Embed Size (px)
Citation preview

lable at ScienceDirect
Food Control 63 (2016) 1e10
Contents lists avai
Food Control
journal homepage: www.elsevier .com/locate/ foodcont
Assessment of antimicrobial activity of Na -lauroyl arginate ethylester(LAE®) against Yersinia enterocolitica and Lactobacillus plantarum byflow cytometry and transmission electron microscopy
J. Coronel-Le�on a, 1, A. L�opez a, M.J. Espuny a, M.T. Beltran b, A. Molinos-G�omez b,X. Rocabayera b, A. Manresa a, *
a Laboratori Microbiologia, Facultat de Farm�acia, Universitat de Barcelona, Joan XXIII s/n, 08028 Barcelona, Spainb Laboratorios Miret, SA, 08228 Terrassa, Barcelona, Spain
a r t i c l e i n f o
Article history:Received 7 July 2015Received in revised form22 October 2015Accepted 31 October 2015Available online 14 November 2015
Keywords:Na -lauroyl arginate ethylesterLAEAntimicrobial activityBiocidesSurfactantsYersinia enterocoliticaLactobacillus plantarumFlow cytometryElectron microscopy
* Corresponding author.E-mail addresses: [email protected] (J. Coro
(A. L�opez), [email protected] (M.J. Espu(M.T. Beltran), [email protected] (A. Molinos-G(X. Rocabayera), [email protected] (A. Manresa).
1 Current address: Escuela Superior Polit�ecnica deIngeniería ec�anica y Ciencias de la Producci�on (ESPGalindo, Guayaquil-Ecuador, Ecuador.
http://dx.doi.org/10.1016/j.foodcont.2015.10.0500956-7135/© 2015 Published by Elsevier Ltd.
a b s t r a c t
Na -lauroyl arginate ethylester, LAE®, which was approved as GRAS by the US Food and Drug Adminis-tration (FDA) and by the European Food Safety Authority (EFSA) in 2007, is a surfactant that exhibitsantimicrobial activity. To assess its antimicrobial effect, treated cell suspensions of Yersinia enterocoliticaand Lactobacillus plantarum were analysed for reduction of cell viability. Membrane dysfunction wasdetermined by staining with bis-oxonol to detect the loss of membrane potential, and with propidiumiodide to detect permeabilized membranes by flow cytometry. LAE® treatment for 30 min induced a 4log10 reduction in cell viability in both bacteria; different subpopulations with variable degrees of cellulardamage were observed by flow cytometry. Permeabilized membranes suggested the leakage of cellularmaterial; this was also indicated by the loss of potassium ion(s), which was higher in L. plantarum than inY. enterocolitica. Structural changes involving collapse of the cytosol and alterations of the cellular en-velopes, mainly in Y. enterocolitica, and the formation of mesosomes in L. plantarum were observed bytransmission electron microscopy.
© 2015 Published by Elsevier Ltd.
1. Introduction
Two approaches to microbiological assessment in food are mi-crobial food safety and microbial food spoilage. The challenge inmicrobial food safety is to protect public health by minimizing themicrobiological risk, an increasingly complex task in the newglobalmarket, characterized by new food vehicles, trade and technologiesas well as population migrations and climate change. Over the lastdecades, microbial food security has attracted considerable atten-tion and their control is mainly based on a preventive approach,
nel-Le�on), [email protected]), [email protected]�omez), [email protected]
l Litoral, ESPOL, Facultad deOL-FIMCP) Campus Gustavo
such as adopting good hygiene practices. In the implementation ofthe Hazard Analysis and Critical Control Points system (HACCP) toreduce the disease burden, microbiological criteria are used in thevalidation and verification of procedures throughout the foodchain. Improvements in microbiological food safety have beenachieved by the implementation of international standards andlegislation, which have had a positive impact on diarrhoeal inci-dence worldwide (Newell et al., 2010). Although it is difficult topredict new trends in food safety issues, continuous microbial riskassessment could reveal new emerging foodborne pathogens.
The predominant causative agents of foodborne disease,50e60% of which are unknown, recently Salmonella, Escherichia coliand Campylobacter, and Listeria have been the focus of attentiondue to an increase of cases (Becerril, Manso, Nerin, & G�omez-Lus,2013; Kennedy, Cronin, & Wilkinson, 2011; Newell et al., 2010;Stopforth, Visser, Zumbrink, Dijk, & Bontenbal, 2010; Suksathit &Tangwatcharin, 2013; Theinsathid, Visessanguan, Kruenate,Kingcha, & Keeratipibul, 2012).
Yersinia enterocolitica is not explicitly mentioned in European

J. Coronel-Le�on et al. / Food Control 63 (2016) 1e102
legislation (Regulation 1441/2007), but according to the HACCPsystem every company should assess hazards and specific criticalpoints of control in processes related to the manufactured,distributed and/or sold product (EFSA, 2007). Therefore,Y. enterocolitica, as an emerging foodborne pathogen with limiteddocumentation (Nesbakken, Eckner, & Røtterud, 2008; Ye, Wu, Hu,Zhang, & Huang, 2016), deserves attention.
Y. enterocolitica is a psychrotrophic bacteriumwith the ability togrow at temperatures below 4 �C. As it can survive more easily infood stored at room and refrigeration temperatures than at an in-termediate temperature, and has a longer persistence in cookedthan in raw foods, due to increased nutrient availability (FDA,2012), the risk of it causing disease increases in ready-to-eatfoods. In fact, yersiniosis is currently the third most predominantzoonose among foodborne illnesses across Europe, with around8000 reported cases to date (ACSA, 2011). EFSA-published dataindicate that, on average, 4.8% of pork meat samples in all EUmember states contained Y. enterocolitica (EFSA, 2014). The mostcommon sources of yersiniosis infection are pork and lamb, andstrict slaughter hygiene is important to limit carcass contaminationwith Y. enterocolitica, as well as other human pathogenic microor-ganisms (Skjerve, Lium, Nielsen,&Nesbakken,1998). Outbreaks areassociated with the consumption of meat, especially of non-heat-treated fermented sausages (Lindqvist & Lindblad, 2009), and un-pasteurized milk and dairy products (FDA, 2012). Other risk factorsinclude cross-contamination through surfaces, utensils or hands,and consumption of untreated well water.
