Embed Size (px)
Citation preview

Vol. 40, No. 3INFECTION AND IMMUNITY, June 1983, P. 977-9840019-9567/83/060977-08$02.00/0Copyright 0 1983, American Society for Microbiology
T and B Lymphocyte Susceptibility to Murine Leukemia VirusMoloney
DALE D. ISAAKl* AND JAN CERNY2Department of Microbiology and Immunology, Kirksville College of Osteopathic Medicine, Kirksville,
Missouri 635011; and Department of Microbiology, University of Texas Medical Branch, Galveston, Texas775502
Received 7 June 1982/Accepted 4 February 1983
The susceptibility ofT and B lymphocytes to productive infection and transfor-mation by murine leukemia virus Moloney was determined by enumeration ofcells producing infectious virus after in vitro infection of mitogen-stimulated,isolated cell populations and by in vivo infection of euthymic BALB/c andthymus-deficient (nude) mice. Our in vitro results demonstrated that the majorityof splenic T cells and thymocytes are resistant to productive infection in vitro; aspecific subpopulation of susceptible nylon-adherent splenic T cells was identi-fied, however. Similarly, surface immunoglobulin-positive B cells also representsusceptible targets in vitro; mature B cells, however, did not represent theprincipal target for transformation in the in vivo experiments. Infected euthymicmice expressed increasing titers of murine leukemia virus and uniformly devel-oped fatal T-cell lymphomas at 10 to 12 weeks postinfection; nude mice, incontrast, maintained high, stable levels of viremia throughout the 28 weeks ofobservation. Infected nude mice remained free of malignancy or developed eithergranulocytic leukemias or, in one case, reticulum cell sarcoma. Collectively, theresults indicate that While the majority of T cells are resistant to productiveinfection, they represent the principle targets for transformation; B cells, howev-er, represent permissive targets for virus replication, but are resistant to transfor-mation.
Murine leukemia virus-Moloney (MuLV-M)induces a fatal lymphatic leukemia within sever-al months after infection of newborn mice (24).The disease is characterized by enlarged thymusglands, hepatosplenomegaly, and enlargementof both peripheral and mesenteric lymph nodes.The neoplastic process begins in the thymus (11,32) and cell lines established from leukemic miceexpress both Thy 1.2 and MuLV-M-specific cell-associated antigens (27, 32), indicating that thevirus infects and transforms T cells.However, several studies have provided evi-
dence that MuLV-M also infects antibody-form-ing cells (i.e., B cells). In mice infected withMuLV-M and then immunized with an unrelatedantigen, individual antibody-forming cells wereshown to contain viral glycoprotein(s) detect-able by immunofluorescence (10), virus particles(26), and virus-specific, cell-associated surfaceantigen(s) detectable by cytotoxic treatmentwith MuLV-M-specific antibody plus comple-ment (5).Thus, it appears that MuLV-M infection may
offer a model of leukemia virus restriction inwhich both T and B lymphocyte classes may beinfected, but only the T cell is transformed.
Since the evidence for B cell permissiveness isindirect, however, and no experiments on trans-formation of B cells by MuLV-M have beenreported, we have examined this issue morethoroughly. First, we determined the ability ofMuLV-M to infect and replicate in cultures ofmitogen-activated, purified splenic T and B cellsand in thymocytes. Second, we examined thevirus replication and the leukemogenicity incongenitally thymus-deficient nude mice, whichhave normal B cells but lack differentiated Tcells. The results of these studies indicate adichotomy between ecotropic MuLV infectionand transformation of target cells within somelymphocyte subclasses.
MATERIALS AND METHODSAnimals. BALB/c as well as nude mice were derived
from breeding stock maintained in our animal facili-ties. BALB/c stock was obtained initially from CharlesRiver Laboratories, Inc., Wilmington, Mass.; nudemice and their phenotypically normal, euthymic litter-mates (LM) were the progeny of heterozygous breed-ing stock (derived from mice supplied by N. D. Reed,Montana State University, Bozeman) backcrossed 10generations on the BALB/c strain.
977
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

978 ISAAK AND CERNY
Virus. The origin of our stock MuLV-M was de-scribed previously (9). Virus passages were preparedas 10% (wt/vol) cell-free homogenates from thespleens of infected, leukemic BALB/c mice and werecharacterized for their content of infectious MuLV bydetermining the number of focus-forming units (FFU)per milliliter on sarcoma virus-positive, leukemia vi-rus-negative (S+L-) indicator fibroblasts (3), as de-scribed previously (6); virus stocks routinely con-tained approximately 106 FFU of ecotropic MuLV perml. In addition, the content of mink lung cytopathicfocus-forming virus, determined by focus induction onCCL-64 indicator cells as previously described (15),was approximately 4.4 x 103 infectious units per ml.
Mitogens. Escherichia coli strain 0127:B8 lipopoly-saccharide (LPS) (Difco Laboratories, Detroit, Mich.)was dialyzed against deionized water, lyophilized,solubilized in 0.15 M NaCl, and boiled for 2 h beforeuse; the optimum mitogenic concentration, 10 p.g/ml ofculture medium, was used. Concanavalin A (ConA)(grade IV; Sigma Chemical Co., St. Louis, Mo.) wasused at its optimum mitogenic concentration, 2 ,ug/mlof culture medium, or as otherwise indicated.
