Embed Size (px)
Citation preview

Biochemistry
Sheng Zhao ( 赵晟 ), Biochemistry and Molecular Department of Medical school in Southeast University
Web: http://teaching.ewindup.info/
Email: [email protected] or [email protected]
QQ /MSN/Skype/gChat: [email protected]
Mobile:18551669724 or 13675130010
Conception, theory, research, and application——Logic and LIY (Learn It Yourself)

Chapter 3: Enzyme
Thermodynamics and Kinetics


Section One
The Molecular Structure and Function of Enzyme

Enzymes /ˈɛnzaɪmz/ are large biological molecules responsible for the thousands of chemical interconversions that sustain life.
What is a Enzyme ?

Etymology and history• In 1833, French chemist Anselme Payen discovered the
first enzyme, diastase. • In 1877, German physiologist Wilhelm Kühne (1837–
1900) first used the term enzyme, which comes from Greek ενζυμον, “in leaven” (in yeast).
• In 1897, Eduard Buchner discovered cell-free fermentation "zymase". (1907 Nobel Prize in Chemistry)
• In 1926, James B. Sumner showed that the enzyme urease was a pure protein and crystallized it (1946 Nobel Prize in Chemistry).
• In 1965, the first X-ray 3D structure of enzyme was done for lysozyme by David Chilton Phillips’ group.
• In 1989, Thomas R. Cech and Sidney Altman won the Nobel Prize in chemistry for their "discovery of catalytic properties of RNA.“ in 1980s.

Substrates
Enzyme
Product
Enzymatic reactions1. In enzymatic reactions, the molecules at the beginning of the
process, called substrates, are converted into different molecules, called products.
2. Enzymes are used commercially, for example, in the synthesis of antibiotics, in biological washing powders break down protein or fat stains on clothes, and in food industry to break down proteins.

Enzymes catalyze the conversion of substrates into products
• What is a substrate?– A substrate is the compound that is converted into
the product in an enzyme catalyzed reaction.– For the reaction catalyzed by aldolase, fructose
1,6-phosphate is the substrate.
Fructose 1,6-phosphate
Glyceraldehyde3-phosphate
Dihydroxyacetone phosphate
+aldolase
Substrate Products

What are enzymes made from?
Protein
1. All enzymes are proteins except some RNAs (Ribozymes) and DNAs (deoxyribozymes)
2. not all proteins are enzymes
NucleicacidEnzyme

Ribozymes
It was assumed that all enzymes are proteins until 1982 when Thomas Cech and Sydney Altman discovered catalytic RNAs (Nobel, 1989 in Chemistry);
The RNA world hypothesis, if true, has important implications for the definition of life, places RNA at center-stage when life originated.

What do enzymes do?• Enzymes are biological catalysts that
accelerate the rates of chemical reactions.
Snail without enzyme catalyst Snail with enzyme catalyst

Enzyme catalyzed reactions are much faster than non-catalytic reactions
Time
[Pro
du
ct]
Enzyme catalyzed reaction
Non-catalytic reaction
00
Reaction Rate =D[Product]
D(time)

General Properties of Enzymes
1. Higher reaction rates :• carbonic anhydrase• CO2 +H2O H2CO3
• nonenzymatic rate constant = 1.3 x 10-1 s-1
• enzymatic rate constant = 1 x 106 s-1 (x 7.7 x 106)
• Staphylococcal nuclease• nonenzymatic rate constant = 1.7 x 10-13 s-1
• enzymatic rate constant = 95 s-1 (x 5.6 x 1014)

Enzymes do more than just increase the rate of a chemical reaction
Control when and where reactions occurRegulate the rate of a reaction
(“controlled combustion”)
Prevent unwanted side reactionsOptimize reaction for specific conditions
(e.g. pH, temperature)

Some definitions of the enzyme
Monomeric enzyme: only one polypeptide chain in
which the active site resides.
Oligomeric enzyme : More than one polypeptide /
subunits 。 Multienzyme system : System of two or more enzymes
functioning sequentially to catalyze the reactions
Multifunctional enzyme or tandem enzyme : Enzyme
with more than one catalytic activity
Isozymes or isoenzymes or multiple forms of enzymes:
enzymes that differ in amino acid sequence but
catalyze the same chemical reaction.

