Embed Size (px)
Citation preview

Archaeology of Eukaryotic DNA Replication
Kira S. Makarova and Eugene V. Koonin
National Center for Biotechnology Information, National Library of Medicine, National Institutesof Health, Bethesda, Maryland 20894
Correspondence: [email protected]
Recent advances in the characterization of the archaeal DNA replication system togetherwith comparative genomic analysis have led to the identification of several previously un-characterized archaeal proteins involved in replication and currently reveal a nearly com-plete correspondence between the components of the archaeal and eukaryotic replicationmachineries. It can be inferred that the archaeal ancestor of eukaryotes and even the lastcommon ancestor of all extant archaea possessed replication machineries that were compa-rable in complexity to the eukaryotic replication system. The eukaryotic replication systemencompasses multiple paralogs of ancestral components such that heteromeric complexes ineukaryotes replace archaeal homomeric complexes, apparently along with subfunctionali-zation of the eukaryotic complex subunits. In the archaea, parallel, lineage-specific dupli-cations of many genes encoding replication machinery components are detectable as well;most of these archaeal paralogs remain to be functionally characterized. The archaeal rep-lication system shows remarkable plasticity whereby even some essential components suchas DNA polymerase and single-stranded DNA-binding protein are displaced by unrelatedproteins with analogous activities in some lineages.
Double-stranded DNA is the molecule thatcarries genetic information in all cellular
life-forms; thus, replication of this genetic ma-terial is a fundamental physiological processthat requires high accuracy and efficiency(Kornberg and Baker 2005). The general mech-anism and principles of DNA replication arecommon in all three domains of life—archaea,bacteria, and eukaryotes—and include recog-nition of defined origins, melting DNA withthe aid of dedicated helicases, RNA primingby the dedicated primase, recruitment of DNApolymerases and processivity factors, replica-tion fork formation, and simultaneous replica-tion of leading and lagging strands, the latter via
Okazaki fragments (Kornberg and Baker 2005;Barry and Bell 2006; Hamdan and Richardson2009; Hamdan and van Oijen 2010). Thus, itwas a major surprise when it became clear thatthe protein machineries responsible for thiscomplex process are drastically different, espe-cially in bacteria compared with archaea andeukarya. The core components of the bacterialreplication systems, such as DNA polymerase,primase, and replication helicase, are unrelatedor only distantly related to their counterparts inthe archaeal/eukaryotic replication apparatus(Edgell 1997; Leipe et al. 1999).
The existence of two distinct molecularmachines for genome replication has raised
Editors: Stephen D. Bell, Marcel Mechali, and Melvin L. DePamphilis
Additional Perspectives on DNA Replication available at www.cshperspectives.org
Copyright # 2013 Cold Spring Harbor Laboratory Press; all rights reserved; doi: 10.1101/cshperspect.a012963
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
1
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

obvious questions on the nature of the replica-tion system in the last universal common ances-tor (LUCA) of all extant cellular life-forms, andthree groups of hypotheses have been proposed(Leipe et al. 1999; Forterre 2002; Koonin 2005,2006, 2009; Glansdorff 2008; McGeoch and Bell2008): (1) The replication systems in Bacte-ria and in the archaeo–eukaryotic lineageoriginated independently from an RNA-ge-nome LUCA or from a noncellular ancestralstate that encompassed a mix of genetic ele-ments with diverse replication strategies andmolecular machineries. (2) The LUCA was atypical cellular life-form that possessed eitherthe archaeal or the bacterial replication appara-tus in which several key components have beenreplaced in the other major cellular lineage. (3)The LUCAwas a complex cellular life-form thatpossessed both replication systems, so that thedifferentiation of the bacterial and the ar-chaeo–eukaryotic replication machineries oc-curred as a result of genome streamlining inboth lines of descent that was accompanied bydifferential loss of components. With regard tothe possible substitution of replication systems,a plausible mechanism could be replicon take-over (Forterre 2006; McGeoch and Bell 2008).Under the replicon takeover hypothesis, mobileelements introduce into cells a new replicationsystem or its components, which can displacethe original replication system through one orseveral instances of integration of the given ele-ment into the host genome accompanied by in-activation of the host replication genes and/ororigins of replication. This scenario is compat-ible with the experimental results showing thatDNA replication DNA in Escherichia coli withan inactivated DnaAgene or origin of replica-tion can be rescued by the replication appara-tus of R1 or F1 plasmids integrated into thebacterial chromosome (Bernander et al. 1991;Koppes 1992). Furthermore, genome analysissuggests frequent replicon fusion in archaeaand bacteria (McGeoch and Bell 2008); in par-ticular, such events are implied by the observa-tion that in archaeal genomes, genes encodingmultiple paralogs of the replication helicaseMCM and origins of replication are associatedwith mobile elements (Robinson and Bell 2007;
Krupovic et al. 2010). Replicon fusion also is aplausible path from a single origin of replicationthat is typical of bacteria to multiple originspresent in archaea and eukaryotes. However,all the evidence in support of frequent repliconfusion and the plausibility of replicon takeovernotwithstanding, there is no evidence of dis-placement of the bacterial replication apparatuswith the archaeal version introduced by mobileelements, or vice versa, displacement of the ar-chaeal machinery with the bacterial version, de-spite the rapid accumulation of diverse bacterialand archaeal genome sequences. Thus, the dis-placement scenarios of DNA replication ma-chinery evolution are so far not supported bycomparative genomic data.
Regardless of the nature of the DNA replica-tion system (if any) in the LUCA and the under-lying causes of the archaeo–bacterial dichotomyof replication machineries, the similarity be-tween the archaeal and eukaryotic replicationsystems is striking (Table 1). Generally, the ar-chaeal replication protein core appears to bethe same as the eukaryotic core, but eukaryotestypically possess multiple paralogous subunitswithin complexes that are homomeric in ar-chaea, many additional components interactingwith the core ones and more complex regulation(Leipe et al. 1999; Bell and Dutta 2002; Bohlkeet al. 2002; Kelman and White 2005; Barry andBell 2006). Thus, the archaeal replication systemappears to be an ancestral version of the eukary-otic system and hence a good model for func-tional and structural studies aimed at gainingmechanistic insights into eukaryotic replication.
In the last few years, there has been substan-tial progress in the study of the archaeal replica-tion systems that has led to an apparently com-plete delineation of all proteins that are essentialfor replication (Berquist et al. 2007; Beattie andBell 2011a; MacNeill 2011). The combination ofexperimental, structural, and bioinformaticsstudies has led to the discovery of archaeal ho-mologs (orthologs) for several components ofthe replication system that have been previouslydeemed specific for eukaryotes (Barry andBell 2006; MacNeill 2010, 2011; Makarovaet al. 2012). Furthermore, complex evolutionaryevents that involve multiple lineage-specific
K.S. Makarova and E.V. Koonin
2 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Table 1. The relationship between archaeal and eukaryotic replication systems
Archaea
(projection for LACA)
Eukaryotes
(projection for LECA) Comments
ORC complexarORC1 Orc1, Cdc6 In LACA the ORC/Cdc6 complex probably
consisted of two distinct subunits, and inLECA of six distinct.
Both complexes might possess additional Orc6and Cdt1 components.
arORC2 Orc2, Orc3, Orc4, Orc5TFIIB or homologa Orc6WhiP or other
wHTH proteinaCdt1
CMG complexArchaeal Cdc45/RecJ Cdc45 In many archaea and eukaryotes, CDC45/RecJ
apparently contain inactive DHHphosphoesterase domains.
The RecJ family is triplicated in euryarchaea,and some of the paralogs could be involved inrepair.
MCM is independently duplicated in severallineages of euryarchaea.
Mcm Mcm2, Mcm3, Mcm4,Mcm5, Mcm6, Mcm7
Gins23 Gins2, Gins3Gins15 Gins1, Gins5Inactivated MCM
homologaMcm10
CMG activation factors— RecQ/Sld2 There is no evidence that kinases and
phosphatases in archaea are directly involvedin replication, although they probablyregulate cell division.
— Treslin/Sld3— TopBP1/Dpb11STK CDK, DDKPP2C PP2C
PrimasesPrim1/p48 PriS In eukaryotes, Pol a is involved in priming by
adding short DNA fragments to RNAprimers.
In archaea, DnaG might be involved in primingspecifically on the lagging strand.
Prim2a/p58 PriLDnaG —
PolymerasesPolB3 Pol a, Pol d, Pol z No eukaryotic homologs of DP2 are known, but
Zn fingers of Pol 1 are apparently derivedfrom DP2.
PolB1 Pol 1DP1 B subunits of Pola, Pol d, Pol
z, Pol 1DP2 —
DNA polymerase sliding clamp and clamp loaderRFCL RFC1 Eukaryotes have additional duplications of both
RFCs and PCNA involved in checkpointcomplexes (Rad27 and Rad1, Rad9, Hus1,respectively).
RFCS RFC2, RFC3, RFC4, RFC4PCNA PCNA
Primer removal and gap closureRNase H2 RNase II There is a triplication of ligases (LigI, LigIII,
LigIV) in eukaryotes, but only LigI is directlyinvolved in replication.
In a few Halobacteria, ATP-dependent ligase isreplaced by NAD-dependent ligase.
Fen1 Fen1/EXO1, Rad2, Rad27Lig1 Lig1
SSBarRPA1_long Rpa1 In Thermoproteales, RPA is displaced by the
non-homologous ThermoSSB; two shortRPA forms in many euryarchaea; expansionof short RPA forms in Halobacteria.
arRPA1_short andRPA2
Rpa2
arCOG05741a Rpa3
For eukaryotic genes in Homo sapiens and Saccharomyces cerevisiae, gene names are indicated. Archaeal genes are denoted as
in Barry and Bell (2006) or as introduced here.aNot confidently traced to LACA.
Evolution of DNA Replication
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

duplications, domain rearrangements, and geneloss, and in part seem to parallel the evolutionof the evolution of the replication system ineukaryotes, have been delineated for a varietyof replication proteins in several archaeal lin-eages (Tahirov et al. 2009; Chia et al. 2010; Kru-povic et al. 2010). Here we summarize thesefindings and present several additional casestudies that show the complexity of evolution-ary scenarios for the components of the archaealreplication machinery and new aspects of theirrelationship with the eukaryotic replicationsystem.
PREREPLICATION COMPLEX
DNA replication begins at specific sites in thegenome known as the origins of replication.Some archaea possess a single and others pos-sess several replication origins, whereas in alleukaryotes replication starts from numerous,independent origins (Robinson and Bell 2005;Coker et al. 2009). Typically, replication originsencompass a distinct sequence signature, theAT-rich box, that can be used to predict originsin silico (Zhang and Zhang 2005).
The origin recognition complex (ORC) is ahexameric protein complex that binds the ori-gins of DNA replication and recruits additionalreplication factors resulting in the formation ofthe prereplicative complex (Fig. 1). EukaryoticOrc1-5 and the closely related CDC6 are paral-ogous proteins that belong to the DnaA/ORCclade of AAAþ ATPases (Iyer et al. 2004) andoften also contain a carboxy-terminal helix–turn–helix (HTH) DNA-binding domain. Allthe six families of eukaryotic ORC subunitshave been traced back to the last eukaryotic com-mon ancestor (LECA) (Makarova et al. 2005).The Orc2 and Orc3 families are highly divergedand contain inactivated ATPase domains.
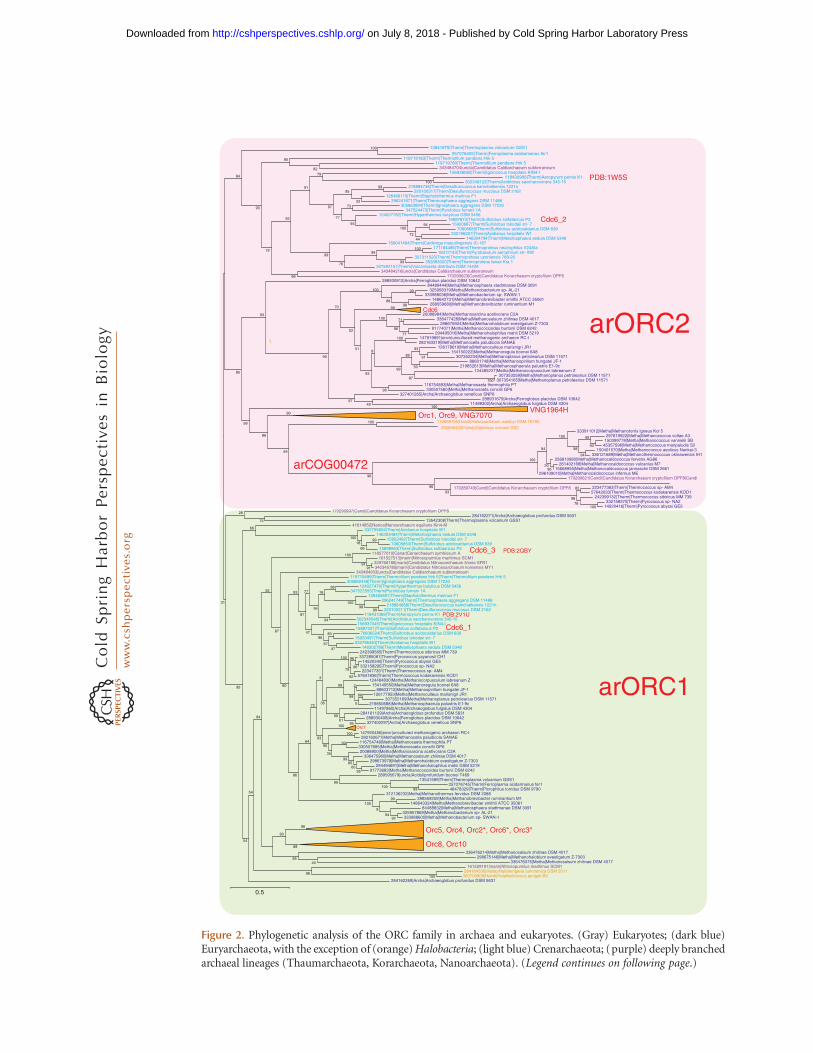
Phylogenetic analysis of archaeal ORC ho-mologs reveals a complex evolutionary scenarioof gene duplications, losses, and extreme diver-gence (Fig. 2). All this complexity notwith-standing, the phylogenetic tree clearly showsthe existence of two major groups of ORC pro-teins, namely, the slow-evolving arORC1 andthe fast-evolving arORC2. The vast majority of
archaea encode representatives of each of thesegroups.
Thermoproteales, Nanoarchaeum equitans,and several Thaumarchaeota possess only oneORC homolog that, in the latter two cases,belongs to the arORC1 group. Haloarchaeashow evidence of ancestral bursts of duplicationevents in both arORC1 and arORC2 clades(Fig. 2). The distinction between the two ances-tral ORC subfamilies persists, but most of thelineage-specific paralogs show accelerated evo-lutionary rates, and some have inactivatedATPase domains. Further duplications are de-tectable in several Halobacterial lineages givingrise to up to 20 ORC paralogs in Haloterrigenaturkmenica. It appears that paralogization ofORC genes is driven by the appearance ofadditional origins of replication including ac-quisition and integration of extra-chromosomalelements (McGeoch and Bell 2008). In Sulfolo-bus, it has been shown that the two origins arerecognized by distinct monomers or dimers ofOrc1/Cdc6 paralogs homologs and that thethree Orc1/Cdc6 genes are differentially ex-pressed in different growth phases and cell cyclestages, suggesting a role for these proteins inmodulating the activity of the origins (Robin-son et al. 2004). However, the functional differ-ences and the causes behind the distinct evolu-tionary constraints in the two major groups ofarchaeal ORCs remain poorly understood.
The Euryarchaeota of the order Methano-coccales are currently thought to lack ORC pro-teins, and Thermococcales are assumed to pos-sess only one ORC that belongs to the arORC1subfamily (Barry and Bell 2006). We identi-fied a subfamily of archaeal AAAþ ATPases(arCOG00472) that are fused to the carboxy-terminal wHTH domain and clusters with thearORC2 branch in the phylogenetic tree. Thetwo major lineages represented in this subfam-ily, Methanococcales and Thermococcales, areexactly those that were missing from the arORC2clade. Moreover, the genetic context of thesegenes in Methanococcales suggests their possi-ble role in replication (Fig. 3). The only archaeonin which we were unable to identify an ORCcandidate is Methanopyrus kandleri AV19. Thus,it is most likely that the last common ancestor
K.S. Makarova and E.V. Koonin
4 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

