Embed Size (px)
Citation preview

Anela Topĉagić
Lejla Klepo
Ismet Tahirović
BIOHEMIJA II
LABORATORIJSKE VJEŽBE
(INTERNA SKRIPTA)
Student:____________________________
Grupa:___________________
Sarajevo, 2016.


BIOHEMIJA II–laboratorijske vjeţbe
2
Vježba broj 1
ODREĐIVANJE SADRŽAJA ASKORBINSKE KISELINE
U UZORKU VOĆNOG SOKA
1. TEORETSKI DIO
Vitamini su spojevi prisutni u hrani ţivotinjskog i biljnog porijekla u malim količinama
koji su neophodni za rast i odrţavanje organizma. Iako su po hemijskom sastavu i fiziološkim
ulogama u organizmu heterogeni spojevi, vitamini ipak imaju nekoliko zajedničkih osobina.
Prema rastvorljivosti u vodi ili uljima dijelimo ih na hidrosolubilne i liposolubilne vitamine.
Vitamini topljivi u vodi (hidrosolubilni) su:
Vitamini B kompleksa (B1, B2, B3, B5, B6, B9, B12)
Vitamin C
Vitamin H
Vitamini topljivi u ulju (liposolubilni) su:
Vitamin A
Vitamin D
Vitamin E
Vitamin K
Vitamini su uglavnom proizvodi biljaka i nekih mikroorganizama. U organizam se
unose u malim količinama – neki i u mikrogramskim količinama. Poslije resorpcije deponuju
se u nizu organa, a prije svega u jetri. Zbog toga i mnoge ţivotinjske namirnice-jetra i drugi
unutrašnji organi, riblje ulje, mlijeko, jaja itd., sluţe kao izvor vitamina. U organizmu vitamini
imaju vaţne fiziološke uloge. Mnogi od njih ulaze u sastav koenzima mnogih enzima u
organizmu ili su neophodni za plodnost, razvoj embriona i fetusa, za procese rasta i razvoja, za
pravilno sazrijevanje eritrocita, za pravilan razvoj i strukturu epiderma i epitela.
Nedostatak vitamina u hrani dovodi do odreĎenih poremećaja u organizmu, koji su
specifični za svaki vitamin. Primijećeno je da koţa osobito osjetljivo reagira na pomanjkanje
različitih vitamina. Hipervitaminoze su poznate kod nekih vitamina topljivih u ulju (A, D). One
su gotovo uvijek posljedica pogrešnog doziranja pri terapiji vitaminima. U normalnim uvjetima
prehrane praktično ne postoje avitaminoze, iako su one uvijek posljedica jednostrane prehrane.
Klinički su vaţnije, hipovitaminoze, tj. relativno stanje pomanjkanja vitamina koje još ne
dovodi do klasične slike bolesti. Kod nekih vitamina organizam sisavaca moţe provesti
posljednji stupanj sinteze, tj. moţe prevesti provitamin u sam vitamin. U tim slučajevima
(vitamin A) potreba se najvećim dijelom pokriva uzimanjem provitamina. Pokriću normalne
potrebe za vitaminima znatno pridonose bakterije crijeva koje moramo smatrati simbiotima.
Npr. potrebu čovjeka za vitaminom K u znatnoj mjeri pokrivaju bakterije.

BIOHEMIJA II–laboratorijske vjeţbe
3
Tabela: Pregled i hemijske strukture vitamina
Naziv
vitamina Hemijska struktura
Hemijski naziv
vitamina
Preporučene
doze Uloga u organizmu
Bolest usljed
nedostatka
vitamina
B1 N
N
NH2
CH3
N+
S
CH3
OH
Cl-
Tiamin 1,2 mg
Dekarboksilacija,
transfer
aldehidne grupe
Beriberi
B2 N N
NNH
O
O
OH
OH
OH
OH
CH3
CH3
H
H
H
HH
Riboflavin 1,3 mg Reakcije
redukcije Ariboflavinoza
B3
N
O
NH2
Nikotinamid,
Nikotinska
kiselina 16,0 mg
Reakcije
redukcije Pelagra
B5
O
ONH
OH
OHCH3
CH3
OH
Pantotenska
kiselina 5,0 mg
Prenos acilne
grupe Parestezija
B6
NOH
CH3
OH
OH
Piridoksin 1,3–1,7 mg Prenos amino
grupe
Anemija
periferne
neuropatije

BIOHEMIJA II–laboratorijske vjeţbe
4
B7 S
NH
NHOH O
H
H
O
Biotin 30,0 µg Reakcija
karboksilacije Dermatitis
B9 N
NH
N
N
NHO
O
O
NH2
NH
O OH
OH
Folna
kiselina 400 µg Prenos C1 grupe
Megaloblastična
anemija
B12
N
Co+
N
N
N
NN
O
PN
OO
-
OO
NH
O
O
O
O
OH
O
O
O
CH3
CH3
CH3
CH3
NH2
NH2
NH2
CH3
NH2
NH2
NH2
CH3CH3
CH3
OH
CH3
CH3
CH3
HH
H
H
Kobaltamin 2,4 µg Intramolekularna
premještanja
Perniciozna
anemija
BIOHEMIJA II–laboratorijske vjeţbe
5
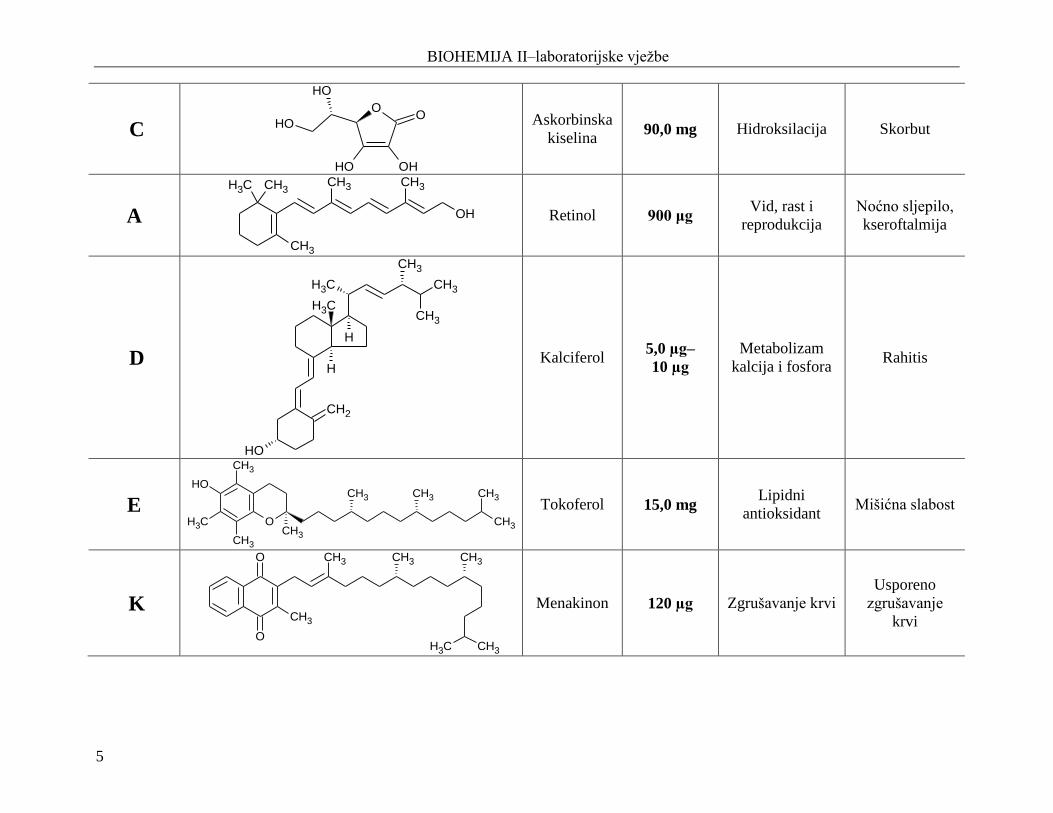
C
O
OH OH
O
OH
OH
Askorbinska
kiselina 90,0 mg Hidroksilacija Skorbut
A OH
CH3
CH3CH3CH3 CH3
Retinol 900 μg Vid, rast i
reprodukcija
Noćno sljepilo,
kseroftalmija
D
CH2
CH3
OH
CH3
CH3
CH3
CH3
H
H
Kalciferol 5,0 µg–
10 µg
Metabolizam
kalcija i fosfora Rahitis
E O
OH
CH3
CH3
CH3
CH3
CH3
CH3CH3
CH3
Tokoferol 15,0 mg Lipidni
antioksidant Mišićna slabost
K
O
O
CH3
CH3
CH3CH3
CH3CH3
Menakinon 120 µg Zgrušavanje krvi
Usporeno
zgrušavanje
krvi

BIOHEMIJA II–laboratorijske vjeţbe
6
L-askorbinska kiselina (vitamin C)
Pod vitaminom C podrazumijeva se L-askorbinska kiselina. Vitamin C je ketolakton
sa šest ugljikovih atoma, pa je po strukturi jako sličan glukozi i topiv je u vodi. To je
kristalni spoj, vrlo kiselog karaktera, ima sposobnost reverzibilnog prelaska iz keto u enolni
oblik, te igra ulogu oksidaciono-redukcionog sistema. Najbogatiji izvori su šipak, limun,
paprika, kao i drugo svjeţe voće i povrće. Osim biljaka, mogu ga sintetizirati i mnoge
ţivotinjske vrste. Ljudski organizam nije u mogućnosti da sintetizira vitamin C, te ga
čovjek mora unositi hranom .
Rastvori askorbinske kiseline su nestabilni, jer podlijeţu razgradnji u prisustvu
oksidacijskog sredstva, a pogotovo molekulskog kisika. Oksidacijom L-askorbinska
kiselina prelazi u formu L-dehidroaskorbinske kiseline, koja zadrţava vitaminska svojstva,
a daljnjom oksidacijom i prelaskom u 2,3-diketogulonsku kiselinu gubi se vitaminska
aktivnost. Oksidaciju L-askorbinske kiseline potiču i svijetlo, reakcije u baznoj sredini i
teški metali (Fe, Cu, Ag). Oksidaciju u biološkom materijalu kataliziraju i razni enzimi.
Obzirom da oksidacijom L-askorbinske kiseline nastaju inaktivni spojevi, koji ne posjeduju
vitaminsko djelovanje, tokom obrade potrebno je paziti na uslove rada i time smanjiti
njenu razgradnju. Kako je L-askorbinska kiselina termolabilna, povišene temperature
takoĎer pospješuju razgradnju. Kuhanjem namirnica pri visokim temperaturama dolazi do
njene razgradnje. Reaguje kao reducent u mnogim biološkim procesima. Vaţna je za
sintezu kolagena i karnitina, kao i za sintezu hormona serotonina i adrenalina, te za
metabolizam masnih kiselina.
Postoji veliki broj metoda odreĎivanja sadrţaja vitamina C. Spektrofluorimetrijska
metoda za kvantifikaciju vitamina C zasniva se na njegovoj oksidaciji u L-
dehidroaskorbinsku kiselinu, koja u reakciji sa o-fenilendiaminom (OFDA) u baznoj sredini
daje N-heterociklični spoj sa velikim brojem konjugiranih π-veza, koji moţe emitovati jaku
fluorescenciju. Intenzitet fluorescencije nastalog spoja proporcionalan je koncentraciji i
mjeri se pri talasnoj duţini ekscitacije oko 350 nm i emisije oko 430 nm.
Mehanizam fluorescirajuće reakcije odreĎivanja vitamina C

BIOHEMIJA II–laboratorijske vjeţbe
7
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
o Askorbinska kiselina (AK) (radna koncentracija): 0,100 g AK otopiti u 100 ml
destilovane vode (1mg/ml) i čuvati u tamnoj reagens boci.
o o-fenilendiamin (OFDA): 0,5 g OFDA otopiti u 100 ml 0,1 mol/l HCl i čuvati u
tamnoj reagens boci na 4 °C.
o Pufer NH3-NH4Cl (pH 9,4): 13,5 g NH4Cl i 15,75 ml amonijaka otopiti u 100 ml
destilovane vode. Zatim se doda rastvor 0,1 mol/l NaOH, da bi se dobio optimalan
pH otopine (provjeriti pH metrom)
Upotrebom radne koncentracije, pripremiti u odmjernim sudovima od 10 ml standardne
otopine AK slijedećih koncentracija: 0,5; 1; 5; 10; 30; 50 i 70 μg/ml.
Izvagati 1g uzorka voća ili povrća i macerirati sa 9 ml destilovane vode, a potom ga
centrifugirati pri 15 000obr/min, 15 minuta. Od dobijenih supernatanata pripremiti deset
puta razblaţeniji uzorak. U slučaju uzorka voćnog soka, uzorak samo centrifugirati prema
istim uslovima i razblaţiti na isti način kao uzorak voća ili povrća.
Ovako pripremljeni rastvori AK i uzoraka se koriste za mjerenje. Rastvori se miješaju po
slijedećem redoslijedu: 1 ml AK, 1,0 ml 0,5% OFDA, 1,5 ml pufera NH3-NH4Cl. Rastvor
se dopuni do 10 ml s destilovanom vodom, dobro izmiješa i ostavi da stoji 5 minuta prije
mjerenja. Intenzitet fluorescencije se mjeri u kvarcnoj kiveti od 1 cm pri talasnoj duţini
ekscitacije od 330 nm i talasnoj duţini emisije od 430 nm.
Izračunati koncentraciju AK u uzorku i rezultat izraziti kao mg/100 ml uzorka ili kao
mg/100 g uzorka.
3. ZADACI I VJEŽBE
1. Koji od navedenih vitamina, kao strukturne elemente, sadrţi izoprenske fragmente:
a) vitamin E
b) vitamin A
c) nikotinska kiselina
d) vitamin K
2. Koji od navedenih vitamina je derivat sterola:
a) vitamin B12
b) vitamin D
c) vitamin A
d) vitamin B2
3. Kakva je hemijska struktura askorbinske kiseline:
a) derivat sterola
b) derivat izoaloksazina
c) amid piridin-3-karbonske kiseline
d) lakton dienilgulonske kiseline (2-keto-L (-)-gulonkiseli-γ-lakton)
4. Spojite vitamine sa ponuĎenim odgovorima
A. Nikotinamid a. Sadrţi prsten tiazola
B. Tiamin b. U organizmu ţivotinja sintetizira se iz triptofana

BIOHEMIJA II–laboratorijske vjeţbe
8
C. Pantotenska kiselina c. Učestvuje kao koenzim u reakcijama transaminacije
i dekarboksilacije aminokiselina
D. Piridoksalfosfat d. Ulazi u sastav koenzima A
5. Vitamini A i D se mogu uzeti odmah, u jednoj dozi, u takvoj količini da je dovoljna
za odrţavanje njihove normalne koncentracije u toku nekoliko sedmica. Vitamine B
grupe neophodno je uzimati znatno duţe i češće. Zašto?
_________________________________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
6. Ako je dnevna potreba za vitaminom C 75 mg, koliko mg ili g ispitivanog uzorka je
potrebno uzeti da bi se zadovoljila dnevna potreba za vitaminom C?

BIOHEMIJA II–laboratorijske vjeţbe
9
4. REZULTATI
Standard / Uzorak Intenzitet fluorescencije
OdreĎivanje sadrţaja vitamina C u uzorku (izraziti kao mg/100 ml uzorka)
OVJERA VJEŽBE:

BIOHEMIJA II–laboratorijske vjeţbe
10
Vježba broj 2
KVANTITATIVNO ODREĐIVANJE UKUPNIH LIPIDA I TRIGLICERIDA U
SERUMU ILI PLAZMI
1. TEORETSKI DIO
Ukupni serumski (plazma) lipidi su sastavljeni od triglicerida, fosfolipida,
holesterola, estera holesterola, neesterificiranih masnih kiselina, glikolipida (cerebrozida),
acetilfosfatida (plazmalogena), fosfatidnih kiselina, viših alkohola, karotinoida, steroidnih
hormona, te vitamina A, D, E. Po svojoj vaţnosti, naročito je bitno u uzorcima odrediti
koncentracije triglicerida i ukupnog holesterola.
U analitici ukupnih lipida opisane su gravimetrijske i turbidimetrijske metode,
računske, te kolorimetrijske metode.
Ukupni lipidi plazme (seruma) se odreĎuju spektrofotometrijski, reakcijom sa
vanilinom i fosfornom kiselinom. Pretpostavka je da pri dodavanju koncentrirane sulfatne
kiseline serumu uz zagrijavanje, svi lipidi koji u svojoj strukturi posjeduju dvostruke veze
daju ketone ili ketole. Ovi potom reagiraju sa vanilinom u fosfornoj kiselini uz nastajanje
ruţičasto crvene boje, čiji intenzitet apsorpcije se mjeri izmeĎu 510-550 nm. Reakcija
odreĎivanja ukupnih lipida se odvija u tri stepena:
1. Nezasićena masna kiselina reagira sa H2SO4 stvarajući karbonium jon:
2. Vanilin reagira sa H3PO4 stvarajući aromatski ester:
3. Karbonijum jon reagira s aktiviranom karbonilnom grupom fosfovanilina stvarajući
nabijeni ruţičasto obojeni kompleks, koji se stabilizira rezonancijom i maksimalno
apsorbira kod 525 nm:
Trigliceridi su esteri glicerola i masnih kiselina. Glavni su sastojak ţivotinjskih i
biljnih masti i ulja. Masti nas opskrbljuju velikom količinom energije. Nalazimo ih u

BIOHEMIJA II–laboratorijske vjeţbe
11
masnom tkivu i u masnim kapljicama ostalih ćelija. Zbog toga su masti i ulja vrlo vaţna
skupina spojeva u ljudskoj prehrani. One su rezerva energije neophodne svakom ţivom
organizmu za njegov razvoj i odrţavanje. Vrlo su široko rasprostranjene u prirodi, a osim
biološkog, imaju i veliko tehničko značenje. One sluţe kao sirovina za proizvodnju sapuna,
margarina, raznih maziva, lakova i boja. Maslac i margarin su vrlo česti izvori masti u
prehrani. U probavi hrane, mast se razgraĎuje tek u tankom crijevu. Ţuč, koja dolazi iz
ţučnog mjehura u duodenum (prvi dio tankog crijeva), zajedno s hranom putuje kroz tanko
crijevo i razgraĎuje mast na njene komponente: glicerol i masnu kiselinu. Masna kiselina se
dalje probavlja, a glicerol se metabolizira u jetri.
Trigliceridi se odreĎuju po hemijskom ili enzimskom principu odreĎivanja
glicerola. Samo odreĎivanje triglicerida se sastoji od ekstrakcije, potom odvajanja
fosfolipida i drugih hromogena, te saponifikacije, odnosno ekstrakcije, nakon čega slijedi
direktno odreĎivanje glicerola. Faza ekstrakcije se izvodi u rastvaračima poput metanola,
etanola, izopropanola ili hloroforma. Svi ovi rastvarači dovode do denaturacije lipoproteina,
dakle disocijacije vezanih triglicerida. Interferirajuće supstance se mogu ukloniti na dva
načina i to korištenjem adsorbensa poput zeolita, salicilne kiseline ili aktivnog magnezij-
silikata ili pak korištenjem nonana ili heksana. Glavne interferirajuće supstance koje se
mogu ukloniti na ovaj način su fosfolipidi i glukoza zajedno sa nekim hromogenima i
ponekad sa slobodnim glicerolom. Druga faza u sklopu ovih postupaka uključuje hidrolizu
triglicerida na glicerol i masne kiseline i obično se izvodi sa etanolnim rastvorom kalij-
hidroksida pri visokim temperaturama (saponifikacija). U trećoj fazi dolazi do oksidacije
glicerola u formaldehid, koji se prevodi u reakciji sa acetilacetonom u obojeni spoj.
OdreĎivanje triglicerida se moţe prikazati hemijskim reakcijama u tri stepena:
Prvi stepen:
CH2
CH
CH2
O
O
O
C
C
C
O
O
O
R
R
R
+ K OH
CH2
CH
CH2
OH
OH
OH
+ RCOOK3 3
Drugi stepen:
CH2
CH
CH2
OH
OH
OH
+ NaJO4 C
O
H H + NaJO3 + H2O3
Treći stepen:
Holesterol je najprisutniji sterol u organizmu, koji po svom hemijskom sastavu
sadrţi ugljikov skelet ciklopentanoperhidrofenantrena.

