Embed Size (px)
Citation preview

A n n a l s o f C l i n i c a l L a b o r a t o r y S c i e n c e , Vol. 3 , N o. 4 Copyright © 1973, Institute for Clinical Science
Biochemical Basis o f Hyaline Membrane Disease
DONALD T. FORMAN, Ph.D.
Division of Clinical Biochemisry, Evanston Hospital,Evanston, 1L 60201
andDepartment of Biochemistry and Pathology,
Northwestern University Medical School, Chicago, IL 60611
ABSTRACT
Pulmonary function in a normal fetus is dependent upon maturation and maintenance of the biochemical synthesis of a distinct surface-active lipid. Two major pathways for the biosynthesis of this major surface-active lipid, dipalmitoyl lecithin, are described and relative activities of each pathway in human lung are discussed. In normal human lung, the CDP-choline pathway is quantitatively most important for total lecithin synthesis. Infants with idiopathic respiratory distress syndrome (RD S) are usually prematurely bom. Their lungs lack adequate surface activity and are deficient in the principle surface-active component lecithin. Stress (hypoxia, acidosis, and hyperthermia) may induce respiratory distress in the fetus. Biochemical methods are available to detect and characterize pulmonary surfactant in amniotic fluid. Studies show that developmental changes in phospholipids in lungs are reflected by the phospholipids in amniotic fluid, and it is possible to predict the fetus whose lung is too prematurely developed for delivery and who may develop RDS.
In trod u ction
The survival of a newly born infant depends upon the development of a respiratory system capable of both pulmonary and cellular oxygenation and carbon dioxide excretion. During the development of this system, there must be both anatomical and biochemical maturation to allow for adequate lung function at birth. Fetal lung maturation serves as an excellent model for showing these important facets of pulmonary function.
In 1959, Avery and Mead1 reported that lung surface tension lowering material was
absent or markedly depleted in infants who died with hyaline membrane disease. Histological studies13 of human lung have shown that the presence of osmiophilic granules correlated well with the presence of surfactant. Both are usually found in fetuses and newborn infants of more than 29 weeks gestation. However, granules and surface tension lowering material are not found in infants dying from hyaline membrane disease (HMD) in the first few days of life. The role of surfactant deficiency in HMD remains controversial although such deficiency is probably responsible for the
2 4 2

B IO C H E M IC A L BA SIS O F H Y A L IN E M E M B R A N E D ISEA SE 2 4 3
progressive atelectasis and lack of alveolar stability found in HMD (figure 1).
B iosynthesis o f Pulm onary Surface- A ctive Lipid
Stability of lung depends upon a surface- active phospholipid rich complex which lines the pulmonary alveoli. This surfactant decreases the alveolar surface tension resulting in a decrease of their radii on expiration and stabilizing the fine air spaces of the lung.10 The unusual chemical, physical and physiologic properties of the principal pulmonary surface-active lipid, dipalmitoyl lecithin (D PL), have stimulated much research since the first description of its occurrence by Clements & coworkers in 1961.17
The basic component of the surfactant is a lipoprotein composed largely of surface active phospholipids and smaller amounts of neutral lipids and proteins. Dipalmitoyl- phosphatidylcholine is a major surface active component of surfactant and comprises about 50 percent of the total surfactant lipids.2 This dipalmitoyl lecithin (D PL) contains a phosphatidic acid structure in which the primary hydroxyl group of glycerol is esterified with phosphoric acid. This tertiary structure has a nitrogen base (choline) esterified to the hydroxyl group of phosphoric acid. Therefore, DPL contains an acidic hydroxyl group of phosphoric acid and a basic hydroxyl of choline and behaves as a zwitterion (internal salt) and has an isoelectric pH at 6.7. When different fatty acid components are substituted on the primary and secondary position of glycerol, lecithins of varying properties are derived. Lecithin containing stearic and oleic acids on the primary and secondary position of glycerol is different from lecithin containing palmitic and oleic acid substitutes. DPL is highly surface active owing to its bipolar chemical configuration, - a hydrophilic choline group and two hydrophobic saturated fatty acid
AERATION (LUNG)
*L IN IN G LAVER
FORMED AT'GAS/LIQUID INTERFACE
*SURFACTANT CONSUMPTION
*[s u r f a c t a n t d e f ic ie n c y ]
/ *ALVEOLAR COLLAPSE 1
iINTERSTITIAL EDEMA -*■ CELL INJURY
*INTERSTITIAL FLUID LEAKS INTO ALVEOLAR SPACE
[HYALINE MEMBRANE; FORMATION!