of the extant archaea and the last archaeo–eu-karyotic ancestor (whatever the exact natureof this entity) already encoded at least two paral-ogous ORC proteins. The nature of the func-tional distinctions between the slow-evolving
and fast-evolving ORC paralogs remains un-clear. In S. solfataricus, both the slow-evolvingarORC1 and two fast-evolving paralogs from thearORC2 branch all recognize origins of replica-tion albeit with different specificities (Robinson
Orc1-5arORC1/arORC2
Mcm2-7
DDKSTK
Sld2RecQ4 treslin/Sld3
TopBP1/Dpb11
GINS1234/Pfs1, Sld1-3GINS(51 and 23)
CDK
PP2CPP2C
Mcm10
Cdc45
Prim1BPrim2
RecJ/CDC45
PCNA
Pol ε
Cdt1
Orc6TFIIIB
Cdc6MCM
Whip
Pol δ
P
PP
PDP2DP1
DnaG
arRPA1
arRPA2
PolB3 PriLPriS
Rfc2-4Rfc1
Fen1
Fen1/EXO1, Rad2, Rad27
RNAseH2
Lig1Lig1
Rpa2Rpa3
Rpa1
PolB1
Pol α
Figure 1. Comparison of the archaeal (reconstructed LACA) and eukaryotic replication systems (reconstructedLECA). Orthologs are shown by shapes of the same color, and paralogs are denoted by similar colors. The geneproduct names are indicated as follows: for most eukaryotic genes, the Homo sapiens nomenclature is used, butin several cases, both H. sapiens and Saccharomyces cerevisiae gene names are given for clarity. Archaeal genes arenamed as in Barry and Bell (2006) or as discussed in the text. The dotted outline in some shapes indicatescomponents that could not be confidently traced to LACA. Small circles with “P” inside indicate phosphory-lation.
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 5
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
13541875|Therm|Thermoplasma volcanium GSS1257076405|Therm|Ferroplasma acidarmanus fer1
119719183|Therm|Thermofilum pendens Hrk 5119719780|Therm|Thermofilum pendens Hrk 5
343484704|uncla|Candidatus Caldiarchaeum subterraneum156938082|Therm|Ignicoccus hospitalis KIN4-I
118430906|Therm|Aeropyrum pernix K1302349122|Therm|Acidilobus saccharovorans 345-15
218884738|Therm|Desulfurococcus kamchatkensis 1221n320100317|Therm|Desulfurococcus mucosus DSM 2162
126466115|Therm|Staphylothermus marinus F1296241871|Therm|Thermosphaera aggregans DSM 11486
305663894|Therm|Ignisphaera aggregans DSM 17230347524473|Therm|Pyrolobus fumarii 1A
124027782|Therm|Hyperthermus butylicus DSM 545615897673|Therm|Sulfolobus solfataricus P2
15920687|Therm|Sulfolobus tokodaii str- 770606689|Therm|Sulfolobus acidocaldarius DSM 639
332796207|Therm|Acidianus hospitalis W1146304794|Therm|Metallosphaera sedula DSM 5348
159041494|Therm|Caldivirga maquilingensis IC-167171184486|Therm|Thermoproteus neutrophilus V24Sta
18312143|Therm|Pyrobaculum aerophilum str- IM2327311520|Therm|Thermoproteus uzoniensis 768-20
352683020|Therm|Thermoproteus tenax Kra 1307594151|Therm|Vulcanisaeta distributa DSM 14429
343484216|uncla|Candidatus Caldiarchaeum subterraneum170290623|Candi|Candidatus Korarchaeum cryptofilum OPF8
288930812|Archa|Ferroglobus placidus DSM 1064284489444|Metha|Methanosphaera stadtmanae DSM 3091325958319|Metha|Methanobacterium sp- AL-21
333988036|Metha|Methanobacterium sp- SWAN-1148642731|Metha|Methanobrevibacter smithii ATCC 35061
288559680|Metha|Methanobrevibacter ruminantium M1
Cdc620088984|Metha|Methanosarcina acetivorans C2A
336477428|Metha|Methanosalsum zhilinae DSM 4017298675924|Metha|Methanohalobium evestigatum Z-7303
91774071|Metha|Methanococcoides burtonii DSM 6242294495016|Metha|Methanohalophilus mahii DSM 5219
147919891|envir|uncultured methanogenic archaeon RC-I282163319|Metha|Methanocella paludicola SANAE
126178619|Metha|Methanoculleus marisnigri JR1154150222|Metha|Methanoregula boonei 6A8
307352234|Metha|Methanoplanus petrolearius DSM 1157188601748|Metha|Methanospirillum hungatei JF-1
219852613|Metha|Methanosphaerula palustris E1-9c124485227|Metha|Methanocorpusculum labreanum Z
307353259|Metha|Methanoplanus petrolearius DSM 11571307354165|Metha|Methanoplanus petrolearius DSM 11571
116754893|Metha|Methanosaeta thermophila PT330507560|Metha|Methanosaeta concilii GP6
327401265|Archa|Archaeoglobus veneficus SNP6288931679|Archa|Ferroglobus placidus DSM 10642
11498302|Archa|Archaeoglobus fulgidus DSM 4304
VNG1964HOrc1, Orc9, VNG7070
110669150|Halob|Haloquadratum walsbyi DSM 16790292656420|Halob|Haloferax volcanii DS2
333911012|Metha|Methanotorris igneus Kol 5297619922|Metha|Methanococcus voltae A3
150399778|Metha|Methanococcus vannielii SB45357596|Metha|Methanococcus maripaludis S2
150401570|Metha|Methanococcus aeolicus Nankai-3336121689|Metha|Methanothermococcus okinawensis IH1
256810980|Metha|Methanocaldococcus fervens AG86261402168|Metha|Methanocaldococcus vulcanius M7
15668955|Metha|Methanocaldococcus jannaschii DSM 2661296109010|Metha|Methanocaldococcus infernus ME
170290621|Candi|Candidatus Korarchaeum cryptofilum OPF8|Candi
170289743|Candi|Candidatus Korarchaeum cryptofilum OPF8 223477363|Therm|Thermococcus sp- AM457642033|Therm|Thermococcus kodakarensis KOD1
242399132|Therm|Thermococcus sibiricus MM 739332158370|Therm|Pyrococcus sp- NA2
14520416|Therm|Pyrococcus abyssi GE5170290997|Candi|Candidatus Korarchaeum cryptofilum OPF8
284162271|Archa|Archaeoglobus profundus DSM 563113542308|Therm|Thermoplasma volcanium GSS1
41614852|Nanoa|Nanoarchaeum equitans Kin4-M332795694|Therm|Acidianus hospitalis W1
146303484|Therm|Metallosphaera sedula DSM 534815922492|Therm|Sulfolobus tokodaii str- 7
70605854|Therm|Sulfolobus acidocaldarius DSM 63915898963|Therm|Sulfolobus solfataricus P2
118577010|Cenar|Cenarchaeum symbiosum A 161527513|marin|Nitrosopumilus maritimus SCM1
329766186|marin|Candidatus Nitrosoarchaeum limnia SFB1 340345785|marin|Candidatus Nitrosoarchaeum koreensis MY1
343484933|uncla|Candidatus Caldiarchaeum subterraneum 119719499|Therm|Thermofilum pendens Hrk 5|Therm|Thermofilum pendens Hrk 5
305662446|Therm|Ignisphaera aggregans DSM 17230 124027479|Therm|Hyperthermus butylicus DSM 5456
347522555|Therm|Pyrolobus fumarii 1A 126465697|Therm|Staphylothermus marinus F1
296241749|Therm|Thermosphaera aggregans DSM 11486 218884688|Therm|Desulfurococcus kamchatkensis 1221n
320100211|Therm|Desulfurococcus mucosus DSM 2162 118431069|Therm|Aeropyrum pernix K1
302348548|Therm|Acidilobus saccharovorans 345-15 156937045|Therm|Ignicoccus hospitalis KIN4-I
15897201|Therm|Sulfolobus solfataricus P2 70606524|Therm|Sulfolobus acidocaldarius DSM 639
15920497|Therm|Sulfolobus tokodaii str- 7 332796464|Therm|Acidianus hospitalis W1
146302786|Therm|Metallosphaera sedula DSM 5348 242399565|Therm|Thermococcus sibiricus MM 739 337285081|Therm|Pyrococcus yayanosii CH1
14520340|Therm|Pyrococcus abyssi GE5 332158295|Therm|Pyrococcus sp- NA2 223477351|Therm|Thermococcus sp- AM4
57641836|Therm|Thermococcus kodakarensis KOD1 124484830|Metha|Methanocorpusculum labreanum Z
154149550|Metha|Methanoregula boonei 6A8 88603712|Metha|Methanospirillum hungatei JF-1
126177953|Metha|Methanoculleus marisnigri JR1 307352169|Metha|Methanoplanus petrolearius DSM 11571
219850688|Metha|Methanosphaerula palustris E1-9c 11497860|Archa|Archaeoglobus fulgidus DSM 4304
284161129|Archa|Archaeoglobus profundus DSM 5631 288930408|Archa|Ferroglobus placidus DSM 10642 327400297|Archa|Archaeoglobus veneficus SNP6
Orc7
Cdc6_1
Cdc6_3 PDB:2QBY
PDB:2V1U
Cdc6_2
147920456|envir|uncultured methanogenic archaeon RC-I 282162671|Metha|Methanocella paludicola SANAE
116754748|Metha|Methanosaeta thermophila PT 330507685|Metha|Methanosaeta concilii GP6 20088900|Metha|Methanosarcina acetivorans C2A
336475960|Metha|Methanosalsum zhilinae DSM 4017 298673979|Metha|Methanohalobium evestigatum Z-7303
294494691|Metha|Methanohalophilus mahii DSM 5219 91773683|Metha|Methanococcoides burtonii DSM 6242
289595679|uncla|Aciduliprofundum boonei T469 13541589|Therm|Thermoplasma volcanium GSS1
257076745|Therm|Ferroplasma acidarmanus fer1 48478329|Therm|Picrophilus torridus DSM 9790
312136232|Metha|Methanothermus fervidus DSM 2088 288559259|Metha|Methanobrevibacter ruminantium M1
148643324|Metha|Methanobrevibacter smithii ATCC 35061 84488832|Metha|Methanosphaera stadtmanae DSM 3091
325957869|Metha|Methanobacterium sp- AL-21 333988600|Metha|Methanobacterium sp- SWAN-1
Orc5, Orc4, Orc2*, Orc6*, Orc3*
Orc8, Orc10 336476214|Metha|Methanosalsum zhilinae DSM 4017
298675146|Metha|Methanohalobium evestigatum Z-7303 336476075|Metha|Methanosalsum zhilinae DSM 4017
161529191|marin|Nitrosopumilus maritimus SCM1 284164538|Halob|Haloterrigena turkmenica DSM 5511
300709908|Halob|Halalkalicoccus jeotgali B3 284162269|Archa|Archaeoglobus profundus DSM 5631
96
88
90
4085
99
6618
100
66
75
26
100
82
95
100
79
99
33
95
73
94
4472
10093
77
97
91
100
99
99
76
99
93
66
79
93
84
99
9888
100
99
71
7796
100
100
91
93
50
89
10087
99
8
98
93
51
52
70
4997
99
93
100
90
100
93
99
94
98
100
9620
94
100
82
10078
98
9396
96
64
86
56
85
31
95
54
10098
54
9191
100
98
9998
100
16
94
94
77
83
9767
9647
87
93
96
96
92
79
100
2
86
0
98
99
9561
99
36
3
100
100
99
9560
8599
35
90
93
72
99100
99
64
99
9294
9
100
86
80
87
92
84
0.5
arORC2
arORC1
arCOG00472
PDB:1W5S
Figure 2. Phylogenetic analysis of the ORC family in archaea and eukaryotes. (Gray) Eukaryotes; (dark blue)Euryarchaeota, with the exception of (orange) Halobacteria; (light blue) Crenarchaeota; (purple) deeply branchedarchaeal lineages (Thaumarchaeota, Korarchaeota, Nanoarchaeota). (Legend continues on following page.)
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

