Embed Size (px)
Citation preview

Eukaryotic DNA Replication

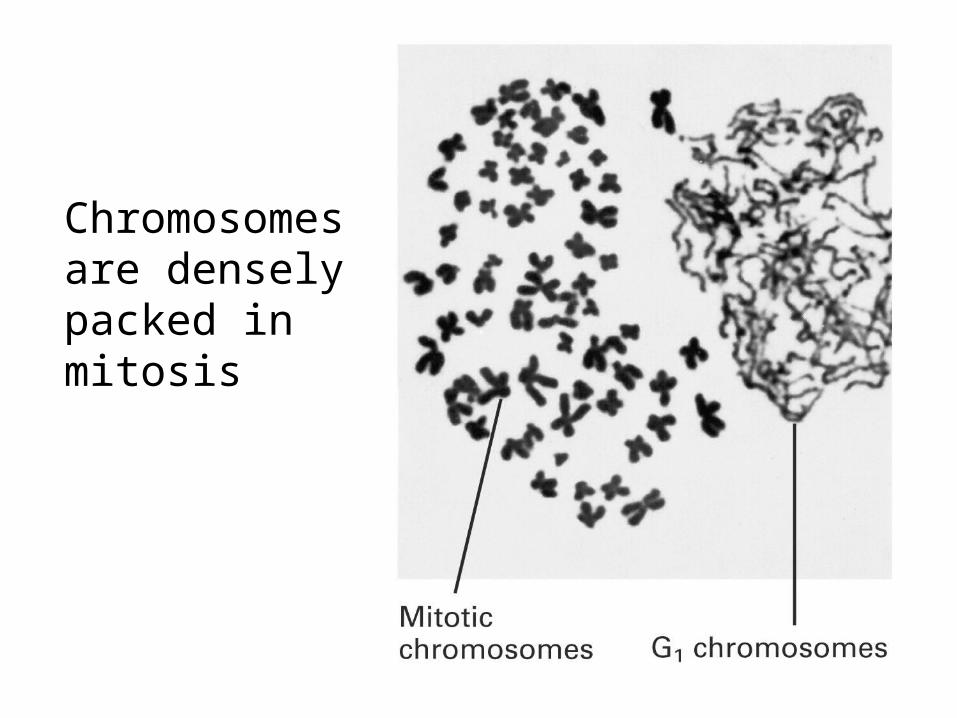
Chromosomesare denselypacked inmitosis

The accuracy of DNA replication is seen in the quality of the product
The accuracy of DNA replication is seen in the quality of the product
Duplication of DNA
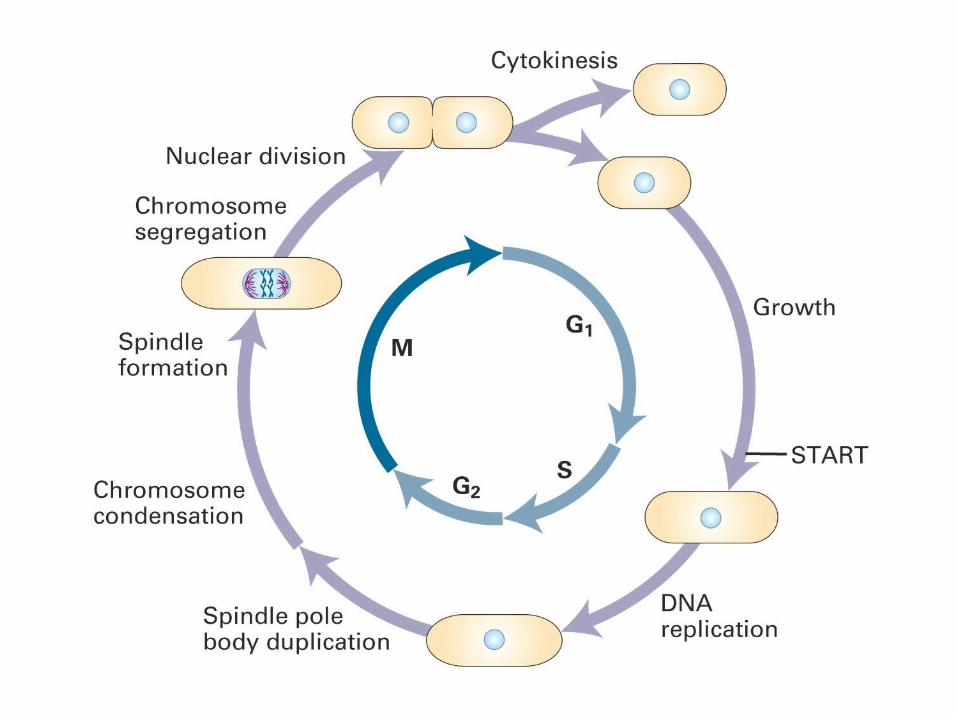
Cell Division
Fertilised EggProduct
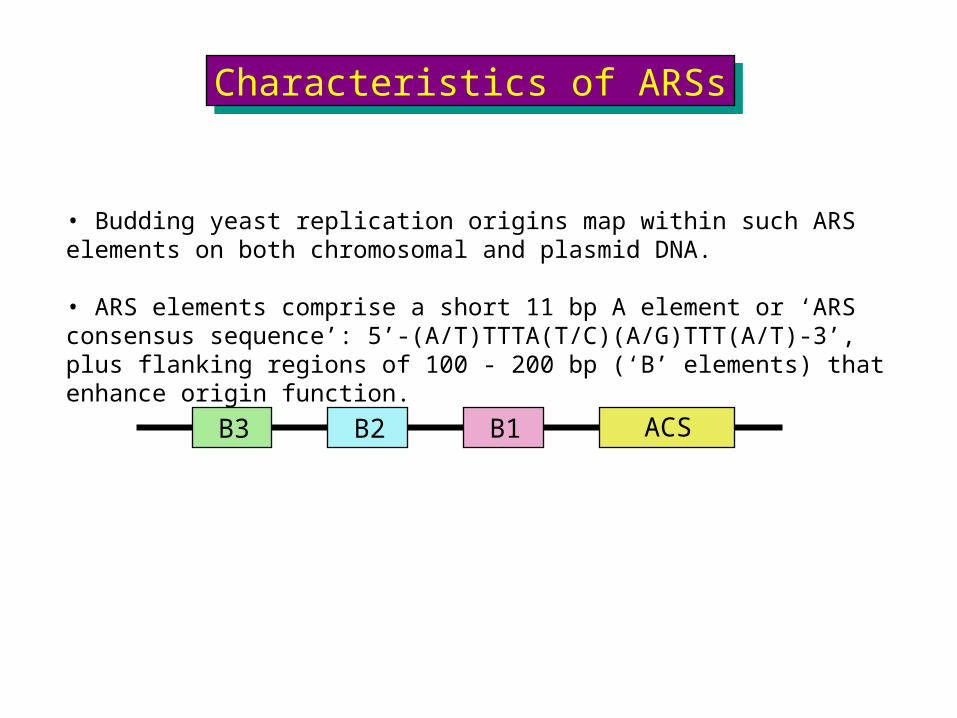
ACSB1B3 B2
• Budding yeast replication origins map within such ARS elements on both chromosomal and plasmid DNA.
• ARS elements comprise a short 11 bp A element or ‘ARS consensus sequence’: 5’-(A/T)TTTA(T/C)(A/G)TTT(A/T)-3’, plus flanking regions of 100 - 200 bp (‘B’ elements) that enhance origin function.
Characteristics of ARSsCharacteristics of ARSs

Which proteins bind to and define eukaryotic replication origins?

Replication origins in metazoans (somatic cells)Replication origins in metazoans (somatic cells)
• The structure of replication origins in higher eukaryotes is unclear.
• Small extrachromosomal DNA sequences replicate poorly, even when carrying >10 kb genomic DNA known to act as origins when in the chromosome.
• Replication initiates at specific regions at a characteristic time in S phase. Both place and timing may change with cell type.
• Replication forks can potentially initiate at a number of different sites throughout an “initiation zone” that may extend over >10 kb.

The ‘Origin Number’ ParadoxThe ‘Origin Number’ ParadoxE. coli:
Genome, 4 Mb = 4 x 106 bpFork rate approx. 800 bp / secReplication time approx. 40 minutes = 2,400 secs
Amount replicated by 2 forks in 40 mins = 2 x 2400 x 800 = 3,840,000 bp (~4 Mb)
Eukaryotes
Genome 20 Mb (yeast) up to 6,000 Mb (human)Fork rate 10 bp / sec (frog) - 50 bp / sec (mammal)
Amount replicated by 2 forks in 8 hr (human cells) = 2 x 50 x 28,800 = 2,880,000 (~ 3 Mb, a 2,000-fold deficit)
46 chromosomes (human cells) - with one origin per chromosome, at least 92 replication forks gives approx. 140 Mb replicated in 8 hours (still a 40-fold deficit)

The solution:- eukaryotes replicate their chromosomes from multiple replication origins
Electron micrograph showing an approx. 300 kb stretch of replicating chromosomal DNA from the yeast S. cerevisiae. Replication forks are indicated by an arrow. (Petes, Newlon, Byers, & Fangman 1974;Cold Spring Harb Symp Quant Biol. 38:9-16 ).

The study of replication origins using DNA fibre autoradiography
The study of replication origins using DNA fibre autoradiography
Interpretation:
Before pulse I:
End of pulse I:
End of pulse II:
Protocol:a) Pulse label proliferating cells with 3H-thymidine for 5 min (pulse I)b) Dilute label to 1/5 activity for further 5 min (pulse II)c) Isolate DNA and spread on a photographic plated) expose for 6 monthse) develop and examine grains under microscope
heavy labellinglight labelling

