Embed Size (px)
Citation preview

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
135
064: cartel ALIMENTACIÓN Y ASPECTOS BIOLÓGICO-PESQUEROS DE ALGUNAS ESPECIES DE IMPORTANCIA
EN LA PESCA ARTESANAL DE PUERTO ÁNGEL, OAXACA
Daniel Mata Martínez e Isaías H. Salgado Ugarte
Laboratorio de Biometría y Biología Pesquera, FES Zaragoza, Campus II. Batalla 5 de mayo s/n, Esq. Fuerte de Loreto, Ejército de Oriente, Iztapalapa, Ciudad de México. C.P. 09230. [email protected] Introducción El objetivo de este estudio es conocer la biología alimenticia del cocinero (Caranx caballus) y de la rabirrubia (Lutjanus inermis) por su importancia ecológica y económica, para su aprovechamiento racional, ante ello es necesario determinar el lugar que ocupan dentro de la red alimenticia por medio del análisis del contenido estomacal. Material y Métodos Para tal fin se efectuaron durante un ciclo anual muestreos mensuales de noviembre del 2013 a enero del 2015. De un total de 240 datos de análisis de estómagos de la captura realizada del cocinero (C. caballus); y de 147 datos de la rabirrubia (L. inermis) en la zona de Costa Chica, Oaxaca, por medio del análisis numérico y gravimétrico, se registró la importancia de 3 grupos o categorías principales que forman parte del componente alimenticio en el cocinero (peces, moluscos y crustáceos) y solamente 2 grupos (peces y crustáceos) en la dieta del rabirrubia; por lo que se clasificaron como especies depredadoras, debido a la diversidad de presas presentes en los estómagos. Resultados En el análisis porcentual numérico (%N), la categoría más numerosa es la de Crustáceos 80.56%, seguida de los Peces 10.34% y por último los Moluscos 2.82%; en el análisis porcentual gravimétrico (%G) la primera categoría la conforman los Peces con 82.46%, seguida de los Crustáceos 6.2% y por último los Moluscos 2.28%, en C. caballus y para L. inermis con el %N, el grupo de los Crustáceos dominan con 84.82%, seguida de los Peces con 7.14%; en el %G el grupo de los Crustáceos dominan con 43.52%, seguida de los Peces con 27.69%. El ANMUCOVA muestra los meses con mayor alimentación, así como también los períodos de desove indicado por los valores medios ajustados más altos de gónadas, ajustadas por la covariada para los datos totales (Salgado-Ugarte et al. 2005). Los meses de noviembre, septiembre y enero son los meses con mayor actividad de alimentación, de las cuales se encuentra las siguientes presas: peces (Engraulidae, Clupeidae, Caragidae); crustáceos (Euphausia, Solenocera, Portunus) y moluscos (Octopus y Nassarius). La alimentación se incrementa en los primeros meses del año cuando en la zona se encuentra el
flujo de la Corriente de California (CC): fría, de baja salinidad y alta en O2, la cual llega hasta Chiapas y la Corriente Norecuatorial (CNE): caliente, alta salinidad y baja en O2. Al encontrarse estas dos corrientes provocan diferencias de temperatura o incrementos dramáticos de salinidad que son capaces de hundir masas de agua. Tabla 1. Coeficiente de alimentación (Q) para hembras de C. caballus.
Tabla 2. Componentes alimenticios por sexo según la longitud de C. caballus.
Otro pico de alimentación comienza en otoño, ya que en la zona se presentan surgencias intensas debido a la interacción de frentes fríos que bajan por el istmo de Tehuantepec, produciendo un aporte masivo de nutrientes que pone en marcha cadenas productivas que benefician el reclutamiento y el crecimiento de las especies.
Ítem Peces Crustáceo Molusco
C. caballus
853.07 (1rio, Preferido)
499.93 (1rio, Preferido)
0.45 (Accidental)
Sexo MONI (cm)
Crustáceos (cm)
Crustáceos/ Peces (cm)
Peces (cm)
Hembras 21-26 26-36 36-42 42-54
Machos 22-24 24-36 35-46 46-56
Indeter. 22-25 25-29 (Moluscos)
29-32 32-44
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
136
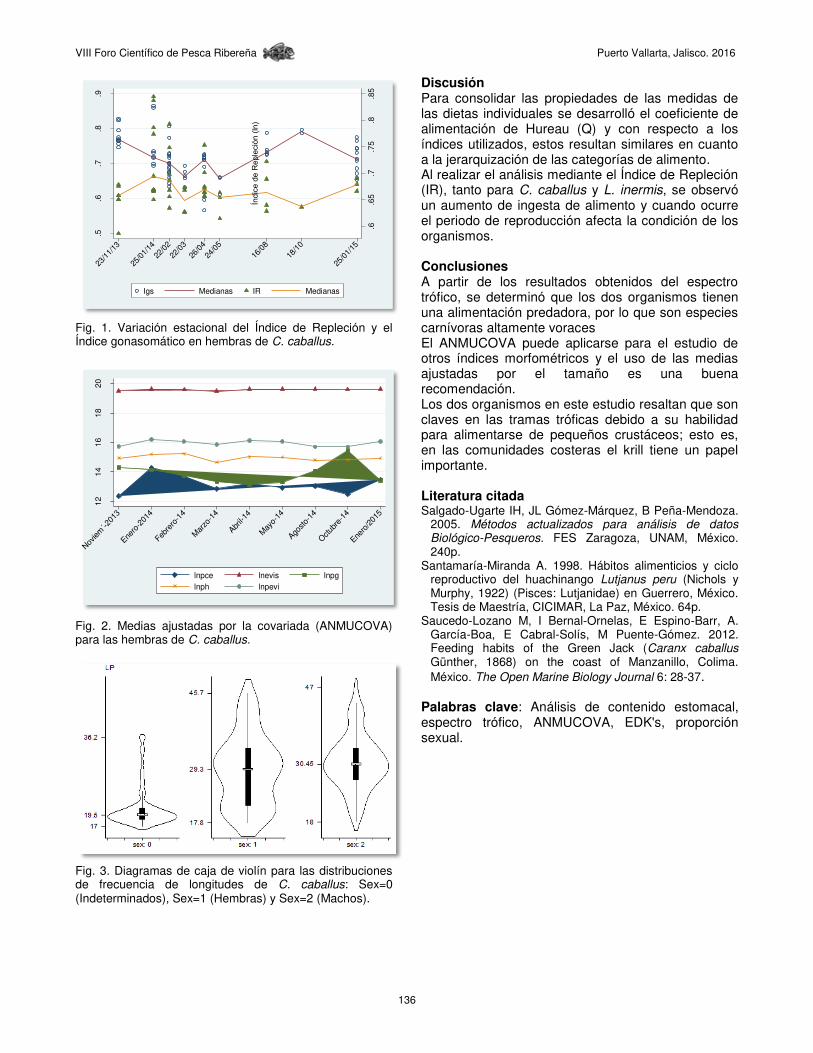
Fig. 1. Variación estacional del Índice de Repleción y el Índice gonasomático en hembras de C. caballus.
Fig. 2. Medias ajustadas por la covariada (ANMUCOVA) para las hembras de C. caballus.
Fig. 3. Diagramas de caja de violín para las distribuciones de frecuencia de longitudes de C. caballus: Sex=0 (Indeterminados), Sex=1 (Hembras) y Sex=2 (Machos).
Discusión Para consolidar las propiedades de las medidas de las dietas individuales se desarrolló el coeficiente de alimentación de Hureau (Q) y con respecto a los índices utilizados, estos resultan similares en cuanto a la jerarquización de las categorías de alimento. Al realizar el análisis mediante el Índice de Repleción (IR), tanto para C. caballus y L. inermis, se observó un aumento de ingesta de alimento y cuando ocurre el periodo de reproducción afecta la condición de los organismos. Conclusiones A partir de los resultados obtenidos del espectro trófico, se determinó que los dos organismos tienen una alimentación predadora, por lo que son especies carnívoras altamente voraces El ANMUCOVA puede aplicarse para el estudio de otros índices morfométricos y el uso de las medias ajustadas por el tamaño es una buena recomendación. Los dos organismos en este estudio resaltan que son claves en las tramas tróficas debido a su habilidad para alimentarse de pequeños crustáceos; esto es, en las comunidades costeras el krill tiene un papel importante. Literatura citada Salgado-Ugarte IH, JL Gómez-Márquez, B Peña-Mendoza.
2005. Métodos actualizados para análisis de datos Biológico-Pesqueros. FES Zaragoza, UNAM, México. 240p.
Santamaría-Miranda A. 1998. Hábitos alimenticios y ciclo reproductivo del huachinango Lutjanus peru (Nichols y Murphy, 1922) (Pisces: Lutjanidae) en Guerrero, México. Tesis de Maestría, CICIMAR, La Paz, México. 64p.
Saucedo-Lozano M, I Bernal-Ornelas, E Espino-Barr, A. García-Boa, E Cabral-Solís, M Puente-Gómez. 2012. Feeding habits of the Green Jack (Caranx caballus Günther, 1868) on the coast of Manzanillo, Colima. México. The Open Marine Biology Journal 6: 28-37.
Palabras clave: Análisis de contenido estomacal, espectro trófico, ANMUCOVA, EDK's, proporción sexual.
.6.6
5.7
.75
.8.8
5
Índi
ce d
e R
eple
ción
(ln)
.5.6
.7.8
.9
Índi
ce G
onas
omát
ico
(ln)
23/1
1/13
25/0
1/14
22/0
222
/03
26/0
424
/05
16/0
818
/10
25/0
1/15
Igs Medianas IR Medianas
1214
1618
20
Var
iabl
es ε
orfo
fisio
lógi
cas
(ln)
Noviem
´-201
3
Enero
-201
4
Febre
ro-1
4
Mar
zo-1
4
Abril-1
4
May
o-14
Agosto
-14
Octubr
e-14
Enero
/201
5
lnpce lnevis lnpglnph lnpevi

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
137
065: oral ESTIMACIÓN DE DENSIDAD Y ABUNDANCIA DE LANGOSTA (Panulirus argus), CARACOL (Lobatus
gigas) Y PEPINO DE MAR (Holothuria mexicana), Y RIQUEZA ESPECÍFICA DE PECES EN EL PARQUE NACIONAL ARRECIFE ALACRANES (PNAA), YUCATÁN, MÉXICO
Gloria Verónica Ríos Lara1, Carlos E. Zetina Moguel2, Juan Carlos Espinoza Méndez1,
Kenneth Cervera Cervera1, Egna Deneb Cervera Paul1, Mariana Uribe Cuevas1 y David E. De Anda Fuentes1
1Centro Regional de Investigación Pesquera de Yucalpetén. INAPESCA. 2Facultad de Ingeniería. Universidad Autónoma de Yucatán. Av. Ind. No Cont. y Anillo Periférico Norte s/n. Mérida, Yuc. e-mail: [email protected] Introducción Desde hace varios años se piensa que el ordenamiento pesquero en el largo plazo requiere de la ubicación de las pesquerías en un contexto ecosistémico. El Parque Nacional Arrecife Alacranes (PNAA) ubicado a 130 km al norte de Progreso, Yucatán (2221´45´´ y 2234´55´´ LN y 8936´47´´ y 8947´53´´LO), tiene gran importancia ecológica y económica, dada la diversidad de especies que alberga y por ser una importante zona de pesca de diversos recursos (langosta, varias especies de escama y en algún tiempo caracol Lobatus gigas), tradicionalmente aprovechados por un grupo limitado de pescadores. Como parte de las investigaciones relacionadas a la pesquería de langosta, se hizo una prospección al finalizar la veda, en la zona central del PNAA. El objetivo principal fue evaluar la población de langosta y obtener indicadores de abundancia de otras especies bentónicas de importancia comercial o potenciales, que se encuentran en los sitios donde se captura la especie. Se estimó la densidad, abundancia y biomasa para la langosta y la densidad y abundancia para el caracol y el pepino de mar. Por otra parte se obtuvo la riqueza específica de peces para la misma zona. Material y métodos La colecta de datos se hizo del 4 al 10 de junio de 2015 a través de muestreo por buceo en cuadrantes, dirigido a sitios con mayor disponibilidad de hábitat preferido por la langosta, dadas sus características arquitectónicas y su diversidad (zona central del arrecife, entre 2 y 15 m de profundidad). Se distribuyeron geográficamente 40 estaciones en tres tipos de fondo: parches de coral duro con arenales intersticiales entre 1-3 m de profundidad (2), parches de coral duro y abundancia de coral blando entre 3-8 m de profundidad (4) y parche de coral duro con paredes de 10 a 15 m (6), según mapa temático de tipos de fondo de Bello-Pineda et al. (2005). Se hicieron transectos y se contabilizaron: número de langostas (P. argus), caracoles (L. gigas) y pepinos de mar (Holothuria mexicana) observados; también se registraron las especies de peces, las cuales fueron identificadas mediante la observación. Se registraron los elementos dominantes del paisaje y se tomaron fotografías del fondo, con el fin de
identificar los principales elementos presentes (corales, esponjas, arena, algas y pastos). Se midieron los parámetros físico-químicos de la columna de agua: profundidad (m), oxígeno disuelto (mg/l), pH, salinidad (ppt) y temperatura (°C). La densidad media de las tres especies fue estimada como:
iii
i
plw
yD
ˆ
Donde: wi = ancho de la iésima área muestreada, li = largo de la iésima área muestreada y pi = detectabilidad (probabilidad de encuentro de langosta) en la iésima área muestreada. La abundancia se estimó como:
ADY *ˆˆ Donde: A = área de interés. En el caso de la langosta se estimó la biomasa, utilizando el peso medio de las langostas observadas. Para la estimación de la riqueza específica (S) máxima se usó el modelo de Clench:
)*(1
*
Eb
EaSE
Donde: SE= Riqueza específica al esfuerzo E; a y b son parámetros a estimar y E = esfuerzo de muestreo (transectos recorridos), Smax = máximo teórico o asintótico de especies en el ecosistema sin considerar las que llegan fuera de las condiciones de observación o las especies accidentales. Se construyó una curva de acumulación área-especie utilizando métodos de remuestreo intensivo y el ajuste de las curvas se hizo a través de regresión no lineal. Resultados Las estimaciones de densidad (DL), abundancia (AL) y biomasa (BL) para langosta, por tipo de fondo fueron: clase 2, DL=53 org/ha, AL=119,531 y BL= 55 t; clase 4, DL= 34 org/ha, AL=81,156 org. y BL= 37 t; clase 6, DL=4, AL=281 y BL= 0.13 t. La AL total para el área de estudio fue de 200,969 y una BL total de
b
aS max
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
138
92 t de langosta entera. Las estimaciones de densidad (DC) y abundancia (AC) para caracol, por tipo de fondo fueron: clase 2, DC=20 org/ha y AC=45,487 org.; clase 4, DC=9 org/ha y AC=21,814 org.; clase 6, DC=7 org/ha y AC=563 organismos. La AC total para el área de estudio fue de 67,864 org. Las estimaciones de densidad (DP) y abundancia (AP) de pepino de mar, por tipo de fondo fueron: clase 2, DP=14 org/ha y AP=31,114 org.; clase 4, DP=3 org/ha y AP=8,159 org.; 6, DP=5 org/ha y AP=427 org. La AP total para el área de estudio fue de 39,699 org. Se elaboraron mapas de distribución de densidad para las tres especies (Fig. 1).
Fig. 1. Distribución de la densidad de langosta (P. argus), caracol (L. gigas) y pepino de mar (H. mexicana) en el área central del AA.
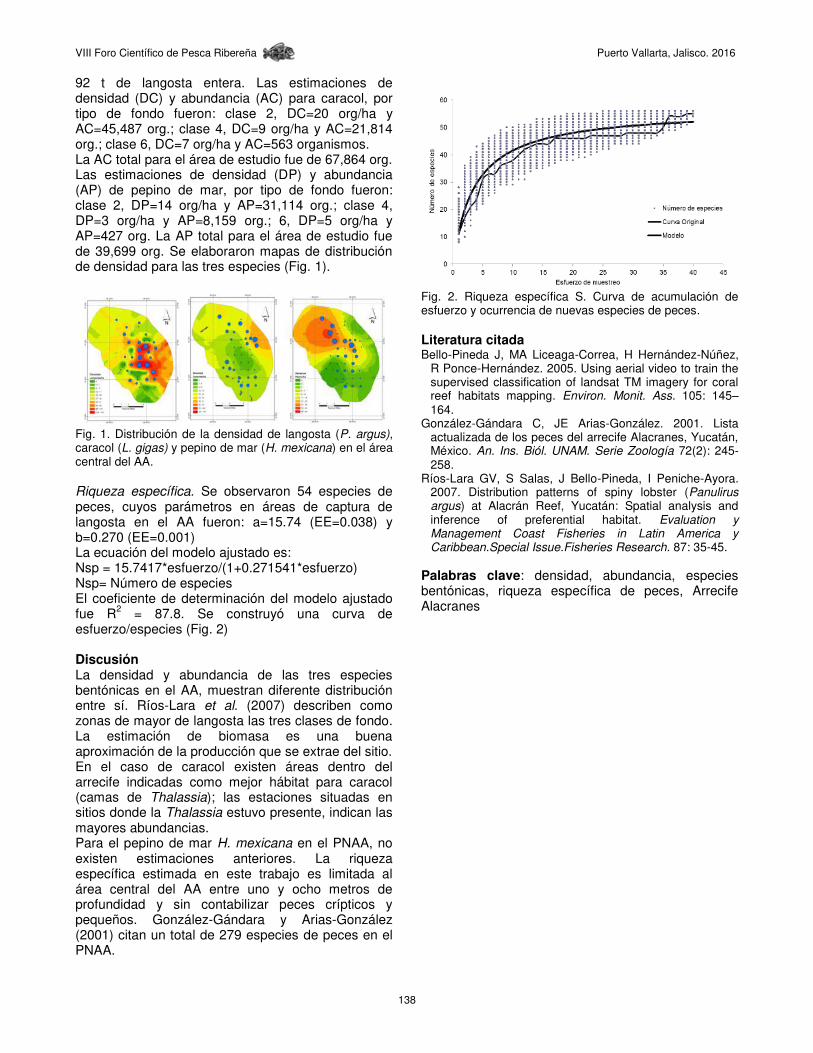
Riqueza específica. Se observaron 54 especies de peces, cuyos parámetros en áreas de captura de langosta en el AA fueron: a=15.74 (EE=0.038) y b=0.270 (EE=0.001) La ecuación del modelo ajustado es: Nsp = 15.7417*esfuerzo/(1+0.271541*esfuerzo) Nsp= Número de especies El coeficiente de determinación del modelo ajustado fue R2 = 87.8. Se construyó una curva de esfuerzo/especies (Fig. 2) Discusión La densidad y abundancia de las tres especies bentónicas en el AA, muestran diferente distribución entre sí. Ríos-Lara et al. (2007) describen como zonas de mayor de langosta las tres clases de fondo. La estimación de biomasa es una buena aproximación de la producción que se extrae del sitio. En el caso de caracol existen áreas dentro del arrecife indicadas como mejor hábitat para caracol (camas de Thalassia); las estaciones situadas en sitios donde la Thalassia estuvo presente, indican las mayores abundancias. Para el pepino de mar H. mexicana en el PNAA, no existen estimaciones anteriores. La riqueza específica estimada en este trabajo es limitada al área central del AA entre uno y ocho metros de profundidad y sin contabilizar peces crípticos y pequeños. González-Gándara y Arias-González (2001) citan un total de 279 especies de peces en el PNAA.
Fig. 2. Riqueza específica S. Curva de acumulación de esfuerzo y ocurrencia de nuevas especies de peces. Literatura citada Bello-Pineda J, MA Liceaga-Correa, H Hernández-Núñez,
R Ponce-Hernández. 2005. Using aerial video to train the supervised classification of landsat TM imagery for coral reef habitats mapping. Environ. Monit. Ass. 105: 145–164.
González-Gándara C, JE Arias-González. 2001. Lista actualizada de los peces del arrecife Alacranes, Yucatán, México. An. Ins. Biól. UNAM. Serie Zoología 72(2): 245-258.
Ríos-Lara GV, S Salas, J Bello-Pineda, I Peniche-Ayora. 2007. Distribution patterns of spiny lobster (Panulirus argus) at Alacrán Reef, Yucatán: Spatial analysis and inference of preferential habitat. Evaluation y Management Coast Fisheries in Latin America y Caribbean.Special Issue.Fisheries Research. 87: 35-45.
Palabras clave: densidad, abundancia, especies bentónicas, riqueza específica de peces, Arrecife Alacranes

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
139
066: oral “SI A LAS TRES PM NO HE VENDIDO EL PRODUCTO, YA NO LO PODRÉ VENDER”. LA
COMERCIALIZACIÓN DEL CAMARÓN: EL CASO DE LOS PESCADORES DE CD. DEL CARMEN, CAMPECHE
Jazmín Villegas Calzada
Facultad Latinoamericana de Ciencias Sociales sede México. Carretera al Ajusco 377, Col. Héroes de Padierna, Del. Tlalpan, Ciudad de México. C.P.14200. Tel. (55)30000200. [email protected] Introducción La Carta Nacional Pesquera (DOF 2012) reconoce la complejidad en el manejo pesquero para el Golfo de México y El Caribe, ante la intervención de aspectos biológicos, tecnológicos, sociales, culturales, ecológicos, económicos y políticos. De esta forma encontramos el argumento de Breton (1989) al considerar que este oficio comprende todo un proceso logístico traducido en la planeación, preparación y ejecución de la travesía, la captura y la venta del producto. Asimismo, no se encuentra aislado, ya que como cualquier otra actividad humana recibe inevitablemente la influencia de los factores contextuales bajo los que se desarrolla. Para Marín Guardado (2007) los pescadores mexicanos se encuentran en medio de enormes desventajas para operar eficientemente en los mercados y sujetos al menoscabo dentro de las políticas de la economía nacional. En esta investigación se analiza el caso del Municipio Del Carmen, Campeche, donde se explora la comercialización del camarón siete barbas y de la escama provenientes de la captura ribereña. La cadena de intermediarios en la comercialización de los productos del mar se muestra en la figura 1.
Fig. 1. Esquema de ejemplo de lo complejo e intrincado de una cadena de distribución. Asimismo, la figura 2 resalta a la vista que el pescador artesanal ocupa el nivel más bajo en la comercialización del producto. Gráficamente se observa una pirámide de intermediación y donde el precio del pescado y/o marisco se incrementa hasta en 300%.
Fig. 2. Ejemplo del incremento del precio a lo largo de la cadena de distribución. Material y métodos Se efectuó una investigación de carácter cualitativo consistente en un estudio de caso mediante la realización de visitas de observación y recolección de datos sociodemográficos y económicos, a través de la toma de imágenes y grabaciones de entrevistas practicadas a pescadores ribereños en Ciudad Del Carmen, Campeche, durante los meses de marzo y junio de 2014. El trabajo es descriptivo y las conclusiones a las que se lleguen tendrán una validez limitada y no general. Las preguntas son relativas al acceso de los medios de producción y conservación, el grado de participación en la comercialización de los productos, así como los lugares de venta. Para el análisis de la información se clasificó a los pescadores que capturan camarón y a aquellos que se dedican a la escama. Esta diferenciación permitió observar la cadena de intermediarios, el equipo de conservación (congeladores) disponible, medios de transporte y el capital social con el que cuentan los pescadores, lo que determina la ganancia obtenida. Resultados y discusión Al efectuar el análisis de las actividades, destacan los siguientes hallazgos:

