (Fig. 2B). Analogous to the case with themethanogen, however, MCB and 3,5-DCP built up duringincubation. The degree of conversion, however, was muchlower. Within 100 days about 6 and 1% of the initialalpha-HCH present was converted to MCB and 3,5-DCP,respectively. The observed intermediate buildup could alsohave resulted from conversion of the small amounts ofgamma-HCH or other soil contaminants that were present(Table 1).No significant alphaLHCH biodegradation could be de-
tected over the 103 days of incubation under denitrifyingconditions (Fig. 2C). In the later stage of the experiment,two intermediates, MCB and 1,4-DCB, were detected at lowconcentrations. A likely explanation for the observed alpha-HCH recalcitrance was the presence of nitrate. Nitrate/hasbeen shown to inhibit bacterial reductive dechlorination ofgamma-HCH and dichlorodiphenyltrichloroethane (24, 30).Aerobic conditions lead to a relatively fast (23 mg/kg of drysoil per day) and complete alpha-HCH bioconversion overthe 100 days of the investigation (Fig. 2D). Only one metab-olite, pentachlorocyclohexene, was identified during incuba-tion, but the concentrations were very low (at most, a fewmicrograms per liter). The observed aerobic behavior wassuperior to the behavior of the methanogen with respect toboth lag time and conversion rates (Fig. 2) and, therefore,was in sharp contrast to previously published data (3-6, 8,17, 18, 31), probably because of the inhibiting effect ofcertain soil contaminants on mixed methanogenic popula-tions. The decrease in alpha-HCH, as determined by anincrease in free chloride, was paralleled by a decrease inoxygen (Fig. 3). A chloride balance showed that stoichio-
1.2
o
C3 a:VaOEE
20- aerobic (soil) - aerobic (HCH)Wt denitrifying (soil)
_methanogenic (HCH)75- '} ^\
50- sulfate reducing (soil)
°_ methanogenic (soil)0~~~~~~~~~~~~~~~~iX
100 200soil concentration 1g dry soil 1
300
12s
0
0 25 50 75 100HCH concentration (mg. (-1)
FIG. 1. Toxic effects of various concentrations of contaminatedsoil and HCH on microbial activities under different redox condi-tions. A maximum rate ratio of 1.0 indicates no toxicity. The HCHconcentration corresponds to the amount of alpha-HCH that was
present in the contaminated soil. Toxicity tests performed withcontaminated soil are indicated "(soil)." "(HCH)" indicates exper-iments in which aerobic and methanogenic seed cultures were
exposed to various concentration of added alpha-HCH in theabsence of soil.
VOL. 54, 1988
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
146 BACHMANN ET AL.
300
200
510Zs
%
o
time (days)
4000'.D
g300 a
_-HCH(controt)
200 \
A\CHI= 100 \
0 20 40 60 80 100time (days)
4-
D8= 8(tI
I
20 40 60 80 10time (days)
12
400 12
300 o oC X t1~~~~~~~~ HCH(catral@ ~ ~~~~~8
200- 6monochdoraobenzere
00 -a //°tF:~~~~~2
0 0 0 0I 100time (days)
FIG. 2. Conversion of alpha-HCH in soil slurries under different redox conditions (methanogenic [A], sulfate reducing [B], denitrifying[C], and aerobic [D]) as a function of time. The slurries that contained the native microbial population were seeded with additional mi-croorganisms from the following sources (0.7 g [dry weight]/liter): anaerobic sewage sludge (A), anaerobic marine sediment (B), and activatedsludge (C and D). HCH was degraded under methanogenic and aerobic conditions. "(Control)" indicates a sterile control. For clarity, thecontrol which follows the same pattern shown in panel C was omitted in panel A. TCP, Trichlorophenol.