Chromosome regions replicate at different timesChromosome regions replicate at different times
BrdU
BrdU
BrdU
BrdU
BrdU
BrdUBrdU
BrdU
BrdU
BrdU
BrdU
BrdU
BrdUBrdUBrdU
BrdU
BrdU
BrdU
BrdU
BrdU
BrdU
BrdU
BrdU
BrdU
Protocol:
a) Pulse cells, at different times, with BrdU for 1 hr.
b) “Chase”, collect chromosomes.
c) Stain with anti-BrdU antibodies.
late S2 hr chase
mid S5 hr chase
early S9 hr chase
~ 8 ~2 ~1Duration (hours)
5 hr
9 hr
2 hrG1 S G2 M
= BrdU pulse
= chase
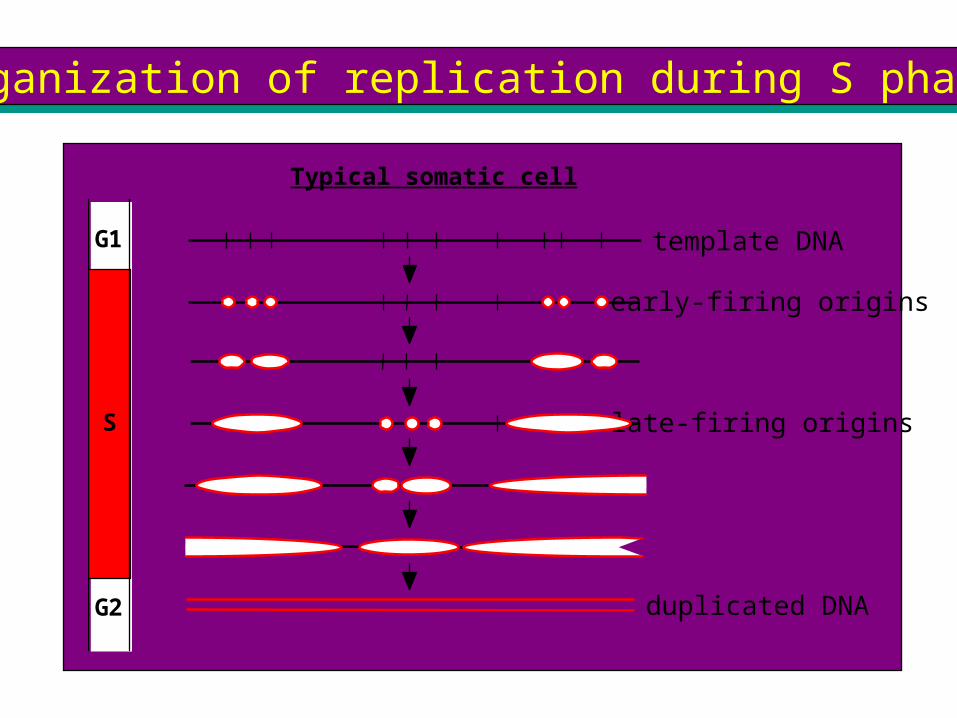
Organization of replication during S phaseOrganization of replication during S phase
G1
S
G2
template DNA
Typical somatic cell
early-firing origins
late-firing origins
duplicated DNA
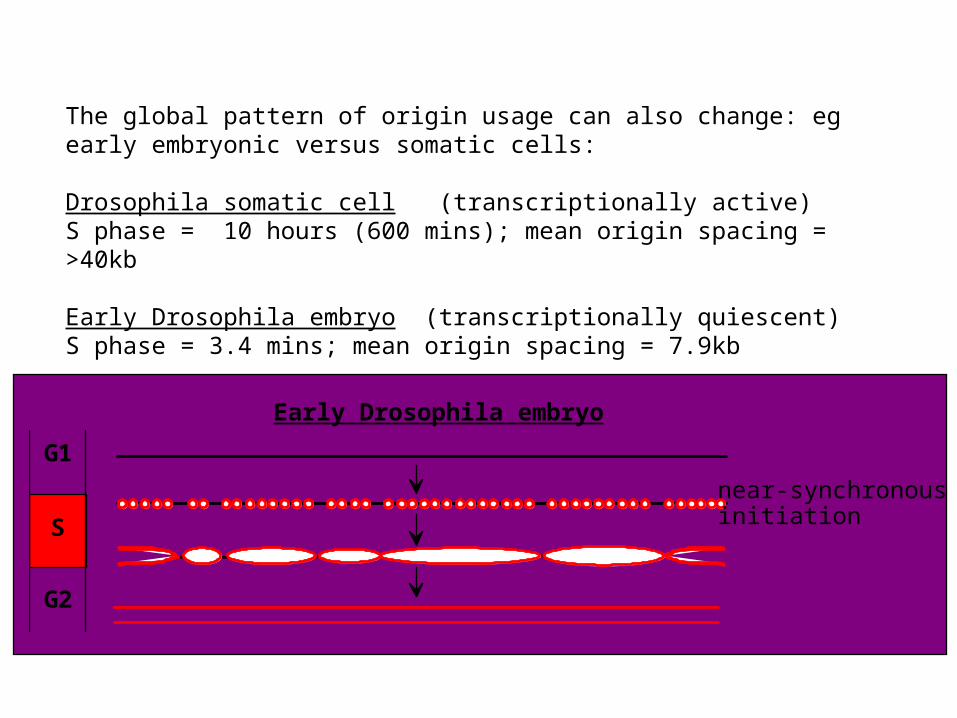
The global pattern of origin usage can also change: eg early embryonic versus somatic cells:
Drosophila somatic cell (transcriptionally active)S phase = 10 hours (600 mins); mean origin spacing = >40kb
Early Drosophila embryo (transcriptionally quiescent)S phase = 3.4 mins; mean origin spacing = 7.9kb
G1
S
G2
near-synchronous initiation
Early Drosophila embryo

Why so many origins?Why so many origins?
Excess origins are used to lower the probability of a lethal ‘double stall’?
To prevent problems if origins do not initiate with 100% probability?
stalled fork
replication completed byother fork of pair
double stall: no way of replicating intervening DNA
To allow different sections of the genome to replicate at different times?
To allow sections of the genome to replicate faster?


Facts IFacts I
• Rate of progression of replication forks is fairly constant for a given organism
• Forks generally stop only when they encounter an oppositely moving fork• Chromosome replication is regulated mainly through control of the initiation of new replication forks
For example:-
-by regulating the number and spacing of origins that fire eg. during development
-by regulating the time during S phase at which different origins are activated

Facts IIFacts II
• In somatic mammalian cells, most inter-origin distances (replicon sizes) are between 30 - 300 kb (ie would take 5 - 50 min to replicate completely).
• Some adjacent origins (“origin clusters”, typically 2 - 5 origins) initiate synchronously
• Different origins / origin clusters initiate at different times during S phase
Typical mammalian cell replicates 6,000 Mb in 8 hr = 6 x 109 ÷ 28,800 bp/sec ie. ~200,000 bp/sec
For fork rate of 50 bp / sec = 200,000 ÷ 50 ~ 4,000 forks active at any given time in S phase
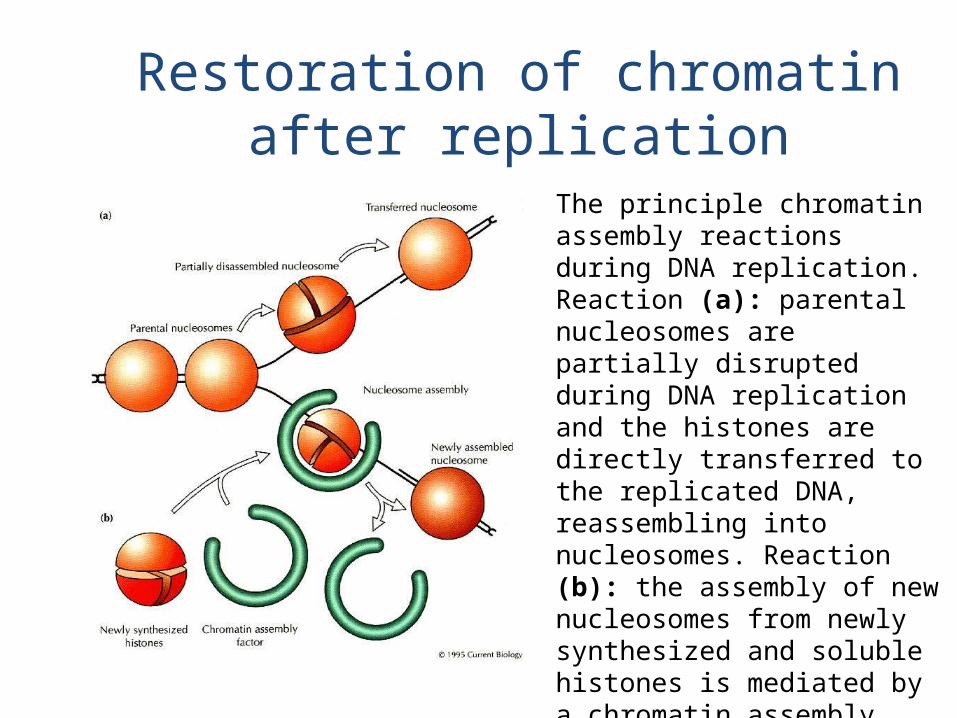
Restoration of chromatin after replication
The principle chromatin assembly reactions during DNA replication. Reaction (a): parental nucleosomes are partially disrupted during DNA replication and the histones are directly transferred to the replicated DNA, reassembling into nucleosomes. Reaction (b): the assembly of new nucleosomes from newly synthesized and soluble histones is mediated by a chromatin assembly factor
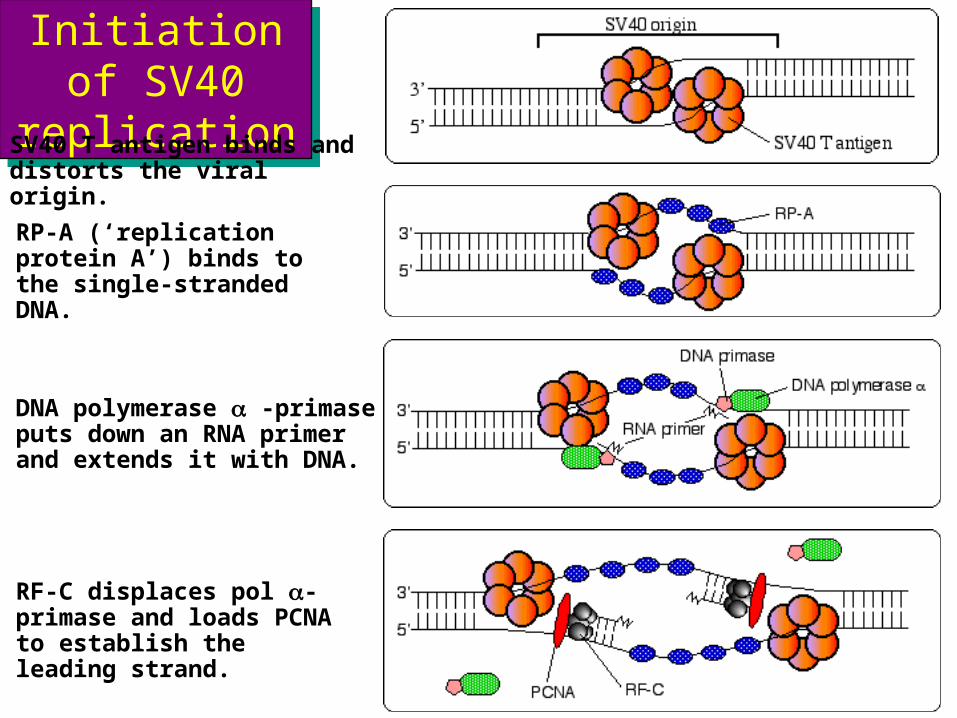
Initiation of SV40 replication
Initiation of SV40 replication
SV40 T antigen binds and distorts the viral origin.
RP-A (‘replication protein A’) binds to the single-stranded DNA.
DNA polymerase a -primase puts down an RNA primer and extends it with DNA.
RF-C displaces pol a-primase and loads PCNA to establish the leading strand.
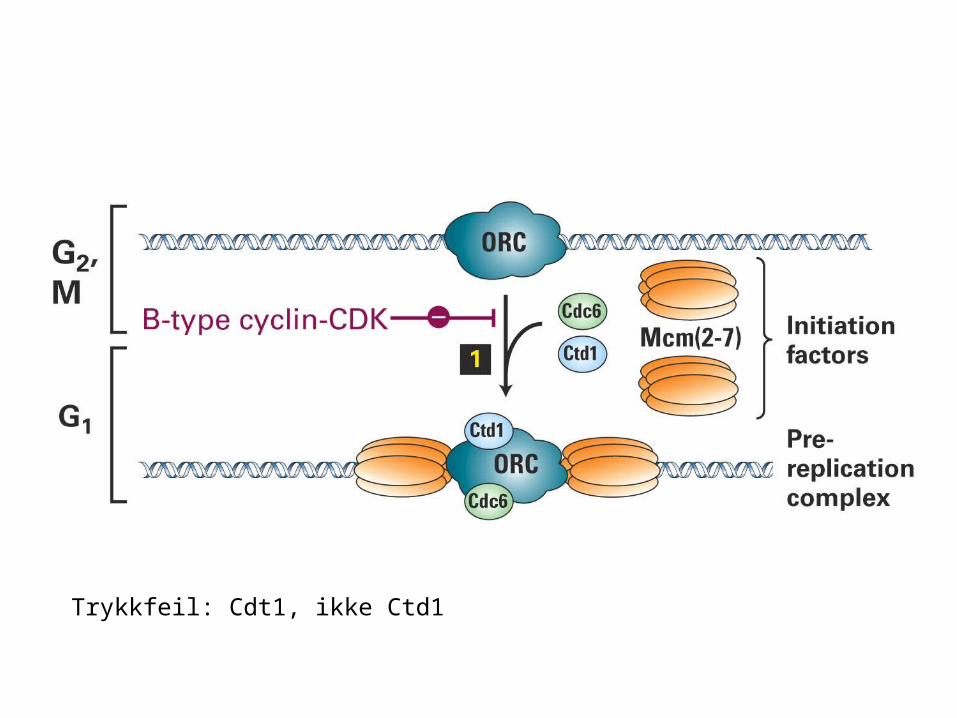
Trykkfeil: Cdt1, ikke Ctd1
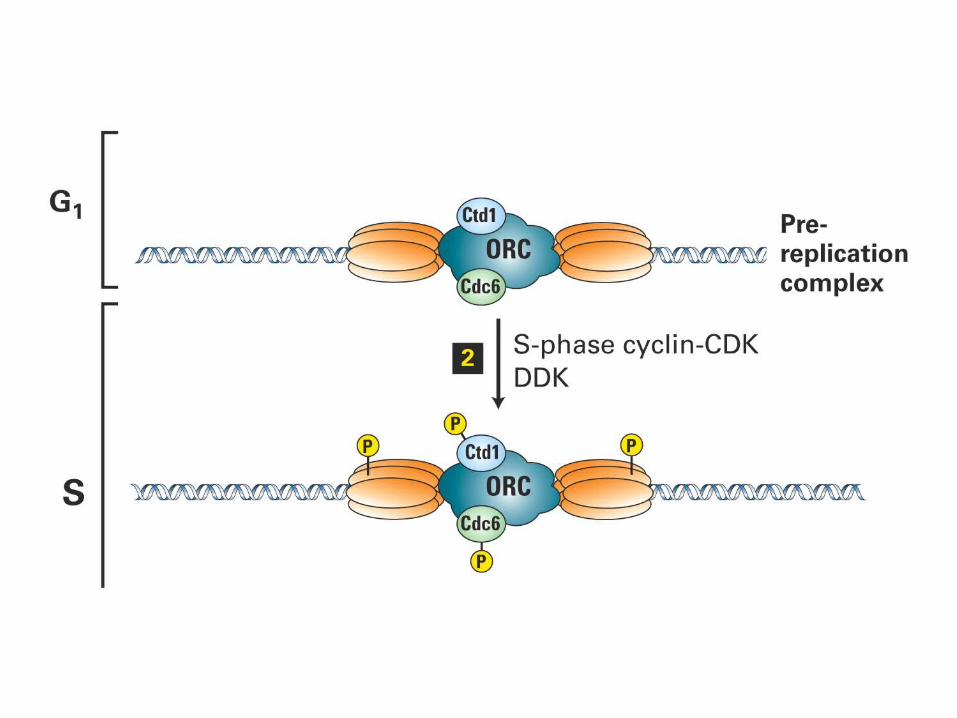

