Embed Size (px)
Citation preview

Architecture and organizationalprinciples of Broca’s regionKatrin Amunts1,2,3 and Karl Zilles1,2,4
1 Research Centre Julich, Institute of Neuroscience and Medicine (INM-1), Julich, Germany2 Julich-Aachen Research Alliance (JARA), Translational Brain Medicine, Julich, Germany3 Section Structural-functional Brain Mapping, Department of Psychiatry, Psychotherapy, and Psychosomatics, Medical School,
RWTH Aachen University, Aachen, Germany4 C. & O. Vogt Institute for Brain Research, University of Dusseldorf, Dusseldorf, Germany
Identifying cortical areas for language and speech pro-cessing is a prerequisite for cognitive neuroscience andclinical research. Although Broca’s region is one of theessential nodes in the language network, its anatomicalconstituents are ill-defined and multiple definitions ofBroca’s region exist. Sanides’ concept of microstructuralgradations interpreted Broca’s region as developingfrom neighboring motor, dorsolateral-prefrontal, andinsular cortices. Recent mapping approaches based oncytoarchitecture, transmitter receptor distribution, andconnectivity revealed a highly differentiated segregationof this region far beyond Brodmann’s classical scheme.This novel segregational concept of structural and func-tional architecture more adequately reflects the variousfunctions of Broca’s region in cognitive and/or linguisticprocesses.
IntroductionThe term Broca’s region originates from a clinico-pathological examination of a patient, Leborgne (‘Tan’),who lost speech. The neurologist Paul Broca [1] delivered ashort presentation at the Anthropological Society of Parisin 1861 after Leborgne’s death, where he reported that theloss of speech, aphemia, was related to a lesion in theposterior part of the left third frontal convolution, latercalled Broca’s region. The lesion was large and occupiedmajor parts of the inferior frontal convolution, as well asparts of the basal ganglia, the white matter, and moreposterior regions [2,3] (Figure 1). Broca’s observation wasnot without antecedents: it was preceded by studies byFranz Josef Gall, Johann Casper Spurzheim, Gustav Dax,Louis P. Gratiolet, Jean Baptiste Bouillaud, Ernest Aubertinand others, who had published earlier observations ofpatients with aphasia after brain injury (see [4,5], for ahistorical review).
Broca’s case did not only provide strong arguments forthe localization of language, but also showed a hemisphericspecialization in the brain. Determining its anatomicalunderpinnings has been the aim of a large number ofstudies. Keller and colleagues [6] provided an overviewof gross anatomical and cytoarchitectonic studies targetingasymmetry. One of the strongest pieces of evidence for
Review
Glossary
Agranular cortex: part of the isocortex, where the internal granular layer (IV)
has been reduced during ontogeny to such a degree that it is no longer visible
as a distinct layer. For example, Brodmann areas 4 and 6 as part of the motor
and premotor cortex.
Convergence zones: Binder and Desai [68] recently proposed high-level
convergence zones in the inferior parietal, lateral temporal, and ventral
temporal cortex. They note that these zones are far from primary sensory
and motor cortices, and appear to be involved in processing general rather
than modality-specific semantic information. Other models of convergence
zones have been proposed elsewhere [91].
Cortical areas and cortical map: the cerebral cortex is composed of areas,
which are characterized by a certain microstructure, for example, cytoarchi-
tecture, connectivity, and brain function. Various cortical maps and parcella-
tion schemes have been put forward during the past century, which differ in
the number of areas, their ontology, and the aspect of brain organization that
they reflect (e.g., cytoarchitecture, myeloarchitecture, molecular architecture,
pigmentoarchitecture).
Cytoarchitecture: organizational principle of the cerebral cortex and subcortical
nuclei, which considers differences in the distribution of cell bodies, their size,
and shape between cortical areas and subcortical nuclei. Cytoarchitecture is
revealed in cell-body stained histological sections (e.g., Nissl-stain with cresyl
violet, Merker-silver stain), which are the basis of the first cortical maps, that is,
that of Brodmann from 1909 [70] and other early maps of Broca’s region [22,23].
Dysgranular cortex: part of the isocortex, where the internal granular layer (IV)
is not clearly visible as a distinct layer due to intermingling of granular cells from
layer IV and pyramidal cells from deep layer III and superficial layer V. Brodmann
area 44, which is located rostrally to the premotor cortex, is an example of
dysgranular cortex. This area is interpreted as a core part of Broca’s region.
Gradations: streams along which cytoarchitectonic features change from one
region to another. The term and the underlying concept were introduced by
Sanides [57,58], and based on earlier observations of the Vogt’s [27,93] and
Brockhaus [94]. Gradations correspond to evolutionary pathways pointing from
phylogenetically older to more recent cortical areas. According to this concept,
Broca’s region originates from motor precentral, opercular/insular and more
dorsally located lateral-frontal areas. This complex composition makes sense
considering the variety of brain functions associated with Broca’s region.
Granular cortex: part of the isocortex with clearly visible internal granular layer
(IV). Brodmann area 45 (another core part of Broca’s region), primary sensory
cortices, fronto-polar cortex, and other associational cortices are examples of
granular cortex.
Isocortex: six-layered cortex covering major parts of the lateral and medial
aspect of the brain surface. It coincides with the phylogenetic term ‘neocortex’.
This type of cortex has the following layers: molecular (I), external granular (II),
external pyramidal (III), internal granular (IV), internal pyramidal (V), and
multiforme (VI). Each layer has a specific connectivity with other layers and
areas. The isocortex is subdivided into several dozens to hundreds of cortical
areas in the various available maps.
Myeloarchitecture: organizational principle of the cerebral cortex and sub-
cortical nuclei, which considers differences in the intracortical distribution of
myelinated nerve fibers. This type of architecture was used to create maps of
the whole brain and Broca’s region, in particular [27,29]. It has attracted
increasing interest in the context of recent high-resolution MRI.
Receptorarchitecture: organizational principle of the cerebral cortex and
subcortical nuclei, which considers differences in the distribution of receptor
types of different transmitter receptors, such as glutamate, acetylcholine or
anatomical asymmetry comes from studies analyzing the serotonin [79]. This architecture is most closely related to brain function, sincetransmitter receptors are key molecules of signal transduction.
Corresponding author: Amunts, K. ([email protected]).
418 1364-6613/$ – see front matter � 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tics.2012.06.005 Trends in Cognitive Sciences, August 2012, Vol. 16, No. 8

(a) (b)
sfs
ifs
ifs
sfs
TRENDS in Cognitive Sciences
Figure 1. Dorsal view of a 3D reconstruction of an MR dataset of Leborgne’s brain (a) and coronal sections through Broca’s region (ellipse) at the level of the posterior part
of the inferior frontal gyrus (b). Note the deformation of the interhemispheric fissure caused by brain fixation and the effects of the large lesion, which included not only
cortical tissue of the posterior part of the inferior frontal gyrus, but also a large portion of the white matter and basal ganglia. The MR data set was provided by Dronkers,
Plaisant, Iba-Zizen and Cabanis [3], and the figure was created in collaboration with Nina Dronkers and Odile Plaisant. Abbreviations: ifs, inferior frontal sulcus; sfs, superior
frontal sulcus.