BIOHEMIJA II–laboratorijske vjeţbe
12
Struktura holesterola
Porijeklo holesterola u organizmu je dvojako, većina ćelija sintetizira holesterol, a drugi
njegov izvor je hrana kojom se unosi. Oko 2/3 holesterola nastaje sintezom u organizmu
(kod odrasle osobe oko 800-900 mg/dan), a svega 1/3 se unosi hranom (oko 150-300
mg/dan). Djeci je potrebna proporcionalno veća količina, što se opravdava njegovom
značajnom ulogom kao strukturnog elementa svih ćelijskih i unutarćelijskih membrana.
Najveći dio holesterola nastaje u jetri, a do njegove sinteze moţe doći i u sluzokoţi crijeva i
nadbubreţnim ţlijezdama. Odatle se putem krvotoka transportuje do ćelija organizma.
Pošto je nerastvorljiv u vodi, u krvi se holesterol transportuje tako što se veţe za proteine
gradeći lipoproteine. Postoji više vrsta ovih lipoproteina i dijele se prema gustoći na:
VLDL (Very Low Density Lipoproteins), lipoproteini vrlo male gustoće
LDL (Low Density Lipoproteins), lipoproteini male gustoće
HDL (Low Density Lipoproteins), lipoproteini velike gustoće
Lipoproteini sa dosta lipida imaju niţu gustoću.
U krvi holesterol je prisutan u slobodnom i esterificiranom obliku vezan sa jednom
molekulom masne kiseline. Esterifikacija holesterola odigrava se u plazmi pod djelovanjem
enzima lecitin-holesterol-acetiltransferaze (LHAT), koji se nalazi u krvnoj plazmi. U
plazmi je pribliţno 75 % ukupnog holesterola esterificirano najčešće nezasićenim masnim
kiselinama, (od toga 55 % linolnom kiselinom). Eliminacija holesterola iz organizma se
vrši preko ţuči (konverzijom u holne kiseline), perutanjem koţe, mala količina se gubi
urinom, dok ţene koje doje gube nešto holesterola preko mlijeka.
Holesterol je neophodan za normalno funkcioniranje svake ćelije, kao bitan
strukturni element. Specifične uloge holesterola su:
Sinteza ţučnih kiselina u hepatocitima
Sinteza steroidnih hormona u kori nadbubreţnih i polnih ţlijezda
Transport liposolubilnih vitamina (A, D, E i K)
Štetno djelovanje holesterola se ispoljava kada je u krvi prisutan u znatno većim
količinama. Povećanje LDL-a, a time i holesterola u krvnoj plazmi dovodi do povišenog
ulaska estera holesterola u ćelije krvnih sudova, gdje dolazi do taloţenja estera holesterola,
što moţe da dovede do začepljenja krvnih sudova, smanjenja njihove prirodne elastičnosti,
a u kasnijim stadijima, formiranja tromba i infarkta miokarda.
Prilikom uklanjanja holesterola iz organizma slobodni holesterol dospijeva u ţuč u
kojoj je nerastvorljiv. On se u ţuči inkorporira u micele koje čine lecitin i ţučne soli. Ove
micele imaju ograničen kapacitet rastvaranja holesterola. Kod ljudi sa kamenom u ţučnoj
kesi dolazi do formiranja abnormalne ţuči, koja postaje prezasićena holesterolom. Pod
djelovanjem različitih faktora dolazi do viška taloţenja holesterola u vidu kristala. Ako se
kristali ne izluče u crijevo, narastaju i formiraju kamenje.
Metode odreĎivanja holesterola su spektrofotometrijske, enzimske i
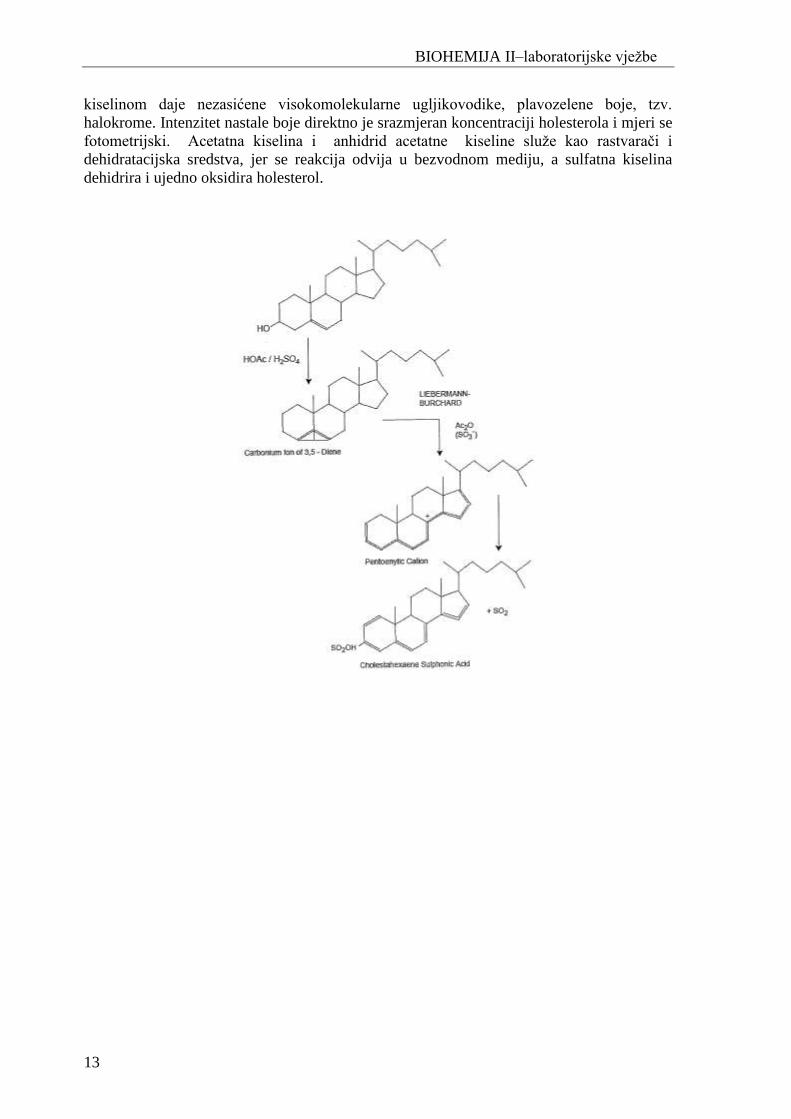
hromatografske. Najčešće se primjenjuje Liberman-Burchard-ova metoda odreĎivanja, gdje
holesterol u bezvodnoj sredini sa anhidridom acetatne kiseline i koncentriranom sulfatnom
BIOHEMIJA II–laboratorijske vjeţbe
13
kiselinom daje nezasićene visokomolekularne ugljikovodike, plavozelene boje, tzv.
halokrome. Intenzitet nastale boje direktno je srazmjeran koncentraciji holesterola i mjeri se
fotometrijski. Acetatna kiselina i anhidrid acetatne kiseline sluţe kao rastvarači i
dehidratacijska sredstva, jer se reakcija odvija u bezvodnom mediju, a sulfatna kiselina
dehidrira i ujedno oksidira holesterol.

BIOHEMIJA II–laboratorijske vjeţbe
14
2. EKSPERIMENTALNI DIO
2.1. ODREĐIVANJE UKUPNIH LIPIDA
Potrebni reagensi:
- Sulfatna kiselina, koncentrirana
- o-fosfatna kiselina, p.a.
- Vanilin 0,008 M vanilina (C8H8O3, Mr 152,15) otopi se u 100 ml redestilirane vode
- Fosfovanilinski reagens: fosfatna kiselina 11,9 M i vanilin 0,008 M, 1 dio fosfatne
kiseline pomiješa se s 1 dijelom (1:1) otopine vanilina. Reagens je stabilan u tamnoj
boci na sobnoj temperaturi 2 sedmice
- Standard maslinova ulja, 1 g maslinova ulja/100 ml etanola.
Otopine su stabilne jednu godinu na sobnoj temperaturi, ako su zatvorene. Otopine
treba čuvati u tamnim bocama.
U epruvete s brušenim staklenim čepom otpipetira se:
_________________________________________________
Proba Standard
_________________________________________________
Serum 50 μl -
Standard lipida - 50 μl
H2SO4 2,0 ml 2,0 ml
Sulfatna kiselina dodaje se tako da se pokupe svi tragovi seruma na zidovima epruvete na
dno. Epruvete se stave 10 minuta u ključalo vodeno kupatilo i ohlade u hladnoj vodi.
U nove epruvete se otpipetira:
Smjesa analize: 100 μl - -
Smjesa standarda lipida: - 100 μl -
Fosfovanilinski reagens: 2,0 ml 2,0 ml 2,0 ml
Miješati i ostaviti da stoji 30 min. a nakon toga se mjeri apsorpcija (A) probe i standarda
lipida prema slijepoj probi kod 530 nm u kiveti od 1 cm.
Odrediti sadrţaj ukupnih lipida u uzorku seruma, a rezultat izraziti kao g/L.
Referentne vrijednosti ukupnih lipida: 4 do 8 g/l.
2.1. ODREĐIVANJE TRIGLICERIDA
Potrebni reagensi:
- n-heptan;
- izopropanol;
- sulfatna kiselina, 0,04 mol/L;
- acetatna kiselina, 1 mol/L;
- kalij-hidroksid 6,25 mol/L;
- rastvor metaperjodata: rastvoriti 3 g natrij-metaperjodata i 25 mL glacijalne acetatne
kiseline u destiliranoj vodi i dopuniti vodom do 500 mL;
- amonij-acetat, 2 mol/L;

BIOHEMIJA II–laboratorijske vjeţbe
15
- rastvor acetilacetona: rastvoriti 0,75 mL acetilacetona i 2,5 mL izopropanola i
dopuniti do 100 mL 2,0 mol/l rastvorom amonij-acetata. Rastvor čuvan u tamnoj
boci stabilan je mjesec dana na +4 °C;
- standard: maslinovo ulje.
Pipetirati u epruvete sa brušenim zatvaračima prema priloţenoj tabeli:
1. Ekstrakcija
Slijepa proba Standard Uzorak
Destilirana voda 0,5 mL
Standard 0,5 mL
Serum 0,5 mL
n-heptan 2,0 mL 2,0 mL 2,0 mL
Izopropanol 3,5 mL 3,5 mL 3,5 mL
Sulfatna kiselina 1,0 mL 1,0 mL 1,0 mL
Miješati na vorteksu oko 30 s i sačekati da se odvoje slojevi.
2. Saponifikacija i oksidacija
Na 0,4 mL heptanskog sloja (gornji sloj) dodati 2 mL izopropanola i kap KOH. Dobro
promiješati i inkubirati na 70 °C u trajanju od 10 min. Nakon toga dodati 0,2 mL reagensa
NaJO4 i 1 mL reagensa acetilacetona. Dobijeni rastvor dobro promiješati i inkubirati na 70
°C u trajanju od 10 min. Nakon toga, smjesu ohladiti na sobnu temperaturu. Dobijenom
rastvoru mjeriti apsorbansu na 425 nm. Boja je stabilna oko sat vremena. Standardne
rastvore čije koncentracije idu do 2 g/L tretirati na isti način. Na osnovu dobijenih
vrijednosti apsorbansi za standard triglicerida, konstruirati kalibracionu krivu i pomoću
jednačine pravca odrediti sadrţaj triglicerida. Sadrţaj triglicerida izraziti kao g/L.
3. ZADACI I VJEŽBE
1. OslobaĎanje masnih kiselina u toku posta ide iz kog organa:
a) Jetre
b) Masnog tkiva
c) Epitela tankog crijeva
d) Limfe
2. Koji se proces naziva saponifikacija masti:
a) Enzimska hidroliza
b) Alkalna hidroliza
c) Hidrogenizacija
d) Reakcija sa olovo-acetatom
e) Emulzifikacija masti
3. Kakvom pretvaranju podlijeţe glicerol koji nastaje u prvoj etapi pri razgradnji
triacilglicerola:
a) Redukciji
b) Oksidaciji
c) Metiliranju
d) Fosforiliranju
e) Aciliranju

BIOHEMIJA II–laboratorijske vjeţbe
16
4. Koji od navedenih lipoproteina se karakterizira kao „loši“ holesterol:
a) LDL
b) HDL
c) Hilomikroni
d) IDL
5. Koji se od vitamina ponašaju kao lipidi:
a) vitamin C
b) vitamin K
c) vitamin A
d) vitamin E
e) vitamin D
6. Koji sastojci ćelije sadrţe fosfolipide:
a) mijelinski omotač nervnih ćelija velikog mozga
b) hijeloplazma
c) kompleks membrana
7. Objasniti ulogu sulfatne kiseline u reakciji odreĎivanja ukupnih lipida?
_________________________________________________________________________
_________________________________________________________________________
_________________________________________________________________________
_________

BIOHEMIJA II–laboratorijske vjeţbe
17
4. REZULTATI
- OdreĎivanje koncentracije ukupnih lipida (rezultat izraziti kao g/L)
Uzorak Apsorpcija (A)
- OdreĎivanje koncentracija triglicerida (rezultat izraziti kao g/L)
Uzorak Apsorpcija (A)
OVJERA VJEŽBE

BIOHEMIJA II–laboratorijske vjeţbe
18
Vježba broj 3
KVANTITATIVNO ODREĐIVANJE HOLESTEROLA PO
LIBERMANN-BURCHRD-U
1. TEORETSKI DIO
Holesterol pripada grupi supstanci koje nazivamo steroidi. Oni u svojoj strukturi imaju
ciklopentanoperhidro fenantrenski prsten. On je veoma vaţan za normalno funkcionisanje
organizma jer predstavlja jednu od esencijalnih komponenti membrana svih ţivotinjskih
ćelija, prekursor je u biosintezi ţučnih kiselina kao i prekursor u biosintezi steroidnih
hormona.
OdreĎivanje ukupne koncentracije holesterola implicira odreĎivanje esterificiranog i
nesterificiranog holesterola. U serumu (plazmi) oko 2/3 ukupnog holesterola se javlja u
esterificiranoj formi, a ostatak u slobodnoj formi.
U analitici holesterola javljaju se tzv. jednostepene, dvostepene, trostepene i četverostepene
metode. Kod jednokratnih metoda ne postoji u klasičnom smislu te riječi ono što se naziva
pripremanje uzorka, to znači da ne postoji izolacija i pročišćavanje steroida. Kod ovih
metoda jedan reagens je dovoljan za manipulaciju sa uzorkom. S toga su ove metode jako
pogodne za automatizaciju.
Kod dvostepenih metoda potrebno je uraditi ekstrakciju u organskoj fazi prije mjerenja
holesterola i drugih njemu sličnih steroida. Ova procedura uklanja nespecifične
hromatogene koji mogu inferferirati.
Trostepene metode se od dvostepenih razlikuju po tome što pored ekstrakcije holesterola
ovdje imamo zastupljen i proces saponifikacije koji obezbjeĎuje hidrolizu
masnokiselinskog ostatka od holesterolskog estra. Uslijed toga mjeri se samo slobodni
holesterol.
Kod četvorofaznih metoda pored metode esterifikacije, saponifikacije i razvijanja boje
imamo i purifikaciju holesterola koja se postiţe dodavanjem saponina, digitronina.
LIEBERMANN-BURCHARD-OVA REAKCIJA
Ova reakcija se izvodi u jako kiseloj sredini koju obezbjeĎuju sulfatna kiselina, acetatna
kiselina i acetat anhidrid. Holesterol u bezvodnoj sredini sa anhidridom acetatne kiseline i
koncentrovanom sulfatnom kiselinom daje nezasićene visokomolekularne ugljikovodike,
plavozelene boje, tzv. halokrome ( Liberman-Burchard-ova reakcija ). Intenzitet nastale
boje direktno je srazmjeran koncentraciji holesterola i mjeri se fotometrijski. Acetatna
kiselina i anhidrid acetatne kiseline sluţe kao otapala i dehidratacijska sredstva jer se
reakcija odvija u bezvodnom mediju, a sulfatna kiselina dehidrira i ujedno oksidira
holesterol.
U Liberman-Burchard-ovoj reakciji nastaju plavozeleni polienski kationi i SO2.
U hemijskom smislu holesterol se podvrgava stepenastoj oksidaciji gdje u svakoj fazi
nastaje holestapoilen molekula, koja uvijek ima jednu dvostruku vezu više od supstance od
koje originalno potiče. U prvoj fazi reakcije uključena je protonacija hidroksline grupe
holesterola i gubitak vode tako da se dobije tzv. karbonijum ion ili 3,5 holestadien, čije
dobivanje predstavlja prvu fazu kod razvijanja boje. Stepenasta oksidacija ovog aliličnog
karbonijum jona putem sulfita uzrokuje stvaranje holestaheksan-sulfonske kiseline kao
hromafora koji pokazuje maksimum apsorpcije na 410nm.
Ova klasična metoda doţivjela je niz promjena u smislu da se na istom principu moglo
vršiti odreĎivanje bilo slobodnog bilo esterificiranog holesterola ili pak oba.

BIOHEMIJA II–laboratorijske vjeţbe
19
Liebermann-Bruchard-ova reakcija je kompleksna i ovisi od više faktora poput
koncentracije sulfatne kiseline, vode, sadrţaja acetatne kiseline u anhidridu, rastvarača,
vremena reakcije te uticaja svjetlosti i temperature.
Slika 1: Mehanizam reakcije po Liebermann-Burhardu
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- holesterol reagens: stabilizirana smjesa acetatne kiseline 5 mol / L, anhidrid
acetatne kiseline 5,87 mol / L, sulfatna kiselina 1,87 mol / L i natrijev sulfat 0,14
mol / L
- Standard holesterola 5,17 mmol/L ( korišten je standard holesterola 2,73 mmol / L
od firme Precinorm U )
Potrebna oprema i pribor:
- spektrofotometar S106 WPA, Velika Britanija
PROCEDURA ZA RAD
a) Priprema reagenasa:
- Holesterol-reagens: u tikvicu od 1 L pomiješa se 150 mL glacijalne acetatne
kiseline i 300 mL anhidrida acetatne kiseline ( CH3CO )2O, Mr 102,09), te se uz
hlaĎenje polahko doda 50 mL koncentrovane sulfatne kiseline. Kad se tekućine
pomiješaju, doda se 10 g bezvodnog natrijevog sulfata (Na2SO4, Mr 142,04) i
miješa se dok se potpuno ne otopi. Reagens se drţi u tamnoj boci. Stabilan je oko
četiri sedmice na + 4 C. Prije upotrebe ga treba dovesti na sobnu temperaturu.
Reagens je bezbojan ; ako se pojavi boja treba ga baciti.
- Standard holesterola: 199,9 mg holesterola (C27H46O, Mr 386,67) otopi se i dopuni
do 100 mL glacijalnom acetatnom kiselinom u odmjernoj tikvici. Neograničeno je
stabilan.