metric amounts of chlorides were freed, indicating thatalpha-HCH is completely dehalogenated. In experimentsdone with radiolabeled alpha-HCH, between 20 and 40% ofthe total radiolabeled carbon was converted to 14C02, indi-cating that alpha-HCH is at least partially mineralized. Withstudies of pure cultures, it has been demonstrated thatalpha-HCH is mineralized under the conditions used in theaerobic experiments (J. L. M. Huntjens, W. Brouwer, K.Grobben, 0. Jansma, F. Scheffer, and A. J. B. Zehnder,manuscript in preparation).To determine whether seeding was necessary for alpha-
HCH degradation in this soil sample, soil samples were
10 60
\ /chloride measured 50
I ~~~~~~-choridecitctd4
4~~~~~~~~~~~~~~~~~0
-o*~~~xygent~~~~~2 _
0 20 40 60 80 10
time (days)
FIG. 3. Chloride production during the degradation of alpha-HCH. The theoretical free chloride concentration was calculatedbased on the disappearance of alpha-HCH. It agreed well with thechloride concentration that was actually measured. The chlorideconcentration that was originally present in the medium was sub-tracted from these values. Oxygen consumption paralleled chlorideproduction. The chloride concentration is given in millimoles per
kilogram of original dry soil.
incubated under the four redox conditions, but the slurrieswere not supplied with additional bacteria (Fig. 4). Thealpha-HCH concentration decreased to about 5 mg/kg of drysoil within 20 days in the presence of oxygen. However, itremained recalcitrant under the other three redox conditionsstudied. This is not surprising since no methanogenic activ-ity could be detected in unseeded soil samples during the 100days of incubation. The very low biochemical oxygen de-mand, the sandy character of the soil, and the constantexposure of the soil to air (top 50 cm of the soil) might havebeen the reason that no anaerobic microniches (23), andthus, no methanogenic populations, developed in this soil.To test the performance of aerobic seed for HCH degra-
dation, a contaminated soil slurry was sterilized by gamma-radiation before it was seeded with activated sludge. Theamount of seed was 0.9% (expressed as the weight of theorganic carbon per the weight of the dry soil). The alpha-HCH degradation behavior of these seeded samples wascomparable to that of nonsterilized samples that contained
I
401
35
3D
29
20
19
o a50
o 20 40 60 80 o
time (days)
FIG. 4. Conversion of alpha-HCH in soil slurries as a function oftime under different redox conditions, i.e., methanogenic (l),sulfate-reducing (x), denitrifying (Lx), and aerobic (0). The slurrieswere not seeded and contained only the native microbial population.
00
~~-HCH~~61 \/°~0- b<ermene -4C
0 2 S00 80CPX10
0 20 40 60 80 100
B.10
.8
30 ~~HCH(canlo ~monodh(orobenzene .4
D0 nt=o3
I°-o*
Ln
E
.i
) t
.!
Fs
It
APPL. ENVIRON. MICROBIOL.
1.t
9
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
HCH BIOCONVERSION IN SOIL SLURRY 147
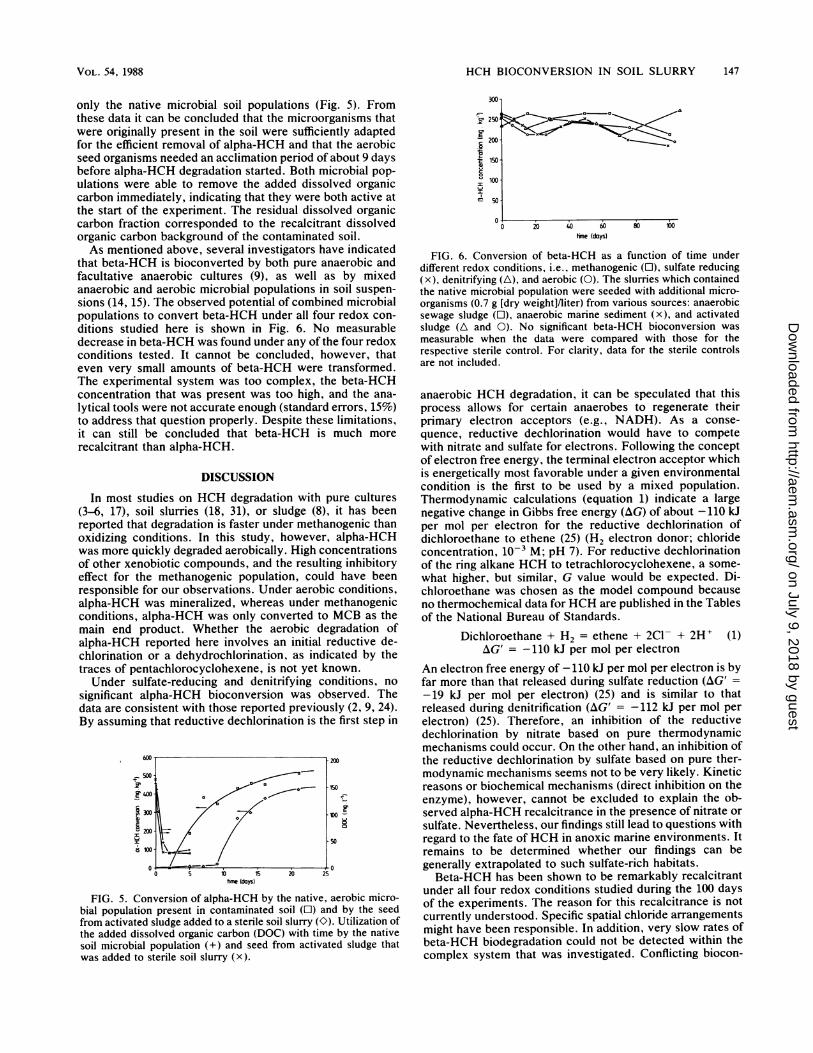