F ig u r e 1. Postulated chain of events resulting in surfactant deficiency and hyaline membrane formation.
side chains. It is remarkably effective in lowering the surface tension of aqueous solutions when chemically combined or adsorbed to proteins.
Dipalmitoyl lecithin (in contrast to total lung lecithin with acyl groups of variable length and unsaturation) may be synthesized in lung by random nonspecific acylation of a, B-diglyceride and subsequent condensation with cytidine dipho- sphocholine (CDP-choline)18 (figure 2 ). An alternate biosynthetic route has been proposed by Bremer and Greenberg8(figure 3). In both phosphatidyl dimethyl- ethanolamine and phosphatidyl choline (PC ), palmitate accounts for 65 to 80 percent of fatty acyl groups, and both lipids possess a remarkable ability to lower sur-
O ' Oc h 2 - o - c - r , CH2- 0 - C - R |I O I O1 n PHOSPHOCHOLiNE 1 h
C H - O - C - R 2 *C 0 P -C H 0 U N £ ---------------------- ► C H - 0 - C - R 2I TRANSFERASE j 0
CHsjOH CH20 -P - 0 -0 H 2 - C H 2N*(CHi)3D - a , R DIGLYCERIDE '
PHOSPHATIDYL CHOLINE (LECITHIN) ♦ CMP
Ri ond Rg » Dipolmiroyl' F ig u r e 2 . Enzymatic pathway f o r de novo
biosynthesis o f lecithin.
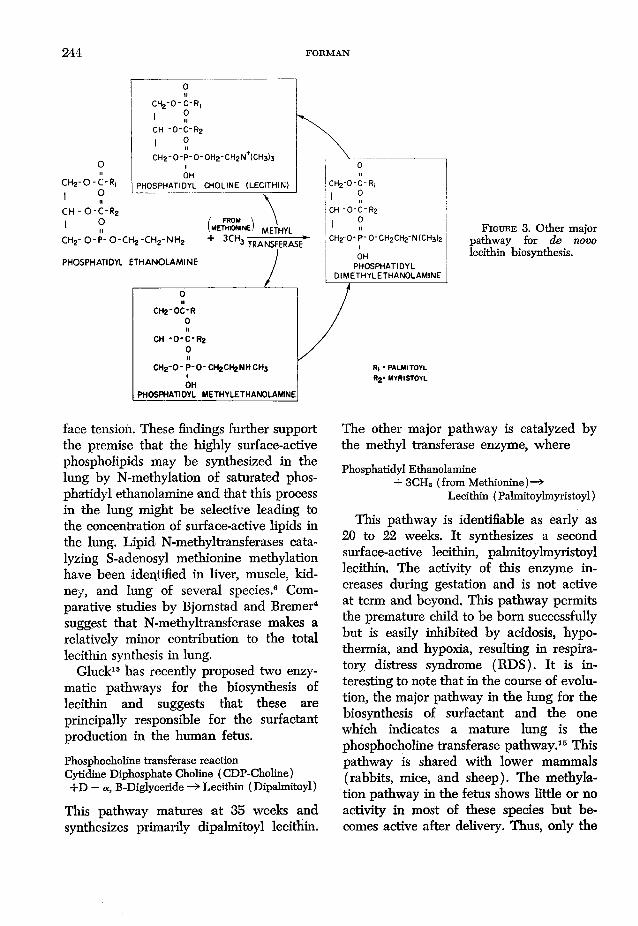
244 FORMAN
CHg-0 " C‘ R,
I 01 nCH -O-C-R2
1 °CH2-0-P-0-0H2-CH2N+{CH3)3
IOH
CH2- 0 - C - R | PHOSPHATIDYL CHOLINE (LECITHIN)I OCH - O -C-Rz I oII
c h 2- o -p - o - c h 2 - c h 2- n h 2
PHOSPHATIDYL ETHANO LAM INE
C H 2 -O C -R 0 11
CH - O - C - R 2 0 11
C H 2 - 0 - P - O - CH 2CH 2N H c h 3IOH
PH O SPH ATID YL M ETH YLETH A N O LAMINE
/ FRO M \I m e t h i o n i n e ) m e t h y l
Qfu ________3 TRANSFERASE
F i g u r e 3. Other major pathway for de novo lecithin biosynthesis.
