Embed Size (px)
Citation preview

Multiple sclerosis (MS) is an inflammatory de-myelinating disease of the central nervous sys-tem (CNS) that takes a relapsing–remitting or aprogressive course (reviewed in Refs 1,2). Itscounterpart in the peripheral nervous system(PNS) is chronic inflammatory demyelinatingpolyradiculoneuropathy (CIDP) (reviewed in Ref.3). In addition, there are acute, monophasic dis-orders, such as the inflammatory demyelinatingpolyradiculoneuropathy termed Guillain–Barrésyndrome (GBS) in the PNS, and acute dissemi-nated encephalomyelitis (ADEM) in the CNS.Both MS and GBS are heterogeneous syn-dromes. In MS different exogenous assaults to-gether with genetic factors can result in a diseasecourse that finally fulfils the diagnostic criteria. Inboth diseases, axonal damage can add to a pri-marily demyelinating lesion and cause permanentneurological deficits. No single animal model ex-ists that mimics all the features of human de-myelinating diseases; rather, the available mod-els reflect specific facets. Here, we focus onexperimental autoimmune encephalomyelitis(EAE) and neuritis (EAN) as models in rat andmouse strains, and discuss their distincthistopathology and the roles played by differentautoantigens.
Experimental autoimmuneencephalomyelitis
Experimental aspects of EAE EAE can be reliably elicited in a number of dif-ferent species by immunization with either CNStissue or with purified components of CNSmyelin, for instance myelin basic protein (MBP)and proteolipid protein (PLP). This type of modelis called ‘actively-induced EAE’. Sensitized T-celllines, propagated in vitro and then injected intra-
venously, generate ‘adoptive-transfer EAE’ (AT-EAE), giving conclusive proof of the pivotal path-ogenic role of T cells. In contrast to human trials,which require inclusion of hundreds of patients todemonstrate statistically significant effects, ex-perimental therapies can be evaluated in the in-bred rodent models with only 6–12 animals pertreatment group, depending on disease variabil-ity. This collectively depends on the antigen(s)used, the strain of rodent, the mode of immun-ization and the respective treatment. In the ATmodels, fewer animals are required because thedisease runs a more synchronous course.Maximum disease expression of EAE in theLewis rat is normally reached two to four daysafter onset, and severely affected rats can evendie as a result of widespread inflammation in thebrainstem. Recovery from the monophasic dis-ease is then associated with enhanced apoptosisof inflammatory T cells in the lesion, which couldalso have therapeutic relevance (reviewed inRef. 4). Only with immunopharmacological mod-ulation, for example by low-dose cyclosporin A,can relapses of active EAE in Lewis rats beachieved.
Lessons from encephalitogenic T-cell lines The generation of antigen-specific, autoaggres-sive T-cell lines (i.e. T-cell lines that attack thehost) was a major step towards better under-standing immunological paradigms of EAE in ro-dents (reviewed in Refs 2,3). Recognition pat-terns of major epitopes of myelin proteins thatgive rise to encephalitis (encephalitogenic pro-teins) were characterized in autoantigen-specific,CD41 T cells in rats and mice and then adaptedto study human T-cell lines. In particular, the find-ing that these T cells used a limited range of dis-
ease-associated T-cell receptor (TCR) variableregions in different rodent strains raised hopes ofdeveloping TCR-specific immunotherapies. Thisconcept has now been abandoned by most re-search groups because TCR usage turned out tobe more heterogenous than originally suspected5.Furthermore, these T-cell lines and clones servedas tools to characterize the secretion pattern ofthe proinflammatory mediators, cytokines andchemokines that orchestrate the autoimmune re-action in nervous tissue.