BIOHEMIJA II–laboratorijske vjeţbe
20
b) Eksperimentalna procedura: Paralelno se odreĎuju: slijepa proba, standard i uzorak.
Smjesa se laganim mućkanjem dobro promiješa i ostavi da stoji 20 minuta na sobnoj
temperaturi te izmjeri apsorpcija ( A ) probe i standarda prema slijepoj probi kod 610 nm.
Boja je postojana 30 minuta.
holesterol (mmol / L) = Euzorka / Estandarda x koncentracija standarda
Referentne vrijednosti: 3,36 do 6,72 mmol / L
Napomene: Metoda je nedovoljno specifična jer pored holesterola reagiraju i neki steroli
analogne strukture. MeĎutim , u rutinskom radu metoda uglavnom zadovoljava jer se
interferirajući spojevi nalaze u serumu u vrlo niskim koncentracijama.Tačnost metode se
smanjuje kod hemoliziranih i ikteričnih seruma. Serumi sa više od 1 g / L hemoglobina nisu
upotrebljivi, dok do 0,5 g / L hemoglobina, hemoglobin ne smeta. Normalne koncentracije
bilirubina ne smetaju, dok koncentracija od 85 mmol / L povećava apsorpciju za oko 0,01.
Takve serume treba razrijediti sa 0,15 mol / L NaCl. U odnosu na druge, ova metoda ima
prednosti u jednostavnosti postupka izvoĎenja, povećanoj stabilnosti holesterol-reagensa
dodatkom natrijevog sulfata, smanjenom utjecaju temperature na reakciju i u povećanoj
stabilnosti apsorpcije nastalog produkta. To su razlozi zbog kojih se ova metoda često
izvodi u rutinskom radu i na automatskim analizatorima.
Uzorak Standard Slijepa proba
Holesterol-reagens 4,0 ml 4,0 ml 4,00 ml
Serum 100 μl - -
Standard
holesterola
- 100 μl -
Destilirana voda - - 100 μl

BIOHEMIJA II–laboratorijske vjeţbe
21
3. REZULTATI
- Izvršiti eksperimentalni dio vjeţbe
- Vrijednosti mjerenja apsorpcije
Uzorak Apsorpcija (A)
- Koncentracija nepoznatog uzorka
OVJERA VJEŽBE

BIOHEMIJA II–laboratorijske vjeţbe
22
Vježba broj 4
RAZDVAJANJE PROTEINA GEL ELEKTROFOREZOM
(RAZDVAJANJE I IDENTIFKACIJA PROTEINA U UZORCIMA GOVEĐEG
SERUMA NA AGAROZNOM GELU UZ PRIMJENU ISTOSMJERNE ELEKTRIČNE
STRUJE)
1. TEORETSKI DIO
Elektroforeza po svojoj definiciji, je tehnika kojom se razdvajaju naelektrisane
čestice pod djelovanjem homogenog električnog polja. Tom prilikom otopljena čestica
efektivnog naboja q putuje u viskoznom puferu pod djelovanjem električnog polja prema
jednoj od elektroda, u ovisnosti od prirode njenog naboja. Ovo je jedna od vaţnijih tehnika
u separaciji bioloških molekula, jer obično ne uzrokuje promjene na nivou strukture
molekula koje se separiraju, a osim toga osjetljiva je na male promjene u naboju i masi
makromolekula.
Kretanje molekula u električnom polju predstavlja funkciju naboja q i koeficijenta trenja f.
Brzina koju ostvaruje jedan molekul sa nabojem u električnom polju je funkcija jačine polja
E tako da imamo odnos:
v=f
Eq
v - brzina kretanja molekule (m/s)
E - jačina električnog polja (V/m)
q - naboj molekule
f - koeficijent trenja (Vs/m2)
Pri tome je razdvajanje čestica u električnom polju ovisno i o vremenu.
Kao analitička metoda, elektroforeza je jednostavna, brza i vrlo osjetljiva. U kombinaciji s
drugim tehnikama molekularne biologije, postala je jedna od najčešće primjenjivanih
metoda.
Postoje dva tipa elektroforeze: slobodna i zonska.
Kod slobodne elektroforeze naelektrisani joni se slobodno kreću kroz rastvor sve
dok postoji djelovanje električnog polja. Ovdje se radi o jednom dinamičkom sistemu koji
direktna mjerenja moţe da vrši samo kod onih čestica koje svojom različitom brzinom
putovanja stvaraju mjesta različite gustoće. Za ovu potrebu koristi se U-cijev koja u donjem
dijelu sadrţi uzorak i krajeve koji su napunjeni elektrolitom kako bi se odrţavale oštre
granice prema uzorku. Kretanje uzorka se mjeri pomoću pomjeranja granica u funkciji
vremena. Na ovaj način dolazi do optičke pojave (tzv. šlira), do prelamanja svjetlosti na
granici dvije različite koncentracije i one se registruju posredstvom mjerenja indeksa
prelamanja svjetlosti. Ova tehnika uglavnom se koristi u analizama fizičkih karakteristika
molekula.
Mnogo pogodnija tehnika je tzv. zonska elektroforeza. Naelektrisane čestice se kod ove
tehnike kreću kroz interni potporni medij koji moţe biti filter papir, celuloza-acetat, škrobni
gel, agar gel, poliakrilamid gel... Prednost ove tehnike je i u tome da se razdvojene
supstance u ţeljenom poloţaju mogu stabilizirati, osušiti i hemijski fiksirati. Potporni medij
pri tome ima ulogu da spriječi sve poremećaje vezane za kretanje uzoraka, a osim toga
predstavlja u izvjesnom smislu i molekulsko sito ili pak stabilizirajući medij za pH
gradijent. Napon koji se koristi kod razdvajanja će ovisiti od tipa uzorka i vrste potpornog

BIOHEMIJA II–laboratorijske vjeţbe
23
medija. Tako npr. visoki naponi se koriste za papirne potporne medije uslijed velikog
otpora samog papira. Visoki napon se takoĎer koristi za izbjegavanje problema koji bi
nastali difuzijom i doveli tako do širenja molekula. Sa velikim naponima stvaraju se i veće
jačine struje, dolazi do stvaranja toplote što zahtijeva hlaĎenje sistema. Dodatni problem u
nekim efektivnim potpornim medijima je fenomen elektroendoosmoze u kojem pufer sam
pokazuje elektroforetski učinak i time maskira pokretljivost molekule koja se prati. Ova
pojava se moţe kao takva pripisati efektima kretanja pufera koji se javljaju uslijed
negativno nabijenog potpornog medija i dovode to toga da pufer, kada se izloţi električnom
polju se kreće prema katodi. Proteini su negativno nabijeni i kreću se prema anodi. Drugim
riječima, kada dolazi do ove pojave dolazi do njihovog kretanja suprotno kretanju pufera.
Brzina kretanja pufera se mijenja sa korištenjem različitih potpornih medija. U izvjesnom
smislu efekat elektroendoosmoze se pokazao korisnim, jer pomaţe kod razdvajanja gama
globulina od ostalih proteina. Ova frakcija proteina ima negativan naboj, ali se radi o
molekulama velike molekulske mase tako da naboj koji oni posjeduju nije dovoljan da
anulira efekat kretanja pufera.
Dostupan je širok raspon opreme za elektroforezu, ali u osnovi postoji zajednički
bazni dizajn. Dobro dizajnirana komora treba da spriječi bilo koji stepen isparavanja sa
potpornih medija, da ima učinkovit sistem hlaĎenja kada se koriste gelovi ili tehnike
visokog napona pri čemu je potrebno posebno voditi računa o sigurnosnim prekidačima.
Tipovi elektroforeze su:
ELEKTROFOREZA NA FILTER PAPIRU
ELEKTROFOREZA NA CELULOZAACETATU
ELEKTROFOREZA NA TANKOM SLOJU
KAPILARNA ELEKTROFOREZA
GEL ELEKTROFOREZA
Idealan potporni medij za elektroforezu je hemijski inertan, sa njima se lahko rukuje i u
svakom trenu je moguće kontrolirati poroznost. Iako se papir smatra pravim nosačem kod
separacije aminokiselina, malih peptida, ovaj medij neosporno pokazuje velike nedostatke,
jer se u ovom slučaju poroznost ne moţe kontrolirati. Supstance sa velikim molekulskim
masama poput proteina i nukleinskih kiselina se ne separiraju dobro na papiru, jer celulozna
vlakna pokazuju veliki otpor prema kretanju. Gelovi sa varirajućom poroznošću su idealni
kod separacije visokomolekularnih supstanci jer dozvoljavaju lagano kretanje
makromolekula uz istovremeno spriječavanje konvekcije i uz minimalnu difuziju. Tri tipa
gela se koriste u ovu svrhu: skrobni gel, poliakrilamid gel i agaroza. Aparati mogu biti za
cilindrične gelove i za gel ploče (gdje broj rupica za uzorke se utvrĎuje na osnovu broja
zuba na plastičnom češlju koji se koristi).
Najpoznatiji gelovi koji se koriste u elektroforezi su:
AGAROZNI
ŠKROBNI
POLIAKRILAMIDNI
AGAROZA KAO GEL
Agarozni gelovi se stvaraju tako što se suha agaroza otopi u vodenom puferu, potom zagrije
dok se ne dobije bistri rastvor, a zatim ohladi na sobnoj temperaturi. Agaroza je prirodni
linearni polisaharid graĎen od galaktoze i 3,6-anhidrogalaktoze. Dobija se iz agara.
Visokoporozni gel koji se dobija na ovaj način je pogodan za elektroforezu i za neke
imunološke tehnike. Agaroza se obično koristi pri koncentracijama izmeĎu 1% i 3%.
Agarozni gel se odlikuje relativno velikom veličinom para nakon njegovog hlaĎenja. Ove
pore su stabilizirane vodikovim vezama. Veličina pore se kontrolira početnom
koncentracijom agaroze. Velike pore se stvaraju pri korištenju niskih koncentracija agaroze,

BIOHEMIJA II–laboratorijske vjeţbe
24
a manje veličine se stvaraju prilikom korištenja velikih koncentracija agaroze. Velike pore i
gelovi niskih koncentracija agaroze omogućavaju separiranje vrlo velikih molekula poput
DNK, lipoproteina, imunoglobulina i enzimatskih kompleksa. Agaroza predstavlja
definitivno metod izbora kod pripreme DNK lanca za sekvencioniranje. Fragmente DNK je
pri tome moguće ekstrahirati direktno sa niskotopive agaroze (62 oC-65
oC) nakon
zagrijavanja na 65 oC. Ovaj tip gela vrlo se često koristi za imunoelektroforezu gdje se
separiraju veliki kompleksi antigen-antitijelo, a takoĎer se koristi i kod izoelektričnog
fokusiranja. Dobra strana ovih gelova je da su oni fizički jači od niskoprocentnih
poliakrilamid gelova.
SDS-PAGE kontinuirana elektroforeza (Weber-Osborn postupak) je tehnika kod koje se
deterdţent natrijumdodecilsulfat (SDS) dodaje na sistem. Uglavnom se koristi u
istraţivačke svrhe za karakterizaciju proteina u toku izolacije. Proteini velikih molekulskih
masa, proteinski kompleksi ili pak nukleinske kiseline se vrlo često razdvajaju u prisustvu
agensa koji djeluju na razbijanje nativne strukture. Natrijum dodecilsulfat (SDS) je
deterdţent koji pokazuje polarne i nepolarne karakteristike. SDS se vezuje za najveći broj
proteina tako da su njegovi nepolarni hidrofobni dijelovi pohranjeni u nepolarni region
proteina, a negativno nabijeni sulfatni izloţen prema rastvaraču. Vezivanje SDS-a ima
dvojake efekte. Prvi su vezani za interferenciju sa nativnim hidrofobnim i jonskim vezama
što dovodi do disocijacije najvećeg broja oligomernih proteina u njihove monomere i do
razaranja njihove sekundarne strukture. Ukoliko su polipeptidni lanci povezani disulfidnim
vezama, potrebno je dodatno uzorke zagrijati u prisustvu disulfid reducirajućih agenasa
(betamerkaptoetanola ili 1,4-ditiotreitola). Da bi se to postiglo, potrebno je da protein
disocira na manje podjedinice. Ovaj drugi efekat se pojavljuje kada proteini postanu
zasićeni sa negativno nabijenim molekulama SDS-a. Tada elektroforetska separacija ovisi
isključivo od molekulske mase, tako monomeri malih molekulskih masa putuju znatno brţe
jer je koeficijent trenja manji. Elektroforeza koja se izvodi pod ovim uvjetima moţe da se
koristi za utvrĎivanje molekulske mase proteina.
Weber i Osborn su pokazali da se molekulske mase najvećeg broja proteina mogu
utvrditi putem mjerenja pokretljivosti na poliakrilamidnom gelu koji sadrţi SDS. Standardi
proteina poznatih molekulskih masa se analiziraju elektroforetski i njihova pokretljivost se
mjeri i nanosi na graf papir i izraţava u obliku logaritma molekulske mase. Ovaj grafikon
daje jednu pravu liniju. Pokretljivost nepoznatog uzorka se moţe odrediti pod identičnim
uslovima, a molekulska masa se moţe dobiti na osnovu grafika pokretljivosti standarda u
odnosu na njihove odgovarajuće logaritme molekulskih masa. Molekulske mase utvrĎene
metodom elektroforeze pod disocirajućim uslovima zadovoljavaju tačnost u rasponu od 5-
10 %.
SDS-PAGE primjena

BIOHEMIJA II–laboratorijske vjeţbe
25
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- GoveĎi serum
- Standard - albumin goveĎeg seruma (BSA)
- Agaroza
- glicin
- tris (hidroksimetil)-aminometan
- natrijum-dodecilsulfat (SDS)
- 2-merkaptoetanol
- glicerol
- brom-fenol plavo
Za analizu uzoraka koristiće se jednodimenzionalna agarozna gel elektroforeza.
Detekcija se vrši pomoću indikatora brom-fenol plavog, komazi briljantno plavog
(coomassie brilliant blue) ili bojenjem sa srebrom.
Intenzitet trake je veličina za relativno odreĎivanje količinskih dijelova neke komponente.
Na osnovu protoka referentnih proteina sa poznatom masom, mogu se odrediti molekulske
mase različitih proteina u uzorku i isti mogu biti identificirani. U slijedećoj tablici navedene
su relativne molekulske mase različitih proteina odreĎene na poliakrilamidnom gelu:
Protein
Mr PreĎena
udaljenost
(mm)
Transferin 78 000 6,0
Albumin goveĎeg seruma 66 000 12,5
Ovalbumin (albumin jajeta) 45 000 32,0
Gliceraldehid-3-fosfat dehidrogenaza 36 000 38,0
Protein
Mr PreĎena
udaljenost
(mm)
Karboanhidraza 29 000 50,0
Tripsinogen 24 000 54,0
Tripsin inhibitor soje 20 100 61,0
-laktoglobulin 18 400* 69,0
Mioglobin 17 800 69,0
Lizozim 14 300 79,0
Citohrom c 12 400 86,5 * -laktoglobulin ima Mr od 36 800 jer je dimer od dvije identične subjedinice sa Mr od 18 400. Pod
reducirajućim uvjetima pufera za uzorak disulfidne veze podjedinica su reducirane (pocijepane) tako da se na
gelu vide monomerni lanci.
1. Priprema potrebnih pufera
a) Elektroforetski pufer
Postoji više pogodnih elektroforetskih pufera za analizu proteina, ali je najčešće u upotrebi
TRIS-Glicin pufer koji se priprema na slijedeći način:
Pripremiti po 1 litar 0,192 mol/dm3 glicina i 0,025 mol/dm
3 tris (hidroksimetil)-
aminometana. Miješanjem tih komponenata dostići pH vrijednost 8,3.

BIOHEMIJA II–laboratorijske vjeţbe
26
b) Pufer za tretman uzorka
400 l elektroforetskog pufera + 100 l 2-merkaptoetanola + 100 l 2 % SDS-a u
elektroforetskom puferu.
c) Pufer za aplikaciju uzorka
300 l elektroforetskog pufera + 700 l 87 %-og glicerola + nekoliko kristalića brom-fenol
plavog ili komazi briljant plavog.
2. Priprema uzorka ili standarda
U kivetu za centrifugu odmjeriti 200 l uzorka ili standarda (pripremljenog tako da se 0,01
g albumina goveĎeg seruma otopi sa 1 ml elektroforetskog pufera), dodati 200 l pufera za
tretman uzorka, promiješati na vorteks miješalici i drţati na ključalom vodenom kupatilu 5
minuta.
3. Eksperimentalna procedura
a) Pripremiti 1% rastvor agaroze ukupnog volumena 150 mL (agarozu otopiti u
elektroforetskom puferu).
b) Miješati na magnetnoj miješalici uz grijanje dok se agaroza ne otopi.
c) Nakon otapanja, ostaviti otopinu na sobnoj temperaturi da se hladi uz miješanje na
drugoj magnetnoj miješalici. Dok je otopina još vrela popuniti s njom šupljine
izmeĎu ivica staklene ploče i okvira koristeći Pasterovu pipetu. Kada se ruka moţe
zadrţati na tikvici sa otopinom, tada dodati 0,1 g SDS-a, promiješati staklenim
štapićem da se otopi i sipati otopinu gela na staklenu ploču.
d) Odmah uroniti češalj u otopinu blizu jednog od krajeva ploče. Češalj podesiti tako
da donji dijelovi njegovih zubaca ne dodiruju površinu ploče (debljina gela na tim
mjestima treba da je 0,5 – 1,0 mm.
e) Nakon što proĎe 30 – 45 minuta paţljivo ukloniti češalj i okvir, te staviti ploču s
gelom u elektroforetsku jedinicu.
f) U udubljenja koja su napravili zupci češlja aplicirati uzorke koji su pripremljeni na
odgovarajući način (pomiješati 75 l pufera za aplikaciju uzorka sa 300 l uzorka ili
standarda).
g) U elektroforetske komore sipati elektroforetski pufer da dodiruje ivice gela.
h) Pokriti sistem sa poklopcem i priključiti kablove sa elektrodama koje su spojene sa
ispravljačem.
i) Uključiti ispravljač uz podešavanje odgovarajućeg napona (u početku dok uzorci ne
uĎu u gel napon moţe biti oko 50-60 V, a kasnije 80-100 V).
NAPOMENA
Prije rada upoznati se sa načinom rukovanja hemikalijama koje se koriste. Podaci o
štetnosti kemikalija, kao i načinu pruţanja prve pomoći nalaze se u laboratoriju
(konsultovati asistenta i laboranta)! Ovdje su navedene neke informacije za supstance
povećanog rizika:
- 2-Merkaptoetanol
Štetan ukoliko se proguta ili udiše. Toksičan u kontaktu sa koţom. Uzrokuje upaljenje.
Toksičan za vodene organizme, moţe uzrokovati dugotrajne štetne efekte u vodenoj
sredini! Pri radu sa ovom supstancom obavezno koristiti rukavice i rad obavljati u
digestoru!
- Natrijum-dodecilsulfat
Štetan ukoliko se proguta ili udiše. Iritira koţu, oči i respiratorni trakt. Moţe uzrokovati
alergiju koţe ili respiratornu reakciju (kašalj i sl.). Zapaljiv u čvrstom stanju!