only the native microbial soil populations (Fig. 5). Fromthese data it can be concluded that the microorganisms thatwere originally present in the soil were sufficiently adaptedfor the efficient removal of alpha-HCH and that the aerobicseed organisms needed an acclimation period of about 9 daysbefore alpha-HCH degradation started. Both microbial pop-
ulations were able to remove the added dissolved organiccarbon immediately, indicating that they were both active atthe start of the experiment. The residual dissolved organiccarbon fraction corresponded to the recalcitrant dissolvedorganic carbon background of the contaminated soil.As mentioned above, several investigators have indicated
that beta-HCH is bioconverted by both pure anaerobic andfacultative anaerobic cultures (9), as well as by mixedanaerobic and aerobic microbial populations in soil suspen-
sions (14, 15). The observed potential of combined microbialpopulations to convert beta-HCH under all four redox con-
ditions studied here is shown in Fig. 6. No measurabledecrease in beta-HCH was found under any of the four redoxconditions tested. It cannot be concluded, however, thateven very small amounts of beta-HCH were transformed.The experimental system was too complex, the beta-HCHconcentration that was present was too high, and the ana-
lytical tools were not accurate enough (standard errors, 15%)to address that question properly. Despite these limitations,it can still be concluded that beta-HCH is much more
recalcitrant than alpha-HCH.
DISCUSSION
In most studies on HCH degradation with pure cultures(3-6, 17), soil slurries (18, 31), or sludge (8), it has beenreported that degradation is faster under methanogenic thanoxidizing conditions. In this study, however, alpha-HCHwas more quickly degraded aerobically. High concentrationsof other xenobiotic compounds, and the resulting inhibitoryeffect for the methanogenic population, could have beenresponsible for our observations. Under aerobic conditions,alpha-HCH was mineralized, whereas under methanogenicconditions, alpha-HCH was only converted to MCB as themain end product. Whether the aerobic degradation ofalpha-HCH reported here involves an initial reductive de-chlorination or a dehydrochlorination, as indicated by thetraces of pentachlorocyclohexene, is not yet known.Under sulfate-reducing and denitrifying conditions, no
significant alpha-HCH bioconversion was observed. Thedata are consistent with those reported previously (2, 9, 24).By assuming that reductive dechlorination is the first step in
s
50
'lxI'4
! 35050-
time (days)
FIG. 5. Conversion of alpha-HCH by the native, aerobic micro-bial population present in contaminated soil (O) and by the seedfrom activated sludge added to a sterile soil slurry (O). Utilization ofthe added dissolved organic carbon (DOC) with time by the nativesoil microbial population (+) and seed from activated sludge thatwas added to sterile soil slurry (x).