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
volume of cytoarchitectonic area 44, which is larger in theleft than in the right hemisphere, in a proportion that issimilar to left-hemispheric dominance for language [7,8]; incontrast, studies of gross anatomical asymmetry of theanterior speech region came to more controversial conclu-sions [6]. More recently, asymmetry has also been demon-strated for the distribution of the cholinergic M2-receptorfor the neurotransmitter acetylcholine [9] and for theunderlying fiber tracts (see below).
Leborgne’s brain was never subjected to any histologicalanalysis, and thus, the extent of the lesion site could beassigned only to surface landmarks. This is a severerestriction for further analysis of structural-functionalcorrelations, since non-primary sulci and borders of micro-structurally defined areas vary to a considerable degreeand independently from each other [8,10]. Moreover, theydiffer between hemispheres [6]. Significant variability isalso found in the localization and extent of brain lesions indifferent cases [11,12]. As a consequence, we can onlyspeculate about the cortical areas (see Glossary) contribut-ing to the speech deficit in the patients described by Brocaand others, since the definition of Broca’s region remainsvague in anatomical terms.
Presently, different definitions of Broca’s region co-exist[6]. Here, we use the term ‘Broca’s region’ for a region onthe inferior frontal gyrus, including parts of surroundingcortices. We will argue that there are also no robustfunctional criteria to define Broca’s region unambiguously.There is an ongoing debate about the selective engagementof this region in language versus a multifunctional involve-ment in cognition [13].
The solution to such controversies requires a clearconcept of the neuroanatomical organization of Broca’sregion and other regions involved in language. To addressthis point, we will briefly review the literature concerningthe architecture of Broca’s region, compare existing mapsand parcellation schemes with their underlying concepts,
analyze their relevance for recent studies, and discuss newanatomical concepts of Broca’s region based on recentstructural, functional, and connectivity studies of themacaque and living human brain.
Classical maps of Broca’s regionCyto- and myeloarchitectonic maps are relevant for thedefinition of Broca’s region, because they represent notonly topographical aspects of the cerebral cortex, but alsoreflect organizational principles, connectivity, cellularcomposition, comparative neuroanatomical concepts, aswell as physiological and pathological aspects. Each archi-tectonic area is a unique part of the brain, comprisingcorrelated structural and functional properties. KorbinianBrodmann, a pioneer of cytoarchitectonics, was convincedthat each of his more than forty cytoarchitectonic areashas a unique function and evolutionary position [14]. Forexample, he proposed area 4 as the anatomical equivalentof the primary motor cortex, which guides voluntarymovements [15]. This concept was already supported byelectrophysiological observations at that time. Brodmannviewed Broca’s region as the anatomical correlate of thefunctionally defined anterior center of speech. However,the relationship between microscopic structure, forinstance, cytoarchitecture, and function had never beenrigorously tested in this region during Brodmann’s time. Aprecise anatomical, that is, microstructurally based, mapis therefore a necessary prerequisite for any topographi-cally specific interpretation of (language-related) brainactivity and for a common structural and functionalconceptualization of brain and language.
Many attempts to map Broca’s region and surroundingcortices were made at the beginning of the 20th century byusing different methods, theoretical concepts of corticalparcellation, and different nomenclature (Table 1). Someof these maps addressed the whole cerebral cortex, forexample, the cytoarchitectonic maps of Brodmann [14],
419
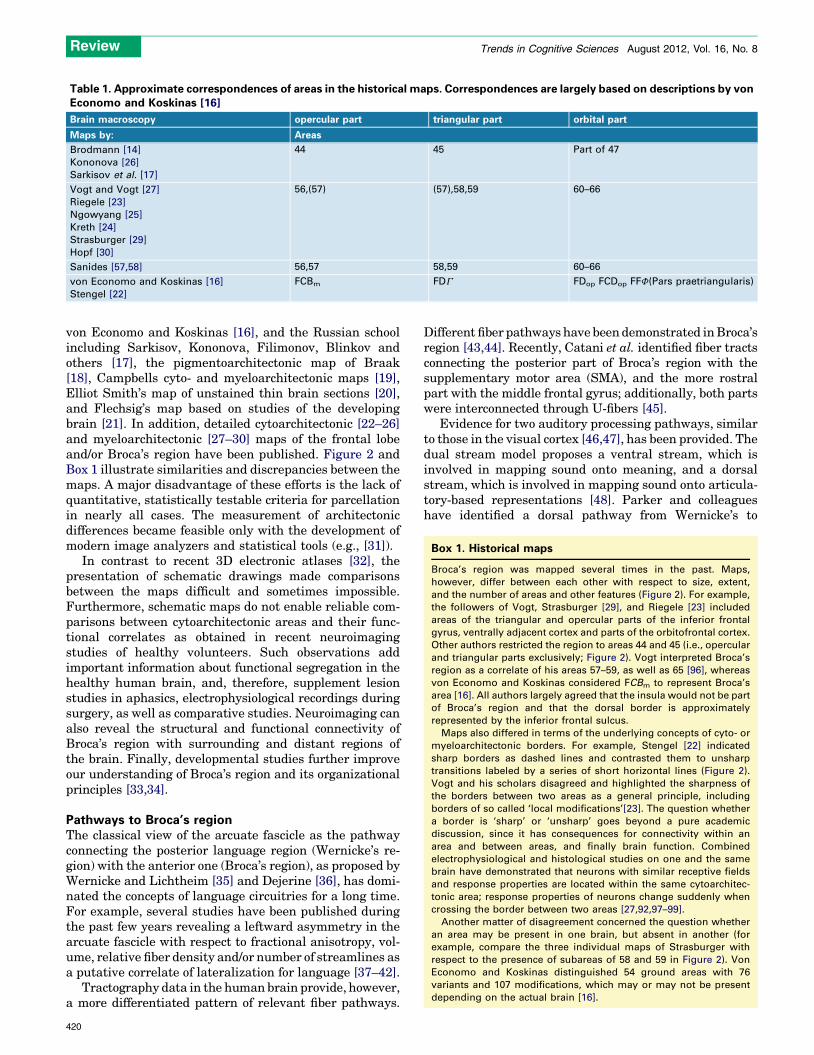
Table 1. Approximate correspondences of areas in the historical maps. Correspondences are largely based on descriptions by vonEconomo and Koskinas [16]
Brain macroscopy opercular part triangular part orbital part
Maps by: Areas
Brodmann [14]
Kononova [26]
Sarkisov et al. [17]
44 45 Part of 47
Vogt and Vogt [27]
Riegele [23]
Ngowyang [25]
Kreth [24]
Strasburger [29]
Hopf [30]
56,(57) (57),58,59 60–66
Sanides [57,58] 56,57 58,59 60–66
von Economo and Koskinas [16]
Stengel [22]
FCBm FDG FDop FCDop FFF(Pars praetriangularis)
Box 1. Historical maps
Broca’s region was mapped several times in the past. Maps,
however, differ between each other with respect to size, extent,
and the number of areas and other features (Figure 2). For example,
the followers of Vogt, Strasburger [29], and Riegele [23] included
areas of the triangular and opercular parts of the inferior frontal
gyrus, ventrally adjacent cortex and parts of the orbitofrontal cortex.