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
140
Camarón No hay un control de los precios de pescados y
mariscos por parte de la autoridad pesquera. Los camaroneros no cuentan con refrigeradores y/o
congeladores que les permitan conservar el producto por uno o más días, colocándolos a merced de las condiciones establecidas por los acaparadores.
Estos trabajadores del mar dependen netamente del precio fijado por los denominados “coyoteros” quienes llevan a cabo las “juntas negras”, reuniones donde acuerdan un precio para el camarón oscilante entre 30 y 32 pesos.
Al no contar con medios de transporte adecuados no pueden trasladar el producto al mercado más importante del lugar, ubicado en la cabecera municipal, restaurantes y empacadoras, limitando el área geográfica de comercialización.
Escama: Acuden al mercado de la cabecera municipal donde
lo venden a los propietarios de los puestos del área de pescado, a los restaurantes del lugar y al mercado de mariscos.
Cuentan con sistemas de congelación que les permiten acumular el producto hasta por 3 días, cuando les resulta redituable realizar el traslado de la mercancía.
Tienen la posibilidad de negociar el precio del producto a los minoristas (propietarios de los puestos del mercado central).
Algunos lo ofertan directamente a la empacadora del lugar.
Entre los mismos pescadores se comunican dónde o quién paga un poco más, de esta manera sus conocidos les ayudan para obtener mayor ganancia.
La venta relativamente más directa del producto (sin intermediarios) es otro factor determinante en las trayectorias laborales de estos trabajadores del mar.
Conclusiones Se confirma que la pesca artesanal en Ciudad del Carmen existe una gran cadena de intermediarios, en donde los trabajadores del mar se encuentran distantes de las tiendas de autoservicio, el mercado a nivel nacional sin siquiera pensar en el internacional, sobre todo por el camarón, producto altamente cotizado en otros países. Asimismo, los factores que favorecen u obstaculizan el precio del pescado y/o marisco es el acceso a los medios de refrigeración y congelación, la participación directa en la comercialización del producto así como el lugar donde se oferta. El capital social también resulta importante para negociar el precio y consecuentemente la ganancia obtenida.
El pescador carmelita es quien arriesga más y quien gana menos. Asimismo, la falta de diversificación del producto no les permite superar los límites del área de venta. La deficiencia en la operación de las cooperativas, la falta de capacitación para desempeñar el oficio, el analfabetismo, la falta de control en precios, la ausencia de políticas públicas entre otros, convergen y agravan la situación de los pescadores carmelitas al constituirse como una sector pobre, marginado y vulnerable. La intervención de las autoridades y la implementación de políticas públicas del sector deben enfocarse a atender los problemas que no permiten que la pesca se constituya como un oficio que garantice el acceso al bienestar. La pesca es un sector cuya derrama económica es relevante, evidentemente hay ganancias pero éstas no son para los que efectúan la captura sino para aquellos que cuentan con la capacidad económica y social para desenvolverse en los mercados. Finalmente, se pone de manifiesto el abandono en el que se encuentra el sector pesquero artesanal. Literatura citada Breton Y, E López Estrada. 1989 Ciencias sociales y
desarrollo de las pesquerías: modelos y métodos aplicados al caso de México Instituto Nacional de Antropología e Historia
DOF. 2012. Carta Nacional Pesquera. Instituto Nacional de Pesca, SAGARPA. Diario Oficial de la Federación.
Marín Guardado G. 2007. Vidas a contramarea: pesca artesanal, desarrollo y cultura en la costa de Michoacán. CIESAS México.
Palabras clave: comercialización, intermediarios,
pesca ribereña, marginalidad y vulnerabilidad.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
141
067: oral DISTRIBUCIÓN ESPACIAL Y ABUNDANCIA DE ELASMOBRANQUIOS EN EL GOLFO DE MÉXICO
DURANTE 2014
Heber Zea De la Cruz, Jorge L. Oviedo Pérez y Pedro C. Reyna González
Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. Av. Ejército Mexicano 106, Col. Ex Hacienda Ylang Ylang, Boca del Río, Veracruz. C.P. 94298. Tel. (229) 1304520. [email protected] Introducción Los elasmobranquios poseen características biológicas que los hacen susceptibles a la explotación, entre ellas sobresalen un relativo lento crecimiento, madurez sexual tardía, baja fecundidad y una posible relación denso-dependencia. La región del Golfo de México presenta una diversidad de elasmobranquios que son capturados de manera incidental en el sistema de arrastre camaronero y ante la importancia que ha adquirido la explotación sustentable de tiburones y rayas, es prioritaria una evaluación de la abundancia de estos recursos. Asimismo se requiere de estimaciones de abundancia, distribución espacio temporal y datos de las poblaciones con información proveniente de muestreos independiente de la pesquería comercial. El objetivo del presente trabajo fue estimar la distribución espacial y abundancia de elasmobranquios a partir de lances de investigación con el sistema de arrastre camaronero en la parte mexicana del Golfo de México durante 2014. Material y métodos Se analizó la información proveniente de 13 cruceros de investigación a bordo de embarcaciones camaroneras en cuatro zonas del Golfo de México; Tamaulipas (26°00' y 22°00' N), Veracruz norte (22°00' y 19°30' N), Veracruz sur (19°30' y 19°00 N' y desde 95°57' y 94°00' O) y Tabasco-Campeche (92°30' y 93°15' O y desde 21°30' y 19°30' N). Al inicio y final de los lances fueron obtenidos los datos de operación de los equipos de pesca: fecha, hora, posición geográfica, profundidad, velocidad, rumbo, tiempo de arrastre. En cada lance fueron separados todos los elasmobranquios, identificados al nivel de especie. Se registró el sexo, el número de organismos y el peso se registró de manera global por especie. Se estimaron los índices de abundancia relativa en Captura por Unidad de Esfuerzo (CPUE) y de densidad en Captura por Unidad de Área (CPUA). La CPUE se expresó para cada lance en kg/hr de arrastre:
fCi /CPUEi
donde: Ci = captura en kg totales de elasmobranquios y f = esfuerzo en horas de arrastre en el lance i respectivamente.
La densidad se expresó como la captura en kg/ha para cada lance. La CPUA se expresó en kg/hectárea:
ii AbC /CPUAi
donde: Ci = captura en kg del lance i y Abi = área barrida del lance. El Abi (Abi= ti vi ahi) se estableció en cada lance como el producto de la velocidad (vi), tiempo efectivo de arrastre (ti) y la abertura operacional de la red (ahi), el factor de ahí fue asumida a 60% (Klima 1976, Sparre y Venema 1997). El promedio del índice CPUE y CPUA se estimó para cada zona bajo el modelo general de Pennington (1996), asumiendo que estos índices presentan una distribución ∆. Se determinó la compocisión específica por especies en número de organismos capturados y peso (kg). Resultados y discusión Se monitorearon 250 lances de investigación, con un tiempo acumulado de arrastre de 475.4hr, 1,269 mn recorridas (2,351 km) y un área barrida de 8,108.8 ha (81.1 km) en profundidades entre 9 y 91 m. Durante los meses de mayo a julio el patrón de distribución espacial muestra a los elasmobranquios hasta en 91 m de profundidad. La mayor abundancia en kg/ha, se encuentra en la zona del litoral de Tamaulipas en el mes de julio, con máximos valores entre 1.51 y 1.89 kg/ha en lances realizados entre 14 y 20 m de profundidad entre la latitud N24° 34’ y 25° 36’ y la longitud λ7° 08’ O y λ7° 33’ O (Fig.1). En los meses de mayo y junio los máximos valores fueron de 0.85 y 1.82 kg/ha, respectivamente. La mayor abundancia relativa promedio en kg/hr de arrastre se estimó para Tamaulipas en 6.31kg/hr (IC= 4.43–8.19), para Veracruz sur se estimó en 5.83 kg/hr (IC= 3.75–7.91), en Veracruz norte 4.02 kg/hr (IC= 2.64–5.40) y en Tabasco-Campeche 1.26 kg/hr (IC= 0.63–1.89). En términos de densidad, se estimó para Tamaulipas el mayor valor con 0.377 kg/ha (IC= 0.264–0.489 kg/ha) Veracruz sur 0.313 kg/ha (IC= 0.204–0.421), Veracruz norte 0.256 kg/ha (IC= 0.168–0.344) y Tabasco-Campeche 0.094 (IC= 0.039–0.149) (Fig. 2).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
142
Fig. 1. Distribución espacial y densidad de elasmobranquios a partir muestreos de investigación con sistema de arrastre camaronero en el Golfo de México.
Fig. 2. Índices de abundancia de elasmobranquios a) kg/hr de arrastre, b) kg/ha en zonas del litoral del Golfo de México durante 2014. Las barras horizontales representan el error estándar. Se registraron un total de 2,383 organismos agrupados en 8 familias, 11 géneros y 14 especies de elasmobranquios: nueve de rayas: G. micrura, D. olseni, N. bancroftii, R. ackleyi, R. texana, U. jamaicensis, R. lentiginosus, R. bonasus; y seis de tiburones: S. tiburo, R. terraenovae, S. dumeril, S. mexicana, M. canis y M. norrisi. En la figura 3 se presenta la composición porcentual numérica de los elasmobranquios.
Fig. 3. Composición específica (%) del número de elasmobranquios monitoreados en lances de investigación con el sistema de arrastre camaronero durante 2014. Conclusiones Los patrones de distribución espacial muestran que los elasmobranquios se distribuyen a lo largo de la plataforma continental del Golfo de México al menos hasta los 91 m de profundidad. En julio se presentaron los valores máximos de densidad: en las zonas someras entre 14 y 20 m y en menor medida en lances hasta las 69 brz de profundidad. Los valores promedio de abundancia fueron estimados para la zona de Tamaulipas. A pesar de ser una estimación de abundancia estacional en primavera-verano, representan un marco de referencia y las bases de una serie temporal para evaluar posibles cambios en la abundancia. Los patrones de distribución espacial y abundancia pueden estar asociados a las características biológicas de las especies como alimentación, periodos y agregaciones de reproducción. Literatura citada Klima EF. 1976. An assessment of the fish stocks and
fisheries of the Campeche Bank. WECAF Stud. 5: 24 p. Pennington M. 1996. Estimating the mean and variance
from highly skewed marine data. Fishery Bulletin 94: 498-505.
Sparre P, SC Venema. 1997. Introduction to tropical fish stock assessment. Part 1. Manual. FAO Fisheries Technical Paper No. 306.1, rev. 2, Rome, FAO, 420 pp.
Palabras lave: Tiburones, rayas, abundancia, profundidad, arrastre camaronero.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
143
068: cartel ANÁLISIS SOCIODEMOGRÁFICO DE LAS COMUNIDADES DEL CORREDOR SAN COSME A PUNTA
COYOTE, B.C.S Y LA INFLUENCIA DE ACCIONES DE MANEJO PESQUERO
Melisa M. Vázquez Garcés, José M. Marrón Espitia, Salvador Rodríguez Van Dyck y Amy Hudson Weaver
Sociedad de Historia Natural Niparajá A. C., Revolución de 1910, núm. 430, Col. Esterito, La Paz, B.C.S. C.P. 23020. [email protected] Introducción El Corredor San Cosme-Punta Coyote se ubica en la costa oriental de Baja California Sur abarcando 150 km de litoral de los municipios de Loreto, Comondú y La Paz. Dentro del área se encuentran 15 comunidades, que en su mayoría realizan actividades de pesca ribereña como principal actividad económica. Dichas comunidades tienen un gran conocimiento de la zona y de los recursos que ahí existen, además de tener un fuerte arraigo a la zona y un gran interés en mejorar la productividad de su actividad por lo que el Corredor representa una gran oportunidad de conservación y manejo pesquero (Sociedad de Historia Natural Niparajá A.C., Pronatura Noroeste A.C. y Iemanya Oceánica A.C., 2009). En 2010 se les invitó a los pescadores del área a participar en un Programa de Ordenamiento Pesquero (POP) y a evaluar la factibilidad de establecer zonas de refugio pesquero (ZDR). Una red de ZDR fue establecida en nov 2012 (DOF 2012). En el presente trabajo se realiza una descripción y comparación de las comunidades y el esfuerzo pesquero. Así mismo, se presentan los cambios percibidos entre 2009 y 2016 para las problemáticas que han estado presentes en el área y el avance en las soluciones propuestas por el sector pesquero. Material y métodos Durante finales de 2015 y principios de 2016 se realizaron tres tipos de encuestas en las comunidades del Corredor San Cosme-Punta Coyote: 1) censo comunitario, 2) censo de esfuerzo pesquero y 3) cuestionario sobre problemática en la zona y propuestas de soluciones. El primero fue respondido por subdelegados, habitantes de las comunidades y observaciones personales. El segundo se aplicó a los capitanes de embarcaciones que viven en el área de manera permanente, y en las comunidades en las que existen Sociedades Cooperativas de Producción Pesquera (SCPP) se aplicó a representantes del consejo de administración. El tercero fue aplicado a capitanes de embarcaciones pesqueras; dentro de éste cada problemática y propuesta fue clasificada con base en escalas mostradas en la tabla 1 y tabla 2.
Para la elaboración del cuestionario sobre la problemática y soluciones, se tomaron como base las identificadas en 2009 por organizaciones de la sociedad civil, las cuales se obtuvieron a través de reuniones con los pescadores de cada localidad de la zona (Sociedad de Historia Natural Niparajá A.C., Pronatura Noroeste A.C. y Iemanya Oceánica A.C., 2009). El valor obtenido en cada una de las problemáticas y soluciones fue promediado entre todos los entrevistados de cada comunidad.
Tabla 1. Escala de valoración empleada en el cuestionario sobre la problemática del área.
Mejoró bastante
Mejoró un poco
Sigue igual
Empeoró un poco
Empeoró bastante
5 4 3 2 1 Tabla 2. Escala de valoración empleada para las soluciones propuestas por el sector pesquero.
Se solucionó
el problema
Se tomaron algunas acciones
al respecto
No se ha hecho
nada/Sigue igual
Hay menor
interés en hacer algo al
respecto
Ya no hay interés o
disposición
5 4 3 2 1 Resultados y discusión La población del Corredor San Cosme-Punta Coyote es de aproximadamente 600 personas que habitan en su mayoría en casas de cemento y concreto, obtienen la energía eléctrica a través de celdas fotovoltaicas y el abastecimiento de agua lo realizan a través de ojos de agua, pozos, de una desalinizadora e inclusive de yates. El acceso a seis de las 15 comunidades se realiza a través de caminos de terracería y al resto sólo es posible acceder a través de alguna embarcación. El esfuerzo pesquero permanente en la zona del Corredor actualmente es de 96 pangas empleadas por 182 pescadores, lo que representa un aumento de 4% en la cantidad de embarcaciones y 8% en el número de pescadores a comparación del 2009 (Sociedad de Historia Natural Niparajá A.C., Pronatura Noroeste A.C. y Iemanya Oceánica A.C., 2009). En ese año sólo 45% de las embarcaciones tenían permiso de pesca comercial, sin embargo hoy en día alrededor de 90% cuentan con al menos un permiso para la extracción ya sea de escama, tiburón