300
250_x
E
2000
c 150r0U 100
C 50
20 60
time (days)
80 100
FIG. 6. Conversion of beta-HCH as a function of time underdifferent redox conditions, i.e., methanogenic (O), sulfate reducing(x), denitrifying (A), and aerobic (0). The slurries which containedthe native microbial population were seeded with additional micro-organisms (0.7 g [dry weight]/liter) from various sources: anaerobicsewage sludge (O), anaerobic marine sediment (x), and activatedsludge (A and 0). No significant beta-HCH bioconversion was
measurable when the data were compared with those for therespective sterile control. For clarity, data for the sterile controlsare not included.
anaerobic HCH degradation, it can be speculated that thisprocess allows for certain anaerobes to regenerate theirprimary electron acceptors (e.g., NADH). As a conse-
quence, reductive dechlorination would have to competewith nitrate and sulfate for electrons. Following the conceptof electron free energy, the terminal electron acceptor whichis energetically most favorable under a given environmentalcondition is the first to be used by a mixed population.Thermodynamic calculations (equation 1) indicate a largenegative change in Gibbs free energy (AG) of about -110 kJper mol per electron for the reductive dechlorination ofdichloroethane to ethene (25) (H2 electron donor; chlorideconcentration, 10'- M; pH 7). For reductive dechlorinationof the ring alkane HCH to tetrachlorocyclohexene, a some-
what higher, but similar, G value would be expected. Di-chloroethane was chosen as the model compound becauseno thermochemical data for HCH are published in the Tablesof the National Bureau of Standards.
Dichloroethane + H2 = ethene + 2Cl- + 2H+ (1)AG' = -110 kJ per mol per electron
An electron free energy of -110 kJ per mol per electron is byfar more than that released during sulfate reduction (AG' =
-19 kJ per mol per electron) (25) and is similar to thatreleased during denitrification (AG' = -112 kJ per mol perelectron) (25). Therefore, an inhibition of the reductivedechlorination by nitrate based on pure thermodynamicmechanisms could occur. On the other hand, an inhibition ofthe reductive dechlorination by sulfate based on pure ther-modynamic mechanisms seems not to be very likely. Kineticreasons or biochemical mechanisms (direct inhibition on theenzyme), however, cannot be excluded to explain the ob-served alpha-HCH recalcitrance in the presence of nitrate or
sulfate. Nevertheless, our findings still lead to questions withregard to the fate of HCH in anoxic marine environments. Itremains to be determined whether our findings can begenerally extrapolated to such sulfate-rich habitats.Beta-HCH has been shown to be remarkably recalcitrant
under all four redox conditions studied during the 100 daysof the experiments. The reason for this recalcitrance is notcurrently understood. Specific spatial chloride arrangementsmight have been responsible. In addition, very slow rates ofbeta-HCH biodegradation could not be detected within the
complex system that was investigated. Conflicting biocon-
002
200i
20~~~~~~~~~
0 0
00 5 10 15 205
VOL. 54, 1988
a..o
0
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

148 BACHMANN ET AL.
version statements have been found in the literature. Insome studies it has been claimed that beta-HCH is biocon-vertible (9, 14, 15), and in others it has been claimed that itis not, even at low concentrations (19).The buildup of intermediates from the bioconversion of
alpha-HCH under methanogenic conditions and the apparentrecalcitrance of beta-HCH under any of the four redoxconditions tested has important environmental and healthimplications. Because of the prevailing high groundwaterlevels in The Netherlands and the higher mobilities ofintermediates such as chlorinated benzenes and chlorinatedphenols, compared with that of HCH, care must be taken toprevent anaerobiosis in HCH-contaminated soils. Becauseof the recalcitrance of beta-HCH, the possibility of ground-water contamination by this chemical cannot be ignored,despite its rather hydrophobic character (log Pow > 4). Littleis known about the ecotoxicological significance of beta-HCH. Results of the few studies that are available show anincrease in enzyme activities in rat liver, effects on lymphoidand endocrine organs, neurological signs leading to death, aswell as several effects that are indicative of an estrogenicaction with doses of beta-HCH from 2 to 250 mg/kg of feed(29). For humans, by assuming an average weight of 70 kg,50% lethal doses of 35, 420, and 9 g for alpha-, beta-, andgamma-HCH, respectively, have been reported (10). Eventhough beta-HCH is less toxic than the other isomers, it stillappears to represent a serious health hazard.From an engineering standpoint, aerobic conditions have
been found to be the most feasible for cleaning up soilcontaminated with alpha-HCH. It appears that for the bio-degradation of this isomer, only sufficient soil homogeneity,water, oxygen, and nutrients are needed for complete min-eralization. With respect to the potential of biotechnologicalprocesses to clean up soils contaminated with beta-HCH, westill have reservations. More work is needed in this area. Forsuccessful biotechnological application of aerobic alpha-HCH biodegradation, more information is needed on theoptimum environmental conditions of both the overall soilenvironment, as well as on the microorganisms that arepresent in the soil.