Other authors restricted the region to areas 44 and 45 (i.e., opercular
and triangular parts exclusively; Figure 2). Vogt interpreted Broca’s
region as a correlate of his areas 57–59, as well as 65 [96], whereas
von Economo and Koskinas considered FCBm to represent Broca’s
area [16]. All authors largely agreed that the insula would not be part
of Broca’s region and that the dorsal border is approximately
represented by the inferior frontal sulcus.
Maps also differed in terms of the underlying concepts of cyto- or
myeloarchitectonic borders. For example, Stengel [22] indicated
sharp borders as dashed lines and contrasted them to unsharp
transitions labeled by a series of short horizontal lines (Figure 2).
Vogt and his scholars disagreed and highlighted the sharpness of
the borders between two areas as a general principle, including
borders of so called ‘local modifications’[23]. The question whether
a border is ‘sharp’ or ‘unsharp’ goes beyond a pure academic
discussion, since it has consequences for connectivity within an
area and between areas, and finally brain function. Combined
electrophysiological and histological studies on one and the same
brain have demonstrated that neurons with similar receptive fields
and response properties are located within the same cytoarchitec-
tonic area; response properties of neurons change suddenly when
crossing the border between two areas [27,92,97–99].
Another matter of disagreement concerned the question whether
an area may be present in one brain, but absent in another (for
example, compare the three individual maps of Strasburger with
respect to the presence of subareas of 58 and 59 in Figure 2). Von
Economo and Koskinas distinguished 54 ground areas with 76
variants and 107 modifications, which may or may not be present
depending on the actual brain [16].
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
von Economo and Koskinas [16], and the Russian schoolincluding Sarkisov, Kononova, Filimonov, Blinkov andothers [17], the pigmentoarchitectonic map of Braak[18], Campbells cyto- and myeloarchitectonic maps [19],Elliot Smith’s map of unstained thin brain sections [20],and Flechsig’s map based on studies of the developingbrain [21]. In addition, detailed cytoarchitectonic [22–26]and myeloarchitectonic [27–30] maps of the frontal lobeand/or Broca’s region have been published. Figure 2 andBox 1 illustrate similarities and discrepancies between themaps. A major disadvantage of these efforts is the lack ofquantitative, statistically testable criteria for parcellationin nearly all cases. The measurement of architectonicdifferences became feasible only with the development ofmodern image analyzers and statistical tools (e.g., [31]).
In contrast to recent 3D electronic atlases [32], thepresentation of schematic drawings made comparisonsbetween the maps difficult and sometimes impossible.Furthermore, schematic maps do not enable reliable com-parisons between cytoarchitectonic areas and their func-tional correlates as obtained in recent neuroimagingstudies of healthy volunteers. Such observations addimportant information about functional segregation in thehealthy human brain, and, therefore, supplement lesionstudies in aphasics, electrophysiological recordings duringsurgery, as well as comparative studies. Neuroimaging canalso reveal the structural and functional connectivity ofBroca’s region with surrounding and distant regions ofthe brain. Finally, developmental studies further improveour understanding of Broca’s region and its organizationalprinciples [33,34].
Pathways to Broca’s regionThe classical view of the arcuate fascicle as the pathwayconnecting the posterior language region (Wernicke’s re-gion) with the anterior one (Broca’s region), as proposed byWernicke and Lichtheim [35] and Dejerine [36], has domi-nated the concepts of language circuitries for a long time.For example, several studies have been published duringthe past few years revealing a leftward asymmetry in thearcuate fascicle with respect to fractional anisotropy, vol-ume, relative fiber density and/or number of streamlines asa putative correlate of lateralization for language [37–42].
Tractography data in the human brain provide, however,a more differentiated pattern of relevant fiber pathways.
420
Different fiber pathways have been demonstrated in Broca’sregion [43,44]. Recently, Catani et al. identified fiber tractsconnecting the posterior part of Broca’s region with thesupplementary motor area (SMA), and the more rostralpart with the middle frontal gyrus; additionally, both partswere interconnected through U-fibers [45].
Evidence for two auditory processing pathways, similarto those in the visual cortex [46,47], has been provided. Thedual stream model proposes a ventral stream, which isinvolved in mapping sound onto meaning, and a dorsalstream, which is involved in mapping sound onto articula-tory-based representations [48]. Parker and colleagueshave identified a dorsal pathway from Wernicke’s to

Brodmann, 1909 Sarkisov, 1949 Vogt, 1910 Vogt, 1951
Ngowyang, 1934Riegele, 1931Sanides, 1962
von Economo and Koskinas, 1925
Strasburger, 1938
Kreht, 1935Hopf, 1956Stengel, 1930
TRENDS in Cognitive Sciences
Figure 2. Overview of historical architectonic maps of the cerebral cortex with a focus on Broca’s region. In order to compare the different maps more easily with each
other, Brodmann areas 44 and 45 and the corresponding areas in the other maps were colored in red and yellow, respectively. The equivalence of Brodmann areas 44 and
45 to areas of researchers applying a different nomenclature and parcellation is based on von Economo and Koskinas, who carefully compared the different architectonic
descriptions and illustrations found in the literature [16]. Other authors sometimes assumed slightly different correspondences, for example, with respect to Vogt’s area 57:
Riegele [23] emphasizes the similarity of area 57 with 58 as compared to 56, whereas Ngowyang [25] interprets 56 as the caudal area 44 of Brodmann, and 57 its rostral part.
In other words, in his understanding the similarity of 57 to 56 is higher than to 58 (which belongs to Brodmann area 45). The map of Sarkisov [17] is, with respect to the
frontal lobe, very much based on the work of Kononova [26].
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
421

Box 2. Sanides’ concept of gradations
The concept is based on earlier short notes by the Vogt’s [27,93] and
later Brockhaus [94]. They observed that certain architectonic
features change from area to area along well-defined developmental
lines in a stepwise manner (Figure 2). Brockhaus called them
directions of differentiation, but did not yet interpret them as a
developmental principle. Gradations, according to Sanides [57,58],
reflect architectonic similarities of a stream of neighboring areas,
which change in a stepwise manner. Sanides subdivided the frontal
lobe into different zones and identified gradations for each of them.
The ‘fronto-opercular zone’ (FoZ; Figure 59 in Sanides [57]; Figure 2
in the present review) roughly corresponds to Broca’s region. It
represents a zone where several gradations converge (Figure 3):
(a) in caudo-rostral direction: from the frontomotor zone (FmZ;
agranular motor and premotor cortex (Vogt area 40/41) to Vogt
dysgranular area 56 and granular area 58;
(b) in ventro-dorsal direction: from the insula via the opercular
cortex to Vogt areas 56 and 58;
(c) in dorso-ventral direction: from more dorsally located prefrontal
cortex of the paraopercular zone (PoZ) to Vogt area 58.