Newer EAE models and histopathologyThe value of the Lewis rat EAE model for the elu-cidation of the pathogenesis of MS is limited by thelack of spontaneous relapses, and also by the ab-sence of primarily demyelinating lesions. In con-trast to MS, the histopathological features of EAEthat is induced by immunization with MBP or bytransfer of specific T cells are dominated by in-flammation and by axonal damage in the spinalcord. A significant amount of demyelination is onlyseen when co-transfer of antibodies specific formyelin-oligodendrocyte glycoprotein (MOG), to-gether with encephalitogenic T cells, is performed,reflecting the contribution of autoantibody andcomplement deposits to demyelination in MS (Ref.6). Despite these limitations, the Lewis rat has stillhelped us to define important disease patterns inwhich a variety of putative autoantigens apparentlydetermine the topography of inflammatory lesionsin the CNS. It came as a great surprise that evenproteins expressed in glial cells, such as S-100,which are not CNS-specific but are also found inthymus and peripheral nerve, can serve as auto-antigens7. S-100 EAE results in widespread T-cellmediated inflammation with pathology in the opticnerve, a prominent feature of early MS1,2.
Disease models MOLECULAR MEDICINE TODAY, FEBRUARY 2000 (VOL. 6)
88 1357-4310/00/$ - see front matter © 2000 Elsevier Science Ltd. All rights reserved. PII: S1357-4310(99)01639-1
Animal models forautoimmune demyelinatingdisorders of the nervoussystem
Ralf Gold, Hans-Peter Hartung and Klaus V. Toyka
There is increasing evidence to suggest that itis not the major myelin proteins like MBP andPLP, but rather the minor constituent MOG, thatplays a key role in CNS autoimmunity. The wholespectrum of MS pathology was closely reflectedin MOG-induced EAE in susceptible rat strains.Even clinical and histopathological subforms ofMS, such as neuromyelitis optica (Devic’s dis-ease), could reproducibly be induced in thismodel6. These findings have already been ex-tended to the analysis of concomitant axonaldamage, which has recently attracted much in-terest in MS. Finally, the role of genetic and envi-ronmental factors that contribute to susceptibility
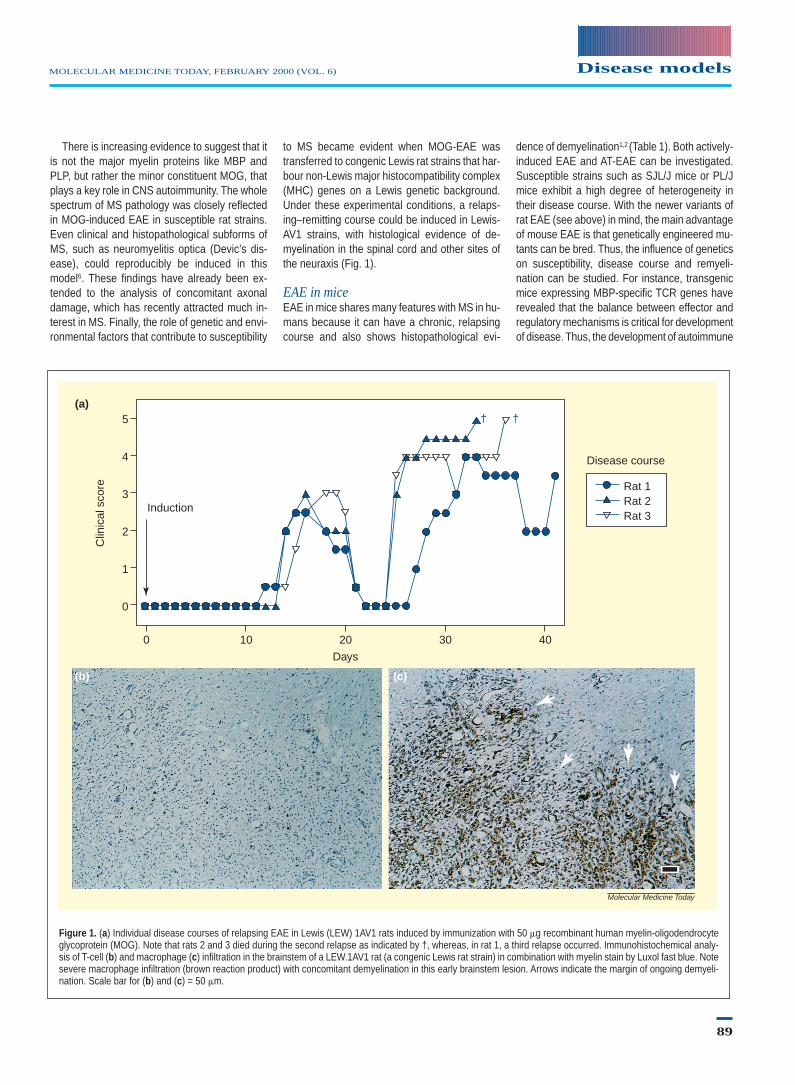
to MS became evident when MOG-EAE wastransferred to congenic Lewis rat strains that har-bour non-Lewis major histocompatibility complex(MHC) genes on a Lewis genetic background.Under these experimental conditions, a relaps-ing–remitting course could be induced in Lewis-AV1 strains, with histological evidence of de-myelination in the spinal cord and other sites ofthe neuraxis (Fig. 1).