ACKNOWLEDGMENTS
We thank M. A. Posthumus, Department of Organic Chemistry,for gas chromatographic, mass spectrometric analyses.
This work was supported by a grant from the Dutch Ministry ofHousing, Physical Planning and Environment.
LITERATURE CITED
1. Engst, T., R. M. Macholz, and M. Kujawa. 1974. Metabolismusdes Lindan. Abbau von Lindan durch Schimmelpilzkulturen.Nahrung. 18:737-745.
2. Gibson, S. A., and J. M. Suflita. 1986. Extrapolation of biodeg-radation results to groundwater aquifers: reductive dehalogena-tion of aromatic compounds. Appl. Environ. Microbiol. 52:681-688.
3. Haider, K. 1979. Degradation and metabolization of lindane andother hexachlorocyclohexane isomers by anaerobic and aerobicsoil microorganisms. Z. Naturforsch. Teil C 34:1066-1069.
4. Haider, K., and G. Jagnow. 1975. Degradation of 14C-, 3H-, and36C1-labelled gamma-hexachlorocyclohexane by anaerobic soilmicroorganisms. Arch. Microbiol. 104:113-121.
5. Haider, K., G. Jagnow, R. Kohnen, and S. U. Lim. 1974.Degradation of chlorinated benzenes, phenols and cyclohexanederivatives by benzene and phenol utilizing soil bacteria underaerobic conditions. Arch. Microbiol. 96:183-200.
6. Heritage, A. D., and I. C. MacRae. 1977. Identification of
intermediates formed during the degradation of hexachlorocy-clohexanes by Clostridium sphenoides. Appl. Environ. Micro-biol. 33:1295-1297.
7. Heritage, A. D., and I. C. MacRae. 1977. Degradation of lindaneby cell-free preparations of Clostridium sphenoides. Appl. En-viron. Microbiol. 34:222-224.
8. Hill, D. W., and P. L. McCarty. 1967. Anaerobic degradation ofselected chlorinated hydrocarbon pesticides. J. Water Pollut.Control Fed. 39:1259-1277.
9. Jagnow, G., K. Haider, and P.-C. Ellwardt. 1977. Anaerobicdechlorination and degradation of hexachlorocyclohexane iso-mers by anaerobic and facultative anaerobic bacteria. Arch.Microbiol. 115:285-292.
10. Kleter, G. 1983. HCH contaminations in Zwolle. In Staatstoe-zicht op de Volksgezondheid, Zwolle. (In Dutch.)
11. Koster, I. W., and H. J. Brons. 1984. Respirometric testingmethod for biodegradability of xenobiotics using compost. En-viron. Sci. Health B19:785-792.
12. Machholz, R. M., and M. Kujawa. 1985. Recent state of lindanemetabolism. Part III. Res. Rev. 94:119-149.
13. MacRae, I. C., K. Raghu, and E. M. Bautista. 1969. Anaerobicdegradation of the insecticide lindane by Clostridium sp. Nature(London) 221:859-860.
14. MacRae, I. C., K. Raghu, and T. F. Castro. 1967. Persistenceand biodegradation of four common isomers of benzene hexa-chloride in submerged soils. J. Agric. Food Chem. 15:911-914.
15. MacRae, I. C., Yamaya, and T. Yoshida. 1984. Persistence ofhexachlorocyclohexane isomers in soil suspensions. Soil Biol.Biochem. 16:285-286.
16. Ohisa, N., N. Kurihara, and M. Nakajiama. 1982. ATP synthesisassociated with the conversion of hexachlorocyclohexane re-lated compounds. Arch. Microbiol. 131:330-333.