BIOHEMIJA II–laboratorijske vjeţbe
27
Pri radu sa ovom supstancom obavezno koristiti rukavice!
3. ZADACI I VJEŽBE
1. Kakav je pravac kretanja peptida (ostaje na startu, kreće se ka anodi ili katodi) u
procesu eletroforeze pri pH 2; 3,5; 6,5; 10:
a) Lys-gly-ala-glu
b) Glu-gly-ala-glu
c) Gly-gly-ala-lys
2. Smjesa glicina, glutaminske kiseline, lizina, arginina i serina razdvaja se metodom
elektroforeze na papiru pri pH 6,0. Navedite koja su se jedinjenja kretala ka anodi, +
(A), katodi, - (K), a koja su ostala na startu C.
A:
B:
C:
3. Navedite pravac kretanja peptida lyz-gly-ala-gly u procesu elektroforeze na papiru
pri pH 1,9; 3,0; 6,5; 10,0.
4. Navesti u koje svrhe se moţe koristiti elektroforetska metoda?
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________

BIOHEMIJA II–laboratorijske vjeţbe
28
5. Koju ulogu u pripremi uzorka za elektroforezu imaju 2-merkaptoetanol i natrijum-
dodecilsulfat?
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________

BIOHEMIJA II–laboratorijske vjeţbe
29
4. REZULTATI
OVJERA VJEŽBE

BIOHEMIJA II–laboratorijske vjeţbe
30
Vježba broj 5
KVANTITATIVNO ODREĐIVANJE UKUPNIH PROTEINA BIURET
METODOM
1. TEORETSKI DIO
Proteini predstavljaju kompleksne polimere koji se sintetiziraju na nivou svih ţivih
bića. MeĎusobno se razlikuju po svojim funkcijama, veličini, obliku molekule te strukturi,
rastvorljivosti, sastavu, elektroforetskoj pokretljivosti. U odnosu na ulogu koju imaju u
organizmu, u principu se mogu podjeliti na proteine katalizatore (enzime), potom
regulatorne proteine (receptori, hormoni, represori, inhibitori), transportne proteine,
strukutrne proteine, proteine odbrane (imunoglobulini), onkofetalne i placentalne proteine i
proteine čija funkcija još nije poznata. Najstariji pristup odreĎivanja ukupnih proteina jeste
utvrĎivanje koncentracije azota proteinskog porijekla. Poznati postupak koji se koristi kod
odreĎivanja ovog tipa je Khjeldahl-ov postupak. Ovom metodom se utvrĎuje koncentracija
ukupnog azota u biološkom materijalu. Princip je da se supstance koje sadrţe azot pretvore
u amonijum jon posredstvom oksidacije u sistemu za razaranje koji sadrţi koncentrovanu
sulfatnu kiselinu kao katalizator i so koja ima funkciju da podigne tačku ključanja smjese.
Amonijum jon se najbolje analizira pretvorbom u amonijak sa dodatkom alkalija. Nakon
destilacije upari se u rastvor borne kiseline i amonijak potom titrira sa standardnim
rastvorom hlorovodonične kiseline. Korekcija za neproteinski azot se ostvari korištenjem
seruma koji ne sadrţi proteine. Metoda odreĎivanja po Khjeldahlu odlikuje visokom
tačnošću i preciznošću, činjenica je da traje dugo i da je dosta komplikovana sa aspekta
izvoĎenja rutinske analize serumskih proteina čak i u slučajevima kada se amonijak
odreĎuje enzimatskim analizama. Postoji takoĎer i nesigurnost oko tačnosti faktora
konverzije azota u proteine. Historijski se još uvijek koristi faktor 6,25 iako se zna da je
vrijednost istog relativno niska. Ova metoda se još uvijek koristi kao standardna metoda sa
kojom se uporeĎuju sve druge metode.
Jedna od najčešće korištenih metoda za utvrĎivanje koncentracije serumskih
proteina je zasnovana na principu Biuret reakcije. U ovoj reakciji bakarni jon reagira sa
peptidnim vezama proteina u osnovnom rastvoru gradeći ljubičasto obojene komplekse koji
pokazuju apsorpcioni maksimum na 540 nm. Biuretska reakcija predstavlja vrlo pogodnu
metodu i odlikuje se dobrom preciznošću (pozitivna bojena reakcija dobija se i pri
razblaţenju 1:104). Količina boje koju daje Biuret reagens u vrlo bliskom je odnosu sa
polipeptidnim dijelom različitih serumskih proteina što je direktna prednost nad Lowry-
evom metodom. Reakcija koristi jedan vrlo stabilan reagens i uslijed toga našla svoju
široku primjenu u automatizaciji. Nedostatak je što pozitivnu reakciju mogu dati i neke
supstance koje ne poseduju peptidnu vezu u molekuli, kao što su histidin, asparagin ili
oksamid.
Kingsley je 1942. godine koristio prvi reagens koji nije bio veoma stabilan. Prvi
stabilan Biuret reagens opisao je Weichselbaum koji je dodao kalij natrij tartarat kao
stabilizator i kalij jodid da bi spreiječio autoredukciju alkalnog bakar tartarata i separaciju
bakar oksida. Ovaj metod se zasniva na interakciji Cu2+
jona sa atomima nitrogena i
oksigena u peptidnoj vezi proteina u alkalnoj otopini, pri čemu nastaje ljubičasto obojeni
kompleks čija apsorpcija se mjeri na 545 nm.
Pored ove metode, najčešće korištene metode za kvantitativno odreĎivanje proteina su:

BIOHEMIJA II–laboratorijske vjeţbe
31
1. Bredford-ova metoda: zasniva se na mjerenju apsorpcije kompleksa proteina sa
Bredfordovim reagensom pri 595 nm. Bredfordov reagens je Coomassie briljantno plavo G
u fosfatnoj kiselini i metanolu. Linearno područje koncentracija proteina: 0,1-1,4 mg/ml.
Ukoliko u uzorku ima tragova deterdţenata prikladnija je BCA metoda koja će biti opisana.
2. Lowry-jeva metoda: zasniva se na redukciji fosfomolibden-volfram mješovitog
kiselog kromogena u fenolskom Folin-Ciocalteu reagensu od strane proteina, što rezultira u
apsorpcionom maksimumu pri 750 nm. Nedostatak je što se na rezultat moţe negativno
odraziti prisustvo ostalih reducirajućih supstanci i što reagens reagira samo sa proteinima
koji sadrţe tirozin.
3. Metoda sa bicinkoninskom kiselinom (BCA metod) zasniva se na redukciji
kupri (Cu2+
) jona do kupro (Cu+) jona proteinom. Bicinkoninska kiselina se koristi za
detekciju Cu+. Ova metoda ima nekoliko interferirajućih supstanci čiji efekat moţe biti u
izvjesnoj mjeri umanjen razrijeĎivanjem ispitivane otopine proteina.
4. Fluorimetrijski metod zasniva se na derivatizaciji proteina sa o-ftalaldehidom
(OPA), koji reagira sa primarnim aminima proteina (N-terminalne aminokiseline i -NH2
grupa lizina). Osjetljivost se moţe povećati prethodnom hidrolizom proteinskog uzorka.
5. Metoda mjerenja refraktivnog indeksa zasniva se na mjerenju prelamanja
ulazne svjetlosti na smjesu uzorka, ali za serum, to će u principu odraţavati masu prisutnih
proteina. Obzirom da serum sadrţi i znatnu količinu ostalih tvari, refraktometar se mora
specifično kalibrirati sa serumom poznate koncentracije proteina.
Vrlo vaţni koraci na nivou standardizacije prilikom odreĎivanja ukupnih serumskih
proteina su poduzeti osamdesetih godina prošlog stoljeća. U tom periodu univerzalno se
prihvatilo korištenje goveĎeg serumskog albumina kao referentnog materijala koji će se
koristiti za odreĎivanje ukupne koncentracije proteina u spektrofotometrijskim metodama
poput biuretske i Lowry-eve metode. On se preporučuje kod svih odreĎivanja koja za
princip imaju biuretsku reakciju.

BIOHEMIJA II–laboratorijske vjeţbe
32
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- 200,00 g 0, 9% vodene otopine NaCl
- 100,00 g 6% vodene otopine NaOH
- 200,00 mL Biuret reagensa koji se priprema na slijedeći način: 3,46 g bakar (II)-sulfata
otopiti u 10,00 ml vruće vode i ostaviti da se ohladi (otopina A). Otopiti 34,6 g
natrijumcitratdihidrata i 20,0 g natrijumkarbonata u 80,0 ml vruće vode i ohladiti (otopina
B). Pomiješati otopine A i B i razrijediti sa vodom do 200,0 ml. Ovakav Biuret reagens
stabilan je pri sobnoj temperaturi 6 mjeseci. Ukoliko se pojavi zamućenje ili talog ne
koristiti takav reagens.
- 0,2% NaOH u 50% etanolu (za uzorke pšenice, raţi, ječma, zobi)
Priprema slijepe probe:
Na 1 ml otopine NaOH dodati 1 ml Biuret reagensa i sadrţaj protresti. Na to dodati 0,4 ml
otopine NaCl i promiješati.
Priprema i upotreba standardne otopine proteina:
Odvagati 0,0500 g albumina goveĎeg seruma (BSA) (liofilizirani prah, nabavljen od
Sigma
) i otopiti u 5,00 ml 0,9 % NaCl. Dobijenu otopinu izraziti u mg/mL, označiti je kao
osnovnu otopinu, te od nje prirediti po 10,00 mL slijedećih radnih otopina: 7.5, 5.0, 2.5, 1.0
i 0.5 mg/ml.
Izmjeriti apsorbansu svih otopina (uključujući i osnovnu) pri 545 nm koristeći se slijedećim
postupkom:
U čiste i označene epruvete (pripremiti istovremeno za standarde, uzorke i slijepu probu)
sipati po 1 ml otopine 6% rastvora NaOH, dodati u svaku 1 ml Biuret reagensa i sadrţaj
protresti. Na to dodati 0,4 ml otopine standarda (ili uzorka) zatvoriti i promiješati. Nakon
15 minuta sadrţaj istresti u kivetu, obrisati kivetu i mjeriti apsorbansu. Konstruirati
kalibracionu krivu A = f (c), nanoseći na apscisu konačne koncentracije standardnih
otopina.
Priprema i rad sa uzorkom proteina
UZORAK 1: Odvagati pribliţno 2 g sirovog bjelanca (sa preciznošću na četiri decimalne
cifre) u prethodno odvaganoj čistoj i suhoj čašici. Pomoću 0,9 % otopine NaCl prebaciti
kvantitativno sadrţaj bjelanca u odmjerni sud od 50 ml. Izmjeriti apsorbansu tako
pripremljenog uzorka pri istim uvjetima korištenim za rad sa standardnom otopinom BSA.
Pomoću jednačine pravca dobijene iz kalibracione krive izračunati koncentraciju ukupnih
proteina u uzorku bjelanca izrazivši konačan rezultat kao mg proteina/g svjeţeg bjelanca.
UZORAK 2: Odmjeriti ispitivanog uzorka po 1-1,5 g materijala koji je prethodno usitnjem
ili izmljeven u staviti u odgovarajuće posebne porculanske avane. Probama dodati 2 g
kvarcnog pijeska i dodati po 3 mL NaOH u alkoholu, a zatim paţljivo rastrljavati uzorak u
trajanju od tri minute kako bi se razorile ćelije i ekstrahovali proteini. Posle isteka 3 minute
dodati u avan još 12 mL rastvarača i ponovo homogenizirati u trajanju od dvije minute.
Tako dobijenu smjesu kvantitativno prebaciti u odmjerni sud od 50 mL; avan 2-4 puta

BIOHEMIJA II–laboratorijske vjeţbe
33
isprati sa po 5 mL rastvarača i prenijeti u odmjerni sud. Odmjerni sud dopuniti sa
rastvaračem do mjerne crte, dobro promućkati i ostaviti sa stoji 1 sat. Sadrţaj profiltrirati u
suhu erlenmajericu od 50 mL. 10 mL takvog filtrata staviti u kivete za centrifugiranje i
dodati po 1 mL 6% NaOH i po 1 mL Biuret reagensa. Smjesu u epruvetama dobro
promućkati i centrifugirati u trajanju pd 5 minuta. Ako rastvor nije proziran, onda je
potrebno da se eksperiment ponovi uz dodavanje po 2 mL baze i reagensa. Poslije
centrifugiranja, rastvor mora biti bistar (proziran).
3. ZADACI I VJEŽBE
1. Prikazati mehanizam Biuret reakcije na primjeru uree?

BIOHEMIJA II–laboratorijske vjeţbe
34
4. REZULTATI
Standard/
Uzorak
Apsorpcija
(A)
Sadrţaj ukupnih protiena (rezultat kao mg proteina/g uzorka)
- Napomena: Priloţiti grafik
OVJERA VJEŽBE:

BIOHEMIJA II–laboratorijske vjeţbe
35
Vježba br. 6
ODREĐIVANJE KONCENTRACIJE ALBUMINA I GLOBULINA
1. TEORETSKI DIO
Prečišćavanje proteina je osnovni korak u izučavanju njihovih fizičkih i bioloških
svojstava i jedan je od najčešćih postupaka u praktičnoj biohemiji. Za postizanje
prečišćavanja protein mora biti osloboĎen od svog biološkog matriksa i selektivno odvojen
od ostalih proteina odgovarajućim postupkom frakcioniranja. Predmeti izučavanja na
prečišćenim proteinima odreĎuju prihvatljivost granica čistoće koju je potrebno dostići.
Tako, ispitivanja aktivnosti ne zahtijevaju 100% čist preparat, dok osnovna strukturna
izučavanja zahtijevaju. Suprotno, ispitivanja aktivnosti zahtijevaju očuvanje funkcije i stoga
smanjenje denaturacije i proteolize, dok strukturna ispitivanja se mogu izvesti na
denaturiranom proteinu. Količina prečišćenog proteina potrebna za izučavanje objektivno
utiče na izbor metoda prečišćavanja. Neke tehnike imaju visok kapacitet i mogu raditi sa
velikim volumenima i koncentracijama proteina, dok druge imaju ograničen kapacitet.
Proteini se razlikuju u njihovoj osjetljivosti na denaturaciju tokom procesa
ekstrakcije i prečišćavanja, posebno u njihovoj osjetljivosti na povišene temperature (iznad
40 C), na prisutnost deterdţenata, teških metala i na ekstremne pH vrijednosti. Ove razlike
često su iskorištene u strategiji prečišćavanja pojedinih proteina i tehnika zasnovanih na
njihovom zajedničkom svojstvu u ranim fazama procesa prečišćavanja. Topivost prirodnih
proteina uvjetovana je pH vrijednošću (topivost postaje minimalna u izoelektričnoj tački),
dodatkom soli kao što su amonijumsulfat i prisutnošću organskih otapala, kao što su aceton
i butanol, pri niskim temperaturama. Male razlike u ovakvim svojstvima enzima takoĎer su
iskorištene u postupcima frakcioniranja i prečišćavanja. Spojevi koji sadrţe tiolnu grupu
kao što su merkaptoetanol i glutation (redukovani oblik) često se dodaju enzimskim
preparatima da spriječe oksidaciju sulfhidrilnih grupa, koja se moţe desiti odmah nakon
razaranja stanice i prethodne denaturacije.
Tokom odvijanja prečišćavanja proteina često se prave razrijeĎene otopine proteina.
Naţalost, razrijeĎene otopine proteina, posebno prečišćenih proteina, često su nestabilne ili
zbog disocijacije subjedinica ili adsorpcije proteina na površini posude. Problemi adsorpcije
mogu biti smanjeni upotrebom silikoniziranih posuda uz dodatak malih koncentracija
deterdţenata kao što su Triton X-100 ili Tween 20. Problemi disocijacije mogu često biti
prevaziĎeni dodatkom glicerola (10% do 40% (v/v)), glukoze ili saharoze ili povremeno
albumina goveĎeg seruma (bovine serum albumin, BSA). Čuvanje uzoraka pri 0-4 C je
takoĎer korisno. Duţe čuvanje moţe biti omogućeno upotrebom visokih koncentracija
amonijsulfata (to objašnjava zašto su trgovački preparati enzima često spremljeni u otopini
amonijsulfata).
Prečišćene proteine moguće je frakcionirati na različite načine:
1. Frakcioniranje denaturacijom koristi razlike u toplotnoj osjetljivosti proteina.
Kada je poznata temperatura denaturiranja, moguće je ukloniti više termolabilnih
kontaminirajućih proteina grijanjem smjese do temperature koja je 5-10 C niţa od
ove kritične temperature u trajanju od 15-30 minuta. Denaturirani, neţeljeni protein
se potom ukloni centrifugiranjem. Prisustvo supstrata, produkta ili inhibitora enzima
često stabilizira enzim i omogućava da se moţe primijeniti čak i viša temperatura
toplotne denaturacije. Na sličan način, proteini se razlikuju u lakoći denaturiranja
pri ekstremnim pH vrijednostima ( 3 ili 10 ). Kompletan proteinski ekstrakt se