To provide an example of a cytoarchitectonic gradation – the
gradation from the motor cortex towards the more rostrally located
frontal areas (a) was characterized by an increasing visibility of the
laminar pattern, a decrease in thickness of layers III and V, an
increasing subdivision of layer V into 2 sublayers, and an increasing
granularity in layer IV. These cytoarchitectonic changes are
accompanied by myeloarchitectonic changes. Interestingly, the
FoZ does not receive any gradations from more rostrally located
areas. Arrows of gradation seem to stop area 53 v, belonging to the
most ventral part of the para-opercular zone (PoZ) (Figure 2).
In addition to the embedding of Broca’s region into neighboring
areas, Sanides’ concept of gradations represents a framework for
understanding the internal organization of Broca’s region. However,
Sanides did not quantify these observations, including the definition
of architectonic borders. Moreover, if gradations are a general
principle of cortical organization, it can be hypothesized that other
aspects of cortical structure follow similar streams of gradations.
Sanides explained the cortical development and areal specialization
from an evolutionary perspective, and he introduced hierarchical
aspects of cortical organization as compared to the mosaic-like
maps of his predecessors (e.g., Brodmann).
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
Broca’s region that includes the arcuate fascicle and con-nectivity to Brodmann area 40, lateral superior and middletemporal gyri, and a ventral pathway that runs via theexternal capsule/uncinate fascicle [41]. A recent reviewdiscusses four major streams: dorsal pathway I connectsthe superior temporal gyrus to the premotor cortex via thearcuate fascicle and the superior longitudinal fascicle;dorsal pathway II connects the superior temporal gyrusto Brodmann area 44 via the arcuate and superior longi-tudinal fascicles; ventral pathway I connects Brodmannarea 45 and the temporal cortex via the extreme capsule,whereas ventral pathway II connects the frontal opercu-lum and the anterior parts of the superior temporal gyrusand sulcus via the uncinate fascicle [49]. Most importantly,these models do not only distinguish fiber pathways con-necting brain regions with each other, but explicitly ascriberoles to these regions in speech processing and language[43]. For example, dorsal pathway I subserves auditory-motor integration, whereas dorsal pathway II seems to befunctionally related to higher-level semantic and syntacticlanguage functions [49].
Limitations with respect to the interpretation of DTIdata arise from the following facts: (i) the directionality offiber tracts cannot be obtained from DTI studies; (ii) thespatial resolution is in the range of millimeters, and track-ing results, therefore, refer to gross macroscopical unitssuch as gyri, but not cortical areas; and (iii) connectivity ina strict sense requires verification at the synaptic level.
These are reasons why studies in experimental animals,in particular the macaque brain, are highly important,although the open question of homologies between ma-caque and human brain areas involved in language isparticularly relevant [50]. Similarities in connectivity be-tween macaque and human brains have been demonstrat-ed, for example, the presence of an anterior and posteriorpart of area 45 [51,52]. (Dis-)similarities between humanand macaque brains have been discussed, as well [53].Differences between human brains and those of non-hu-man primates concern, for instance, the ability to processcomplex sentence structures. This difference seems to beneuroanatomically linked to the dorsal pathway connect-ing the posterior part of Broca’s area and the posteriorsuperior temporal gyrus/sulcus [54,55].
Petrides and Pandya [51] have shown that area 44 of themacaque brain receives strong input from area PFG of theinferior parietal lobule, an area that corresponds to aregion in the caudal human supramarginal gyrus. Bycontrast, the caudally adjacent premotor area 6 mainlyhas connections with area PF, which is located morerostrally in the inferior parietal lobule [56], and seemsto control orofacial muscles. In addition, area 44 receivesinput from the caudal inferior parietal lobule, primarilyarea PG, which is located at the angular gyrus in thehuman brain [56]. Both subdivisions of area 45 receiveparietal input from PFG and PG via the second and thirdsuperior longitudinal fascicles (SLF II and SLF III). Someof the axons originating from the most ventral part of areaPG belong to the arcuate fascicle. In contrast to this dorsalstream, the ventral pathway is linked to the extremecapsule, originates from areas of the superolateral tempo-ral cortex, and terminates primarily in area 45 [51]. The
422
authors of this study concluded that precursor circuitshave been identified in the nonlinguistic human primatebrain, which help to understand the circuits, mechanisms,and evolutionary development of language processing inthe human brain. Is such an evolutionary perspective alsoinherent in the architectonic segregation of Broca’s region?Does the architectonic segregation of the human cerebralcortex enable the identification of developmental lines,along which evolutionarily more recent areas differentiatedfrom older ones?
Sanides’ gradations and Broca’s regionSanides contributed a new view on principles ofcytoarchitectonic segregation of the cortex, the conceptof ‘gradations’ [57,58] (Box 2). Gradations are spatiallydirected sequences of cortical areas, which originate, inmost cases, in phylogenetically old regions of the cortex.The gradation theory provides a framework for the identi-fication of the source region, from which language-relatedcortical areas originated. Three gradation streams arrive atBroca’s region (Figure 3 and Box 2):(a) Caudo-rostrally directed gradation from the fronto-
motor zone to Broca’s region. The close topographicaland functional relationship between motor and
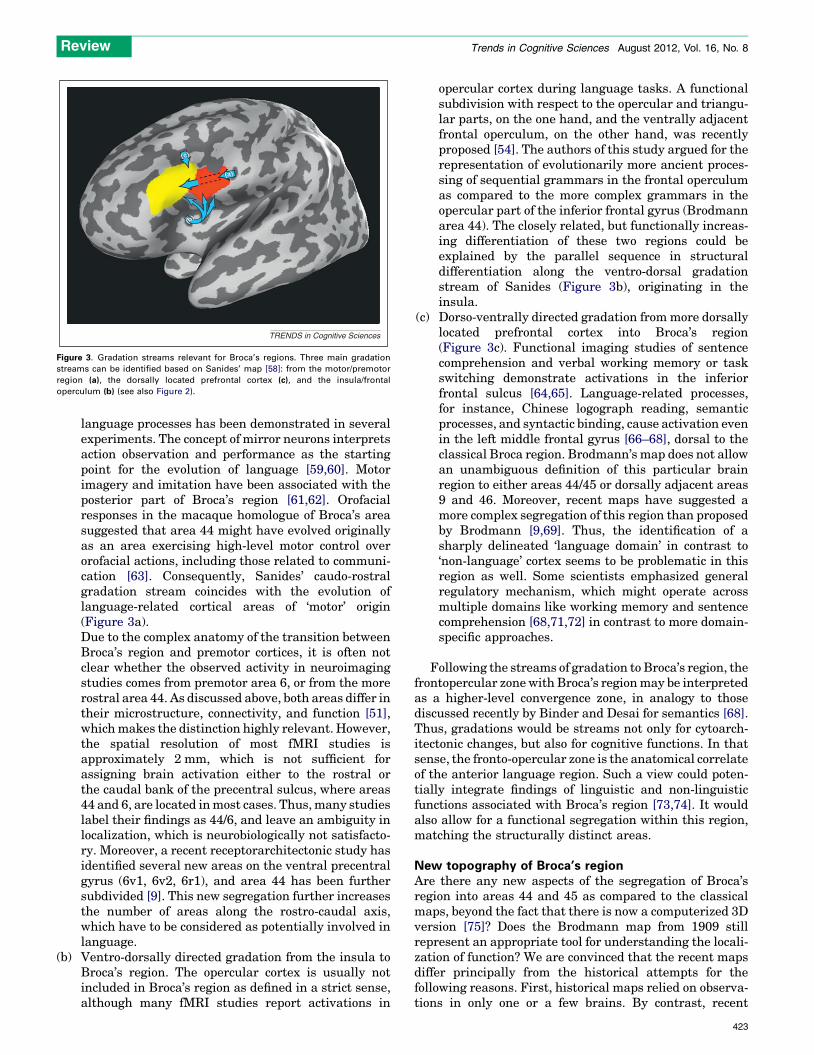
TRENDS in Cognitive Sciences
(c)(c)
(a)(a)
(b)
(b)
Figure 3. Gradation streams relevant for Broca’s regions. Three main gradation
streams can be identified based on Sanides’ map [58]: from the motor/premotor
region (a), the dorsally located prefrontal cortex (c), and the insula/frontal
operculum (b) (see also Figure 2).