The economic losses caused by microbial spoilage of foods areanother serious concern for the industry. Risk factors in food pro-cessing include undercooking, cross-contamination from rawmeat,food handlers or food contact surfaces, as well as poor temperatureand time control, and storage conditions tomaintain the qualitativecharacteristics of the product.
Lactic acid bacteria are involved in diverse meat fermentationprocesses and are recognized as important starter cultures. Theircellular state under different stress conditions has recently beenstudied (Bonomo, Milella, Martelli, & Salzano, 2013). Lactobacillusplantarum is a lactic acid bacterium associated with spoilage in awide range of products, found in sugar distilleries (Oliva Neto et al.,2014), and particularly in ready-to-eat meat (C�amara-Martos,Zurera-Cosano, Moreno-Rojas, García-Gimeno, & P�erez-Rodríguez,2012; Fern�adez-Ramírez., Smid, Abee, & Nierop Groot, 2015). Anindicator of food quality, but not safety, is the Aerobic Colony Count(ACC). An ACC of less than 106 cfu/g is usually associated with amixed flora. Above this level there is usually one predominant or-ganism, and the acceptability and organoleptic quality of the foodwill depend onwhich type of organism predominates. Spoilage willeventually occur due to the production of lactic acid related withpredominant bacteria (mainly lactobacilli and streptococci) at alevel of around 109 cfu/g (Health Protection Agency (HPA, 2009).
L. plantarum has been associated with biofilm formation, whichis a potential source of contamination. Information on environ-mental conditions that influence biofilm formation by lactobacilli isof relevance to food-producing industries and could assist with thedesign of new intervention strategies to prevent biofilm formation(Fern�adez-Ramirez, Smid, Abee, & Nierop Groot, 2015). To this end,besides rational equipment designed to minimize microbial risk byfacilitating cleaningeineplace procedures and good manufacturepracticum, biocides might be added to packaging polymers (Lebert,Leroy, & Talon, 2007; Muriel-Galet, L�opez-Carballo, Gavarra, &Hern�andez-Mu~noz, 2015).
Na -lauroyl arginate ethylester, LAE®, is a surfactant derivedfrom lauric acid and arginine that exhibits antimicrobial activity(Ma, Davidson, & Zhong, 2013; Martin et al., 2009; Muriel-Galetet al., 2015; Pattanayaiying, H-Kittikun, & Cutter, 2014; Rodríguez,
Seguer, Rocabayera,&Manresa, 2004; Suksathit& Tangatwacharin,2013). It is metabolized by humans to natural dietary componentsand thus has been considered as GRAS by the FDA since 2005.Another advantage is that it is odour-free and colourless, whichfavours its application as an additive (EC, 2005; EFSA, 2012) in thecosmetic and food industries (EFSA, 2012; Otero et al., 2014;Stopforth, Visser, Zumbrink, Dijk & Bontenbal 2010; Theinsathidet al., 2012).
Traditionally, cell count has been used to assess the effect ofantimicrobial treatment. A complementary technique is flowcytometry, which allows the rapid measurement of several pa-rameters within the cell. Antimicrobial treatments frequentlytarget the membrane, leading to alteration of membrane func-tionality and permeability and eventually the loss of cellular ma-terial and structural changes. Flow cytometry is a sensitivetechnique for assessing membrane functionality and integritywhen combined with specific cell staining to determine differenttypes of cellular damage (Kennedy et al., 2011).
In this work we explored the antibacterial effect of LAE® as afood additive against Y. enterocolitica, a model of an emergingfoodborne bacteria, and L. plantarum, active in food spoilage. Toassess the biocidal activity of LAE® against these bacteria, we usedflow cytometry, ion efflux and transmission electron microscopy.
2. Material and methods
2.1. Materials
Lauroyl arginate ethyl (LAE®), CAS registration number 60372-77-2 of 91.7% purity was supplied by VEDEQSA SA (Barcelona,Spain). A stock solution of powdered LAE® was prepared before usein bi-distilled water (Millipore, MA, USA). Molecular dyes weresupplied by Molecular Probes Europe (BV, Leiden. Netherlands).Microbiological products were supplied by Pronadisa (Barcelone,Spain) and Oxoid (Oxoid Ltd, Basingstoke, UK). All chemicals andreagents were of analytical grade and supplied by Panreac (Barce-lona, Spain) or SigmaeAldrich (St Louis, USA).
2.2. Microorganisms
Y. enterocolitica ATCC 9610 and L. plantarum ATCC 8014 strainswere maintained frozen in cryovials (AES Laboratoire, Combourg,France) at �80 �C and sub-cultured weekly on trypticase soya agar(TSA; Pronadisa, Barcelona, Spain).
2.3. Antimicrobial susceptibility test
The minimal inhibitory concentration (MIC) of LAE® wasdetermined using a broth microdilution assay (Woods &Washington, 2003). Serial dilutions of LAE®, between 64 and4 mg ml�1
final concentration, in MullereHinton Broth (Oxoid Ltd,Basingstoke, UK) were dispensed (200 mL) in the correspondingwells of a 96-well polypropylene microtitre plate (Costar; CorningIncorporated, Corning, NY, USA). For the inoculum, a colony pickedfrom a TSA agar plate was suspended in Mueller-Hinton Broth andincubated for 18 h; an aliquot was then inoculated in Mueller-Hinton Broth and further incubated for 2 h. The turbidity wasadjusted to the 0.5 McFarland barium sulphate standard. The cor-responding dilutions (200 mL) were inoculated with 10 mL suspen-sion of the test organism in MuellereHinton Broth to a finalconcentration of ca 105 CFU ml�1. The MIC was defined as thelowest concentration of antibacterial agent that inhibited devel-opment of visible growth after 24 h of incubation at 37 �C. TheMBC,defined as the antimicrobial concentration corresponding to atleast 3 log reductions of viable cells, was determined by spreading

Fig. 1. Effect of LAE on bacterial count on L. plantarum (◊) and Y. enterocolitica (▫) atthe respective MIC concentration. Bacteria were suspended in buffered peptone waterand mainrained at 25 �C.
J. Coronel-Le�on et al. / Food Control 63 (2016) 1e10 3
100 ml aliquots from negative wells on trypticase soy agar (TSA,Pronadisa, Barcelona, Spain), and the plates were incubated for 48 hat 37 �C (Branen & Davidson, 2004; Swenson, Hindler, & Jorgensen,2003). Experiments were conducted in triplicate.