EAE in miceEAE in mice shares many features with MS in hu-mans because it can have a chronic, relapsingcourse and also shows histopathological evi-
dence of demyelination1,2 (Table 1). Both actively-induced EAE and AT-EAE can be investigated.Susceptible strains such as SJL/J mice or PL/Jmice exhibit a high degree of heterogeneity intheir disease course. With the newer variants ofrat EAE (see above) in mind, the main advantageof mouse EAE is that genetically engineered mu-tants can be bred. Thus, the influence of geneticson susceptibility, disease course and remyeli-nation can be studied. For instance, transgenicmice expressing MBP-specific TCR genes haverevealed that the balance between effector andregulatory mechanisms is critical for developmentof disease. Thus, the development of autoimmune
Disease modelsMOLECULAR MEDICINE TODAY, FEBRUARY 2000 (VOL. 6)
89
Figure 1. (a) Individual disease courses of relapsing EAE in Lewis (LEW) 1AV1 rats induced by immunization with 50 mg recombinant human myelin-oligodendrocyteglycoprotein (MOG). Note that rats 2 and 3 died during the second relapse as indicated by †, whereas, in rat 1, a third relapse occurred. Immunohistochemical analy-sis of T-cell (b) and macrophage (c) infiltration in the brainstem of a LEW.1AV1 rat (a congenic Lewis rat strain) in combination with myelin stain by Luxol fast blue. Notesevere macrophage infiltration (brown reaction product) with concomitant demyelination in this early brainstem lesion. Arrows indicate the margin of ongoing demyeli-nation. Scale bar for (b) and (c) = 50 mm.
Molecular Medicine Today
5
4
3
2
1
0
Clin
ical
sco
re
(a)
(b) (c)
0 10 20 30 40
Days
Induction
† †
Disease course
Rat 1Rat 2Rat 3

disease depends not only on the existence of self-reactive T cells, but also on the presence of the ap-propriate number of functional regulatory T cells8.Furthermore, in some cases, such as Bcl-2 knock-out mice, the genetic defect can interfere with theability to mount a significant T-cell response afteractive induction and require the use of adoptive-transfer models to generate pathology.
EAE in primatesThe identification of mechanisms that cause auto-immune disorders of the CNS is essential for thesuccessful application of novel immunotherapies.Because of the species barrier, non-human pri-mate models were developed that more closelyreflect human MS. The general fundamental prin-ciples obtained in rodent EAE have been con-firmed and thus underscore the importance of
EAE for the evaluation of novel immunotherapies(reviewed in Ref. 9). Recently, detailedhistopathological analyses have also shown strik-ing parallels between autoantibody deposition inprimate EAE and in human MS lesions.
Models of virus-induced demyelinationTheiler’s murine encephalomyelitis viruses(TMEV) belong to the Picornaviridae and arenatural pathogens of mice. In susceptible strains,such as SJL or DBA/1, persistent infection of theCNS with TMEV leads to chronic progressive, im-mune-mediated demyelination (reviewed in Ref.1). Histological features of the pathological lesionin TMEV infection and MS are very similar, al-though chronic demyelination in TMEV is inducedin the absence of antimyelin responses by B cells.The Theiler’s virus model has served as a useful
tool to study immunotherapies targeted at remyelination10.