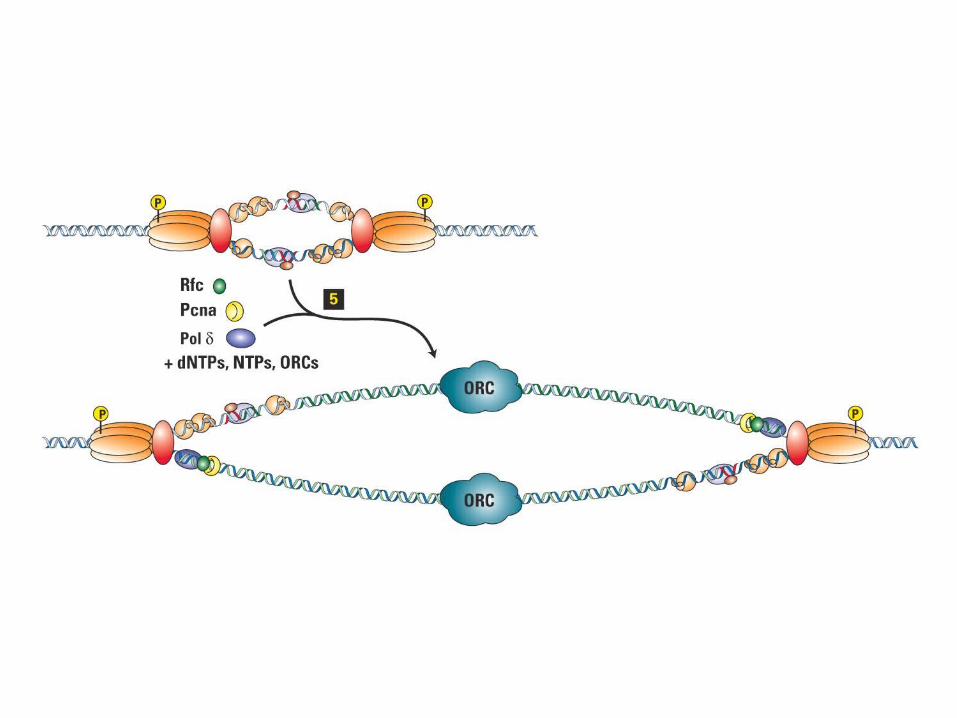


Mcm2-7Mcm2-7
Mcm2-7 (mini-chromosome maintenance) proteins were originally identified in yeast because as mutants affecting replication origin usage.
Fractionation showed them to be a key component of Licensing Factor.
They are loaded onto DNA in anaphase and are removed from chromatin during S phase.
They form a hexameric ring, capable of encircling double-stranded DNA.
Highly conserved throughout eukaryotes; archaea also possess an Mcm2-7 homologue
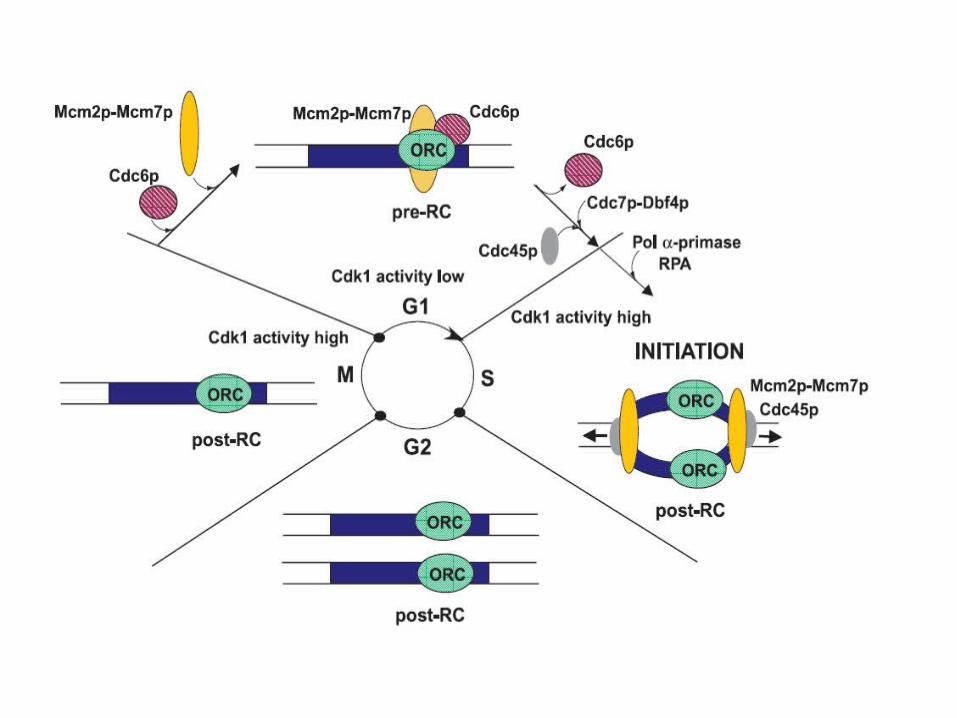

Recap
Whether a cell has only one chromosome (as in prokaryotes) or has many chromosomes (as in eukaryotes), the entire genome must be replicated precisely once of every cell division.
Two general principles: (1) Initiation of DNA replication commits the cell to a further division. Replication is controlled at the stage of initiation. Once replication has started, it continues until the entire genome has been duplicated. (2) If replication proceeds, the consequent division cannot be permitted to occur until the replication event has been completed.
Repliconthe unit of DNA in which an individual act of replication occurs.Originsite at which replication is initiated.Terminussite at which replication stops.

A genome in a prokaryotic cell constitutes a single replicon; thus the units of replication and segregation coincide.
A plasmid is an autonomous circular DNA genome that constitutes a separate replicon; may show single copy control or under multicopy control. Any DNA molecule that contains an origin can be replicated autonomously in the cell.
Each eukaryotic chromosome contains a large number of replicons; each must be activated no more than once in each cell cycle.
The DNA of mitochondria and chloroplasts may be regulated more like plasmids that exist in multiple copies per bacterium.

Replicons Can Be Linear or Circular
A replicated region appears as an eye within nonreplicated DNA.
A replication fork is initiated at the origin and then moves sequentially along DNA.
Replication is unidirectional when a single replication fork is created at an origin.
Replication is bidirectional when an origin creates two replication forks that move in opposite directions.
Key Concepts
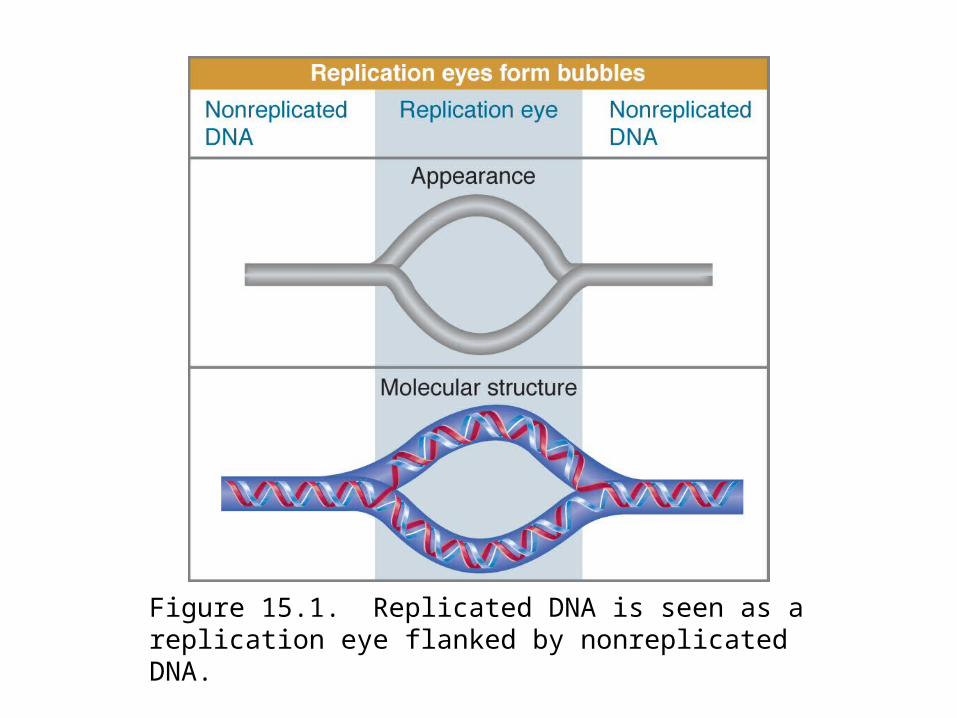
Figure 15.1. Replicated DNA is seen as a replication eye flanked by nonreplicated DNA.

Figure 15.2. Replicons may be unidirectional or bidirectional, depending on whether one or two replication forks are formed at the origin.

Figure 15.3. A replicatin eye forms a θ structure in circular DNA.