2.4. Exposure to LAE®
Suspensions of microorganisms were obtained from an over-night culture of each strain of L. plantarum and Y. enterocolitica onTSA at 30 �C. Four colonies were suspended in 10 ml of peptonebuffered solution, pH 7.0. Each bacterial suspensionwas pelleted bycentrifugation at 8000 � g for 20 min, washed in the sterile filteredpeptone buffered solution, pH 7.0, centrifuged again, and finallysuspended to obtain a concentration of 107e108 CFU/mL. Anappropriate volume (250 mL) of the respective cell suspensions wasused to inoculate flasks containing 24ml of buffered peptonewater(pH 7) to obtain a cell density of ca 105e106 CFU/mL. To flaskscontaining 24ml of the respective bacterial suspensions, LAE® stocksolution (1 mL) was added to reach a final concentration of LAE®
corresponding to theMIC and 2/3 of theMIC for Y. enterocolitica andL. plantarum. The inoculated flasks were kept in darkness at roomtemperature. The contact time established for FC was 30 min. Fortransmission electronmicroscopy (TEM) observations, contact timewas 3 h, then suspensions were centrifuged at 8000 � g for 30 min.The sediment was resuspended in 2 ml of filtered peptone bufferedwater, pH 7.0.
In parallel control experiments, cells were incubated in LAE®-free buffer solution and treated under the same conditions. At theend of the contact time, the bacterial suspensions were dilutedwith the same buffer, centrifuged at 8000 � g for 30 min andwashed to eliminate the LAE®.
2.5. Bacterial counts
After contact with LAE®, reduction in bacterial viability wascalculated by viable cell counts, calculated from the colony formingunits (CFUml�1) obtained on TSA. Rapid separation of bacteria fromLAE® was achieved by centrifugation at 8000 g for 10 min in abench centrifuge, after an appropriate dilution in Ringer's solution,0.1 mLwas inoculated on plates and incubated at 30 �C for 24e48 h.Cell counting was performed per triplicate and the mean wascalculated. Based on these data, growth inhibition was calculatedaccording to the viability reduction as follows (Bouhdid, Abrini,Zhiri, Espuny, & Manresa, 2009):
(1�NT/NC) � 100
where NT is the bacterial count in the treated sample and Nc is thebacterial count in the control sample.
2.6. Flow cytometry
The staining protocols for FC experiments were as follows: 10 mLof a 1 mg ml�1 stock solution of propidium iodide (PI) in distilledwater was added to 1 ml of the bacterial suspension, containing107e108 CFU/mL prepared as described earlier in filtered bufferedpeptone water. Staining was carried out at room temperaturebefore the FC analysis. To evaluate membrane potential, 2 mL of a250 mmol/L stock solution of bis-oxonol in ethanol was added to1 ml of the bacterial suspension. Heat-killed cells (30 min at 70 �C)were used as a positive control for PI and bis-oxonol staining pro-tocols. Experiments were performed in duplicate.
Flow cytometry experiments were carried out using a CytomicsFC500 MPL flow cytometer (Beckman Coulter, Inc., Fullerton, CA,USA). Samples were excited using a 488-nm air-cooled argon-ion
laser at 15mWpower. The instrument was set upwith the standardconfiguration: green (525 nm) fluorescence for bis-oxonol, red(675 nm) for PI. The results were collected on logarithmic scales.Optical alignment was checked using 10-nm fluorescent beads(Flow-Check fluorospheres, Beckman Coulter). The cell populationwas selected by gating in a FS vs SS dot plot, excluding aggregatesand cell debris. Fluorescence histograms were represented insingle-parameter histograms (1024 channels). The windows usedto calculate percentages were set using living and dead cell pop-ulations for each strain. Data were analysed using Summit® ver. 3.1software (Cytomation, Fort Collins, CO, USA).
2.7. Potassium ion leakage
Potassium ion(s) (Kþ) leakage was determined as follows(Rodriguez et al., 2004). Briefly, the bacterial strains were grownovernight in Trypticase soy broth (TSB) medium at 30 �C in ashaking incubator (120 rev/min). Cells were then washed threetimes in 0.9% NaCl by centrifugation at 8000 � g for 15 min and re-suspended in 30 mL of 1 mmol/L glycyl-glycine (Sigma, USA) buffersolution, pH 6.8, to obtain a cell density of 7.4 � 107 CFU/mL forY. enterocolitica and 8.8 � 107 CFU/mL for L. plantarum. The bacteriawere treated with LAE® at the MIC and at 2/3 MIC and incubated at30 �C in a shaking incubator. Samples (5 mL) of cell suspensionwere removed at 0, 10, 15, 30, 60 and 120 min, diluted and filteredthrough a 0.22-mm pore-size membrane (Merck-Millipore, Darm-stadt, Germany) to remove bacteria. LAE®-free controls were pre-pared in the same conditions to determine normal Kþ
flux over thetime course of the experiment. Heat treatment was also conductedby incubating the cell suspension in a water bath at 70 �C. The Kþ
concentration in the supernatant was measured using an atomicabsorption spectrophotometer (UNICAM 939⁄959, Cambridge, UK).The instrumental parameters were as follows: Kþ hollow cathodelamp, wavelength of 766.5 nm, band pass of 0.5 nm, air-acetyleneflame and fuel flow rate of 1 L/min. Absorbance values were con-verted to Kþ concentration (ppm) by reference to a curve previouslyestablished using standard Kþ solutions of 0, 0.05, 0.1, 0.3, 0.5 and1 ppm. Experiments were conducted in triplicate, and means andstandard deviations were calculated.
2.8. Transmission electron microscopy
After treatment of cell suspensions with LAE® for 3 h at the MICand 2/3 MIC for each microorganism, the bacterial pellets were

Table 1Bacterial counts of Yersinia enterocolitica and L. plantarum treated at MIC concen-tration of LAE®.