face tension. These findings further support the premise that the highly surface-active phospholipids may be synthesized in the lung by N-methylation of saturated phosphatidyl ethanolamine and that this process in the lung might be selective leading to the concentration of surface-active lipids in the lung. Lipid N-methyltransferases catalyzing S-adenosyl methionine méthylation have been identified in liver, muscle, kidney, and lung of several species.6 Comparative studies by Bjomstad and Bremer4 suggest that N-methyltransferase makes a relatively minor contribution to the total lecithin synthesis in lung.
Gluck15 has recently proposed two enzymatic pathways for the biosynthesis of lecithin and suggests that these are principally responsible for the surfactant production in the human fetus.
Phosphocholine transferase reaction Cytidine Diphosphate Choline (CDP-Choline)+ D — a, B-Diglyceride —> Lecithin (Dipalmitoyl)
This pathway matures at 35 weeks and synthesizes primarily dipalmitoyl lecithin.
The other major pathway is catalyzed by the methyl transferase enzyme, where
Phosphatidyl Ethanolamine+ 3CHS (from Methionine)—*
Lecithin (Palmitoylmyristoyl)
This pathway is identifiable as early as 20 to 22 weeks. It synthesizes a second surface-active lecithin, palmitoylmyristoyl lecithin. The activity of this enzyme increases during gestation and is not active at term and beyond. This pathway permits the premature child to be bom successfully but is easily inhibited by acidosis, hypothermia, and hypoxia, resulting in respiratory distress syndrome (RDS). It is interesting to note that in the course of evolution, the major pathway in the lung for the biosynthesis of surfactant and the one which indicates a mature lung is the phosphocholine transferase pathway.15 This pathway is shared with lower mammals (rabbits, mice, and sheep). The methyla- tion pathway in the fetus shows little or no activity in most of these species but becomes active after delivery. Thus, only the
BIOCHEMICAL BASIS OF HYALINE MEMBRANE DISEASE 245
human and other primates can be bom prematurely. Maturation of the phospho- choline transferase system takes place in all humans and animals at about 90 percent of their gestation period.
E ffect o f Oxygen an d C arbon D ioxide T en sion on Sy n th esis o f S u r fa c ta n t
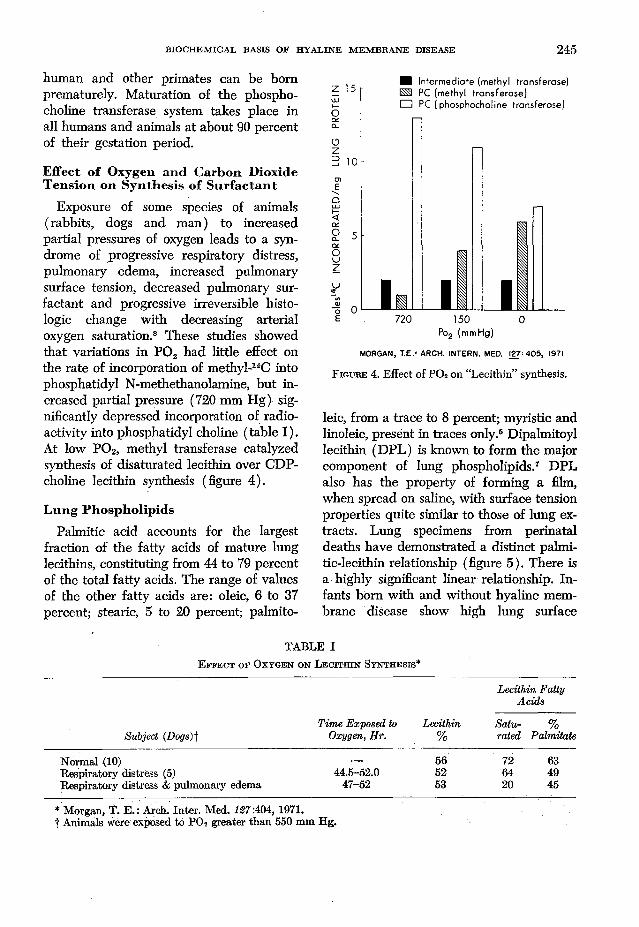
Exposure of some species of animals (rabbits, dogs and man) to increased partial pressures of oxygen leads to a syndrome of progressive respiratory distress, pulmonary edema, increased pulmonary surface tension, decreased pulmonary surfactant and progressive irreversible histologic change with decreasing arterial oxygen saturation.8 These studies showed that variations in P 02 had little effect on the rate of incorporation of methyl-14C into phosphatidyl N-methethanolamine, but increased partial pressure (720 mm Hg) significantly depressed incorporation of radioactivity into phosphatidyl choline (table I). At low P 02, methyl transferase catalyzed synthesis of disaturated lecithin over CDP- choline lecithin synthesis (figure 4).