Origins Can Be Mapped by Autoradiography and Electrophoresis
Replication fork movement can be detected by autoradiography using radioactive pulses.
Replication forks create Y-shaped structures that change the electrophoretic migration of DNA fragments.
Key Concepts
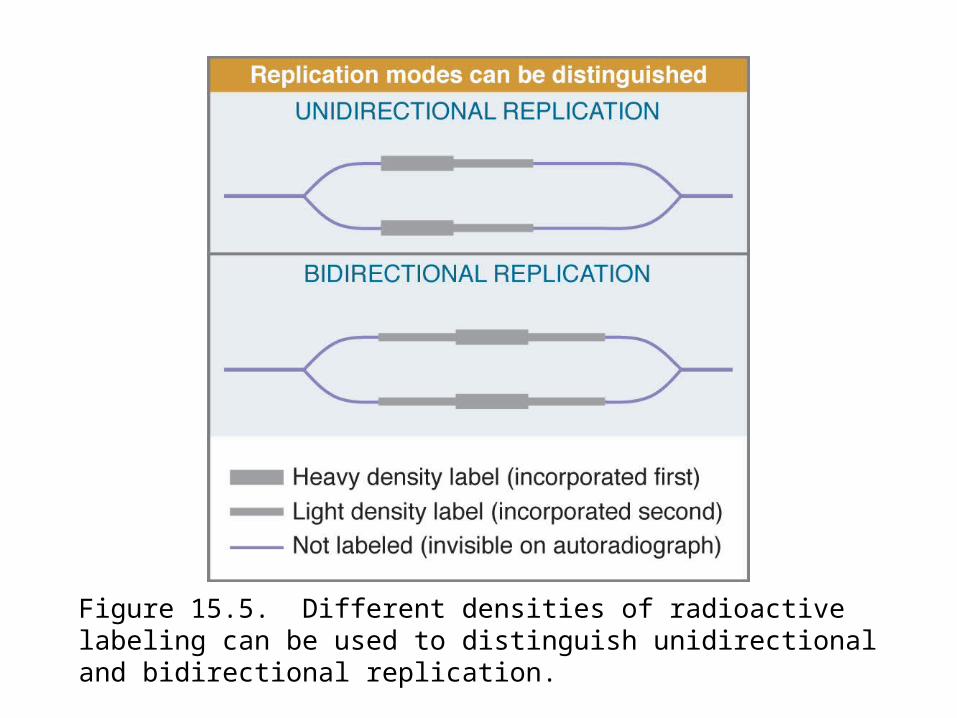
Figure 15.5. Different densities of radioactive labeling can be used to distinguish unidirectional and bidirectional replication.
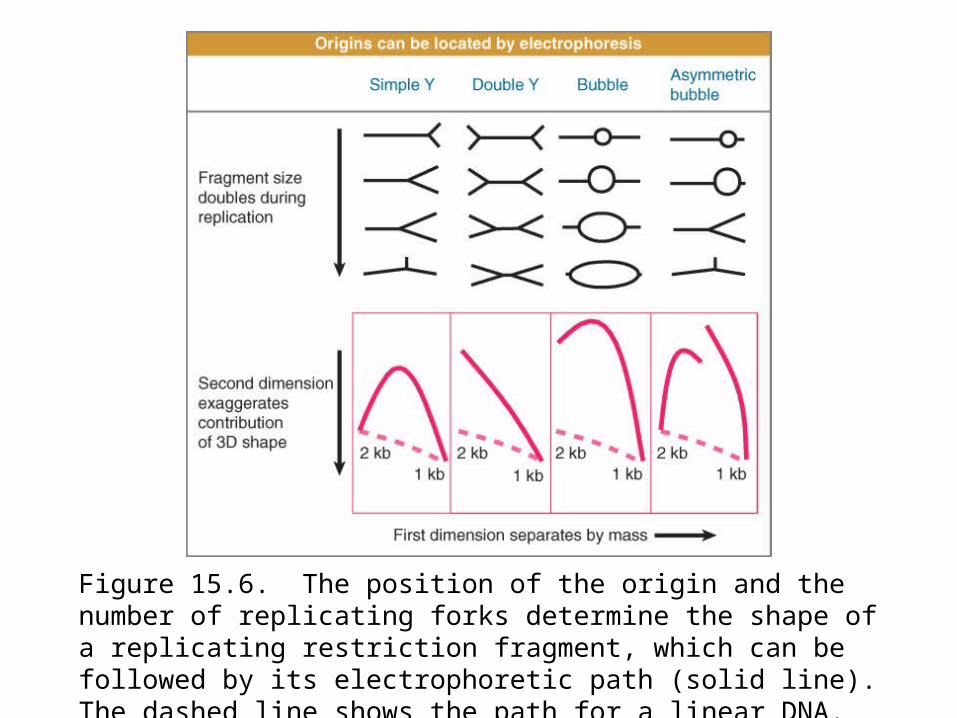
Figure 15.6. The position of the origin and the number of replicating forks determine the shape of a replicating restriction fragment, which can be followed by its electrophoretic path (solid line). The dashed line shows the path for a linear DNA.

15.4 Does Methylation at the Origin Regulate Initiation?
oriC contains eleven GATC/CTAG repeats that are methylated on adenine on both strands.
Replication generates hemimethylated DNA, which cannot initiate replication.
There is a 13-minute delay before the GATC/CTAG repeats are remethylated.
Key Concepts

What feature of a bacterial (or plasmid) origin ensures that it is used to initiate replication only once per cycle?
Some sequences that are used for this purpose are included in the origin. oriC contains eleven copies of the sequence GATC/CTAG, which is a target for methylation at the N6 position of adenine by the Dam methylase (Figure 15.7).
If the plasmid is methylated it undergoes a single round of replication, and then the hemimethylated products accumulate (Figure 15.8). Hemimethylated origins cannot initiate again until the Dam methylase has converted them into fully methylated origins.
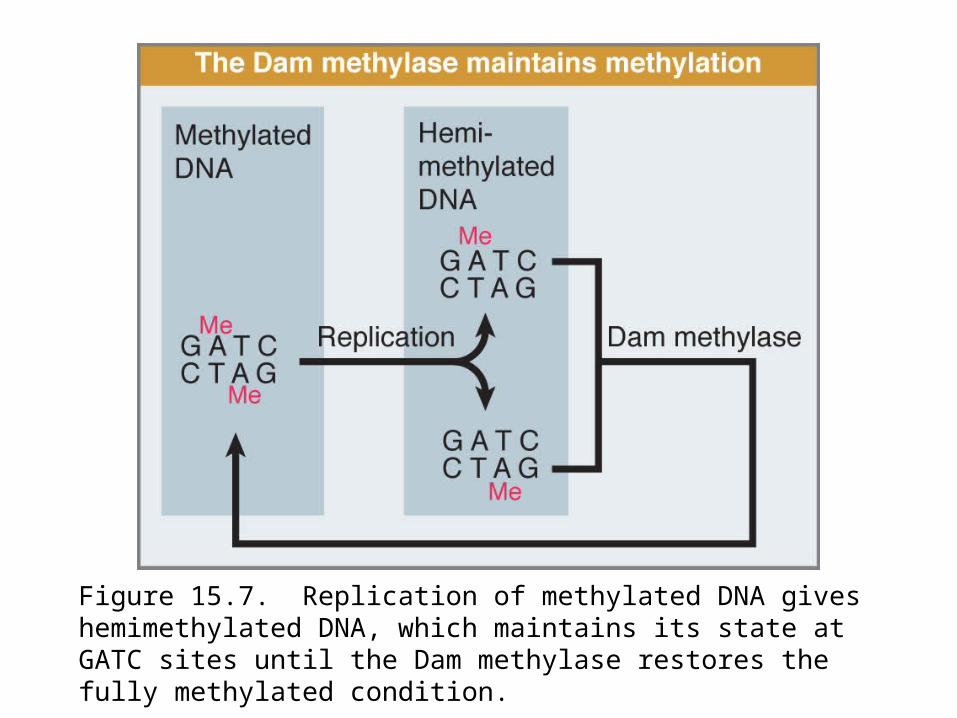
Figure 15.7. Replication of methylated DNA gives hemimethylated DNA, which maintains its state at GATC sites until the Dam methylase restores the fully methylated condition.
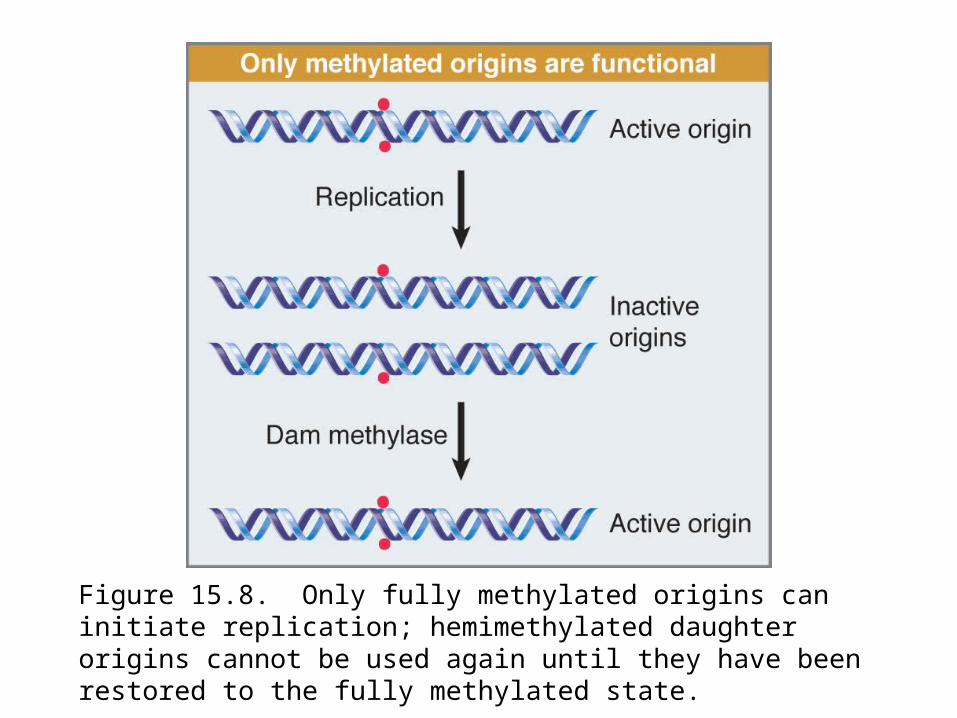
Figure 15.8. Only fully methylated origins can initiate replication; hemimethylated daughter origins cannot be used again until they have been restored to the fully methylated state.
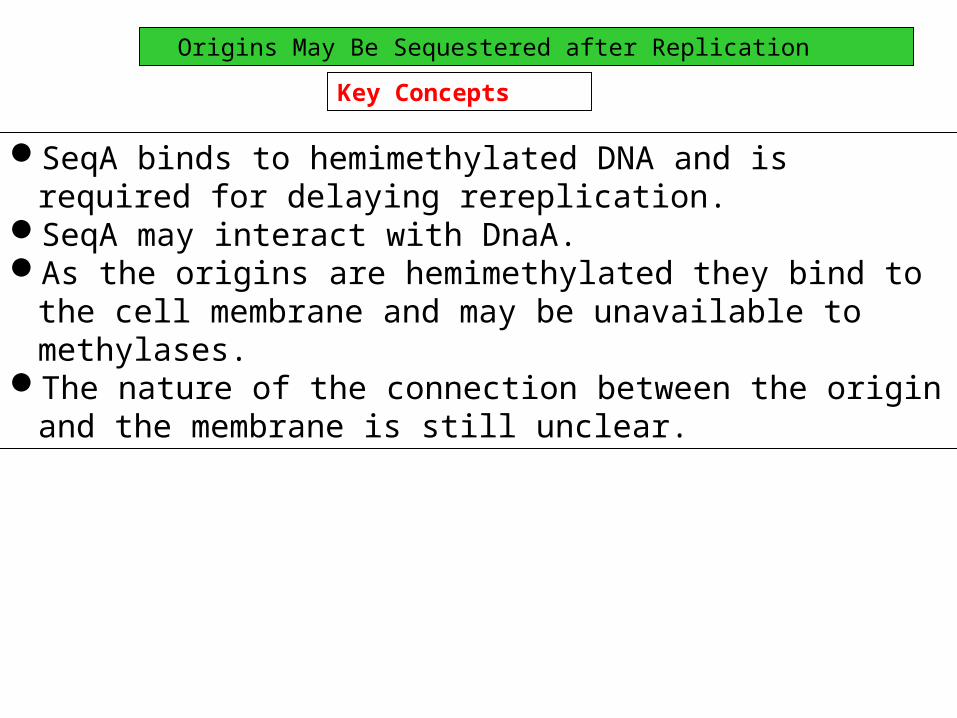
Origins May Be Sequestered after Replication
SeqA binds to hemimethylated DNA and is required for delaying rereplication.
SeqA may interact with DnaA.As the origins are hemimethylated they bind to the cell membrane
and may be unavailable to methylases.The nature of the connection between the origin and the membrane
is still unclear.
Key Concepts
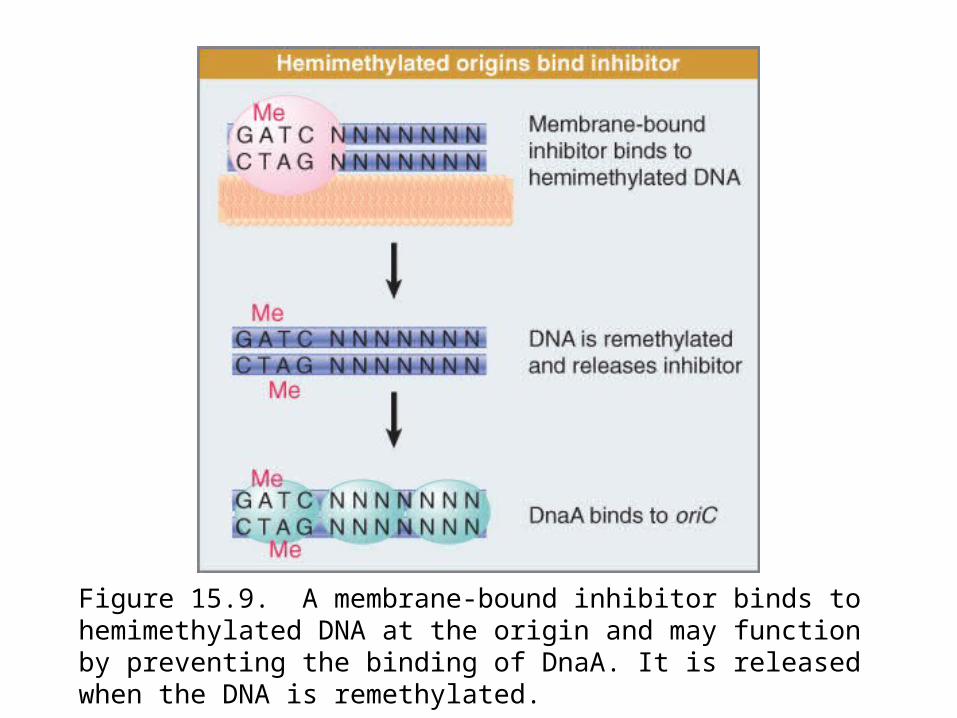
Figure 15.9. A membrane-bound inhibitor binds to hemimethylated DNA at the origin and may function by preventing the binding of DnaA. It is released when the DNA is remethylated.