Experimental autoimmune neuritisThe first successful attempt to induce experi-mental autoimmune neuritis (EAN) was con-ducted by B. Waksman’s immunization of rabbitswith PNS tissue in adjuvant (reviewed in Ref. 3).Like EAE, well-defined neuritogenic componentsof PNS myelin, such as P2 protein or P0 protein,were then defined concomitantly with the de-scription of an AT-EAN. The methodology of iso-lating encephalitogenic T-cell lines could be eas-ily transferred to establish P2 specific,neuritogenic T cells. Nevertheless, EAN hasnever reached the popularity of EAE; this mightalso be due to the lower incidence of GBS andCIDP compared with MS. In our opinion, EAN
90
Disease models MOLECULAR MEDICINE TODAY, FEBRUARY 2000 (VOL. 6)
Table 1. Antigen-induced animal models for autoimmune demyelinating disorders a
Model Similarities to human disease Differences from human disease Further comments
Lewis rat
Active EAE T-cell inflammation and weak antibody Axonal damage, monophasic; Reliable model, commonly used for (CNS myelin, MBP, MOG, PLP) response secondary demyelination therapeutic studies
Adoptive-transfer EAE T-cell inflammation; topography of Axonal damage (MBP) Homogenous course, rapid onset; (MBP, S-100) lesions differential recruitment of T cells
and macrophages depending on autoantigen
Active EAE + co-transfer of T-cell inflammation and demyelination Only transient demyelination First model to demonstrateanti-MOG antibodies pathogenic role of antibodies
Active EAN Strong demyelination With myelin inoculation: Only mild disease with PMP-22(PNS myelin, P2, PMP22) also CNS inflammation
Adoptive-transfer EAN (P2, P0) Severe T-cell inflammation Only moderate demyelination Superimposed ischemic Wallerian degeneration
Congenic Lewis (DA, BN strains)
Active EAE (MOG) Relapsing–remitting disorders mimicking Increasing severity of relapses Pathology in the optic nerve, axonal histopathology of MS and its subtypes damage similar to MS
Murine EAE (SJL, PL/J, C57/Bl6 strains)
Active EAE (MBP, MOG, PLP) Relapsing–remitting and chronic disease Severe effects on the animals’ health- In many cases, pertussis (toxin) courses with demyelination and condition resulting from non-specific required; variability of susceptibility axonal damage systemic disease (cytokine stress?) and disease course
aAbbreviations: EAE, experimental autoimmune encephalomyelitis; EAN, experimental autoimmune neuritis; MBP, myelin basic protein; MOG, myelin-oligodendrocyte glycoprotein; PLP,proteolipid protein; MS, multiple sclerosis.

serves as an invaluable adjunct to EAE to char-acterize mechanisms of T-cell mediated autoim-munity.
Like EAE in the Lewis rat, EAN is monophasic.A relapsing disease course is only observed whenimmunopharmacological modifiers such as cyclo-sporin A are given, or different strains are chal-lenged. In EAN, specific neurophysiological meth-ods can be applied to yield functional informationon myelin integrity and axonal damage, which issimilar to GBS and CIDP. This allows for a bettermonitoring of immunotherapies, which can be di-rected at the induction or at the effector phase ofthe disease. Chronic EAN induced by immunizationof rabbits with galactocerebroside underscores thepathogenetic contribution of circulating autoanti-bodies for the induction of demyelinating lesions.
Concluding remarksIn the CNS, interest is focused on MS as a chronicdemyelinating disease, whereas, in the PNS, themonophasic acute form GBS attracts greatest at-tention. Animal models for autoimmune, demyeli-nating disorders provide a rational basis for study-ing mechanisms of pathogenesis and newimmunotherapeutic strategies for MS, GBS andCIDP (reviewed in Refs 3,9). Owing to the com-plexities of human disease, it is apparent that thereis no single model (Table 1). Instead, the adaptationof different approaches will finally help us to developnew and more effective therapeutic strategies.