Time (min) Y. enterocolitica L. plantarum
0 1.0 � 107 2.1 � 107
5 2.8 � 104 3.1 � 104
10 1.1 � 104 1.3 � 104
30 1.7 � 103 3.5 � 103
60 7.0 � 100 3.0 � 100
90 e e
120 e e
J. Coronel-Le�on et al. / Food Control 63 (2016) 1e104
rinsed with buffered peptone water, pH 7.0. Y. enterocolitica andL. plantarum pellets were transferred to planchettes (1 mm diam-eter and 200 mm deep) and immediately cryoimmobilized using aLeica EMPact high pressure freezer (Leica Vienna, Austria). Plan-chettes were cryofixed with a pressure of about 2100 bars and areduction in temperature of 8 �C/s and then stored in liquid ni-trogen until further use. Frozen samples were freeze-substituted ina Leica EMAFS (automatic freeze substitution system, Leica, Vienna,Austria) where the substitution was performed in pure acetonecontaining 2% of osmium tetroxide and 0.1% of uranyl acetate for
Fig. 2. Dual parameter of Y. enterocolitica stained cells (bis-oxonol and PI) at different LAEpopulation; c) population treated with LAE at 2/3 MIC concentration (5.33 mg ml�1); d) po
72 h at �90 �C. The temperature was gradually increased(Dt ¼ 5 �C h�1) to 4 �C and held constant for 2 h and then finallyincreased to room temperature and maintained for 1 h. Sampleswere washed twice with acetone at room temperature and infil-trated in a graded series of Epon-acetone mixtures, and finallysamples were embedded in fresh Epon and polymerized at 60 �C for48 h. Ultrathin sections were prepared with a Leica UCT ultrami-crotome and mounted on Formvar carbon-coated copper grids.Sections were post-stained with 1% uranyl acetate and lead citrate.Sections were examined with a Phillips EM 30 (Eindhoven,Holland) microscope with an acceleration of 60 kV.
3. Results
3.1. Antimicrobial activity of LAE®
The MIC of LAE® was 8 mg ml�1 against Y. enterocolitica and32 mg/mL against L. plantarum. The effect of LAE® over time onbacterial suspensions in buffered peptone water is presented inFig. 1. After 5 min of exposure, Y. enterocolitica suffered 3 log10 unitsloss of viability which reached 4 log10 units, at 30 min. In
concentration a) positive control, untreated population; b) negative thermal treatedpulation treated with LAE at MIC concentration (8 mg ml�1).

Fig. 3. Dual parameter of L. plantarum stained cells (bis-oxonol and PI) at different LAE concentration a) positive control, untreated population; b) negative thermal treatedpopulation; c) population treated with LAE at 2/3 MIC concentration (21.33 mg ml�1); d) population treated with LAE at MIC concentration (32 mg ml�1).
Table 2Percentage of stained cells measured by flow cytometry analysis and viability reduction in Yersinia enterocolitica treated with LAE®.
Treatment Cell number Unstained cells (%) % Of stained cells % Reduction viability by cell counts
Bis-oxonol PI
positive control none 5.0 � 104 98.7 0.60 0.7 0LAE® 5.3 mg/mL 2.0 � 104 0.4 3.70 95.80 98.33
8.0 mg/mL 2.0 � 104 0.5 1.80 97.80 100negative control 70 �C 2.0 � 104 0.4 6.6 93. 0 0
J. Coronel-Le�on et al. / Food Control 63 (2016) 1e10 5
L. plantarum, after treatment at MIC (32 mg/mL), the reduction inviability was 3 log10 units after 5 min, increasing to 4 log10 unitsafter 30 min. In both cases, there was a 100% reduction in viabilityafter 60 min of contact (Fig. 1).
The minimal bactericidal concentration (MBC) of LAE® wasidentical to that of the corresponding MIC: Y. enterocolitica (8 mg/mL) and L. plantarum (32 mg/mL). A decrease of 99.7% and 99.8%,
respectively, in cell viability was observed after 5 min of contact,and a decrease of 99.9% after 30 min was observed in both micro-organisms assayed (Table 1).
3.2. Flow cytometry
In order to study the effect of LAE® on bacterial suspensions, the
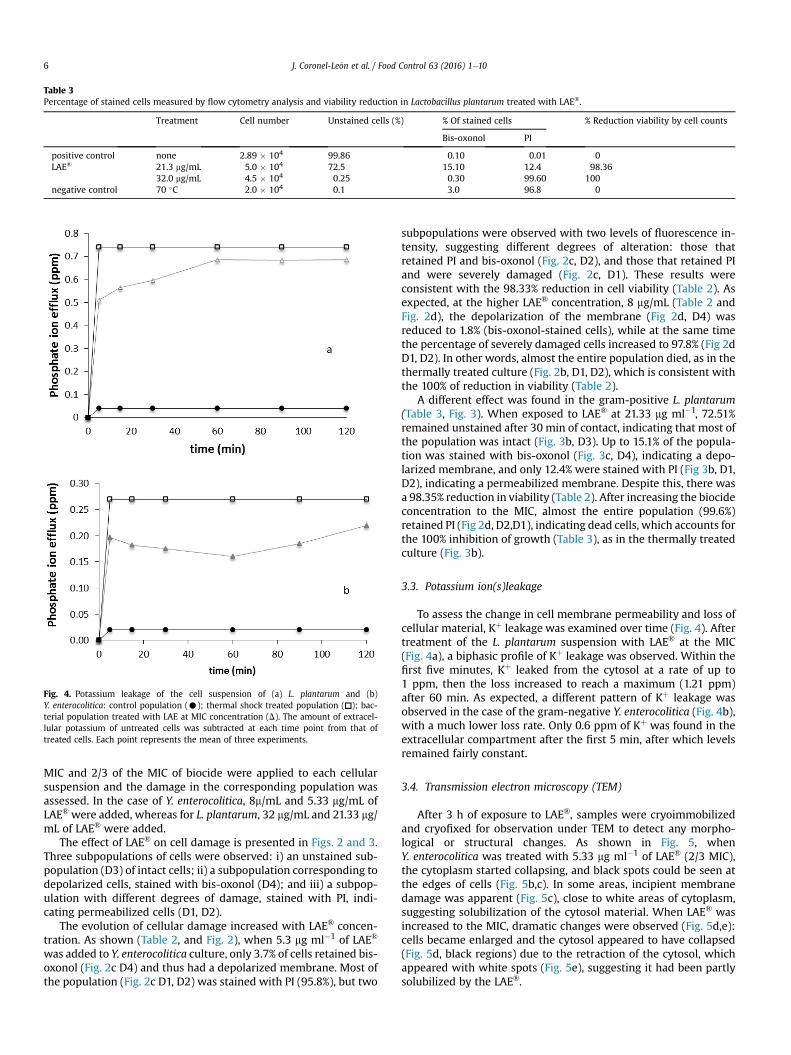
Table 3Percentage of stained cells measured by flow cytometry analysis and viability reduction in Lactobacillus plantarum treated with LAE®.