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
language processes has been demonstrated in severalexperiments. The concept of mirror neurons interpretsaction observation and performance as the startingpoint for the evolution of language [59,60]. Motorimagery and imitation have been associated with theposterior part of Broca’s region [61,62]. Orofacialresponses in the macaque homologue of Broca’s areasuggested that area 44 might have evolved originallyas an area exercising high-level motor control overorofacial actions, including those related to communi-cation [63]. Consequently, Sanides’ caudo-rostralgradation stream coincides with the evolution oflanguage-related cortical areas of ‘motor’ origin(Figure 3a).Due to the complex anatomy of the transition betweenBroca’s region and premotor cortices, it is often notclear whether the observed activity in neuroimagingstudies comes from premotor area 6, or from the morerostral area 44. As discussed above, both areas differ intheir microstructure, connectivity, and function [51],which makes the distinction highly relevant. However,the spatial resolution of most fMRI studies isapproximately 2 mm, which is not sufficient forassigning brain activation either to the rostral orthe caudal bank of the precentral sulcus, where areas44 and 6, are located in most cases. Thus, many studieslabel their findings as 44/6, and leave an ambiguity inlocalization, which is neurobiologically not satisfacto-ry. Moreover, a recent receptorarchitectonic study hasidentified several new areas on the ventral precentralgyrus (6v1, 6v2, 6r1), and area 44 has been furthersubdivided [9]. This new segregation further increasesthe number of areas along the rostro-caudal axis,which have to be considered as potentially involved inlanguage.
(b) Ventro-dorsally directed gradation from the insula toBroca’s region. The opercular cortex is usually notincluded in Broca’s region as defined in a strict sense,although many fMRI studies report activations in
opercular cortex during language tasks. A functionalsubdivision with respect to the opercular and triangu-lar parts, on the one hand, and the ventrally adjacentfrontal operculum, on the other hand, was recentlyproposed [54]. The authors of this study argued for therepresentation of evolutionarily more ancient proces-sing of sequential grammars in the frontal operculumas compared to the more complex grammars in theopercular part of the inferior frontal gyrus (Brodmannarea 44). The closely related, but functionally increas-ing differentiation of these two regions could beexplained by the parallel sequence in structuraldifferentiation along the ventro-dorsal gradationstream of Sanides (Figure 3b), originating in theinsula.
(c) Dorso-ventrally directed gradation from more dorsallylocated prefrontal cortex into Broca’s region(Figure 3c). Functional imaging studies of sentencecomprehension and verbal working memory or taskswitching demonstrate activations in the inferiorfrontal sulcus [64,65]. Language-related processes,for instance, Chinese logograph reading, semanticprocesses, and syntactic binding, cause activation evenin the left middle frontal gyrus [66–68], dorsal to theclassical Broca region. Brodmann’s map does not allowan unambiguous definition of this particular brainregion to either areas 44/45 or dorsally adjacent areas9 and 46. Moreover, recent maps have suggested amore complex segregation of this region than proposedby Brodmann [9,69]. Thus, the identification of asharply delineated ‘language domain’ in contrast to‘non-language’ cortex seems to be problematic in thisregion as well. Some scientists emphasized generalregulatory mechanism, which might operate acrossmultiple domains like working memory and sentencecomprehension [68,71,72] in contrast to more domain-specific approaches.
Following the streams of gradation to Broca’s region, thefrontopercular zone with Broca’s region may be interpretedas a higher-level convergence zone, in analogy to thosediscussed recently by Binder and Desai for semantics [68].Thus, gradations would be streams not only for cytoarch-itectonic changes, but also for cognitive functions. In thatsense, the fronto-opercular zone is the anatomical correlateof the anterior language region. Such a view could poten-tially integrate findings of linguistic and non-linguisticfunctions associated with Broca’s region [73,74]. It wouldalso allow for a functional segregation within this region,matching the structurally distinct areas.
New topography of Broca’s regionAre there any new aspects of the segregation of Broca’sregion into areas 44 and 45 as compared to the classicalmaps, beyond the fact that there is now a computerized 3Dversion [75]? Does the Brodmann map from 1909 stillrepresent an appropriate tool for understanding the locali-zation of function? We are convinced that the recent mapsdiffer principally from the historical attempts for thefollowing reasons. First, historical maps relied on observa-tions in only one or a few brains. By contrast, recent
423

ifj2
ifj1
ifs1
ifs2
ifs
ifs
cs
prcsprcsds
ab
hb
44v6r1
6v1
6v2
4
47
44d
45p
45a
op8
op6
op9
op4
Amunts et al., 2010
Amunts et al., 1999
(a) (b)
(c)
0.2 0.3 0.4 0.5 0.6 0.7 0.8Distance
Cor
tical
are
a
47
4
6r1
6v1
op9
op8
45
44
45
TRENDS in Cognitive Sciences
Figure 4. (a) The map of Broca’s region based on the distribution of receptors of neurotransmitters and modulators shows a complex segregation of the inferior frontal
gyrus and adjacent cortices [9]. In particular, it shows the presence of a series of new areas in the inferior frontal sulcus (ifs) labeled as areas ifs1, ifs2, ifj1 and ifj2, as well in
the frontal operculum (‘op’-areas). The areas of this region can be clustered based on their receptorarchitectonic pattern as shown in (b). Transmitter receptors are key
molecules for neurotransmission, and therefore, indicative of function. Areas 44 and 45 of the classical Broca region are most similar to each other and they cluster with
areas op8 and op9 of the frontal operculum. Interestingly, area 47 is the most different of all analyzed areas, suggesting distinct function [9]. (c) Cytoarchitectonic
probabilistic map of right area 45 [8] in the standard reference space of the single subject template from the MNI [95]. The color indicates the overlap of individually mapped
area 45 in common reference space. Note the high intersubject variability (labeled in blue and green) of area 45 in regions outside the free surface of the triangular part
(orange and red colors).
Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
cytoarchitectonic probabilistic maps are based on anobserver-independent mapping in a sample of ten brains;they provide population-based information of an area instandard reference space, enabling a reliable comparison ofthe cytoarchitectonic areas with language function (e.g.,[76]) in different brains. This is relevant, since (i) inter-subject variability in brain anatomy is significant, andadds to variability in behavior and brain function, and(ii) relating microstructural areas to specific functions iscrucial for understanding brain organization.
Regional differences in connectivity and cytoarchitec-tonics of Broca’s region have been amplified by differencesin molecular organization, leading to new parcellationschemes. A recent study on the distribution of receptorsof different neurotransmitters and modulators has shownthat Broca’s region can be subdivided into more than adozen areas [9]. These areas include subdivisions of area 44and 45, a series of areas located in the depth of the inferiorfrontal sulcus, the frontal operculum, and at the point oftransition to the premotor cortex (Figure 4). The degree ofdetail of the parcellation of human inferior frontal gyrusand adjacent sulci matches that of the macaque brain [77],and goes beyond that in other maps [51]. Area F5a ofBelmalih and co-authors seems to be a privileged gateway
424
for the access of prefrontal and rostral opercular frontalinputs to the ventral premotor cortex [78], and may corre-spond to human area 6r1 [9]; the complete homology ofareas in human and macaque brains, however, is stillunresolved.
Apart from the mapping aspect, receptorachitectonicstudies also demonstrated that Broca’s region is organizedin a hierarchical way (Figure 4), similar to other corticalregions [79–81]. Hierarchical organization has also beenfound for other aspects of organization at a systemic level,including cytoarchitecture [82], connectivity [83,84], andgenetic topography [85]. Sanides’ concept of architectonicgradation is in line with these arguments [86]. Speechproduction, planning behavior, and executive control havebeen conceptualized as hierarchically organized, to provideonly a few examples [87–89]. These findings support thenotion of nested, hierarchical levels of organization [90].At the same time, the model does not exclude parallelprocessing and constant modification [88,90].
Concluding remarksThe data reviewed in this article indicate a complex segre-gation of Broca’s region. Its areas are unique with respectto their cellular architecture, myeloarchitecture, molecular

Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
signature, connectivity, and function. In addition to thesubdivision of ‘classical’ constituents of Broca’s region, thatis, areas 44 and 45, further areas of inferior frontal sulcus,frontal operculum and ventral premotor cortex have to betaken into account when searching for the anatomicalcorrelates of language processing. Area 47, which occupiesthe orbital part of the inferior frontal gyrus and adjacentorbitofrontal gyri, is receptorarchitectonically remarkablydistinct from areas 44/45 and the frontal operculum.Whether activations of this area during language proces-sing are correctly assigned to this area, or whether theybelong to one of the newly identified areas of the frontaloperculum, remains a project for future research.
Although Brodmann’s map looks simple and convincing,it does not reflect the richness of functional segregation asseen in functional imaging studies of language. The novelparcellation of Broca’s region is in accordance with thenotion that cortical areas do not simply form a mosaic,but are hierarchically organized. The understanding ofstructure-function relationships for language processingrequires both more comprehensive and differentiatedarchitectonic maps that takes into account their complexand nested hierarchical organization, and precise andreliable matching procedures between microstructuraldata and functional analyses.
References1 Broca, P. (1861) Remarques sur le siege de la faculte du langage articule,
suivies d’une observation d’aphemie (Perte de la Parole). Bull. Mem. Soc.Anat. Paris 36, 330–357 (in French)
2 Signoret, J-L. et al. (1984) Rediscovery of Leborgne’s brain: anatomicaldescription with CT scan. Brain Lang. 22, 303–319
3 Dronkers, N.F. et al. (2007) Paul Broca’s historic cases: high resolution MRimaging of the brains of Leborgne and Lelong. Brain 130, 1432–1441
4 Finger, S. (ed.) (1994) Origins of Neuroscience. A History of Explorationsinto Brain Function, Oxford University Press
5 Grodzinsky, Y. and Amunts, K., (eds) (2006) Broca’s Region, OxfordUniversity Press
6 Keller, S.S. et al. (2009) Broca’s area: nomenclature, anatomy, typologyand asymmetry. Brain Lang. 109, 29–48
7 Galaburda, A.M. (1980) La region de Broca: observations anatomiquesfaites un siecle apres la mort de son decoveur. Rev. Neurol. (Paris) 136,609–616 (in French)
8 Amunts, K. et al. (1999) Broca’s region revisited: cytoarchitecture andintersubject variability. J. Comp. Neurol. 412, 319–341
9 Amunts, K. et al. (2010) Broca’s region: novel organizational principlesand multiple receptor mapping. PLoS Biol. 8, e1000489
10 Tomaiuolo, F. et al. (1999) Morphology, morphometry and probabilitymapping of the pars opercularis of the inferior frontal gyrus: an in vivoMRI analysis. Eur. J. Neurosci. 11, 3033–3046
11 Dronkers, N.F. (1996) A new brain region for coordinating speecharticulation. Nature 384, 159–161
12 Schulze, H.A.F. (1966) Architektonische Untersuchungen zur Frageder ideokinetischen motorischen Aphasie. J. Hirnforsch. 8, 111–127(in German)
13 Fedorenko, E. et al. (2011) Functional specificity for high-levellinguistic processing in the human brain. Proc. Natl. Acad. Sci.U.S.A. 108, 16428–16433
14 Brodmann, K. (ed.) (1909) Vergleichende Lokalisationslehre derGroßhirnrinde in ihren Prinzipien dargestellt auf Grund desZellenbaues, Barth JA (in German)
15 Fritsch, G. and Hitzig, E. (1870) Uber die elektrische Erregbarkeit desGrosshirns. Arch. Anat. Physiol. Wiss. Med. 300–332 (in German)