Isozymes: An automotive analogy
1. Physically distinct forms of the same enzyme.2. Differ in amino acid sequences or posttranslational
modifications3. Different tissues or subcellular organelles

Example: Lactate Dehydrogenase is composed of four monomers
Homo or hetero tetramers composed of muscle (M) and heart (H) protein subunits encoded by the LDHA and LDHB genes, respectively:
LDHx—A third isoform, coded by LDHC or LDHX gene, is expressed only in the testis
LDH-1 (4H)—in the heart and in RBC (red blood cells) LDH-2 (3H1M)—in the reticuloendothelial system LDH-3 (2H2M)—in the lungs LDH-4 (1H3M)—in the kidneys, placenta, and pancreas LDH-5 (4M)—in the liver and striated muscle
H H
H H
H H
H M
H H
MM
H
MM
M
MM
MM
LDH1
(H4)LDH2
(H3M) LDH3
(H2M2)LDH4
(HM3)LDH5
(M4)

Components of the enzyme
Apoenzyme: the protein part of an enzyme
Cofactor
Inorganic: Metal ions
Organic
Holoenzyme
Simple enzyme Conjugated enzyme
Prosthetic group: emphasizes the nature of the binding of a cofactor to a protein (tight or covalent)
Coenzymes: additional substance required for enzyme activity

Active site of Enzyme
• Definition: The small 3D groove or pocket of an enzyme where substrate molecules bind and undergo a chemical reaction.
• The active site displays highly specific substrate binding
• The active site is responsible for whether there is ordered or random binding of substrates and release of products

The active site of the enzyme
Active site ofChymotrypsin(a digestive enzyme component of pancreatic juice acting in the duodenum where it performs proteolysis, the breakdown of proteins and polypeptides.)

Important sites of Enzyme
• Active site residues: residues that directly participate in the catalytic reaction mechanism in active site of enzyme.
• Essential groups: Side chain groups required for the catalytic ability of enzymes Inside active site: for reaction
binding group catalytic group
Outside active site: for conformation

Substrate
Outside the active site
Binding group
Catalytic groups
Active site


Specificity of Enzyme Absolute specificity: specific substrate
Relative specificity : a class of bond or group
Stereospecificity : specific stereoisomer

Enzyme active sites can distinguish between stereoisomers
H
CCO2-
CH3
NH2
H
C
CH3
CO2-
NH2 L-Alanine D-Alanine
Stereoisomers are non-superimposable mirror images of each other

.
L-AlanineAminotransferase
H
CCO2-
CH3
NH2D-Alanine
L-Alanine
H
C CO2-
CH3
NH2
Enzymes contain binding sites that recognize the substrates of the reaction

Enzyme Nomenclature and Classes
Oxidoreductases (EC Class 1)Transfer electrons (RedOx reactions)
Transferases (EC Class 2)Transfer functional groups between molecules
Hydrolases (EC Class 3)Break bonds by adding H2O
Lyases (EC Class 4)Elimination reactions to form double bonds
Isomerases (EC Class 5)Intramolecular rearangements
Synthetase or Ligases (EC Class 6)Join molecules with new bonds

Oxidoreductases catalyze the transfer of hydrogen atoms and electrons
Example - Lactate Dehydrogenase
O O-
C
C
CH3
O + NADH + H+
O O-
CH
C
CH3
HO + NAD+
pyruvate L-lactate
lactatedehydrogenase

Transferases catalyze the transfer of functional groups from donors to acceptors
Example - Alanine aminotransferase
O O-
C
C
CH3
O
pyruvate
O O-
CH
C
CH2
H2N
CH2
O O-C
glutamate
O O-
CH
C
CH3
H2N
L-alanine
O O-
C
C
CH2
CH2
O O-C
O
a-ketoglutarate
+ +
alanineaminotransferase

Hydrolases catalyze the cleavage of bonds by the addition of water (hydrolysis)
Example - Trypsin
NH
C
NH2(CH2)4H
O
C
O
HH
CNH
NH
C
CH3H
O
C
O
H
CNH
CC
CH(CH3)2
NH
C
NH2(CH2)4H
O
C
O
HH
CNH
O -C
NH
C
CH3H
O
C
O
CH(CH3)2H
CH 3N
+ C
+ H2O
trypsin
+
Gly-Lys-Val-Ala
Val-Ala
Gly-Lys