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
144
y raya o almeja. Actualmente existen siete SCPP conformadas en el área, de las cuales cinco están agremiadas en una Federación. La problemática que de acuerdo a la percepción del sector pesquero ha disminuido en mayor medida, es la falta de permisos para la pesca de escama, lo cual fue resultado de la participación de los pescadores en el POP, mientras que por otro lado una de las problemáticas que ha empeorado un poco es la presencia de pescadores que emplean equipo de buceo y arpones para la captura de escama y de algunas otras especies de invertebrados durante la noche (Tabla 3), aun con el establecimiento de ZDR. Actualmente ha disminuido la disponibilidad que existía en 2009 por compartir sus zonas de pesca con pescadores de otros estados.
Tabla 3. Principales problemáticas del sector pesquero en el Corredor San Cosme-Punta Coyote y propuestas para solucionarlas.
Problemáticas identificadas Promedio en la escala
Falta de permisos para pesca de escama. 4.3
No pueden acceder a créditos ni programas de apoyo gubernamental por falta de permisos.
4.1
Barcos de La Paz, Sonora y Sinaloa arrastrando en las zonas de pesca y o crianza de escama.
3.7
Competencia entre los diferentes artes de pesca por los recursos en una zona 3.2
Disminución de los recursos pesqueros.
2.9
Presencia de pistoleros en la zona. 2.8
Soluciones propuestas Promedio en la escala
Prohibir las actividades de los pescadores que trabajan con compresor
4
Involucrar a las autoridades en los problemas de pesca del Corredor 3.8
Involucrar a las comunidades para colaborar en las labores de vigilancia 3.7
Obtener permisos para otras especies diferentes a la escama y al tiburón 3.7
No aumentar el esfuerzo pesquero 2.7
Establecer áreas compartidas de trabajo con pescadores de otros estados que sigan las reglas de pesca en la zona
2.1
Resulta importante prestar atención a la problemáticas dentro y entre las comunidades ya que las reservas marinas, como son las zonas de refugio pesquero, pueden promover la cooperación y la
competencia entre los pescadores, y mientras se mantengan en equilibrio pueden conducir a una mejor conservación de los recursos marinos. Sin embargo, si alguna de las dos aumenta mientras que la otra disminuye, el éxito y trascendencia del área protegida y de la biodiversidad y comunidades que dependen de ella se verán amenazados (Basurto et al. 2016). Conclusiones Las comunidades ubicadas en la zona del Corredor San Cosme-Punta Coyote son poblados pequeños con acceso limitado a los servicios de vivienda y salud cuya economía depende en gran medida de la pesca. A comparación del 2009 el esfuerzo pesquero ha aumentado, tanto en número de embarcaciones como en cantidad de pescadores debido a que las nuevas generaciones se sumaron a la actividad. La problemática continua siendo la misma que en 2009 con diferentes matices en cada una de las comunidades, sin embargo se han realizado algunos avances al respecto y varias de las soluciones propuestas por los pescadores se han concretado. Literatura citada Basurto X, E Blanco, M Nenadovic, B Vollan. 2016.
Integrating simultaneous prosocial and antisocial behavior into theories of collective action. Science Advances 2: 1-10
DOF. 2012. Acuerdo por el que se establece una red de zonas de refugio en aguas marinas de jurisdicción federal frente a la costa oriental del Estado de Baja California Sur, en el corredor marino de San Cosme a Punta Coyote. Diario Oficial de la Federación, 16 de noviembre.
Sociedad de Historia Natural Niparajá A.C., Pronatura Noroeste A.C. y Iemanya Oceánica. 2009. Conociendo el Corredor una descripción de las comunidades pesqueras, su problemática y posibles soluciones. Reporte interno. http://npj.niparaja.org/wp-content/uploads/2015/06/Conociendo_Corredor_2009.pdf
Palabras Clave: Pesca ribereña, esfuerzo pesquero, refugio pesquero, problemáticas, soluciones.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
145
069: oral ¿ES LA CAPTURA DE CAMARON EN LA ZONAS LAGUNARES Y COSTERAS LA CAUSANTE DEL
DETERIORO DE LA PESQUERÍA DE CAMARON ROSADO EN LA SONDA DE CAMPECHE?
Armando T. Wakida Kusunoki1, Ramón Isaac Rojas Gónzalez2, Alejandra Toro Ramírez1, Humberto Armando Medina Quijano1, José Luis Cruz Sánchez1, Luis Daniel Santana2 e Ismael CarrilloNolasco2
1Centro Regional de Investigación Pesquera de Yucalpetén. INAPESCA. Boulevard del Pescador S/N, Esq. Con antigua Carretera a Chelem , Puerto de Abrigo, Yucalpetén, Progreso, Yucatán. C.P. 97320. [email protected]; 2Centro Regional de Investigación Pesquera de Lerma. INAPESCA. Carretera Campeche-Lerma Km. 5. Lerma, Campeche, México. C.P. 24500. Introducción La Carta Nacional Pesquera menciona que el camarón rosado está en deterioro (sobreexplotado), ya que su captura se encuentra en los niveles de producción históricamente más bajos (DOF 2012). Entre las causas consideradas para explicar este deterioro de la población de camarón rosado Farfantepenaeus duorarum, destaca el incremento desmedido del esfuerzo de pesca en las décadas de los setenta y ochenta y las elevadas tasas de mortalidad asociadas, la sobrepesca de juveniles en la zona costera y la Laguna de Términos (Gracia 1995). En este trabajo se presenta un análisis de la pesca de camarón en la zona costera de Campeche y Yucatán durante un año completo. Para cada comunidad se describen: a) el número de pescadores dedicados a esta actividad, b) las artes de pesca empleadas, c) su uso, d) las distribuciones de longitudes de las capturas en las zonas, y e) el posible efecto de esta actividad en las capturas en altamar. Materiales y métodos Para el desarrollo de esta investigación se realizaron visitas mensuales de enero a diciembre del 2015 a las principales comunidades pesqueras de Campeche y Yucatán, que abarcaron desde Sabancuy en el estado de Campeche hasta Las Coloradas en Yucatán. Los muestreos consistieron en conteos de pescadores dedicados a la pesca de camarón desde diferentes puntos de observación fijos y móviles, en recorridos a pie y a bordo de vehículos terrestres y acuáticos. Se tomaron las características de las artes de pesca observadas y se estimaron los rendimientos pesqueros, así como las distribuciones de longitudes, para cada localidad. Con lo anterior, se calcularon los cambios de longitudes y número de organismos en el tiempo, utilizando el crecimiento individual y mortalidad natural. Con cada composición mensual de longitudes se estimó la máxima biomasa que alcanzaría y la captura en altamar que se tendría.
Resultados y discusiones Se realizaron en total 74 censos, donde se observó que sólo en tres localidades se captura camarón durante todo el año: Champotón e Isla Arena en Campeche y Celestún en Yucatán. En el resto, sólo se captura de camarón de forma esporádica e intensa, en periodos cortos principalmente durante la presencia de “nortes”. Esta época coincide con los periodos de mayor reclutamiento del camarón rosado (Wakida-Kusunoki et al. 2008). La mayor presencia de pescadores se registró en Champotón donde se detectó un máximo de 82 y un mínimo con dos pescadores en Chabihau, Yucatán. Se observó el uso de ocho tipos de artes de pesca. En la mayoría de las zonas, la actividad se realiza sin el uso de embarcaciones motorizadas, que sólo se registraron en Río Lagartos. Algunas de las artes de pesca producen mayor impacto ecológico que otras, un ejemplo de ellas son las artes de pesca pasivas tales como las redes de corriente y los triángulos pasivos, ya que por la forma en que se utilizan y los lugares donde se colocan, es difícil que exista escape de los organismos y producen una mortandad de peces y crustáceos en estado juvenil. La composición de longitudes de las diferentes zonas presentaron diferencias, se observó que las modas en Champotón, Isla Arena y Celestún son más pequeñas comparadas con las demás zonas (entre 35 y 45 mm de LT). La comparación de longitudes, artes de pesca y forma de operación demostró diferencias significativas entre las localidades (ANOVA F8,545 = 118.24, P˂0.00). Estas diferencias en las distribuciones son resultado del tipo de artes utilizadas. Los rendimientos de pesca fueron altamente variables por época del año y por el arte de pesca utilizado, pero los mayores se observaron en las artes de pesca pasivas en Celestún y Progreso con máximos entre 150 a 300 kilos por noche y en Río Lagartos con redes de arrastres, sus capturas pueden ser de hasta 60 kilogramos. El estimado de la pérdida por cada kilo que se captura por las artes de pesca utilizadas en las diferentes zonas, se estimó que está entre cinco y siete kilogramos que se dejarían de pescar y entre 8 y 11 kilogramos que se dejarían de tener en el mar.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
146
El análisis del efecto de la captura de juveniles de camarón rosado, da como resultado un estimado de captura anual de 92 t de camarón entero, que equivalen a 333 t de camarón sin cabeza que se dejaron de capturar en altamar, estimando un decremento del 26% con respecto a la captura reportada por la flota. Conclusiones a) En Champotón, Isla Arena y Celestún la captura de camarón se realiza todo el año; en las demás zonas se realiza principalmente en épocas de nortes aprovechando la migración que genera el reclutamiento en altamar. b) La zona de pesca donde se aplica mayor esfuerzo pesquero en número de pescadores es Champotón. c) La captura se realiza con por lo menos ocho artes de pesca distintas. d) Las longitudes mayores se registraron con artes de pesca que capturan camarones que se encuentran en migración, estas son las artes pasivas como el mundo y redes de corriente. El efecto de sólo las zonas donde se captura camarón rosado es de un estimado de 333 t de camarón sin cabeza que se dejaron de capturar en altamar y un decremento estimando del 26% con respecto a la captura reportada por la flota Literatura Citada DOF. 2012. Acuerdo por el que se da a conocer la
Actualización de la Carta Nacional Pesquera. Diario Oficial de la Federación. México. 24 de agosto del 2012.
Gracia A. 1995. Impact of artisanal fishery on production of the pink shrimp Penaeus Farfantepenaeus duorarum Burkenroad, 1939. Ciencias Marinas 21(3):343-359.
Wakida-Kusunoki AT, L Garcia-Solorio, NG Vázquez Benavides. 2008. Abundancia de juveniles de camarones peneidos comerciales en la zona norte de Laguna Madre, México. Hidrobiológica 18 (1): 85-88.
Palabras Clave: Pesca ribereña, camarón rosado, Campeche y Yucatán.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
147
070: cartel ESTRUCTURA DE LONGITUDES y EDADES DE Rhinoptera steindachneri, Dasyatis dipterura y D. longa CAPTURADAS INCIDENTALMENTE DURANTE ARRASTRES DE CAMARÓN EN LA ENTRADA AL GOLFO
DE CALIFORNIA (2011–2016)
Lucía Elizabeth López López1, Javier Tovar Ávila2 y Darío A. Chávez Arrenquín3
1CUCBA, UDG. Camino Ramón Padilla Sánchez 2100, Las Agujas, Zapopan, C.P. 44171, Jalisco, México. [email protected]. 2Centro Regional de Investigación Pesquera de Bahía de Banderas. INAPESCA. Tortuga 1, La Cruz de Huanacaxtle, Nayarit, México. C. P. 63732. 3C entro Regional de Investigación Pesquera de Mazatlán. INAPESCA.
Mazatlán, México. Introducción Numerosas especies de importancia comercial para las pesquerías ribereñas son capturadas como fauna de acompañamiento en la pesquería industrial de arrastre de camarón, representando un impacto para sus poblaciones y los pescadores ribereños (García-Caudillo y Gómez-Palafox 2005). Entre las especies capturadas se encuentran Rhinoptera steindachmneri, Dasyatis dipterura y D. longa. El conocimiento de la composición de longitudes y edades de las capturas incidentales es de gran importancia, ya que pueden ayudar a evaluar el impacto de esta pesquería en sus poblaciones. El método más utilizado para estimar la edad en los elasmobranquios es el conteo de bandas de crecimiento en sus vértebras (Cailliet y Goldman 2004). En el presente estudio se estima por primera vez la estructura de longitudes y edades de las rayas antes mencionadas en las capturas de prospecciones de camarón en la entrada del Golfo de California (Sur de Sinaloa y Nayarit) durante 2011–2016. Materiales y métodos Los organismos analizados fueron capturados durante arrastres de camarón a bordo del BIP XII e INAPESCA I de 2011 a 2016. Las muestras fueron congeladas abordo y transportadas al CRIP Bahía de Banderas, Nayarit, para su procesamiento. En el laboratorio se identificó la especie, se determinó el sexo, se obtuvo el ancho de disco (AD) y se extrajeron de 5-10 vértebras de la región abdominal de cada organismo. Las vértebras se limpiaron con un bisturí e hipoclorito de sodio, de éstas se obtuvieron cortes sagitales de 300–500 micras con una sierra de baja velocidad Isomet (Buehler). Los cortes fueron observados en un microscopio estereoscópico con luz transmitida, contando los pares de bandas de crecimiento (opaca y translúcida) tres veces de manera no consecutiva por dos lectores independientes. Para estimar la precisión de los conteos se utilizó el coeficiente de variación (CV) (Chang 1982) y el porcentaje de error promedio (APE) (Beamish y Fournier 1981). La edad de los organismos se estimó a partir del número de bandas contadas menos uno, considerando la primera marca
translúcida como la marca de nacimiento. Se asumió que las bandas se forman de manera anual, como ha sido comprobado en la mayoría de los elasmobranquios (Cailliet y Goldman 2004). Se determinó la proporción sexual de las capturas para cada especie analizada. La estructura de longitudes y edades por sexo se analizó mediante histogramas de frecuencia, determinando si existían diferencias significativas mediante análisis de varianza (ANDEVA) o pruebas de t-Student, cuando no se cumplió el supuesto de homocedasticidad. Resultados y discusión Se analizaron 409 organismos pertenecientes a las tres especies: 315 de R. steindachneri, 104 D. dipterura y 20 D. longa. La precisión de los conteos de las bandas de crecimiento fue alta para un mismo lector (APE: 5.3, 11.3 y 10.9% y CV: 3.77, 4.18 y 5.21 para R. steindachneri, D. longa y D. dipterura respectivamente) y entre ambos lectores (APE: 4.3, 6.81 y 10.18% y CV: 3.04, 4.82 y 6.14%). Similares niveles de precisión han sido reportados para R. steindachneri (Pabón-Aldana 2015) y D. dipterura (Smith et al. 2007), no existiendo estudios para D. longa. Se registró un mayor número de hembras que machos de R. steindachneri (1.26H:1M), no así para D. dipterura (1H:1.31M) y D. longa (1H:1.5M) en las cuales el número de machos superó al de las hembras. No se encontraron diferencias significativas entre las longitudes de machos y hembras de las tres especies: R. steindachneri (ANDEVA295,296= 2.043, P>0.05), D. dipterura (ANDEVA 92,93= 3.241, P>0.05) y D. longa (t-student13.591,18= 0.839, P>0.05), ni entre las edades de ambos sexos de R. steindachneri (ANDEVA216,217= 2.645, P>0.05) y D. dipterura (ANDEVA 70,71= 2.567, P>0.05) (Tabla 1, Figs. 1 y 2).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
148
Tabla 1. Intervalo de longitudes registradas y edades estimadas de las rayas capturadas en los arrastres camaroneros (sexos combinados)
Longitud (ancho de disco, cm) Edad (años)
R. steindachneri
23.7–61.5 (X=38.4, D.E.= 5) 0–3 (X=0.5, D.E.=0.7)
D. dipterura
13–53.7 (X=31.2, D.E.=10.1) 0–11 (X=2.7, D.E.=2.2)
D. longa
18–52.8 (X=35.2, D.E.=9.5) 1–11 (X=5.3, D.E.=3.4)
Fig. 1. Frecuencia de longitudes registradas.
Fig. 2. Frecuencia de edades estimadas. Se determinó que los organismos capturados durante los arrastres de camarón en la zona de estudio son en su mayoría juveniles, siendo las longitudes y edades registradas en el presente estudio menores a las reportadas por diversos autores para R. steindachneri (de 0 a 7 años en organismos de 28–96 cm de AD) (Gutiérrez-Rivera et al. 2016) y D. dipterura (de 0 a 28 años en organismos de 26–83 cm de AD) (Smith et al. 2007). Cabe destacar que los organismos de estos estudios son capturados por pesquerías ribereñas de manera dirigida.
Conclusiones Las vértebras de las tres especies estudiadas presentan bandas de crecimiento con alta precisión. Las rayas capturadas en los arrastres de camarón son de menor longitud y edad que las capturadas en la pesca dirigida. Todos los organismos capturados de estas especies en los arrastres camaroneros son juveniles. Literatura citada Beamish RJ, DA Fournier. 1981. A method for comparing
the precision of a set of age determinations. Can. J. Fish Aquat. Sci. 38:982-983.
Cailliet GM, KJ Goldman. 2004. Age Determination and Validation in Chondrichthyan Fishes. En: Carrier JC, JA Musick, MR Heithanus (Eds). Biology of sharks and their relatives. CRC Press, Boca Raton, Florida, E.U. pp: 399–447.
Chang WYB. 1982. A statical method for Evaluating the reproducibility of age determination. Can. J. Fish Aquat. Sci. 39:1208-1210.
García-Caudillo JM, JV Gómez-Palafox. 2005. La pesca industrial del camarón en el Golfo de California: Situación económico-financiera e impactos socio-ambientales. Conservación internacional. Región Golfo de California. Guaymas, Sonora, México. 104 p
Gutiérrez-Rivera OA, D Voltolina-Lobina, D Corro-Espinosa, A Muhlia-Melo, G Andrade-Domínguez, A Torres. 2016. Edad y crecimiento de la raya tecolote Rhinoptera steindachneri (Evermann y Jenkins 1891) en las costas de Sonora y Sinaloa, México. En: Aguilar C, A Díaz-Sánchez (Eds.). VII Simpoisum Nacional de Tiburones y Rayas. SOMEPEC 180p
Pabón-Aldana KA, FN Mello-Barrera, PA Mejía-Falla, VH Cruz-Escalona. 2016. Estimación de la edad de la Raya Tecolote (Rhinoptera steindachneri) en la Bahía de La Paz, BCS, México. En: Aguilar C, A Díaz-Sánchez (Eds.). VII Simposium Nacional de Tiburones y Rayas. SOMEPEC 180p
Smith WD, GM Cailliet, EM Melendez. 2007. Maturity and growth characteristics of a commercially exploited stingray, Dasyatis dipterura. Mar. Fresh. Res. 58: 54–66.
Palabras clave: Batoideos, vértebras, fauna de acompañamiento.
0
20
40
60
80
100
120
140
160
0
5
10
15
20
25
30
35
10 15 20 25 30 35 40 45 50 55 60 65
Fre
cue
nci
a R
. st
ei
Fre
cue
nci
a (
D.
dip
t y
D.
lon
g)
Ancho de Disco (cm)
Dasyatis longa
Dasyatis dipterura
Rhinoptera steindachneri
0
20
40
60
80
100
120
140
160
0
2
4
6
8
10
12
14
16
0 1 2 3 4 5 6 7 8 9 10 11 12
Fre
cue
nci
a R
. st
ei
Fre
cue
nci
a (
D.
lon
g y
D.
dip
t)
Edad
Dasyatis longa
Dasyatis dipterura
Rhinoptera steindachneri

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
149
071: oral ANÁLISIS DE LA PESQUERÍA DE ESCAMA MARINA EN LA ZONA NORTE DE CAMPECHE
Rosa Guadalupe Morales Martínez1 y Josefina Santos Valencia 2
1Centro Regional de Investigación Pesquera de Lerma. Carretera Campeche Lerma, s/n. 2Centro Regional de Investigación Pesquera de Yucalpetén. INAPESCA. Blvd. del pescador s/n Puerto de Abrigo, Yucalpetén, Yuc. C.P. 97320. [email protected]; [email protected] Introducción Campeche es uno de los principales productores de especies marinas en el Golfo de México, cuyo principal componente es la captura la escama marina. En la Zona Norte de Campeche (ZNC) se producen alrededor de 10 mil toneladas anuales de peces marinos. La captura se compone de poco más de 100 especies comerciales, con siete de ellas de mayor importancia por su volumen y valor: jurel (Caranx hippos, y C. latus), corvina (Cynoscion griseus y C. arenarius), pargo mulato (Lutjanus griseus), cojinúa (Caranx crysos), huachinango (Lutjanus campechanus), sierra (Scomberomorus maculatus) y carito (Scomberomorus cavalla. La pesca de escama tiene un gran impacto social en la ZNC donde se registran 2,772 embarcaciones menores y más de cinco mil pescadores. Se utilizan cuatro sistemas de pesca: redes de enmalle, palangre, línea de mano y buceo, siendo el primero el más utilizado por la flota ribereña. El objetivo de este trabajo fue caracterizar la pesquería de escama marina, analizando la estructura de tallas de las especies principales, la captura por unidad de esfuerzo (CPUE) y los rendimientos económicos por viaje de pesca. Materiales y métodos El área de estudio se localiza en el litoral norte de Campeche en cuatro principales localidades pesqueras: Champotón, Seybaplaya puerto de Campeche e Isla Arena. Se monitoreó la captura de 141 embarcaciones con redes de enmalle como arte de pesca. Se midieron 11 mil organismos (8,954 kg) de las seis principales especies: se les tomó el peso (g) y longitud (cm). La CPUE se estimó en kilogramos por viaje diario de pesca y se estimaron los costos variables de los viajes de pesca por medio de entrevistas aplicadas a los pescadores de escama marina. Resultados y discusión En 2015 la captura de escama se compone de 58 especies comerciales, siete representan 50%: jurel (16%), corvina (13%), pargo (7%), cojinúa (5%), huachinango (5%), sierra (3%) y carito (1%). El 50% restante en conjunto corresponde a las otras 52 especies (Fig. 1).
Fig. 1. Capturas (%) de las principales especies en la zona norte de Campeche, 2015.
Fig. 2. Distribución de las frecuencias de longitudes La estructura de longitudes de las especies analizadas (Fig. 2) muestra que la captura está compuesta tanto por juveniles como por adultos en proporciones variables según la especie. En la distribución de longitudes del jurel se observan dos cohortes: la primera entre 24 y 38 cm LT y la

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
150
segunda entre 48 y 64 cm LT. La corvina tuvo una cohorte de 18 a 64 cm, igual que el pargo mulato de 21 a 52.5 cm. Para esta especie, Claro et al. (2001) reportan que la longitud mínima de madurez sexual es de 22 cm en hembras y 23 cm en machos. Con base en lo anterior, 3.9% de las longitudes capturadas en la ZNC se encuentran por debajo de la mínima a la madurez. La cojinuda es un pez de alta demanda en el mercado por su bajo costo. En su estructura de longitudes, la moda se ubica a los 34 cm, con una cohorte de entre 30 y 40 cm LT. El Huachinango presenta un intervalo más amplio de entre 16 y 78 cm LT. Byron y Palmer (2004) establecieron la longitud de primera madurez a los 37.8 cm en hembras y 22.3 cm en machos, por lo cual 67.7% de los organismos capturados se encuentran por debajo de la talla mínima de captura. En los peces pelágicos migratorios se observó diferente estructura de longitudes: la sierra presenta una sola cohorte entre 40 y 60 cm LT, el carito dos cohortes: la primera entre 44 y 66 cm LT y la segunda entre 74 y 90 cm LT. La longitud de primera madurez de la sierra es de 26 cm LP (Collette y Nauen 1983), por lo que la totalidad de los ejemplares capturados tienen tallas superiores a ésta.
Fig. 3. CPUE mensual en la flota ribereña de la zona norte de Campeche.
La CPUE total de los viajes de pesca de la flota menor en 2015 se muestra en la figura 3, los mayores rendimientos se registraron en septiembre, con más de 95 kg por día efectivo de pesca, y rendimiento promedio de 39 kg por día de pesca. Los costos totales por viaje de pesca de la flota ribereña que opera con redes de enmalle en la ZNC van de $100 a $250.00, de los cuales alrededor de 80% representa el costo del combustible. Los ingresos por venta del producto varían entre $277 y $2,156 pesos y el beneficio neto (cuasi renta) generados por viaje de pesca van de $184.00 a $1,900.00, siendo marzo, abril y mayo cuando se obtienen ingresos superiores a $1,500.00, mismos que normalmente son repartidos entre dos pescadores que participan en la jornada de pesca (Fig. 4). La menor cuasi renta se obtiene entre junio-
julio y en los meses posteriores gran parte de la flota ribereña de la ZNC cambia sus redes por jimbas para la captura de pulpo.
Fig. 4. Costos variables en la operación de viaje de pesca, ingresos totales por viaje. Conclusión En la Zona Norte de Campeche la pesca ribereña de escama es de gran importancia social que aporta casi 10 mil toneladas anuales, de las cuales 50% la constituyen siete especies comerciales. Las operaciones de pesca son diarias y los rendimientos por viaje de pesca son muy variables, igual que los beneficios netos a lo largo del año, siendo de marzo a mayo los meses de mayores ingresos, mientras que en junio y julio se perciben los menores bajos ingresos. Literatura citada Claro R, R Baisre, K Lindeman, P García-Arteaga. 2001.
Cuban fisheries: historical trends and currentstatus. In: Claro, R., K.C. Lindeman K. y L.R. Parenti (Eds.). Ecology of the marine Fish of Cuba. Smithsonian Institution, Washington DC.194-219.
Collette BB, CE Nauen. 1983. FAO species catalogue. Vol. 2. Scombrids of the world. An annotated and illustrated catalogue of tunas, mackerels, bonitos and related species known to date. FAO Fish. Synop. 125. Vol. 2, 137 p.
Palabras clave: Pesca ribereña, Zona norte de Campeche, rendimientos, Cuasi renta.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
151
072: cartel CALIDAD DE VIDA, PESCA RIBEREÑA Y TURISMO: CASO COSTA DE JALISCO, MÉXICO
Rosa María Chávez Dagostino, Myrna Leticia Bravo Olivas y Oscar A. Maldonado Ibarra
1Centro Universitario de la Costa, Universidad de Guadalajara, Puerto Vallarta, Jalisco, México. [email protected] ,2Instituto Tecnológico de Bahía de Banderas, Nayarit, México. 3Universidad Tecnológica de Escuinapa, Sinaloa, México. Introducción La pesca ribereña es una actividad importante para la seguridad alimentaria y la reducción de la pobreza (FAO 2012), sin embargo, está en crisis. Para la región Centro Pacífico Mexicano (incluye la costa de Jalisco), se ha reportado que 68.7% de las pesquerías están colapsadas o sobrexplotadas, con una tendencia de captura de especies de altos niveles tróficos y se considera la más deteriorada del país (Arreguín-Sánchez y Arcos-Huitrón 2011). Por otro lado, la pobreza de las comunidades pesqueras persiste. El turismo se considera la actividad económica más importante del mundo debido a su crecimiento, ingresos y empleos que genera. Ambos sectores conviven en las regiones costeras, creando sinergias o conflictos por el uso de los recursos en general, situación que debe afectar la calidad de vida de los locales. Puede afirmarse que el desarrollo turístico beneficia a la pesca y viceversa, pero pronto la sinergia entre “el bello pueblo de pescadores, la disponibilidad de alimento fresco y de bajo costo” y la actividad turística, es aniquilada por el crecimiento turístico voraz (Cabrera-Socorro y Cabrera-Socorro 2004). El desplazamiento humano para dar paso a macro-proyectos turísticos es muy frecuente (Alcalá-Moya 2003). La transformación del paisaje bajo el influjo del turismo fue consecuencia de la puesta en marcha del modelo neoliberal a nivel nacional que provocó ajustes estructurales en la economía de poblados especialmente en los dedicados a la pesca artesanal, donde el mercado tiene un papel preponderante en el uso del territorio costero, disminuyendo la participación del Estado (Cruz-Coria et al. 2013). El objetivo de este trabajo fue analizar la calidad de vida los pescadores ribereños en la costa de Jalisco, y relacionarlo con la actividad turística, bajo la hipótesis de que una relación no conflictiva con la actividad turística produce niveles mayores de calidad de vida en los pescadores. Material y métodos En ocho cooperativas de las localidades de Puerto Vallarta, Chimo, Cruz de Loreto, Punta Pérula, Barra de Navidad, Careyitos, Rebalsito y Chamela en la costa de Jalisco, se aplicaron 105 encuestas a pescadores, con preguntas cerradas sobre ingresos, importancia y grado de satisfacción de 11 indicadores (Tabla 1) para obtener la calidad de vida (CV)
promedio por cooperativa (Bravo-Olivas et al. 2015) y su y relación con la actividad turística (Tabla 2). Los valores de importancia y satisfacción de cada pescador fueron interpretados y convertidos en puntuación de calidad de vida (de +10 a -10) según lo propuesto por Brown et al. (1998) donde los valores arriba de 4.5 fueron considerados como excelente; los valores entre 1.5 y 4.5 bueno, de -1.5 a 1.5 como adecuado; problemático de -1.5 a 1.5 y, muy problemático los valores menores que -4.5 y promediada por cooperativa. Con base en la respuesta de su relación con la actividad turística, se establecieron tres grupos y se codificaron: los que tenían relación sinérgica (0), conflictiva (1) o ninguna (2). Se asociaron las variables de calidad de vida y su relación con el turismo, mediante un análisis de varianza de dos vías (SigmaPlot 11.0). Resultados La CV promedio de los pescadores fue de 4.23 (desviación estándar d.e. = 2.6), donde lo más importante y satisfecho es la familia (Tabla 1). Debajo del nivel aceptable se encuentran el costo de la vida y las vacaciones. Transporte, compras y nivel escolar. La relación con el turismo percibida por los pescadores fue sinérgica si se beneficiaban, mutuamente. Las cooperativas con relación sinérgica con el turismo fueron tres y las conflictivas, fueron cuatro (Tabla 2). El análisis de varianza no reveló una interacción significativa (p >0.05) entre la calidad de vida y el tipo de relación con las diferentes cooperativas. Tabla 1. Calidad de Vida de pescadores con base a la importancia (I) y satisfacción (S) por indicador
Indicador I S CV
Salud 5.0 4.0 6.2 Costo de la vida 4.53 3.26 1.31 Oport. de trabajo 4.5 3.86 4.25 Vivienda 4.72 4.15 5.59 Familia 4.96 4.81 9.07 Amigos 4.19 4.31 6.23 Compras 4.21 3.51 2.47 Transportación 4.4 3.31 1.56 Vacaciones 3.7 3.02 0.71 Agua 4.84 4.26 6.2 Nivel escolar 4.54 3.62 3.1