Acknowledgments. We apologize to those whosework has not been cited owing to space limitations in
this short article. We thank Professor Hans Lassmann(Vienna) for many stimulating discussions, Drs MartinaStienekemeier, Lisa Schönrock and Stefan Jung forsharing unpublished data, Professors Rudolf Martiniand Michael Sendter for helpful suggestions. Our workis supported by grants from the DeutscheForschungsgemeinschaft, Bundesministerium fürForschung und Technik, Gemeinnützige Hertie-Stiftung,Wilhelm-Sander-Stiftung, German Multiple SclerosisSociety, Austrian Multiple Sclerosis Society, AustrianResearch Foundation, and funds from the State ofBavaria and the University of Graz.
References01 Raine, C.S. et al. (1997) Multiple Sclerosis. Clinical
and Pathological Basis. Chapman & Hall02 Compston, A. et al. (1998) McAlpine’s Multiple
Sclerosis, (3rd edn), Churchill Livingstone03 Antel, J. et al. eds (1998) Clinical Neuroimmunology.
Blackwell Science04 Gold, R. et al. (1997) T-cell apoptosis in autoimmune
diseases: termination of inflammation in the nervoussystem and other sites with specialized immune-de-fense mechanisms. Trends Neurosci. 20, 399–404
05 Hafler, D.A. et al. (1996) TCR usage in human andexperimental demyelinating disease. Immunol.Today 17, 152–159
06 Storch, M.K. et al. (1998) Autoimmunity to myelinoligodendrocyte glycoprotein in rats mimics thespectrum of multiple sclerosis pathology. BrainPathol. 8, 681–694
07 Kojima, K. et al. (1994) Experimental autoimmunepanencephalitis and uveoretinitis transferred to theLewis rat by T lymphocytes specific for the S100
beta molecule, a calcium binding protein of as-troglia. J. Exp. Med. 180, 817–829
08 Lafaille, J.J. et al. (1994) High incidence of sponta-neous autoimmune encephalomyelitis in immuno-deficient anti-myelin basic protein T cell receptortransgenic mice. Cell 78, 399–408
09 Hohlfeld, R. (1997) Biotechnological agents for theimmunotherapy of multiple sclerosis – Principles,problems and perspectives. Brain 120, 865–916
10 Miller, D.J. et al. (1996) Central nervous system re-myelination clinical application of basic neuro-science principles. Brain Pathol. 6, 331–344
Ralf Gold MD*Consultant in Neurology and Head
Clinical Research Group for Multiple Sclerosisand Neuroimmunology
Hans-Peter Hartung MDProfessor and Chairman
Department of Neurology, University of Graz,Austria.
Klaus V. Toyka MDProfessor and Chairman
Department of Neurology, University ofWurzburg, Neurologische Universitätsklinik,
Josef-Schneider-Str. 11, 97080 Würzburg,Germany.
Tel: 149 931 201 5755Fax: 149 931 201 3488
e-mail: [email protected]
Disease modelsMOLECULAR MEDICINE TODAY, FEBRUARY 2000 (VOL. 6)
91
http://tto.trends.comEditor: Adrian Bird, Institute for Cell and Molecular Biology at the University of Edinburgh
New Technical Tip articles published recently in Technical Tips Online include:Tanaka, K.J. and Nishikata, T. (1999) A non-radioactive gel shift protocol enables recovery of RNA-binding proteins(http://tto.trends.com) t01794
Goldberg, M. (1999) The use of green fluorescent fusion proteins for efficient and reliable in vitro binding assays(http://tto.trends.com) t01801
Petek, E. and Wagner, K. (1999) A rapid and efficient PCR approach for the generation of direct-labeled DNA probes forFISH analysis (http://tto.trends.com) t01757
Kuschak, T.I. et al. (1999) FISH on purified extrachromosomal DNA molecules (http://tto.trends.com) t01669
Coleman, A.E. et al. (1999) Use of B1-repeat supplemented Cot-1 DNA to enhance background suppression in FISH(http://tto.trends.com) t01769





