L u n g P h osph olip ids
Palmitic acid accounts for the largest fraction of the fatty acids of mature lung lecithins, constituting from 44 to 79 percent of the total fatty acids. The range of values of the other fatty acids are: oleic, 6 to 37 percent; stearic, 5 to 20 percent; palmito-
Po2 (mmHg)
MORGAN, T.E.' ARCH. INTERN. MED. 127-405, 1971
F i g u r e 4. Effect of P 02 on “Lecithin” synthesis.
leic, from a trace to 8 percent; myristic and linoleic, present in traces only.® Dipalmitoyl lecithin (DPL) is known to form the major component of lung phospholipids.7 DPL also has the property of forming a film, when spread on saline, with surface tension properties quite similar to those of lung extracts. Lung specimens from perinatal deaths have demonstrated a distinct palmi- tic-lecithin relationship (figure 5). There is a highly significant linear relationship. Infants bom with and without hyaline membrane disease show high lung surface
TABLE IE f f e c t o f O x y g e n o n L e c i t h i n S y n t h e s i s *
Subject (Dogs)fTime Ex-posed to
Oxygen, Hr.Lecithin
%
Lecithin Fatty Acids
Satu- % rated Palmitate
Normal (10) — 56 72 63Respiratory distress (5) 44.5-52.0 52 64 49Respiratory distress & pulmonary edema 47-52 53 20 45
* Morgan, T. E .: Arch. Inter. Med. 127:404, 1971. t Animals were exposed to P 0 2 greater than 550 mm Hg.

24<j FO RM A N
TOTAL LECITHIN (g/100g)
F i g u r e 5 . Relation of palmitic-lecithin to total lecithin in 29 perinatal deaths.
tension values even when lung lecithin is low.5 No such relationship is present in a comparable number of stillbirths. Therefore, the lecithin content of lung must provide some measure of reserve from which surfactant is formed. An infant born with relatively small amounts of lecithins may be at risk for developing surfactant deficiency but would only do so if additional factors were present (acidosis, massive lung hemorrhage).
Results of studies using radioactive palmitic acid to label DPL, has demonstrated that the surfactant has a rapid turnover with a half-life of about 14 hours.20 Recent studies, however, have shown that the rate of turnover of surfactant is considerably slower.19 The biological half-life for both choline and phosphate incorporated into lecithin of surfactant is 43 hours
F i g u r e 6 . A . Expanded normal alveolus surrounded by small interstitial space and drained by lymphatic. B. Collapsed alveolus lacking surfactant with increased interstitial fluid and decreased lymphatic drainage.
and the biological half-life for leucine incorporated into the protein of surfactant is 45 hours. This similarity in the biological half-life for choline, phosphate and leucine suggests that surfactant is a lipo-protein which turns over as a single unit.2 These latter findings intimate that a transient or relatively acute decrease of surfactant biosynthesis would not result in measurable surfactant deficiency. Numerous biochemical and ultrastructural observations9 postulate that the pulmonary surfactant is synthesized by Type II alveolar epithelial cells, stored in the inclusion bodies of these cells and then are secreted into the alveolar space where it forms a layer on the surface of the alveolar wall. The previous concept that the surfactant layer is formed from a single phospholipid film interspaced between air and liquid phase has been challenged recently by a concept12 which views the surfactant layer as a multimolecular gel structure which protects the air spaces against transudation of fluid since gels and certain liquid-crystaline states can tolerate wide variations in water content ( figure 6).