Separation of spleen cells on nylon fiber columns. Bcell-enriched and T cell-enriched fractions were ob-tained from BALB/c spleen cell suspensions by amodification (6) of the procedure of Handwerger andSchwartz (14). The principal modification was thatcells were filtered through two consecutive nylon fibercolumns. The adherent fraction from the second col-umn was enriched for B cells in that >85% of therecovered cells were surface immunoglobulin-positive(sIg+) after membrane immunofluorescence stainingwith goat anti-mouse immunoglobulin conjugated tofluorescein isothiocyanate, but reproducibly containeda low percentage of nylon wool-adherent T cells. Thenonadherent fraction from the second column wasjudged to be greater than 90%o T cells by sensitivity tolysis with AKR/J anti-C3H/HeJ Thy 1.2 plus comple-ment treatment; it contained <10% sIg+ cells.
Separation of sIg+ and sIg- ceils on immunoadsor-bent columns. Immunoadsorbent columns preparedwith rabbit anti-mouse F(ab')2-conjugated SephadexG200 were used to obtain sIg+ and sIg- cells aspreviously described (30). Greater than 98% of therecovered adherent cells were sIg+, whereas less than2% of the effluent, nonadherent cells were sIg+ afterstaining with fluoresceinated rabbit anti-mouse(F(ab')2 antiserum.
Cytotoxic treatment. sIg+ cells were removed fromspleen cell suspensions via a two-step cytolytic proce-dure involving treatment with rabbit polyvalent anti-mouse immunoglobulin and guinea pig complement aspreviously described (19). The T cell origin of cellsreleasing MuLV (infectious centers) was tested byspecific cytotoxic treatment with anti-Thy 1.2 anti-body plus complement.
Cell cultures. Single cell suspensions of normal,uninfected lymphocytes were prepared in RPMI 1640medium supplemented with 2 mM HEPES (N-2-hy-droxyethylpiperazine-N'-2-ethanesulfonic acid) buff-er, 2 mM glutamine (Microbiological Associates, Be-thesda, Md.), 5% fetal calf serum, antibiotics (100 U ofpenicillin, 100 ,ug of streptomycin, and 25 ,ug offungizone per ml; GIBCO Laboratories, Grand Island,N.Y.) and 2 ,ug of polybrene (Aldrich Chemical Co.,Milwaukee, Wis.) per ml of culture medium.
For in vitro infection of cells, 107 pelleted cells weresuspended in 0.2 ml of a 1:2 dilution of MuLV-M (-105FFU; multiplicity of infection, 102 FFU ofMuLV pernucleated cell); supplemented RPMI medium with 4,ug of polybrene per ml was used to dilute the virusstocks. Cell-virus mixtures were incubated at 4°C for40 min and then suspended in 4 ml of medium ormedium containing LPS or ConA; cultures were incu-bated at 37°C in humidified air with 5% CO2 until thecells were harvested and assayed for the presence ofcells releasing infectious MuLV. Cultures were set induplicate in 17- by 100-mm plastic culture tubes (Fal-con Plastics, Oxnard, Calif.). The mitogen stimulationof lymphocytes was assayed by [3H]thymidine uptakeas previously described (9). Data are expressed as thestimulation index, the mean counts per minute incultures of 106 cells with mitogen divided by the meancounts per minute in unstimulated cultures.
Infection of mice. Within 24 h of birth, nude and LMmice were injected intraperitoneally with 1 x 104 to 2x 104 FFU of MuLV-M. On various days postinfec-tion, peripheral blood was harvested from a small cutof the ventral tail artery by using calibrated glassmicrocapillary tubes rinsed in complete medium con-taining 10 U of heparin per ml. The microliter sampleswere diluted 10-fold in culture medium and assayedfresh or after storage at -70°C.
Assay for IC and MuLV. Cell suspensions recoveredfrom in vitro infection cultures were assayed for thepresence of MuLV-releasing cells in an infectiouscenter (IC) assay on S+L- indicator fibroblasts aspreviously described (6). Recovered cells from dupli-cate cultures were washed three times in supplement-ed medium, counted, and assayed at a minimum of twodilutions (each in duplicate) on dishes seeded previ-ously with S+L- fibroblasts. Individual centers offibroblast transformation, reflecting the number ofMuLV-releasing cells, were scored on day 5 and areexpressed as IC/106 viable lymphocytes plated. Thenumber of IC was proportional to the lymphocytedilution. Data represent means from dishes with opti-mal IC numbers (30 to 70 IC/dish).
In addition, serial dilutions of peripheral blood re-covered from MuLV-M-infected nude and LM micewere assayed on S+L- cells. The foci of transforma-tion were expressed as the number of FFU of MuLVper ml of blood assayed. Since whole blood wasassayed, the observed foci were the result of freeMuLV and circulating, virus-releasing blood cells.
RESULTSThe results from representative experiments
on the infection of various lymphocyte popula-tions by MuLV-M in vitro are shown in Table 1.The infection of normal spleen cells was en-hanced almost 10-fold by the addition of ConA,a T cell-specific mitogen, into the culture medi-um. The effect seen on day 3 after infectionappeared to be transitory, however, since thenumber of IC in ConA-stimulated cultures onday 6 decreased to the level observed in unsti-mulated cultures. Stimulation with the B cell-specific mitogen LPS also resulted in enhancedinfection, but the effect was not transitory. LPS
INFECT. IMMUN.