Treatment Cell number Unstained cells (%) % Of stained cells % Reduction viability by cell counts
Bis-oxonol PI
positive control none 2.89 � 104 99.86 0.10 0.01 0LAE® 21.3 mg/mL 5.0 � 104 72.5 15.10 12.4 98.36
32.0 mg/mL 4.5 � 104 0.25 0.30 99.60 100negative control 70 �C 2.0 � 104 0.1 3.0 96.8 0
Fig. 4. Potassium leakage of the cell suspension of (a) L. plantarum and (b)Y. enterocolitica: control population (C); thermal shock treated population (▫); bac-terial population treated with LAE at MIC concentration (D). The amount of extracel-lular potassium of untreated cells was subtracted at each time point from that oftreated cells. Each point represents the mean of three experiments.
J. Coronel-Le�on et al. / Food Control 63 (2016) 1e106
MIC and 2/3 of the MIC of biocide were applied to each cellularsuspension and the damage in the corresponding population wasassessed. In the case of Y. enterocolitica, 8m/mL and 5.33 mg/mL ofLAE® were added, whereas for L. plantarum, 32 mg/mL and 21.33 mg/mL of LAE® were added.
The effect of LAE® on cell damage is presented in Figs. 2 and 3.Three subpopulations of cells were observed: i) an unstained sub-population (D3) of intact cells; ii) a subpopulation corresponding todepolarized cells, stained with bis-oxonol (D4); and iii) a subpop-ulation with different degrees of damage, stained with PI, indi-cating permeabilized cells (D1, D2).
The evolution of cellular damage increased with LAE® concen-tration. As shown (Table 2, and Fig. 2), when 5.3 mg ml�1 of LAE®
was added to Y. enterocolitica culture, only 3.7% of cells retained bis-oxonol (Fig. 2c D4) and thus had a depolarized membrane. Most ofthe population (Fig. 2c D1, D2) was stained with PI (95.8%), but two
subpopulations were observed with two levels of fluorescence in-tensity, suggesting different degrees of alteration: those thatretained PI and bis-oxonol (Fig. 2c, D2), and those that retained PIand were severely damaged (Fig. 2c, D1). These results wereconsistent with the 98.33% reduction in cell viability (Table 2). Asexpected, at the higher LAE® concentration, 8 mg/mL (Table 2 andFig. 2d), the depolarization of the membrane (Fig 2d, D4) wasreduced to 1.8% (bis-oxonol-stained cells), while at the same timethe percentage of severely damaged cells increased to 97.8% (Fig 2dD1, D2). In other words, almost the entire population died, as in thethermally treated culture (Fig. 2b, D1, D2), which is consistent withthe 100% of reduction in viability (Table 2).
A different effect was found in the gram-positive L. plantarum(Table 3, Fig. 3). When exposed to LAE® at 21.33 mg ml�1, 72.51%remained unstained after 30 min of contact, indicating that most ofthe population was intact (Fig. 3b, D3). Up to 15.1% of the popula-tion was stained with bis-oxonol (Fig. 3c, D4), indicating a depo-larized membrane, and only 12.4% were stained with PI (Fig 3b, D1,D2), indicating a permeabilized membrane. Despite this, there wasa 98.35% reduction in viability (Table 2). After increasing the biocideconcentration to the MIC, almost the entire population (99.6%)retained PI (Fig 2d, D2,D1), indicating dead cells, which accounts forthe 100% inhibition of growth (Table 3), as in the thermally treatedculture (Fig. 3b).
3.3. Potassium ion(s)leakage
To assess the change in cell membrane permeability and loss ofcellular material, Kþ leakage was examined over time (Fig. 4). Aftertreatment of the L. plantarum suspension with LAE® at the MIC(Fig. 4a), a biphasic profile of Kþ leakage was observed. Within thefirst five minutes, Kþ leaked from the cytosol at a rate of up to1 ppm, then the loss increased to reach a maximum (1.21 ppm)after 60 min. As expected, a different pattern of Kþ leakage wasobserved in the case of the gram-negative Y. enterocolitica (Fig. 4b),with a much lower loss rate. Only 0.6 ppm of Kþ was found in theextracellular compartment after the first 5 min, after which levelsremained fairly constant.
3.4. Transmission electron microscopy (TEM)
After 3 h of exposure to LAE®, samples were cryoimmobilizedand cryofixed for observation under TEM to detect any morpho-logical or structural changes. As shown in Fig. 5, whenY. enterocolitica was treated with 5.33 mg ml�1 of LAE® (2/3 MIC),the cytoplasm started collapsing, and black spots could be seen atthe edges of cells (Fig. 5b,c). In some areas, incipient membranedamage was apparent (Fig. 5c), close to white areas of cytoplasm,suggesting solubilization of the cytosol material. When LAE® wasincreased to the MIC, dramatic changes were observed (Fig. 5d,e):cells became enlarged and the cytosol appeared to have collapsed(Fig. 5d, black regions) due to the retraction of the cytosol, whichappeared with white spots (Fig. 5e), suggesting it had been partlysolubilized by the LAE®.

J. Coronel-Le�on et al. / Food Control 63 (2016) 1e10 7
When L. plantarum was treated with 21.33 mg ml�1 LAE® (2/3MIC), an increase in size of the cellular envelope was observed,compared with the untreated population (Fig. 6a), along with se-vere damage to the bacterial membrane (Fig. 6b). Intracellularmesosomes were observed invading the cytoplasm (Fig. 6b) andapparent alterations in the cytosol could be observed. At the MIC ofLAE® (Fig. 6d,e), cells developed a granular appearance, with thecollapse of the cytoplasm.
4. Discussion
Na lauroyl-L-arginine ethyl ester monohydrochloride has aknown antimicrobial effect against some gram-positive and gram-negative bacteria. The mechanism of action of such compoundsdepends on the interaction of the alkyl chain and the hydrophobicmembrane of microorganisms (Becerril et al., 2013; Rodríguez et al.,2004). To gain new insight into the spectrum of activity of LAE®,two food-related bacteria were assayed for susceptibility to thiscompound. Given that contamination of food products is a majorconcern in the food industry, we studied the effect of the biocideLAE® on two bacteria that severely effect product quality: theemerging foodborne bacteria Y. enterocolitica, and the food spoilageL. plantarum.