19112617|Fungi|Schizosaccharomyces pombe 972h
19112164|Fungi|Schizosaccharomyces pombe 972h
19112702|Fungi|Schizosaccharomyces pombe 972h
Orc1
Cdc6
Orc5
Orc2
Orc3
Orc4
118380400|Cilio|Tetrahymena thermophila66358398|Apico|Cryptosporidium parvum Iowa II
84995186|Apico|Theileria annulata strain Ankara348681925|Oomyc|Phytophthora sojae
66824523|Mycet|Dictyostelium discoideum AX4308809383|Chlor|Ostreococcus tauri
49659270|Strep|Arabidopsis thaliana156389428|Metaz|Nematostella vectensis32454746|Metaz|Homo sapiens
348518229|Metaz|Oreochromis niloticus291236736|Metaz|Saccoglossus kowalevskii
198426976|Metaz|Ciona intestinalis71004418|Fungi|Ustilago maydis 521
6325420|Fungi|Saccharomyces cerevisiae S288c
340522426|Fungi|Trichoderma reesei QM6a302813646|Strep|Selaginella moellendorffii
17136788|Metaz|Drosophila melanogaster167527239|Choan|Monosiga brevicollis MX1
221130070|Metaz|Hydra magnipapillata32483369|Metaz|Homo sapiens
115948481|Metaz|Strongylocentrotus purpuratus145614652|Fungi|Magnaporthe oryzae 70-15
6323025|Fungi|Saccharomyces cerevisiae S288c50311281|Fungi|Kluyveromyces lactis NRRL Y-1140
307104133|Chlor|Chlorella variabilis145354524|Chlor|Ostreococcus lucimarinus CCE9901
167998931|Strep|Physcomitrella patens subsp- patens66153722|Strep|Oryza sativa Japonica Group15224316|Strep|Arabidopsis thaliana
5453830|Metaz|Homo sapiens198428189|Metaz|Ciona intestinalis
167525942|Choan|Monosiga brevicollis MX16319534|Fungi|Saccharomyces cerevisiae S288c
326469111|Fungi|Trichophyton tonsurans CBS 11281819112935|Fungi|Schizosaccharomyces pombe 972h-
118384008|Cilio|Tetrahymena thermophila17534571|Metaz|Caenorhabditis elegans
268532148|Metaz|Caenorhabditis briggsae66826059|Mycet|Dictyostelium discoideum AX4
281207872|Mycet|Polysphondylium pallidum PN500118398183|Cilio|Tetrahymena thermophila
66800637|Mycet|Dictyostelium discoideum AX4
6324068|Fungi|Saccharomyces cerevisiae S288c30688490|Strep|Arabidopsis thaliana
308801743|Chlor|Ostreococcus tauri71999676|Metaz|Caenorhabditis elegans
198426177|Metaz|Ciona intestinalis115935317|Metaz|Strongylocentrotus purpuratus
156357653|Metaz|Nematostella vectensis4505525|Metaz|Homo sapiens
348685832|Oomyc|Phytophthora sojae 17506005|Metaz|Caenorhabditis elegans
268566871|Metaz|Caenorhabditis briggsae 218189401|Strep|Oryza sativa Indica Group
224059306|Strep|Populus trichocarpa 30680045|Strep|Arabidopsis thaliana
198418008|Metaz|Ciona intestinalis 156407454|Metaz|Nematostella vectensis
47550985|Metaz|Strongylocentrotus purpuratus 4502703|Metaz|Homo sapiens 47214086|Metaz|Tetraodon nigroviridis
71018825|Fungi|Ustilago maydis 521
340521289|Fungi|Trichoderma reesei QM6a 6322266|Fungi|Saccharomyces cerevisiae S288c
159113089|Diplo|Giardia lamblia ATCC 50803 17555708|Metaz|Caenorhabditis elegans
309357462|Metaz|Caenorhabditis briggsae AF16 66356538|Apico|Cryptosporidium parvum Iowa II
118387267|Cilio|Tetrahymena thermophila 85000897|Apico|Theileria annulata strain Ankara
15235420|Strep|Arabidopsis thaliana 224106169|Strep|Populus trichocarpa
115466830|Strep|Oryza sativa Japonica Group 308803492|Chlor|Ostreococcus tauri
76803807|Metaz|Homo sapiens 115709869|Metaz|Strongylocentrotus purpuratus
4007803|Fungi|Schizosaccharomyces pombe 71024009|Fungi|Ustilago maydis 521
6323475|Fungi|Saccharomyces cerevisiae S288c 6323575|Fungi|Saccharomyces cerevisiae S288c
66811242|Mycet|Dictyostelium discoideum AX4
920
0
100
9799
92
100
100
98
52
81
99
69
85
99
3292
86
87
72
17
57
83
94
7762
25
90
87
10024
100
9799
12
68
88
49
60
10091
89
42
95
70
94
100
99
75
11
3999
91
2
82
38
95
80
86
100
72
3285
30
100
88
99
993
98
64
90
91
90
83
99
94
89
12
0.5
Figure 2. (Continued) The MUSCLE program (Edgar 2004) was used for construction of sequence alignments.The tree was reconstructed using the FastTree program (Price et al. 2010) (293 sequences and 186 alignedpositions for the archaeal tree and 84 sequences and 380 aligned positions for the eukaryotic tree). For Hal-obacteria, the branches are collapsed. For characterized genes of Halobacteria, Sulfolobus, and yeast, the conven-tional gene name or protein identifies are indicated in red on the right of the corresponding sequence or thecorresponding branch. For two archaeal sequences for which the structure is solved, its PDB code is indicated. (�)The corresponding genes are missing in some Halobacteria. (Red) Fast-evolving branches; (green) slow-evolvingbranches.
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 7
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

et al. 2004). Furthermore, the loss of ORC1 andthe colocalization of the ORC2 gene with repli-cation machinery components in Methanococ-cales imply that on some occasions in the evo-lution of archaea, ORC2 could take over the
essential function of ORC1 in replication. Inaddition, the absence of recognizable ORC ho-mologs in M. kandleri suggests that alternative,uncharacterized mechanisms of archaeal repli-cation initiation might exist.
Thermoproteales
Thermoproteales
Methanococcales
Desulfurococcales
Sulfolobales, Staphylothermus
Methanobacteria, Methanomicrobia
Methanomicrobia, Halobacteria
Halobacteria
Methanobacteria, Methanomicrobia, Methanococcales
ORC
DnaG
TFIIB (cyclin)
PolB2 (inactivated polymerase)
RPA
arORC1
arORC1
arORC2 (472)
HTH(4056)
HTH+Znf(5049)
HTH(5050)
GTPase(358)
MCM
PolB2
PolB2
DnaG
RecA
RFCL S7
TFIIB HIT
CC1DUF66Endo V
DUF66Endo V
RFCS
RFCS
arORC2
(2253)
(5526) (5525)
arRPA1_short
arRPA1_long
arRPA1_long HHT1
RecA
arRPA2MPP(1150) Lhr-like helicase
(7300)
Methanomicrobia, Halobacteria
Deacetylase(324)
Figure 3. Genomic context of selected genes for proteins involved in replication in archaea. Homologous genesare shown by arrows of the same color; genes are shown approximately to scale. (Blue) Replication genes;(magenta) genes coding for chromatin-binding protein and the respective modification enzymes; (orange)genes for translation system components; (gray) uncharacterized genes. Genes for which the neighborhoodis discussed in the text are marked by an outline. The arCOG numbers to which uncharacterized proteins areassigned are shown in parentheses. HTH, helix–turn–helix domain; S7, ribosomal protein S7; CC1, DNA-binding protein CC1; Endo V, homolog of endonuclease V; Znf, Zn finger; TFIIB, transcription initiation factorTFIIB; HIT, HIT family hydrolase; MPP, metallophosphatase superfamily protein; HHT1, histone.
K.S. Makarova and E.V. Koonin
8 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

The phylogenetic tree of the eukaryoticORC/CDC6 family also divides into two bran-ches: the slow-evolving ORC1/CDC6 and fast-evolving ORC2-3-4-5 (Fig. 2). Owing to theextreme sequence divergence, it is difficult toprecisely decipher the relationships between ar-chaeal and eukaryotic ORC families, but itseems likely that arORC1 is the ancestor ofOrc1/CDC6, whereas arORC2 is the ancestorof the ORC2-3-4-5 branch. However, in eukary-otes, the functional division between the slow-evolving and fast-evolving ORC subunits is dif-ferent from that in archaea because all of theseproteins are components of the heteromericcomplex involved in replication.
The sixth component of the ORC complex,Orc6, is shown to be required for DNA replica-tion in eukaryotes, and it is critical for ORCfunction (Duncker et al. 2009). Orc6 does notdisplay any sequence similarity with ATPases.Structural modeling of the amino-terminal do-main of metazoan Orc6 that is essential for rep-lication (Balasov et al. 2009) and the recentlyresolved structure of the middle portion ofhuman Orc6 both show that Orc6 contains twocyclin-like domains similar to those in the tran-scription factor TFIIB that belongs to a con-served achaeo–eukaryotic protein family (Liuet al. 2011). Unlike Orc6, the TFIIB family pro-teins, in addition to cyclin domains, also containan amino-terminal Zn finger. However, in sev-eral archaea, the Zn-binding ligands are lackingor the finger module is lost completely. Interest-ingly, in several genomes of Thermoproteales,the gene for TFIIB is encoded in a predictedoperon with the small subunit of clamp loader,replication factor C (RFC) (Fig. 3). Further-more, several archaeal lineages encode addition-al, functionally uncharacterized TFIIB paralogsthat typically contain a single cyclin-like do-main. Therefore, it cannot be ruled out thatTFIIB and/or its paralog perform dual functionsin both transcription and replication (Fig. 1).
In eukaryotes, the hexameric ORC recruitstwo additional components, namely, Cdc6,which is an apparent product of ancestral du-plication of Orc1 gene in eukaryotes (Fig. 2),and Cdt1 (Duncker et al. 2009). Both the mid-dle and the carboxy-terminal, MCM-interact-
ing, domains of Cdt1 adopt the winged HTHfold (Lee et al. 2004; De Marco et al. 2009). Inthe Crenarchaeota Aeropyrum pernix and sever-al Sulfolobus species, a gene adjacent to the or-igin of replication region (ORI) encodes awinged-helix initiator protein (WhiP) (Robin-son and Bell 2007). Given that WhiP containstwo winged HTH domains, it has been pro-posed that this protein is the ortholog and func-tional analog of Cdt1 (Robinson and Bell 2007).Typical WhiP proteins with two HTH domainsso far have been detected only in Desulfuro-coccales, although many other archaea encodeclosely related homologs of the amino-terminalHTH domain of WhiP, and some of theseproteins might interact with ORI regions.Moreover, in several archaeal genomes, HTH-containing proteins are encoded adjacent toORC subunits (Fig. 3), suggesting that theseproteins also could perform specific roles inreplication.
The resulting complete ORC–Cdc6–Cdt1complex is responsible for the ATP-dependentloading of the replicative helicase, the minichro-mosome maintenance (MCM) complex, in eu-karyotes and probably in some if not most ar-chaea and by inference in LACA (Fig. 1).
THE CMG COMPLEX
The eukaryotic MCM complex consists of sixparalogous proteins (Mcm2–7), all of whichare essential for cell viability and are requiredfor the initiation of DNA replication and repli-cation fork progression (Bochman and Schwa-cha 2009). Additional eukaryotic MCM proteinfamilies, Mcm8 and Mcm9, might be ances-tral but are not present in all species, and theirfunction in replication is unclear (Bochmanand Schwacha 2009). Genetic, biochemical, andstructural studies have shown that the hexa-meric MCM complex is the replicative helicasethat separates DNA strands during chromosom-al replication (Bochman and Schwacha 2009).The conserved core structure of all MCMproteins consists of an a-helical amino-termi-nal subdomain, an OB fold domain with aninserted Zn finger, an AAAþ ATPase domain,and a carboxy-terminal HTH domain. The
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 9
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

HTH domain can be confidently detected onlyin Mcm6. The structure of this domain has beensolved, and it has been shown to bind Cdt1 (Weiet al. 2010). The same core structure is con-served in MCM proteins from archaea includ-ing the carboxy-terminal HTH domain. It ap-pears that all eukaryotic MCM paralogs areproducts of ancestral duplications arising froma single ancestral MCM protein (Chia et al.2010; Krupovic et al. 2010). All archaea encodeat least one MCM protein homolog, and serialduplication of the MCM genes occurred in mul-tiple lineages. At least two paralogs could betraced back to common ancestor of Halobacter-iales and independently to the common ances-tor of Methanococcales (Chia et al. 2010; Kru-povic et al. 2010). Other duplications and lossesfollowed the ancestral duplication in Methano-coccales giving rise to up to eight MCM paral-ogs in Methanococcus maripaludis C6 (Chiaet al. 2010; Walters and Chong 2010). Duplica-tions occurred also within a few other narrowarchaeal lineages. It has been shown that allfour MCM paralogs of M. maripaludis S2 couldform a heterohexameric complex (Walters andChong 2010). In many archaea, some of theparalogous MCM genes are associated with mo-bile elements or viruses and show acceleratedevolution (Chia et al. 2010). To date, a singlegenetic study has been published showing thatonly one of the three MCM paralogs in the eu-ryarchaeon T. kodakarensis is essential for cellviability (Pan et al. 2011b). Interestingly, an in-tein inserted into one of the MCM genes, ap-parently early in the evolution of euryarchaea,followed by inactivation of the ATPase domainin several intein-containing MCM homologs;all archaea that possess this inactivated MCMprotein also encode an intact paralog.
In eukaryotes, in vitro and in vivo experi-ments have shown that the MCM complex, onits own, is not the active helicase but requires theassociation with two accessory factors, the tet-rameric GINS complex (Sld5, Psf1-3) and theCdc45 protein (Moyer et al. 2006; Pacek et al.2006; Labib and Gambus 2007). This complex isreferred to as the CMG (Cdc45, MCM, GINS)complex and is thought to be the active replica-tive helicase unit in vivo (Moyer et al. 2006;
Pacek et al. 2006; Labib and Gambus 2007). Inaddition to binding MCM proteins, the GINScomplex also associates with Pol a-primase, theprotein complex that synthesizes the primers onthe lagging strand, and with the leading andlagging strands polymerases, Pol 1 and Pol d,respectively (MacNeill 2010). In contrast tothe eukaryotic Mcm2–7 complex, which doesnot show helicase activity without the associat-ed GINS and Cd45 proteins, in vitro experi-ments with the archaeal MCM homohexamersfrom several organisms have revealed robusthelicase activity that did not require additionalsubunits (Sakakibara et al. 2009).
Highly diverged archaeal GINS homologshave been originally identified in silico (Maka-rova et al. 2005) and have been subsequentlyshown to interact with MCM (Marinsek et al.2006; Bell 2011). Archaea encode two distinctforms of the GINS proteins, one of which ap-pears to have been derived from the other bycircular permutation of a small domain (Marin-sek et al. 2006). These two forms are also pres-ent in eukaryotes, which have two ancestralgenes with one domain configuration (Psf2and Psf3) and two genes with the permuteddomain arrangement (Psf1 and Sld5). Althoughmany archaea encode only one GINS proteinthat belongs to the Psf1/Sld5 subfamily, Cren-archaeota, Thaumarchaeota, Korarchaeaon, andThermococci encode proteins of both subfam-ilies, suggesting that eukaryotes inherited bothforms from their archaeal ancestor (Marinseket al. 2006). Recently, the structure of twoGINS protein from T. kodakaraensis has beensolved, revealing that the backbone structureof each subunit and the tetrameric assemblyclosely resemble those of the human GINS com-plex (Oyama et al. 2011).
Until very recently, it was believed thatCdc45, which is present in all eukaryotes inone copy, is specific to eukaryotes (MacNeill2010). However, an in-depth computationalanalysis has shown that the amino-terminalregion of Cdc45 contains a DHH phosphoes-tarase domain, suggesting that Cdc45 is theeukaryotic ortholog of archaeal and bacterialRecJ nucleases (Sanchez-Pulido and Ponting2011). Furthermore, the GINS complex of
K.S. Makarova and E.V. Koonin
10 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