BIOHEMIJA II–laboratorijske vjeţbe
36
dovede na pH ne manji od jedne pH jedinice od pH unutar kojeg se ispitivani
protein taloţi. Osjetljiviji proteini će se istaloţiti i ukloniti centrifugiranjem.
2. Frakcioniranje solima izvodi se postepenim dodavanjem odgovarajuće soli. U
praksi amonijsulfat je najčešće korišten jer je dobro topiv u vodi, moţe se dobiti u
visokom stepenu čistoće, jeftin je i nema štetnog uticaja na strukturu proteina.
Nakon svakog dodatka soli mora se osigurati potpuno otapanje soli i nastajanje
homogenog otapala. Istaloţeni protein se odcentrifugira, otopi u svjeţem puferu i
ispita u odnosu na ukupni protein i proteinsku aktivnost. Sve faze se izvode pri 0-10
C radi smanjenja denaturacije. Dati protein se normalno isoli u okviru uskog
područja koncentracije amonijsulfata. To je posljedica činjenice da protein-protein
udruţivanje za dati protein naglo postaje dominantno nad protein-voda i protein-so
interakcijama. To je zbog toga što dodatak soli uklanja sloj molekula vode koji
okruţuje hidrofobne grupe na površini proteina, što omogućava hidrofobnim
grupama da uzrokuju udruţivanje proteina i stoga taloţenje.
3. Frakcioniranje organskim otapalom zasniva se na razlikama u topivosti proteina
u vodenim otopima organskih otapala kao što su etanol, aceton i butanol. Organsko
otapalo sniţava dielektričnu konstantu medija putem povećanja privlačnosti izmeĎu
nabijenih molekula proteina i smanjenja njihove interakcije sa vodom. Topivost
proteina se stoga smanjuje. Frakcioniranje organskim otapalom u biti je suprotno
procesu obuhvaćenim frakcioniranjem solima. Kod prvog procesa, hidrofobne grupe
proteina su znatno zaštićene molekulama organskog otapala, a jonske grupe postaju
dominantne, dok kod drugog procesa hidrofobne grupe su jako izloţene.
4. Frakcioniranje organskim polimerom slično je frakcioniranju organskim
otapalom u mehanizmu djelovanja, ali zahtijeva niţe koncentracije za izazivanje
taloţenja proteina. Najčešće korišteni polimer je polietilenglikol (PEG) sa
relativnom molekulskom masom u području 6 000 - 20 000.
5. Izoelektriĉno frakcioniranje (frakcioniranje taloţenjem u izoelektričnoj tački)
zasnovano je na tome da proteini imaju minimalnu topivost u izoelektričnoj tački.
Pri ovom pH postoji jednak broj pozitivnih i negativnih naboja proteinske molekule
i intermolekulska odbijanja su minimalna, a intermolekulska privlačenja
maksimalna, što rezultira u nastajanju netopivih agregata. Osnovno što se moţe
iskoristiti je ili uklanjanje neţeljenog proteina, podešavanjem pH ekstrakta proteina
tako da se izazove taloţenje ovih proteina, ali ne i ispitivanog proteina, ili
uklanjanje ispitivanog proteina, podešavanjem pH ekstrakta na pHi tog proteina.
Centrifugiranje je moćna i opće primjenjiva metoda za razdvajanje i analizu stanica,
organela i bioloških makromolekula. Čestica koja se kreće kruţno sa poluprečnikom r,
ugaonom brzinom , izloţena je centrifugalnom polju koje iznosi 2r.
Brzina taloţenja tj. brzina sedimentacije čestice ovisi o nekoliko činioca:
o masi čestice – teţe čestice brţe se taloţe od onih lakših;
o obliku čestice – koeficijent trenja kompaktne čestice manji je nego u slučaju
izduţene čestice iste mase, što znači da će se kompaktnije čestice brţe taloţiti od
izduţenih;
o gustoći čestice – gušće čestice se kreću mnogo brţe od onih manje gustoće;
o gustoći otopine u kojoj se čestica nalazi.
Krvna plazma je svijetlo-ţuta tečnost, a najveći dio nje čini voda (oko 90%) u kojoj su
otopljene različite organske i anorganske tvari, oko 7% proteina, 1% raznih anorganskih

BIOHEMIJA II–laboratorijske vjeţbe
37
soli i 0,1% glukoze. MeĎu organskim tvarima najvaţnije su proteini plazme. U 1 litri
plazme ima ih oko 73 grama.
Te proteine moţemo svrstati u 3 velike grupe:
- Albumini – 45 g/l – to su proteini plazme s najmanjom molekulskom masom, te su vrlo
prikladni za odrţavanje osmotskog tlaka. Osmotski tlak je vrlo vaţan, jer odrţava protuteţu
hidrostatskom tlaku krvi koji stvara srce. Albumini se sintetiziraju u jetri i odatle se
otpuštaju u krv. Sudjeluju u prijenosu raznih hormona, različitih jona, aminokiselina,
masnih kiselina, lijekova itd.
- Globulini – 25 g/l – vaţni su nositelji odbrane organizma u obliku različitih antitijela.
Dijele se na alfa, beta i gama globuline. Uloga alfa i beta globulina je slična ulozi albumina,
a gama globulini su zapravo imunoglobulini (Ig), a to su molekule koje obavljaju zaštitne
funkcije.
- Fibrinogeni – 3 g/l – vrlo vaţni proteini u procesu zgrušavanja krvi, te osiguravaju
zaustavljanje krvarenja. Fibrinogeni pomoću trombocita stvaraju «mreţu» u kojoj se onda
ulove eritrociti i nastaje ugrušak (tromb). Krv bez fibrinogena je defibrinirana krv.

BIOHEMIJA II–laboratorijske vjeţbe
38
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- 0, 9 % vodena otopina NaCl
- 2, 0 % otopina trihloracetatne kiseline u 0,9 %-om NaCl
- Biuret reagens
- 6,0 % vodena otopina NaOH
U uzorku goveĎeg seruma izmjeriti koncentraciju ukupnih proteina koristeći se Biuret
testom (100 l goveĎeg seruma razrijediti sa 900 l 0,9 % -ne otopine NaCl, a za daljnju
proceduru vidjeti vjeţbu „Kvantitativno odreĎivanje ukupnih proteina Biuret metodom“).
Podatak zabiljeţiti u radni dnevnik i preračunati na sadrţaj proteina izraţen u mg/100 ml
seruma.
Nakon toga u dvije ependorf epruvete (centrifugalne kivete) odmjeriti po 100 l uzorka
goveĎeg seruma i na to dodati po 900 l 2,0 % otopine trihloracetatne kiseline. Propisno
zatvoriti kivete, okrenuti ih nekoliko puta da se sadrţaj izmiješa i ostaviti ih 10 minuta da se
izvrši taloţenje. Za to vrijeme podesiti centrifugu na 3 500 o/min i centrifugiranjem u
trajanju 5 minuta oboriti globuline na dno kiveta. U supernatantima odrediti sadrţaj
albumina Biuret testom. Iz razlike koncentracija ukupnih proteina i albumina izračunati
koncentraciju globulina.
3. ZADACI I VJEŽBE
1. Glavne osobine albumina (A), globulina (B) i prolamina (C) su:
a) Nerastvorljivi u vodi, rastvorljivi u 70-80% alkoholu
b) Dobro rastvorljivi u vodi
c) Nerastvorljivi u vodi i u rastvorima soli umjerene koncentracije

BIOHEMIJA II–laboratorijske vjeţbe
39
4. REZULTATI
Standard/
Uzorak
Apsorpcija
(A)
Sadrţaj ukupnih proteina, albumina i globulina (rezultat izraziti kao mg/100 ml seruma)
- Napomena: Priloţiti grafik
OVJERA VJEŽBE:

BIOHEMIJA II–laboratorijske vjeţbe
40
Vježba broj 7
ENZIMI
1. TEORETSKI DIO
Enzimi su globularni proteini tercijarne ili kvarterne strukture, koji na površini
imaju većinu polarnih aminokiselinskih ostataka, dok su nepolarni ostaci okrenuti prema
unutrašnjosti molekule. Ovako organizirane proteinske molekule nazvane enzimi imaju
visoku katalitičku moć i specifičnost za pojedine hemijske reakcije i supstrate u ţivim
organizmima. Katalitička moć enzima se manifestira njegovom sposobnošću da smanjuje
energiju aktivacije potrebnu za odvijanje hemijskih reakcija. Enzimi ubrzavaju reakcije i
vrlo često kataliziraju samo jednu hemijsku reakciju ili skup vrlo srodnih reakcija, a
specifičnost za supstrat je najčešće potpuna.
U trenutku katalitičke promjene supstrat je s enzimom povezan uglavnom slabim
nekovalentnim vezama (vodikove, hidrofobne ili Van der Waalsove veze), što omogućava
brzu izmjenu molekula supstrata na enzimu. Ta izmjena se dogaĎa u jednom dijelu enzima
nazvanom aktivno mjesto. Aktivno mjesto predstavlja mali broj aminokiselina smještenih u
unutrašnjosti u hidrofobnom dijelu proteinske molekule, čije prostorno ureĎenje odgovara
molekuli supstrata. Taj prostor u hidrofobnoj unutrašnjosti je odgovoran za njegovu
katalitičku moć.
Internacionalnom konvencijom enzim se svrstava u jednu od šest vrsta na osnovu
hemijske reakcije koju katalizira. Svaka vrsta se dijeli prema prirodi hemijske grupe,
koenzima i drugih grupa uključenih u reakciju. Saglasno pravilima Evropske komisije (EC),
svaki enzim moţe se označiti jedinstvenim kodom od četiri broja i nedvosmislenim
sistematskim imenom zasnovanim prema kataliziranoj reakciji.
Šest vrsta enzima su:
Oksidoreduktaze, koje prenose atome hidrogena, oksigena ili elektrone od jednog
substrata na drugi;

BIOHEMIJA II–laboratorijske vjeţbe
41
Transferaze, koje prenose hemijske grupe izmeĎu substrata;
Hidrolaze, koje kataliziraju hidrolitičke reakcije;
Liaze, koje razlaţu substrate reakcijama različitim od hidrolize;
Izomeraze, koje pretvaraju izomere jedan u drugi intermolekularnim
premještanjem;
Ligaze (sintetaze), koje kataliziraju nastajanje kovalentne veze, uz cijepanje
odgovarajućeg nukleotid-trifosfata.
Jedinica za enzimsku aktivnost
Jedna enzimska jedinica definira se kao količina enzima koja pri optimalnim uvjetima (25
°C, 1 bar), razloţi 1 μmol supstrata za jednu minutu. Specifična aktivnost enzima moţe se
izraziti kao broj enzimskih jedinica po miligramu proteina.
Nova internacionalna jedinica (IU) odreĎena od strane EC je katal (kat). Definira se kao
količina enzima koja pri optimalnim uvjetima (25 °C, 1 bar) razloţi 1 mol supstrata za
jednu sekundu. Iz praktičnih razloga u upotrebi su manje jedinice kao mikrokatal (μkat),
pikokatal (pkat), nanokatal (nkat) itd.
Amilaza spada u grupu hidrolaza (karbohidraze), jer kataliziraju hidrolizu oligo- i
polisaharida. Hidroliza se vrši razlaganjem glikozidnih veza izmeĎu monosaharida, koji
izgraĎuju oligo- i polisaharide. α-amilaza se nalazi u svim organima čovjeka i drugih sisara
(ubikvitaran enzim), a najveća aktivnost je u pljuvački (gdje se naziva ptijalin) i u
pankreasnom soku (gdje se naziva dijastaza). Specifična je za α1-4 glikozidnu vezu u
polisaharidima (škrob i glikogen). Pošto ne moţe da cijepa α1-6 glikozidnu vezu, razlaţe se
oko 80% molekule škroba tj. potpuno hidrolizira amilozu i amilopektin. Krajnji produkti
koji nastaju hidrolizom škroba ovim enzimom su maltoza i granični dekstrini koji nastaju
na mjestima grananja u amilopektinu, tj. gdje se nalazi α1-6 glikozidna veza. Zato se α-
amilaza još naziva i endoamilaza (unutrašnja) ili dekstrinogena amilaza. Za razliku od α-
amilaze, β-amilaza je biljnog porijekla. Specifična je, takoĎer, za α1-4 glikozidnu vezu, ali
djeluje na krajevima polisaharidnih lanaca. Zato ima naziv-egzoamilaza (spoljašnja) i krajni
produkti njenog djelovanja su molekule maltoze.
Pepsin je takoĎer enzim iz grupe hidrolaza (III grupa enzima) iz grupe peptidhidrolaza. To
je proteolitički enzim koji cijepa peptidne veze u denaturiranim proteinima u ţelucu. Kada
se proteini u ţelucu denaturiraju hloridnom kiselinom iz ţelučanog soka, dobiju naziv acid
albumini (acid proteini). Pepsin prvenstveno cijepa peptidne veze izmeĎu aromatskih i
dikarboksilnih aminokiselina tako da nastaju, kao produkti, duţi ili kraći peptidni lanci.
Kraći lanci se nazivaju peptoni, a duţi albumoze. Smjesa peptona i albumoza se koristi kao
podloga za bakterije. Pepsin luče glavne ćelije ţelučane sluznice u obliku proenzima-
pepsinogena koji je neaktivan što se tiče proteolize. Pepsinogen se pri pH ispod 3
reverzibilno aktivira promjenom konformacije, a zatim autokatalizom odcjepljuje dva
peptida sa N-terminalnog kraja, prelazeći ireverzibilno u pepsin.
Schardingerov enzim katalizira oskidaciju aldehida i otuda se naziva aldehid-dehidrogenaza
Schardingera (aldehid: O2-oksidoreduktaza 1.2.3.1.). Pored toga, ovaj enzim ubrzava
oksidaciju hipoksantina i ksantina u mokraćnu kiselinu. Zahvaljujući drugoj reakciji koju
katalizira, dobio je naziv ksantin-oksidaza (ksantin: O2-oksido-reduktaza-1.2.3.2). Dugo se
mislilo da su to dva odvojena enzima. Svijeţe mlijeko sadrţi dosta Schardingerovog
enzima. Jetra čovjeka takoĎer je bogata ovim ezimom. Schardingerov enzim oksidira
podjednako efikasno i aromatske i alifatske aldehide. Da bi se ostvarila oksidacija (putem
oduzimanja hidrogena) in vitro, potrebno je reakcionoj smjesi dodati supstancu-akceptor

BIOHEMIJA II–laboratorijske vjeţbe
42
hidrogena. U tu svrhu moţe da posluţi metilensko plavo, jer ono prima hidrogen do
oksidiranog supstrata i odmah se obezboji-prelazi u leuko formu. Ako duţe stoji,
metilensko plavo moţe ponovo da se oboji, jer lahko predaje hidrogen oksigenu iz zraka. In
vivo, hidrogen oduzet od supstrata predaje se direktno oksigenu, ili preko NAD-a, ako je
hipoksantin supstrat oksidacije. Schardingerov enzim spada u aerobne dehidrogenaze u oba
slučaja: i kada djeluje na aldehide i kada djeluje na purine. Ako upotrebljavamo mlijeko
kao izvor enzima, neophodno je da bude svjeţe. Temperatura ključanja brzo razara enzim.
2. EKSPERIMENTALNI DIO
Specifiĉnosti enzima
Specifičnost enzimske aktivnosti moţe se jednostavno promjeriti sa enzimima amilazom i
saharazom, od koji je prva specifična za škrob, koga hidrolizom škrob prevodi u maltozu, a
druga vrši hidrolitičko razlaganje saharaze do fruktoze i glukoze.
- 2 % ratvor saharoze
- 1% rastvor škroba u 0,3% rastvoru NaCl
- Rastvor saharaze (100 g suhog kvasca usitniti u avanu do praha, preliti sa 450-500 mL
vode, ekstrahovati u toku 1,5-2 sata, zatim filtrirati polako (obično preko noći) i dobiveni
filtrat čuvati u boci na hladnom)
- Amilaza
- 10% NaOH
- 5% rastvor CuSO4
- Vodeno kupatilo
Specifičnost amilaze. U jednu epruvetu dodati 5 mL rastvora škroba, a u drugu 5 mL
saharoze. Dodati u svaku epruvetu po 1-1,5 mL rastvora amilaze, pomješati i uroniti
epruvetu u vodeno kupatilo, čija je temperatura 38-40°C. Posle isteka od 8-10 minuta
sadrţaj obje epruvete ispitati na prisustvo redukujućih šećera (maltoza iz škroba, glukoza i
fruktoza iz saharoze) pomoću Fehling-ove reakcije. Epruvete sa škrobom će dati pozitivnu
reakciju, jer je nastala maltoza koja ima redukujuća svojsta, pa redukuje rastvor Fehling-a
do Cu2O (crvena boja). U drugoj epruveti pozitivna reakcija izostaje jer je saharoza
neredukujući šećer.
Specifičnost saharaze. Postupak je isti kao u prvom slučaju, samo se umjesto amilaze
dodaje rastvor saharaze. Posle inkubacije u trajanju od 15-20 minuta izvrši se testiranje obje
epruvete pomoću Fehling-ovo reagensa. Epruveta koja je sadrţavaja saharozu i saharazu će
dati pozitivnu reakciju uslijed reakcije izmeĎu reagensa i produkata hidrolize saharoze
(glukoze i fruktoze).

BIOHEMIJA II–laboratorijske vjeţbe
43
Dokazivanje aktivnosti i termolabilnosti ptijalina (α-amilaze iz pljuvaĉke)
Sakupiti 2-3 ml pljuvačke, podijeliti na dva dijela. Jedan dio proključati na plameniku ili
rešou. U tri epruveta pripremiti smjese prema datoj tabeli:
Epruveta I II III
Škrob
2-3 ml 2-3 ml 2-3 ml
Neprokuhana
pljuvačka
Nekoliko
kapi
Prokuhana
pljuvačka
Nekoliko
kapi
Lugolov reagens
2-3 kapi 2-3 kapi 2-3 kapi
Sve epruvete staviti u vodeno kupatilo na 37 ºC na inkubiranje od 30 minuta. Nakon tog
vremena izvaditi epruvete i zaključiti šta se desilo u epruvetama.
Izvesti Fehling-ovu reakciju na sadrţaje u drugoj i trećoj epruveti. U svaku dodati po 1-2 ml
pripremljenog Fehlingovog reagensa i zagrijavati do ključanja. Zaključiti šta se desilo u
ovim epruvetama.
Optimalni pH za djelovanje amilaze
Postupak: U četiri epruvete dodati po 5ml fosfatnog pufera pripremljenog na slijedeći
način:
Epruveta 1
pH=5,59 Epruveta 2
pH=6,64 Epruveta 3
pH=7,17 Epruveta 4
pH=8,04
Na2HPO4 (0,067 M) 0,25 mL 2,00 mL 3,00 mL 4,75 mL
KH2PO4 (0,067 M) 4,75 mL 3,00 mL 2,00 mL 0,25 mL
0,5% škrob 1 mL 1 mL 1 mL 1 mL
Amilaza (1:15) 1 mL 1 mL 1 mL 1 mL
Rastvore Na2HPO4 (0,067 M) i KH2PO4 (0,067 M) paţljivo sipati iz birete i dobro
izmješati. U svaku epruvetu dodati 0,5 % rastvor škroba i razrijeĎene amilaze. Sadrţaj
promješati i staviti u vodeno kupatilo. Ostaviti 10-15 min na 40°C. Epruvete izvaditi,
ohladiti i u svaku dodati po 3-4 kapi joda u kaloj-jodidu (Lugolov reagens). Prema boji joda
moţe se orediti stepen razgradnje škroba u zavisnosti od pH sredine. Epruveta u kojoj ne
nastane plava boja sa micelama škrba imala je optimalan pH za djelovanje amizale.
Intezivna plava boja indicira neakrivnost amilaze (izvan dometa pH za djelovanje amilaze),
dok blijedoplava boja indicira aktivnost u sredini udaljenoj od optimalnog pH, ali još uvijek
u domenu djelovanja pluvačne amilaze.
U domenu optimalnog pH za djelovanje amilaze (u epruveti u kojoj nema plave boje sa
Lugolovim reagensom) škrob se razlaţe na sastojke koji ne daju plavu boju sa jodom, uz
povaju pozitivne redukujuće reakcije.
U domenu djelovanja enzima izvan optimalnog pH (niţi ili viši pH) škrob se djelimično
razgraĎuje, a za potpunu razgradnju treba više vremena, pa je i rekacija joda sa preostalim
škkrobom manje intezivna. Izvan domena djelovanja enzima, pri pH izrazitije udaljenom od