Microbiocides are chemical compounds with a wide range ofapplications, including in food and healthcare consumer products.LAE® has been approved as GRAS by the US Food and DrugAdministration (USDA, 2005), and by the European Food SafetyAuthority (EFSA, 2007). Interest in microbiocides reflects theincreasing emergence of pathogen resistance and a better under-standing of microbial pathogens on the part of the public (Maillardet al., 2013. Biocides have multiple targets and thus the develop-ment of resistance is less likely. Among the parameters used toevaluate the effect of an antimicrobial compound is the degree ofmetabolic activity: the lower the metabolic activity, the greater thechance of surviving exposure to an antimicrobial agent (Matos &Lopes Da Silva, 2013). In this context, the cellular effect of expo-sure to LAE®was assessed in non-proliferating cells in an attempt toreduce the metabolic activity of the bacterial population. Despitebeing non-proliferating cells, the reduction in viability in bothbacteria after 5 min of contact at the MBC was 3 log10 units,increasing to 4 log10 units after 30 min. A similar reductionalthough after 24 h of growth (about 4 log10 units) has been re-ported in several gram-positive and gram-negative bacteria whentreated at MIC (which was similar to the MBC) (Becerril, Manso,Nerín, & Gomez Lus, 2013). In contrast, Ma et al. (2013) reporteda reduction in viability of only 2 log units when LAE® was usedagainst Listeria monocytogenes. They suggest that LAE® antimicro-bial activity could be boosted by combining it with an essential oil(Ma et al., 2013).
Flow cytometry can be applied to measure physical and chem-ical parameters of individual cells, providing information bylabelling cells with specific fluorochromes (molecular probes) andproducing fluorescence signals that allow the detection of certainindicators of functionality of the cell. The anionic dye bis-oxonolwas used to assess loss of membrane potential, by entering depo-larized cells where it binds to lipid-rich components. Propidiumiodide (PI), a positively charged dye that enters the cell throughpermeabilizedmembranes and intercalates between RNA and DNA,was used to assess membrane permeability (Comas-Riu & Rius,2009). Given that the electrochemical membrane potential in
Fig. 5. Transmission electron microscopy of Yersinia enterocolitica (a) control, (b)treated with LAE 2/3 CMI concentration for 3 h (b, 30 K; c 50 K). Cells treated with LAECMI concentration 3 h (d, 75 K; e, 50 K). Arrows indicated alteration of cell membrane.

J. Coronel-Le�on et al. / Food Control 63 (2016) 1e108
viable cells should be intact, its magnitude is a tentative measure ofmicroorganism health. The dual staining with a cell-permeant dyeand a dye detecting loss in the membrane potential is a robustapproach that can distinguish between different degrees of dam-age. While Y. enterocoliticawas strongly affected by sub-MIC valuesof LAE®, the bactericidal action of LAE® increased with its concen-tration (Table 2, Fig. 2), which was evidenced by the reduction inviability. A similar effect of LAE® concentration was observed inL. plantarum, but in this case, a high proportion of unstained pop-ulation was found after sub-MIC treatment, despite the highreduction in viability (Table 3). This suggests that the effect of LAE®
on L. plantarum not only caused depolarization and permeabilitybut also some other unspecific effect inside the cell, where acollapse of the cytoplasmic material could be observed (Fig. 5).
Novo et al. reported a discrepancy between the drop in mem-brane potential and cell viability, suggesting a recovery in thebacterial population after the initial antimicrobial impact. More-over, they concluded that, although nucleic acid dyes can be used asan indicator of cell death, this criterion should not be applied in allsituations (Novo, Perlmutter, Hunt, & Shapiro, 2000). This was thecase of LAE®-treated L. plantarum, since most of the populationremained unstained (72%), while the reduction in viability was98.6%. According to Novo et al., this might be due to the multi-etarget sites of the biocide, or to a reversion to an impermeablestate (Novo et al., 2000). Thus, several parameters should bemeasured to have a comprehensive understanding of the mecha-nism of action. The effect of another surface active agent, bis(Na-caproyl-L-arginine)-1,3-propanediamine dihydrochloride, has alsobeen described as concentration-dependent, although in this casewith a low reduction in viability, suggesting that the damage wasreversible (Castillo, Clap�es, Infante, Comas, & Manresa, 2006).When Bouhdid et al. assessed the effect of Origanum compactum oilon Pseudomonas aeruginosa and Staphylococcus aureus, a highreduction in viability was found at MIC, but a subpopulation with adifferent degree of PI staining suggested a different level of damagecaused by the essential oil (Bouhdid et al., 2009).
Kþ is a major cation involved in the maintenance of membranepotential and thus the functionality of the cytoplasmic membraneand the turgor of the cell. Being a small ion, the leakage of Kþ is asign of the permeabilization of the membrane and loss of cyto-plasmic material. Lower intracellular Kþ leakage was observed inboth strains compared with the controls, but the leakage fromL. plantarum was twice that from Y. enterocolitica, which might bedue to the thicker cell envelope, as observed by TEM. In the case ofY. enterocolitica, the loss of Kþ was monophasic, suggesting thatonce the outer membrane is destabilized, LAE® acts on the cellularmembrane, causing Kþ leakage. In L. plantarum, however, the loss ofKþ followed a biphasic profile. Similarly, the leakage of Kþ wasabout 2-fold greater in Salmonella Typhimurium than S. aureus(Rodríguez et al., 2004). These results are consistent with the dataof Pattanayaiying et al., who reported that LAE® leads to more Kþ
leakage than nisin Z in E. coli 0157:H7, L. monocytogenes and Bro-chothrixt thermosphacta (Pattanayaiying et al., 2014). In agreementwith these authors, we observed by TEM that LAE® caused Kþ effluxwithout cell lysis.
Using cryo-electron microscopy, which allows a wide range ofcellular structures to be identified, our observations (Fig. 5 and 6) ofmembrane damage induced by the cationic surfactant (LAE®) areconsistent with previous reports (Becerril et al. 2013;
Fig. 6. Transmission electron micrographies of Lactobacillus plantarum. Control cells (a,40 K), treated with LAE 2/3 CMI concentration for 3 h (b, 75 K; c, 50 K) cells treatedwith LAE CMI concentration for 3 h (d, 40 K; e, 75 K). Arrows indicated change sinbacterial envelope and changes in the cytoplasm.