16 von Economo, C. and Koskinas, G.N., (eds) (1925) DieCytoarchitektonik der Hirnrinde des Erwachsenen Menschen,Springer (in German)
17 Sarkisov, S.A. et al., (eds) (1949) Cytoarchitecture of the Human CortexCerebri (Russ.), Medgiz
18 Braak, H. (ed.) (1980) Architectonics of the Human TelencephalicCortex, Springer
19 Campbell, A.W. (ed.) (1905) Histological Studies on the Localisation ofCerebral Function, Cambridge University Press
20 Elliot Smith, G. (1907) A new topographical survey of the humancerebral cortex, being an account of the distribution of theanatomically distinct cortical areas and their relationship to thecerebral sulci. J. Anat. 41, 237–254
21 Flechsig, P. (ed.) (1927) Meine Myelogenetische Hirnlehre mitBiographischer Einleitung, Springer (in German)
22 Stengel, E. (1930) Morphologische und cytoarchitektonische studienu ber den Bau der unteren Frontalwindung bei Normalen undTaubstummen. Ihre individuellen und Seitenunterschiede. Z. Exp.Angew. Psychol. 130, 630–677 (in German)
23 Riegele, L. (1931) Die Cytoarchitektonik der Felder der Broca’schenRegion. J. Psychol. Neurol. 42, 496–514 (in German)
24 Kreth, H. (1936) Cytoarchitektonik und motorisches Sprachzentrum.Zeitschrift fu r mikroskopisch-anatomische Forschung 39, 331–354 (inGerman)
25 Ngowyang, G. (1934) Die Cytoarchitektonik des menschlichenStirnhirns. I. Teil. Cytoarchitektonische Felderung der Regiogranularis und Regio dysgranularis. In Monographs of the NationalResearch Institute of Psychology, Academia Sinica, pp. 1–69, TheNational Research Institute of Psychology Shanghai, China
26 Kononova, E.P. (1935) Structural variability of the cortex cerebri.Inferior frontal gyrus in adults (Russian). In Annals of theBrain Research Institute. Vol. I. (Sarkisov, S.A. and Filimonoff, I.N.,eds), pp. 49–118, State Press for Biological and Medical Literature
27 Vogt, C. and Vogt, O. (1919) Allgemeinere Ergebnisse unsererHirnforschung (English translation: results of our brain research ina broader context). J. Psychol. Neurol. 25, 292–398
28 Knauer, A. (1909) Die Myeloarchitektonik der Broca’schen Region.Neurol. Centralbl. 28, 1240–1243 (in German)
29 Strasburger, E.H. (1938) Vergleichende myeloarchitektonischeStudien an der erweiterten Brocaschen Region des Menschen. J.Psychol. Neurol. 48, 477–511 (in German)
30 Hopf, A. (1954) Zur architektonischen Gliederung der menschlichenHirnrinde. J. Hirnforsch. 1, 442–496 (in German)
31 Haug, H. (1979) The evaluation of cell-densities and nerve-cell sizedistribution by stereological procedures in a layered tissue (cortexcerebri). Microsc. Acta 82, 147–161
32 Toga, A.W. et al. (2006) Towards multimodal atlases of the humanbrain. Nat. Rev. Neurosci. 7, 952–966
33 Leroy, F. et al. (2011) Early maturation of the linguistic dorsal pathwayin human infants. J. Neurosci. 31, 1500–1506
34 Amunts, K. et al. (2003) Broca’s region: cytoarchitectonic asymmetryand developmental changes. J. Comp. Neurol. 465, 72–89
35 Lichtheim, L. (1885) On aphasia. Brain 7, 433–48436 Dejerine, J. (ed.) (1901) Anatomie des Centres Nerveux, Rueff (in
French)37 Wan, C.Y. et al. (2012) Atypical hemispheric asymmetry in the arcuate
fasciculus of completely nonverbal children with autism. Ann. N. Y.Acad. Sci. 1252, 332–337
38 Takao, H. et al. (2011) White matter asymmetry in healthy individuals:a diffusion tensor imaging study using tract-based spatial statistics.Neuroscience 193, 291–299
39 Powell, J.L. et al. (2012) The effect of sex and handedness on whitematter anisotropy: a diffusion tensor magnetic resonance imagingstudy. Neuroscience 207, 227–242
40 Thiebaut de Schotten, M. et al. (2011) Atlasing location, asymmetryand inter-subject variability of white matter tracts in the human brainwith MR diffusion tractography. Neuroimage 54, 49–59
41 Parker, G.J.M. et al. (2005) Lateralization of ventral and dorsalauditory-language pathways in the human brain. Neuroimage 24,656–666
42 Bu chel, C. et al. (2004) White matter asymmetry in the human brain: adiffusion tensor MRI study. Cereb. Cortex 14, 945–951
43 Friederici, A.D. (2009) Pathways to language: fiber tracts in the humanbrain. Trends Cogn. Sci. 13, 175–181
44 Anwander, A. et al. (2006) Connectivity-based parcellation of Broca’sarea. Cereb. Cortex 17, 816–825
45 Catani, M. et al. (2012) Short frontal lobe connections of the humanbrain. Cortex 48, 273–291
425

Review Trends in Cognitive Sciences August 2012, Vol. 16, No. 8
46 Rauschecker, J.P. (1998) Cortical processing of complex sounds. Curr.Opin. Neurobiol. 8, 516–521
47 Rauschecker, J.P. and Tain, B. (2000) Mechanisms and streams forprocessing of ‘what’ and ‘where’ in auditory cortex. Proc. Natl. Acad.Sci. U.S.A. 97, 11800–11806
48 Hickok, G. and Poeppel, D. (2004) Dorsal and ventral streams: aframework for understanding aspects of the functional anatomy oflanguage. Cognition 92, 67–99
49 Friederici, A.D. (2011) The brain basis of language processing: fromstructure to function. Physiol. Rev. 91, 1357–1392
50 Rauschecker, J.P. and Scott, S.K. (2009) Maps and streams in theauditory cortex: nonhuman primates illuminate human speechprocessing. Nat. Neurosci. 12, 718–724
51 Petrides, M. and Pandya, D.N. (2009) Distinct parietal and temporalpathways to the homologues of Broca’s area in the monkey. PLoS Biol.7, e1000170
52 Gerbella, M. et al. (2010) Cortical connections of the macaque caudalventrolateral prefrontal areas 45A and 45B. Cereb. Cortex 20, 168
53 Petrides, M. (2005) Lateral prefrontal cortex: architectonic andfunctional organization. Philos. Trans. R Soc. Lond. B: Biol. Sci.360, 781–795
54 Friederici, A.D. et al. (2006) The brain differentiates human and non-human grammars: functional localization and structural connectivity.Proc. Natl. Acad. Sci. U.S.A. 103, 2458–2463
55 Fitch, W.T. and Hauser, M.D. (2004) Computational constraints onsyntactic processing in a nonhuman primate. Science 303, 377380
56 Caspers, S. et al. (2006) The human inferior parietal cortex:cytoarchitectonic parcellation and interindividual variability.Neuroimage 33, 430–448
57 Sanides, F. (ed.) (1962) Die Architektonik des Menschlichen Gehirns,Springer-Verlag (in German)