BIOHEMIJA II–laboratorijske vjeţbe
44
optimalnog pH, nema promjena na škrobu, ostaje nerazgraĎen, i reakcija sa jodom je
maksimalnog inteziteta.
Prikaţi grafički djelovanje enzima u zavisnosti od pH sredine.
Inhibitorsko djelovanje hlorida na dehidrogenazu
U krompiru se nalazi dehidrogenaza, čije se djelovanje na zraku zapaţa tamnjenjem
razrezane površine krompira. Hloridni joni inhibiraju djelovanje dehidrogenaze i izostaje
efekat tamnjenja (oksidacije) na površini razrezanog krompira.
Postupak: Jedan krompir razrezati na polovine i označiti ih brojevima 1 i 2.
Polovina br. 1: Kontrola
Polovina br. 2: Posuti kristalima natrij-hlorida.
Stajanjem na zraku uočite razliku izmeĎu polovica krompira.
Ureaza
Ureaza procesom hidrolize razlaţe ureu na amonijak i ugljendioksid.
(NH2)2CO + H2O → CO2 + 2NH3
U rastvoru osloboĎeni amonijak se bolje razlaţe, nastaje bazna sredina, pri kojoj
fenolftalein prelazi u crvenu boju.
Postupak: Na oko 1g uree dodati 5 mL destilovane vode. U dvije epruvete nasuti po 2 mL
rastvora uree i po 3 kapi fenolftaleina. U prvu epruvetu na vrhu kašičice dodati malo
sojinog brašna (koje sadrţi enzim ureazu) i obje epruvete drţati 30 minuta na 38°C. U prvoj
epruveti će se pojaviti crvena boja, a u drugoj neće.
Pepsin
Treba pripremiti kuhano bjelance kokošijeg jajeta isjeckano na kockice koje će posluţiti
kao supstrat. Uzeti četiri epruvete i dodati u njih:
Epruveta Bjelance Pepsin Prokuhani pepsin 0,2% HCl 1% Na2CO3
I 2-3 kockice 2-3 kapi 5 mL
II 2-3 kockice 5 mL
III 2-3 kockice 2-3 kapi 5 mL
IV 2-3 kockice 2-3 kapi 5 mL
Sve četiri epruvete staviti u vodeno kupatilo, na 37 °C i inkubirati 30 minuta. Pripremiti
nove četiri epruvete i nakon inkubacije sadrţaje epruveta dekantirati u nove pripremljene
epruvete. U izdvojene filtrate dodati 1-2 ml Biuret reagensa. Uočiti gdje se desila Biuret
reakcija.
Dehidrogenaza mlijeka
Ovaj enzim sadrţi mlijeko, a funkcija mu je oksidacija nekih aldehida u odgovarajuće
kiseline. Reakcija se odvija u anaerobnim ulovima i tada neko organsko jedinjenje sluţi kao
primalac vodika. Bojeni indikator oksido-redukcije je jedinjenje koje gubi boju primajući
vodik (nastaje redukovani derivat). Često se u ogledima koristi metilensko-plavo:

BIOHEMIJA II–laboratorijske vjeţbe
45
MP + 2H+ → MPH2
Nastali derivat je autoksidabilan, pa se na vazduhu prevodi u svoj oksidovani oblik, pa
treba izbjegavati duţe izlaganje zraku.
MPH2 + O2 → MP + H2O2
Postupak: U tri epruvete staviti:
- epruveta 1: 5 mL svijeţeg, nekuhanog mlijeka + 0,5 mL metilenplavog (0,05%)
- epruveta 2: 5 mL svijeţeg, nekuhanog mlijeka + 0,5 mL rastvora metilenplavog
(0,05%) + 1 mL 0,4% rastvora formaldehida
- epruveta 3: 5 mL svijeţeg, kuhanog mlijeka + 0,5 mL rastvora metilenplavog
(0,05%) + 1 mL 0,4% rastvora formaldehida
Svaku epruvetu treba dobro promješati (izbjeći, po mogućnosti, veća izlaganja zraku), pa
staviti u vodeno kupatilo na 42°C. Zapaziti da se u epruveti br. 2 metilenplavo dekoloriše,
jer prima vodik iz aldehid-hidrata i prelazi u leukoderivat:
Napomena: Uslijed neizbjeţnog kontakta sa kisikom iz zraka i na površini epruvete br. 2
moţe se zadrţati nešto plave boje (autooksidacija redukovanog enzima).
Dehidrogenaza djeluje i na ksankin i hipoksantin prevodeći ih u mokraćnu kiselinu, pa se
često naziva i ksantin-oksidaza.
Tirozinoksidaza krompira
Ovaj enzim, sadrţan izmeĎu ostalog i u krompiru, oksidira tirozin koristeći kisik. Tirozin se
postepeno prevodi u oksidacione produkte, koji imaju crvene, odnosno, daljom
oksidacijom-crnu boju.
Postupak: U dvije epruvete staviti po nekoliko kapi 1% rastvora tirozina. U epruvetu 1
dodati 0,5 mL filtriranog, vodenog ekstrakta krompira (narendani krompir-ekstrahovati
vodom) (Narendani krompir staviti u gazu, zatim uroniti u laboratorijsku čašu od 500 mL,

BIOHEMIJA II–laboratorijske vjeţbe
46
koja sadrţi 200 mL destilovane vode. Lagano istisnuti škrob. Promjeniti vodu i ponoviti
ekstrakciju. Pustiti da se iz ekstrakta istaloţi škrob i filtrirati supernatant. Dobijeni filtrat
koristiti za uzorak u epruveti 1). U epruvetu 2 dodati 0,5 mL vode. Staviti obje epruvete u
vodeno kupatilo na 40°C, i tokom inkubacije sadrţaj epruveta nekoliko puta promješati
rada inteziviranja aeracije. Utvrditi da tokom inkubacije djelovanjem tirozinoksidaze, uz
korištenje kisika, tirozin prelazi (u prvoj epruveti) u crveno obojeni derivat.
Stajanjem, tj. inkubacijom u roku 1-2 sata tirozin prelazi u crni pigment melanin.
U drugoj epruveti nema promjene, jer nije dodan enzim, pa tirozin ostaje nepromjenjen.
Katalaza
Ova grupa enzima razlaţe vodikperoksid (nastao oksidacijom-dehidrogenacijom-kada se
ona vrši u aerobnim uslovima).
2H2O2 → 2H2O + O2
Izdvaja se molekularni kisik.
Postupak: U epruvetu staviti 1 mL 6% rastvora vodikperoksida i 1 kap krvi. Odmah se
izdvajaju mjehurići kisika, tečnost se zapjeni i pjena puni epruvetu.
Peroksidaze
Peroksidaze kataliziraju oksidacije fenola i polifenola vodikperoksidom ili organskim
peroksidima. Prenose nascentni kisik iz peroksida na primaoca, koji se oksidira.
Peroksidazna aktivnost hemoglobina
Hemoglobin i citohromi imaju izraţenu peroksidaznu aktivnost, što se moţe potvrditi
reakcijom sa benzidinom i H2O2.
Postupak: Rastvoriti malo benzidina (ima ulogu primaoca aktivnog-nascentnog kisika) u 3-
4 mL 25% sirćetne kiseline. Promućkati, pa dodati 1 mL razblaţenog vodikperoksida i
jednu kap razblaţene, defibrirane krvi. Rastvor postaje vrlo plav ( u prisustvu tragova krvi),
jer se benzidin oksidiše i prelazi u plavo, odnosno tamno-zeleno obojene oksidacione
derivate.
Peroksidaza krompira
Postupak: Ponoviti postupak sa benzidinom, ali umjesto krvi uzeti 1mL vodenog ekstrakta
krompira. Utvrditi da benzedin prelazi u oksidacioni produkt plave boje.

BIOHEMIJA II–laboratorijske vjeţbe
47
Peroksidaze hrena
Postupak: Napraviti ekstrakt hrena u vodi (nastrugati korjen hrena u 5-10 mL vode), pa ga
poslije 60 minuta stajanja filtrirati. Filtrat sadrţi peroksidazu.
U dvije numerisane epruvete uzeti po 20 kapi 2% pirogalola u vodi, pa zatim:
- epruvetu 1 staviti 20 kapi ekstrakta hrena i 2 kapi vode
- u peruvetu 2 staviti 20 kapi ekstrakta hrema i 2 kapi 1 % rastvora vodikperoksida
Ekstrakt hrema sadrţi peroksidazu, koja u drugoj epruveti koristeći dodani vodikperoksid,
prevodi pilogalol u purpurgalin.
Purpurgalin je nerastvorljiv i izdvaja se kao talog crvene boje.
3. ZADACI I VJEŽBE
1. Koji enzim pokazuje apsolutnu specifičnost prema supstratu:
a) Himotripsin
b) Papain
c) Ureaza
d) Arginaza
e) Lizozom
2. Koji od proteolitičkih enzima pokazuje esteraznu aktivnost:
a) Tripsin
b) Karboksipeptidaza
c) Aminopeptidaza
d) Himotripsin
e) Pepsin
3. Na kojoj temperaturi se enzimi denaturiraju:
a) 0 °C
b) 80-100 °C
c) 20-30 °C
d) 30-40 °C
4. Koja je temperatura optimalna za djelovanje većine enzima:
a) 50-60 °C
b) 15-20 °C
c) 80-100 °C
d) 35-40 °C
5. Koji optimum pH ima enzim pepsin:
a) 1,5-2,5
b) 4-5
c) 6-7
d) 8-9
e) 10-11

BIOHEMIJA II–laboratorijske vjeţbe
48
6. Kinetika enzima je riješena preko Michael-Mentenove konstante. Napisati jednačinu
i prikazati je grafički?
4. REZULTATI
Specifiĉnost enzima
Specifiĉnost
amilaze Rezultat Fehling-ova proba Komentar
Epruveta I
Epruveta II
Specifiĉnost
saharaze Rezultat Fehling-ova proba Komentar
Epruveta I
Epruveta II

BIOHEMIJA II–laboratorijske vjeţbe
49
Dokazivanje aktivnosti i termolabilnosti ptijalina (α-amilaze iz pljuvaĉke)
Broj epruvete
Rezultat
Fehling-ova proba
Komentar
Optimalni pH djelovanja amilaze
Broj epruvete
Rezultat
Komentar
Inhibitorsko djelovanje hlorida na dehidrogenazu
Broj epruvete
Rezultat
Komentar
Ureaza
Broj epruvete
Rezultat
Komentar

BIOHEMIJA II–laboratorijske vjeţbe
50
Praćenje aktivnosti pepsina
Broj epruvete
Biuret proba
Komentar
Dehidrogenaza mlijeka
Broj epruvete
Rezultat
Komentar
Tirozinoksidaza krompira
Broj epruvete
Rezultat
Komentar

BIOHEMIJA II–laboratorijske vjeţbe
51
Katalaza
Broj epruvete
Rezultat
Komentar
Peroksidaze
Peroksidna
aktivnost
hemoglobina
Peroksidna
aktivnost
krompira
Peroksidna aktivnost hrena
Broj epruvete
Rezultat
Komentar
OVJERA VJEŽBE:

BIOHEMIJA II–laboratorijske vjeţbe
52
Vježba broj 8
ODREĐIVANJE AKTIVNOSTI ENZIMA: α-AMILAZE I KATALAZE
1. TEORETSKI DIO
Postoji niz metoda za odreĎivanje aktivnosti amilaze. Prema njihovom principu mogu
se podijeliti na slijedeće grupe:
o Viskozimetrijske metode (zasnivaju se na smanjenju viskoznosti rastvora škroba
zbog hidrolitičkog djelovanja amilaze);
o Saharogene metode, zasnivaju se na odreĎivanju reduktivnih spojeva koji nastaju
hidrolizom škroba. Ove metode nisu precizne;
o Hromogene metode, koriste se u proteklih 20-tak godina, a zasnivaju se na
djelovanju amilaze na škrob ili amilozu ili amilopektin koji su kovalentno vezani
(eterski ili esterski) za neku boju koja je derivat trijazina, koja se oslobaĎa pri
hidrolizi škroba (amiloze ili amilopektina) , centrifugiranjem odvaja i mjeri
fotometrijski;
o Nefelometrijske metode, zasnivaju se na smanjenju zamućenosti reakcione smjese
škroba i ispitivanog materijala (urin, plazma, pljuvačka) nakon hidrolize škroba;
o Spektrofotometrijske i fluorimetrijske enzimske metode u kojima se prati
oksidoredukcija nikotinamidadenin dinukleotida (NAD-a);
o Fotometrijske metode sa sintetskim supstratima čiji se razgradni produkti mogu
mjeriti fotometrijski, kontinuirano. Takvi su supstrati razni: p-nitrofenilni i Cl-p-
nitrofenilni maltozidi (u obliku testova). To su najnovije i dosta precizne metode.
Zamjenjuju klasične metode zbog toga, što hidrolizom škroba pod djelovanjem
amilaze nastaju produkti koji nisu tačno definirani u datom momentu (amilo-, eritro-
, ahromodekstrini);
o Amiloklasične metode-zasnivaju se na svojstvu škroba ili amiloze da sa jodom daju
plavi produkt (amilopektin sa jodom daje ljubičastu boju). Djelovanjem amilaze na
škrob nastaju prvo amilodekstrini (sa jodom daju plavoljubičastu boju), zatim
eritrodekstrini (sa jodom daju crvenu boju), ahromodekstrini (ahrodekstrini) sa
jodom se ne boje
U grupu amiloklasičnih metoda ubraja se i metoda odreĎivanja amilaze po Wohlgemuth-u.
Prva metoda za odreĎivanje aktivnosti amilaze koja se zasniva na tome koje razrijeĎenje
uzorka još razgraĎuje škrob, pa se dodatkom joda (Lugolovog rastvora) ne javlja plava
boja. Metodu je uveo Wohlgemuth kao prvu metodu odreĎivanja enzima u dijagnostičke
svrhe. Najveću dijagnostičku vrijednost metoda je pokazala kod oboljenja pankreasa i
parotitisa. Tako α-amilaza iz pljuvačke (ptijalin) ima povećanu aktivnost u pljuvačci,
serumu i urinu kod akutnog parotitisa (zaušnjaka) ili kod čira (ulcusa) na ţelucu. Amilaza
iz pankreasa (dijastaza) ima povećanu aktivnost u serumu i urinu kod akutne nekroze
pankreasa u početnom stadiju, akutnog pankreatitisa. Značajno povišenje aktivnosti
dijastaze u urinu javlja se kod upala ţučne kese, uremije, perforacije čira na ţelucu,
postoperativnih stanja.
Katalaza je široko rasprostanjen enzim i sreće se kod prokariaota i eukariota. Predstavlja
dio antioksidativne zaštite organizma. Katalaza razlaţe toksični vodikperoksid na molekul
kisika i vodu. Katalaza je tetramer sastavljen od četiri polipeptidna lanca, svaki sadrţi preko
500 aminokiselina. Sadrţi četiri porfirinska hema, grupe koje omogućavaju enzimu da
reaguje sa vodikperoksidom.

BIOHEMIJA II–laboratorijske vjeţbe
53
Zastupljen je gotovo kod svih organizama, počev od mikroorganizama (aerobnih i
anaerobnih), biljaka, ţivotinja do čovjeka. Kod čovjeka je široko zastupljenja posebno u
ćelijama jetre u peroksizomima i crvenim krvnim zrncima. Optimalni pH za ljudsku
katalazu je oko 7, dok kod ostalih organizama, u zavisnosti od vrste pH se kreće izmeĎu 4-
11. Ljudska katalaza djeluje na 37°C, dok kod nekih jednoćelijskih organizama optimalna
temperatura djelovanja je 90°C. Osnovna uloga ovog enzima je razgradnja vodikperoksida.
Katalaza je veoma aktivan enzim, jedna molekula katalaze konvertuje million molekula
vodikperoksida u vodu i kisik svake sekunde. Katalaza moţe katalizirati i oksidaciju,
pomoću vodikperoksida, različitih metabolita i toksina, kao što su formaldehid, mravlja
kiselina, fenoli, acetilaldehid i alkoholi. Dešava se slijedeća reakcija, čiji mehanizam je i
dalje nepoznat.
H2O2 + H2R → 2H2O + R
Joni teških metala (kao što su bakarni joni) mogu da se ponašaju kao nekompeticijski
inhibitori katalaze. Otrovi, kao što su cijanidi su kompeticijski inhibitori katalaze, snaţno se
vezuju za hem katalaze i zaustavljaju enzimsku reakciju.
Kod čovjeka i drugi enzimi mogu da razgrade vodikperoksid. Glutationperoksidaza je u
fiziološkim uslovima (mala koncentracija vodikperoksida) je čak aktivnija od katalaze.
Kada koncentracija vodikperoksida poraste uključuje se katalaza. Katalaza test je jedan od
tri glavna testa koji se koristi u mikrobiologiji za identifikaciju bakterija. Prisustvo enzima
katalaze se detektuje upotrebom vodikperoksida. Bakterije koje su katalaza-pozitivne
dodavanjem vodikperoksida „oslobaĎaju” kisik.