Each Eukaryotic Chromosome Contains Many Replicons
Eukaryotic replicons are 40 to 100 kb in length.A chromosome is divided into many replicons.Individual replicons are activated at characteristic times during S
phase.Regional activation patterns suggest that replicons near one another
are activated at the same time.
Key Concepts

S phase usually lasts a few hours in a higher eukaryotic cell.
Individual replicons in eukaryotic genomes are relatively small, typically ~40 kb in yeast or fly and ~ 100 kb in animal cells. The rate of replication is ~ 2000 bp/min, which is much slower than the 50,000 bp/min of bacterial replication fork movement.
A mammalian genome could be replicated in ~1 hour if all replicons functioned simultaneously. S phase actually lasts for >6 hours in a typical somatic cell, implying that no more than 15% of the replicons are likely to be active at any given moment.
Visualization of replicating forks by labeling with DNA precursors identifies 100 to 300 “foci” instead of uniform staining; each focus shown in Figure 15.11 probably contains >300 replication forks.
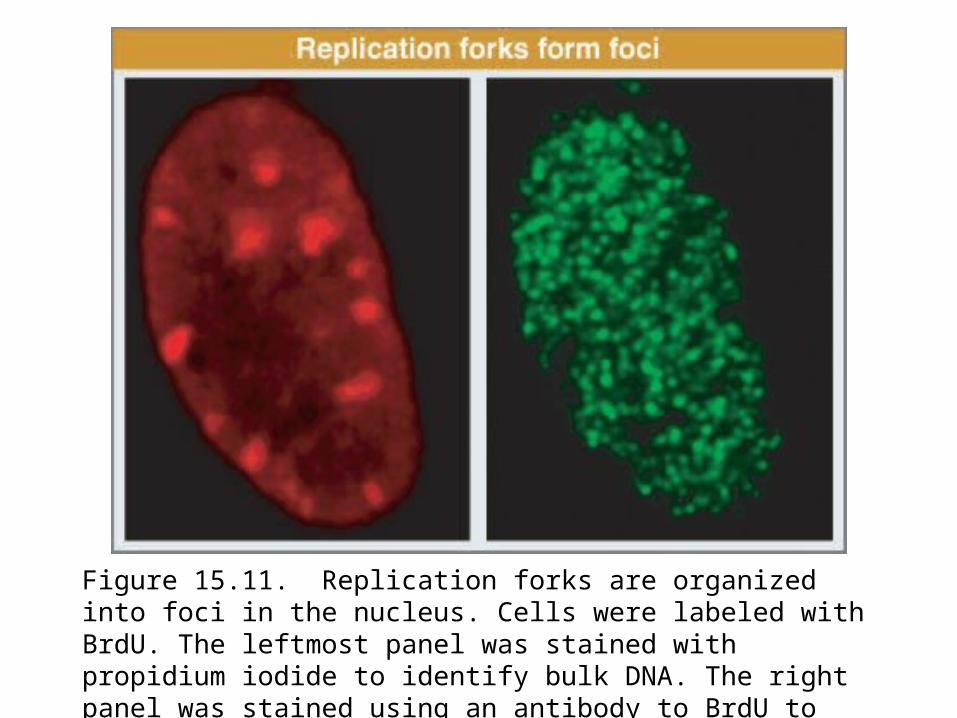
Figure 15.11. Replication forks are organized into foci in the nucleus. Cells were labeled with BrdU. The leftmost panel was stained with propidium iodide to identify bulk DNA. The right panel was stained using an antibody to BrdU to identify replicating DNA.

15.7 Replication Origins Can Be Isolated in Yeast
Origins in S. cerevisiae are short A-T-rich sequences that have an essential 11-bp sequence.
The ORC is a complex of six proteins that binds to an ARS.
Key Concepts

Any segment of DNA that has an origin should be able to replicate, so although plasmids are rare in eukaryotes, it may be possible to construct them by suitable manipulation in vivo. This has been accomplished in yeast, although not in higher eukaryotes.
The discovery of yeast origins resulted from the observation that some yeast DNA fragments (when circularized) are able to transform defective cells very efficiently. These fragments can survive in the cell in the unintegrated (autonomous) state, that is, as self-replicating plasmids.
This segment is called as ARS (for autonomously replicating sequence). ARS elements are derived from origins of replication.
An ARS element consists of an A-T-rich region. Figure 15.12: shows a systematic mutational analysis along the
length of an origin.

Origin function is abolished completely by mutations in a 14-bp “core” region, called the A domain, which contains an 11-bp consensus sequence consisting of A-T base pairs.
This consensus sequence (called ACS for ARS Consensus Sequence) is the only homology between known ARS elements.
Mutations in three adjacent elements, numbered B1 to B3, reduce origin function. An origin can function effectively with any two of the B elements, so long as a functional A element is present.
The ORC (origin recognition complex) is a complex of six proteins with a mass of ~400 kD. ORC binds to the A and B1 elements.
There are about 400 origins in the yeast genome, meaning that the average length of a replicon is ~ 35,000 bp.
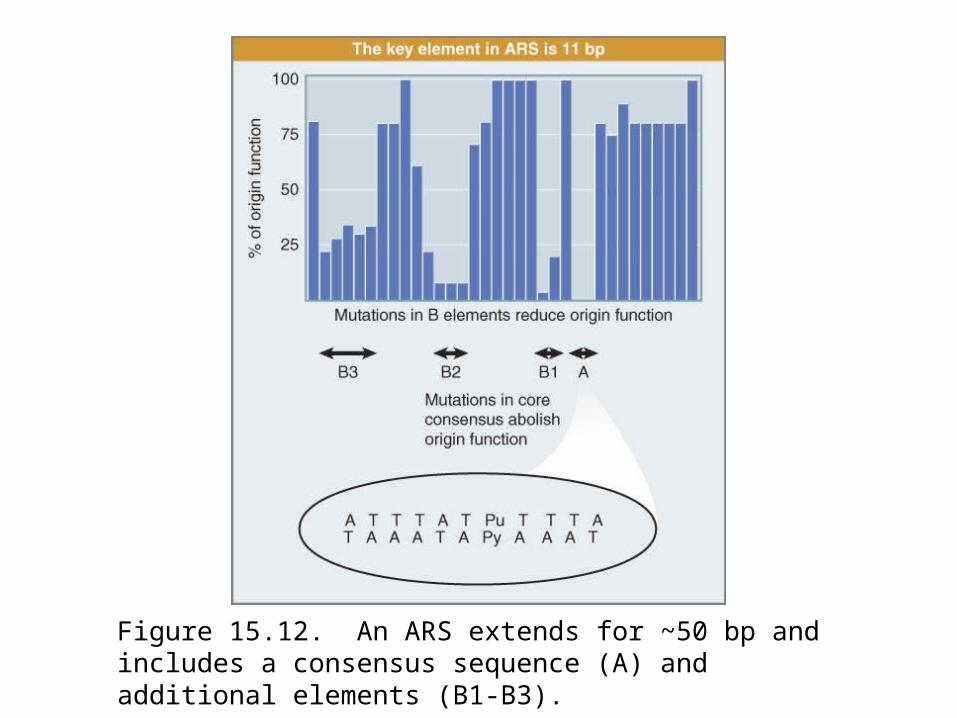
Figure 15.12. An ARS extends for ~50 bp and includes a consensus sequence (A) and additional elements (B1-B3).

Licensing Factor Controls Eukaryotic Replication
Licensing factor is necessary for initiation of replication at each origin.
It is present in the nucleus prior to replication, but is inactivated or destroyed by replication.
Initiation of another replication cycle becomes possible only after licensing factor reenters the nucleus after mitosis.
Key Concepts

A eukaryotic genome is divided into multiple replicons, and the origin in each replicon is activated once and only once in a single division cycle.
Figure 15.13: >1 replication cycle needs cytoplasmic factors.Figure 15.14: explains the control of reinitiation by proposing that
this protein is a licensing factor.It is present in the nucleus prior to replication. One round of
replication either inactivates or destroys the factor, and another round cannot occur until further factor is provided. Factor in the cytoplasm can gain access to the nuclear material only at the subsequent mitosis when the nuclear envelope breaks down.
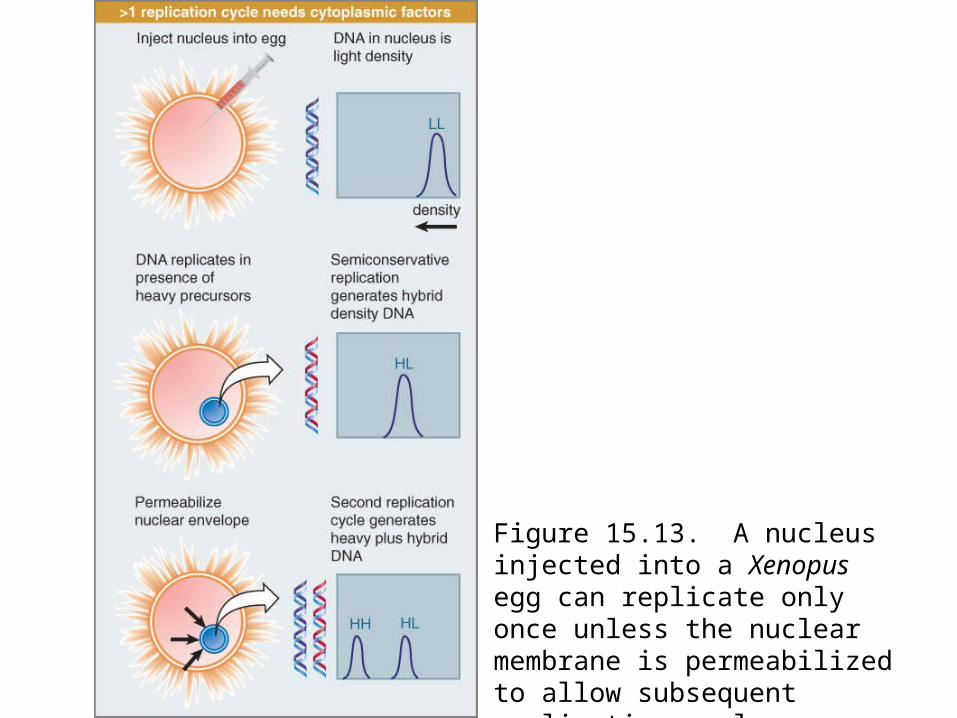
Figure 15.13. A nucleus injected into a Xenopus egg can replicate only once unless the nuclear membrane is permeabilized to allow subsequent replication cycles.
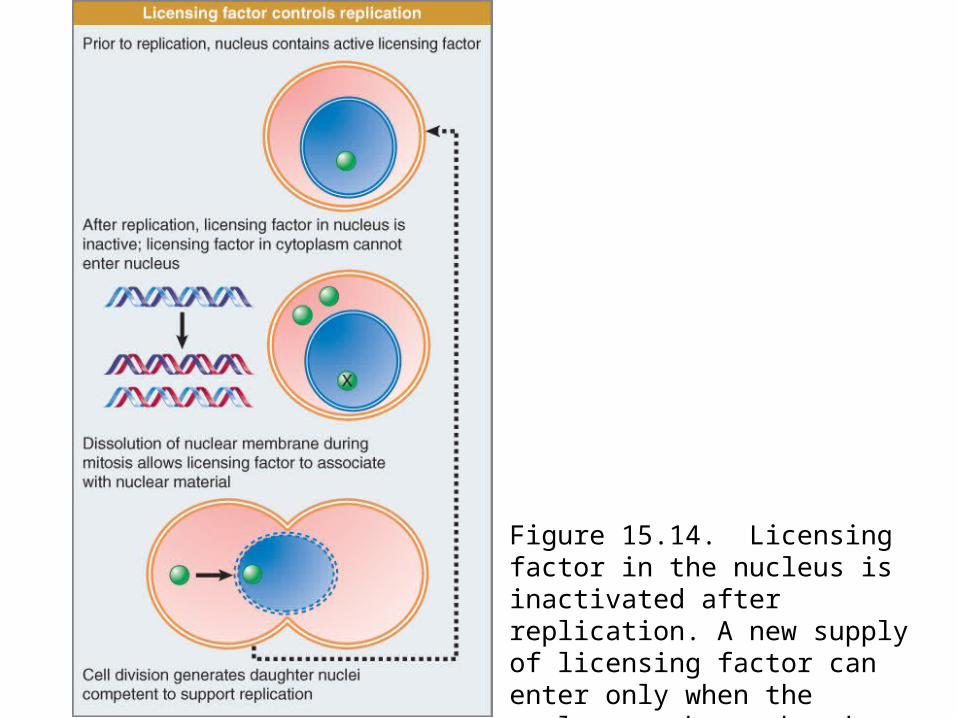
Figure 15.14. Licensing factor in the nucleus is inactivated after replication. A new supply of licensing factor can enter only when the nuclear membrane breaks down at mitosis.