J. Coronel-Le�on et al. / Food Control 63 (2016) 1e10 9
Pattanayaiying et al., 2014; Rodríguez et al., 2004). InY. enterocolitica, the integrity of the cellular body was maintainedeven under a higher LAE® concentration (Fig 5c,d), and no generaldisruption of the outer membrane was observed, similar to thatreported in E. coli O157:H7 (Pattanayaining, H-Kittikun, & Cutter,2014). Instead, a severe collapse of the cytoplasm was evident, incontrast with Salmonella Rissen, in which disruption of the cellularmembranes as well as collapse of the cellular material was reported(Suksathit & Tangwatcharin, 2013). Detailed observation of theeffect of LAE® on L. plantarum revealed that the thick peptidoglycanenvelope protected the cell. Two relevant effects were observed(Fig. 6): a thickening of the envelope and coagulation of the cytosol,which was consistent with the reduction in viability. Similarly,when L. monocytogeneswas treated with LAE®, no disruption of thecellular envelope was observed (Pattanayaiying et al. 2014).Membrane mesosome formation, as reported in S. aureus(Rodriguez et al., 2004), was also evident (Fig. 6b,c) at low con-centrations of LAE®. However, despite the loss of Kþ, no massiveleakage of cellular material was observed.
5. Conclusions
In this study we report the effect of LAE® on cell functionality,such as loss of membrane potential and Kþ leakage, cell viabilityand cellular ultrastructure. Our results demonstrate that LAE®
produces lethal effects in food-related bacteria such asY. enterocolitica and L. plantarum. Uptake of PI and bis-oxonol andTEM observations showed that concentrations lower than thecorrespondingMIC of LAE® greatly reduce viability but do not causecell lysis.
Acknowledgements
We would like to acknowledge Carmen Lopez-Iglesias and J.Comas-Riu from the Serveis Científic-T�ecnics, Universitat de Bar-celona, for advice and excellent technical assistance in electronmicroscopy and flow cytometry analysis.
References
Agencia Catalana de Seguridad Alimentaria (ACSA). (2011). Riesgos emergentes.marzo-Abril, 1e3.
Becerril, R., Manso, S., Nerin, C., & G�omez-Lus, R. (2013). Antimicrobial activity oflauroyl arginate ethyl (LAE), against selected food-borne bacteria. Food Control,32(2), 404e408.
Bonomo, M. G., Milella, L., Martelli, G., & Salzano, G. (2013). Stress responseassessment of Lactobacillus sakei strains selected as potential autochthonousstarter cultures by flow cytometry and nucleic acid double-staining analyses.Journal of Applied Microbiology, 115(3), 786e795.
Bouhdid, S., Abrini, J., Zhiri, A., Espuny, E., & Manresa, A. (2009). Investigation offunctional and morphological changes in Pseudomonas aeruginosa andStaphylococcus aureus cells induced by Origanum compactum essential oil.Journal of Applied Microbiology, 106(5), 1558e1568.
Branen, J. K., & Davidson, P. M. (2004). Enhancement of nisin, lysozime, and mon-olaurin antimicrobial activities by ethylendiaminetetracetic acid and lactoferrin.International Journal of Food Microbiology, 90(1), 63e74.
C�amara-Martos, F., Zurera-Cosano, G., Moreno-Rojas, R., García-Gimeno, R. M., &P�erez-Rodríguez, F. (2012). Identification and quantification of lactic acid bac-teria in a water-based matrix with near-infrared spectroscopy and multivariateregression modeling. Food Analytical Methods, 5(1), 19e28.
Castillo, J. L., Clap�es, P., Infante, M. R., Comas, J., & Manresa, A. (2006). Comparativestudy of the antiumicrobial activity of bis(Na-caproyl-L-arginine)-1,3-pro-panediamine dihydrochloride and closhexidine dihydrochloride againstStaphylococcus aureus and Escherichia coli. Journal of Antimicrobial Chemo-tyherapy, 57, 691e698.
Comas-Riu, J., & Rius, N. (2009). Flow cytometry applications in the food industry.Journal of Industrial Microbiology & Biotechnology, 36(8), 999e1011.
European Comission Scientific Comitee on Consumer Products. (2005). Opinion onethyl lauroyl arginate SCCP 0837/04 (pp. 1e43).
European Food Safety Authority. (2007). Opinion of the scientific panel on foodadditives, flavourings, processing aids and materials in contact with food (AFC)related to an application on the use of ethyl lauroyl arginate as a food additive.
EFSA Journal, 511, 1e27.European Food Safety Authority. (2012). Revised exposure assesment for ethyl
lauroyl arginate for the proposed uses as food additive. EFSA Journal, 10(4),2452.
European Food Safety Authority. (2014). Scientific Opinion on the risk posed bypathogens in food of non-animal origin. Part 2 (Salmonella, Yersinia, Shigellaand Norovirus in bulb and stem vegetables, and carrots). EFSA Journal, 12,91e115.
European Union. Comission Regulation (EC) 1441/2007. Ammending Regulation(EC) on microbiological criteria for foodstuffs. Official Journal of the EuropeanUnion L- 322/12.
Fern�adez-Ramirez, M. D., Smid, E. J., Abee, T., & Nierop Groot, M. N. (2015). Char-acterisation of biofilms formed by Lactobacillus plantarum WCFS1 and foodspoliage isolates. International Journal of Food Microbiology, 207, 23e29.
Food and Drug Administration. (2012). Hadbook of food bad bug book, foodbornepathogenic microorganisms and natural toxins. In K. Lampel, S. Al-Khaldi, &S. A. Assimon (Eds.), Pathogenic bacteria (2nd ed., pp. 8e81). US: FDA.
Health Protection Agency (HPA). (2009). Guidelines for assessing the microbiologicalsafety of ready-to-eat foods placed on the market. London: Health ProtectionAgency (November), 33.
Kennedy, D., Cronin, U. P., & Wilkinson, M. G. (2011). Responses of Escherichia coli,Listeria monocytogenes, and Staphylococcus aureus to simulated food pro-cessing treatments, determined using fluorescence-activated cell sorting andplate counting. Applied and Environmental Microbiology, 77(13), 4657e4668.
Lebert, I., Leroy, S., & Talon, R. (2007). Effects of industrial and natural biocides onsopiage, pathogenic and technological strains grown in biofilm. Food Microbi-ology, 24, 281e287.