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
LEUKEMIA TARGET CELLS 979
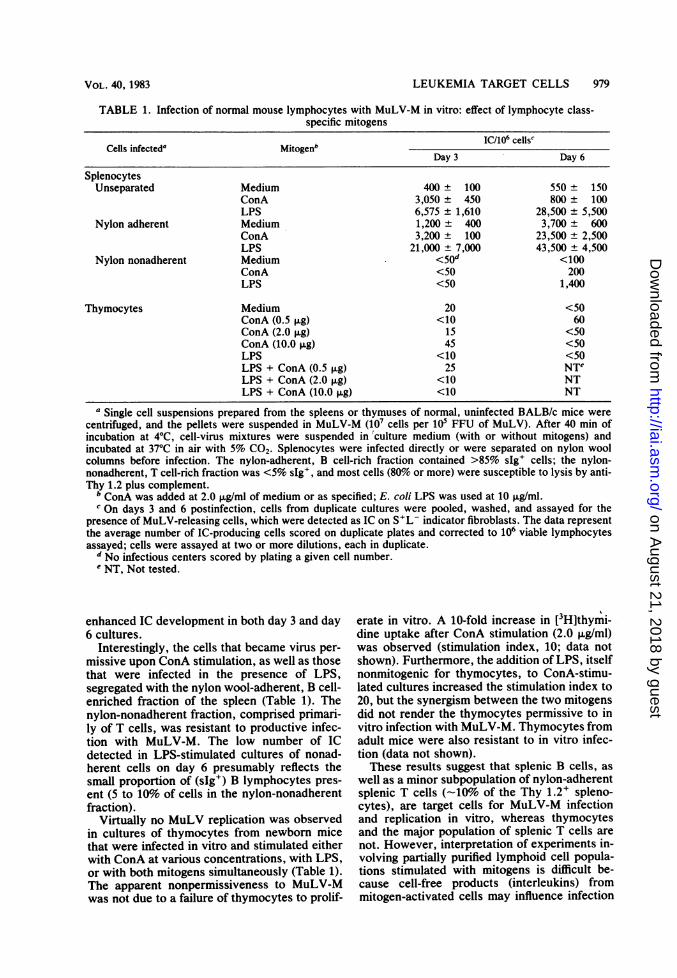
TABLE 1. Infection of normal mouse lymphocytes with MuLV-M in vitro: effect of lymphocyte class-specific mitogens
Cells infected' Mitogenb IC/10' cells'Day 3 Day 6
SplenocytesUnseparated Medium 400 ± 100 550 ± 150
ConA 3,050 ± 450 800 ± 100LPS 6,575 ± 1,610 28,500 ± 5,500
Nylon adherent Medium 1,200 ± 400 3,700 ± 600ConA 3,200 ± 100 23,500 ± 2,500LPS 21,000 ± 7,000 43,500 ± 4,500
Nylon nonadherent Medium <50d <100ConA <50 200LPS <50 1,400
Thymocytes Medium 20 <50ConA (0.5 1lg) <10 60ConA (2.0 jig) 15 <50ConA (10.0 pg) 45 <50LPS <10 <50LPS + ConA (0.5 F±g) 25 NTeLPS + ConA (2.0 ,ug) <10 NTLPS + ConA (10.0 ,ug) <10 NT
a Single cell suspensions prepared from the spleens or thymuses of normal, uninfected BALB/c mice werecentrifuged, and the pellets were suspended in MuLV-M (107 cells per 10' FFU of MuLV). After 40 min ofincubation at 4°C, cell-virus mixtures were suspended in'culture medium (with or without mitogens) andincubated at 37°C in air with 5% CO2. Splenocytes were infected directly or were separated on nylon woolcolumns before infection. The nylon-adherent, B cell-rich fraction contained >85% sIg+ cells; the nylon-nonadherent, T cell-rich fraction was <5% sIg', and most cells (80% or more) were susceptible to lysis by anti-Thy 1.2 plus complement.
b ConA was added at 2.0 p.g/ml of medium or as specified; E. coli LPS was used at 10 ,ug/ml.c On days 3 and 6 postinfection, cells from duplicate cultures were pooled, washed, and assayed for the
presence of MuLV-releasing cells, which were detected as IC on S+L- indicator fibroblasts. The data representthe average number of IC-producing cells scored on duplicate plates and corrected to 106 viable lymphocytesassayed; cells were assayed at two or more dilutions, each in duplicate.
d No infectious centers scored by plating a given cell number.e NT, Not tested.
enhanced IC development in both day 3 and day6 cultures.
Interestingly, the cells that became virus per-missive upon ConA stimulation, as well as thosethat were infected in the presence of LPS,segregated with the nylon wool-adherent, B cell-enriched fraction of the spleen (Table 1). Thenylon-nonadherent fraction, comprised primari-ly of T cells, was resistant to productive infec-tion with MuLV-M. The low number of ICdetected in LPS-stimulated cultures of nonad-herent cells on day 6 presumably reflects thesmall proportion of (sIg+) B lymphocytes pres-ent (5 to 10% of cells in the nylon-nonadherentfraction).
Virtually no MuLV replication was observedin cultures of thymocytes from newborn micethat were infected in vitro and stimulated eitherwith ConA at various concentrations, with LPS,or with both mitogens simultaneously (Table 1).The apparent nonpermissiveness to MuLV-Mwas not due to a failure of thymocytes to prolif-
erate in vitro. A 10-fold increase in [3H]thymi-dine uptake after ConA stimulation (2.0 ,ug/ml)was observed (stimulation index, 10; data notshown). Furthermore, the addition of LPS, itselfnonmitogenic for thymocytes, to ConA-stimu-lated cultures increased the stimulation index to20, but the synergism between the two mitogensdid not render the thymocytes permissive to invitro infection with MuLV-M. Thymocytes fromadult mice were also resistant to in vitro infec-tion (data not shown).These results suggest that splenic B cells, as
well as a minor subpopulation of nylon-adherentsplenic T cells (-10% of the Thy 1.2+ spleno-cytes), are target cells for MuLV-M infectionand replication in vitro, whereas thymocytesand the major population of splenic T cells arenot. However, interpretation of experiments in-volving partially purified lymphoid cell popula-tions stimulated with mitogens is difficult be-cause cell-free products (interleukins) frommitogen-activated cells may influence infection
VOL. 40, 1983
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

980 ISAAK AND CERNY
TABLE 2. Effect of treatment with rabbit anti-mouse immunoglobulin plus complement on mitogen-enhanced in vitro infection of BALB/c spleen cells
with MuLVWMaTreatmentb Mitogenc IC/106 cellsd
Untreated Medium 100 ± 0
ConA 1,075 ± 258LPS 6,300 ± 1,745
Rabbit anti-immuno- Medium 5 ± 5globulin plus com- ConA 1,500 ± 552plement LPS 650 ± 150
Normal rabbit serum Medium 250 ± 50plus complement ConA 950 ± 150
LPS 2,725 ± 858
a Treated or untreated spleen cells from normal,noninfected B3ALB/c donor mice were adjusted to 107cells per culture, infected in vitro with MuLV-M, andcultured as in Table 1.