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
152
Tabla 2. Calidad de vida (CV) por cooperativa y tipo de relación con la actividad turística
Cooperativa Rel. CV Rosita 0 6.47 Punta Pérula 0 3.9
Barra de Navidad 0 4.95
Cruz de Loreto 1 3.71
Rebalsito 1 5.22
Careyes 1 5.37
Chamela 1 5.6
Rivera de Suchitlán 2 2.45
Discusión y conclusiones Los recursos costeros se consideran estratégicos por sus características ambientales y socio-espaciales, por lo que la pesca se convierte en un obstáculo para la inversión extranjera y la apropiación del territorio por parte del capital turístico, deteriorando la calidad de vida, sin embargo, estadísticamente no se pudo probar esta asociación por lo que deberán incluirse otras variables. Literatura citada Alcalá-Moya G. 2003. Políticas pesqueras en México
(1946-2000). Contradicciones y aciertos en la planificación de la pesca nacional. México: El Colegio de México.
Arreguín-Sánchez F y E Arcos-Huitrón. 2011. La pesca en México: estado de la explotación y uso de los ecosistemas. Hidrobiológica 21(3): 431-462.
Bravo-Olivas ML, RM Chávez-Dagostino, CD Malcolm, R Espinoza. 2015. Notes on the quality of life of artisanal small-scale fishermen along the Pacific coast of Jalisco, México. Sustainability 7: 6046-6068.
Brown I, D Raphael, R Renwick. 1998. Quality of Life Profile, Adults, Quality of Life Resources, Adult Series; Item #2–1; Quality of Life Research Unit, Centre for Health Promotion, University of Toronto: Toronto, ON, Canada
Cabrera-Socorro G y A Cabrera-Socorro. 2004. Turismo versus pesca artesanal. A propósito de la Reserva Marina de la Isla La Graciosa y los Islotes del Norte de Lanzarote. Pasos. Revista de Turismo y Patrimonio Cultural 2 (1):1-16
Cruz-Coria E, L Zizumbo-Villarreal, N Monterroso-Salvatierra y A Quintanilla. 2013. La transformación de los espacios de vida y la configuración del paisaje turístico en Puerto Morelos, Quintana Roo, México. POLIS 12(34): 307-332
FAO. 2012. Examen mundial de la Pesca y Acuicultura. Parte I. http://www.fao.org/docrep/016/i2727s/i2727s01.pdf
Palabras clave: Satisfacción, comunidades pesqueras, cooperativas, sinergia, desarrollo turístico.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
153
073: cartel HUELLA ECOLÓGICA MARINA DE LAS PESQUERÍAS RIBEREÑAS EN LA COSTA DE NAYARIT
Myrna Leticia Bravo Olivas1, Jocelin González Cedano1, Karen Yareth López Álvarez1 y
Rosa María Chávez Dagostino2
1Instituto Tecnológico de Bahía de Banderas. Bahía de Banderas, Nayarit. C.P 63734; 2Centro Universitario de la Costa, Universidad de Guadalajara, Puerto Vallarta, Jalisco. C.P 48280; [email protected] Introducción El uso de los recursos pesqueros destinados al consumo humano en el mundo ha aumentado a 130.8 millones de toneladas debido a la acuicultura, aumento de la pesca y la mejora de los canales de distribución (FAO 2012). Dada la necesidad de comprender los impactos de la pesca y su estado, se han desarrollado algunos indicadores. La huella ecológica es un indicador que mide las necesidades humanas en términos de área requerida para la generación de productos y absorción de desechos durante el curso del proceso de producción (Wackernagel y Rees 1996), y se puede utilizar para muchos propósitos. La huella ecológica de la pesca, una herramienta para medir la extensión espacial de la apropiación humana de los ecosistemas marinos en base a la productividad primaria requerida en relación con la captura, se puede utilizar para establecer los impactos ecológicos y la sustentabilidad de la producción y el consumo de pescado en los diferentes niveles. El objetivo de esta investigación es aplicar la metodología de la huella ecológica de la pesca a las capturas ribereñas de la costa de Nayarit, con el fin de conocer los impactos ecológicos y sustentabilidad de la producción pesquera en el área. Material y métodos Se calculó la huella ecológica de la pesca a las capturas obtenidas entre el 2005 y 2014 por la flota ribereña de la costa de Nayarit. Estos se agruparon por grupos taxonómicos y se obtuvo el peso húmedo captura total en toneladas por año para cada grupo. A continuación, cada uno fue asignado a una categoría de nivel trófico, según lo propuesto por Froese y Pauly (2010), Galván-Piña y Arreguín-Sánchez (2004). Se calculó la productividad primaria requerida para las capturas (Pauly et al. 1998). Finalmente se calculó la huella ecológica tomando en cuenta la productividad primaria requerida, el factor de rendimiento y el factor de equivalencia de las pesquerías marinas. Se comparó el resultado de la huella ecológica de la pesca con el de la biocapacidad del área de pesca y se pudo así determinar si existe un déficit ecológico en las capturas ribereñas de la costa de Nayarit. Resultados La captura total en el periodo de 2005 a 2014 fue de 125,857 toneladas, clasificadas en 34 recursos
pesqueros, los cuales en su mayoría fueron peces (79%), seguido por moluscos (12%) y en menor proporción crustáceos. Los mayores registros se obtuvieron en 2011 y los menores en 2006 (Fig. 1)
Fig. 1. Registro de capturas de la costa de Nayarit. El principal recurso capturado es el camarón, el cual representa 42% de los registros (Fig. 2).
Fig. 2. Registros de captura de la flota ribereña en la costa de Nayarit. Se establecieron 23 grupos tróficos, con niveles de entre 2.01 (lisa) y 4.45 (cintilla). La productividad primaria requerida para la captura de estos recursos fue de 2,700,374 g C. La tabla 1 muestra los resultados del cálculo de huella ecológica de la pesca. Indican que en ninguno

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
154
de los casos se sobrepasó la biocapacidad del área de pesca. Tabla 1. Huella ecológica de la pesca y balance ecológico en la costa de Nayarit.
Año FP (ha) Biocapacidad Nayarit (hag)
Balance ecológico (hag)
2005 146,129 6,188,797 6,042,668
2006 127,189 6,073,460 5,946,271
2007 141,599 6,050,541 5,908,942
2008 180,274 5,977,923 5,797,649
2009 225,871 5,886,197 5,660,326
2010 232,191 5,816,140 5,583,949
2011 543,929 5,737,100 5,193,172
2012 559,896 5,680,092 5,120,196
2013 553,534 6,050,541 5,497,007
2014 522,285 5,538,110 5,015,824 Discusión La herramienta “huella de la pesca” permite a los usuarios cuantificar, con un enfoque no tradicional, el impacto sobre los ecosistemas marinos y puede ser utilizado como una herramienta para evaluar la sustentabilidad de los niveles de captura en países o regiones, teniendo en cuenta los efectos del nivel trófico y la biocapacidad de un área específica del océano, donde la captura se expresa en términos de peste de pequeños rumiantes, que es en gran medida una función del nivel trófico de las especies capturadas. En términos de captura, es importante tener en cuenta la falta de fiabilidad de los datos oficiales de captura en México, como en muchos otros países, debido a los datos de las capturas no declaradas e ilegales por parte de pescadores, lo cual era una estimación de 15 % de la captura total en 2000-2003 período en la región oriental del Pacífico central (Agnwe et al. 2009). Este hecho afectaría a la productividad primaria requerida, así como la huella de la pesca. Sin embargo, el equilibrio ecológico aún sería positivo. Aun suponiendo un número mayor del 20% de la captura ilegal o no declarada para esta región, los resultados no se modificarían significativamente.
Conclusiones Con base a los resultados obtenidos, se concluye que en la costa de Nayarit se lleva a cabo una pesquería ribereña sustentable, sin embargo, aunque los resultados indican la pesca sostenible en la región no sólo puede ser un resultado de la evaluación de huella de la pesca, sino que también debe incluir los resultados de otros indicadores
socioeconómicos y ambientales, pero resulta una herramienta importante para la gestión de la pesca. Literatura citada Agnew DD, J Pearce, G Pramod, T Peatman, R Watson,
JR Beddington, TJ Pitcher. 2009. Estimating the worldwide extent of illegal fishing. PLoS One 4: 1–8.
FAO. 2012. The State of World Fisheries and Aquaculture. Food and Agriculture Organization. Fisheries and Aquaculture Department: Rome, Italy. 209 p.
Froese R, D Pauly, 2010. FishBase: Conceptos, estructura y fuentes de datos. ICLARM: Manila, Philippines. 322 p.
Galván-Piña VH, F Arreguín-Sánchez. 2004. Interacting industrial and artisanal fisheries and their impact on the ecosystem of the continental shelf on the Central Pacific coasts of Mexico. In: Proceedings of the 4th World Fisheries Congress, Vancouver, BC, Canada, 2–6 May pp. 587–600.
Pauly D, V Christensen, J Dalsgaard, R Froese, F Torres. 1998. Fishing down marine food webs. Science 272: 860–863.
Wackernagel M, W Rees. 1996. Our ecological footprint. Reducing human impact on the Earth. New Society Publishers: Philadelphia, PA, USA, 160 p.
Palabras clave: nivel trófico, biocapacidad, productividad primaria, balance ecológico, camarón.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
155
074: oral CICLO REPRODUCTIVO DE LA LISA BLANCA Mugil curema EN LAS COSTAS DE MAZATLÁN, MÉXICO:
AVANCES
Irene Marlen Soberano Palomares1, Luis Antonio Salcido Guevara1, Guillermo Rodríguez Domínguez1 y Rebeca Sánchez Cárdenas2
1CONACyT, 2Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa, Paseo Claussen s/n, Col. Los Pinos, Mazatlán, Sinaloa, México. C. P. 82000. [email protected], [email protected], [email protected], [email protected] Introducción Los miembros de la familia Mugilidae forman parte importante en la captura pesquera en gran número de países. M. curema se encuentra en ambas costas de América; en el Atlántico desde Cabo Code hasta Brasil y en el Pacífico desde el Golfo de California hasta Chile (Yáñez-Arancibia 1975). La producción de M. curema en México la posiciona en el lugar 17 en volumen y en lugar 27 respecto a su valor. Sinaloa es el mayor productor, aportando 31.4% en el 2013 (CONAPESCA 2013). M. curema es una de las especies más utilizadas como carnada para la pesquería de peces demersales, de algunos crustáceos (como jaibas y langostas) y para la pesca deportiva (picudos), debido a que se ofrece a un precio bajo y es altamente aceptada por las especies objetivo. Actualmente la temporada de veda reproductiva de M. curema que habita en la costa de Mazatlán, Sinaloa es del 01 de abril al 30 de junio de cada año (DOF 01/12/2015). Sin embargo, esta medida debe ser revisada, dado el constante señalamiento de los pescadores sobre su ineficiencia. Este estudio presenta un avance de la evaluación del ciclo reproductivo de M. curema en Mazatlán, Sinaloa, con el fin de proporcionar información biológica básica para un posible reajuste de la regulación de su veda reproductiva temporal. Esto evitará que la población de lisa blanca para esta zona sea afectada por la extracción de organismos maduros y por lo tanto siga siendo una pesquería sustentable. Material y métodos La información se generó a partir de muestreos sobre las capturas de la pesquería ribereña de Mazatlán, de febrero 2015 a abril 2016. Semanalmente se obtuvieron un promedio de ocho ejemplares al azar con diferentes intervalos de longitud. Los ejemplares fueron procesados inmediatamente en el campo pesquero; se midió la longitud total (LT, cm) y el peso total (PT, g). Los organismos se diseccionaron, se extrajo la gónada y se pesó (PG, g). Se observaron y anotaron en bitácora las características de cada gónada en fresco (color, textura, consistencia, presencia de ovocitos a simple vista), se definió el sexo y se fotografiaron.
A partir de esta información se definió la fase morfocromática de desarrollo gonadal (FMDG) de acuerdo con la escala de Solomon y Ramnarine (2007). El ciclo reproductivo (CR) fue descrito a partir de las frecuencias mensuales de las FMDG y el índice gonadosomático (IGS = 100PG/PT). Además se exploró el CR con respecto a la presencia de precipitaciones (CONAGUA 2016) y el fotoperiodo (González 2011). Resultados y discusión Las frecuencias de las FMDG indican la presencia de organismos en fase de madurez casi todo el año, pero la mayor proporción se presentó en el periodo abril-agosto de 2015, coincidiendo con los promedios más altos de fotoperiodo. Durante este periodo, el IGS mostró un incremento y un declive notable en las hembras y en los machos. Los resultados preliminares sugieren que M. curema presenta un ciclo reproductivo anual en la costa de Mazatlán. Sin embargo, es necesario confirmar mediante un estudio histológico las FMDG, para definir de manera precisa el desarrollo del ciclo y la época de desove. Por ejemplo, es necesario corroborar la presencia de madurez y desove entre septiembre y marzo. Solomon y Ramnarine (2007) reportaron que la época de desove de M. curema en la costa de Venezuela coincide con las precipitaciones. Similarmente, en el presente estudio, la disminución del IGS coincide con el incremento de las precipitaciones en Mazatlán. El valor más alto del IGS para hembras se presentó en los meses de junio y julio y el valor más bajo en diciembre. En el caso de los machos, el valor más alto registrado fue en el mes de marzo y el menor en el mes de diciembre. Los datos de precipitaciones en Sinaloa muestran que los meses con mayor precipitación en 2015 y hasta abril de 2016, se registraron a partir de junio hasta octubre 2016. Los datos de fotoperiodo en Mazatlán indican que los meses con mayores horas luz fueron mayo, junio y julio en el 2015 y en mayo de 2016.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
156
POSTDESOVE DESOVE MADURO DESARROLLO AVANZADO
DESARROLLO INICIAL
INMADURO
FO
TO
PE
RIO
DO
10.0
10.5
11.0
11.5
12.0
12.5
13.0
13.5F
recu
enci
a re
lativ
a (%
)
0
20
40
60
80
100
MARZO
ABRIL
MAYO
JUNIO
JULI
O
AGOSTO
SEPTIEM
BRE
OCTUBRE
NOVIEM
BRE
DICIE
MBRE
ENERO
FEBRERO
1MARZO
1ABRIL
0
20
40
60
80
100
IGS
0
2
4
6
8
10
12
FO
TO
PE
RIO
DO
10.0
10.5
11.0
11.5
12.0
12.5
13.0
13.5
IGS
0
1
2
3
4
5
PR
EC
IPIT
AC
ION
ES
(m
m)2
0
50
100
150
200
250
300
PR
EC
IPIT
AC
ION
ES
(m
m)2
0
50
100
150
200
250
300
IGS FOTOPERIODO PRECIPITACIONES
Fig. 1. FMDG e IGS de M. curema de la costa de Mazatlán, México, en correspondencia con las precipitaciones y el fotoperiodo. La veda reproductiva de M. curema está protegiendo el periodo de abril-junio (DOF 01/12/2015), y considerando el periodo de abril-agosto de las mayores frecuencia de la fase de madurez, quedan desprotegidos los meses de julio y agosto. Estos meses también parecen ser importantes para el desove, ya que el IGS de las hembras muestra su mayor inclinación. Conclusión La mayor intensidad de madurez está sincronizada con los mayores fotoperiodos, y el desove con las precipitaciones. La veda reproductiva no protege los meses de julio y agosto, que son importantes para la madurez y el desove de M. curema. Literatura itada CONAGUA. 2016. Resúmenes mensuales de temperaturas
y lluvia. En línea: http://smn.cna.gob.mx/es/climatologia/temperaturas-y-lluvias/resumenes-mensuales-de-temperaturas-y-lluvias
CONAPESCA. 2013. Anuario estadístico de acuicultura y pesca. SAGARPA. México, D.F. 299 pp.
DOF. 2015. NOM-016-SAG/PESC.2014. SAGARPA. Publicado el 01de diciembre del 2015.
Gonzalez JI. 2011. Predicción de mareas en México. MAR V1.0. En línea: http://oceanografía.cicese.mx/predmar.
Solomon FN, IW Ramnarine. 2007. Reproductive biology of white mullet, Mugil curema (Valenciennes) in the Southern. Carib. Fish. Res. 88: 133–138.
Yáñez-Arancibia A. 1975. Observaciones sobre Mugil curema (Valenciennes) en áreas naturales de crianza, México. Alimentación, crecimiento, madurez y relaciones ecológicas. Anales del Centro de Ciencias del Mar y Limnología (UNAM). México.
Palabras clave: reproducción, índice gonadosomático, precipitaciones, fotoperiodo, medida de manejo

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
157
075: oral DESCRIPCIÓN DEL CICLO REPRODUCTIVO DE Paralabrax nebulifer, EN LA COSTA OCCIDENTAL DE
BAJA CALIFORNIA SUR, MÉXICO
Laura C. Zúñiga Pacheco1, Marcela Zuñiga Flores1, Carmen G Gómez Gauna2 y José A. Gracia Olea2
1Centro Regional de Investigación Pesquera de La Paz. INAPESCA. [email protected]. 2Pronatura Noroeste A.C. Introducción El verdillo Paralabrax nebulifer es un serránido muy abundante en las costas de Baja California Sur. Se posiciona como el de mayor importancia en la pesquería de escama, representando 29% de la captura total de B.C.S. Sin embargo, existe poca información acerca de su biología reproductiva, lo cual es un aspecto fundamental para poder optimizar su sustentabilidad. El objetivo de esta investigación es conocer la proporción de sexos, describir el ciclo reproductivo mediante observación directa, obtener el índice gonadosomático y calcular la longitud de primera madurez. Material y métodos Se realizaron muestreos mensuales de febrero de 2013 a enero de 2014, en la costa occidental de BCS principalmente las zonas (Guerrero Negro, Punta Abreojos, San Juanico, Laguna San Ignacio, Las Barrancas y Cabo San Lázaro). Se estimó el grado de madurez de las gónadas mediante observación directa, utilizando la escala propuesta en Nikolsky (1963). Se realizó el cálculo del índice gonadosomático (IG) en términos porcentuales (� = ��� ∗ ), se colectaron gónadas en formaldehído al 10% para su posterior análisis en laboratorio. El análisis histológico se realizó por el Centro Inetrdiciplinario de Ciencias Marinas (CICIMAR), en el Departamento de Pesquerías y Biología Marina. Resultados Se colectaron un total de 302 muestras biométricas/biológicas de verdillo, de los cuales 125 fueron machos, 174 hembras y 3 indiferenciados; lo que da una proporción sexual total de 1.2H:1M. Se generó un gráfico con los promedios mensuales obtenidos de la escala Nikolsky (1963). Los valores más altos fueron para los meses de junio, julio, agosto y septiembre (marcados en la figura 1 de color naranja).
Fig. 1. Promedio del estadio Nikolsky de manera mensual (línea verde: no diferenciado sexo; línea azul: machos; línea roja: hembras). Tabla 1. Proporción de sexos mensual y total de P. nebulifer en la costa occidental de B.C.S. El asterisco* indica diferencia significativa con X2=3.841, P<0.05 Mes Hembra
(%) Macho
(%) H:M X2 Valor
P Feb (2013) 23 (64.5) 11 (35.5) 1.8:1 8.4 0.004*
Mar 18 (62.1) 11 (37.9) 1.6:1 5.8 0.016*
Abr 8 (25.0) 24 (75.0) 0.3:3 25.0 0.000*
May 13 (61.9) 8 (38.1) 1.6:1 5.8 0.017*
Jul 11 (32.4) 23 (67.6) 0.5:2 12.5 0.000*
Ago 17 (54.8) 14 (45.2) 1.2:1 0.94 0.331
Sep 13 (48.1) 14 (51.9) 1.0:1 0.15 0.701
Oct 18 (60.0) 12 (40.0) 1.5:1 4.0 0.045*
Nov 17 (56.7) 13 (43.3) 1.3:1 1.8 0.181
Dic 17 (58.6) 12 (41.4) 1.4:1 3.0 0.084
Ene (2014) 26 (41.1) 33 (58.9) 0.7:1 2.0 0.174
Total 181 (50.8)
175 (49.2)
1.2:1 3.0 1.56
El índice gonadosomático coincide con el ciclo reproductivo de la especie, por lo que es un buen indicador de su actividad reproductiva.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
158
Fig. 2. Variación mensual de los valores medios del Índice Gonadosomático. Conclusiones La escala Nikolsky es un buen indicador para obtener información preliminar del ciclo reproductivo, y al compararlo con las estimaciones del IG se encontró coincidencia con las temporadas reproductivas. La actividad reproductiva de P. nebulifer es estacional y se presenta de mayo a septiembre, con un pico máximo en agosto. Literatura citada Nikolsky GV. 1963. The ecology of fishes. Academic
press. 352pp. Palabras clave: verdillo, índice gonadosomático, ciclo reproductivo,