BIOHEMIJA II–laboratorijske vjeţbe
54
2. EKSPERIMENTALNI DIO
OdreĊivanje aktivnosti α-amilaze
U stalak se pripremi 10 čistih epruveta i u svakoj odpipetira po 1 ml fiziološkog
rastvora (0,90% NaCl). U posebnu, čistu epruvetu se sakuplja profiltrirana pljuvačka bez
dodatka vode radi lakšeg i brţeg filtriranja. Sakupi se nešto više od 1 ml, a potom
odpipetira 1 ml i stavi u prvu epruvetu.
Sadrţaj epruvete se dobro promiješa i na taj način se dobije dva puta razblaţena pljuvačka.
Nakon toga se odpipetira 1 ml tako razblaţene pljuvačke i stavi u drugu epruvetu. Sadrţaj i
ove epruvete se dobro promiješa i dobije četiri puta razblaţena pljuvačka. Postupak se
ponavlja sve do desete epruvete iz koje se 1 ml pljuvačke izbaci. Potrebno je prilikom ovog
razblaţivanja pipetu kojom se pipetira i prebacuje sadrţaj jedne epruvete u drugu, nakon
svakog prebacivanja sadrţaja isprati vodom. Kada se naprave razblaţenja pljuvačke, u
svaku epruvetu se odpipetira po 1 ml 0,1% koloidnog rastvora škroba. Sadrţaji u svakoj
epruveti se promiješaju, sve se vrati u stalak i termostatira na sobnoj temperaturi. Poslije 15
minuta termostatiranja u svaku epruvetu (od desete do prve) se doda po jedna kap
Lugolovog reagensa. U epruvetama u kojima je škrob hidroliziran nema plave boje, a u
kojima nije hidroliziran pojavljuje se plava boja. IzmeĎu obojenih i neobojenih sadrţaja u
epruveta javlja se jedan koji sa Lugolovim rastvorom daje crvenu boju. Boja potiče od
razgradnih produkata škroba-eritrodekstrina sa jodom iz Lugolovog reagensa. U toj
epruveti razblaţena pljuvačka sadrţi jednu enzimsku jedinicu α-amilaze, po Wohlgemuth-u.
Jedna enzimska jedinica α-amilaze po Wohlgemuth-u je ona aktivnost ovog enzima pri
kojoj se na temperaturi od 38 C hidrolizira 1 mg škroba za 30 minuta, odnosno 0,00055
mg škroba za jednu sekundu. Najmanje razblaţenje uzorka u kome se za 30 minuta na
temperaturi od 38 C hidrolizira jedan mg škroba do produkata koji sa Lugolovim
rastvorom ne daju plavu boju (eritrodekstrini-daju crvenu boju) sadrţi jednu jedinicu α-
amilaze po Wohlgemuth-u (1 WJ).
Pretvaranje IU u katale:
1 kat = 1 mol/s = 60 mol/min = 60x106 μmol/min = 6x10
7 IU
1 IU = 1 μmol/min = 1/60 μmol/s = 1/60 μkat = 16,67 nkat
Ukoliko se kao supstrat na koji djeluje amilaza koristi škrob koji ima molekulsku masu
55000, onda 1 WJ α-amilaze iznosi 0,01 nkat.
OdreĊivanje aktivnosti katalaze
Princip određivanja: Prema Bah-u i Oparin-u kvantitativno odreĎivanje katalaze zasnovano
je na osobini H2O2 koji zaostaje nakon razloţnog djelovanja katalaze, da reaguje sa KMnO4
uz obrazovanje slobodnog kisika. Jednačina ove reakcije je slijedeća:
2KMnO4 + 5H2O2 + 4H2SO4 → 2MnSO4 + 2KHSO4 + 8 H2O + 5O2
Na osnovu razlike u mL rastvora KMnO4 utrošenog za titraciju kontrolne probe i ogledne
probe dolazi se do količine H2O2 koja je razloţena od strane enzima.
Osim KMnO4, kao sredstvo za razlaganje viška H2O2 posle inkubacije sluţi J2.
Reakcija se odvija po po jednačini:

BIOHEMIJA II–laboratorijske vjeţbe
55
H2O2 + 2KJ H2SO4 → K2SO4 + J2 + 2H2O
Višak joda se odreĎuje sa Na2S2O3.
Reagensi i pribor:
1. Preparat katalaze: Odmjeriti na analitičkoh vagi 2g brašna ili drugog izmljevenog biljnog
materijala, staviti u balon od 100 mL, dodati vode i dobro promućkati, zatim dodati 2-3
kapi toluola, dopuniti balon destilovanom vodom do markice i ostaviti smjesu na sobnoj
temperaturi u toku 2 sata. Posle ekstrakcije, sadţaj profilitrirati kroz naboran filter papir
(moţe i centrifugirati) i filtrat koristiti kao ekstrakt katalaze.
Za ekstrakciju enzima moţe se koristiti i krompir. 20g krompira isitniti i izmrviti, staviti u
balon, dodati vode, promućkati, dopuniti vodom do 50 mL i profiltirrati.
Za ispitivanje katalaze moţe se koristiti i krv u razblaţenju 1:1000.
- 1% rastvor H2O2
- 10% H2SO4
- 0,1 M KMnO4
- 0,1 M NaOH
Tok odreĎivanja: U četiti tikvice nasuti pipetom po 20 mL filtrata, zatim u posljednje dvije
(3 i 4) koje sluţe kao kontrolne zagrijati da sadrţaj proključa (kuhati 5 minuta) radi
inaktivacije enzima, a onda ohladiti.
Posle toga u sve tikvice dodati po 20 mL destilovane vode, po 5 mL 1% rastvora H2O2, koji
je prethodno neutraliziran rastvorom NaOH i ostaviti ih na sobnoj temperaturi u toku 30
minuta. Posle tog roka dodati u probe po 5 mL 10% H2SO4 i smjesu titrirati rastvorom
KMnO4.
Aktivnost katalaze se odreĎuje po količini mg H2O2 koji se u toku 30 minuta razloţi
pomoću katalaze koja sadrţi 1 g ispitivanog materijala (1 mL 0,1 M KMnO4 ekvivalentan
je 1, 7 mg H2O2).
A = g
xba 7,1)(
Gdje je:
a- broj mL 0,1 M KMnO4 utrošenog za kontrolnu probu
b - broj mL 0,1 M KMnO4 utrošenog za uzorak
g – količina ispitivanog uzorka u g

BIOHEMIJA II–laboratorijske vjeţbe
56
3. ZADACI I VJEŽBE
1. Koji pH optimum ima enzim amilaza:
a) 1,5-2
b) 7-7,5
c) 8-9
d) 3,5-4
e) 4,5-5
2. Za koje enzime su Mg2+
aktivatori:
a) Fosforilaze
b) Amilaze
c) Heksokinaze
d) Kreatinkinaze
e) Karboksipeptidaze
3. Za koje enzime su joni Zn2+
aktivatori:
a) Karboksipeptidazu
b) Karboanhidrazu
c) Glutamatdehidrogenazu
d) Laktatdehidrogenazu
e) Aminooksidazu
4. Koji je jon neophodan za djelovanje kinaza (fosfotransferaza) koje kataliziraju
graĎenje O-fosfatnih estera:
a) Mg2+
b) Co2+
c) Ca2+
d) Na+
e) K+
7. Kod kojih enzima u aktivnom centru su dokazane aminokiseline glutamat i aspartat:
a) Piruvatdehidrogenaze
b) Lizozomu
c) Pepsinu
d) Ureazi
e) Karboksipeptidazi
8. Koji enzimi pripadaju grupi „serinskih“ enzima:
a) Pepsin
b) α-himotripsin
c) tripsin
d) acetilholinesteraza
e) lizozom
9. Dokazati da 1 WJ iznosi 0,01 nkat?

BIOHEMIJA II–laboratorijske vjeţbe
57

BIOHEMIJA II–laboratorijske vjeţbe
58
4. REZULTATI
Izračunati aktivnost uzorka amilaze i prikazati u WJ, kat i IU.
Broj
epruvete
Razblaţenje
pljuvačke
WJ

BIOHEMIJA II–laboratorijske vjeţbe
59
Izračunati aktivnost katalaze
OVJERA VJEŽBE:

BIOHEMIJA II–laboratorijske vjeţbe
60
Vježba broj 9
SPEKTROFOTOMETRIJSKO ODREĐIVANJE SADRŽAJA
UKUPNIH FENOLA U UZORKU PĈELINJEG MEDA
1. TEORETSKI DIO
Postoji više načina da se odredi ukupna količina fenolskih spojeva, uz napomenu da
nijedna od ovih tehnika ne detektuje sve fenolske spojeve. Jedna od najčešće korištenih
metoda za odreĎivanje ukupnog sadrţaja fenola je stara oko 100 godina i u početku je
razvijena za istraţivanje metabolizma proteina kod ljudi. Folin i Denis (1912) su koristili
kolorimetrijski metod detekcije tirozina prilikom hidrolize proteina. Aktivni reagens je ţut i
sastavljen je od kompleksnih polimernih jona formiranih od fosfomolibdenove i
fosfovolframove kiseline. Čini se da ovaj rastvor sadrţi integrirane polimerne serije i da
imaju formu tetraedra okruţenu sa oktaedarskim molibden oksikiselinskim jedinicama u
kojoj volfram moţe lahko supstituirati molibden. Ovaj reagens oksidira fenolate i
heteropolarna kiselina biva reducirana iz stanja +6 u smjesu stanja +6 i +5 uz nastanak
plavog molibden-volframovog kompleksa. Metod je modificiran od strane Folin-a i
Ciocalteu-a (1927). Modifikacija uključuje dodatak litijuma u fosfovolframat /
fosfomolibdat reagens na kraju procesa kuhanja. Dodatak litijuma spriječava precipitiranje i
smetnje prilikom kvantifikacije u odnosu na intenzitet boje. Intenzitet boje moţe biti
korišten za kvantitativnu analizu na osnovu vrijednosti apsorbanse. Koncentracija fenola se
izraţava kao ekvivalenti galne, odnosno hlorogenske kiseline koje se koriste kao standardi.
Od ranije je evidentan uticaj fenolskih komponenti na ljudsko zdravlje.
Najvjerovatnije najstarija medicinska primjena fenolskih komponenti je upotreba fenola
kao antiseptika. Zbog negativnog uticaja koji ima na tkiva, posebno nastajanje rana pri
velikim koncentracijama, nije više u upotrebi. Do danas su se zadrţale u upotrebi 0,5%
otopina fenola protiv bakterija Staphylococcus aureus i 1,5% otopina kao oralni anestetik.
Druga česta upotreba fenolskih komponenti je kao zaštita od UV zračenja. Prisustvo
aromatskog prstena doprinosi efektivnoj apsorpciji UV zračenja (izmeĎu 280-315 nm) i na
taj način štiti od opekotina. Najčešći sastojak krema za sunčanje je p-aminobenzojeva
kiselina (nije fenolski spoj). Ovaj sastojak se koristio do 1970-ih kada je otkriveno da
izaziva osip i akne. Alternative aktivne komponente su salicilati, kao što je oktilsalicilat,
cimetati kao što je oktilmetilcimetat (nerastvoran u vodi i koristi se za kreme nerastvorne u
vodi), benzofenon, oksibenzon, antraanilati kao što je metilantranilat. Imaju ulogu pri
stresnim situacijama u okolini, što uključuje zaštitu od visokih nivoa vidljive i UV
svjetlosti, odbrana od patogena i opšta zaštita od oksidativnog stresa, ozona. TakoĎer, utiču
na ljudsko zdravlje uz moguće djelovanje kao antioksidanti, antikancerogeni i
karvoprotektivni agens. Fenoli u biljkama nalaze slijedeće uloge: zaštita od štetnog UV
zračenja, odbrana od biljojeda, odbrana od patogena, zaštita od fitoinhibije i zaštita od
reaktivnih kisikovih čestica (ROS-reactive oxygen species).
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- Uzorci za analizu-Pčelinji med
- Standard-galna kiselina, 98 %, - Folin-Ciocalteu's reagens
- Natrij-karbonat

BIOHEMIJA II–laboratorijske vjeţbe
61
Za analizu će biti korištena metoda po Singleton-Rossiju.
a) Priprema standardne otopine galne kiseline
U čistoj i suhoj čašici odvagati 0,0250 g galne kiseline, dodati 200 l 95% etanola, lagano
protresti rukom da se supstanca otopi, te dodavanjem destilirane vode špric balonom
kvantitativno prenijeti sadrţaj (ispirati čašicu destiliranom vodom) u odmjerni sud od 50
ml. Postojanost ove otopine je 2 dana na 4 C. Odgovarajuća razrijeĎenja ove standardne
otopine koriste se za pripremu kalibracionog pravca (npr. pripremiti po 10,00 ml 200, 150,
100, 50 i 10 mg/l galne kiseline).
b) Priprema uzorka
U čistoj i suhoj čašici odvagati na analitičkoj vagi oko 1 g uzorka meda, zapisati tačnu
masu odvage, dodati na uzorak 10 ml destilirane vode i miješati staklenim štapićem do
otapanja. Otopinu sa uzorkom prenijeti kvantitativno u odmjerni sud od 25 ml uz ispiranje
čaše sa još 10 ml (u porcijama po 5-6 ml) destilirane vode. Nakon što se i taj dio prenese u
odmjerni sud, otopina se protrese i dopuni do oznake od 25,00 ml pomoću špric balona.
Ukoliko se primijete čvrsti ostaci u otopini, kompletan sadrţaj profiltrirati preko običnog
filtar papira u čistu čašu ili drugi odmjerni sud. Nakon toga, odreĎeni volumen uzorka
razrijediti u omjeru 1 : 10 (npr. na 1,00 ml uzorka dodati 9,00 ml destilirane vode).
c) Eksperimentalna procedura
Na 2,00 ml otopine uzorka ili standarda (razrijeĎenih na gore opisani način) doda se 10,00
ml Folin-Ciocalteu reagensa razrijeĎenog u omjeru 1:10 i promiješa. Ova smjesa se ostavi
da odstoji oko 10 minuta. Zatim se doda 8,0 ml 7,5 %-og Na2CO3. Nakon 30 minuta
stajanja na sobnoj temperaturi očita se apsorbansa pri 743 nm.
Priprema slijepe probe obavlja se na isti način, s tom razlikom što se umjesto uzorka ili
standarda uzima 2,00 ml destilirane vode
3. REZULTATI
Uzorak γ(mg/L) Apsorbansa (A)

BIOHEMIJA II–laboratorijske vjeţbe
62
- Koncentracija nepoznatog uzorka (rezultat izraziti kao mg galne kiseline/g
uzorka)
- Uz izvještaj priloţiti i grafik kalibracione krive
OVJERA VJEŽBE
BIOHEMIJA II–laboratorijske vjeţbe
63
Vježba broj 10
SPEKTROFOTOMETRIJSKO ODREĐIVANJE UKUPNOG
HLOROFILA
1. TEORETSKI DIO
Hlorofili apsorbuju svjetlost u plavom (430-490 nm) i crvenom (630-760 nm) dijelu
spektra, a zelenu svjetlost propuštaju pa otuda imaju zelenu boju. Hlorofil ima sposobnost
fluorescencije, što znači da svjetlost koju apsorbuje moţe ponovo da otpusti, ali kao
svetlost veće valne duţine, dalekocrvene. Zbog toga je rastvor hlorofila, posmatran u
epruveti u odbijenoj svetlosti, tamnocrvene boje.
Apsorbcioni spekrar hlorofila a b hlorofila b
Količinski odnos hlorofila a i b u tijelu viših biljaka zavisi od količine svjetla kojom su
izloţene. Tako dobro osjvetljene biljke imaju tri puta više hlorofila a nego hlorofila b, dok
one koje ţive u sjenci imaju više hlorofila b. Razlog tome je pojava označena kao
komplementarna hromatična adaptacija prema kojoj biljke stvaraju pigmente čija je boja
podudarna boji raspoloţive svjetlosti. Hlorofil, hemoglobin i vitamin B12 imaju sličnu
hemijsku strukturu, ali im se uloge u ćelijama biljaka i ţivotinja bitno razlikuju. Hlorofil je
po hemijskoj graĎi porfirinski spoj izgraĎen od:
porfirinskog prstena u čijem se centru nalazi atom magnezijuma (Mg); porfirinski
prsten je aktivni dio molekula;
fitolnog lanca u vidu produţetka (rep hlorofila) lipofilnih svojstava preko koga se
molekul pričvršćuje za tilakoidnu membranu.
Strukture hlorofila a i hlorofila b

BIOHEMIJA II–laboratorijske vjeţbe
64
Hlorofili a i b se meĎusobno razlikuju po tome što je kod hlorofila a na poziciji 3 u
porfirinskom prstenu metil grupa (CH3), dok je na istoj poziciji kod hlorofila b aldehidna
grupa (CHO).Na tilakoidnim membranama pigmenti se udruţuju sa proteinima gradeći
naročite makromolekulske komplekse pigment/protein koji se označavaju kao:
fotosistem I koji sadrţi hlorofil a i hlorofil b u odnosu 12:1, β-karoten (pripada
karotenoidima) i polipeptida; loptastog je oblika i velike molekulske mase od 110
000 Da;
fotosistem II se sastoji od hlorofila a i b u jednakim količinama (1:1), ksantofila,
feofitina i kompleksa mangana koji oksiduje vodu; ima jajast oblik i manju
molekulsku masu od fotosistema I (32 000 Da)
U fotosistemu I glavni pigment je molekul hrorofila a koji apsorbuje svetlost maksimalne
valne duţine 700nm (ozanačava se kao P700), dok je u fotosistemu II hlorofil a koji upija
svjetlost talasne duţine 680 (označava se kao P680). Oba fotosistema funkcionišu po istom
principu: apsorbuju sunčevu svjetlost i pretvaraju je u hemijsku energiju. Prema najnovijim
istraţivanjima makromolekulskih kompleksa tilakoidne membrane smatra se da su
fotosistemi nešto sloţenije graĎe i da obuhvataju kompleks, skraćeno nazvan LHC (Light
Harvesting Complex = kompleks koji žanje svetlost), u kome je skoncentrisano 99%
hlorofila, praktično sav hlorofil koji je vezan za tilakoidnu membranu.
LHC kompleks je izgraĎen od dva osnovna dela:
antene koja žanje sunčevu svjetlost i
reakcionog centra koji je hvata.
Funkcija fotosistema svodi se na slijedeće: u nivou antena se apsorbuje svjetlosna energija,
a zatim pomoću elektrotransportnog lanca (lanac prenosilaca elektrona) se prenosi do
reakcionog centra pri čemu dolazi do oslobaĎanja energije koja se iskoristi za izbacivanje
protona u lumen tilakoida. To pokreće sa jedne strane sintezu ATP, a sa druge strane
dovodi do redukcije NADP i oksidacije vode do molekulskog kiseonika koji se oslobaĎa u
atmosferu. Svaka vrsta hlorofila apsorbuje svetlost tačno odreĎene valne duţine u
zavisnosti od svog apsorpcionog spektra. Pri tome elektroni molekula hlorofila postaju
bogatiji energijom, koja je jednaka apsorbovanom kvantu svetlosti (fotonu) i prelaze u
pobuĎeno (ekscitovano) stanje, odnosno, na slijedeću orbitalu koja odgovara povišenoj
energiji. U molekulu hlorofila elektron moţe da preĎe na dijve moguće orbite pobuĎenog
stanja jednu koja odgovara fotonu crvene (niţa), a drugu koja odgovara fotonu plave
svetlosti (viša). Ekscitirani elektron napušta molekul hlorofila i prihvata ga akceptor
elektrona (feredoksin) koji se pri tome redukuje, dok sam molekul hlorofila ostaje
oksidovan (pozitivno naelektrisan). Oksidovani molekuli hlorofila izgubljene elektrone
nadoknaĎuju od tzv. donora (davaoca) elektrona. Hlorofili fotosistema II nadoknaĎuju
elektrone iz vode, a fotosistema I to čine iz fotosistema II.

BIOHEMIJA II–laboratorijske vjeţbe
65
2. EKSPERIMENTALNI DIO
Hlorofil iz biljnog materijala se kvantitativno ekstrahuje 96 % etanolom i u dobijenom
ekstraktu se kolorimetrijsi odreĎuje njegov sadţaj. OdreĎivanje koncentracije hlorofila u
ispitivanom uzorku moţemo odrediti na principu zavisnosti inteziteta propuštene svijetlosti
od koncentracije rastvora i debljine njegovog sloja.
Za pravljenje standardnog rastova upotrebljavaju se slijedeći spojevi: CuSO4, K2Cr2O7 i
NH4OH koji daju absorbciju koju bi dao rastvor 8,5 hlorofila u 100 mL 96% etanola.
Postupak: Uzeti 0,5-5 g biljnog materijala i staviti u čašu sa ključalom vodom kojoj je
dodato oko 0,1 g CaCO3. Posle kuhanja koje traje oko 1 minut, materijal se vadi i stavlja na
saharno stanko da se odstrani suvišna voda. Zatim, material prenjeti u tarionik u kome se
nalazi nešto kvarcnog pijeska i CaCO3 i paţljivo homogenizira uz dodavanje nekoliko
mililitara 96% etanola. Dobije ekstrakt se prenosi u lijevak sa filter papirom, a masi u
tarioniku dodaje se nova količina alkohola i nastavlja homogeniziranje i ekstrakcija.
Postupak se ponavlja sve dok se ne izgubi zelena boja ekstrakta koja otiče iz lijevka. Po
završenoj ekstrakciji filtrat se dopun 96% alkoholom do 100 mL. Taj rastvor sluţi da se u
njemu kolorimetrijski odredi sadrţaj hlorofila.
Kao standard sluţi rastvor koji u 100 mL sadrţi: 28,5 mL 1% rastvora CuSO4, 50 mL 2%
rastvora K2Cr2O7 i 10 mL 7% rastvora NH4OH i sve to dopuni destilovanom vodom do 100
mL. Ovakav rastvor po intezitetu boje odgovara rastvoru hlorofila (a+b) koncentracije 8,5
mg/100 mL.
Izračunati sadrţaj hlorofila kao mg/mL i kao sadrţaj mg hlorofila/ g biljnog materijala.