b Surface immunoglobulin-positive cells were de-pleted from spleen cell suspensions in a two-stepcytolytic treatment with rabbit anti-mouse immuno-globulin plus complement. Cells were resuspended inantiserum appropriately diluted in saline containing0.01 M sodium azide, incubated at 0°C for 45 min,centrifuged, and suspended in guinea pig complementdiluted 1:3. After 45 min at 37°C, the cells werewashed and infected. Control cells were treated withnormal rabbit serum plus complement or left untreat-ed.
c LPS was used at 10 ,ug/ml of culture medium, andConA was used at 2 ,ug/ml of culture medium.d On day 3 postinfection, recovered cells were
washed, and known numbers of cells were assayed forthe presence of MuLV-releasing cells. The data repre-sent average numbers of IC scored on duplicate plates,corrected to 106 viable lymphocytes assayed.
of other cell types. To confirm that LPS-stimu-lated B cells were the target cells in question,two approaches were used. First, normal spleencells were treated with rabbit anti-mouseimmunoglobulin antiserum plus complement be-fore infection. Such pretreatment reduced thenumber of IC seen in LPS-stimulated cultures by89% compared with untreated spleen cell cul-tures (650 versus 5,700 IC/106 cells) and by 77%compared with treatment with normal rabbitserum and complement (650 versus 2,725 IC/106)(Table 2).
In a second approach, splenic B cells werepurified with immunoadsorbent columns pre-
pared with goat anti-mouse F(ab')2-conjugatedSephadex G200. The elution of adherent cellswith mouse F(ab')2 resulted in the recovery ofhighly purified B cells (>98% of the cells were
sIg+). LPS enhanced the in vitro infection ofboth unseparated spleen cells and the sIg+ Bcells (10,000 and 14,538 IC/106 cells, respective-ly) (Table 3). The enrichment of permissive cellsin the sIg+ fraction (45%) was less than atwofold increase expected after elimination of
TABLE 3. LPS-enhanced MuLV-M infection in Bcells purified on immunoadsorbent columnsaCells infectedb Mitogen IC/106 cellsc
Unseparated spleen Medium 250 ± 20LPS 10,000 ± 2,000
Immunoglobulin-positive Medium NDdcells LPS 14,538 ± 1,125a Unseparated spleen cells or purified splenic B
cells isolated on immunoadsorbent columns preparedwith goat anti-mouse F(ab')2 conjugated to SephadexG200 were infected in vitro with MuLV-M and incu-bated in medium with or without LPS.
b Cells adhering to the immunoadsorbent columnswere eluted with mouse F(ab')2 and assayed for puritybefore in vitro infection. Greater than 98% of the cellswere positive for membrane immunoglobulin afterstaining with fluorescinated goat anti-mouse immuno-globulin.
I On day 4 postinfection cells from duplicate cul-tures were pooled, washed, and assayed for the pres-ence of MuLV-releasing cells (detected as infectiouscenters on S+L- cells). Cells were assayed at two ormore dilutions, each in duplicate.
d ND, Not done.
50% of spleen cells as sIg-. This reflects theexistence of sIg-, LPS-responsive target cells inthe spleen which we observed in our earlierexperiments (19). Thus the data show that puri-fied sIg+ B cells are a major, but not the only,target for MuLV-M.A specific cytotoxic treatment was used to
demonstrate that the splenocytes that becamepermissive to MuLV-M upon ConA stimulationwere indeed T cells (Table 4). Normal spleencells were infected and cultured in medium withConA for 3 days. Before the assay on S'L-fibroblasts, the infected lymphocytes were incu-bated with anti-Thy 1.2 antibody plus comple-ment. The treatment reduced the number ofdetectable IC by 75%, compared with controlcells treated with normal mouse serum pluscomplement (500 versus 2,000 IC/106 cells). Be-cause cells in each group were adjusted to equalnumbers of viable cells before assay, this repre-sents a minimum estimate of Thy 1.2+ targets.The observation that cultured B lymphocytes
are permissive for MuLV-M (Tables 2 and 3)prompted studies on MuLV-M replication andits leukemogenic potential in vivo with nudemice, which have normal B lymphocytes butlack both thymocytes and mature T cells. Pre-liminary experiments established that the in vi-tro infection of LPS-stimulated spleen cells fromnude mice by MuLV-M was quantitatively simi-lar to that observed with cells from euthymicmice (unpublished results from our laboratory).Nude mice and LM mice were infected with
MuLV-M within 24 h of birth. At various times
INFECT. IMMUN.