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
159
076: oral EVALUACIÓN PESQUERA DE Oreochromis aureus EN LA PRESA EL GALLO,
GUERRERO-MICHOACÁN, MÉXICO
Claudio Osuna Paredes, Ezequiel Arredondo Vargas, Yesenia Fuerte Velázquez, Atzimba Pérez Juárez, Claudia de Jesús Avendaño y Berenice Vital Rodríguez
Centro Regional de Investigación Pesquera de Pátzcuaro. INAPESCA. Calz. Ibarra No. 28, Pátzcuaro, Michoacán, México. C.P. 61609. Tel: 01(434)3421184; [email protected], [email protected], [email protected], [email protected] [email protected] Introducción La pesca continental difiere de las demás pesquerías por su alto grado de interrelación con otros usuarios del recurso acuático. La mayor parte de las aguas continentales son controladas por otros intereses diferentes a los de la pesca, como: la generación de energía eléctrica, la navegación, el saneamiento de marismas para la agricultura, la urbanización, la extracción de agua y la evacuación de residuos. Esto implica que la mayoría de las políticas para asignar los recursos acuáticos y su contexto físico están sujetos a decisiones que se toman fuera de la pesca. Por ello, la ordenación pesquera debe realizarse en medio de las limitaciones impuestas por estos sectores externos, observando las técnicas aplicables para reducir o contrarrestar los efectos provenientes del exterior. El objetivo del presente estudio es proporcionar las bases técnicas sobre la dinámica poblacional de los principales recursos pesqueros explotados en la presa El Gallo. Material y métodos El material biológico utilizado procedió de 8 muestreos mensuales de la captura comercial de tilapia realizados entre abril y diciembre 2015. Cada muestreo consideró un promedio de 179 organismos, que se midieron: la longitud patrón (Lp), longitud total (Lt) y pesaron: peso total (Pt). Se anotó el sexo y el estado de madurez gonádica. Se determinó la estructura de la población y definieron los principales parámetros poblacionales como crecimiento, por el método indirecto ELEFAN I (Electronic Length Frecuency Analysis), contenido en el programa FISAT (Gayanilo et al. 1996), mortalidad total (por la curva de captura a edades relativas, Gayanilo et al. 1996), la natural a partir de la ecuación empírica de Pauly (1983) y por pesca. Se estimó el rendimiento por recluta empleando el modelo de Beverton y Holt (1957 en Pauly 1983). Resultados La composición de la captura de tilapia se encuentra sesgada por la selectividad de las redes de enmalle que se utilizan, 97% de la captura se basa en las longitudes de 18.0 a 23.0 cm Lt, pesos entre 99.9 y 407.3 g. En la muestra, la tilapia alcanzó una longitud
máxima de 38.4 cm Lt, y peso de 348.06 g. Los valores mínimos fueron de 17.4 cm Lt, 243 g Pt. La proporción de sexos fue: 70.01% machos y 29.98% hembras, es decir, 2 machos por hembra anualmente (Tabla 1). La relación longitud-peso fue Pt = 0.1603 Lt2.4905. Tabla 1. Valores morfométricos de tilapia.
Longitud promedio
(cm)
Peso (g)
Lt-Pt H:M N
a b
25.76 282.15
0.1603 2.4905 2.3:1 1,614 Mín 17.4 243
Máx 38.5 348.06
d.s. 1.90 47.19
La población de tilapia presenta una relación alométrica negativa. Se analizó el grado de madurez, y su comportamiento en el tiempo, se encontró que se reproduce todo el año. La etapa II se muestra durante todos los meses pero existen dos picos, uno en septiembre y otro en diciembre que muestran mayor incidencia en la fase de desarrollo gonádico. La longitud de primera captura es de 24.66 cm Lt según la frecuencia acumulada. Los parámetros de crecimiento se calcularon k = 0.44 año-1 y longitud máxima L∞ = 39 cm Lt. La mortalidad natural M = 0.98, la total Z = 2.36 (Fig. 1) y por pesca F = 1.90. Por lo que la tasa de explotación es E = 0.59, que indica que el recurso rebasa los límites de su nivel.
Fig. 1. Curva de captura a edades relativas de Oreochromis spp.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
160
De acuerdo al modelo de rendimiento (B-H), la pesquería de tilapia registró un rendimiento por recluta de r/r = 166.31 g, con una Fact = 1.90 y una edad de primera captura de 1.93 años (24.7 cm) (Fig. 2). Por otra parte, se calculó el rendimiento por recluta variando la edad de primera captura (Fig. 3), al aplicar el modelo indica que la edad óptima de captura es de 2.5 años (27.9 cm Lt), porque a esta edad el rendimiento puede aumentar hasta 222.64 g por recluta, medida que puede mejorar el rendimiento económico de los usuarios
Fig. 2. Rendimiento por recluta de Oreochromis spp. Discusión y conclusiones La captura de tilapia (Oreochromis aureus) se basa en longitudes de 18.0 a 23.0 cm Lt, el promedio de los organismos capturados fue de 25.76 cm Lt, Arredondo-Vargas et al. (2014) reportaron una longitud promedio de 28.88 cm Lt para la presa El Cajón en Nayarit y Arredondo-Vargas et al. (2014) obtuvieron una longitud promedio de 30.88 cm Lt en la Presa Aguamilpa, ambas para la pesquería de O. aureus. Al aplicar el modelo de crecimiento se obtuvo una L = 39 cm Lt, inferior a la obtenida por Arredondo-Vargas et al. (2005) en este mismo embalse y que fue de 39.84 cm Lp para esta misma especie. La tasa de crecimiento de k = 0.44 fue superior a la obtenida en 2004 para este mismo embalse por Arredondo-Vargas et al. (2005). El análisis revela que la tilapia se reproduce prácticamente todo el año, con picos máximos en junio y julio. La tasa de mortalidad natural M = 0.97 año-1 y la mortalidad por pesca F = 1.39 año-1, por lo que se tiene una mortalidad de Z = 2.36 año-1. Estos valores de mortalidad se reflejan en la tasa de explotación que en la presa fue de E = 0.59, lo que indica que el recurso rebasa el límite de explotación óptimo. Para el recurso tilapia se obtuvo un rendimiento por recluta de r/r = 166.31 g, para aumentar el rendimiento a 222.64 g/r se debe incrementar la edad de primera captura a 2.5 años, que sería la edad óptima de captura.
Literatura citada Arredondo-Vargas E, C Osuna-Paredes, N Hernández-
Zárate, D Hernández-Montaño. 2005. Evaluación limnológica y biológico-pesquera de la Presa El Gallo, Municipio de Cutzamala de Pinzón, Guerrero, México. Informe de Investigación (documento interno), Instituto Nacional de Pesca, Centro Regional de Investigación Pesquera-Pátzcuaro. 35 p. Inédito.
Arredondo-Vargas E, C Osuna-Paredes, C Meléndez-Galicia, C De Jesús-Avendaño, ML Bravo-Olivas, D Hernández-Montaño, MG Juárez-Avilés, CA Calderón-Campos. 2014. Estado actual de las pesquerías de la Presa Leonardo Rodríguez Alcaine (El Cajón), Nayarit, México. Informe de Investigación (documento interno), Instituto Nacional de Pesca, Centro Regional de Investigación Pesquera-Pátzcuaro. 69 p. Inédito.
Gayanilo F.C., P. Sparre, D. Pauly. 1996. The FAO-ICLARM Stock assessment Tools (FISAT) User´s Guide. FAO Computarized Information Series (Fisheries). No. 8 Rome, FAO 126 p.
Pauly, D. 1983. Some simple methods for the assessment of tropical fish stock. FAO Fish Technical Paper (234):52 pp.
Palabras clave: tilapia, Oreochromis spp., Presa El Gallo, crecimiento, tasa de explotación, rendimiento

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
161
077: oral ANÁLISIS DE LA PESQUERÍA Oreochromis aureus DE LA PRESA LA YESCA,
JALISCO-NAYARIT, MÉXICO
Ezequiel Arredondo Vargas, Claudio Osuna Paredes, Atzimba Pérez Juárez, Yesenia Fuerte Velázquez, Claudia De Jesús Avendaño y Berenice Vital Rodríguez
Centro Regional de Investigación Pesquera de Pátzcuaro. INAPESCA. Calz. Ibarra No. 28, Pátzcuaro, Michoacán, México. C.P. 61609. Tel: 01(434)3421184; [email protected], [email protected], [email protected], [email protected] [email protected] Introducción La pesca en los diversos embalses del país constituye un recurso importante para las comunidades, debido a que genera fuentes de alimento, empleo y desarrollo. Ante este panorama, el INAPESCA ha procurado la evaluación biológico-pesquera en diversos cuerpos de agua con la finalidad de proponer medidas de manejo de los recursos. El embalse objeto de estudio es de reciente creación y presenta potencial para el desarrollo de la actividad pesquera, pero podría enfrentar problemáticas particulares como modificación de hábitat, fallas en la cadena productiva y conflictos sociales en el corto o mediano plazo, constituyendo amenazas para la sostenibilidad. Por ello, es necesario conducir estudios en diversos ámbitos como el limnológico, el socioeconómico y el desarrollo social para caracterizar de manera integral el aprovechamiento de la Presa La Yesca y poder por un lado, proveer indicadores de desarrollo y vulnerabilidad del sistema y, por otra parte, medidas de regulación pesquera en beneficio de los principales usuarios. Material y métodos Se llevaron a cabo 9 muestreos mensuales de la captura comercial de tilapia entre abril y diciembre de 2015, con un promedio de 231 organismos, de los cuales se les registraron los principales datos morfométricos: longitud patrón (Lp), longitud total (Lt) y peso total (Pt), sexo y estado de madurez gonádica. Se determinó la estructura de la población, parámetros de crecimiento, por el método indirecto ELEFAN I (Electronic Length Frecuency Analysis,), contenido en el programa FISAT (Gayanilo et al. 1996), mortalidad total (Z), estimada por la curva de captura a edades relativas (Gayanilo et al. 1996), natural (M), a partir de la ecuación empírica de Pauly (1983) y por pesca (F). Se estimó el rendimiento por recluta (r/r) de Beverton y Holt (1957, en Pauly 1983). Resultados El 97.59% de la captura se basa en las longitudes de 23.0 a 29.0 cm Lt, y 167.6 a 464.7 g Pt. En la muestra, la tilapia alcanzó una longitud máxima de 33.0 cm Lt, con un peso de 667.6 g. Mientras que los valores mínimos que se registraron fueron de 20.5
cm Lt, con un peso de 167.6 g. La relación Macho-Hembra para esta especie es de nueve machos por cada hembra, aproximadamente (Tabla 1). El modelo de la relación longitud-peso fue Pt = 0.1031 Lt2.4682. Tabla 1.- Valores morfométricos de tilapia.
Longitud total (cm) Peso (g)
Lt-Pt M:H N
a b
25.81 316.73
0.1031 2.4682 9.3:1 2080 Min 20.5 167.6
Max 33.0 667.6
d.s. 1.237 51.28
La población de tilapia presenta una relación alométrica negativa. Se analizó el grado de madurez, y su comportamiento en el tiempo, se encontró que la tilapia se reproduce prácticamente todo el año, pero en muy baja proporción en la muestra analizada. La etapa II se muestra durante todos los meses pero existen tres picos, uno en mayo, otro en julio-agosto y, finalmente, otro en octubre-noviembre. El 50% de la frecuencia acumulada muestra que la longitud de primera captura es de 25.5 cm Lt. Para este ciclo de muestreo se calculó un parámetro de curvatura de crecimiento k = 0.45 año-1 alcanzando una longitud máxima L∞ = 39.30 cm Lt. Se presentó una mortalidad natural M = 0.98, una mortalidad total Z = 3.69 (Fig. 1) y una mortalidad por pesca F = 2.71. Por lo que la tasa de explotación es E = 0.73, lo que indica que el recurso rebasa el límite de su nivel óptimo de explotación.
Fig. 1. Curva de captura a edades absolutas de Oreochromis spp.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
162
La figura 2 muestra que con la mortalidad por pesca actual F = 2.71 se obtiene un rendimiento de 223.8 g/r con una edad de primera captura de 1.84 años. Por otra parte, se calculó el rendimiento por recluta variando la edad de primera captura, al aplicar el modelo indica que la edad óptima de captura es 2.5 años, a esta edad de primera captura el rendimiento puede aumentar hasta 318.93 gramos por recluta, medida que puede mejorar el rendimiento económico de los usuarios.
Fig. 2. Rendimiento por recluta de Oreochromis spp. Discusión y conclusiones La composición de la captura de tilapia fue de 20.5 a 33.0 cm Lt, con un promedio de 28.88 cm Lt, longitudes similares también son reportadas por Silva (2009) para la Presa El Cajón (25.0 a 42.0 cm Lt). Arredondo-Vargas et al. (2013) obtuvieron una longitud promedio de 30.88 cm Lt para la pesquería de Oreochromis aureus en la Presa Aguamilpa, lo que indica que las longitudes están condicionadas por el tamaño de malla utilizado. La tasa de crecimiento fue k = 0.45 años-1, ligeramente inferior a la reportada por Arredondo-Vargas et al. (2014) para la presa El Cajón de 0.51. Esta tasa de crecimiento es acorde a la obtenida en otros estudios para Oreochromis aureus y ligeramente inferior a las reportadas por Silva-García (2009) y Arredondo-Vargas et al. (2014) para la presa El Cajón de 0.47 y 0.51. Tasas de crecimiento dentro de este intervalo son características de esta especie. Se analizó el grado de madurez, y su comportamiento en el tiempo, se encontró que la tilapia se reproduce prácticamente todo el año, pero a muy bajo proporción con un periodo máximo en el mes de abril. La tasa de mortalidad natural M = 0.98 año-1 fue mucho más baja que la mortalidad por pesca F = 2.71 año-1, estos valores de mortalidad se reflejan en la tasa de explotación que en la presa La Yesca fue de E = 0.73, que indica que el recurso ya rebasó el límite de explotación óptimo.
Para el recurso tilapia, se obtuvo un rendimiento por recluta de 223.8 g; para aumentar el rendimiento a 318.93 g/r se debe incrementar la edad de primera captura a 2.5 años, que sería la edad óptima de captura. Literatura citada Arredondo-Vargas E, C Osuna-Paredes, C Meléndez-
Galicia, C De Jesús-Avendaño, F Cerna-Aguirre, IE Serrato-Mireles, D Hernández-Montaño. 2013. Estado actual de las pesquerías de la Presa Aguamilpa, Nayarit, México. Informe de Investigación (documento interno), Instituto Nacional de Pesca, Centro Regional de Investigación Pesquera-Pátzcuaro. 58 p. Inédito.
Arredondo-Vargas E, C Osuna-Paredes, C Meléndez-Galicia, C De Jesús-Avendaño, ML Bravo-Olivas, D Hernández-Montaño, MG Juárez-Avilés, CA Calderón-Campos. 2014. Estado actual de las pesquerías de la Presa Leonardo Rodríguez Alcaine (El Cajón), Nayarit, México. Informe de Investigación (documento interno), Instituto Nacional de Pesca, Centro Regional de Investigación Pesquera-Pátzcuaro. 69 p. Inédito.
Gayanilo FC, P Sparre, D Pauly. 1996. The FAO-ICLARM Stock assessment Tools (FISAT) User´s Guide. FAO Computarized Information Series (Fisheries). No. 8 Rome, FAO 126 p.
Pauly D. 1983. Some simple methods for the assessment of tropical fish stock. FAO Fish Technical Paper (234):52 pp.
Silva-García RE. 2009. Evaluación biológica-pesquera de la tilapia Oreochromis aureus en el embalse de la Presa Hidroeléctrica El Cajón, Nayarit. Tesis para obtener el grado de Ingeniería Pesquera. Escuela Nacional de Ingeniería Pesquera. Universidad Autónoma de Nayarit. 46 p.
Palabras clave: tilapia, Oreochromis spp., Presa La Yesca, crecimiento, tasa de explotación, rendimiento

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
163
078: oral MADUREZ GONÁDICA DEL OSTIÓN DE ROCA Crassostrea iridescens,
DE LA COSTA DE MICHOACÁN, MÉXICO
Carlos Meléndez Galicia, Flor D. Estrada Navarrete, Vicente Hernández Covarrubias, Andrés A. Arellano Torres y Daniel Hernández Montaño.
Centro Regional de Investigación Pesquera de Pátzcuaro. INAPESCA. Ibarra 28, Col. Ibarra, Pátzcuaro, Michoacán. Tel. 01(434)3420087. [email protected]. Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Calzada Sábalo Cerritos S/N, Col. Estero del Yugo, Mazatlán, Sinaloa. C. P. 82010. Tel/Fax: 01(669)9880049. [email protected] Introducción En el Pacífico mexicano es posible encontrar tres especies de ostras silvestres: el ostión de roca Crassostrea iridescens (Hanley 1854), el ostión de placer C. corteziensis (Hertlein 1951) y el ostión de mangle C. palmula (Keen 1971) (Páez-Osuna et al. 1995). En México, este recurso ha soportado volúmenes elevados de explotación en varios puntos de la costa del Pacífico y actualmente se han obtenido cifras mayores a 42 945 t de ostión (captura), con un promedio de producción de 48 138 t al año (SAGARPA 2013). De la producción nacional, 13.8% proviene de las costas del Pacífico mexicano. El ciclo reproductivo en moluscos bivalvos es uno de los temas de estudio más importantes en la actualidad para el desarrollo de prácticas de manejo tendientes a ser sustentables (Cuevas-Guevara y Martínez-Guerrero 1978, Clédon et al. 2004). Los bivalvos bentónicos marinos presentan un patrón cíclico de reproducción que puede ser dividido en tres fases: a) gametogénesis y vitelogénesis; b) desove y fertilización y c) desarrollo larvario y crecimiento (Newell et al. 1982). Los principales factores que influyen en su ciclo reproductivo son: temperatura, profundidad, salinidad, alimento disponible y contaminación (Galtsoff 1964). Cabe mencionar que los estudios de reproducción son esenciales debido a que proporcionan datos relacionados con la distribución y la estructura de las poblaciones, permite efectuar predicciones acerca del reclutamiento y proporcionan la información necesaria para el manejo de las pesquerías. Material y métodología El litoral michoacano posee una plataforma continental muy estrecha y talud continental con pendiente muy pronunciada (Guzmán et al. 1985). La extensión litoral es de 246 km y su plataforma continental abarca 1 372 km2 (SEPESCA 1994). El tipo de clima es Aw (w), cálido subhúmedo, con lluvias en verano y precipitación invernal menor a 5%. La temperatura media anual es de 26°C a 28°C. La época de lluvias abarca los meses de junio a octubre. La precipitación media anual es de 894.8 mm (García 1973).
Los lugares de muestreo se seleccionaron de acuerdo con la capacidad de captura del ostión y la abundancia de este recurso; así, los más importantes son el puerto de Lázaro Cárdenas y la comunidad del Zapote de Huahua, Michoacán, ubicados entre 102°12’35’’ O y 17°57’02’’ N. Los muestreos se realizaron mensualmente de febrero a diciembre de 2010. Las muestras se obtuvieron de manera aleatoria mediante la técnica de buceo a pulmón y semiautónomo y en cada sitio se consideraron diez organismos por mes; in situ se registró la longitud total (lt), el ancho (a) y el peso total (p) de cada organismo. Los ostiones se lavaron y se desconcharon para obtener los tejidos blandos, se realizaron cortes entre el músculo aductor y el umbo, área ocupada por la gónada y la glándula digestiva. Después del desconche, los ostiones se fijaron en solución de formol buferado a 10% y se depositaron en frascos de 250 ml con el fin de preservar convenientemente los tejidos por 48 horas. El análisis del tejido se realizó con la técnica histológica de Hematoxilina-Eosina de Humason (1979), Luna (1968) y Howard y Smith (1983). Las muestras procesadas fueron analizadas cualitativamente en un microscopio compuesto, con objetivos de 10x y 40x donde se determinó la predominancia de los estadios de maduración, así como la respectiva abundancia de los diferentes estadios y el grado de madurez gonádica del organismo, con respecto a cada mes de colecta. Para la obtención de las imágenes se utilizó una cámara (Infinity 1 digital). Resultados y discusión La variación mensual de la relación peso de cuerpo blando de la gónada entre el peso total de la concha varió durante el proceso de muestreo, observándose el mayor porcentaje en mayo con 18.1%, mientras que la menor en marzo con 4.6%. La mayor variación fue en agosto con 13.4% y la menor en febrero de 6.3% (Fig. 1). La media varió de 6.3 a 13.4% durante el muestreo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
164
Fig. 1. Valores mínimos, máximos y media mensual de la relación peso blando de la gónada en relación al peso total de la concha de Crassostrea iridescens en la costa de Michoacán. El análisis de desarrollo gonádico por cortes histológicos se realizó en 100 organismos: 31 eran machos, 24 hembras y 2 hermafroditas, además de 43 individuos indiferenciados.
Fig. 2. Porcentaje de las fases de madurez gonádica del ostión de roca Crassostrea iridescens de la costa de Michoacán. El desarrollo gonádico porcentual e individual del ostión de roca C. iridescens durante el periodo de muestreo febrero a diciembre de 2010, permitió reconocer cinco fases diferenciables en los que se encontraban los organismos en el momento de su captura (Fig. 2). El ciclo reproductivo en muchos bivalvos está definido por el conjunto de eventos que inicia con la activación de la gónada y pasa por la gametogénesis, la madurez, el desove y la recesión de la gónada (García-Domínguez 2002). De manera que el análisis de las gónadas en estos organismos es básico para comprender la biología reproductiva de C. iridescens y de otras especies de bivalvos (Gosling 2003), especies con un excelente potencial acuícola (DOF 2012). Del análisis cuantitativo como cualitativo de los cortes histológicos de la gónada del ostión de roca C. iridescens indican que la especie tiene un período de reproducción corta, se observó la liberación de
gametos, en los meses de septiembre y octubre principalmente, es importante tener muestras durante los meses de verano, período en el que se pueden presentar desoves parciales los cuales coincidan con la presencia de altas temperaturas. Literatura citada Cuevas-Guevara CA, A Martínez-Guerrero. 1978. Estudio
gonádico de Crassostrea corteziensis Hertlein, C. palmula Carpenter y C. iridescens Hanley de San Blas, Nayarit, México. An. Inst. Cienc. Del Mar y Limnol. UNAM 6: 81-98.
Palabras clave: ostión de roca Crassostrea iridescens, reproducción, cortes histológicos, madurez gonádica, tinción, Michoacán.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
165
079: oral EFECTO DE LA PROFUNDIDAD EN LAS ESPECIES ASOCIADAS AL PALANGRE TIBURONERO EN LA
PESQUERÍA ARTESANAL DE TIBURÓN EN LA COSTA CENTRAL DE VERACRUZ, MÉXICO
Leticia González Ocaranza1, Jorge Luis Oviedo Pérez2. Heber Zea de la Cruz2, Luis Enrique Martínez Cruz3 y Pedro Reyna González2
1Universidad Veracruzana. Posgrado Ecología y Pesquerías. Hidalgo 617, Boca del Río, Veracruz.. C. P. 94290. 2Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. 3Centro Regional de Investigación Pesquera de Lerma. INAPESCA. [email protected] Introducción La pesca ribereña se define como la actividad de extracción de recursos acuáticos ejecutada con embarcaciones menores, que no cuentan con maquinaria de cubierta accionada con fuerza electromotriz para el auxilio de las operaciones de pesca, y utilizan el hielo para la conservación del producto con una autonomía que les permite operar por tres o cinco días. Los organismos capturados por este tipo de pesca se dividen en dos grupos, al primero se les denomina especies objetivo, las cuales se definen por su valor económico en el mercado, logrando su captura por temporadas o a lo largo del año; al segundo, se les denomina especies asociadas, las cuales son aquellas que comparten el hábitat y pertenecen a la misma comunidad o ensamble, formando un grupo funcional, lo que las hace susceptibles al mismo arte de pesca, en la mayoría de los casos también son aprovechadas comercialmente y pueden representar un recurso pesquero potencial (DOF 2012). En México, el sector de ribera representa poco más de 80% del total de pescadores del país y contribuye con 65% de la producción para consumo humano directo. La producción nacional obtenida por pesca ribereña en el 2013 fue de 69,162 toneladas. Lo que representa una derrama económica de $1,172,266.00 (CONAPESCA 2013). El objetivo del presente trabajo es determinar las principales especies de teleosteos capturadas por el palangre tiburonero y la relación de la composición de la captura por estrato de operación en la flota artesanal en la zona central del estado de Veracruz, México. Materiales y métodos Los sitios de pesca se localizan en el suroeste del Golfo de México, en el área de influencia del Sistema Arrecifal Veracruzano y de las desembocaduras del río Antigua Datos biológico-pesqueros. Se analizaron datos biológico-pesqueros provenientes del Proyecto Tiburón, CRIP Veracruz del Instituto Nacional de Pesca (INAPESCA), recabados en muestreos aleatorios mensuales desarrollados en el Municipio de Alvarado, Veracruz entre julio de 2001 y diciembre de 2010. Los muestreos en cuestión cubrieron aproximadamente
50% de la flota pesquera dedicada a la captura de tiburón con palangre en dicha localidad (Oviedo et al. 2012). Los datos analizados incluyeron información de fecha de muestreo, localidad de muestreo, zona de captura, arte de pesca utilizado, profundidad y forma de operación del arte de pesca, número de anzuelos utilizados en el viaje de pesca, tamaño de anzuelo, tipo de carnada, especies capturadas en cada viaje, longitud y estado de madurez. Análisis de datos. De la base de datos construida, se separaron los registros de la captura objetivo (tiburones y rayas) y posteriormente se llevó a cabo un análisis exploratorio de los datos, de las especies de escama, con el fin de conocer cuales especies aparecían con más frecuencia asociadas a la captura de especies de elasmobranquios (Nelson 2006). Resultados y discusión Composición específica de la captura. Se registró una captura de 47,320 organismos, compuesta por 66.97% de tiburones, 6.39% de rayas y 26.64% de peces óseos. Las especies más importantes en número de organismos y valor comercial fueron Scomberomorus cavalla 18.39%, Caranx latus 17.25%, C. hippos 6.04%, Seriola fasciata 5.90%, Ocyurus chrysurus 5.47%, Lutjanus cyanopterus 3.67, L. analis 3.13%, Scomberomorus maculatus 2.62%, Rachycentron canadum 2.50%, Mycteroperca phenax 2.16%. En total estas 10 especies representan 67.13% de los peces óseos asociados a la pesca de tiburones con palangre. Operación del palangre tiburonero. Se registraron variantes en la forma de operación del palangre, que pueden ser a superficie, media agua y fondo. Estas formas de operación presentaron variantes combinadas como: a media agua–fondo, a superficie-fondo y a superficie-media agua. Se registró un esfuerzo pesquero total de 1’152,096 anzuelos, agrupados en seis diferentes formas de operación (Fig. 1).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
166
Fig. 1. Esfuerzo pesquero por estratos de operación del palangre tiburonero de la flota artesanal tiburonera en Antón Lizardo, Veracruz.
Fig. 2. Abundancia relativa de las familias de peces óseos capturados por el palangre en Antón Lizardo, Veracruz.
Composición de la captura. Para describir la composición de la captura del palangre por estrato de operación, se utilizaron solamente las variantes a superficie, media agua y a fondo, que en su conjunto equivalen a 66.2% del esfuerzo pesquero aplicado. No se incluyeron las variantes combinadas de operación del palangre pues la captura no puede ser relacionada en una sola forma de operación. La captura total fue de 33,175 organismos, conformada por 74.7% de tiburones, 6.7% de rayas y 18.6% de peces óseos (Fig. 2). La composición de la captura cambia si el arte de pesca opera a fondo, media agua o superficie. La captura total del palangre a fondo fue de 10,109 organismos con 53.4% de tiburones, 19.3% de rayas y 27.3% de peces óseos. Operado a media agua la captura fue de 2,054 organismos, con 74.9% de tiburones, 2.1% de rayas y 22.9% de peces óseos. Utilizado a superficie la captura fue de 21,402 organismos, con 85.1% de tiburones, 1.0% de rayas y 13.9% de peces óseos (Fig. 3). La captura de tiburones presentó mayor porcentaje cuando fue operado a superficie, las rayas tienen mayor porcentaje de captura cuando el palangre es operado a fondo.
Fig. 3. Composición porcentual de la captura de tiburones, rayas y peces óseos del palangre con respecto a su estrato de operación en Antón Lizardo, Veracruz.
Conclusiones El jurel blanco se capturó en todos los estratos de operación; a fondo se registraron especies demersales como cubera caballera y pargo habanero; a media agua y superficie se registraron especies pelágicas como peto, sierra, huachinango. No existe registro en las estadísticas oficiales del esfuerzo que se ejerce en las especies cuando son consideradas especies asociada o incidentales. Es necesario cuantificar este esfuerzo para incluirlo en las evaluaciones de las pesquerías de escama. Literatura citada CONAPESCA. 2013. Anuario estadístico de acuacultura y
pesca. SAGARPA. DOF. 2012. Carta Nacional Pesquera. Diario Oficial de la
Federación Oviedo-Pérez JL, H Zea de la Cruz, LE Martínez-Cruz.
2012. Caracterización de la pesquería de tiburones y rayas en el estado de Veracruz. Informe Técnico. SAGARPA –INAPESCA-CRIP-Veracruz.
Nelson JS. 2006. Fishes of the world. 4th. Ed. Edit. John Wiley and sons. Inc. Hoboken, New Jersey, U.S.A. 2006.
Palabras clave: Teleósteos, Veracruz, palangre, profundidad, pesca ribereña
FONDO 28.8%
MEDIA AGUA 5.3%
SUPERFICIE 32.1%
MEDIA AGUA, FONDO 27.4% SUPERFICIE,
FONDO 1.5%
SUPERFICIE, MEDIA AGUA
0.3%
SIN REGISTRO 4.6%
Carangidae 28%
Lutjanidae 24%
Scombridae 26%
Rachycentridae 10%
Serranidae 6% Sphyraenidae
2% Megalopidae
2%
Tetraodontidae 0.1%
Balistidae 0.2%
Coriphaenidae 1%
Istiophoridae 0.1%
Ariidae 1%
0
20
40
60
80
100
FONDO MEDIA AGUA SUPERFICIE
% C
ap
tura
Estrato de operación del palangre
Peces óseos Rayas Tiburones