Licensing Factor Consists of MCM Proteins
The ORC is a protein complex that is associated with yeast origins throughout the cell cycle.
Cdc6 protein is an unstable protein that is synthesized only in G1.
Cdc6 binds to ORC and allows MCM proteins to bind.When replication is initiated, Cdc6 and MCM proteins are
displaced. The degradation of Cdc6 prevents reinitiation.Some MCM proteins are in the nucleus throughout the
cycle, but others may enter only after mitosis.
Key Concepts

The key event in controlling replication is the behavior of the ORC complex at the origin. The origin (ARS) consists of the A consensus sequence and three B elements. The ORC complex of six proteins binds to the A and adjacent B1 element. The transcription factor ABF1 binds to the B3 element; this assists initiation.
Most origins are localized in regions between genes.Figure 15.15; summarizes the cycle of events at the origin.In yeast, Cdc6 is a highly unstable protein, with a half-life of <5
minutes. It is synthesized during G1 and typically binds to the ORC between the exit from mitosis and late G1.
In yeast the presence of Cdc6 at the origin allows MCM (mini-chromosome maintenance) proteins to bind to the complex. The origin therefore enters S phase in the condition of a prereplication complex, which contains ORC, Cdc6, and MCM proteins. When initiation occurs, Cdc6 and MCM are displaced, returning the origin to the state of the postreplication complex, which contains only ORC.
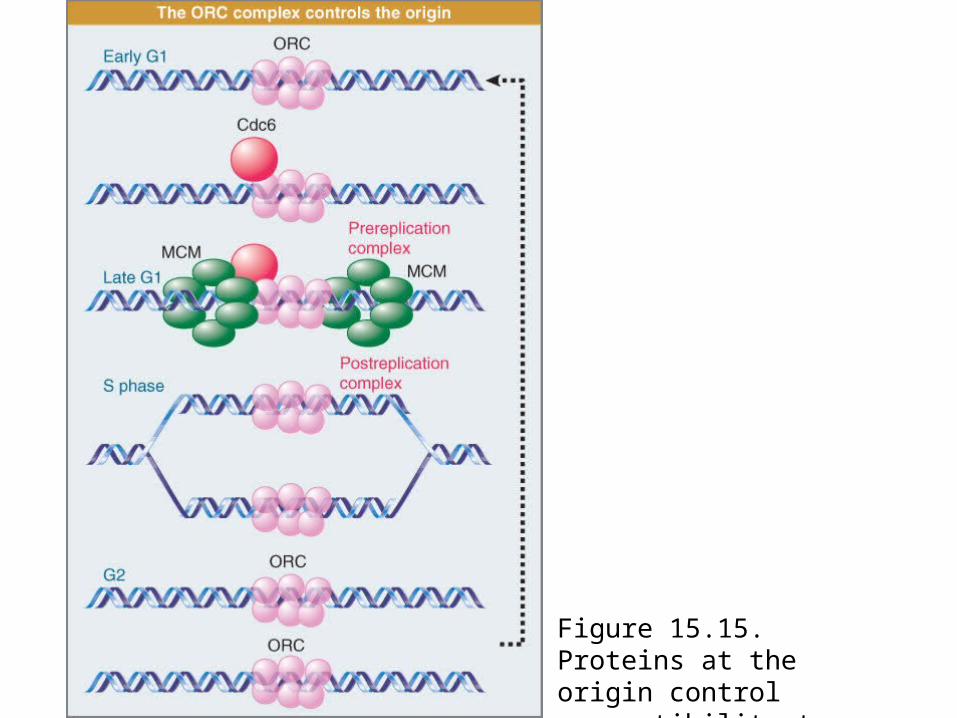
Figure 15.15. Proteins at the origin control susceptibility to initiation.

D Loops Maintain Mitochondrial Origins
Mitochondria use different origin sequences to initiate replication of each DNA strand.
Replication of the H strand is initiated in a D loop.Replication of the L strand is initiated when its origin is
exposed by the movement of the first replication fork.
Key Concepts

Initiation requires separating the DNA strands and commencing bidirectional DNA synthesis. A different type of arrangement is found in mitochondria.
Replication starts at a specific origin in the circular duplex DNA. Initially, though, only one of the two parental strands (the H strand in mammalian mitochondrial DNA) is used as a template for synthesis of a new strand. Synthesis proceeds for only a short distance, displacing the original partner (L) strand, which remains single-stranded, as illustrated in Figure 15.16. The condition of this region gives rise to its name as the displacement loop, or D loop.

Figure 15.16. The D loop maintains an opening in mammalian mitochondrial DNA, which has separate origins for the replication of each strand.

The existence of D loops exposes a general principle: An origin can be a sequence of DNA that serves to initiate DNA synthesis using one strand as template.
The opening of the duplex does not necessarily lead to the initiation of replication on the other strand. In the case of mitochondrial DNA replication, the origins for replicating the complementary strands lie at different locations.
Origins that sponsor replication of only one strand are also found in the rolling circle mode of replication.
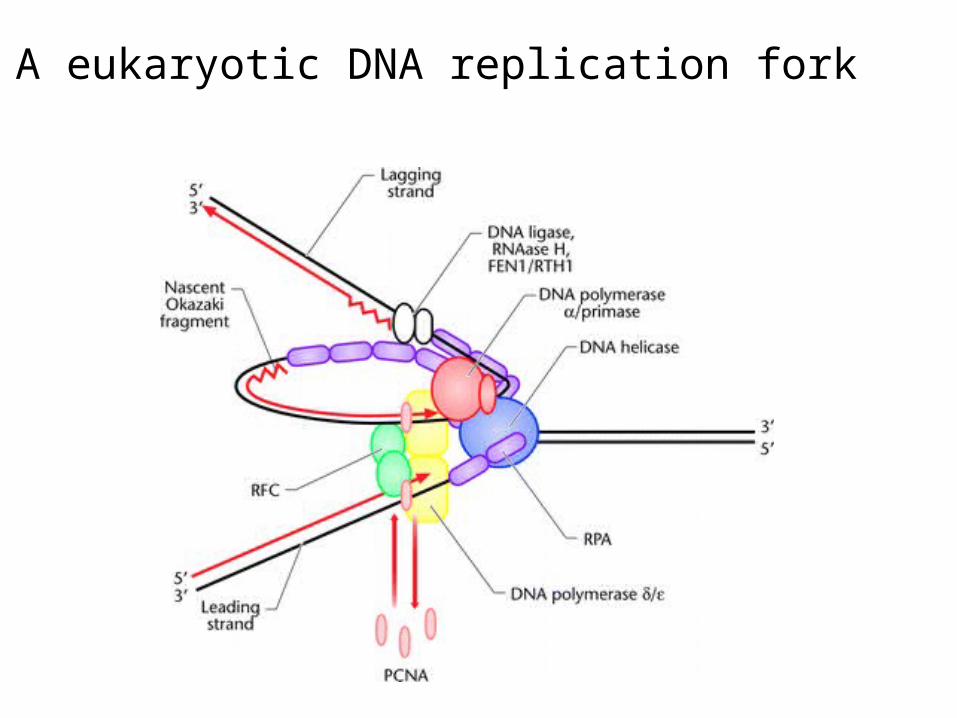
A eukaryotic DNA replication fork

Teleomeres
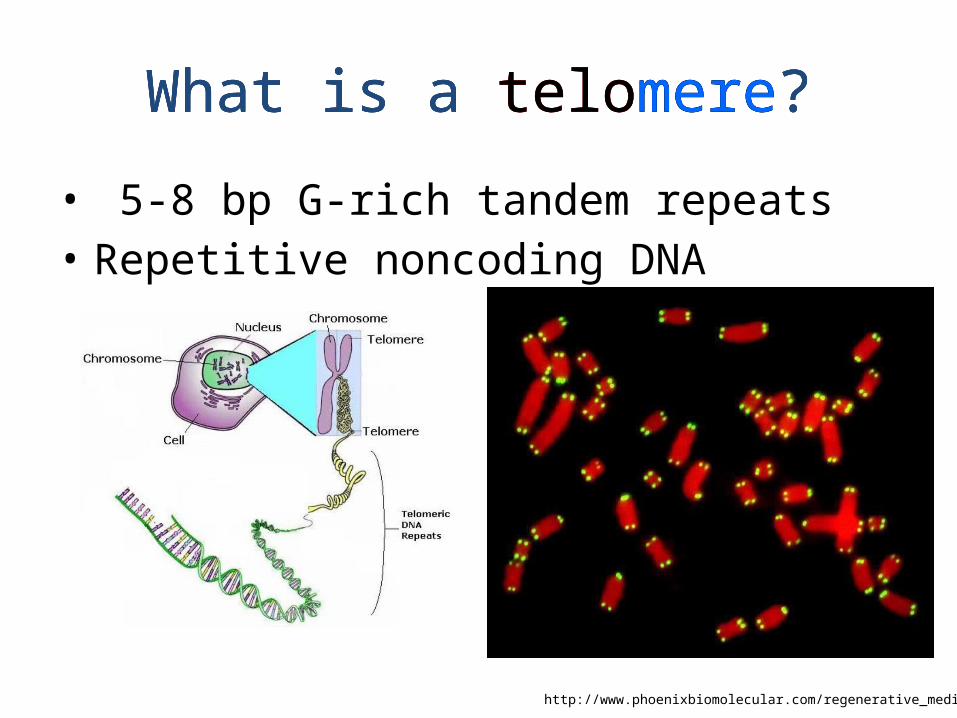
What is a telomere?
• 5-8 bp G-rich tandem repeats• Repetitive noncoding DNA
http://www.phoenixbiomolecular.com/regenerative_medicine.html
What is a telomere?What is a telomere?
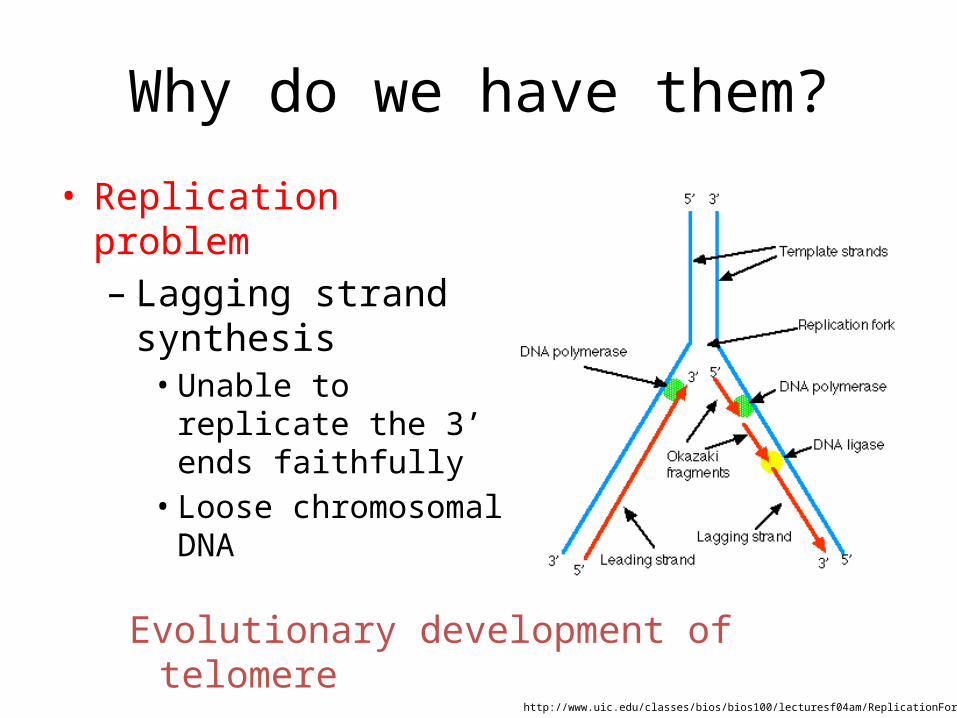
Why do we have them?
• Replication problem– Lagging strand synthesis
• Unable to replicate the 3’ ends faithfully
• Loose chromosomal DNA
http://www.uic.edu/classes/bios/bios100/lecturesf04am/ReplicationFork.gif
Evolutionary development of telomere