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
LEUKEMIA TARGET CELLS 981
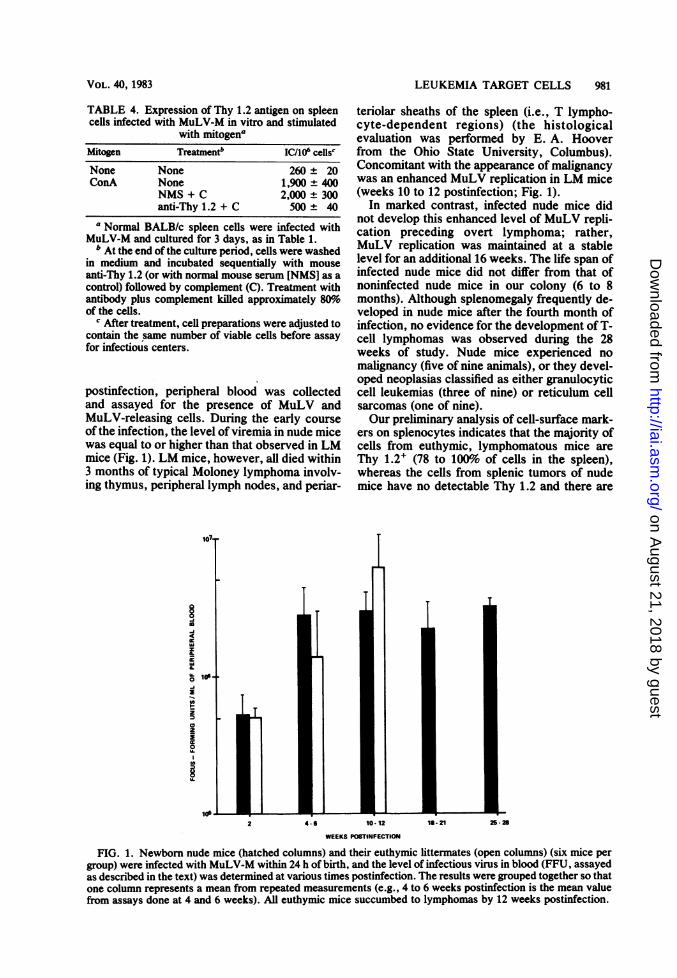
TABLE 4. Expression of Thy 1.2 antigen on spleencells infected with MuLV-M in vitro and stimulated
with mitogenaMitogen Treatmenth IC/106 celis'None None 260 t 20ConA None 1,900 t 400
NMS + C 2,000 t 300anti-Thy 1.2 + C 500 ± 40
a Normal BALB/c spleen cells were infected withMuLV-M and cultured for 3 days, as in Table 1.
b At the end of the culture period, cells were washedin medium and incubated sequentially with mouseanti-Thy 1.2 (or with normal mouse serum [NMS] as acontrol) followed by complement (C). Treatment withantibody plus complement killed approximately 80%oof the cells.
c After treatment, cell preparations were adjusted tocontain the same number of viable cells before assayfor infectious centers.
postinfection, peripheral blood was collectedand assayed for the presence of MuLV andMuLV-releasing cells. During the early courseof the infection, the level of viremia in nude micewas equal to or higher than that observed in LMmice (Fig. 1). LM mice, however, all died within3 months of typical Moloney lymphoma involv-ing thymus, peripheral lymph nodes, and periar-
io7
oU.
U.
teriolar sheaths of the spleen (i.e., T lympho-cyte-dependent regions) (the histologicalevaluation was performed by E. A. Hooverfrom the Ohio State University, Columbus).Concomitant with the appearance of malignancywas an enhanced MuLV replication in LM mice(weeks 10 to 12 postinfection; Fig. 1).
In marked contrast, infected nude mice didnot develop this enhanced level of MuLV repli-cation preceding overt lymphoma; rather,MuLV replication was maintained at a stablelevel for an additional 16 weeks. The life span ofinfected nude mice did not differ from that ofnoninfected nude mice in our colony (6 to 8months). Although splenomegaly frequently de-veloped in nude mice after the fourth month ofinfection, no evidence for the development of T-cell lymphomas was observed during the 28weeks of study. Nude mice experienced nomalignancy (five of nine animals), or they devel-oped neoplasias classified as either granulocyticcell leukemias (three of nine) or reticulum cellsarcomas (one of nine).Our preliminary analysis of cell-surface mark-
ers on splenocytes indicates that the majority ofcells from euthymic, lymphomatous mice areThy 1.2+ (78 to 100% of cells in the spleen),whereas the cells from splenic tumors of nudemice have no detectable Thy 1.2 and there are
FIG. 1. Newborn nude mice (hatched columns) and their euthymic littermates (open columns) (six mice pergroup) were infected with MuLV-M within 24 h of birth, and the level of infectious virus in blood (FFU, assayedas described in the text) was determined at various times postinfection. The results were grouped together so thatone column represents a mean from repeated measurements (e.g., 4 to 6 weeks postinfection is the mean valuefrom assays done at 4 and 6 weeks). All euthymic mice succumbed to lymphomas by 12 weeks postinfection.
VOL. 40, 1983
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

982 ISAAK AND CERNY
very few sIg+ cells (Isaak, Asjo, and Cerny,experiments in progress). It should be noted thatthe injection of newborn LM mice with MuLVpresent in the blood of infected, viremic, butlymphoma-free, nude mice resulted in the devel-opment of typical Moloney lymphomas (8).