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
167
080: oral GENERACIÓN DE INFORMACIÓN Y MONITOREO PESQUERO DE ESCAMA: CASO DEL CORREDOR SAN
COSME – PUNTA COYOTE, BCS, MÉXICO
Tomás Plomozo Lugo y Amy Hudson Weaver
Sociedad de Historia Natural Niparajá A. C., Revolución de 1910 / Vicente Gro. y H. Colegio Militar, Col. El Esterito, La Paz, Baja California Sur, México. C. P. 23020. Tel: 01(612)1221171 (Ext) 106; [email protected]; [email protected] Introducción Actualmente en México la pesca ribereña necesita de información para actualizar su status y potencial, específicamente en los recursos de escama, los cuales han estado generalizados y se manejan como uno solo. La información que se utiliza para el manejo de los recursos muchas veces es limitada, lo cual dificulta una mejor seguridad en la toma de decisiones. El Corredor costero San Cosme-Punta Coyote se localiza dentro del Golfo de California en la parte sur-oriental de la península de Baja California Sur, comprende alrededor de 150 km de línea de costa, en donde se consideran existen 11 comunidades consolidadas en la costa y 2 pequeños asentamientos en las islas (La Palma Sola en Isla San José y El Pardito). La población oscila alrededor de 600 personas, en donde la fuente principal de empleo es la actividad pesquera y menos de 15% de los habitantes se dedica a la ganadería. Las características de lejanía, poca densidad poblacional y esfuerzo limitado de flota local, hacen del Corredor una zona productiva en cuanto a la pesca de escama y por ende una oportunidad para estudiarlo (Fig. 1). Material y métodos Desde 2010 la Sociedad de Historia Natural Niparajá y Pronatura Noroeste realizaron la capacitación de un grupo de pescadores como técnicos pesqueros para la generación de información pesquera del área. En esta capacitación se generaron los protocolos de monitoreo para la descripción y conocimiento de la pesca (línea base). Posteriormente en 2013, se establecieron criterios de selección de especies de importancia comercial, número de organismos que se necesitan medir (información de pesquerías con pocos datos) y la adecuación de la metodología en campo. En 2015 el grupo de técnicos pesqueros recibió la capacitación en técnicas de comunicación, para poder expresar y transmitir la información que están generando en sus comunidades, así como la entrega de los resultados de un año y medio, las conclusiones que el mismo grupo puede dialogar y cuestionar.
Fig. 1 Corredor costero San Cosme-Punta Coyote. Resultados y discusión Se capacitaron 37 pescadores, hijos o esposas de pescadores como técnicos pesqueros en la zona del Corredor San Cosme-Punta Coyote, para obtener información fidedigna y de primera mano de la actividad pesquera de escama en la región. De acuerdo al monitoreo pesquero se encontró que el huachinango, cadernal, pierna, jurel y cabrilla sardinera son las especies más representativas en las comunidades pesqueras. La información de la producción recabada en las bitácoras pesqueras representa 50% existente en el Corredor de San Cosme-Punta Coyote. Sólo cuatro especies de las 13 más importantes presentan capturas por debajo de su longitud de primera madurez sexual, en el caso de huachinango

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
168
y pierna, estos presentan las capturas justo en el límite de longitud de primera madurez sexual de la especie. La información que se genera en el proyecto está limitada al esfuerzo aplicado en cada monitoreo, describe y explica de alguna forma la pesca en la zona, pero no se tiene toda la información que se necesita para poder describir la pesca por completo. Conclusiones La información que se genera con los técnicos pesqueros es sumamente importante dado que ellos son los custodios de la zona, conocen, practican y viven de la pesca. La información se complementa de monitoreo pesquero y monitoreo de producción (bitácoras), una describe cómo se hace la pesca y otra como se compone y entrega el producto. Al mantener un monitoreo continuo y sistemático en la zona, da la oportunidad de observar los cambios de la pesca a través del tiempo, esto ha ocurrido en el Corredor y los pescadores han sido partícipes. Este grupo de técnicos ha motivado a un sector del Corredor a participar, actualizarse y a involucrarse en los temas de la pesca de la región. Al involucrar al pescador en los temas de estudio de la pesca, tienen la oportunidad de proponer cómo manejar sus recursos para un futuro mejor de las próximas generaciones. Literatura citada Fischer W, F Krupp, W Schneider, C Sommer, KE
Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de pesca, Pacífico centro-oriental. Vertebrados-parte 1. FAO. Roma. Vol. II: 647-1200.
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de pesca, Pacífico centro-oriental. Vertebrados-parte 2. FAO. Roma. Vol. III: 1201-1813.
Human P, N Deloach. 2004. Reef fish identification, Baja to Panama. New world publications, INC. U.S.A. 343 pp.
Froese R. 2014. Keep it simple: three indicators to deal with overfishing. Fish and Fisheries 5: 86-91.
Mateo I. 2013. Relative vulnerability of commercially important fish stock in the El Corredor región of the Gulf of California. Enviromental Defense Found. 13 pp.
Mateo I. 2013. Application of sustainibility indicators to asses to population status of six species of commercial importance in the El Corredor region of the Gulf of California. Enviromental Defense Found. 22 pp.
Weaver AH, T Plomozo-Lugo, PN Moreno-Castro, C Santa Ana. 2012. Información sobre la pesquería de escama del Corredor San Cosme-Punta Coyote, B. C. S., México. Resultados de monitoreos realizados por técnicos pesqueros comunitarios entre 2011 y 2012.
Palabras clave: Corredor San Cosme-Punta Coyote, generación de información pesquera, monitoreo pesquero, evaluaciones pesqueras.
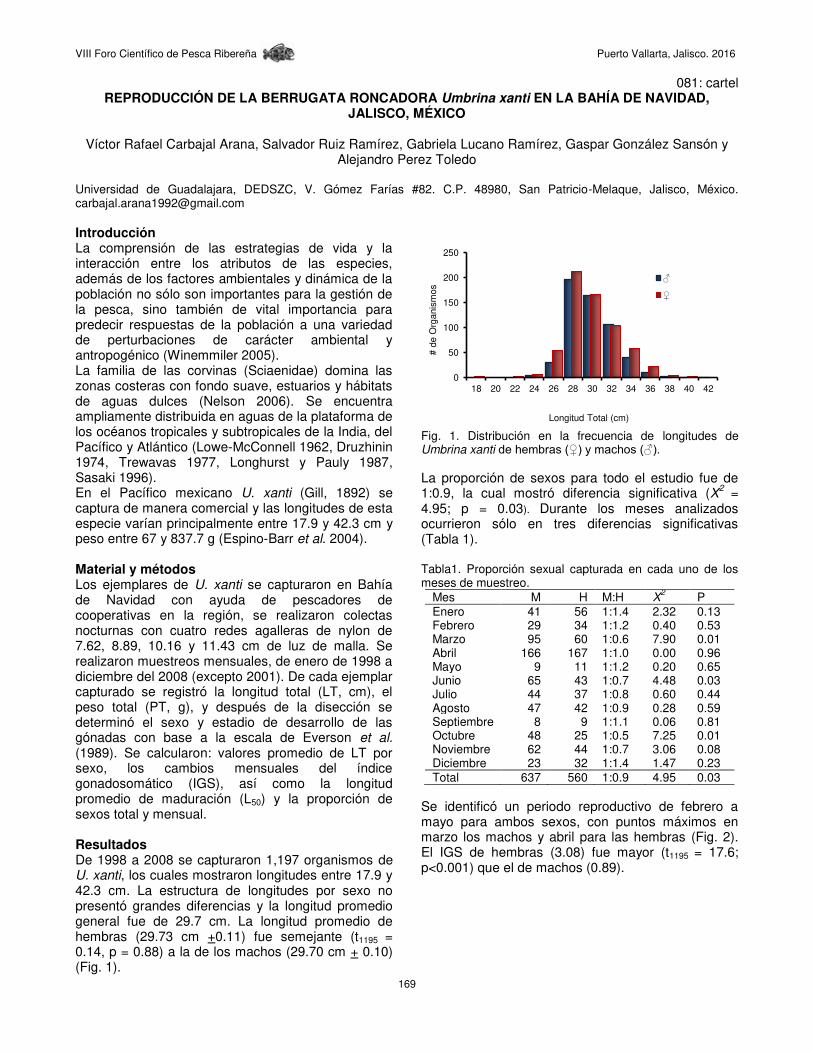
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
169
081: cartel REPRODUCCIÓN DE LA BERRUGATA RONCADORA Umbrina xanti EN LA BAHÍA DE NAVIDAD,
JALISCO, MÉXICO
Víctor Rafael Carbajal Arana, Salvador Ruiz Ramírez, Gabriela Lucano Ramírez, Gaspar González Sansón y Alejandro Perez Toledo
Universidad de Guadalajara, DEDSZC, V. Gómez Farías #82. C.P. 48980, San Patricio-Melaque, Jalisco, México. [email protected] Introducción La comprensión de las estrategias de vida y la interacción entre los atributos de las especies, además de los factores ambientales y dinámica de la población no sólo son importantes para la gestión de la pesca, sino también de vital importancia para predecir respuestas de la población a una variedad de perturbaciones de carácter ambiental y antropogénico (Winemmiler 2005). La familia de las corvinas (Sciaenidae) domina las zonas costeras con fondo suave, estuarios y hábitats de aguas dulces (Nelson 2006). Se encuentra ampliamente distribuida en aguas de la plataforma de los océanos tropicales y subtropicales de la India, del Pacífico y Atlántico (Lowe-McConnell 1962, Druzhinin 1974, Trewavas 1977, Longhurst y Pauly 1987, Sasaki 1996). En el Pacífico mexicano U. xanti (Gill, 1892) se captura de manera comercial y las longitudes de esta especie varían principalmente entre 17.9 y 42.3 cm y peso entre 67 y 837.7 g (Espino-Barr et al. 2004). Material y métodos Los ejemplares de U. xanti se capturaron en Bahía de Navidad con ayuda de pescadores de cooperativas en la región, se realizaron colectas nocturnas con cuatro redes agalleras de nylon de 7.62, 8.89, 10.16 y 11.43 cm de luz de malla. Se realizaron muestreos mensuales, de enero de 1998 a diciembre del 2008 (excepto 2001). De cada ejemplar capturado se registró la longitud total (LT, cm), el peso total (PT, g), y después de la disección se determinó el sexo y estadio de desarrollo de las gónadas con base a la escala de Everson et al. (1989). Se calcularon: valores promedio de LT por sexo, los cambios mensuales del índice gonadosomático (IGS), así como la longitud promedio de maduración (L50) y la proporción de sexos total y mensual. Resultados De 1998 a 2008 se capturaron 1,197 organismos de U. xanti, los cuales mostraron longitudes entre 17.9 y 42.3 cm. La estructura de longitudes por sexo no presentó grandes diferencias y la longitud promedio general fue de 29.7 cm. La longitud promedio de hembras (29.73 cm +0.11) fue semejante (t1195 = 0.14, p = 0.88) a la de los machos (29.70 cm + 0.10) (Fig. 1).
Fig. 1. Distribución en la frecuencia de longitudes de Umbrina xanti de hembras (♀) y machos (♂). La proporción de sexos para todo el estudio fue de 1:0.9, la cual mostró diferencia significativa (X2 = 4.95; p = 0.03). Durante los meses analizados ocurrieron sólo en tres diferencias significativas (Tabla 1). Tabla1. Proporción sexual capturada en cada uno de los meses de muestreo.
Mes M H M:H X2 P Enero 41 56 1:1.4 2.32 0.13 Febrero 29 34 1:1.2 0.40 0.53 Marzo 95 60 1:0.6 7.90 0.01 Abril 166 167 1:1.0 0.00 0.96 Mayo 9 11 1:1.2 0.20 0.65 Junio 65 43 1:0.7 4.48 0.03 Julio 44 37 1:0.8 0.60 0.44 Agosto 47 42 1:0.9 0.28 0.59 Septiembre 8 9 1:1.1 0.06 0.81 Octubre 48 25 1:0.5 7.25 0.01 Noviembre 62 44 1:0.7 3.06 0.08 Diciembre 23 32 1:1.4 1.47 0.23 Total 637 560 1:0.9 4.95 0.03
Se identificó un periodo reproductivo de febrero a mayo para ambos sexos, con puntos máximos en marzo los machos y abril para las hembras (Fig. 2). El IGS de hembras (3.08) fue mayor (t1195 = 17.6; p<0.001) que el de machos (0.89).
0
50
100
150
200
250
18 20 22 24 26 28 30 32 34 36 38 40 42
# de
Org
anis
mos
Longitud Total (cm)
♂
♀

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
170
Fig. 2. Índice gonadosomático mensual de hembras y machos de U. xanti La L50 de hembras fue de 28.03 cm y de 29.03 cm los machos (Fig. 3).
Fig. 3. Proporción de hembras y machos maduros de U. xanti. Discusión y conclusiones Este es el primer trabajo que analiza la biología reproductiva de U. xanti y es uno de los pocos generados en el Pacífico sobre las curvinas. En este estudio no se encontró diferencias importantes en la estructura de longitudes entre hembras y machos; y la proporción de sexos presentó poca variación tanto de manera total como mensual. La mayor actividad reproductiva tanto de hembras como de machos ocurrió en marzo-abril y otro menos pronunciado en noviembre-diciembre. Cárdenas (2012) menciona que la mayoría de los corvinas son especies iteróparas, con desoves parciales, la reproducción ocurre en primavera, lo cual coincide con lo observado para U. xanti. También los valores relativamente bajos del IGS en ambos sexos de U. xanti, son típicos de especies que realizan varios desoves con temporadas reproductivas relativamente más extensas. Debido al periodo reproductivo largo que presenta la especie, se puede concluir que tiene desoves parciales, además los valores del IGS de hembras son mayores al de los machos, característica que presentan varias especies de peces.
Literatura citada Cárdenas S. 2012. Acuicultura de corvinas (Pisces:
Sciaenidae) en América y en el mundo. Universidad Nacional Agraria La Molina.
Druzhinin AD. 1974. Distribución, biología y pesquerías de las corvinas. Pishevaia Promishlennost, Moscú, 119 p.
Espino-Barr E, EG Cabral-Solis, A García-Boa, M Puente-Gómez. 2004. Especies marinas con valor comercial de la costa de Jalisco, México. SAGARPA.
Everson AR, HA Williams, BM Ito. 1989. Maturation and reproduction in two hawaiian eteline snappers, uku, Aprion virescens, and onaga, etelis. Fishery Bulletin. 87(4): 877-888.
Longhurst A, D Pauly. 1987. Dynamic of tropical fish populations. Ecology of tropical oceans. Academic Press. San Diego. pp. 309-368.
Lowe-McConnell RH. 1962. The fishes of the British Guiana continental shelf, Atlantic coast of South America, with notes on their natural history. Zoological Journal of the Linnean Society 44(301): 667-700.
Nelson JS. 2006. Fishes of the world. 4th. Ed. Edit. John Wiley and sons. Inc. Hoboken, New Jersey, U.S.A. 2006.
Sasaki K. 1996. Sciaenid fishes of the Indian Ocean (Teleostei, Perciformes). Memoirs of the Faculty of Science Kyushu University 16 (17): 83-95.
Trewavas E. 1977. The sciaenid fishes (croakers or drums) of the Indo-west Pacific. Transactions of the Zoological Society of London 33: 253-541.
Winemiller KO. 2005. Life history strategies, population regulation and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences 68: 872-885.
Palabras clave: Umbrina xanti, Scianidae, pesca artesanal, longitud de madurez, índice gonadosomático, proporción sexual, costa de Jalisco.
-0.5
0.0
0.5
1.0
1.5
2.0
2.5
0.5
1.0
2.0
4.0
8.0
En Fe Mr Ab My Jn Jl Ao Sp Oc Nv Dc
IGS
Machos IG
S H
embr
as
Hembra
Macho
0
0.25
0.5
0.75
1
15 25 35 45
Pro
po
rció
n d
e M
ad
ure
z
Longitud Total (cm)
Machos Observados
Hembras Observadas
Macho
Hembra

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
171
082: oral ANÁLISIS DE LAS CAPTURAS DEL TIBURÓN AZUL Y MAKO DE LA FLOTA RIBEREÑA DE LAGUNA
MANUELA, BAJA CALIFORNIA
Carlos Javier Godínez Padilla, José Leonardo Castillo Geniz e Itzel Ortega Salgado
Centro Regional de Investigación Pesquera de Ensenada. INAPESCA. Carretera Tijuana-Ensenada Km 97.5, Parque Industrial Fondeport, El Sauzal de Rodríguez, Ensenada, Baja California. C.P. 22760. [email protected] Introducción Para determinar la estructura de longitudes-sexos y la abundancia relativa de las capturas que se obtienen de tiburones pelágicos (azul y mako) en la costa occidental de Baja California (BC), y que sostienen la pesquería ribereña de tiburón del estado, el Instituto Nacional de Pesca (INAPESCA) a través de su Centro Regional de Investigación Pesquera (CRIP) de Ensenada ha desarrollado a partir del año 2007 muestreos biologicos-pesqueros de los desembarques de la flota tiburonera de uno de los principales campamentos pesqueros (Laguna Manuela, BC), haciendo énfasis que la Bahía Sébastian Vízcaino (BSV) podría representar el área de crianza de tiburones más importante de la región del noroeste Pacífico mexicano en términos de manejo y conservación de este recurso. Los monitoreos de las especies azul y mako, principalmente capturados con palangre, contribuyen a las mayores capturas númericas de la composición de tiburones en este estudio (Castillo-Géniz et al. 2016). Material y métodos Los tiburones desembarcados por la flota de Laguna Manuela (LM) durante 2007−2015, fueron identificados a nivel de especie, se registraron las medidas morfométricas: longitud total extendida (LT), longitud furcal (LF) y la longitud precaudal (LPC). El sexado se llevó a cabo por medio de la presencia-ausencia de los órganos copuladores myxopterigios o gonopodios. El estado de madurez se realizó siguiendo una escala subjetiva descrita por Castro (1993) y cuyos estadios son: neonato, juvenil, maduro, y preñez en hembras. El índice de abundancia relativa de las especies de tiburones, fue la captura por unidad de esfuerzo: CPUE = 100 ∙ (núm. ind/núm. anzuelos) por viaje de pesca. La comparación entre los promedios de sexos y longitudes se realizó mediante pruebas t. Además se realizaron los análisis de la relación peso-longitud por sexos, su comparación mediante pruebas de ANCOVA. La proporción sexual se analizó mediante pruebas de chi cuadrada (Castillo et al. 2015). Resultados y discusión En total durante 2007−2015 se hn documentaron 955 viajes de pesca en LM, 26.5% (253) fueron viajes dirigidos a la pesca de tiburones con palangres de superficie, en los que se incluyen los 79 viajes
realizados por el personal técnico del CRIP para pescar y marcar tiburones. Los meses en donde se observaron el mayor número de viajes de pesca con palangre fueron agosto con 150 y octubre con 26 (Fig. 1). A partir de la entrada en vigor en 2012 de la veda de tiburones y rayas en el Pacífico mexicano, se redujo el esfuerzo de pesca en los meses de mayo, junio y julio.
Fig. 1. Distribución mensual de los viajes de pesca de la flota ribereña monitoreados durante 2007-2015 por equipos de pesca (redes y palangres) documentados en Laguna Manuela, BC. En el periodo de 9 años se documentó la captura de 7,214 tiburones de 19 especies. Las especies más representativas en desembarques de la flota ribereña fueron el tiburón azul Prionace glauca (29%), mako Isurus oxyrinchus (36%), la cornuda prieta Sphyrna zygaena (10%), el tiburón sulfin Galeorhinus galeus (8%), y el cazón mamón o café Mustelus henlei (5%). Estas especies de tiburones se han documentado principalmente durante el periodo de marzo a octubre, aunque el mes en el que se documentaron las mayores capturas numéricas fue agosto. En los años de capturas (2007−2013) de tiburones realizadas por las embarcaciones menores de LM, la especie dominante fue el tiburón azul, sin embargo en los años 2014 y 2015 la especies dominantes han sido el mako (1,406 ind), cornuda prieta (261 ind), y tiburón azul (141 ind) que representaron el 95.3% del total de tiburones documentados y el resto fueron 90 individuos de 10 especies (4.7%). La composición de las capturas de tiburón azul estuvo compuesta por hembras de 80−242 cm LT con un promedio de 159 ± 2.0 cm, mientras que los machos tuvieron un intervalo de 83−290 cm, con una
0
50
100
150
200
250
300
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC
Palangre
Red
n= 955
No. via
jes
Temporada de pesca de
Langosta