17. Ohisa, N., and M. Yamaguchi. 1978. Gamma-BHC degradationaccompanied by the growth of Clostridium rectum isolated frompaddy field soil. Agric. Biol. Chem. 42:1819-1823.
18. Ohisa, N., and M. Yamaguchi. 1978. Degradation of gamma-BHC in flooded soil enriched with peptone. Agric. Biol. Chem.42:1983-1987.
19. Ohisa, N., and M. Yamaguchi. 1979. Clostridium species andgamma-BHC degradation in paddy soil. Soil Biol. Biochem.11:645-649.
20. Ohisa, N., M. Yamaguchi, and N. Kurihara. 1980. Lindanedegradation by cell-free extracts of Clostridium rectum. Arch.Microbiol. 125:221-225.
21. Owen, W. F., D. C. Stuckey, J. B. Healy, Jr., L. Y. Young, andP. L. McCarty. 1979. Bioassay for monitoring biochemicalmethane potential and anaerobic toxicity. Water Res. 13:485-492.
22. Postgate, J. R. 1969. Viable counts and viability, p. 611-628. InJ. F. R. Norris and D. W. Ribbons (ed.), Methods in microbi-ology. Academic Press, Inc., London.
23. Rosswall, T. 1981. The biogeochemical nitrogen cycle. In G. E.Likens (ed.), Some perspectives of the major biogeochemicalcycles. John Wiley & Sons, Inc., New York.
24. Sethunaton, N., E. Baustida, and T. Yoshida. 1969. Degradationof benzenehexachloride by soil bacteria. Can. J. Microbiol.15:1349-1354.
25. Stumm, W., and J. J. Morgan. 1981. Aquatic chemistry, 2nded., p. 58-68. John Wiley & Sons, Inc., New York.
26. Sweeney, R. A. 1969. Metabolism of lindane of unicellular algae.Proc. Conf. Great Lakes Res. 12:98-102.
27. Trueper, H. G., and H. G. Schlegel. 1964. Sulphur metabolism inthiorhodaceae. I. Quantitative measurements on growing cellsof Chromatium okenii. Antonie van Leeuwenhoek 30:225-238.
28. Tu, C. M. 1976. Utilization and degradation of lindane by soilmicroorganisms. Arch. Microbiol. 108:259-263.
29. Van Velsen, F. L., L. H. J. C. Danze, F. X. R. van Leeuwen,J. A. M. A. Dormans, and M. J. van Logten. 1986. The subchro-nic toxicity of the beta-isomer of hexachlorocyclohexane inrats. Fund. Appl. Toxic. 6:697-712.
30. Wedemeyer, G. 1966. Dechlorination of DDT by Aerobacteraerogenes. Science 152:647.
31. Yoshida, T., and T. F. Castro. 1970. Degradation of gamma-
APPL. ENVIRON. MICROBIOL.
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

HCH BIOCONVERSION IN SOIL SLURRY
BHC in rice soils. Soil Sci. Soc. Am. Proc. 34:440-442.32. Yoshinari, T., R. Hynes, and R. Knowles. 1977. Acetylene
inhibition of nitrous oxide reduction and measurement of deni-trification and nitrogen fixation in soil. Soil Biol. Biochem. 9:177-183.
33. Zehnder, A. J. B., and T. D. Brock. 1980. Anaerobic methane
oxidation: occurrence and ecology. Appl. Environ. Microbiol.39:194-204.
34. Zvyagintsev, D. G., S. A. Lukin, G. A. Lisichkina, P. A.Kozhevin, and M. L. Lomonosov. 1984. Method for more accu-rate quantitative determination of soil microorganisms. Mikro-biologiya 53:665-668. (Translated from Russian.)
VOL. 54, 1988 149
on July 9, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
















![Prediction of Pavement Surface Skid Resistance [Read-Only]€¦ · measured skid resistance with ti tftESRhb 3) Both the greywacke and basalt aggregate demonstrated a taminants after](https://img.dokumen.tips/doc/110x75/6060cf662242d63a9507c8d2/prediction-of-pavement-surface-skid-resistance-read-only-measured-skid-resistance.jpg)