BIOHEMIJA II–laboratorijske vjeţbe
66
3. REZULTATI
- Koncentracija nepoznatog uzorka (rezultat izraziti kao mg hlorofila/ g biljnog
materijala)
OVJERA VJEŽBE

BIOHEMIJA II–laboratorijske vjeţbe
67
Vježba broj 11
PRIMJENA ADSORPCIONE HROMATOGRAFIJE U SEPARACIJI
BILJNIH PIGMENATA
1. TEORETSKI DIO
U osnovu adsorpcione hromatografije raznih spojeva prisutan je princip različite
polarnosti. Spojevi male i srednje polarnosti razdvajaju se najbolje od polarnih spojeva. Za
formiranje kolone koriste se razni adsorbensi, sa nejednakim adsorpcionim osobinama.
Takvi su silika gel, glina, kalcijum fosfat, aktivni ugalj (biljni ili ţivotinjski) i saharoza.
Adsorpciona sredstva pojedinih adsorbenasa mogu se u značajnom stepenu modificirati ili
izmjeniti putem raznog prethodnog tretiranja ili obradom, na primjer sa kiselinama ili
zagrijanjem. Ova obrada poznata je kao aktivacija ili aktiviranje adsobenasa.
Prije nanošenja uzorka redukovati nivo tečnosti u koloni neposredno iznad diska filter
papira. Koncentrovani uzorak pipetom paţljivo naneti na vrh kolone. Zatim dodati solvent
postepeno i uspostaviti jedan konstantni nivo tečnosti na vrhu kolone. Ako je brzina
proticanja solventa kroz kolonu suviše mala, ista se moţe povećati spajanjem vrha kolone i
vakuum pumpe. U takvom slučaju postupani paţljivo. Razdvajanje komponenata iz uzorka
nanetog na kolonu adsorbensa zavisi, izmeĎu ostalog, od izbora odgovarajućeg solventa za
svaki pojedinačni slučaj, što je u saglasnosti sa svim mogućim interakcijama izmeĎu tri
komponente sistema: rastvorena supstanca, solvent, adsorbens, a što se mora imati u vidu
pri svakom empirijskom zahvatu.
Eluirajuća moć solventa raste sa njihovom polarnošću. Ona se uopšteno moţe izraziti
sljedećim rasporedom:
voda ˃ etanol ˃ aceton ˃ hloroform ˃ petrol eter
Jačina adsorpcije komponente na koloni je u zavisnosti sa polarnošću. Više polarna
jedinjenja se jače adsorbuju. Ovo se moţe izraziti na slijedeći način: kiseline ˃ alkoholi ˃
ketoni ˃ ugljikovodici. Razdvajanje eluiranjem ostvaruje se u slučaju kad se solvent
adsorbuje mnogo slabije od komponente, te on u svom kretanju jednostavno „prekoračuje“
adsorbovane spojeve duţ kolone. Na ovaj način moţe se postići dobro razdvajanje
komponenti iz uzorka, ali je u sistemu „angaţovan“ samo jedan dio ukupnog kapaciteta
kolone.
Razdvajanje pomicanje (potiskivanjem) javlja se u slučaju mnogo jačeg adsorbovanja
solventa u poreĎenju sa spojevima, tako da se velike količine materijala mogu razdvojiti, a i
sama separacija je adekvanta.
Kod adsorpcione hromatografije obojenih spojeva obično se primjenjuje istiskivanje kolone
iz nosača nakon separacije traka, a zatim se ova isječe na sektore sa trakama, koji se
pomoću solventa eluiaju i spojevi odreĎuju na podesan način.

BIOHEMIJA II–laboratorijske vjeţbe
68
2. EKSPERIMENTALNI DIO
Razdvajanje pigmenata zelenih listova na koloni sa više asdorbenata
a) Ekstrakcija pigmenata
Suši se 4-5 svjeţih listova špinata 1-2 sata na 40°C ili 24 sata na 20°C. Listovi se izreţu na
trake i ekstrahuju 1 sat sa smjesom od 10ml benzena, 30m etanola i 90ml petroletera.
Filtritra se i filtrat ispere 4 puta vodom, suši natrij-sulfatom i upari na vodenom kupatilu do
zapremine od 5ml.
b) punjenje kolone
U erlenmajerovu tikvicu staviti 8g Al2O3 II stepena aktivnosti i 15 ml petroletera
U erlenmajerovu tikvicu staviti 5,3g kalcij-karbonata (sušenog na 150°C) i 30ml
petroletera
U erlenmajerovu tikvicu staviti 7g fino sprašene saharoze i 20 ml petroletera
Kolonu 1/18 dobro oprati, osušiti i na donjem dijelu staviti tampon od staklene vune.
Kolona se učvrsti na stativ da stoji okomito. Al2O3 se dobro izmješa sa petroleterom.
Dobivena suspenzija se ulije u kolonu tako da se sloj adsorbenta razmjesti ravnomjerno, a
nakon toga se na površinu adsorbenta stavi tačno izrezani kruţić filter papira. Zatim se na
isti način unosi suspenzija CaCO3 i filter papir, i kao zadnji adsorbent suspenzija saharoze.
Kod pakovanja kolone pri dodavanju svakog adsorbenta lupka se drvenim štapićem po
koloni da se adsorbent slegne. Gornji kraj adsorbenta na vrhu kolone mora biti potpuno
ravan jer je u protivnom hromatogram iskrivljen.
c) razvijanje hromatograma
Na kolonu se kroz čep postavi lijevak za odvajanjeu i u njega sipa 100-200ml petroletera,
pa polako propusti kroz kolonu. Drugi kraj kolone se spoji preko čepa sa vakuum bocom.
Kolona nikada ne smije biti suha. Nakon toga, aplicirati pripremljeni ekstrakt. Nakon toga
se razdvajanje hromatograma vrši propuštanjem smjese petroleer-benzen (4:1). U koloni se
pojavljuju obojene zone. Na gornjem dijelu sloja Al2O3 javlja se narandţasti sloj od
izdvojenih karotena; na CaCO3 ţuti sloj od ksantofila, a na saharozi dva sloja, plavo zelen
od hlorofila a i tamno zelen od hlorofila b. Kad se sve komponente dobro razdvoje, kolona
se odsisše pomoću vakuum pumpe i oprezno istrese istisne sadrţaj iz cijeli na filter papir.
Pojedine zone se mehanički odvoje, prenesu u elrenmajerice i ekstrahiraju pigmenti
smjesom eter-metanol (50:1). Adsorbenti se filtriratu, a rastvarač upari na vodenom
kupatilu.
Ako razdvajanje ksantofila i karotina ne bude potpuno, moguće ih je razdvojiti
ekstrakcjiom sa smjesom petroletera i 90% metanola. Karotin zaostaje u petroleteru, a
ksantofl prelazi u metanol.
Karotin – C40H56
Ksantofil – C40H56O2
Hlorofil a – C32H30N4OMg(COOCH3)COOC20H39
Hlorofil b - C32H28N4O2Mg(COOCH3)COOC20H39

BIOHEMIJA II–laboratorijske vjeţbe
69
3. REZULTATI
OVJERA VJEŽBE

BIOHEMIJA II–laboratorijske vjeţbe
70
Vježba broj 12
SPEKTROFLUORIMETRIJSKO ODREĐIVANJE
ANTIOKSIDATIVNOG KAPACITETA U UZORKU PĈELINJEG
MEDA
1. TEORETSKI DIO
Fluorimetrija je analitička metoda odreĎivanja supstanci zasnovana na
fluorescenticiji.Fluorescencija kao fizikalno-kemijski fenomen predstavlja proces koji se
dešava u vrlo kratkom vremenskom periodu (< 10-8
s) kada elektroni vraćajući se sa
najniţeg pobuĎenog energetskog nivoa u osnovno energetsko stanje emitiraju pri tome
energiju u vidu svjetlosti (fotona). Mnoge organske molekule apsorbiraju u UV ili pak
vidljivom valnom području, ali ne fluoresciraju. Ipak, većina od onih koje fluoresciraju, od
biološke su vaţnosti.
Dijagram energijskih razina koji pokazujeneke od energijskih promjena što se dogaĎaju pri (a) apsorpciji, (b)
relaksaciji bez pojave zračenja i (c) fluorescenciji molekulskih vrsta.
Supstance koje fluoresciraju traba da sadrţe fluoroforne grupe čijim je prisustvom
uslovljena fluorescencija. UvoĎenjem nekog supstituenta u jedinjenje moţe da pojača ili
oslabi intezitet fluorescencije. Supstituenti koji pojačavaju intezitet fluorescencije nazivaju
se auksofuori, a oni koji slabe intezitet diminofluori. Auksofluori su supstituenti koji se kad
se uvedu povećavaju delokalizaciju π elektrona. Obično se uvode ovi supstituenti sa orto i
para direkcionim grupama kao sto su fluoro, amino, alkilamino, dialkilamini, hidroksi,
metoksi grupe itd. Planarna rigidna struktura molekule ne dozvoljava pojavu vibracija i
rotacija koje rezultiraju bezemisionim gubitkom energije ekscitacije, kao što su trans
izomeri. Diminofluori djeluju kao meta direkcioni supstituenti koji pokazuju tendanciju da
lokaliziraju π elektrone kao što su karboksi, cijano, nitro i aldehidna grupa. Supstituenti kao
što su hlor, brom i jod, iako su orto i para direkcioni supstituenti, smanjuju intezitet
fluorescencije jer povećavaju unutar sistemsko ukrštanje.

BIOHEMIJA II–laboratorijske vjeţbe
71
Primjena fluorescencije je ograničena na spojeve koji posjeduju sistem dvostrukih
veza. Najintezivnije i najprimjenjljivie fluoresciraju spojevi koji imaju aromatske
prstenove, dok je veoma mali broj alifatskih i aromatskih jedinjenja sa karbonilnom grupom
koja mogu da fluoresciraju.
Razni faktori mogu uticati na intezitet fluorescencije a to su: koncentracija uzorka
koji fluorescira, prisustvo drugih supstanci koje fluoresciraju ili ne fluoresciraju, prisustvo
supstanci koje izazivaju gašenje fluorescencije, uticaj koncentracije vodikovih jona, uticaj
temperature, uticaj rastvarač, uticaj razgradnje uzorka.
Postoje dvije vrste instrumenata koje se koriste za ovu metodu: fluorimetri i
spektrofluorimetri. Razlika izmeĎu ova dva instrumenta je neznatna, fluorimetar ima filtere,
a spektrofluorometar ima prizme (monohromator) za selekciju valnih duţina pri
pobuĎivanju uzorka i prizme za selekciju valne duţine prilikom registriranja emisije
fluorescencije. Slika 2 prikazuje tipičan raspored sastavnih dijelova fluorimetra i
spektrofluorimetra.
Tipičan raspored sastavnih dijelova fluorimetra i spektrofluorimetra.
Fluorescencijski instrumenti su uglavnom dvosnopni instrumenti radi kompenzacije
fluktuacije zračenja izvora. Snop za uzorak najprije prolazi kroz primarni filter ili
monohromator , pri čemu se propušta zračenje koje uzrokuje fluorescenciju, a ograničuje ili
isključuje zračenje koje odgovara fluorescencijskim valnim duţinama. Fluorescencijsko
zračenje uzorka širi se u svim smjerovima, ali ga je najpogodnije posmatrati pod pravim
uglom u odnosu prema pobudnom snopu. Pri drugim uglovima raspršenje u otopini i na
stijenkama kivete moţe prouzročiti velike pogreške u mjerenjima inteziteta. Emitirano
zračenje nakon prolaza kroz sekundarni filter ili monohromator, kojima se za mjerenje
izolira fluorescencijski maksimum, dolazi do fotoelektričnog detektora. Referentni snop
prolazi prigušivačem kojime se postiţe smanjenje njegove snage na pribliţno onu
vrijednost koju ima fluorescencijsko zračenje. Signali referentnog fotodetektora i onog za
uzorak procesiraju se diferencijalnim pojačalom čiji se signal očituje na mjeraču ili pisaču.
Fluorimetri su osjetljiviji od spektrofluorimetara jer filteri imaju veću propustljivost
zračenja od monohromatora. Izvor i detektor se u jednostavnijim instrumentima mogu
smjestiti pono bliţe uzorku, čime se postiţe veća osjetljivost.
Širina primjene fluorimetrijskih metoda za analizu organskih tvari je veoma impresivna.

BIOHEMIJA II–laboratorijske vjeţbe
72
Fluorescencija kao pojava našla je primjenu npr. u medicini, forenzici, mineralogiji i
kristalografiji, proizvodnji svijetlećih tijela itd. Velike biomolekule mogu imati
fluorescentnu kemijsku grupu vezanu kemijskom reakcijom, a fluorescencija te grupe
omogućava vrlo osjetljivu detekciju molekule.
2. EKSPERIMENTALNI DIO
Potrebni reagensi:
- Uzorak za analizu-Pčelinji med
- Standardi-6-hidroksi-2,5,7,8-tetrametilkroman-2-karboksilna kiselina (troloks),
«Sigma Aldrich
» Chemie GmbH, Njemačka,(sintetski vodotopivi analog vitamina
E.
- fluorescein «FLUKA» Chemika GmbH, Steinheim, Germany;
- hidrogenperoksid, 33-35 %, ''Semikem'', Sarajevo
- bakar(II)-sulfat-pentahidrat, p.a. Kemična tovarna podnart Slovenija
Potrebna oprema i pribor:
- vaga
- menzura
- čašice ili posudice za vaganje
- odmjerni sudovi
- pH-metar
- mikropipete
- termostat
- kivete za uzorke
Za analizu će biti korištena metoda oksigen radikal apsorpcijskog kapaciteta
(ORAC). Ova metoda moţe kvantitativno mjeriti ukupni AC biološkog uzorka kao npr.
ljudskog i ţivotinjskog seruma, tkiva, poljoprivrednih i prehrambenih proizvoda, sastojaka
hrane, farmaceutskih proizvoda itd. Metoda mjeri efektivnost različitih prirodnih
antioksidanata prisutnih u uzorku, u spriječavanju gubitka intenziteta fluorescencije
fluoresceina (FL) ili fluorescentnog marker proteina -pikoeritrina (beta-PE) tokom
njihovog izlaganja djelovanju FR.
Struktura fluoresceina
Rezultati ORAC metodom postiţu se dovodeći reakciju do kraja i integrirajući
površinu ispod kinetičke krive relativno prema reakciji slijepe probe koja ne sadrţi dodane
antioksidante. Površina ispod krive srazmjerna je koncentraciji svih antioksidanata
prisutnih u uzorku. Svaka reakcija se kalibrira upotrebom standarda troloksa (Trolox
), koji
je sintetski vodotopivi vitamin E analog.

BIOHEMIJA II–laboratorijske vjeţbe
73
Podaci se proslijeĎuju elektronski od Perkin-Elmer luminiscentnog spektrometra LS
55 na računarski sistem opremljen softverskim programom FL WinLab i analiziraju
programom Microsoft Excel 2000 primjenjujući jednačine (1) i (2) za površinu ispod
fluorescentne opadajuće krive:
..../, / PSSPSUgmolTE SSSSkORAC
gdje je : ORAC - oksigen radikal apsorpcijski kapacitet izraţen u troloks ekvivalentima
(TE, mol/g)
k - faktor razrijeĎenja otopine;
US - površina ispod fluorescentne opadajuće krive za uzorak
..PSS - površina ispod fluorescentne opadajuće krive za slijepu probu
SS - površina ispod fluorescentne opadajuće krive za standard
Površina integrala ispod fluorescentne opadajuće krive za uzorak, standard ili slijepu probu
izračunava se po obrascu za izračunavanje površine trapeza na slijedeći način:
iiii ffttffttffttS 1112120101 ...2
1
gdje je:
S - površina integrala ispod fluorescentne opadajuće krive za uzorak, standard ili
slijepu probu
t - vrijeme inkubacije
f - relativni intenzitet fluorescencije
a) Priprema uzorka
U čistoj i suhoj čašici odvagati na analitičkoj vagi oko 1 g uzorka meda, zapisati tačnu
masu odvage, dodati na uzorak 10 ml destilirane vode i miješati staklenim štapićem do
otapanja. Otopinu sa uzorkom prenijeti kvantitativno u odmjerni sud od 25 ml uz ispiranje
čaše sa još 10 ml destilirane vode. Nakon što se i taj dio prenese u odmjerni sud, otopina se
protrese i dopuni do oznake od 25,00 ml pomoću špric balona. Od tako pripremljenog
uzorka uzeti 100 l, razrijediti ih sa 900 l destilirane vode i nakon miješanja na vorteksu
koristiti za analizu.
b) Priprema standarda, fluoresceina i generatora peroksidnih radikala
Pripremiti slijedeće otopine:
4,00 ml 1 mM otopine troloksa (na odvagu troloksa dodati 100 l etanola, a zatim
3,90 ml destilirane vode); zatim razrijeĎivanjem u vodi pripremiti 1,00 ml 20 M
otopine troloksa.

BIOHEMIJA II–laboratorijske vjeţbe
74
10,00 ml 0,64 M otopine fluoresceina (FL);
1,00 ml 2,2 M H2O2;
10,00 ml 72 mM CuSO4 u destiliranoj vodi.
c) Eksperimentalna procedura
U kivetu za mjerenje dodati 50,00 l 0,64 M otopine FL, 100 l 20 M otopine troloksa
ili uzorka meda (razrijeĎenog na odgovarajući način), 1750 l destilirane vode, 50 l 72
mM otopine Cu2+
, postaviti kivetu u aparat za mjerenje,a odmah zatim 50 l 2,2 M otopine
H2O2 i promješati sadrţaj. Nakon toga mjeriti intezitet fluorescencije svakih 10 minuta u
roku od sat vremena.
Slijepa proba:
50,00 l 0,64 M otopine FL, 1850 l destilirane vode i nakon termostatiranja 50 l 72
mM otopine Cu2+
+ 50 l 2,2 M otopine H2O2).
NAPOMENA
Prije rada upoznati se sa načinom rukovanja kemikalijama koje se koriste. Podaci o
štetnosti kemikalija, kao i načinu pruţanja prve pomoći nalaze se u laboratoriju
(konsultovati asistenta i laboranta)! Ovdje su navedene neke informacije za supstance
povećanog rizika:
- Hidrogenperoksid Hidrogenperoksid izaziva oštećenja na koži i stoga je upotreba zaštitnih rukavica obavezna pri radu sa ovom supstancom!
3. ZADACI I VJEŽBE
1. Šta je fluorescencija, a šta fosforescencija?
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
2. Koji su glavni nedostaci spektrofluorimetrije kao metode?
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________
3. Iz kojih razloga se Cu2+
i H2O2 u korištenoj metodi ne dodaju u smjesi već posebno?
_______________________________________________________
_______________________________________________________
_______________________________________________________
_______________________________________________________

BIOHEMIJA II–laboratorijske vjeţbe
75
4. REZULTATI
OVJERA VJEŽBE
Slijepa
proba
Standard
Površina
ORAC





