Telomeres
Ends of linear chromosomes
Centromere
TelomereTelomere
Repetitive DNA sequence(TTAGGG in vertebrates)
Specialized proteins
Form a 'capped' end structure
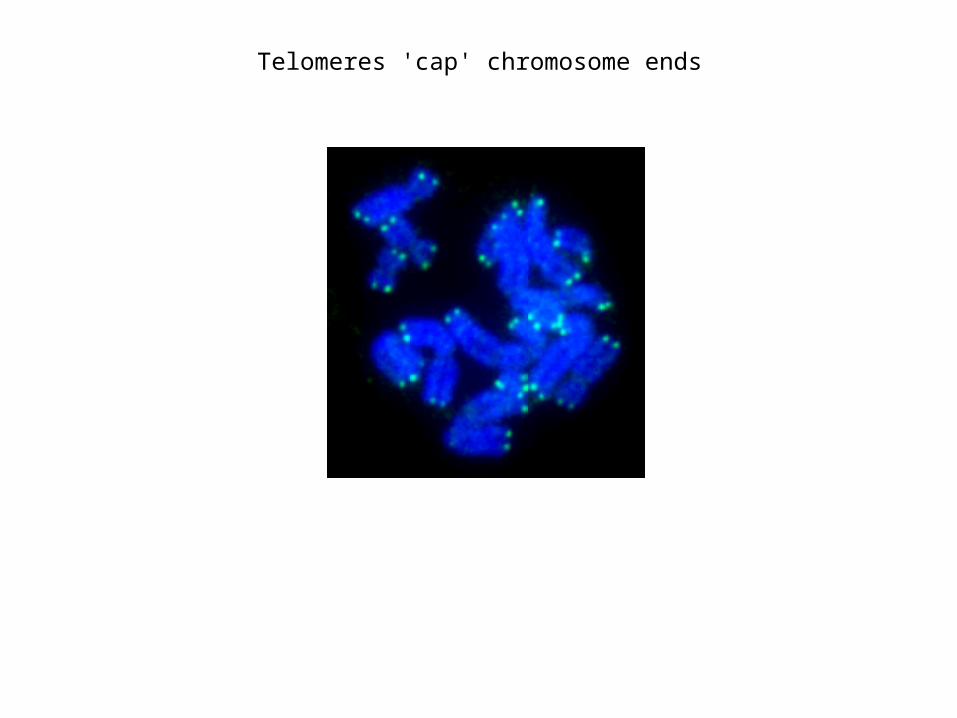
Telomeres 'cap' chromosome ends
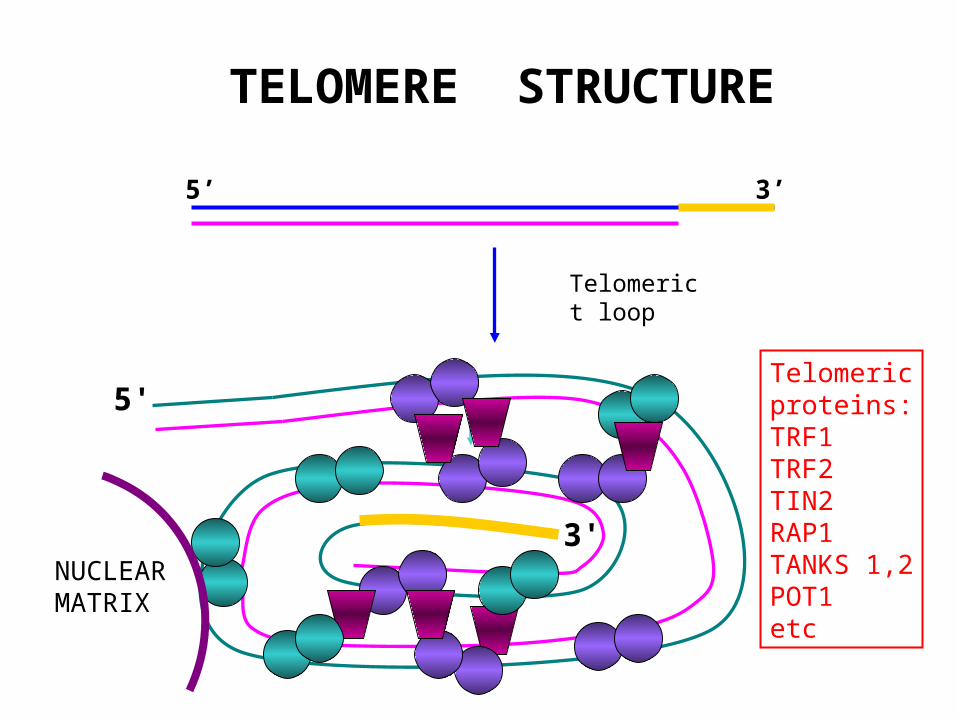
TELOMERE STRUCTURE
5’ 3’
5'
3'
Telomerict loop
Telomericproteins:TRF1TRF2TIN2RAP1TANKS 1,2POT1etc
NUCLEARMATRIX
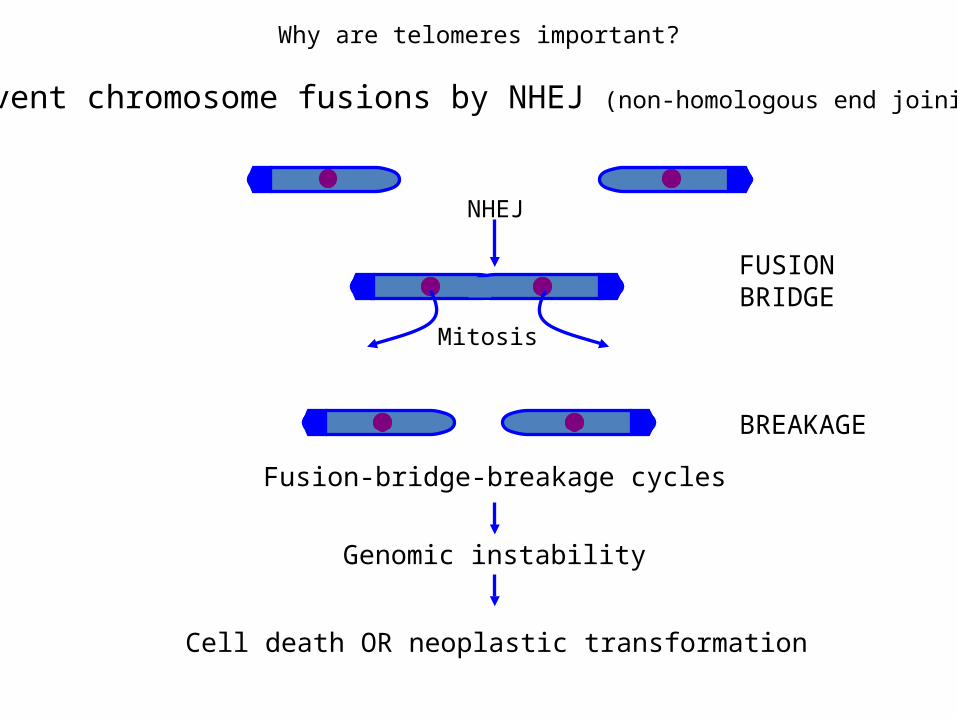
Why are telomeres important?
Prevent chromosome fusions by NHEJ (non-homologous end joining)
NHEJ
Mitosis
FUSIONBRIDGE
BREAKAGE
Fusion-bridge-breakage cycles
Genomic instability
Cell death OR neoplastic transformation
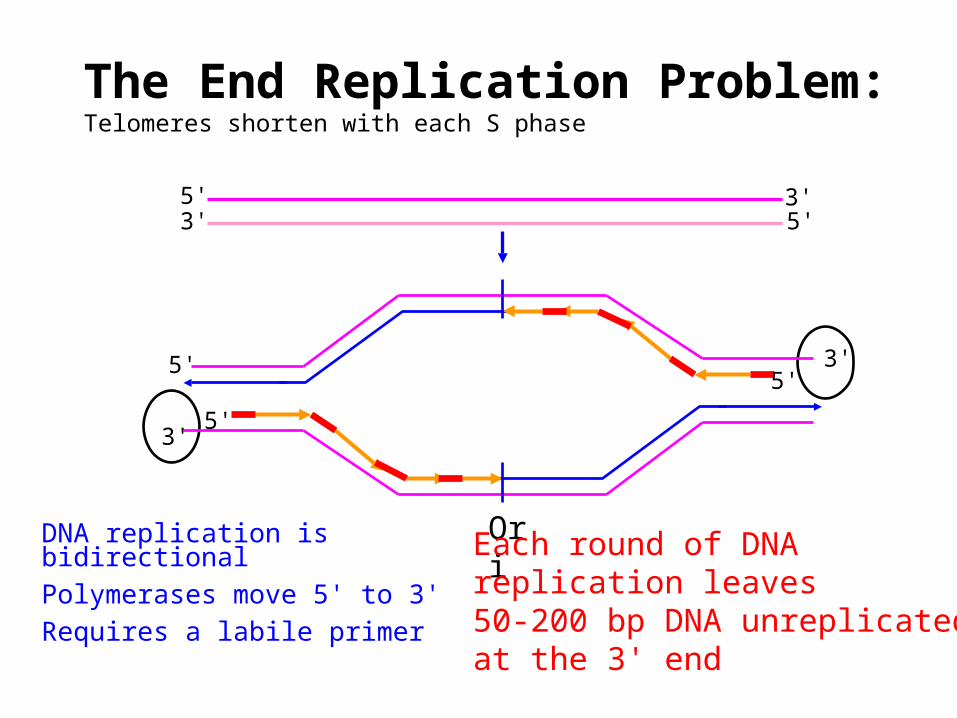
The End Replication Problem:Telomeres shorten with each S phase
OriDNA replication is bidirectionalPolymerases move 5' to 3'Requires a labile primer
3'5'
3'5'
5'
5' 3'3' 5'
Each round of DNAreplication leaves50-200 bp DNA unreplicatedat the 3' end
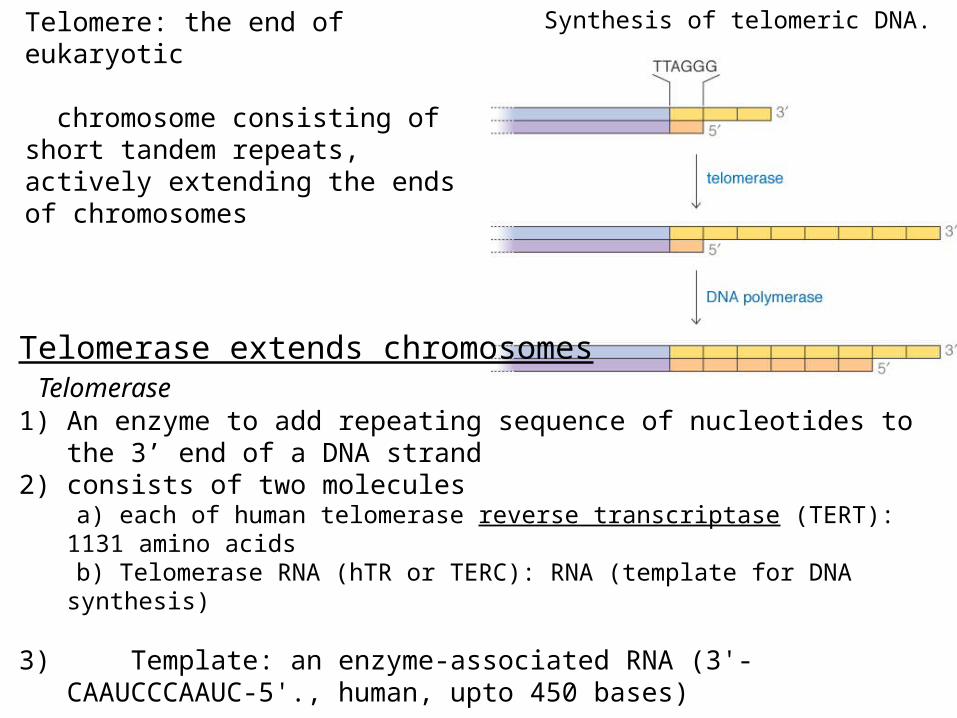
Synthesis of telomeric DNA.
Telomerase extends chromosomes Telomerase1) An enzyme to add repeating sequence of nucleotides to the 3’ end of a DNA
strand2) consists of two molecules a) each of human telomerase reverse transcriptase (TERT): 1131 amino acids b) Telomerase RNA (hTR or TERC): RNA (template for DNA synthesis)
3) Template: an enzyme-associated RNA (3'-CAAUCCCAAUC-5'., human, upto 450 bases)
4) Telomere: Repeated sequence of TTAGGG upto 5-15 kb
Telomere: the end of eukaryotic chromosome consisting of short tandem repeats, actively extending the ends of chromosomes
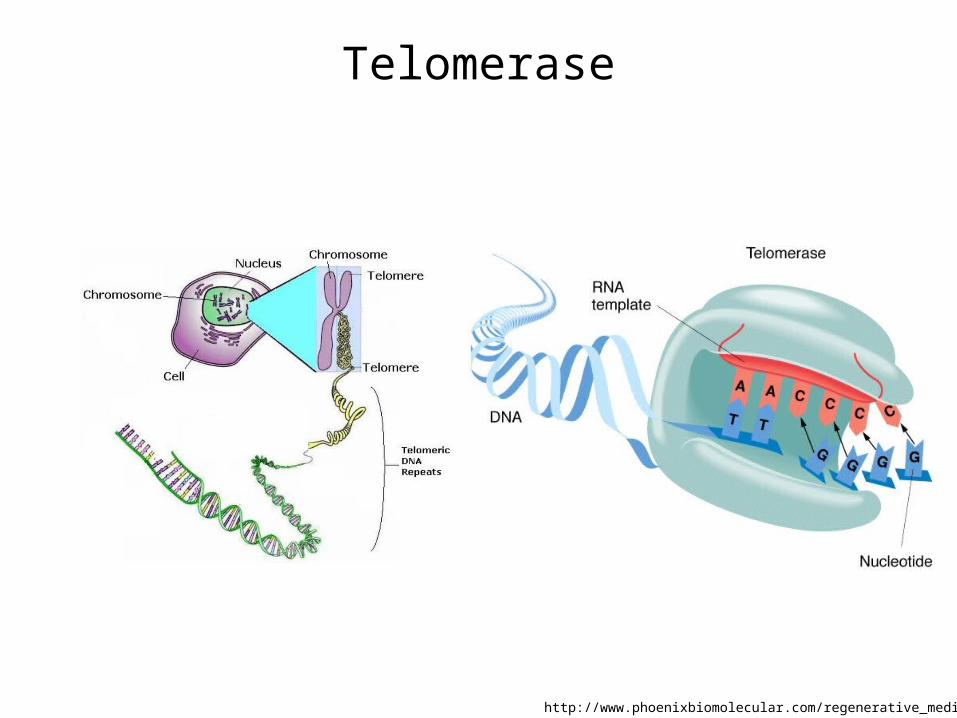
Telomerase
http://www.phoenixbiomolecular.com/regenerative_medicine.html
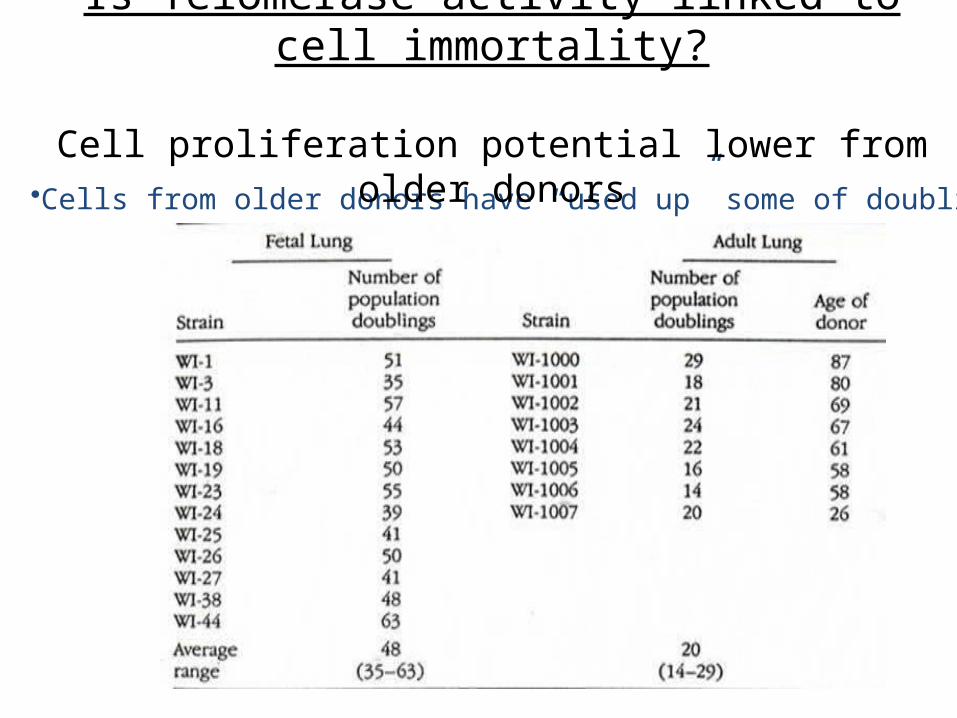
•Cells from older donors have “used up” some of doublings
Is Telomerase activity linked to cell immortality?
Cell proliferation potential lower from older donors