S. solfataricus, in addition to the associationwith MCM, binds a protein denoted RecJdbd(RecJ-like DNA-binding domain), which is ho-mologous to the carboxy-terminal domain ofbacterial RecJ but lacks the nuclease domain(Marinsek et al. 2006).
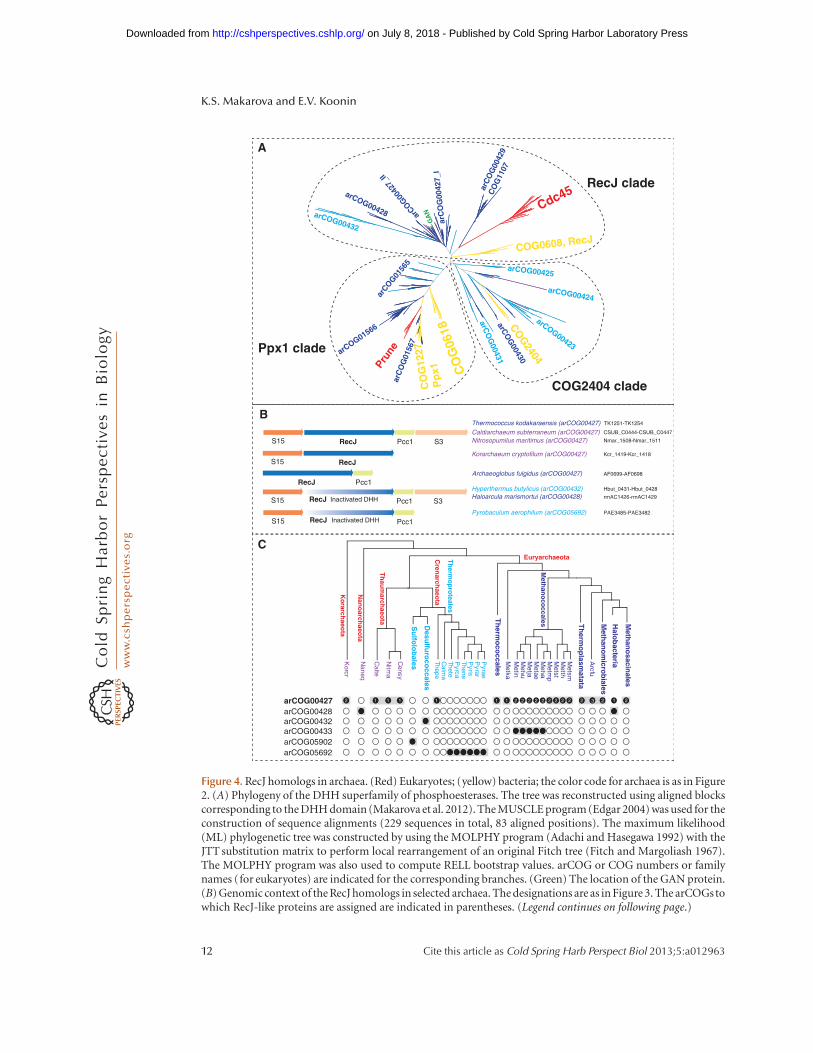
Recently, the GINS complex of the euryar-chaeon T. kodakarensis has been shown to inter-act with primase, MCM, DNA polymerase D,PCNA, and the GINS-associated nuclease(GAN) (Li et al. 2010, 2011). Unlike the RecJdbdof S. solfataricus, GAN is a bona fide ortholog ofbacterial RecJ containing a DHH phosphoester-ase domain with all the essential catalytic resi-dues. Recent phylogenomic analysis of RecJ ho-mologs in archaea led to the identification ofpreviously unsuspected homologs in Thermo-proteales and revealed a complex scenario ofRecJ family evolution in archaea and the originof Cdc45 (Fig. 4A) (Makarova et al. 2012).
Under this scenario, the last common ances-tor of all extant archaea possessed a single RecJortholog that was encoded in the conservedneighborhood including also the S15, S3, andPcc1 genes (Fig. 4B). This ancestral protein wasan active DHH nuclease and an essential com-ponent of the replication machinery. However,the experiments with the S. solfataricus replica-tion system indicate that the nuclease domain isnot required for replication; thus, the ancestralRecJ protein might have performed additionalfunctions, for example, in repair, that requiredthe nuclease activity. In Crenarchaeota, theDHH domain partially deteriorated, losing thenuclease activity, and the RecJ homolog ap-parently became a dedicated replication systemcomponent; the subsequent routes of evolutionwere notably different between the three majorcrenarchaeal branches—Sulfolobales, Desulfur-ococcales, and Themrorpoteales—resulting inextreme sequence divergence. The archaeal an-cestor of eukaryotes, the exact nature of whichremains elusive, also retained the RecJ ortholog(Cdc45) in which some but not all catalytic res-idues of the DHH domain are conserved; so far,to our knowledge, there are no experimentaldata showing a nuclease activity of Cdc45. InEuryarchaeota, the RecJ gene seems to have un-dergone triplication. One clade evolved very fast
and developed some specialized function (ar-COG00429, COG1107). Two otherclades (with-in arCOG00427) retained significant levels ofsequence similarity. Most of these proteins con-tain an active DHH nuclease domain, suggest-ing that they are active nucleases. Only one ofthese paralogs (arCOG00427_II) is often en-coded in a conserved neighborhood with theribosomal proteins S15 and S3, and the Pcc1subunit of the KEOPS complex (Fig. 4B).Thermococci might have lost one paralog(arCOG00427_I). Methanococcales encode anadditional paralog (arCOG00433) that has lostthe DHH domain. These genes are located ingenomic neighborhoods that encode no otherproteins involved in replication; thus, it is un-clear whether they are components of replica-tion systems. In Halobacteria, the RecJ orthologs(arCOG00428), which are encoded in the sameconserved neighborhood, contain an inactivat-ed DHH domain. Thus, inactivation of the RecJ-like nuclease that apparently became a dedicatedreplication protein seems to have occurred atleast twice independently in different archaeallineages (Fig. 4C).
In eukaryotes, MCM10 is an essential repli-cation protein that interacts with the assem-bled preRC and is required for the recruitmentof Cdc45 (Wohlschlegel et al. 2002). It staysin the active replisome complex throughoutthe S phase and appears to be important forboth replisome assembly and fork progression.Mcm10 physically interacts with both PCNAand Pol a (Chattopadhyay and Bielinsky 2007).The structure of the conserved middle domainof Mcm10 revealed a unique arrangement of aCCCH-type Zn-binding module and an OB-fold that jointly form a DNA-binding interface(Warren et al. 2008). The carboxy-terminal do-main of MCM10 is composed of a second,CCCC-type Zn-binding module that is not in-volved in DNA binding and a HTH domain. TheCCCC-type Zn-binding module of MCM10 isstructurally similar to a corresponding modulein the amino-terminal oligomerization domainof eukaryotic and archaeal MCM helicases, someof which also possess a carboxy-terminal HTHdomain (Robertson et al. 2010). In several fun-gi, Mcm10 additionally contains an amino-
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 11
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from
Prun
e
Cdc45
COG0608, RecJ
CO
G06
18G
AN
CO
G12
27
Pp
x1
COG
2404
arC
OG
0042
9C
OG
1107
arCOG00428 arCOG00
427_
II
arC
OG
0042
7_I
arCOG00432
arCOG00425
arCO
G00431
arCO
G00430
arCOG01566
arC
OG
0156
7
arCOG01
565
arCOG00424
arCOG00423
RecJ clade
COG2404 clade
Ppx1 clade
Pyrobaculum aerophilum (arCOG05692)
RecJ Inactivated DHHS15 Pcc1 S3
PAE3485-PAE3482
Hyperthermus butylicus (arCOG00432) Hbut_0431-Hbut_0428
RecJ Inactivated DHHS15 Pcc1
Thermococcus kodakaraensis (arCOG00427)Caldiarchaeum subterraneum (arCOG00427)
RecJS15 Pcc1 S3
TK1251-TK1254
Archaeoglobus fulgidus (arCOG00427) AF0699-AF0698
Haloarcula marismortui (arCOG00428) rrnAC1426-rrnAC1429
CSUB_C0444-CSUB_C0447
Korarchaeum cryptofilum (arCOG00427) Kcr_1419-Kcr_1418
RecJ Pcc1
RecJS15
Nitrosopumilus maritimus (arCOG00427) Nmar_1508-Nmar_1511
A
B
C
Halo
bacteria
Meth
ano
micro
biales
Th
ermo
plasm
atata
Meth
ano
coccales
Th
ermo
coccales
Th
ermo
pro
teales
Desu
lfuro
coccales
Su
lfolo
bales
Th
aum
archaeo
ta
Cren
archaeo
ta
Euryarchaeota
Ko
rarchaeo
ta
Nan
oarch
aeota
Meth
ano
sacinales
Arcfu
Metsm
Metth
Metst
Metm
pM
etvaM
etaeM
etjaM
etvuM
etinM
etka
Pyrae
Pyrar
Pyris
TheneP
yrcaTheteC
alma
Thepe
Censy
Nitm
a
Calte
Naneq
Korcr
arCOG00427arCOG00428arCOG00432arCOG00433arCOG05902arCOG05692
3 11 1 1 2 2 2 2 2 2 2 2 2 2 2 21112
Figure 4. RecJ homologs in archaea. (Red) Eukaryotes; (yellow) bacteria; the color code for archaea is as in Figure2. (A) Phylogeny of the DHH superfamily of phosphoesterases. The tree was reconstructed using aligned blockscorresponding to the DHH domain (Makarova et al. 2012). The MUSCLE program (Edgar 2004) was used for theconstruction of sequence alignments (229 sequences in total, 83 aligned positions). The maximum likelihood(ML) phylogenetic tree was constructed by using the MOLPHY program (Adachi and Hasegawa 1992) with theJTT substitution matrix to perform local rearrangement of an original Fitch tree (Fitch and Margoliash 1967).The MOLPHY program was also used to compute RELL bootstrap values. arCOG or COG numbers or familynames (for eukaryotes) are indicated for the corresponding branches. (Green) The location of the GAN protein.(B) Genomic context of the RecJ homologs in selected archaea. The designations are as in Figure 3. The arCOGstowhich RecJ-like proteins are assigned are indicated in parentheses. (Legend continues on following page.)
K.S. Makarova and E.V. Koonin
12 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

terminal P-loop ATPase domain (Fien et al.2004; Fien and Hurwitz 2006). The humanMCM10 protein forms a hexameric structuresimilar to the structure formed by other MCMproteins (Okorokov et al. 2007). Taken together,these observations suggest that Mcm10 might bederived from the MCM protein family. Archaeaalso encode several families of modified MCMhomologs. For example, inactivated MCM para-logs originated independently in M. kandleriand in Thermococci (arCOG05761). In bothcases, Zn-binding amino acids are also substi-tuted, but the carboxy-terminal HTH domain ispresent. The function of these MCM homologsin archaea is unknown. Several Methanosarci-nales and Halobacteriales possess a protein fam-ily with a CCCC-type Zn finger clearly related tothat in MCM and an amino-terminal HTH do-main (arCOG02259 and arCOG02260, respec-tively), resembling the configuration of the car-boxy-terminal portion of eukaryotic MCM10.Some of these proteins also contain an addition-al carboxy-terminal RepH/I/J family domainthat is involved in replication of plasmidpNRC100 in Haloferax volcanii (Ng and Das-Sarma 1993). Thus, a homolog of Mcm10 mighthave been present already in the archaeal ances-tor of eukaryotes (Fig. 1).
In eukaryotes, activation of the replicativehelicase and loading of the replisome dependon two serine–threonine kinases, CDK (cyclin-dependent kinase) and DDK (Dbf4-dependentkinase) (Tanaka et al. 2007; Araki 2010; Sheu
and Stillman 2010). The DDK phosphorylatesone of the Mcm2/4/6 proteins (Sheu andStillman 2010), whereas CDK phosphorylatesMcm5, Sld2, Sld3, and Mcm10 (Tanaka et al.2007). This phosphorylation results in bindingof Cdc45 and Dpb11 proteins and activation ofthe Mcm2–7 helicase (Sclafani and Holzen2007). The Sld2, Sld3, and Dpb11 proteins areconserved in most eukaryotes and are essentialregulators of replication initiation (Pospiechet al. 2010).
Dpb11 (known as TopBP1 in human) is amember of the BRCT domain superfamily thatcontains several BRCTrepeats (Bork et al. 1997;Garcia et al. 2005). In prokaryotes, the BRCTdomain is present only in NAD-dependentDNA ligase (Bork et al. 1997). The NAD-depen-dent ligase is an essential enzyme of DNA rep-lication and repair that is present in almost allbacteria (except for some intracellular symbi-onts with the smallest genomes) but only in afew archaea, which probably acquired this genevia horizontal transfer from bacteria. The eu-karyotic BRCT domains most likely evolvedfrom the bacterial ones.
The Sld3-like proteins show poor sequenceconservation even among eukaryotes. Recently,however, it has been shown that an a-helicalconserved domain of Sld3 is present in the ma-jority of eukaryotes (in animals and plants, thisprotein is called treslin); however, homologs inprokaryotes so far have not been detected (San-chez-Pulido et al. 2010).
Figure 4. (Continued) The protein IDs for this region in the corresponding genomes are indicated. (C) Phyleticpatterns of RecJ-related arCOGs. The phyletic patterns for indicated arCOGs ([filled circles] presence; [emptycircles] absence) are superimposed over the phylogenetic tree of archaea. The circles for proteins implicated inreplication are shaded. The number of paralogs is indicated inside the circles for arCOG00427 (all other sub-families have one representative in each genome). The tree is a modified version of the consensus phylogeny ofarchaea (Makarova et al. 2010) with Caldiarchaeum subterraneum included in the Thaumarchaeota branch andseveral branches with the same distribution of RecJ-related subfamilies collapsed. Arcfu, Archaeoglobus fulgidus;Metsm, Methanobrevibacter smithii ATCC 35061; Metth, Methanothermobacter thermautotrophicus; Metst,Methanosphaera stadtmanae; Metmp, Methanococcus maripaludis S2; Metva, Methanococcus vannielii SB; Metae,Methanococcus aeolicus Nankai-3; Metja, Methanocaldococcus jannaschii; Metvu, Methanocaldococcus vulcaniusM7; Metin, Methanocaldococcus infernus ME; Metka, Methanopyrus kandleri; Pyrae, Pyrobaculum aerophilum;Pyrar, Pyrobaculum arsenaticum DSM 13514; Pyris, Pyrobaculum islandicum DSM 4184; Thene, Thermoproteusneutrophilus V24Sta; Pyrca, Pyrobaculum calidifontis JCM 11548; Thete, Thermoproteus tenax; Calma, Caldivirgamaquilingensis IC-167; Thepe, Thermofilum pendens Hrk 5; Censy, Cenarchaeum symbiosum; Nitma, Nitro-sopumilus maritimus SCM1; Naneq, Nanoarchaeum equitans; Korcr, Candidatus Korarchaeum cryptofilumOPF8; Calte, Candidatus Caldiarchaeum subterraneum.
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 13
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