Lyases catalyze the cleavage of C-C, C-O, or C-N bonds(addition of groups to double bonds or formation of double bonds by removal of groups)
Example - ATP-citrate lyase
O O-
C
C
CH2
O
O O-C
oxaloacetate
O O-
CH2
C
CH2
O O-C
CO
O-
CHO
citrate
O CH3C
O-
acetate
ATP-citrate lyase
+
ATP ADP + PiCoenzyme A

Isomerases catalyze the transfer of functional groups within the same molecule
Example - Phosphoglucose isomerase
O H
HC
C
CHHO
HC
O OPO32-
C
OH
HC OH
OH
glucose 6-phosphate
C
CH2OH
CHHO
HC
C
O
HC OH
OH
OPO32-
O
fructose 6-phosphate
phosphoglucose isomerase

Ligases use ATP to catalyze the formation of new covalent bonds
Example - DNA ligase
OP-O
O
O-
HO
3'
5' 3'
5'
T G C A GT CG
O P O-
O
O-
OH3'
3'
5'
GCT CACGA
5'
+O P
O
O O
O
O-
P
O O-
G CTGACGT
5'
3'5'
3'
A G CA CT C G
5'
3'
3'
5'
DNA ligase
ATP ADP + Pi

Substrate
+
Enzyme ES complex
Enzyme
Substrate
+
ES complex
Lock and Key - Emil Fischer (1890)
Induced Fit - Daniel E. Koshland Jr. (1958)

An Example: Induced conformational change in hexokinaseAn Example: Induced conformational change in hexokinase
Catalyzes phosphorylation of glucose to glucose 6-phosphate during glycolysis
such a large change in a protein’s conformation is not unusual
BUT: not all enzymes undergo such large changes in conformation

Advantage of the induced fit mechanism The active site can be open to allow
substrates to bind, then close over the substrates to provide optimum transition state stabilization
Disadvantage of the induced fit mechanism Energy that would otherwise be
used to help stabilize the transition state of the reaction must be used to induce the conformational change in the enzyme.

Greater capacity for regulation of Enzymes
• Enzymes can respond to allosteric compounds that alter their kinetic properties.
• Enzymes can also be regulated by covalent modification : e.g. phosphorylation can inactivate or activate an enzyme.
• Through increased transcription of the gene encoding a particular enzyme, the level of mRNA for the enzyme can increased. This can increase levels of this enzyme.

Important things to remember about enzymes(just like other catalysts)
1. Enzymes are not consumed or altered by the reaction they catalyze.
Just as a construction worker can take a pile of lumber and build a home without being physically changed by the process, an enzyme takes substrates and converts them into products without being physically changed or consumed.

This is an important point. An enzyme does not determine which direction the reaction goes, it only increases the rate at which the reaction approaches equilibrium.
2. Enzymes catalyze both the forward and the reverse reaction.

3. Enzymes do not alter the equilibrium (or equilibrium constant) between substrates and products.
At equilibrium, the ratio of substrates to products is the same regardless of whether an enzyme catalyst is present. Although the final equilibrium ratio of reactants to products is not altered by an enzyme, the rate at equilibrium is achieved is increased.
Glucose +ATP Glucose 6-phosphate + ADP
HexokinaseGlucose +ATP Glucose 6-phosphate + ADP
Keq = 103
Keq = 103

G < 0 for the conversion of diamond into graphite
Thermodynamics vs. Kinetics• Thermodynamics tells us whether a reaction
can occur.• Kinetics tells us whether the reaction will
occur in our lifetimes.

Section two: Thermodynamics and Kinetics of Enzyme-Catalyzed Reaction
Yes/No?
Now/Later?

What is the transition state?
The transition state is the most unstable species on the reaction coordinate (i.e. the species with the highest energy)
Substrates
Products
Transition State
DGrxnFre
e E
ner
gy
(DG
)
Reaction Coordinate

Transition State TheoryReactants are required to reach a high-energy
(unstable) state referred to as the transition state. Residence may last only 10-13 to 10-14s
Only a limited number of molecules will possess sufficient energy to reach this transition state .
Increasing temperature or other conditions may facilitate reactants reaching the transition state

The transition state is not an intermediate species
The transition state cannot be trapped or isolated. Intermediates can be trapped or isolated.
Products
Transition State
Fre
e E
ne
rgy
(DG
)
Reaction Coordinate
SubstratesIntermediate

The Transition StateUnderstand the difference between G
and G‡ (Gibbs energy)The overall free energy change for a
reaction (∆G) is related to the equilibrium ratio of [S] and [P]
The free energy of activation for a reaction (G‡) is related to the reaction rate
It is extremely important to appreciate this distinction!