DISCUSSIONOur results demonstrate that sIg+ lympho-
cytes (B cells) are permissive to ecotropicMuLV-M. Although the number of cells thatbecame infected after in vitro infection and LPSstimulation varied from one experiment to an-other, up to 5% of lymphocytes in the B cell-richfraction released infectious MuLV (Table 1).Because the host environment supports cell-virus interactions better than culture tubes (7),and because cells are continuously exposed tohigh titers of MuLV, it is reasonable to assumethat the proportion of B cells infected in vivowould be even higher. Indeed, when lympho-cytes from the spleen and bone marrow ofMuLV-M-infected, viremic nude or euthymicmice were assayed on S+L- indicator cells,between 20 and 80% of cells released infectiousMuLV (Isaak and Cerny, manuscript in prepara-tion). The replication of infectious MuLV inmice with either phenotype was comparable(Fig. 1); furthermore, the virus from the blood ofnude mice readily induced lymphomas in euthy-mic recipients (8). Yet nude mice did not devel-op lymphoblastic leukemias involving mature Bcells, based on morphological and histologicalcriteria as well as on preliminary screenings forsIg. Thus sIg+ B cells represent permissivetarget cells for MuLV-M with respect to virusinfection, integration, and replication, but notwith respect to transformation. A possiblemechanism of this restriction in mature B cells isdiscussed below. The development of granulo-cytic leukemias was observed in a proportion ofnude mice. In addition, 1 of 9 infected nude micedeveloped a reticulum cell sarcoma, which isthought to be related to the B cell lineage (22).This indicates that some bone marrow-derivedcells may represent alternative targets for trans-formation by MuLV-M, but the uniform devel-opment of T-cell lymphomas in euthymic miceemphasizes the preference of the virus for thy-mus-derived cells.
Ricciardi-Castagnoli et al. (29) reported thedevelopment of lymphomas in nude mice infect-ed with radiation leukemia virus (Rad LV), anagent that typically induces thymic lymphomas(13, 21). Cell lines established from these lym-phomas expressed Thy 1.2 antigen, suggestingthe infection with Rad LV may lead to differenti-ation of immature T cell precursors in nude miceand their transformation in the absence of thy-mus. We have not observed the development of
T-cell lymphomas in MuLV-M-infected nudemice during their life span (8 months), and ourpreliminary studies failed to demonstrate thepresence of Thy 1.2 on cells from the enlargedspleens of MuLV-M-infected nude mice thatwere evaluated as lymphoma negative by histo-logical criteria.The target cells for transformation by MuLV-
M are thought to be lymphocytes of the T-celllineage (11, 20, 24, 27, 28, 33), in part becausetransformed thymus cells express a receptor forMuLV-M (2) and produce infectious virus (27).Therefore, we expected a large proportion ofthymocytes or splenic T cells to be permissive tothe virus after in vitro infection. We found,however, that less than 0.005% of cells in cul-tures of metabolically active thymocytes (<50infectious centers per 106 cells; Table 1) werepermissive. This corresponds with the finding ofBaird et al. (2) that only 2% or less of normalthymocytes have specific binding sites forMuLV-M, whereas >90% of MuLV-M-trans-formed cells bind the virus. Baird et al. speculat-ed that MuLV-M-induced lymphomas may orig-inate from a rare subpopulation of permissivecells. In addition to thymocyte resistance, themajority of splenic T cells, recovered as nylon-nonadherent cells, were also nonpermissive toMuLV-M infection in our primary lymphocytecultures. Similarly, Horak et al. found that celllines established from nylon-nonadherent T cellswere resistant to infection with various ecotro-pic MuLV, including MuLV-Moloney (16). Thelack of virus replication in T cells may in part bedue to a transcriptional block, based on theresults of Jaenisch et al. (20), which demonstrat-ed the presence of multiple copies of virusgenome in various tissues of mice infected withMuLV-M at the embryonic stage, but high levelsof viral RNA were found only in overt thymoma.
In contrast to the resistance observed in ny-lon-nonadherent T cells, our present resultsdemonstrate the existence of a quantitativelyminor subpopulation of Thy 1.2+, ConA-respon-sive splenic T cells that are susceptible to pro-ductive infection. Subsequent characterizationindicated that these susceptible targets segregat-ed in the nylon-adherent fraction, but directattempts to demonstrate Thy 1.2 on the surfaceof nylon-adherent, ConA-responsive, suscepti-ble targets by lysis with anti-Thy 1.2 plus com-plement resulted only in a 40% reduction in IC(data not shown). Previous demonstration of thevery low density of Thy 1.2 on nylon wool-adherent, ConA-responsive splenic T cells byother investigators (23) may in part explain ourinability to effectively inhibit IC development bysuch cytotoxic treatments.
Recently it has been pointed out that Thy 1antigen expression in cultures of mouse hemo-
INFECT. IMMUN.
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

LEUKEMIA TARGET CELLS 983
poietic cells is not limited to T cells and in factmay be induced, in the presence of mitogen-activated T cell culture supernatants, on cellsknown not to be T cells (31); in this system, bonemarrow cells depleted of T cells and cultured inconditioned medium derived from ConA-acti-vated T-cell cultures subsequently developedThy 1+ cells related to the granulocytic series.Thus, we must consider the possibility that thesusceptible Thy 1.2' cells we have identified arenot T cells. In addition to expression of Thy 1.2,however, the targets described herein share anumber of other characteristics with T cells;they are ConA-responsive, and they appear tobe thymus derived since they are absent in nudemice. Also, we have not been able to enhanceinfection of nude mouse spleen cells by usingconditioned media from ConA-activated T cellcultures. The latter observations suggest that weare not dealing with an unusual, non-T-cell tar-get induced by lymphokines secreted from mito-gen-activated T cells.