Organism + Life Span: Hayflick Limit:-mouse about 3 years -doublings about 20-human about 100 -doublings about 40-60-Galapagos tortoise about 150 -doublings about 140
Cell proliferation potential greater in long-lived species
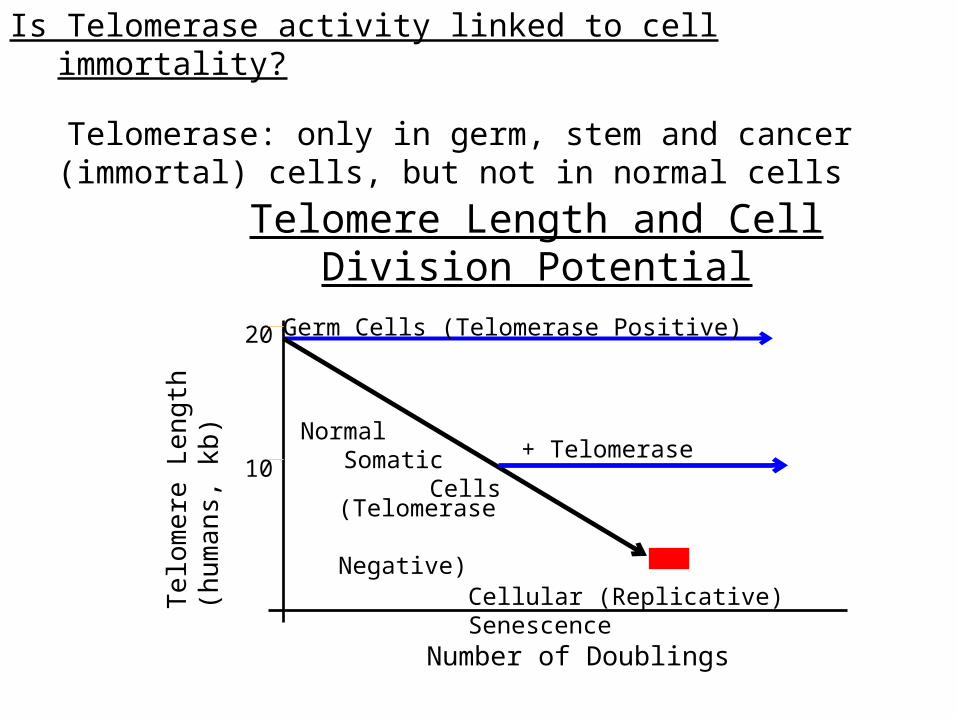
Is Telomerase activity linked to cell immortality?
Telomerase: only in germ, stem and cancer (immortal) cells, but not in normal cells
Telomere Length
(humans, kb)
Number of Doublings
20
10
Cellular (Replicative) Senescence
Normal Somatic Cells
(Telomerase Negative)
Germ Cells (Telomerase Positive)
+ Telomerase
Telomere Length and Cell Division Potential

Cellular senescence• Once the telomere shrinks to a certain extent,
the cell stops dividing. – ~4kb in human cells triggers end to cell division.
• This leads to other changes called cellular senescence:– Cell morphology changes.– Gene expression changes.

• Telomerase mutants have a short lifespan: when telomeres shorten to a critical point, yeast cells stop dividing.
• Overexpression of telomerase:• Longer telomeres.• Increased replicative lifespan.
Yeast replicative lifespan regulated by telomere length
• Lab strains of mice have very long telomeres.• 30-40kb telomeres.
• Tert knock-out mice:• Normal for four generations as their telomeres shorten,• Premature aging phenotypes present in the 5th generation
Telomeres in mice

Telomerase:
Biomedical uses
Expand cells for replacement therapies(burns, joint replacements, etc)
Telomerase inhibitors to selectively kill cancer cells

The telomere hypothesis of aging
Telomeres shorten with each cell division and therefore with ageTRUE
Short telomeres cause cell senescence andsenescent cells may contribute to agingTRUE
HYPOTHESIS:Telomere shortening causes aging andtelomerase will prevent agingTRUE OR FALSE?

The telomere hypothesis of aging
Telomere length is not related to life span(mice vs human; M musculus vs M spretus)
Telomeres contribute to aging ONLY if senescent cells contribute to aging
Telomerase protects against replicativesenescence but not senescence induce byother causes

Telomeres and Cellular Aging
• Cells removed from a newborn infant and placed in culture will go on to divide almost 100 times. Well before the end, however, their rate of mitosis declines (to less than once every two weeks).
• Cells from an 80 year old would manage only a couple of dozen mitoses before they ceased dividing and died out.
• Telomeres are important so their steady shrinking with each mitosis might impose a finite life span on cells.
• This phenomenon is called replicative senescence • Could shrinkage of telomeres be a clock that determines the
longevity of a cell lineage and thus is responsible for replicative senescence?





