Free energy tells us nothing about the rate at which the reaction occurs
The height of the transition state relative to the reactants determines how rapidly substrates are converted to product
Substrates
Products
Transition State
DGrxnFre
e E
ner
gy
(DG
)
Reaction Coordinate
With EnzymeG‡

General Properties of Enzymes
Enzymes bind substrates to their active site and stabilize the transition state of the reaction.

Enzyme Kinetics• The rate of unimolecular reaction is proportional
to the concentration of the reactant. Thus rate is linearly dependent on [A].
• But if this reaction is catalyzed by an enzyme, the rate shows saturation behavior. Why?
PA ][][
Akdt
Adv
v
[A]
PA Enzyme v
[A]

[S]
VVmax
[S]
VVmax
[S]
VVmax
Early stage
Middle stage
Saturated stage

Leonor Michaelis (1875-1949) Maud Menten (1879-1960)

The Michaelis–Menten Equation
You’d better know how this is derived
This is the complete chemical formula for an enzyme-catalyzed (E) reaction of substrate, S and product, P;
Michaelis–Menten equation describes the relationship between reaction rate and substrate concentration.
Assumptions for Mechaelis-Menten:– Initial velocity assumption– Rate law– State steady assumption
PEESSE k1
K-1
k2
K-2

Initial Velocity Assumption
• In the beginning of the reaction, there is very little product, or [P] is small. So the amount of [ES] contributed by E+P is negligible.
• Thus, the MM equation concerns the reaction rate that is measured during early reaction period.
• In which case, the enzyme catalyzed reaction can be simplified to:
PEESSE k1
K-1
k2
K-1
PEESSE k1 k2
K-2

Rate Law and Steady state in Enzyme Catalyzed Reactions
Rate law still applies in enzyme catalyzed reactions. The forward velocity, or rate, vf is,
The reverse velocity or rate, or the rate of disappearance vd is,
At steady state, there is no accumulation of [ES], thus:
PEESSE k1
K-1
k2
SEkv f 1
ESkkESkESkvd )( 2121
df vv

Derivation of Michaelis-Menten Equation
• We need one more condition, that is, the total enzyme concentration, [Et] is the sum of that of enzyme-substrate complex, [ES], and that of free enzyme, [E]:
• At steady state, the forward rate should equal to the reverse rate:
• Rate of production formation (rate law), v = k2[ES]. So:
m
ttt
tt
tt
KSSE
ES
kkkS
SEkkSk
SEkES
kkSkESSEkESkkSESkEk
ESkkSESkSEkESkkSESEk
1
21211
1
21112111
2111211
)())((
))(()(
)()()(
EESEt SESEkSEkv tf )(11
m
t
KS
SEkv
2
df vv
ESkkESkESkvd )( 2121

Understanding Km
• Km is a constant derived from rate constants
• Km is, under true Michaelis-Menten conditions, an estimate of the dissociation constant of E from S, because
at equilibrium,
Reversible reaction, dissociation constant is
So small Km means tight substrate binding; high Km means weak substrate binding.
• Km equals to the substrate concentration at which v=vmax/2
1
21
k
kkK m
ESSE k1
k-1
1
1
][
][
k
k
ES
SEK d
][][ 11 ESkSEk

Notes on the MM Equations
• The rate of production formation can usually be measured experimentally by monitoring the progress curve of production formation.
• The maximum rate can be reached at saturating substrate concentration, or when [S]
So MM equation can be re-written as:
mKS
Svv
max
max2][
2][ 1
vEkEk
v tS
Kt
Sm
Enzyme-catalyzed rate is saturated

Understanding Vmax
The theoretical maximal velocity
Vmax is a constant
Vmax is the theoretical maximal rate of the reaction - but it is NEVER achieved in reality
To reach Vmax would require that ALL enzyme molecules are tightly bound with substrate
Vmax is asymptotically approached as substrate is increased

The dual nature of the Michaelis-Menten equation The Michaelis-Menten equation describes a rectangular
hyperbolic dependence of v on S!