The Sld2 sequence is poorly conserved aswell, and this protein so far has been identifiedonly in Fungi. However, the animal RecQ4helicase contains an amino-terminal Sld2 do-main (Capp et al. 2010). We found that the ami-no-terminal domain of plant RecQ (e.g., GI:308804427) shows weak sequence similarityand compatible secondary structure with Sld2(KS Makarova, unpubl.). This observation sug-gests that the Sld2 domain is involved in theregulation of prereplication complex formationin all eukaryotes. Moreover, the LECA probablycontained the Sld2 domain fused with theRecQ-like helicase that might be directly in-volved in replication (Capp et al. 2010; Pospiechet al. 2010). Most bacteria encode a RecQ heli-case, whereas only a few archaea do. In Metha-nomicrobia, the recQ genes are clearly trans-ferred from bacteria. In contrast, Acidilobussaccharovorans, Aeropyrum pernix, and Korar-chaeum cryptofilum possess a small subfamilyof RecQ homologs that are highly divergedand contain a long amino-terminal region thatin K. cryptofilum encompasses a DEDDh-like30 –50 exonuclease domain and in the othertwo species probably is an inactivated derivativeof this nuclease. This domain architecture re-sembles the animal Werner syndrome RecQ-like helicase (Mushegian et al. 1997; Bernsteinet al. 2010). It seems likely that this particulararchaeal subfamily of RecQ helicases is ances-tral to the eukaryotic RecQ. An interesting ques-tion that probably can be addressed only bystructure comparison is whether the extendedamino-terminal region of these archaeal RecQ-like proteins also contains a domain homo-logous to Sld2 and if they are involved in repli-cation.
Homologs of CDK and DDK (COG0515)serine–threonine kinases are present in mostmajor archaeal lineages (with the exception ofThaumarchaeota and Nanoarchaeota) and thuscan be confidently projected to the archaealancestor of eukaryotes as well. Most likely, ser-ine/threonine protein phosphatase(s) might bepresent in this ancestral form as well. A function-al link between kinase and phosphatase and celldivision and membrane-remodeling systems inarchaea is strongly suggested by conserved geno-
mic contexts (Makarova and Koonin 2010;Makarova et al. 2010), but at this point, thereis no evidence of the involvement of these ki-nases in regulation of archaeal DNA replication.
THE ACTIVE REPLISOME
Once the CMG complex is assembled and acti-vated, it begins separating the two DNA strandsand forming a replication “bubble” from whichthe two replication forks, each containing aleading and a lagging strand, move away (Moyeret al. 2006; Pacek et al. 2006; Labib and Gambus2007; MacNeill 2010). Once replication is inprogress, other proteins required for the bulkDNA synthesis and protection of single-strand-ed DNA can be recruited to form the full repli-some complex at each replication fork (Fig. 1).
Primases
Similarly to the eukaryotic replication system,archaeal DNA replication involves a two-sub-unit primase (PriS, the catalytic subunit, andPriL, the polymerase-interacting subunit) thatsynthesizes an 8- to 12-nt RNA primer (Kuchtaand Stengel 2010). In eukaryotes, the primer isthen elongated by Pol a complexed with thecognate B (small) subunit to form a covalentDNA–RNA hybrid that is required for the rep-licative polymerase to start elongation (Kuchtaand Stengel 2010). In archaea, the RNA primerseems to be sufficient for the activation of thereplicative polymerase (Barry and Bell 2006).The evolution of these proteins in both eu-karyotes and archaea appears to have involvedprimarily simple vertical descent because themajority of archaeal and eukaryotic genomesencompass a single gene for each subunit. Theonly deviation from this simple pattern is foundin N. equitans, which encodes a hybrid proteincontaining regions approximately correspond-ing to the amino-terminal domain of the smallsubunit and the carboxy-terminal domain ofthe large subunit (NEQ395).
In addition to the large and small subunitsof the archaeo–eukaryotic primase (PriSL), allarchaea possess at least one gene for the bac-terial-type DnaG primase, although in contrast
K.S. Makarova and E.V. Koonin
14 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

to the bacterial homologs, archaeal DnaG lacksthe amino-terminal Zn-finger domain and thecarboxy-terminal domain that binds the bac-terial replicative helicase DnaB (Makarova etal. 1999). The recently characterized DnaG pro-tein of S. solfataricus has been shown to possesssimilar properties to the Escherichia coli primaseand shows a fourfold faster rate of DNA primingthan SsoPriSL (Zuo et al. 2010). It has beenproposed that DnaG might be involved in theinitiation of Okazaki fragment synthesis on thelagging strand (Zuo et al. 2010). The dnaG geneis often colocalized with the gene for the majorform of archaeal superfamily B DNA polymer-ase, although these genes might not be co-reg-ulated given that they are oriented convergently(Fig. 5). In many methanogens, dnaG is encod-ed in a predicted operon with uncharacterizeda-helical, potentially metal-binding protein ofarCOG02254 that by inference might have a rolein replication (Fig. 3). Most eukaryotes (but notanimals in which this gene has been inactivated)encode a DnaG protein that is typically fused toa DnaB-like helicase domain and is not directlyrelated to archaeal DnaG. Instead, this gene ap-pears to have been derived from the mitochon-
dria and has a dedicated function in mitochon-drial genome replication (Shutt and Gray 2006).
DNA Polymerase Sliding Clampand Clamp Loader
The PCNA (proliferating cell nuclear antigen)functions as the sliding clamp, a trimer that en-circles double-stranded DNA and freely slidesalong the DNA molecule (Beattie and Bell2011b). The sliding clamp is required for theprocessivity of the DNA polymerase and coor-dinates the function of multiple binding part-ners that are also required for replication andrepair processes. The sliding clamp is one ofthe few universally conserved proteins involvedin replication (the bacterial ortholog is knownas the DNA polymerase b subunit). In eukary-otes and many archaea, there is a single PCNAgene the product of which forms a homotrimer;this gene can be projected back to the lastcommon ancestor of archaea and the archaealancestor of eukaryotes. However, in eukar-yotes and independently in Crenarchaea, thePCNA gene is duplicated. Apparently there hadbeen an ancestral duplication in Crenarchaea
arORC1 DP 1
PriS GINS15
GINS23
GINS23
MCM
MCM
DP 2
PolB3 (major)RimI
GTPase
GTPase
DnaG
PCNA
arORC2 RFCL RFCS
CDC45/RecJS15 Pcc1 S3
(4059)
L44 S27
eIF-2β
eIF-2α Nop10
Figure 5. Conserved genomic context of selected genes encoding replication proteins. The designations are as inFigure 3. S3, S15, S27, L44, The respective ribosomal proteins; RimI, GNAT family acetyltransferase; eIF-2a andeIF-2b, the respective translation initiation factors; Nop10, an RNA-binding protein; Pcc1, subunit of theKEOPS complex.
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 15
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

followed by an additional duplication in theDesulfurococcales lineage (Chia et al. 2010).Some species of Crenarchaea encode multiplecopies of PCNA that can form either homo-trimers or heterotrimers (Barry and Bell 2006;Pan et al. 2011a). In eukaryotes, the ancestralduplication yielded at least three additionalPCNA-like families, all of which are subunitsof the checkpoint 9-1-1 complex (componentsHUS1, RAD1, and RAD9), which is involved inDNA damage checkpoint control (Aravind et al.1999; Majka and Burgers 2004).
Replication factor C, the clamp loader, isanother universally conserved protein that isrequired for PCNA assembly around a DNAmolecule at template–primer junctions (Bloom2009). In both archaea and eukaryotes, RFC is apentamer that consists of one large subunit(RFCL) encoded by a single gene and four smallsubunits (RFCS) that are identical in most ar-chaea but are represented by four distinct para-logs encoded by ancestral genes in eukaryotes(Makarova et al. 2005; Chia et al. 2010). Bothlarge and small subunits are paralogs and belongto the AAAþ superfamily of ATPases (Iyer et al.2004). In addition, early in eukaryotic evolution,further duplications gave rise to other families,such as Rad24, which were recruited for distinctroles in checkpoint complexes in parallel withthe duplication of the corresponding PCNAsand Chl12, a specific chromatid cohesion clamploader (Majka and Burgers 2004). In archaea,independent duplications of RFCS occurredin Methanomicrobia/Halobacteria, Thermopro-teales, and a few smaller lineages independently,often followed by acceleration of the evolution-ary rate of the “extra” copies (Chia et al. 2010).
Notably, in archaeal genomes, the “original”paralogs of both PCNA and RFC that retain theancestral roles in replication are encoded in con-served gene contexts adjacent to other proteinsinvolved in replication, although some addi-tional paralogs are encoded in suggestive con-texts as well (Figs. 3 and 5).
DNA Polymerases
In archaea and eukaryotes, the main replicativepolymerases belong to the B family of Palm-
domain polymerases (Burgers et al. 2001). Inaddition to the polymerase core domain, allthese proteins contain an amino-terminal 30 –50 exonuclease domain. All eukaryotes possessfour paralogous B-family polymerases denotedPol a, Pol d, Pol 1, and Pol z that are involved inboth DNA replication and repair (Makarovaet al. 2005; Kunkel and Burgers 2008). Archaeaencode at least two paralogous B-family poly-merases: the “major” one (present in all ar-chaea), and PolB3 and the minor one (severallineages of methanogens lack this gene), whichare both projected to LACA PolB1 (Edgell et al.1998; Rogozin et al. 2008; Tahirov et al. 2009).A small subset of archaea possesses another, di-vergent B-family polymerase (arCOG04926),which seems to contain active exonuclease andPalm domains. This family is represented bythree paralogs in Methanococcoides burtoniiand appears to be prone to duplication andHGT. In addition, several archaea encode a de-rivative B-family polymerase, PolB2, in whichboth the exonuclease and the Palm domain ap-pear to be inactivated (Edgell et al. 1998; Rogo-zin et al. 2008). In Crenarachaeota, this geneoften colocalizes with genes encoding a smalla-helical protein from arCOG07300 and RadAprotein, whereas in Euryarchaeota, it colocalizeswith the arORC2 gene, suggestive of involve-ment in replication (Fig. 3).
In addition to the B-family polymerases,many archaea encode the unique D-family po-lymerase (Cann et al. 1998), which is absent inCrenarachaea but present in all deeply branch-ing lineages (Thaumarchaea, Nanoarchaeon,Korarchaeon), suggesting that this polymerasewas present in LACA. The D-family polymer-ases consist of two subunits. The large subunitDP2 is a large multidomain protein that forms ahomodimer that is responsible for the polymer-ase activity (Shen et al. 2001; Matsui et al. 2011).The DP2 protein does not display any sequencesimilarity with other protein families (exceptfor two Zn fingers), but examination of con-served motifs suggests that it might be a highlydiverged Palm-domain polymerase (Cann et al.1998). The small subunit DP1 contains at leasttwo domains, an ssDNA-binding OB-foldand the 30 –50 exonuclease domain of the
K.S. Makarova and E.V. Koonin
16 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

metallophosphatase (MPP) family. The DP1protein is the ancestor of the small B subunitof eukaryotic replicative polymerases of the Bfamily that, however, have lost the catalytic ami-no acid residues of the 30 –50 exonuclease (Ara-vind and Koonin 1998; Klinge et al. 2009). Evi-dence has been presented that the D-familypolymerase specializes in the replication of thelagging strand, whereas the B-family polymeraseis involved in the leading-strand replication(Henneke et al. 2005). Interestingly, all Crenar-achaea that have no family-D polymerase pos-sess at least one additional active polymerase ofthe B family, suggesting that the two distinct B-family polymerases specialize on the leading-and lagging-strand replication, respectively, asis the case in eukaryotes.
Phylogenetic analysis of archaeal, eukaryot-ic, and bacterial B-family DNA polymerasescombined with an analysis of domain archi-tectures indicates that eukaryotic B-family po-lymerases, most likely, originated from twodistinct archaeal ancestors, the major archaealform that gave rise to the catalytically activeamino-terminal domain of Pol 1, and the minorform that became the common ancestor of Pola, Pol d, and Pol z. All eukaryotic B-family po-lymerases contain two carboxy-terminal Zn-finger modules. Interestingly, the carboxy-ter-minal module appears to be derived from theZn finger in the DP2 subunit of archaeal D-family DNA polymerases that are unrelated tothe B family, at least as judged by sequence com-parison (Tahirov et al. 2009). The Zn finger ofPol 1 shows greater similarity to the counterpartin archaeal DP2 than the Zn fingers of othereukaryotic B-family polymerases. The car-boxy-terminal portion of eukaryotic Pol 1 con-sists of two additional polymerase and exonu-clease domains, both inactivated; there areindications that this module could be of bacte-rial or bacteriophage origin (Tahirov et al.2009). The presence of an inactivated exonucle-ase–polymerase module in Pol 1 parallels a sim-ilar inactivation of both enzymatic domains in adistinct subfamily of inactivated archaeal B-family polymerases (Rogozin et al. 2008).
The apparent derivation of the large sub-units of eukaryotic B-family polymerases from
the major (PolB3) and minor (PolB1) formsof archaeal polymerases, and the origin of thesmall subunit from DP1 and the origin of thecarboxy-terminal Zn finger from DP2 empha-size the joint contributions of the B-family andD-family archaeal polymerases to the evolutionof the eukaryotic replication machinery. Fur-thermore, these findings suggest that the ar-chaeal ancestor of eukaryotes possessed a highlycomplex replication apparatus, possibly morecomplex than any known extant archaeon.
Primer Removal and Gap Closure
In both eukaryotes and archaea, RNase HII andFEN1 flap endonuclease are responsible for re-moval of RNA primers during replication (Fig.1) (Barry and Bell 2006). Both enzymes arepresent in all archaea with only a few duplica-tions observed. Eukaryotes have a single genefor RNase II, but ancestral duplications haveled to at least three FEN1-like families (Rad2,Rad27, EXO1), which, as shown in yeast, pos-sess essentially the same activities and can com-plement each other in both replication and re-pair processes (Sun et al. 2003).
Another essential replication enzyme isDNA ligase, which is responsible for joining ofthe Okazaki fragments during lagging-strandreplication. The main ligase involved in DNAreplication in archaea is an ATP-dependent li-gase (arCOG01347), which is encoded in thevast majority of archaeal genomes and is anapparent ortholog of the three eukaryotic li-gases (I, III, and IV) that evolved throughancestral duplications (Ellenberger and Tom-kinson 2008; Yutin and Koonin 2009). Theseenzymes share a common DNA-binding do-main, a catalytic nucleotidyltransferase domain,and an OB-fold domain (Martin and MacNeill2002; Ellenberger and Tomkinson 2008). OnlyLigase I in eukaryotes is directly involved inreplication, whereas Ligase III and Ligase IVhave adopted different roles in DNA repairand homologous recombination. Ligase IIIand Ligase IV contain carboxy-terminal BRCTdomains that are involved in protein–pro-tein interactions via phosphoserine recognition(Martin and MacNeill 2002). As pointed out
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 17
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