58 Sanides, F. (1964) The cyto-myeloarchitecture of the human frontallobe, and its relation to phylogenetic differentiation of the cerebralcortex. J. Hirnforsch. 6, 269–282
59 Rizzolatti, G. and Arbib, M.A. (1998) Language within our grasp.Trends Neurosci. 21, 188–194
60 Corballis, M.C. (2009) The evolution and genetics of cerebralasymmetry. Philos. Trans. R. Soc. Lond. B: Biol Sci. 364, 867–879
61 Binkofski, F. et al. (2000) Broca’s region subserves imagery of motion: acombined cytoarchitectonic and fMRI study. Hum. Brain Mapp. 11,273–285
62 Iacoboni, M. (2009) Neurobiology of imitation. Curr. Opin. Neurobiol.19, 661–665
63 Petrides, M. et al. (2005) Orofacial somatomotor responses in themacaque monkey homologue of Broca’s area. Nature 435, 1235–1238
64 Friederici, A.D. et al. (2009) The role of the posterior superior temporalcortex in sentence comprehension. Neuroreport 20, 563–568
65 Philipp, A.M. et al. (2012) Differential roles of inferior frontal andinferior parietal cortex in task switching: evidence from stimulus-categorization switching and response-modality switching. Hum.Brain Mapp. http://dx.doi.org/10.1002/hbm.22036
66 Tan, L.H. et al. (2001) The neural system underlying Chineselogograph reading. Neuroimage 13, 836–846
67 Grodzinsky, Y. and Friederici, A.D. (2006) Neuroimaging of syntax andsyntactic processing. Curr. Opin. Neurobiol. 16, 240–246
68 Binder, J.R. and Desai, R.H. (2011) The neurobiology of semanticmemory. Trends Cogn. Sci. 15, 527–536
69 Rajkowska, G. and Goldman-Rakic, P.S. (1995) Cytoarchitectonicdefinition of prefrontal areas in the normal human cortex: I.remapping of areas 9 and 46 using quantitative criteria. Cereb.Cortex 5, 307–322
70 Brodmann, K. (ed.) (1909) Vergleichende Lokalisationslehre derGrosshirnrinde in ihren Prinzipien dargestellt auf Grund desZellenbaues (English Translation by Garey, L.J.: Brodmann’sLocalization in the Cerebral Cortex; Smith Gordon, London, 1994), Barth
71 Thompson-Schill, S.L. et al. (2005) The frontal lobes and the regulationof mental activity. Curr. Opin. Neurobiol. 15, 219–224
72 Aboitiz, F. et al. (2006) Cortical memory mechanisms and languageorigins. Brain Lang. 98, 40–56
426
73 Grodzinsky, Y. and Santi, A. (2008) The battle for Broca’s region.Trends Cogn. Sci. 12, 474–480
74 Meyer, M.E. and Jancke, L. (2006) Fontal oprculum in preception andproduction. In Broca’s Region (Grodzinsky, Y. and Amunts, K., eds),pp.218–241, Oxford University Press
75 Lancaster, J.L. et al. (2001) Automated Talairach atlas labels forfunctional brain mapping. Hum. Brain Mapp. 10, 120–131
76 Amunts, K. et al. (2004) Analysis of the neural mechanisms underlyingverbal fluency in cytoarchitectonically defined stereotaxic space - therole of Brodmann’s areas 44 and 45. Neuroimage 22, 42–56
77 Belmalih, A. et al. (2009) Multimodal architectonic subdivision of therostral part (area F5) of the macaque ventral premotor cortex. J. Comp.Neurol. 512, 183–217
78 Gerbella, M. et al. (2011) Cortical connections of the anterior (F5a)subdivision of the macaque ventral premotor area F5. Brain Struct.Funct. 216, 43–65
79 Zilles, K. and Amunts, K. (2009) Receptor mapping: architecture of thehuman cerebral cortex. Curr. Opin. Neurol. 22, 331–339
80 Palomero-Gallagher, N. et al. (2009) Receptor architecture of humancingulate cortex: evaluation of the four-region neurobiological model.Hum. Brain Mapp. 30, 2336–2355
81 Eickhoff, S.B. et al. (2008) Organizational principles of humanvisual cortex revealed by receptor mapping. Cereb. Cortex 18, 2637–2645
82 Amunts, K. et al. (2007) Cytoarchitecture of the cerebral cortex - morethan localization. Neuroimage 37, 1061–1065
83 Fellemann, D.J. and van Essen, D.C. (1991) Distributed hierarchicalprocessing in the primate cerebral cortex. Cereb. Cortex 1, 1–47
84 Passingham, R.E. et al. (2002) The anatomical basis of functionallocalization in the cortex. Nat. Rev. Neurosci. 3, 606–616
85 Chen, C-H. et al. (2012) Hierarchical genetic organization of humancortical surface area. Science 335, 1634–1636
86 Sanides, F. (1962) The architecture of the human frontal lobe and therelation to its functional differentiation. Int. J. Neurol. 5, 247–261
87 Koechlin, E. and Jubault, T. (2006) Broca’s area and the hierarchicalorganization of human behavior. Neuron 50, 963–974
88 Hickok, G. (2012) Computational neuroanatomy of speech production.Nat. Rev. Neurosci. 13, 135–145
89 Dehaene, S. and Changeux, J.P. (1997) A hierarchical neuronalnetwork for planning behavior. Proc. Natl. Acad. Sci. U.S.A. 94,13293–13298
90 Changeux, J.P. and Dehaene, S. (2000) Hierarchical neuronalmodeling of cognitive functions: from synaptic transmission to theTower of London. Int. J. Psychophysiol. 35, 179–187
91 Damasio, A.R. (1989) Time-locked multi-regional retroactivation: asystems-level proposal for the neural substrates of recall andrecognition. Cognition 33, 25–62
92 Luppino, G. et al. (1991) Multiple representations of body movementsin mesial area 6 and the adjacent cingulate cortex: an intracorticalmicrostimulation study in the macaque monkey. J. Comp. Neurol. 311,463–482
93 Vogt, O. (1910) Die myeloarchitektonische Felderung desmenschlichen Stirnhirns. J. Psychol. Neurol. XV, 221–232 (in German)
94 Brockhaus, H. (1940) Die Cyto- und Myeloarchitektonik des Cortexclaustralis und des Claustrum beim Menschen. J. Psychol. Neurol. 49,249–348 (in German)
95 Collins, D.L. et al. (1994) Automatic 3D intersubject registration of MRvolumetric data in standardized Talairach space. J. Comp. Ass.Tomogr. 18, 192–205
96 Vogt, O. (1951) Die anatomische Vertiefung der menschlichenHirnlokalisation. Verhandlungen der Gesellschaft deutscherNaturforscher und Arzte. Versammlung 96, 15–29 (in German)
97 Matelli, M. et al. (1991) Architecture of superior and mesial area 6 andthe adjacent cingulate cortex in the macaque monkey. J. Comp. Neurol.311, 445–462
98 Tanji, J. and Kurata, K. (1989) Changing concepts of motor areas of thecerebral cortex. Brain Dev. 11, 374–377
99 Nelissen, K. et al. (2011) Action observation circuits in the macaquemonkey cortex. J. Neurosci. 31, 3743–3756





