The turnover number
A measure of catalytic activity
kcat, the turnover number, is the number of substrate molecules converted to product per enzyme molecule per unit of time, when E is saturated with substrate.
If the M-M model fits, k2 = kcat = Vmax/Et
Values of kcat range from less than 1/sec to many millions per sec

The catalytic efficiencyName for kcat/Km
An estimate of "how perfect" the enzyme is kcat/Km is an apparent second-order rate constant
It measures how the enzyme performs when S is low
The upper limit for kcat/Km is the diffusion limit -
the rate at which E and S diffuse together

1
V= 1
VMAX
+
KM
VMAX
•1
[S]y = m x + b
x intercept = - 1 / KM
0 1 / [S]
1 / V
y intercept = 1 / VMAX
Determining the VMAX and KM : Lineweaver-Burk plot
Take reciprocal for both sides
Denominators !

Enzyme catalyzed reactions are optimized for specific values of
temperature and pH
.
5 6 7 8 9pH
Rea
ctio
n R
ate
Enzyme catalyzed reaction
Non-catalytic reaction
.
20 30 40 50 60Temperature (oC)
Rea
ctio
n R
ate
Enzyme catalyzed reaction
non-catalytic reaction

Enzyme Inhibition Reversible inhibitors associate with enzymes through non-
covalent interactions. Reversible inhibitors include three kinds: Competitive inhibitors; Non-competitive inhibitors; Un-competitive inhibitors
Irreversible inhibitors associate with enzymes through covalent interactions. Thus the consequences of irreversible inhibitors is to decrease in the concentration of active enzymes.

Competitve Inhibitors
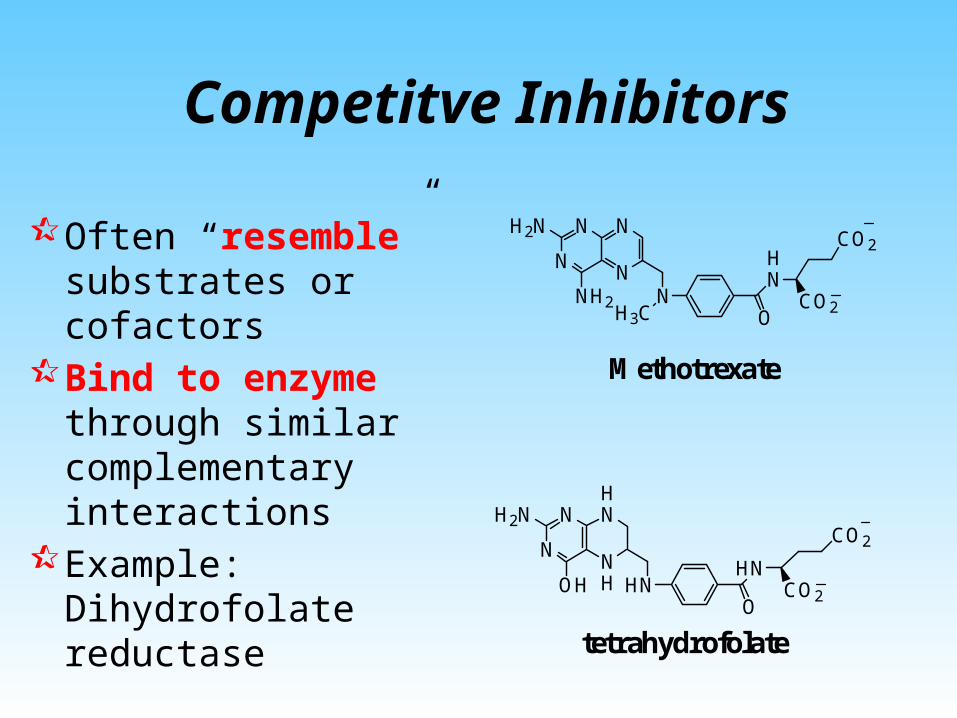
Competitve Inhibitors
Often “resemble” substrates or cofactors
Bind to enzyme through similar complementary interactions
Example: Dihydrofolate reductase
N
NN
N
NH3C
NH2
H2N
HN
OCO2
NH
HNN
N
HNOH
H2N
HN
CO2
OCO2
CO2
Methotrexate
tetrahydrofolate