B ioch em ical E va lu ation o f A m nio tic F lu id P h osph olip ids
Survival of the prematurely delivered human fetus depends upon its ability to synthesize surface-active alveolar lecithin after breathing begins.14 Previous reports by Biezenski3 attempting to correlate lipids in amniotic fluid with abnormal pregnancies showed no clearly defined patterns. However, concentrations of total lipids rise as gestation progresses and relationships between lecithin and sphingomyelin concentrations permit evaluation of pulmonary maturity. Prior to 35 weeks gestation, sphingomyelin concentrations are either greater than or almost identical to those of lecithin (figure 7). Statistically, week 35 of gestation is the turning point when a sudden increase in lecithin/sphingomyelin ratio occurs. By week 36, this ratio is 3: 1 or
Blood A
Surfactant

BIOCHEMICAL BASIS OF HYALINE MEMBBANE DISEASE 247
greater and thereafter the lung function becomes mature. An infant bom spontaneously or by Cesarean section or induction at this time should have no RDS. Where the ratio of lecithin/sphingomyelin in amniotic fluid is one or less than one, RDS is probable, should birth take place at that time. Findings on thin-layer chromatography11 (TLC) lend themselves to helpful interpretation by visual inspection. The TLC examination of amniotic fluid indicates whether or not biochemical development of the lung has progressed to the point of alveolar stability. When the lung has matured sufficiently, RDS will not occur, should the fetus be delivered at that time or later.
The changes in both concentrations of lecithin in amniotic fluid and fatty acids esterified on a- and B-carbons of lecithin parallel changes observed in developing fetal lung and in lungs of infants after birth.16 The fatty acids from surface active lecithin are predominantly a-palmitic/B- palmitic. The alveolar stability seen much earlier in the human fetus is due to production from about week 20 to 22 of less stable surface active palmitoyl-myrisoyl lecithin.16 This is synthesized by the second (methyla- tion) pathway for lecithin biosynthesis in human fetal lung. This pathway is extremely sensitive to metabolic insults, such as acidosis, hypoxia and hypothermia, which accounts for failure of the premature infant to survive.
A major advantage for examining amniotic fluid for lecithin/sphingomyelin is that it is reliable even when the fetus is dysma- ture. The timetable for lung maturity as reflected by a rise in lecithin is apparently unaffected when there is a deviation of growth in utero. This is exemplified by the baby who has a low birth weight and characteristically does not have RDS.16 When decisions to interrupt a pregnancy have to be made, other procedures for assessing fetal maturity (creatinine, lactic
* PO SS IB LE PULMONARY MATURITY
F i g u r e 7. Gestational changes in amniotic fluid phospholipids indicative of pulmonary maturity.
dehydrogenase) may be of little help when the fetus is dysmature.
This procedure for detecting adequate pulmonary surfactant promises to be helpful in timing elective interruption of pregnancy, in deciding on inhibition of uncomplicated premature labor, and in predicting the clinical course of the infant whose birth is expected within hours. It can also help guide pharmacologic manipulation of the fetus.
Acknowledgment
Sincere thanks are extended to Dr. John U. Balis of the Department of Pathology, Loyola University Stritch School of Medicine, Chicago, II for scientific materials which were generously made available.
R eferences1. A v e r y , M . E. a n d M e a d , J.: Surface prop
erties in relation to atelectasis and hyaline membrane disease. Amer. J. Dis Child 97: 517-523, 1959.
2. B a l i s , J. U. a n d S h e l l e y , S . A.: Quantitative evaluation of the surfactant system of the lung. Ann. Clin. Lab. Sci. 2:410—419, 1972.