Thus, it seems that the susceptible Thy 1.2+target cells we have identified represent a uniquesubpopulation ofT cells. It is interesting that we(18) and other investigators (23) have describeda similar pattern of susceptibility versus resist-ance in nylon-adherent versus nonadherent Tcells using an unrelated ecotropic MuLV,MuLV Friend. If productive infection is a neces-sary step in transformation by MuLV, as sug-gested by Ihle (17), such a virus-permissivesubset of T cells may represent the actual targetcells for lymphomagenesis as well as for produc-tive infection by MuLV-M.
Collectively, the data indicate that althoughcells from both T and B lineages are permissiveto MuLV-M infection, T cells become trans-formed but B cells do not. An interpretation ofthe apparent cellular restriction of transforma-tion should take into consideration that MuLV-M belongs to the group of leukemia-inducingretroviruses that are apparently incapable ofdirect cell transformation. Rather, in the courseof prolonged replication in the host, these virus-es may recombine with endogenous xenotropicretroviruses to result in transforming agents (12,15, 25, 34). One can speculate that there is aheterogeneity among the endogenous viruses indifferent lymphocyte classes and that a givenexogenous MuLV may form a recombinant withonly some of them. Thus, MuLV-M may formthe "appropriate" transforming recombinantwith endogenous virus in T cells, but not B cells.We have examined the expression of endoge-nous, xenotropic virus (X) in various lymphoidtissues of MuLV-M-infected mice. In apparentaccordance with the pathogenesis, euthymicmice expressed the highest values of X in thethymus, an observation consistent with that of
Asjo et al. (1), whereas nude mice had thehighest level ofX in bone marrow (8) (Isaak andCerny, manuscript in preparation). The fact thatsome nude mice developed granulocytic leuke-mias fits the pattern of X expression, but it alsoshows that the restriction of transformation inMuLV-M-infected mice is relative in that cellsother than T lymphocytes may become in-volved. A phenotypic heterogeneity of leuke-mias in MuLV-M-infected BALB/c mice wasrecently reported by Boyer et al. (4), who foundthe leukemic cells to be Thy 1.2+ in some mice,whereas they appeared to be "null" cells (Thy1.2-, sIg-) in other mice. The two observationson the diversity of leukemogenesis among ge-netically homogenous mice and the absence oftransformation in subsets of lymphocyte that arepermissive to the virus infection represent apuzzling aspect of the ecotropic MuLV patho-genesis.
ACKNOWLEDGMENTS
We thank Carol L. Reinisch for her assistance with the anti-mouse F(ab')2 immunoadsorbent columns, Edward A. Hooverfor his evaluation of the histologic sections prepared fromleukemic tissues, and Judy Adam for typing of the manuscript.
This work was supported by American Osteopathic Associ-ation grant 79-04-38, National Science Foundation grant7922943, Public Health Service grants CA 15277 and CA 14922from the National Institutes of Health, and by AmericanCancer Society grant IM 35.
LITERATURE CITED1. Asjo, B., E. M. Fenyo, J. Spira, and G. Klein. 1980.
Appearance and distribution of virally determined anti-gens in lymphoid organs of mice during leukemogenesisby Moloney leukemia virus. Leuk. Res. 4:89-103.
2. Baird, S., W. Raschke, and I. L. Weismann. 1977. Evi-dence that MuLV-induced thymic lymphoma cells pos-sess specific cell membrane binding sites for MuLV. Int.J. Cancer 19:403-413.
3. Bassin, R. H., H. Tuttle, and P. J. Fischinger. 1971. Rapidcell culture assay technique for murine leukemia viruses.Nature (London) 229:564-566.
4. Boyer, B., S. Gisselbrecht, P. Debre, I. McKenzie, andJ. P. Levy. 1980. Genetic control of sensitivity to Moloneyleukemia virus in mice. IV. Phenotypic heterogeneity ofthe leukemic mice. J. Immunol. 125:1415-1420.
5. Celada, F., B. Asjo, and G. Klein. 1970. The presence ofMoloney virus induced antigen on antibody-producingcells. Clin. Exp. Immunol. 6:317-323.
6. Cerny, J., S. H. Fistel, and P. A. Hensgen. 1976. Interac-tions of murine leukemia virus (MuLV) with isolatedlymphocytes. I. Virus replication in lymphocytes infectedwith Friend virus and cultured in diffusion chambers invivo. Int. J. Cancer 18:176-188.
7. Cerny, J., P. A. Hensgen, S. H. FIstel, and L. Mastalir-Demler. 1976. Interactions of murine leukemia virus(MuLV) with isolated lymphocytes. II. Infections of Band T cells with Friend virus complex in diffusion cham-bers and in vitro: effect of polyclonal mitogens. Int. J.Cancer 18:189-196.
8. Cerny, J., D. D. Isaak, and E. A. Hoover. 1979. Interac-tions of murine leukemia viruses (Friend and Moloney)with lymphocyte subpopulations. Studies on the cellularrestriction of virus replication, transformation and activa-tion of endogenous viruses, p. 149. In M. R. Proffit (ed.),Virus-lymphocyte interactions: implications for disease.Elsevier/North Holland, New York.
VOL. 40, 1983
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

984 ISAAK AND CERNY
9. Ceruy, J., and R. A. Stiller. 1975. Immunosuppression byspleen cells from Moloney leukemia. Comparison of thesuppressive effect on antibody response and on mitogen-induced response. J. Immunol. 115:943-949.
10. Cremer, N. E., D. 0. Taylor, and E. H. Lennett. 1969.Localization of immunoglobulin and viral antigen in ratsinfected with Moloney virus. J. NatI. Cancer Inst. 42:695-707.
11. Dunn, T. B., J. B. Moloney, A. W. Green, and B. Arnold.1961. Pathogenesis of a virus-induced leukemia in mice. J.Natl. Cancer Inst. 26:189-221.