above, a BRCT domain is also present in bacte-rial NAD-dependent DNA ligase, which is alsofound in several archaea. So far only two ar-chaea, Halalkalicoccus jeotgali and Halorhabdusutahensis, lack the ATP-dependent ligase butencode the NAD-dependent ligase, which inthese organisms can be predicted to functionin replication.
Single-Stranded DNA-Binding Proteins
Single-stranded DNA-binding proteins (SSBs)are essential components of the replication ma-chinery that prevent reannealing of the growingDNA chain with the template and protectssDNA from degradation. In all three domainsof life, the major SSB proteins possess an OB-fold (Murzin 1993). In eukaryotes, the SSB,known as RPA, is a heterotrimer composed ofthree subunits of 70, 32, and 14 kDa (denotedRPA1, RPA2, and RPA3 in yeast) that containfour, one, and one OB-fold units, respectively.All three subunits are essential for the formationof a stable, functional RPA complex (Bochkarevand Bochkareva 2004). Archaea show multiplevariations of RPA domain organization, num-ber of homologs, and modes of interaction.Among the experimentally characterized RPAcomplexes in archaea, there is a homotetramercontaining a single OB-fold in S. solfataricus(Wadsworth and White 2001); single-subunitRPAs containing multiple OB-folds in metha-nogens Methanosarcina acetivorans, Methano-caldococcus jannaschii, and Methanothermobac-ter thermautotrophicus (Robbins et al. 2005, andreferences therein); and a heterotrimeric RPA inPyrococcus furiosus that consists of three distant-ly related subunits: RPA41 (COG1599), RPA32(COG3390), and RPA14 (arCOG05741), eachcontaining an OB-fold; in addition, RPA41 con-tains a Zn-finger-like motif (Komori and Ishino2001).
To characterize general trends in the evolu-tion of RPA in archaea, we reconstructed phy-logenetic trees for COG1599 and COG3390(Fig. 6A). The COG1599 subtree (which wehere denote arRPA1) divides into two clades:arRPA1_long with several OB-fold domainsand often a Zn finger homologous to that in
eukaryotic RPA1; and arRPA1_short, typicallywith a single OB-fold domain.
The diverged COG3390 (arRPA2) consistsof short proteins most of which contain a singleOB-fold domain and a carboxy-terminal wHTHdomain, the same domain arrangement as in theeukaryotic RPA2 protein (Mer et al. 2000). Sofar, RPA2 has been detected only in Euryar-chaeota (Fig. 6B). It seems likely that arRPA2evolved by duplication of the short form ofarRPA1 followed by accelerated evolution. Sim-ilar events have been detected in eukaryotes. Forexample, fungal CDC13, Stn1, and Ten1, thesubunits of a heterotrimeric complex essentialfor telomere maintenance, are paralogs of theRpa1–Rpa2–Rpa3 complex subunits but showonly remote sequence similarity to RPA1, RPA2,and RPA3, respectively (Sun et al. 2009, 2011).
Thus, most archaea possess at least two RPAgenes, one long (arRPA1_long) and one orseveral short ones (arRPA1_short or arRPA2),which are apparent ancestors of eukaryoticRPA1 and RPA2, respectively, and can be con-fidently projected to LACA (Fig. 6B). Whetheror not LACA encoded a third short RPA (ar-COG05741) remains uncertain. but, given thepresence of this gene in Korarchaeaon andseveral recently sequenced genomes of deeplybranching archaea (Fig. 6B) (Narasingarao etal. 2012), it seems likely that this RPA is ancestralin archaea as well (Fig. 1). The major exceptionsfrom this pattern of multiple RPA forms areSulfolobales/Desulfurococcales, which possessonly RPA1_short, and Thermoproteales, whichare the only known organisms without detect-able OB-fold-containing SSBs.
The apparent absence of a canonical RPA/SSB in Thermoproteales stimulated the searchfor an alternative, which resulted in the iden-tification of a distinct ssDNA-binding proteinin T. tenax that is unrelated to RPA and hasbeen denoted ThermoSSB (Paytubi et al. 2012).ThermoSSB (arCOG05578)contains anssDNA-binding domain with a novel fold and a leucine-zipperdomainthatmediatesdimerizationofthisprotein (Paytubi et al. 2012). ThermoSSB is pre-sent in all Thermoproteales with the exceptionof T. pendens, which encodes two RPA-like pro-teins. Thus, ThermoSSB perfectly complements
K.S. Makarova and E.V. Koonin
18 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Halo_RPA2_1
Halo_RPA2_2
13542016|Therm|Thermoplasma volcanium GSS1
48478491|Therm|Picrophilus torridus DSM 9790257076905|Therm|Ferroplasma acidarmanus fer1
55376365|Halob|Haloarcula marismortui ATCC 43049288931348|Archa|Ferroglobus placidus DSM 10642
118577123|Cenar|Cenarchaeum symbiosum A161527601|marin|Nitrosopumilus maritimus SCM1
340343954|marin|Candidatus Nitrosoarchaeum koreensis MY1329766730|marin|Candidatus Nitrosoarchaeum limnia SFB1
20094877|Metha|Methanopyrus kandleri AV19327401950|Archa|Archaeoglobus veneficus SNP6
312136409|Metha|Methanothermus fervidus DSM 208815679384|Metha|Methanothermobacter thermautotrophicus str- Delta H
288559372|Metha|Methanobrevibacter ruminantium M1148643392|Metha|Methanobrevibacter smithii ATCC 35061325957925|Metha|Methanobacterium sp- AL-21
333988515|Metha|Methanobacterium sp- SWAN-141614994|Nanoa|Nanoarchaeum equitans Kin4-M
170290896|Candi|Candidatus Korarchaeum cryptofilum OPF857641896|Therm|Thermococcus kodakarensis KOD1
223478091|Therm|Thermococcus sp- AM4242398695|Therm|Thermococcus sibiricus MM 739337284758|Therm|Pyrococcus yayanosii CH1
14520503|Therm|Pyrococcus abyssi GE5332158150|Therm|Pyrococcus sp- NA2
124485459|Metha|Methanocorpusculum labreanum Z 88604174|Metha|Methanospirillum hungatei JF-1
11497994|Archa|Archaeoglobus fulgidus DSM 4304 297619538|Metha|Methanococcus voltae A3
45358595|Metha|Methanococcus maripaludis S2 150399109|Metha|Methanococcus vannielii SB 333910219|Metha|Methanotorris igneus Kol 5
296109213|Metha|Methanocaldococcus infernus ME 15669348|Metha|Methanocaldococcus jannaschii DSM 2661
256810698|Metha|Methanocaldococcus fervens AG86 261403319|Metha|Methanocaldococcus vulcanius M7
150400778|Metha|Methanococcus aeolicus Nankai-3 336121122|Metha|Methanothermococcus okinawensis IH1
116754196|Metha|Methanosaeta thermophila PT 330508607|Metha|Methanosaeta concilii GP6
Halo_RPA1_1 294494712|Metha|Methanohalophilus mahii DSM 5219
298674005|Metha|Methanohalobium evestigatum Z-7303 91773712|Metha|Methanococcoides burtonii DSM 6242 20093424|Metha|Methanosarcina acetivorans C2A
336475982|Metha|Methanosalsum zhilinae DSM 4017 282165660|Metha|Methanocella paludicola SANAE
289596922|uncla|Aciduliprofundum boonei T469 333910932|Metha|Methanotorris igneus Kol 5
297620031|Metha|Methanococcus voltae A3 45357685|Metha|Methanococcus maripaludis S2
150399872|Metha|Methanococcus vannielii SB 305663511|Therm|Ignisphaera aggregans DSM 17230
70606760|Therm|Sulfolobus acidocaldarius DSM 639 15920719|Therm|Sulfolobus tokodaii str- 7 15899120|Therm|Sulfolobus solfataricus P2
332797833|Therm|Acidianus hospitalis W1 146303452|Therm|Metallosphaera sedula DSM 5348
170289718|Candi|Candidatus Korarchaeum cryptofilum OPF8 302348760|Therm|Acidilobus saccharovorans 345-15
124027704|Therm|Hyperthermus butylicus DSM 5456 126466308|Therm|Staphylothermus marinus F1
296242163|Therm|Thermosphaera aggregans DSM 11486 320100590|Therm|Desulfurococcus mucosus DSM 2162
218883974|Therm|Desulfurococcus kamchatkensis 1221n 156937744|Therm|Ignicoccus hospitalis KIN4-I
118431409|Therm|Aeropyrum pernix K1 118576582|Cenar|Cenarchaeum symbiosum A
329764702|marin|Candidatus Nitrosoarchaeum limnia SFB1 340344367|marin|Candidatus Nitrosoarchaeum koreensis MY1 161528013|marin|Nitrosopumilus maritimus SCM1
289596472|uncla|Aciduliprofundum boonei T469 13542067|Therm|Thermoplasma volcanium GSS1
48477745|Therm|Picrophilus torridus DSM 9790 257076028|Therm|Ferroplasma acidarmanus fer1
343485818|uncla|Candidatus Caldiarchaeum subterraneum 119719124|Therm|Thermofilum pendens Hrk 5 146303241|Therm|Metallosphaera sedula DSM 5348
302348560|Therm|Acidilobus saccharovorans 345-15 289597160|uncla|Aciduliprofundum boonei T469
Halo_RPA1_2 282163689|Metha|Methanocella paludicola SANAE
147921611|envir|uncultured methanogenic archaeon RC-I 20089479|Metha|Methanosarcina acetivorans C2A
298675833|Metha|Methanohalobium evestigatum Z-7303 336476389|Metha|Methanosalsum zhilinae DSM 4017
91773348|Metha|Methanococcoides burtonii DSM 6242 219850711|Metha|Methanosphaerula palustris E1-9c
307352311|Metha|Methanoplanus petrolearius DSM 11571 126180294|Metha|Methanoculleus marisnigri JR1
124486473|Metha|Methanocorpusculum labreanum Z 154149727|Metha|Methanoregula boonei 6A8
Halo_RPA1_3
Halo_RPA_4116754739|Metha|Methanosaeta thermophila PT
330507585|Metha|Methanosaeta concilii GP6147921018|envir|uncultured methanogenic archaeon RC-I
282165519|Metha|Methanocella paludicola SANAE294496321|Metha|Methanohalophilus mahii DSM 5219298675819|Metha|Methanohalobium evestigatum Z-7303
91773338|Metha|Methanococcoides burtonii DSM 6242336477023|Metha|Methanosalsum zhilinae DSM 4017
20091837|Metha|Methanosarcina acetivorans C2A284162746|Archa|Archaeoglobus profundus DSM 5631
327400196|Archa|Archaeoglobus veneficus SNP6288931687|Archa|Ferroglobus placidus DSM 1064211498386|Archa|Archaeoglobus fulgidus DSM 4304
71
91
8794
90
84
6067
5399
74
83
79
94
61
87
90
98
14
98
92
29
91
89
9436
81
93
83
7780
8896
97
92
93
100
9682
98
87
63
993
80
54
44
9477
95
62
92
51
26
97
6993
99
8282
81
7593
26
51
76
59
28
96
99
8389
95
80
94
77
8435
21
90
98
99
75
89
97
47
7786
92
20
93
7593
82
84
89
98
78
83
98
99
67
72
52
11
79
75
85
84
41
78
0.5
13542016|Therm|Thermoplasma volcanium GSS1
48478491|Therm|Picrophilus torridus DS257076905|Therm|Ferroplasma acidarmanus fer93
100
57641896|Therm|Thermococcus kodakarensis K223478091|Therm|Thermococcus sp- AM4
242398695|Therm|Thermococcus sibiricus MM 739337284758|Therm|Pyrococcus yayanosii CH1
14520503|Therm|Pyrococcus abyssi GE5332158150|Therm|Pyrococcus sp- NA2
91
8794
90
84
13542067|Therm|Thermoplasma volcanium 48477745|Therm|Picrophilus torridus DSM 9790
257076028|Therm|Ferroplasma acidarmanus 822
Halobacteria
Methanomicrobiales
Thermoplasmatata
Thermococcales
Methanococci
Methanobacteria
Thermoproteales
Desulfurococcales
Sulfolobales
Thaumarchaeota
Thaumarchaeota
Crenarchaeota
Eu
ryar
chae
ota
Korarchaeota
Nanoarchaeota
Methanosarcinales
Archaeoglobi
Metka
Thepe
Calte
Naneq
Korcr
arR
PA
1_lo
ngar
RP
A1_
shor
tar
RP
A2
arC
OG
0557
8ar
CO
G05
741
arC
OG
0377
2
arC
OG
0226
1
3 22
2
A B
arRPA1_long
arRPA1_short arRPA2
13542015|Therm|Thermoplasma volcanium GSS1 48478490|Therm|Picrophilus torridus DSM 9790
257076906|Therm|Ferroplasma acidarmanus fer1 20094402|Metha|Methanopyrus kandleri AV19
223478132|Therm|Thermococcus sp- AM4 57641894|Therm|Thermococcus kodakarensis KOD1
337284760|Therm|Pyrococcus yayanosii CH1 332158152|Therm|Pyrococcus sp- NA2 14520501|Therm|Pyrococcus abyssi GE5
242398697|Therm|Thermococcus sibiricus MM 739 45357683|Metha|Methanococcus maripaludis S2
150399870|Metha|Methanococcus vannielii SB 333910930|Metha|Methanotorris igneus Kol 5
297620033|Metha|Methanococcus voltae A3 126179197|Metha|Methanoculleus marisnigri JR1
154151219|Metha|Methanoregula boonei 6A8 219852691|Metha|Methanosphaerula palustris E1-9c
307353131|Metha|M.petrolearius DSM 11571
124485338|Metha|Methanocorpusculum labreanum Z 11498385|Archa|Archaeoglobus fulgidus DSM 4304
288930854|Archa|Ferroglobus placidus DSM 10642 284162747|Archa|Archaeoglobus profundus DSM 5631
288931688|Archa|Ferroglobus placidus DSM 10642 327400197|Archa|Archaeoglobus veneficus SNP6
147921017|envir|uncultured methanogenic archaeon RC-I 282165520|Metha|Methanocella paludicola SANAE
116754740|Metha|Methanosaeta thermophila PT 330507584|Metha|Methanosaeta concilii GP6
336477022|Metha|Methanosalsum zhilinae DSM 4017 294496320|Metha|Methanohalophilus mahii DSM 5219
298675818|Metha|Methanohalobium evestigatum Z-7303 91773337|Metha|Methanococcoides burtonii DSM 6242
20091836|Metha|Methanosarcina acetivorans C2A 307352310|Metha|Methanoplanus petrolearius DSM 11571
219850710|Metha|Methanosphaerula palustris E1-9c 126180295|Metha|Methanoculleus marisnigri JR1 154149726|Metha|Methanoregula boonei 6A8
124486472|Metha|Methanocorpusculum labreanum Z 88603781|Metha|Methanospirillum hungatei JF-1
88601895|Metha|M.hungatei JF-1
147921610|envir|uncultured methanogenic archaeon RC-I 282163688|Metha|Methanocella paludicola SANAE
91773347|Metha|Methanococcoides burtonii DSM 6242 20089480|Metha|Methanosarcina acetivorans C2A
336476390|Metha|Methanosalsum zhilinae DSM 4017 298675832|Metha|Methanohalobium evestigatum Z-7303
289597159|uncla|Aciduliprofundum boonei T469
7530
42
85
100
98
98
49
91
51
95
83
89
99
596
98
7560
8085
97
77
98
60
90
87
84
99
81
8794
87
83
96
79
63
95
1475
90
69
99
95
72
92
61
98
0.2
219850726|Metha|Methanosphaerula palustris E1-9c 154151637|Metha|Methanoregula boonei 6A8 126179860|Metha|Methanoculleus marisnigri JR1
Figure 6. Phylogenetic analysis of the RPA family in archaea and eukarya. The designations and the method oftree reconstruction are as in Figure 2. (Green) The Thermococcales branches; (pink) the Thermoplasmatabranches. Halobacterial branches are collapsed and numbered as follows: for arRPA1 from Halo_RPA1_1 toHalo_RPA1_4 and for arRPA2 from Halo_RPA2_1 to Halo_RPA2_2. “Long” RPAs are outlined by the reddotted line and “short” RPAs by the green dotted line. (A) Phylogenetic trees of the RPA1 and RPA2 families.RPA1 corresponds to COG1599 (167 sequences in total, 89 aligned positions), and RPA2 corresponds toCOG03390 (76 sequences in total, 149 aligned positions). (B) The phyletic patterns of SSB/RPA proteinsand their homologs in archaea. The designations are as in Figure 4C. (In panel B, circles with a yellow outlinedenote) RPA proteins from several organisms that could not be confidently aligned and thus are not present inthe corresponding tree but included into the phyletic pattern. (Green outline) Those that do not include allrepresentatives in the corresponding lineage.
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