3 . B i e z e n s k i , J. J., P o m e r a n c e , W., a n d G o o d m a n , J.: Studies on the origin of amniotic

248 FORMAN
fluid lipids—I. Normal Composition. Amer. J. Obstet. Gynec. 105:853-861, 1968.
4. B j o r n s t a d , P. a n d B r e m e r , J.: In vivo studies on pathways for the biosynthesis of lecithin in the rat. J. Lipid Res. 7:38-45, 1966.
5. B o u g h t o n , K., G a n d y , G ., a n d G a i r d n e r , D.: Hyaline membrane disease. II. Lung lecithin. Arch. Dis. Child. 45:311-20, 1970.
6. B r e m e r , J. a n d G r e e n b e r g , D. M.: Methyl transferring enzyme system of microsomes in the biosynthesis of lecithin (phosphatidyl choline). Biochim. Biophys. Acta 46:205-216, 1961.
7. B b o w n , E. S.: Chemical identification of a pulmonary surface active agent. Fed. Proc. 22:438, 1972.
8. B r u m l e y , G . W., C h e r n i c k , V., a n d H o d s o n , W. A.: Correlation of mechanical stability, moiphology, pulmonary surfactant, and phospholipid content in the developing lamb lung. J. Clin. Invest. 46:863-873, 1967.
9. B u c k in g h a m , S ., H e i n e m a n n , H . O., S o m e r s , S . C., a n d M c N a r y , W. F.: Phospholipid synthesis in the pulmonary alveolar cell. Amer. J. Path. 48:1027-1041, 1966.
10. C l e m e n t s , J. A.: Surface tension of lung extracts. Proc. Soc. Exp. Biol. Med. 95:170-172, 1957.
11. F o r m a n , D. T. a n d B a l i s , J. U.: Measurement of phospholipids in amniotic fluid for the assessment of pulmonary maturity. Manual of Procedures of the Seminar on Clinical Pathology of Lipids, Institute for Clinical Science, Philadelphia, pp. 1-8, 1971.
12. F r o s o l o n o , M. F . , C h a r m s , B . L., P a w l o w - s k i , R., a n d S l i n k a , S . : Isolation, characteriza
tion & surface chemistry of a surface active fraction from dog lung. J . Lipid Res. 11:439- 457, 1970.
13. G a n d y , G ., J a c o b s o n , W., a n d G a i r d n e r , D.: Hyaline membrane disease. I. Cellular changes. Arch. Dis. Child. 45:289-294, 1970.
14. G l u c k , L., K u l o v ic h , M. V., B o r e r , R. C., Jr., B r e n n e r , P. H., A n d e r s o n , G. G., a n d S p e l l a c y , W. N.: The diagnosis of the respiratory distress syndrome (RD S) by amniocentesis. Amer. J. Obstet. Gynec. 109:440- 445, 1971.
15. G l u c k , L.: Surfactant: 1972 Symposium on recent clinical advances. Pediat. Clin. N. Amer. 19:325-330, 1972.
16. G l u c k , L., K u l o v ic h , M. V., E i d e l m a n , A. I., C o r d e r o , L., a n d K h a z i n , A. F .: Biochemical development of surface activity in mammalian lung. IV. Pulmonary lecithin synthesis in the human fetus and newborn and etiology of the respiratory distress syndrome. Pediat. Res. 6:81-99, 1972.
17. K l a u s , M. H., C l e m e n t s , J. A., a n d H a v e l , R. J .: Composition of surface-active material isolated from beef lung. Proc. Nat. Acad. Sci. 47:1858-1859, 1961.
18. M o r g a n , T. E . : Biosynthesis of pulmonary surface-active lipid. Arch. Intern. Med. 127: 401-407, 1971.
19. S p i t z e r , H. L. a n d N o r m a n , J . J . : The biosynthesis and turnover of surfactant lecithin and protein. Arch. Intern. Med. 127:429-435, 1971.
20. T i e r n e y , D. F., C l e m e n t s , J . A., a n d T r a h a n , H. J . : Rates of replacement of lecithins and alveolar instability in rat lungs. Amer. J . Physiol. 213:671-676, 1967.