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
172
media de 148.7 ± 2.7 cm. La prueba de t-Student mostró que las longitudes promedio entre sexos fueron diferentes estadísticamente (t(0.05)(521) = 3.16, p= 0.004). La tendencia en las capturas fue que las hembras fueron más grandes que los machos. (Fig. 2b). La estructura de longitudes de P. glauca capturado por la flota artesanal de LM estuvo compuesta por juveniles e individuos inmaduros, con excepción de algunos machos maduros, y una hembra preñada.
Fig. 2. Estructura en longitudes y sexos de las capturas de las principales especies de tiburones capturadas por la flota artesanal de LM, BC durante 2007-2015; a) tiburón azul, Prionace glauca en la temporada 2015; b) tiburón azul durante 2007-2015; c) mako, Isurus oxyrinchus en la temporada 2015 y d) mako durante 2007-2015. Se examinaron 378 hembras y 428 machos tiburones makos. Las hembras fueron de 67−209 cm LT, con promedio de 128.8 ± 1.2 cm. Los machos de 68−212 cm con una media aritmética de 130.9 ± 1.4 cm. La prueba de t-Student demostró que no hay diferencias significativas entre las longitudes de ambos sexos (t = 1.59, gl = 804, p = 0.111) (Fig. 2d). La estructura de longitudes de las capturas de tiburones mako en BSV estuvo compuesta principalmente por individuos juveniles, incluyendo algunos neonatos. No hubo registros de makos maduros, ni machos, ni hembras. Para estimar la CPUE del tiburón azul y el tiburón mako de aletas cortas se emplearon las capturas numéricas de 250 viajes de pesca con palangre por parte de las embarcaciones menores de LM durante el periodo 2007−2015, incluyendo los viajes que se realizaron para la captura de individuos que fueron marcados desde el 2010. Esto porque el equipo de pesca y las unidades de pesca empleados en ambos casos (viajes comerciales y viajes de marcado) fueron los mismos. A partir de las entrevistas realizadas a los pescadores artesanales de LM se documentó que el número total de anzuelos por palangre es de alrededor de 300, cifra que se empleó para estandarizar las capturas numéricas de tiburones por cada 100 anzuelos.
Del total de viajes monitoreados, 79.7% registró capturas de P. glauca (lances positivos) y en 78.8% capturas de I. oxyrinchus. Ordenando las capturas por intervalo de capturas numéricas, se observó que el mayor número de viajes de pesca obtuvieron de 1 a 5 tiburones azules (37.9%) (Fig. 3a). El 81% de los viajes de pesca con captura de mako, tuvieron capturas numéricas de 1-5 individuos (Fig. 3b).
Fig. 3. CPUE ordenada por intervalos de captura numérica documentados en Lε, BC, durante 2007−2015; a) tiburón azul; b) tiburón mako. El “0” representan viajes de pesca con cero capturas. Conclusión Las especies de tiburón P. glauca e I. oxyrinchus son las más capturadas en BSV, con una composición de madurez juvenil. Literatura citada Castillo-Géniz JL. 2015. Estudios sobre las pesquerías de
tiburones y rayas del noroeste del Pacífico mexicano: Investigaciones aplicadas para el mejoramiento de su manejo pesquero y conservación, 2015. Informe Final POA 2015 (Documento Interno). CRIP-Ensenada. INAPESCA.
Castro JI. 1993. The shark nursery of Bull Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States. Enviromental Biology of Fishes. Netherlands 38:37-48.
Palabras clave: Tiburones, pesca ribereña, palangre, CPUE.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
173
083: oral DIAGNÓSTICO DE LOS PRINCIPALES RECURSOS DE ESCAMA MARINA QUE SE PESCAN
EN LA ZONA DE CABO DEL ESTE, BAJA CALIFORNIA SUR
Pablo Noé Castro Moreno, Norma Alejandra Sánchez Reyes y Gustavo D. Danemann
Pronatura Noroeste. Programa de Conservación Marina y Pesca Sustentable. Av de las Ballenas s/n, Int A-5 Col. Fidepaz, La Paz, B.C.S. C.P. 23090. [email protected]. Introducción La pesca a nivel estatal como actividad primaria tiene importancia fundamental por su valor económico, social y alimentario (Cortés-Ortiz et al. 2006, SEPESCA 2011). Baja California Sur (BCS) con 23% de los litorales del país, se distingue a nivel nacional en el contexto pesquero por la captura de especies de escama con importancia económica como pargos, huachinango, cabrillas y jurel, entre otras (Luque-Agraz y Gómez 2007, Cortés-Ortiz et al. 2006). La zona de Cabo del Este es un área de gran importancia para la pesca ribereña y deportiva que ahí se realiza (CONABIO-CONANP-TNC-PRONATURA 2007). La coexistencia de ambos tipos de pesca, deportiva y comercial, ha generado una presión considerable sobre los recursos ribereños de valor comercial; tanto que en la actualidad los mismos pescadores de la zona han observado la reducción del tamaño y calidad de sus capturas. Por lo cual este documento tiene como objetivo caracterizar y generar un diagnóstico de las principales pesquerías de escama en la región, que permita la toma de las medidas necesarias para asegurar la sustentabilidad y el mejoramiento de las condiciones de vida de los pescadores y sus comunidades. Material y métodos El estudio se realizó en el corredor de Cabo del Este, e incluye las comunidades de Boca de Álamo Punta Pescadero, Los Barriles, Buena Vista, La Ribera, y Los Frailes. Para la descripción de los aspectos socioeconómicos y diagnóstico de la pesquería se utilizó información de 4 diferentes fuentes: 1) censos de población y vivienda de INEGI (2012); 2) registros oficiales de las capturas arribadas en las localidades de Cabo del Este del 2003-2013; 3) entrevistas semiestructuradas a pescadores, permisionarios y presidentes de cooperativas y 4) muestreos prospectivos de las capturas en playa. Resultados y discusión Indicadores socioeconómicos. Población: De acuerdo a datos del INEGI (2012) hay un total de 4,033 habitantes en las comunidades pesqueras de Cabo del Este; los centros poblacionales con mayores densidades son: La Ribera, Los Barriles y Buena Vista, existen varias
comunidades costeras con un número bajo de habitantes (<250) (Fig. 1).
Fig. 1. Habitantes por comunidad pesquera en Cabo del Este (Fuente INEGI 2012). A través de entrevistas realizadas entre Duke University en 2013 y Pronatura Noroeste en 2014 a pescadores y permisionario se determinó que el sector pesquero ribereño se encuentra conformado por hombres entre los 18 y los 71 años de edad (Fig. 2).
Fig. 2. Estructura de edades de los pescadores entrevistados en la zona de Cabo del Este. Educación: El nivel educativo alcanzado por los pescadores es bajo, encontrando que del total de entrevistados 64% estudiaron al menos algún año de primaria, 19% cursaron algún año de secundaria y 13% algún año de preparatoria (Fig. 3).
0
500
1000
1500
2000
2500
Boca delÁlamo
LosAlgodones
LosBarriles
BuenaVista
La Ribera LosFrailes
N°
de h
abita
ntes
0%
5%
10%
15%
20%
25%
30%
<20 20-30 31-40 41-50 51-60 61-70 >70
Edades (años)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
174
Fig. 3. Nivel de escolaridad alcanzado por los pescadores entrevistados en cabo del Este. Diagnóstico de la pesquería. Importancia: Con base en información de los avisos de arribo del 2003-2013 se determinó que el volumen promedio anual de escama desembarcado en la última década es de 214 toneladas (t), lo que equivale a un valor promedio anual pagado en playa de $4,280,000 pesos (valor promedio de $20 pesos por kilogramo); la composición de las capturas está integrada por 46 especies de importancia socioeconómica; principalmente las familias Lutjanidae (pargos y huachinango), Serranidae (cabrillas), Carangidae (jureles), Balistidae (cochito), Scombridae (Sierra) y Malacanthidae (pierna y conejo). Grupos de Interés: En esta área existen alrededor de 157 pescadores de escama, de los 91 entrevistados, 67% están afiliados a una cooperativa, 18% trabajan con algún permisionario. En la zona se encuentran registradas alrededor de diez Sociedades Cooperativas de Producción Pesquera (SCPP) las cuales utilizan el área para la captura comercial y arribo. Tendencia histórica: Durante el periodo 2003-2013, la flota de pesca arribó un total 2,358 toneladas del recurso escama, destacando el huachinango (Lutjanus peru) el cual aportó 53.31% de la producción total. Al analizar la serie histórica se puede observar que la pesquería presenta una tendencia a la baja; la captura total promedio de la serie es de alrededor de las 214 t (Fig. 4).
Fig. 4. Tendencia histórica de las capturas del recurso escama periodo 2003-2013 (fuente avisos de arribo). Conclusiones El huachinango es el recurso más importante que se captura por sus volúmenes de captura y valor económico. La adopción de artes de pesca más eficientes (redes de enmalle) han modificado las pesquerías con cambios importantes en sus tasas de capturas. La actividad pesquera en la región ha seguido la misma tendencia que en otras regiones del Golfo de California donde la captura actual está orientada a recursos pesqueros de menor tamaño y valor comercial. Literatura citada CONABIO-CONANP-TNC-PRONATURA. 2007. Análisis de
vacíos y omisiones en conservación de la biodiversidad marina de México. The Nature Conservancy-Programa México, Pronatura, A.C., México, D.F. 130 pp.
Cortés-Ortiz RA, G Ponce-Díaz, M Ángeles-Villa. 2006. El sector pesquero en Baja California Sur: un enfoque de insumo-producto. Región y Sociedad XVIII (35): 107-129.
INEGI. 2012. Censos y conteos de población y vivienda. Página de internet: http://www.inegi. org.mx/est/contenidos/proyectos/ccpv/default.aspx.
Luque-Agraz D, E Gómez. 2007. La construcción de la región del Golfo de California desde lo ambiental y lo indígena. Ra Ximhai 3(1): 83-116.
SAGARPA. Avisos de arribo del 2003-2013 SEPESCA. 2011. Programa Sectorial de Pesca y
Acuacultura 2011-2015 de Baja California Sur. http://fnz.bcs.gob.mx/fnz/wp-content/themes/fnz_bcs/assets/images/transparencia/marco_program/programa_sectoriales/Programa%20Sectorial%20Pesca%20y%20Acuacultura.pdf
Palabras clave: Pesca ribereña, recursos pesqueros, arte de pesca, socioeconómico y escama
0%
5%
10%
15%
20%
25%
30%
35%
40%
45%

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
175
084: cartel ESTRUCTURA DE EDADES DEL TIBURÓN PILOTO, Carchahinus falciformis, CAPTURADO POR LA
PESQUERIA ARTESANAL EN PUERTO MADERO, CHIAPAS
Pablo Delmar Hernández Morales¹, Javier Tovar Ávila2, Edgar Tovar Juárez1 y Felipe Galván Magaña3.
1Instituto de Biociencias. Universidad Autónoma de Chiapas. Puerto Madero, Chiapas. C.P. 30830. [email protected]. 2Centro Regional de Investigación Pesquera de Bahía Banderas. INAPESCA. Calle Tortuga núm. 1, La Cruz de Huanacaxtle, Nayarit, México. C.P. 63732. 3CICIMAR, IPN, Av. I.P.N. s/n, Colonia Playa de Santa Rita. A.P. 592. La Paz, Baja California Sur, México. C.P.23000. Introducción Carcharhinus falciformis es una especie objetivo e incidental de distintas pesquerías artesanales (Dulvy et al. 2008). Se comercializa en el mercado nacional dentro de la categoría “primera especial”, mientras que las aletas son consideradas de primera calidad, siendo exportadas en su totalidad al mercado oriental (CONAPESCA-INP 2004), provocando que esta especie se encuentre actualmente bajo una intensa presión de pesca. Debido a sus características biológicas, como crecimiento lento, fecundidad baja, madurez sexual tardía y gran longevidad que le otorgan una productividad biológica relativamente baja, esta especie es vulnerable a la sobreexplotación. Por lo anterior es de gran utilidad llevar a cabo estudios de edad y crecimiento, que aporten información para su evaluación y determinación de su potencial de explotación. La determinación de la estructura de edades en las capturas puede ayudar también a detectar cambios poblacionales (Cailliet y Goldman 2004). Se ha reportado la disminución en las capturas de la pesquería de Puerto Madero, Chiapas, (Soriano-Velázquez et al. 2006), sin embargo, se han realizado pocas evaluaciones poblacionales de esta pesquería. El objetivo del presente estudio fue comparar la estructura de longitudes y edades de las capturas actuales con la estimada previamente, a fin de detectar posibles disminución en la longitud y edad media de captura. Materiales y métodos Los organismos se obtuvieron de las capturas de la flota artesanal tiburonera en Puerto Madero, Chiapas, provenientes del Golfo de Tehuantepec. Cada organismo fue sexado y se obtuvo su longitud total (LT). Se extrajeron cuando fue posible una muestra de vértebras dorsales. La determinación de la edad de los tiburones se realizó por medio del conteo de las bandas de crecimiento en las vértebras, enteras y cortadas sagitalmente con una cortadora de baja velocidad Isomet. Las bandas de crecimiento fueron contadas con la ayuda de un microscopio estereoscópico con luz transmitida y con luz reflejada. Para resaltar las marcas de crecimiento y hacer los conteos comparables a los de estudios previos de la especie
en la zona (Zea-de la Cruz 2007). Se utilizó el método de tinción con rojo de alizarina 0.01% (La Marca 1996). Se realizaron tres conteos independientes de cada muestra (vértebra completa, cortes sagitales sin y con tinción) y como indicador de la precisión de los conteos se utilizó el coeficiente de variación (CV) (Campana 2001). Los datos de LT fueron agrupados en intervalos de 10 cm y se elaboraron histogramas de frecuencia para conocer la estructura de longitudes de la captura. Se realizó un análisis de varianza (ANOVA) para identificar posibles diferencias significativas entre las longitudes de las hembras y los machos. Finalmente se comparó la estructura de longitudes y edades estimadas con las reportadas en estudios previos de la zona (Soriano et al. 2005, Zea-de La Cruz 2007) mediante varios ANOVA. Resultados y discusión Se registró un total de 175 organismos de C. falciformis. Los machos presentaron longitudes de 54 a 155 cm LT (promedio=104.5 cm) y las hembras de 51 a 195 cm LT (promedio=123 cm) (Fig. 1A). No se encontraron diferencias significativas entre las longitudes en hembra y machos (ANOVA, F = 2, 642, P>0.05). La precisión de los conteos de las bandas de crecimiento en las vértebras de C. falciformis fue aceptable, tanto para los conteos que se les realizaron a las (A) vértebras completas, (B) cortes y (C) cortes con tinción (Fig. 2).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
176
Fig. 1. A) Frecuencias de longitudes. B) Distribución de edades C. falciformis, capturado por la pesquería artesanal de Puerto Madero, Chiapas, durante 2014-2015.
Fig.·2. A) Vértebra completa, B) corte sagital de una vértebra, C) corte sagital de una vértebra con tinción de rojo de alizarina. La precisión intralector del conteo para vértebras enteras (CV = 15.7 %) fue menor a la del conteo de los cortes (CV = 9.79 %) y los cortes con tinción (CV = 9.22 %). Estos dos últimos métodos presentaron por lo tanto precisión similar a la reportadas en otros estudios con vértebras de tiburones (CV >10%) (Campana 2001). Las edades determinadas para C. falciformis abarcaron un intervalo de 0-18 años para sexos combinados (Fig. 1B). La hembra de menor edad (<1 año) tuvo una longitud de 51 cm LT, mientras que la de mayor edad (18 años) tuvo una longitud de 195 cm LT. En el caso de los machos, el organismo de menor edad (1 año) tuvo una longitud de 54 cm LT, mientras que el de mayor edad (11 años) tuvo una longitud de 155 cm LT. La estructura de longitudes y edades determinada para la especie indicó la tendencia a capturar organismos inmaduros de poca longitud y edad, similar a los estudios previos (Zea-de la Cruz 2007). Este autor reportó que más del 85% de los organismos capturado son inmaduros, con longitudes
menores a 160 cm de longitud total. Una adecuada estimación de la edad y de los parámetros de crecimiento es de vital importancia en la investigación pesquera de esta zona, lo que contribuirá al ordenamiento de su explotación para un manejo óptimo. Sí hay diferencias con las longitudes y edades estimadas en los estudios previos que soporten la idea de un impacto negativo de la pesca sobre la especie en la última década. Literatura citada Campana SE. 2001. Accuracy, precision and quality control
in age determination, including review of the use and abuse of age validation methods. J Fish Biol 59:197–241.
Cailliet GM, KJ. Goldman. 2004. Age determination and validation in chondrichthyan fishes. En: Carrier JC, Musick JA y MR Heathus (Eds). Biology of sharks and their relatives. CRC Press, USA, Pp 399–439.
CONAPESCA-INP, 2004. Plan de Acción Nacional para el Manejo y Conservación de Tiburones, Rayas y Especies Afines en México. SAGARPA, México. 85 p.
Dulvy KN, JK Baum, S Clarke, LJV Compagno, E Cortés, A Domingo, S Fordham, S Fowler, MP Francis, C Gibson, J Martínez, JA. Musick, A Soldo, JD Stevens, S Valenti. 2008. You can swim but you can´t hide: the global status and conservation of oceanic pelagic sharks and rays. Aquatic Conservation: Mar. Freshwater Ecosyst. Pub. online in Wiley InterScience. Canadá. 24 pp.
LaMarca MJ. 1966. A simple technique for demonstrating calcified annuli in nthe vertebrae of large elasmobranchs. Ichthyological Copeia (2):351–352.
Soriano-Velásquez S. R., Acal-Sánchez D. E., Castillo-Geniz J. L., Vásquez-Gómez N. y C. E. Ramírez-Santiago. 2006. Tiburón del Golfo de Tehuantepec. Pp. 323–363. En: Sustentabilidad y pesca responsable en México: Evaluación y manejo. INP, SAGARPA.
Zea-de la Cruz H. 2007. Edad y crecimiento del tiburón aleta de cartón Carcharhinus falciformis (Bibron 1839) registrado en el Golfo de Tehuantepec. Tesis de Licenciatura. Instituto Tecnológico de Boca del Río. Boca del Río, Veracruz. 64p.
Palabras clave: Banda de crecimiento, vértebra, borde opaco, longevidad, edad.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
177
085: cartel CARACTERIZACIÓN OPERATIVA DE LAS EMBARCACIONES PRESTADORAS DE SERVICIOS PARA LA
PESCA DEPORTIVA RECREATIVA EN EL ESTADO DE COLIMA
Salvador Ramírez Montes De Oca, Miguel Ángel Carrasco Águila y Mauricio Salas Maldonado
Centro Regional de Investigación Pesquera de Manzanillo. INAPESCA. Playa Ventas S/N, Manzanillo, Colima. México. C.P. 28200. Tel.013143323751, [email protected] Introducción La impresionante población de pez vela en las costas de Manzanillo le ha hecho merecedor a este puerto el título de “Capital εundial del Pez Vela”. Aprovechando la situación, en 1954, a instancias del Club Deportivo de Pesca de Manzanillo A.C., se celebró el Primer Torneo Internacional de Pesca de Pez Vela. Con el tiempo este Torneo ha tenido gran trascendencia en todo el mundo. Asimismmo, en 1957, siendo Gobernador del Estado el General Jesús González Lugo y Presiente Municipal Don Alfredo Woodward Téllez, se le dio a Manzanillo la titularidad de “Capital εundial del Pez Vela” (H. Congreso del Estado de Colima 2004). La pesca deportiva recreativa tiene una gran influencia en el desarrollo del sector pesquero, ya que es una gran fuente generadora de ingresos significativa en la economía del estado entre otros aspectos como lo es la generación de empleos y así mismo impulsa el sector turístico debido a la gran demanda de turistas amantes de este deporte, tomando importancia y convirtiéndose en una de las actividades más importantes y con grandes beneficios para el estado. En general los meses de mayor actividad para las embarcaciones deportivas recreativas son la temporada de Semana Santa por la atracción que representa para el turismo y los meses de octubre a febrero. Las especies destinadas para esta actividad son el pez vela, el marlin (azul, negro y rallado), el pez espada, el dorado, el pez gallo y sábalo, siendo especies restringidas dentro de una franja de 50 millas náuticas a partir de la línea base donde se mide el mar territorial. La pesca deportiva es regulada para así preservar las especies y sea una pesca sustentable (NOM-017-PESC-1994) (SEMARNAT 1995). Se define en la ley, que la pesca deportiva recreativa: es la que se practica con fines de esparcimiento o recreación con las artes de pesca previamente autorizados (SAGARPA 2007). En este trabajo se pretende hacer una caracterización de las embarcaciones dedicas a la pesca deportiva recreativa conociendo sus capturas, fechas de mayor actividad y tiempos de navegación. Material y métodos Se recabó información mediante muestreos semanales durante el 2015 referente a
embarcaciones de pesca deportiva recreativa como: nombre de la embarcación, eslora, tipo de motor y combustible, número de pasajeros, tripulación, horas de navegación, número de cañas, tipo de carnada y organismos capturados, pesos y longitudes. Se clasificaron las embarcaciones y se realizó un análisis de las capturas y horas de navegación. Resultados El muestreo se inició a finales de febrero hasta los últimos días de diciembre de 2015. Se registraron un total de 26 embarcaciones, con un promedio de eslora de 9 m ycapacidad de 10 pasajeros. Se dividieron en base al número de viajes en dos grupos: 13 registraron más de 20 viajes mientras que el restante presentaron por debajo de los 19 viajes siendo el valor máximo 47 y valor mínimo 1 viaje para todo el año. La figura 1 muestra el número de viajes por embarcación.
Fig. 1. Viajes realizados por embarcaciones deportivas recreativas de Manzanillo, Col., en 2015. Durante el año se observó que marzo y abril fueron los de mayor número de viajes con las embarcaciones de pesca deportiva recreativa (Fig. 2).
0
2
4
6
8
10
12
5 10 15 20 25 30 35 40 45 50
Em
barc
aci
ones
Viajes