the phyletic pattern of RPA such that SSB pro-teins now have been identified in all archaea.
Some potential archaeal SSBs remain un-assigned. In particular, an uncharacterized pa-ralog of ThermoSSB (arCOG03772) shows abroader phyletic distribution being present, inaddition to Thermoproteales, in Sulfolobales/Desulfurococcales, Thermococcales, and Ar-chaeoglobi (Paytubi et al. 2012). In addition,there is at least one more protein family con-taining OB-fold domains and distantly similarto archaeal RPA2 (arCOG02261) that is presentin most Methanococcales and Nanoarchaeumequitans.
GENOMIC CONTEXT AND PREDICTION OFNEW COMPONENTS OF THE REPLICATIONMACHINERY
The results of comparative genomics outlinedhere show that despite the fundamental impor-tance of DNA replication, the protein machineryresponsible for this process shows remarkableevolutionary plasticity that involves multipleevents of gene loss, non-orthologous displace-ment, and lineage-specific duplication, evenamong the genes encoding core replication pro-teins. Examples of such events leading to com-plex evolutionary scenarios are the substitutiona B-family polymerase for the D-family poly-merase in Crenarchaeota and substitution ofThermoSSB for RPA in Thermoproteales. More-over, some components of replication systemsevolve rapidly and often lose sequence similarity,which obscures their origin. However, in manysuch cases, the rapidlyevolving gene remains in aconserved and functionally coherent genomicneighborhood (Figs. 3 and 5). One such exam-ple is the extremely divergent RecJ homolog inThermoproteales and another is the numerousGINS proteins that are still annotated as “hypo-thetical” in sequenced archaeal genomes. Thesehighly divergent proteins are encoded in thesame neighborhoods as their better conservedhomologs, which facilitates functional predic-tion. Moreover, using the “guilt by association”principle (Aravind 2000; Galperin and Koonin2000), conservation of gene neighborhoods canbe used for prediction of new genes associated
with replication. However, this approach re-quires caution because some housekeepinggenes encoded in the same locus might not befunctionally related but rather are highly ex-pressed genes that “hitchhike” with genes thatare not directly functional but are expressed sim-ilarly (Rogozin et al. 2002). In particular, genesinvolved in replication are often associated withgenes coding for components of the translationsystem (Fig. 5) (Berthon et al. 2008). Thus, un-characterized genes in the respective operonsmight not be involved in replication directlybut are nevertheless interesting targets for exper-imental study (Figs. 3 and 5; Table 1). Recently,for example, analysis of gene neighborhoods ledto the prediction that such genes as PACE12,a member of the GPN-loop GTPase family;and NudF, a NUDIX pyrophosphatase familymember are involved in DNA replication and/or repair in archaea (Berthon et al. 2008). Herewe also identified several uncharacterized genesthat could be involved in these processes (Figs. 3and 5; Table 1). In addition, because replicationis a complex, metabolically costly process, manyother proteins and cellular systems could be di-rectly or indirectly involved in replication. Re-cent analysis of proteins interacting with corereplication proteins in T. kodakarensis suggeststhat there could be dozens of such partners, andfor many of these, the specific function is notknown (Li et al. 2010). Thus, all recent advancesnotwithstanding, the current understanding ofthe archaeal replication system is still far frombeing complete.
GENERAL TRENDS IN THE EVOLUTIONOF ARCHAEAL AND EUKARYOTICREPLICATION SYSTEMS
The study of archaeal replication is current-ly a dynamic research field that continuesto identify previously uncharacterized proteincomponents of the replication machinery,most of which show new connections with theeukaryotic replication machinery. Comparativegenomic analysis reveals a (nearly) precise cor-respondence between the components of the ar-chaeal and eukaryotic replication systems, witha few notable exceptions such as the CMG-
K.S. Makarova and E.V. Koonin
20 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

activating proteins for which archaeal counter-parts have not been detected (Fig. 1; Table 1).Thus, it appears most likely that the archaealancestor of eukaryotes possessed a DNA repli-cation apparatus that was as complex in its mainfeatures as the eukaryotic replication apparatus.Given this essentially precise correspondencebetween the archaeal and eukaryotic replicationsystems, it can be expected that additional rep-lication proteins shared between archaea andeukaryotes eventually will be discovered (Fig.1; Table 1). A major innovation in eukaryotescould be the regulation of replication by phos-phorylation of replisome subunits catalyzed bydedicated kinases. Whether a counterpart tothis regulatory circuit exists in archaea remainsto be determined.
The close correspondence between the com-ponents of the archaeal and eukaryotic rep-lication systems notwithstanding, eukaryoticreplication does show increased complexitycompared with archaeal replication. The prin-cipal modality of this greater complexity is du-plication of the replication machinery compo-nents followed by subfunctionalization (Lynchand Force 2000; Ward and Durrett 2004) where-by heteromeric complexes in eukaryotes re-place homomeric complexes in archaea. Theobvious cases in point are the ORC, MCM,GINS, and RFC complexes. This evolutionarytrend in the replication machinery is part of ageneral relationship between eukaryotes andtheir prokaryotic ancestors as exemplified bythe proteasome, the exosome, and many othermacromolecular complexes (Makarova et al.2005).
Although, on the whole, the eukaryotic rep-lication system is more complex than the ar-chaeal counterparts, complicated histories oflineage-specific expansion of gene families ac-companiedby functional diversification aboundin archaea. Unlike eukaryotes, where the mainroute to increased complexity is serial intra-genomic gene duplication, in archaea, repliconfusion and more generally horizontal genetransfer are major factors of evolution, leadingin particular to pseudoparalogy (Makarova etal. 2005). Strikingly, in some families of proteinsinvolved in replication, archaea attain far greater
diversity than eukaryotes, the RecJ family beingthe prime example. In other gene families, mul-tiple, independent duplications are traceable ineukaryotes and in different lineages of archaea.The degree of paralogization of the archaeal rep-lication system remains underappreciated, andmany divergent paralogs still await functionalcharacterization.
In addition to the diversification via paral-ogization, archaeal replication systems show lessuniformity and a greater plasticity than the eu-karyotic counterparts. The striking examples in-clude non-orthologous displacement of DNApolymerases, SSB, and ligases. The actual scopeof this plasticity is still unexplored because evenamong the relatively few archaeal genomes se-quenced to date, some gaps within the core ofthe replication machinery are apparent such asthe lack of ORC subunits in M. kandleri, sugges-tive of additional displacements of essentialcomponents.
In summary, the recent experimental andphylogenomic advances in the study of eukary-otic and archaeal replication systems clearlycomplement each other and jointly are yieldingan increasingly complete picture of the organi-zation and evolution of the core replication ma-chinery. This obvious progress notwithstand-ing, the identity and roles of more peripheralcomponents of the replication apparatus, thefunctions of paralogs of replicative genes, andespecially, the ultimate origins of the complexreplication machinery that is inferred to haveexisted in LACA, remain poorly understoodand are targets for future investigation.
ACKNOWLEDGMENTS
The authors’ research is supported by the intra-mural funds of the U.S. Department of Healthand Human Services (to the National Library ofMedicine).
REFERENCES
Adachi J, Hasegawa M. 1992. MOLPHY: Programs for mo-lecular phylogenetics. In Computer science monographs27. Institute of Statistical Mathematics, Tokyo.
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 21
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Araki H. 2010. Cyclin-dependent kinase-dependent initia-tion of chromosomal DNA replication. Curr Opin CellBiol 22: 766–771.
Aravind L. 2000. Guilt by association: Contextual informa-tion in genome analysis. Genome Res 10: 1074–1077.
Aravind L, Koonin EV. 1998. Phosphoesterase domains as-sociated with DNA polymerases of diverse origins. Nu-cleic Acids Res 26: 3746–3752.
Aravind L, Walker DR, Koonin EV. 1999. Conserved do-mains in DNA repair proteins and evolution of repairsystems. Nucleic Acids Res 27: 1223–1242.
Balasov M, Huijbregts RP, Chesnokov I. 2009. Functionalanalysis of an Orc6 mutant in Drosophila. Proc Natl AcadSci 106: 10672–10677.
Barry ER, Bell SD. 2006. DNA replication in the Archaea.Microbiol Mol Biol Rev 70: 15: 614–619.
Beattie TR, Bell SD. 2011a. Molecular machines in archaealDNA replication. Curr Opin Chem Biol 15: 614–619.
Beattie TR, Bell SD. 2011b. The role of the DNA slidingclamp in Okazaki fragment maturation in archaea andeukaryotes. Biochem Soc Trans 39: 70–76.
Bell SD. 2011. DNA replication: Archaeal oriGINS. BMCBiol 9: 36.
Bell SP, Dutta A. 2002. DNA replication in eukaryotic cells.Annu Rev Biochem 71: 333–374.
Bernander R, Dasgupta S, Nordstrom K. 1991. The E. colicell cycle and the plasmid R1 replication cycle in theabsence of the DnaA protein. Cell 64: 1145–1153.
Bernstein KA, Gangloff S, Rothstein R. 2010. The RecQDNA helicases in DNA repair. Annu Rev Genet 44:393–417.
Berquist BR, DasSarma P, DasSarma S. 2007. Essential andnon-essential DNA replication genes in the model halo-philic Archaeon, Halobacterium sp. NRC-1. BMC Genet8: 31.
Berthon J, Cortez D, Forterre P. 2008. Genomic contextanalysis in Archaea suggests previously unrecognizedlinks between DNA replication and translation. GenomeBiol 9: R71.
Bloom LB. 2009. Loading clamps for DNA replication andrepair. DNA Repair (Amst) 8: 570–578.
Bochkarev A, Bochkareva E. 2004. From RPA to BRCA2:Lessons from single-stranded DNA binding by the OB-fold. Curr Opin Struct Biol 14: 36–42.
Bochman ML, Schwacha A. 2009. The Mcm complex: Un-winding the mechanism of a replicative helicase. Micro-biol Mol Biol Rev 73: 652–683.
Bohlke K, Pisani FM, Rossi M, Antranikian G. 2002. Archae-al DNA replication: Spotlight on a rapidly moving field.Extremophiles 6: 1–14.
Bork P, Hofmann K, Bucher P, Neuwald AF, Altschul SF,Koonin EV. 1997. A superfamily of conserved domainsin DNA damage-responsive cell cycle checkpoint pro-teins. FASEB J 11: 68–76.
Burgers PM, Koonin EV, Bruford E, Blanco L, Burtis KC,Christman MF, Copeland WC, Friedberg EC, Hanaoka F,Hinkle DC, et al. 2001. Eukaryotic DNA polymerases:Proposal for a revised nomenclature. J Biol Chem 276:43487–43490.
Cann IK, Komori K, Toh H, Kanai S, Ishino Y. 1998. Aheterodimeric DNA polymerase: Evidence that membersof Euryarchaeota possess a distinct DNA polymerase.Proc Natl Acad Sci 95: 14250–14255.
Capp C, Wu J, Hsieh TS. 2010. RecQ4: The second replica-tive helicase? Crit Rev Biochem Mol Biol 45: 233–242.
Chattopadhyay S, Bielinsky AK. 2007. Human Mcm10 reg-ulates the catalytic subunit of DNA polymerase-a andprevents DNA damage during replication. Mol Biol Cell18: 4085–4095.
Chia N, Cann I, Olsen GJ. 2010. Evolution of DNA replica-tion protein complexes in eukaryotes and Archaea. PLoSONE 5: e10866.
Coker JA, DasSarma P, Capes M, Wallace T, McGarrity K,Gessler R, Liu J, Xiang H, Tatusov R, Berquist BR, et al.2009. Multiple replication origins of Halobacterium sp.strain NRC-1: Properties of the conserved orc7-depen-dent oriC1. J Bacteriol 191: 5253–5261.
De Marco V, Gillespie PJ, Li A, Karantzelis N, Christo-doulou E, Klompmaker R, van Gerwen S, Fish A,Petoukhov MV, Iliou MS, et al. 2009. Quaternary struc-ture of the human Cdt1–Geminin complex regulatesDNA replication licensing. Proc Natl Acad Sci 106:19807–19812.
Duncker BP, Chesnokov IN, McConkey BJ. 2009. The originrecognition complex protein family. Genome Biol 10: 214.
Edgar RC. 2004. MUSCLE: Multiple sequence alignmentwith high accuracy and high throughput. Nucleic AcidsRes 32: 1792–1797.
Edgell DR, Doolittle WF. 1997. Archaea and the origin(s) ofDNA replication proteins. Cell 89: 995–998.
Edgell DR, Malik SB, Doolittle WF. 1998. Evidence of inde-pendent gene duplications during the evolution of ar-chaeal and eukaryotic family B DNA polymerases. MolBiol Evol 15: 1207–1217.
Ellenberger T, Tomkinson AE. 2008. Eukaryotic DNA ligas-es: Structural and functional insights. Annu Rev Biochem77: 313–338.
Fien K, Hurwitz J. 2006. Fission yeast Mcm10p containsprimase activity. J Biol Chem 281: 22248–22260.
Fien K, Cho YS, Lee JK, Raychaudhuri S, Tappin I, HurwitzJ. 2004. Primer utilization by DNA polymerase a-pri-mase is influenced by its interaction with Mcm10p. JBiol Chem 279: 16144–16153.
Fitch WM, Margoliash E. 1967. Construction of phyloge-netic trees. Science 155: 279–284.
Forterre P. 2002. The orgin of DNA genomes and DNAreplication proteins. Curr Opin Microbiol 5: 525–532.
Forterre P. 2006. Three RNA cells for ribosomal lineages andthree DNA viruses to replicate their genomes: A hypoth-esis for the origin of cellular domain. Proc Natl Acad Sci103: 3669–3674.
Galperin MY, Koonin EV. 2000. Who’s your neighbor? Newcomputational approaches for functional genomics. NatBiotechnol 18: 609–613.
Garcia V, Furuya K, Carr AM. 2005. Identification and func-tional analysis of TopBP1 and its homologs. DNA Repair(Amst) 4: 1227–1239.
Glansdorff N, Xu Y, Labedan B. 2008. The last universalcommon ancestor: Emergence, constitution and geneticlegacy of an elusive forerunner. Biol Direct 3: 29.
K.S. Makarova and E.V. Koonin
22 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