Competitive Inhibitors
)][
1(][
][max
Im K
IKS
Svv
v
[S]
vmax
Km1/[S]
1/v
1/vmax-1/Km
Slope=Km/vmax
PEESSE k1
K-1
k2
EI
I
KI
Km increasesvmax unchanged
+inhibitor
Km(1+[I]/KI)
-1/(Km(1+[I]/KI))
Slope= Km(1+[I]/KI)/vmax
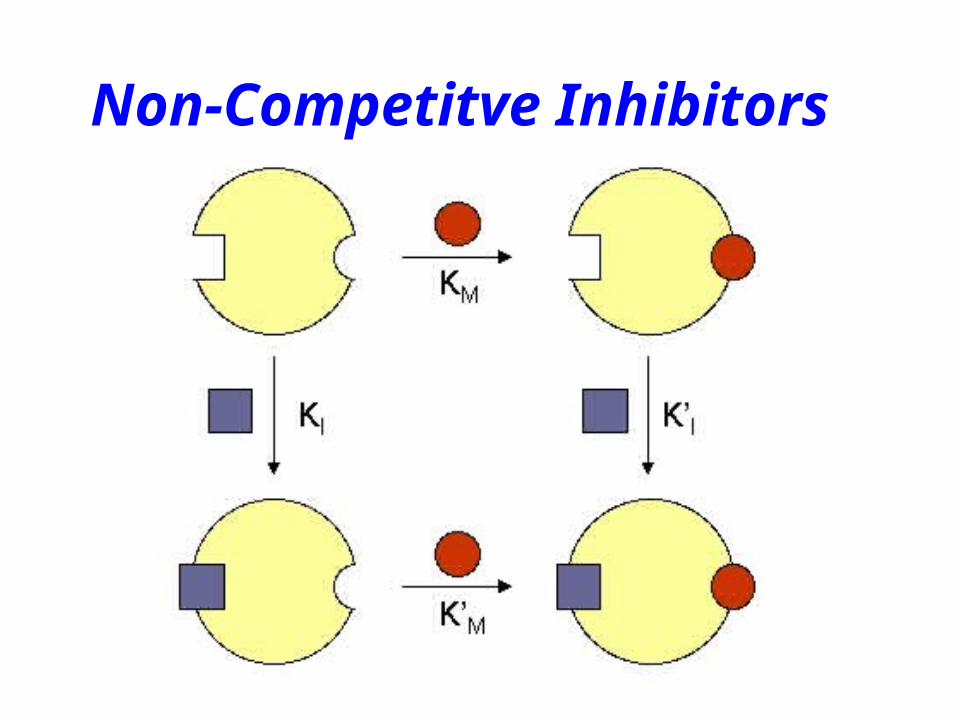
Non-Competitve Inhibitors

Noncompetitive Inhibitors
)][
1(][
][ max
I
m
K
Iv
KS
Sv
PEESSE k1
k-1
k2
EI
I
KI
Km unchangedvmax decreases
I
EISKI’
+inhibitor
1/[S]
1/v
1/vmax-1/Km
Slope=Km/vmax(1+[I]/KI)/VmaxSlope= Km(1+[I]/KI)/vmax
v
[S]
vmax
Km
Km
Vmax/(1+[I]/KI)
S

Uncompetitive Inhibitors
)][
1()
][1(
][
][ max
I
Kv
I
KK
S
Sv
I
I
m
PEESSE k1
k-1
k2
Km decreasesvmax decreasesSlope unchanged
+inhibitor
I
EISKI’
1/[S]
1/v
1/vmax-1/Km
Slope=Km/vmax(1+ KI/[I])/VmaxSlope= Km/vmax
v
[S]
vmax
KmKm/(1+ KI/[I])
Vmax/(1+KI/[I])
- (1+ KI/[I])/Km

Summary of Classes of reversible Inhibitors
• Competitive inhibition - inhibitor (I) binds only to E, not to ES • Noncompetitive inhibition - inhibitor (I) binds either to E
and/or to ES • Uncompetitive inhibition - inhibitor (I) binds only to ES, not to
E. This is a hypothetical case that has never been documented for a real enzyme, but which makes a useful contrast to competitive inhibition.
• Mixed inhibition-when the dissociation constants of (I) to E and ES are different. The inhibition is mixed.