12. Fischinger, P. J., J. N. Ihle, F. De Noronha, and D. P.Bolognesi. 1977. Oncogenic and immunogenic potential ofcloned HIX viruses in mice and cats. Med. Microbiol.Immunol. 164:119-129.
13. Haas, M., T. Sher, and S. Smolinsky. 1977. Leukemogene-sis in vitro induced by thymus epithelial reticulum cellstransmitting murine leukemia viruses. Cancer Res.37:1800-1807.
14. Handwerger, B. S., and R. H. Schwartz. 1974. Separationof murine lymphoid cells using nylon wool columns.Recovery of the B cell-enriched population. Transplanta-tion 18:544-548.
15. Hartley, J. W., N. K. Wolford, L. J. Old, and W. P.Rowe. 1977. A new class of murine leukemia virus associ-ated with development of spontaneous lymphomas. Proc.Natl. Acad. Sci. U.S.A. 74:789-792.
16. Horack, I., L. Enjuanes, J. C. Lee, and J. N. Ihle. 1981.Resistance of cultures of normal T cells to infection withmurine type C viruses. J. Virol. 37:483-487.
17. Ihle, J. N. 1978. Experimental models and conceptualapproaches to studies of lymphomas and leukemia: etiolo-gy, biology and control. Semin. Hematol. 15:95-115.
18. Isaak, D. D., and J. Cerny. 1981. Target cell heterogeneityin murine leukemia virus infection. II. Demonstration ofFriend leukemia virus-permissive and non-permissivesubsets of splenic T cells. Int. J. Cancer 27:505-512.
19. Isaak, D. D., J. A. Price, C. L. Reinisch, and J. Cerny.1979. Target cell heterogeneity in murine leukemia virusinfection. I. Differences in susceptibility to infection withFriend leukemia virus between B lymphocytes fromspleen, bone marrow and lymph nodes. J. Immunol.123:1822-1828.
20. Jaenisch, R., H. Fan, and B. Croker. 1975. Infection ofpreimplantation mouse embryos and of newborn micewith leukemia virus: tissue distribution of viral DNA andRNA and leukemogenesis in the adult animal. Proc. Natl.Acad. Sci. U.S.A. 272:4008-4012.
21. Kaplan, H. S., and M. B. Brown. 1954. Development oflymphoid tumors in nonirradiated thymic grafts in thy-
mectomized irradiated mice. Science 119:439-440.22. Katz, I. R., R. Asofsky, and G. J. Thorbecke. 1980. Sup-
pression of spontaneous reticulum cell sarcoma develop-ment and of syngeneic stimulator cells by anti-pL treatmentof SJL/J mice. J. Immunol. 125:1355-1359.
23. Kumar, V., T. Caruso, and M. Bennett. 1976. Mechanismsof genetic resistance to Friend virus leukemia. III. Sus-ceptibility of mitogen-responsive lymphocytes mediatedby T cells. J. Exp. Med. 143:728-740.
24. Moloney, J. B. 1960. Biological studies on a lymphoid-leukemia virus extracted from sarcoma 37. I. Origin andintroductory investigations. J. NatI. Cancer Inst. 24:933-951.
25. Nowinski, R. C., and E. F. Haws. 1978. Oncogenicity ofAKR endogenous leukemia viruses. J. Virol. 27:13-18.
26. Oshiro, L. S., N. E. Cremer, D. 0. Taylor, and E. H.Lennette. 1969. Electron microscopic studies on the local-ization of antibodies in rat lymph node cells producingMoloney virus. J. Natl. Cancer Inst. 43:1109-1118.
27. Proffitt, M. R., M. S. Hirsch, I. F. McKenzie, B. Gheri-dian, and P. H. Black. 1975. Immunological mechanismsin the pathogenesis of virus-induced murine leukemia. II.Characterization of autoreactive thymocytes. Int. J. Can-cer 15:230-240.
28. Reddy, E. P., C. Y. Dunn, and S. A. Aaronson. 1980.Different lymphoid cell targets for transformation byreplication-competent Moloney and Rauscher mouse leu-kemia viruses. Cell 19:663-669.
29. Ricciardi-Castagnoli, P., M. Lieberman, 0. Finn, andH. S. Kaplan. 1978. T-cell lymphoma induction by irradia-tion leukemia virus in athymic nude mice. J. Exp. Med.148:1291-1310.
30. Schlossman, S. F., and L. Hudson. 1973. Specific purifica-tion of lymphocyte populations on a digestible immunoad-sorbent. J. Immunol. 110:313-315.
31. Schrader, J. W., F. Battye, and R. Scollay. 1982. Expres-sion of Thy-1 antigen is not limited to T cells in cultures ofmouse hemopoietic cells. Proc. Natl. Acad. Sci. U.S.A.79:4161-4165.
32. Shevach, E. M., J. D. Stobo, and I. Green. 1972. Immuno-globulin and 0-bearing murine leukemias and lymphomas.J. Immunol. 108:1146-1151.
33. Siegler, R. 1968. Pathology of murine leukemias, p. 51. InR. A. Rich (ed.), Experimental leukemia. Appleton-Cen-tury-Crofts, New York.
34. Troxier, D. H., and E. M. Scolnick. 1978. Rapid leukemiainduced by cloned Friend strain of replicating murine typeC virus. Association with induction of xenotropic-relatedRNA sequences contained in spleen focus-forming virus.Virology 85:17-27.
INFECT. IMMUN.
on August 21, 2018 by guest
http://iai.asm.org/
Dow
nloaded from





