Hamdan SM, Richardson CC. 2009. Motors, switches, andcontacts in the replisome. Annu Rev Biochem 78: 205–243.
Hamdan SM, van Oijen AM. 2010. Timing, coordination,and rhythm: Acrobatics at the DNA replication fork. JBiol Chem 285: 18979–18983.
Henneke G, Flament D, Hubscher U, Querellou J, Raffin JP.2005. The hyperthermophilic euryarchaeota Pyrococcusabyssi likely requires the two DNA polymerases D andB for DNA replication. J Mol Biol 350: 53–64.
Iyer LM, Leipe DD, Koonin EV, Aravind L. 2004. Evolution-ary history and higher order classification of AAAþ
ATPases. J Struct Biol 146: 11–31.
Kelman Z, White MF. 2005. Archaeal DNA replication andrepair. Curr Opin Microbiol 8: 669–676.
Klinge S, Nunez-Ramirez R, Llorca O, Pellegrini L. 2009. 3Darchitecture of DNA Pol a reveals the functional core ofmulti-subunit replicative polymerases. EMBO J 28:1978–1987.
Komori K, Ishino Y. 2001. Replication protein A in Pyrococ-cus furiosus is involved in homologous DNA recombina-tion. J Biol Chem 276: 25654–25660.
Koonin EV. 2006. Temporal order of evolution of DNA rep-lication systems inferred by comparision of cellular andviral DNA polymerases. Biol Direct 1: 39.
Koonin EV. 2009. On the origin of cells and viruses: primor-dial virus world scenario. Ann NY Acad Sci 1178: 47–64.
Koonin EV, Martin W. 2005. On the origin of genomes andcells within inorganic compartments. Trends Genet 21:647–654.
Koppes LJ. 1992. Nonrandom F-plasmid replication in Es-cherichia coli K-12. J Bacteriol 174: 2121–2123.
Kornberg A, Baker TA. 2005. DNA replication, 2nd ed. Uni-versity Science Books, Sausalito, CA.
Krupovic M, Gribaldo S, Bamford DH, Forterre P. 2010. Theevolutionary history of archaeal MCM helicases: A casestudy of vertical evolution combined with hitchhiking ofmobile genetic elements. Mol Biol Evol 27: 2716–2732.
Kuchta RD, Stengel G. 2010. Mechanism and evolution ofDNA primases. Biochim Biophys Acta 1804: 1180–1189.
Kunkel TA, Burgers PM. 2008. Dividing the workload at aeukaryotic replication fork. Trends Cell Biol 18: 521–527.
Labib K, Gambus A. 2007. A key role for the GINS complexat DNA replication forks. Trends Cell Biol 17: 271–278.
Lee C, Hong B, Choi JM, Kim Y, Watanabe S, Ishimi Y,Enomoto T, Tada S, Cho Y. 2004. Structural basis forinhibition of the replication licensing factor Cdt1 bygeminin. Nature 430: 913–917.
Leipe DD, Aravind L, Koonin EV. 1999. Did DNA replica-tion evolve twice independently? Nucleic Acids Res 27:3389–3401.
Li Z, Santangelo TJ, Cubonova L, Reeve JN, Kelman Z. 2010.Affinity purification of an archaeal DNA replication pro-tein network. MBio 1: e00221-10.
Li Z, Pan M, Santangelo TJ, Chemnitz W, Yuan W, Ed-wards JL, Hurwitz J, Reeve JN, Kelman Z. 2011. A novelDNA nuclease is stimulated by association with the GINScomplex. Nucleic Acids Res 39: 6114–6123.
Liu S, Balasov M, Wang H, Wu L, Chesnokov IN, Liu Y. 2011.Structural analysis of human Orc6 protein reveals a ho-
mology with transcription factor TFIIB. Proc Natl AcadSci 108: 7373–7378.
Lynch M, Force A. 2000. The probability of duplicate genepreservation by subfunctionalization. Genetics 154: 459–473.
MacNeill SA. 2010. Structure and function of the GINScomplex, a key component of the eukaryotic replisome.Biochem J 425: 489–500.
MacNeill SA. 2011. Protein–protein interactions in the ar-chaeal core replisome. Biochem Soc Trans 39: 163–168.
Majka J, Burgers PM. 2004. The PCNA-RFC families ofDNA clamps and clamp loaders. Prog Nucleic Acid ResMol Biol 78: 227–260.
Makarova KS, Koonin EV. 2010. Two new families of theFtsZ-tubulin protein superfamily implicated in mem-brane remodeling in diverse bacteria and archaea. BiolDirect 5: 33.
Makarova KS, Aravind L, Galperin MY, Grishin NV, Ta-tusov RL, Wolf YI, Koonin EV. 1999. Comparative geno-mics of the Archaea (Euryarchaeota): Evolution of con-served protein families, the stable core, and the variableshell. Genome Res 9: 608–628.
Makarova KS, Wolf YI, Mekhedov SL, Mirkin BG,Koonin EV. 2005. Ancestral paralogs and pseudoparalogsand their role in the emergence of the eukaryotic cell.Nucleic Acids Res 33: 4626–4638.
Makarova KS, Yutin N, Bell SD, Koonin EV. 2010. Evolutionof diverse cell division and vesicle formation systems inarchaea. Nat Rev Microbiol 8: 731–741.
Makarova KS, Koonin EV, Kelman Z. 2012. The archaealCMG (CDC45/RecJ, MCM, GINS) complex is a con-served component of the DNA replication system in allarchaea and eukaryotes. Biol Direct 7: 7.
Marinsek N, Barry ER, Makarova KS, Dionne I, Koonin EV,Bell SD. 2006. GINS, a central nexus in the archaeal DNAreplication fork. EMBO Rep 7: 539–545.
Martin IV, MacNeill SA. 2002. ATP-dependent DNA ligases.Genome Biol 3: REVIEWS3005.
Matsui I, Urushibata Y, Shen Y, Matsui E, Yokoyama H. 2011.Novel structure of an N-terminal domain that is crucialfor the dimeric assembly and DNA-binding of an archae-al DNA polymerase D large subunit from Pyrococcus ho-rikoshii. FEBS Lett 585: 452–458.
McGeoch AT, Bell SD. 2008. Extra-chromosomal elementsand the evolution of cellular DNA replication machiner-ies. Nat Rev Mol Cell Biol 9: 569–574.
Mer G, Bochkarev A, Gupta R, Bochkareva E, Frappier L,Ingles CJ, Edwards AM, Chazin WJ. 2000. Structural ba-sis for the recognition of DNA repair proteins UNG2,XPA, and RAD52 by replication factor RPA. Cell 103:449–456.
Moyer SE, Lewis PW, Botchan MR. 2006. Isolation of theCdc45/Mcm2–7/GINS (CMG) complex, a candidatefor the eukaryotic DNA replication fork helicase. ProcNatl Acad Sci 103: 10236–10241.
Murzin AG. 1993. OB(oligonucleotide/oligosaccharide bind-ing)-fold: Common structural and functional solutionfor non-homologous sequences. EMBO J 12: 861–867.
Mushegian AR, Bassett DE Jr, Boguski MS, Bork P,Koonin EV. 1997. Positionally cloned human disease
Evolution of DNA Replication
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 23
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

genes: Patterns of evolutionary conservation and func-tional motifs. Proc Natl Acad Sci 94: 5831–5836.
Narasingarao P, Podell S, Ugalde JA, Brochier-Armanet C,Emerson JB, Brocks JJ, Heidelberg KB, Banfield JF,Allen EE. 2012. De novo metagenomic assembly revealsabundant novel major lineage of Archaea in hypersalinemicrobial communities. ISME J 6: 81–93.
Ng WL, DasSarma S. 1993. Minimal replication origin of the200-kilobase Halobacterium plasmid pNRC100. J Bacter-iol 175: 4584–4596.
Okorokov AL, Waugh A, Hodgkinson J, Murthy A, HongHK, Leo E, Sherman MB, Stoeber K, Orlova EV,Williams GH. 2007. Hexameric ring structure of humanMCM10 DNA replication factor. EMBO Rep 8: 925–930.
Oyama T, Ishino S, Fujino S, Ogino H, Shirai T, Mayanagi K,Saito M, Nagasawa N, Ishino Y, Morikawa K. 2011. Ar-chitectures of archaeal GINS complexes, essential DNAreplication initiation factors. BMC Biol 9: 28.
Pacek M, Tutter AV, Kubota Y, Takisawa H, Walter JC. 2006.Localization of MCM2–7, Cdc45, and GINS to the site ofDNA unwinding during eukaryotic DNA replication.Mol Cell 21: 581–587.
Pan M, Kelman LM, Kelman Z. 2011a. The archaeal PCNAproteins. Biochem Soc Trans 39: 20–24.
Pan M, Santangelo TJ, Li Z, Reeve JN, Kelman Z. 2011b.Thermococcus kodakarensis encodes three MCM homo-logs but only one is essential. Nucleic Acids Res 39:9671–9680.
Paytubi S, McMahon SA, Graham S, Liu H, Botting CH,Makarova KS, Koonin EV, Naismith JH, White MF.2012. Displacement of the canonical single-strandedDNA-binding protein in the Thermoproteales. ProcNatl Acad Sci 109: E398–E405.
Pospiech H, Grosse F, Pisani FM. 2010. The initiation step ofeukaryotic DNA replication. Subcell Biochem 50: 79–104.
Price MN, Dehal PS, Arkin AP. 2010. FastTree 2—Approx-imately maximum-likelihood trees for large alignments.PLoS ONE 5: e9490.
Robbins JB, McKinney MC, Guzman CE, Sriratana B, Fitz-Gibbon S, Ha T, Cann IK. 2005. The Euryarchaeota, na-ture’s medium for engineering of single-stranded DNA-binding proteins. J Biol Chem 280: 15325–15339.
Robertson PD, Chagot B, Chazin WJ, Eichman BF. 2010.Solution NMR structure of the C-terminal DNA bindingdomain of Mcm10 reveals a conserved MCM motif. J BiolChem 285: 22942–22949.
Robinson NP, Bell SD. 2005. Origins of DNA replication inthe three domains of life. FEBS J 272: 3757–3766.
Robinson NP, Bell SD. 2007. Extrachromosomal elementcapture and the evolution of multiple replication originsin archaeal chromosomes. Proc Natl Acad Sci 104: 5806–5811.
Robinson NP, Dionne I, Lundgren M, Marsh VL, Ber-nander R, Bell SD. 2004. Identification of two origins ofreplication in the single chromosome of the archaeonSulfolobus solfataricus. Cell 116: 25–38.
Rogozin IB, Makarova KS, Murvai J, Czabarka E, Wolf YI,Tatusov RL, Szekely LA, Koonin EV. 2002. Connectedgene neighborhoods in prokaryotic genomes. Nucleic Ac-ids Res 30: 2212–2223.
Rogozin IB, Makarova KS, Pavlov YI, Koonin EV. 2008. Ahighly conserved family of inactivated archaeal B familyDNA polymerases. Biol Direct 3: 32.
Sakakibara N, Kelman LM, Kelman Z. 2009. Unwinding thestructure and function of the archaeal MCM helicase.Mol Microbiol 72: 286–296.
Sanchez-Pulido L, Ponting CP. 2011. Cdc45: The missingRecJ ortholog in eukaryotes? Bioinformatics 27: 1885–1888.
Sanchez-Pulido L, Diffley JF, Ponting CP. 2010. Homologyexplains the functional similarities of Treslin/Ticrr andSld3. Curr Biol 20: R509–R510.
Sclafani RA, Holzen TM. 2007. Cell cycle regulation of DNAreplication. Annu Rev Genet 41: 237–280.
Shen Y, Musti K, Hiramoto M, Kikuchi H, Kawarabayashi Y,Matsui I. 2001. Invariant Asp-1122 and Asp-1124 areessential residues for polymerization catalysis of familyD DNA polymerase from Pyrococcus horikoshii. J BiolChem 276: 27376–27383.
Sheu YJ, Stillman B. 2010. The Dbf4–Cdc7 kinase promotesS phase by alleviating an inhibitory activity in Mcm4.Nature 463: 113–117.
Shutt TE, Gray MW. 2006. Twinkle, the mitochondrial rep-licative DNA helicase, is widespread in the eukaryoticradiation and may also be the mitochondrial DNA pri-mase in most eukaryotes. J Mol Evol 62: 588–599.
Sun X, Thrower D, Qiu J, Wu P, Zheng L, Zhou M, Bachant J,Wilson DM 3rd, Shen B. 2003. Complementary func-tions of the Saccharomyces cerevisiae Rad2 family nucle-ases in Okazaki fragment maturation, mutation avoid-ance, and chromosome stability. DNA Repair (Amst) 2:925–940.
Sun J, Yu EY, Yang Y, Confer LA, Sun SH, Wan K, Lue NF,Lei M. 2009. Stn1–Ten1 is an Rpa2–Rpa3–like complexat telomeres. Genes Dev 23: 2900–2914.
Sun J, Yang Y, Wan K, Mao N, Yu TY, Lin YC, DeZwaan DC,Freeman BC, Lin JJ, Lue NF, et al. 2011. Structural bases ofdimerization of yeast telomere protein Cdc13 and its in-teraction with the catalytic subunit of DNA polymerasealpha. Cell Res 21: 258–274.
Tahirov TH, Makarova KS, Rogozin IB, Pavlov YI, KooninEV. 2009. Evolution of DNA polymerases: An inactivatedpolymerase–exonuclease module in Pol 1 and a chimericorigin of eukaryotic polymerases from two classes of ar-chaeal ancestors. Biol Direct 4: 11.
Tanaka S, Umemori T, Hirai K, Muramatsu S, Kamimura Y,Araki H. 2007. CDK-dependent phosphorylation of Sld2and Sld3 initiates DNA replication in budding yeast. Na-ture 445: 328–332.
Wadsworth RI, White MF. 2001. Identification and proper-ties of the Crenarchaeal single-stranded DNA bindingprotein from Sulfolobus solfataricus. Nucleic Acids Res29: 914–920.
Walters AD, Chong JP. 2010. An archaeal order with multi-ple minichromosome maintenance genes. Microbiology156: 1405–1414.
Ward R, Durrett R. 2004. Subfunctionalization: How oftendoes it occur? How long does it take? Theor Popul Biol 66:93–100.
Warren EM, Vaithiyalingam S, Haworth J, Greer B,Bielinsky AK, Chazin WJ, Eichman BF. 2008. Structural
K.S. Makarova and E.V. Koonin
24 Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

basis for DNA binding by replication initiator Mcm10.Structure 16: 1892–1901.
Wei Z, Liu C, Wu X, Xu N, Zhou B, Liang C, Zhu G. 2010.Characterization and structure determination of theCdt1 binding domain of human minichromosomemaintenance (Mcm) 6. J Biol Chem 285: 12469–12473.
Wohlschlegel JA, Dhar SK, Prokhorova TA, Dutta A,Walter JC. 2002. Xenopus Mcm10 binds to origins ofDNA replication after Mcm2–7 and stimulates originbinding of Cdc45. Mol Cell 9: 233–240.
Yutin N, Koonin EV. 2009. Evolution of DNA ligases ofnucleo-cytoplasmic large DNA viruses of eukaryotes: Acase of hidden complexity. Biol Direct 4: 51.
Zhang R, Zhang CT. 2005. Identification of replication or-igins in archaeal genomes based on the Z-curve method.Archaea 1: 335–346.
Zuo Z, Rodgers CJ, Mikheikin AL, Trakselis MA. 2010.Characterization of a functional DnaG-type primase inArchaea: Implications for a dual-primase system. J MolBiol 397: 664–676.
Cite this article as Cold Spring Harb Perspect Biol 2013;5:a012963 25
Evolution of DNA Replication
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from

23, 20132013; doi: 10.1101/cshperspect.a012963 originally published online JulyCold Spring Harb Perspect Biol
Kira S. Makarova and Eugene V. Koonin Archaeology of Eukaryotic DNA Replication
Subject Collection DNA Replication
Barr Viral DNA−Replication of EpsteinWolfgang Hammerschmidt and Bill Sugden
Endoreplication
DePamphilisNorman Zielke, Bruce A. Edgar and Melvin L.
Replication Proteins and Human Disease
Nicholas ColemanAndrew P. Jackson, Ronald A. Laskey and
Replication-Fork Dynamics
Antoine M. van Oijen, et al.Karl E. Duderstadt, Rodrigo Reyes-Lamothe,
Break-Induced DNA Replication
HaberRanjith P. Anand, Susan T. Lovett and James E.
ReplicationReplication Forks at Chromosomal Origins of Helicase Activation and Establishment of
Seiji Tanaka and Hiroyuki ArakiRegulating DNA Replication in Eukarya
Khalid Siddiqui, Kin Fan On and John F.X. DiffleyPoxvirus DNA Replication
Bernard MossArchaeology of Eukaryotic DNA Replication
Kira S. Makarova and Eugene V. Koonin HelicaseThe Minichromosome Maintenance Replicative
Stephen D. Bell and Michael R. BotchanTranslesion DNA Polymerases
Myron F. Goodman and Roger WoodgateDNA Replication Origins
Alan C. Leonard and Marcel Méchali
Cancer?Human Papillomavirus Infections: Warts or
Louise T. Chow and Thomas R. BrokerBacteria, Archaea, and EukaryaPrinciples and Concepts of DNA Replication in
StillmanMichael O'Donnell, Lance Langston and Bruce
Chromatin and DNA ReplicationDavid M. MacAlpine and Geneviève Almouzni
DNA Replication TimingNicholas Rhind and David M. Gilbert
http://cshperspectives.cshlp.org/cgi/collection/ For additional articles in this collection, see
Copyright © 2013 Cold Spring Harbor Laboratory Press; all rights reserved
on July 8, 2018 - Published by Cold Spring Harbor Laboratory Press http://cshperspectives.cshlp.org/Downloaded from





