Lindqvist, R., & Lindblad, M. (2009). Inactivation of Escherichia coli, Listeria mon-ocytogenes and Yersinia enterocolitica in fermented sausages during matura-tion/storage. International Journal of Food Microbiology, 129(1), 59e67.
Ma, Q., Davidson, M., & Zhong, Q. (2013). Antimicrobial properties of lauric arginatealone or in combimation with essential oils in tryptic soy broth and 2% reducedmilk. International Journal of Food Microbiology, 166, 77e84.
Maillard, J.-Y., Bloomfield, S., Coelho, J. R., Collier, P., Cookson, B.,Fanning, S.…Threlfall, J. (2013). Does microbicide use in consumer productspromote antimicrobial resistance? A critical review and recommendations for acohesive approach to risk assessment. Microbial Drug Resistance (Larchmont,N.Y.), 19(5), 344e354.
Martin, E. M., Griffis, K. J. S., O'Bryan, C. A., Friedly, E. C., Marcy, J. A., Ricke, S. C., et al.(2009). Control of Listeria monocytogenes by lauric arginate on franckfurtesformulated with or without lactate/Diacetate. Journal of Food Science, 74(6),M237eM241.
Matos, C. T., & Lopes Da Silva, T. (2013). Using multi-parameter flow cytometry as anovel approach for physiological characterization of bacteria in microbial fuelcells. Process Biochemistry, 48(1), 49e57.
Muriel-Galet, V., L�opez-Carballo, G., Gavarra, R., & Hern�andez-Mu~noz, P. (2015).Antimicrobial effectiveness of Lauroyl arginate incorporated into ethylene vinylalcohol copolymers to extend the self-life of chicken stock and surimi sticks.Food Bioprocess Technol, 8, 208e217.
Nesbakken, T., Eckner, K., & Røtterud, O. J. (2008). The effect of blast chilling onoccurrence of human pathogenic Yersinia enterocolitica compared toCampylobacter spp. and numbers of hygienic indicators on pig carcasses. In-ternational Journal of Food Microbiology, 123(1e2), 130e133.
Newell, D. G., Koopmans, M., Verhoef, L., Duizer, E., Aidara-Kane, A., Sprong, H., et al.(2010). Food-borne diseases - the challenges of 20years ago still persist whilenew ones continue to emerge. International Journal of Food Microbiology,139(Suppl. 1), S3eS15.
Novo, D. J., Perlmutter, N. G., Hunt, R. H., & Shapiro, H. M. (2000). Multiparameteflow cytometry analysis of antibiotic effects on membrane potential, membranepermeability, and bacterial counts of Staphylococcus aureus and Microcodccusluteus. Antimicrobial Agents and Chemotherapy, 44(4), 827e834.
Oliva Neto, P., Aliaga de Lima, F., da Silva, C., Fernandes da SIlva, D., AzevedoCarvhalo, A. F., et al. (2014). Chemical inhibition of the contaminant Lactoba-cillus fermentum from distikkeries producing fuel bioethanol. Brazilian Archivesof Biology and Technology, 573, 441e447.
Otero, V., Becerril, R., Santos, J. A., Rodríguez-Calleja, J. M., Nerín, C., & Garcíia-L�opez, M. L. (2014). Evaluation of two antimicrobial packaging films againstEscherichia coli O157:H7 strain in vitro and during storage os a Spanish ripenedsheep cheese (Zamorano). Food Control, 42, 296e302.
Pattanayaiying, R., H-Kittikun, A., & Cutter, C. N. (2014). Effect of lauric arginate,nisin Z, and a combination against several food-related bacteria. InternationalJournal of Food Microbiology, 188, 135e146.
Rodríguez, E., Seguer, J., Rocabayera, X., & Manresa, A. (2004). Cellular effects ofmonohydrochloride of L-arginine, Na- lauroyl ethylester (LAE) on exposure toSalmonella Typhimurium and Staphylococcus aureus. Journal of AppliedMicrobiology, 96, 903e912.
Skjerve, E., Lium, B., Nielsen, B., & Nesbakken, T. (1998). Control of Yersinia enter-ocolitica in pigs at herd level. International Journal of Food Microbiology, 45(3),195e203.
Stopforth, J. D., Visser, D., Zumbrink, R., Dijk van, L., & Bontebal, E. W. (2010). Controlof Listeria monocytogenes on cooked cured ham by formulation with a lactate-diacetate blend and surface treatment with lauric arginate. Jorunal of FoodProtection, 33(3), 522e555.
Suksathit, S., & Tangwatcharin, P. (2013). Activity of organic acid salts in combi-nation with lauric arginate against Listeria monocytogenes and Salmonella

J. Coronel-Le�on et al. / Food Control 63 (2016) 1e1010
Rissen. Science Asia, 39, 345e355.Swenson, J. M., Fick Hindler, J., & Jogernsen, J. H. (2003). Special phenotipic methods
for detecting antibacterial resistance. In P. R. Murray, E. J. Baron, M. A. Pfaller,J. H. F. C. Jorgensen, & R. H. Yolken (Eds.),Manual of clinical microbiology (8th ed.,pp. 1178e1195). Washington: ASM Press.
Theinsathid, M. P., Visessanguan, W., Kruenate, J., Kingcha, Y., & Keeratipibul, S.(2012). Antimicrobial activity of lauric arginate-coated polylactic acid filmsagainst Listeria monocytogenes and Salmonella Thiphimurium on cooked slicedham. Journal of Food Science, 77(2), M143eM149 (Washington, ASM Press).
USDA. (2005). Agency response letter GRAS notice No 000164. Available at: http//
www.accessdata.fda.org/scripts/fcn/gras_notices/grn000164.pdf (Last accessedon 09.15).
Woods, G. L., & Washington, J. A. (2003). Susceptibility test methods: dilution anddisk diffusion methods. In P. R. Murray, E. J. Baron, M. A. Pfaller, J. H.F. C. Jorgensen, & R. H. Yolken (Eds.), Manual of clinical microbiology (8th ed., pp.1108e1125). Washington: ASM Press.
Ye, Q., Wu, Q., Hu, H., Zhang, J., & Huang, H. (2016). Prevalence and characterizationof Yersinia enterocolitica isoalted from retail foods in China. Food Control, 61,20e27.





