Irreversible Inhibitor
Combines with or destroys an essential functional group on the enzyme (e.g. forms covalent bonds)
Inhibit enzymes irreversibly3 different types:
– Group Specific Reagent: • - inhibitor does not resemble substrate
– Substrate Analogue:• - inhibitor resembles substrate
– Suicide Inhibitors:• - inhibitor resembles substrate, turns "dangerous" after
processed by enzyme

E
S
S
As CH CHCl +
CH2 SH
CH SH
CH2 OH
E
SH
SH
+
CH2 S
CH S
CH2 OH
As CH CHCl
Cl
As
Cl
CH CHCl + E
SH
SHE
S
As
S
CH CHCl + 2HCl
Lewisite
Enzyme lost activity
desulfhydrase Enzyme lost activity
BAL desulfhydrase
Toxication and detoxication——Irreversible does not mean No cure!

Group Specific Reagent Does not resemble substrate
irreversibly inactivates enzyme by modifying an essential R group
e.g. DIPF (potent nerve gas) blocks acetylcholinesterase, which is essential for proper neural transmission
active site serine

Substrate Analogue
Binds at enzyme active siteThen irreversibly modifies (or binds to) to the active site
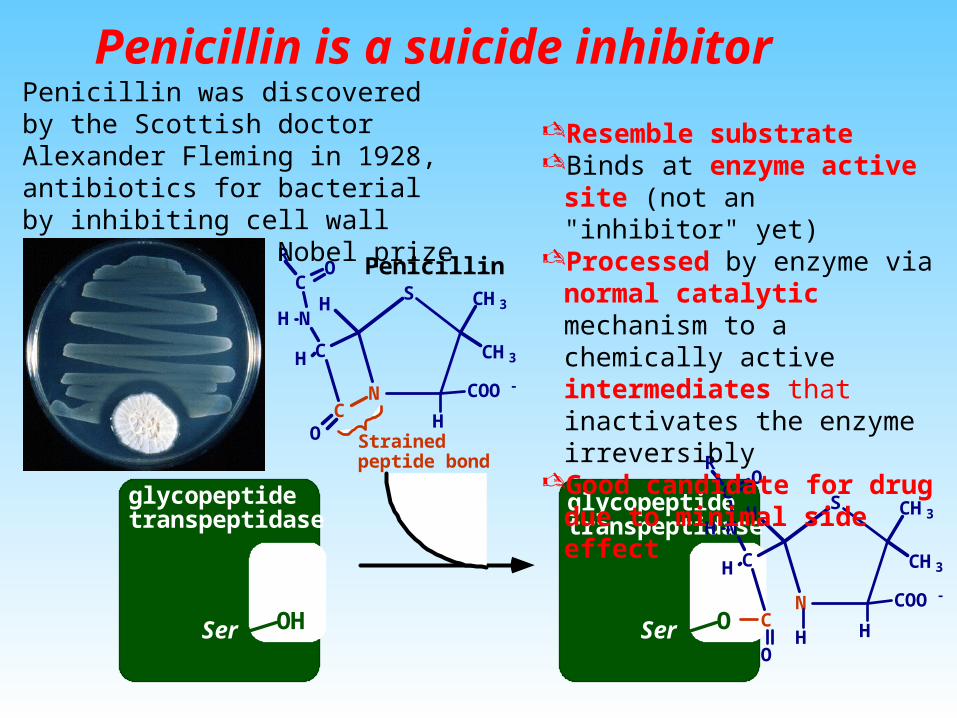
Penicillin is a suicide inhibitor
glycopeptidetranspeptidase
OHSer O
glycopeptidetranspeptidase
SerN
S CH3
CH3
COO-
H
H
C
C
O
H
N
CO
H
H
R
N
S
CH3
COO-
H
H
C
CO
H
N
CO
H
Strainedpeptide bond
PenicillinCH3
R
Resemble substrateBinds at enzyme active site (not
an "inhibitor" yet)Processed by enzyme via normal
catalytic mechanism to a chemically active intermediates that inactivates the enzyme irreversibly
Good candidate for drug due to minimal side effect
Penicillin was discovered by the Scottish doctor Alexander Fleming in 1928, antibiotics for bacterial by inhibiting cell wall formation. (the Nobel prize in Medicine).





