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
178
Fig. 2. Número de viajes realizados por embarcaciones deportivas recreativas en Manzanillo, Col., en 2015. Durante este periodo se encontró que mayo y diciembre fueron los de mayor captura, siendo el pez vela el más abundante (Fig. 3).
Fig. 3. Meses de mayor captura de embarcaciones de pesca deportiva recreativa de Manzanillo, Colima El pez vela fue el mayor número de organismos capturados con 75.6%, seguido del dorado con 15.1%, el marlin con 8.1% (rayado y negro) y el atún con 1.2% (Fig. 4).
Fig. 4. Especies capturadas por embarcaciones de pesca deportiva recreativa en Manzanillo, Col., en 2015. Conclusiones La mitad de la flota de pesca deportiva recreativa realizó más de 20 viajes durante este periodo de estudio.
Las fechas que corresponden a la Semana Santa y Pascua fueron donde se registró el mayor número de viajes. Para el año 2015 los meses de mayo y diciembre fueron los de mayor captura que incluye a todos los organismos. La especie que presenta la mayor captura en el 2015 fue el pez vela. Literatura citada H. Congreso del Estado de Colima. 2004. Acuerdo que
declara al εunicipio de εanzanillo como “Capital εundial del Pez Vela”. http://www.ordenjuridico.gob.mx/Estatal/COLIMA/Municipios/Manzanillo/MZLAcu4.pdf.
SAGARPA. 2007. Ley General de Pesca y Acuacultura Sustentable. México D.F. Mayo de 2007. 119 p.
SEMARNAT. 1995. Norma Oficial Mexicana NOM-017-PESC-1994, Para regular las actividades de pesca deportivo recreativa en las aguas de jurisdicción federal de los Estados Unidos Mexicanos. Diario Oficial de la Federación el 28 de diciembre de 1994.
Palabras clave: Pesca deportiva recreativa, pez vela, viajes en lancha, caracterización.
0
10
20
30
40
50
60
70
80
90
100
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
Núm
ero
de v
iaje
s
Tiempo (mes)
0
5
10
15
20
25
30
35
40
45
50
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC
Num
. Org
anis
mos
Tiempo (mes)
0
10
20
30
40
50
60
70
VELA DORADO M. RAYADO M. NEGRO ATUN
Num
. Org
anis
mos
ESPECIES

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
179
086: oral DISTRIBUCIÓN, ABUNDANCIA Y HÁBITAT DEL PULPO ROJO Octopus maya EN LA PENÍNSULA DE
YUCATÁN
José Luis Cruz Sánchez1, Josefina Santos Valencia1, Juan C. Espinoza Méndez1, Edgar Cob Pech1 y Diego A. Hau Pacheco2
1Centro Regional de Investigación Pesquera de Yucalpetén. INAPESCA. Carretera a Chelem, Blvd. del pescador s/n Puerto de Abrigo, Yucalpetén, Yucatán. 2Centro Regional de Investigación Pesquera de Lerma. INAPESCA. Carretera Campeche-Lerma, s/n, Campeche. [email protected]. Introducción La pesquería de pulpo que se desarrolla en el Golfo de México aporta 70% de la producción nacional. La captura de pulpo en la península de Yucatán la sustentan dos especies: Octopus maya y O. vulgaris, con producción cercana a 20 mil toneladas en el periodo 2000-2015 (CONAPESCA 2014). El pulpo rojo O. maya es la especie de mayor importancia para la pesquería, misma que contribuye con 60% de la producción del Golfo de México. Es una especie endémica de la península de Yucatán que habita principalmente en cuevas desde la zona costera hasta profundidades de 27 metros (Fig. 1).
Fig. 1. Pulpo Octopus maya en cueva. El pulpo es un recurso abundante, fácil de extraer y su alta demanda en el mercado nacional e internacional ha generado un aumento en la presión de pesca. Por la importancia de la pesquería, además de mantener actualizado el conocimiento de los parámetros poblacionales, es necesario darle un enfoque ecosistémico analizando las características de su hábitat y la estructura del fondo. Materia y métodos Se realizó la evaluación poblacional de O. maya mediante muestreos directos por medio de transectos lineales. La densidad poblacional (ind/m2) se estimó por medio de cuadrantes. Se caracterizó el hábitat en los estados de Campeche y Yucatán. Se realizó un total de 50 video-transectos de 30 m cada uno del fondo marino (junio-julio de 2015), de los cuales 23 se situaron en el estado de Campeche y 27 en
Yucatán, mediante la apreciación in-situ de buzos-pescadores se definió el tipo de fondo en los que se observó algún ejemplar, posteriormente los videos-transectos se analizaron identificando los sustratos vivos (algas, pastos, corales y esponjas) y no vivos (arena, conchuela, fondo duro y cordillera). Se estimaron índices de diversidad de la estructura de fondo y mediante un análisis estadístico se observó si existen diferencias entre el número de organismos con el sustrato de fondo comparando el resultado a un nivel de significancia α = 0.05 (Zar 1996). Resultados La densidad promedio en toda esta área de ocurrencia del pulpo O. maya fue de 1,818 individuos/km2, densidad total de 2λ’7λ6,1λ2 organismos y biomasa de 22,348 t. Se observó un gradiente en la distribución de sur a norte, con mayor densidad de organismos pequeños en Champotón y Seybaplaya (Fig. 2).
Fig. 2. Mapa de distribución y densidad de Octopus maya en la Península de Yucatán, 2015. En los video-transectos en Yucatán se observaron un total de 108 pulpos con una mayor presencia en junio en la comunidad de Río Lagartos, mientras que en Campeche fueron 91 ejemplares con una mayor presencia en junio en el puerto de Campeche y Seybaplaya (Fig. 3). El índice de la diversidad estructural del fondo para el área de Champotón fue
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
180
de 0.397 (Varianza = 0.002) y para el área de Campeche-Seyba de 0.681 (Varianza= 0.001) y con el área de Tenabo de 0.632 (Varianza = 0.002), por lo que en los dos casos son significativamente diferentes en las coberturas del fondo marino, mientras que estas dos últimas áreas no rechazan la hipótesis nula, lo que significa que existen similitudes en las coberturas del fondo marino. El análisis entre el número de pulpos con el fondo mostró que en Campeche la arena tuvo una mayor significancia con la aparición de pulpos, mientras que en Yucatán la presencia de los pulpos se asemejó al fondo duro, resultando no tener diferencias significativas (Fig. 3 y 4).
Fig. 3. Relación de tipos de fondo con el número de pulpos Octopus maya en Campeche.
Fig. 4. Relación de tipos de fondo con el número de pulpos Octpus maya en Yucatán. Discusión Los organismos de pulpo rojo O. maya de mayor longitud se capturan mayormente en los puertos de Yucatán. La presencia y la selección de estos para vivir en agujeros o cuevas les permiten esconderse de sus depredadores (Solís-Ramírez 1967). Sin embargo, la cobertura entre los litorales de Campeche y Yucatán fue diferente en cuanto a sustrato vivo y no vivo, teniendo una mayor presencia de los organismos adultos en zonas donde se presentaron mayormente los fondos duros y mayor presencia de juveniles en zonas predominantes de arena.
La estructura de los fondos puede ser un parámetro importante en la distribución por gradiente de longitudes ampliamente observado en la península de Yucatán (SAGARPA 2014, INAPESCA 2014) y puede estar asociado a zonas de reproducción y reclutamiento. Conclusión Al integrarle un enfoque eco sistémico a los estudios poblacionales de pulpo permitirán tener mayores análisis holísticos que favorezcan a resultados eficaces. El conocer el hábitat de las especies objetivo, es un generador importante en la sistemática del comportamiento y en la distribución de las especies. Literatura citada http://dof.gob.mx/nota_detalle.php?codigo=5338727&fecha
=28/03/2014 CONAPESCA. 2014. Anuario estadístico de acuacultura y
pesca 2012. Comisión Nacional de Acuacultura y Pesca. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA).
INAPESCA. 2014. Dictamen técnico sobre el establecimiento de la cuota de captura de Pulpo Octopus maya para la temporada de pesca 2014. SAGARPA, http://www.conapesca.sagarp a.gob.mx/wb/cona/anuario
Solís-Ramírez M.J. 1967. Aspectos biológicos del pulpo, Octopus maya Voss y Solís. Publ. Inibp 18:1-90.
Zar JH. 1996. Biostatistical analysis. 3rd ed. Prentice Hall. USA., 662p.
Palabras clave: pulpo rojo, hábitat, distribución Octopus maya, Yucatán.
0
10
20
30
40
50
0
20
40
60
80
Champoton Seybaplaya Isla Jaina
Nú
m.
tota
l d
e O
. m
ay
a
% F
on
dia
Zonas de estudio
ARENA CONCHUELA FONDO DUROCORDILLERA No. Pulpos
0
5
10
15
20
25
0
20
40
60
80
Rio lagarto Dzilam Progreso Celestun
% d
e f
on
do
Zonas de estudio
Arena Conchuela Fondo duro
Cordillera No. Pulpos

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
181
087: oral UNIDADES OPERACIONALES PARA EL MANEJO DE PESQUERÍAS RIBEREÑAS EN EL GOLFO DE
ULLOA, BAJA CALIFORNIA SUR, MÉXICO
Julio César Parra Alaniz y Mauricio Ramírez Rodríguez
Centro Interdisciplinario de Ciencias Marinas. Instituto Politécnico Nacional. Av IPN s/n, Col Playa Palo de Santa Rita, La Paz, Baja California Sur. C. P. 23096. [email protected] Introducción Las pesquerías ribereñas, multiespecíficas o de pequeña escala son relevantes por la generación de empleos, productos alimenticios y oportunidades de desarrollo para las comunidades costeras (Morán-Angulo y Cruz-Torres 2007, Ramírez-Rodríguez 2011). Aun así, los estudios dirigidos a esta actividad son pocos y la mayoría se enfocan principalmente en la biología y dinámica poblacional de las especies objetivo (Espino-Barr 2000, Díaz-Uribe et al. 2006, Montoya-Campos 2009), ignorando parcialmente el comportamiento de las flotas. Este último refiere a la distribución espacio-temporal del esfuerzo pesquero en función de la disponibilidad de los recursos, mercados y precios, y de no considerarlo se podrían generar percepciones imprecisas del estado de las pesquerías y, consecuentemente recomendaciones inapropiadas para su manejo (Hilborn y Walters 1992). Analizar y comprender la dinámica de flotas en las pesquerías multiespecíficas es un reto complejo, no obstante, una manera de hacerlo es desagregar las flotas pesqueras en unidades operativas, es decir, grupos de embarcaciones con un patrón de operación similar de acuerdo a los artes de pesca, especies objetivo, lugares y temporadas de pesca (Tzanatos 2006, Monroy et al. 2010, Ojeda-Ruíz 2012). En México, los estudios con este enfoque son escasos. Por esto y con el fin de contribuir al desarrollo de medidas de manejo pesquero, la presente investigación analiza la pesca ribereña en el Golfo de Ulloa, Baja California Sur, México (Fig. 1). El objetivo es definir las flotas pesqueras en unidades operativas útiles para la administración, según su patrón de operación con artes de pesca y especies objetivo, en tiempo y espacio. Material y métodos Se utilizó información sobre sistemas de pesca recabada a través de encuestas a pescadores de la región del Golfo de Ulloa y datos de avisos de arribo de 1998 a 2013 con referencias de fecha, empresa, lugar de pesca y desembarco, captura en kilogramos y precio en playa por especie a nombre común facilitados por la CONAPESCA. La identificación de especies a nombre común, lugares de pesca y desembarco se realizó utilizando el
Atlas de Localidades Pesqueras (Ramírez-Rodríguez et al. 2005) y el catálogo de especies de interés pesquero (Ramírez-Rodríguez et al. 2013).
Fig. 1. Ubicación del Golfo de Ulloa. Para definir especies objetivo se aplicó el índice de importancia relativa en función del aporte en kilogramos, valor económico por especie y su frecuencia de registro:
% IIR = %Peso + %Valor + %Frecuencia Con la información de las encuestas se complementaron los datos de avisos de arribo y se identificaron unidades operativas con base a los artes de pesca y sus especies objetivo. Para esto se asignó el arte de pesca utilizado en cada registro considerando el tipo de especie. Para la definición de zonas de pesca se aplicó un análisis de conglomerado a las distancias en kilómetros entre los lugares de captura y los puertos de desembarco de mayor importancia. Se analizó la importancia de las unidades operativas en espacio y tiempo a través del índice de importancia relativa. Resultados En los avisos de arribo se identificaron más de 86 especies. Las más importantes de acuerdo al IIR fueron verdillo, langosta roja, lenguado, corvina, tiburón, abulón y jurel. Se identificaron más de 70 lugares de desembarco, los más importantes de acuerdo a su frecuencia fueron Puerto Adolfo López Mateos, Punta Abreojos, Estero El Cardón, Laguna La

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
182
Escondida, Las Barrancas, Laguna San Ignacio y San Juanico. Se registraron más de 112 sitios de captura, sin embargo, los de mayor importancia se localizan en áreas aledañas a los puertos de desembarco. Se identificaron 8 unidades operativas por sus artes de pesca y a especies objetivo. Están las de redes de fondo con objetivo de peces demersales, redes pelágicas de malla grande y redes pelágicas de malla mediana para peces pelágicos y tiburones. Otras unidades utilizan trampas y capturan langostas, jaibas y verdillo. La unidad de buceo se dirige a moluscos como abulón, callo de hacha y almeja Catarina. Para la pesca de camarón utiliza redes de arrastre tipo Magdalena I. La pesca de almeja pata de mula se efectúa recolectando manualmente los individuos. El análisis de conglomerados permitió identificar 4 zonas de pesca (Fig. 2).
Fig. 2. Zonas de pesca y puertos de desembarco de mayor importancia. La zona 1 (Punta Abreojos) y la 4 (López Mateos) son de mayor importancia para las unidades operativas que trabajan a través de buceo y con redes la mayor parte del año. En las zonas 1, 2 y 4 destaca la unidad que operan con trampas de junio a diciembre. En las zonas 3 y 4 se pesca camarón durante los primeros 3 meses de la temporada de pesca. Discusiones y conclusiones Existen diferencias en la operación espacial (4 zonas) y temporal (de acuerdo a la disponibilidad de la especie) de las 8 unidades operacionales definidas. Estas deben ser consideradas en la evaluación de efectos de medidas de manejo como vedas temporales (camarón, langosta, abulón y tiburón) y facilitar el manejo basado en enfoques integrados (Monroy et al. 2010). Con el enfoque de unidades operacionales, se pueden definir y cuantificar los impactos que ocasionan los traslapes o interacciones entre las flotas (Ojeda-Ruíz de la Peña 2012).
Se observaron traslapes espaciales y temporales entre las unidades que utilizan trampas, redes, buceo y Magdalena I. El enfoque de la dinámica de flotas con base en las unidades operacionales abre un camino a la comprensión de actividades de las pesquerías ribereñas, y es importante probar y desarrollar métodos que contribuyan a la definición de esas unidades y sus oportunidades de uso en el marco de programas de ordenamiento y planes de manejo pesquero. Literatura citada Díaz-Uribe JG, F Arreguín-Sánchez, MA Cisneros-Mata.
2006. Multispecies perspective for small-scale fisheries management: a trophic analysis of La Paz Bay in the Gulf of California, México. Ecol. Mod. 201: 205-222.
Espino-Barr E. 2000. Criterios biológicos para la administración de la pesca multiespecífica artesanal en la Costa de Colima, México. Tesis de Dr. UdeC, Tecomán, Colima, México. 162 p.
Hilborn R, CJ Walters. 1992. Quantitative fisheries stock assessment: Choice, dynamics and uncertainty. Chapman and Hall. 570p.
Montoya-Campos M. 2009. Cambios en la composición específica de la captura artesanal de escama al sur de la Isla San José, Baja California Sur, México. Tesis de Maestría. CICIMAR. La Paz, Baja California Sur, México, 102 p.
Morán Angulo RE, MC Cruz Torres. 2007. Pesca, Medio Ambiente y Sustentabilidad en Sinaloa. UAS. Culiacán, Sin., México. 2007. 13-39 p.
Ojeda-Ruíz MA. 2012. Interacciones entre pesquerías ribereñas en Bahía Magdalena-Almejas, BCS, México. Tesis Doctorado. Centro interdisciplinario de Ciencias Marinas. Instituto Politécnico Nacional. 127p.
Monroy C, S Salas, J Bello-Pineda. 2010. Dynamics of fishing gear and spatial allocation of fishing effort in a multispecies fleet. North American Journal of Fisheries Management 30:1187-1202.
Ramírez-Rodriguez M. 2011. Data collection on the small –scale fisheries of México. ICES Journal of Marine Science 68(8): 1611-1614.
Palabras clave: Pesquería multiespecífica, dinámica de flotas, manejo pesquero, artes de pesca, especies objetivo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
183
088: oral EDAD Y CRECIMIENTO DEL TIBURÓN PILOTO, Carcharhinus falciformis, EN EL OCÉANO PACÍFICO
ORIENTAL TROPICAL
Fabián Cervantes Gutiérrez1, Felipe Galván Magaña1; Felipe Neri Melo Barrera1 y Javier Tovar-Ávila2
1Centro Interdisciplinario de Ciencias Marinas. IPN. Av. IPN s/n. Col. Playa Palo de Santa Rita. La Paz, Baja California Sur, C.P. 23096. Apartado Postal 592. 2Centro Regional de Investigación Pesquera de Bahía de Banderas. INAPESCA. Calle Tortuga Núm. 1, La Cruz de Huanacaxtle, Nayarit, México. C.P. 63732. [email protected] Introducción Carcharhinus falciformis habita en aguas tropicales y subtropicales del mundo (Compagno 1990), siendo la principal especie en las capturas de tiburón en la zona tropical. En México esta especie representa 51% de las capturas de tiburones y es la más frecuente en la captura incidental de los barcos atuneros en el Pacífico Oriental tropical (POT) (FAO 1998). A pesar de su importancia y de la preocupación por el estatus de sus poblaciones a nivel mundial, los trabajos sobre su biología son escasos (Bonfil et al. 1993). Existen diversos estudios de edad y crecimiento individual en el Pacífico Occidental y en el Golfo de México (Sánchez-de Ita et al. 2011, Branstetter 1987). Sin embargo, no hay información al respecto para la zona oceánica del POT. Por lo que se presenta como objetivo estimar la edad y crecimiento de C. falciformis en esta región. Material y Métodos Se recolectaron datos biométricos de 499 organismos capturados incidentalmente en barcos atuneros, que operan en aguas oceánicas entre México y Ecuador. Los organismos fueron medidos, se les determinó el sexo y se obtuvo una muestra de vértebras cervicales de 347 tiburones, que se etiquetaron y congelaron para su procesamiento en el laboratorio. Las vértebras se limpiaron y cortaron longitudinalmente, para estimar la edad a partir del conteo de bandas de crecimiento. Se determinó la proporción de sexos, la estructura de longitudes, la relación longitud total-radio de la vértebra (LT-RV), la periodicidad de formación de las bandas, la estructura de edades y el crecimiento individual mediante los modelos de von Bertalanffy y de Fabens (1965). Resultados y discusión Se encontró una proporción sexual de 1:0.94 (237 machos y 223 hembras). El intervalo de longitudes para hembras fue de 56.6-240.3 cm LT y los machos de 59.5-206 cm LT (Fig. 1), no se detectaron diferencias significativas en la longitud debidas al sexo (p>0.05). δos adultos (≥180 cm) representaron 9.8% de la muestra, la mayor incidencia fueron juveniles, lo cual concuerda con Watson et al. (2008) y con Amandè et
al. (2008), con predominancia de organismos menores a 90 cm en el este del Océano Pacífico (<λ0 cm δT) y en el Océano Índico (≤100cm) respectivamente. De acuerdo con la longitud de madurez reportada por Hoyos-Padilla et al. (2012), sólo 10% de las muestras obtenidas en el presente estudio corresponde a organismos adultos, lo cual implica que el impacto ocasionado por la pesca con redes de cerco de atún en el OPO sobre C. falciformis puede ser aún mayor con el tiempo. Para reducir esta incidencia, Watson et al. (2008) proponen cierres espaciales. El período de formación de las bandas de crecimiento fue anual, lo cual concuerda con lo reportado por otros autores (Bonfil et al. 1993, Oshitani et al. 2003, Shoou-Jeng et al. 2008, Sánchez-de Ita et al. 2011) para esta especie, a pesar de que ha sido por métodos distintos, mencionan que la banda opaca se forma en verano-otoño y la banda translúcida en invierno-primavera.
Fig. 1. Estructura de edades de Carcharhinus falciformis capturado por la flota atunera del Pacífico oriental tropical. La estructura de edades fue de 0 a 19 años para hembras y de 0 a 18 años para machos (Fig. 1), sin observarse diferencias significativas (p>0.10). Los valores de la ecuación de von Bertalanffy (MCVB) fueron: L∞ = 340.45 cm LT, k = 0.04 (año-1) y t0 = -6.53 años y los resultados del modelo de Fabens (1965) fueron: L∞ = 269.32 cm LT, k = 0.068 (año-1). En cuanto a los valores de los parámetros del MCVB se puede observar que la L∞ (340.45 cm LT) es similar a la longitud máxima reportada (330 cm LT) (Garrick 1982). En cuanto al modelo de Fabens

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
184
(1965), la L∞ (269.32 cm LT) resultó menor que la longitud máxima reportada, pero cercana a la longitud máxima observada en el presente estudio (240.3 cm LT). Los valores del parámetro k observados en el presente estudio, tanto en el MCVB como en el modelo de Fabens (1965) son menores a los reportados estudios anteriores para esta especie, teniendo una mayor similitud con el valor observado por Shoou-Jeng et al. (2008) en los tres casos los valores son menores a 0.10, lo cual sugiere que esta especie es de lento crecimiento, como la mayoría de los elasmobranquios. En el presente estudio, los resultados de los parámetros del ajuste que utiliza L0 (Fabens 1965) fueron útiles, ya que la L∞ fue más cercana a la longitud de máxima observada que la que resultó del MCVB, además de que el valor de L0 es considerablemente grande (entre 50 y 80 cm LT). En cuanto al valor del parámetro k los valores de ambos ajustes se encuentran entre los reportados para otras especies de la familia Carcharhinidae. Watson et al. (2008) y Amandè et al. (2008) reportan que existe una gran preocupación por la conservación y manejo de C. falciformis, ya que tanto en el Océano Pacífico como en el Índico, suponen que se está sobreexplotando, por lo que se requiere la elaboración de un plan de manejo internacional adecuado para esta especie, para que el recurso se mantenga sustentable. Conclusiones La pesquería de atún que opera en el OPO tropical incide sobre organismos juveniles de C. falciformis principalmente (90%). C. falciformis es una especie longeva. El valor de k y la alta incidencia de organismos juveniles de C. falciformis es una alerta para crear medidas regulatorias. Literatura citada Amandè MJ, E Chassot, P Chavance, R Pianet. 2008. Silky
shark (Carcharhinus falciformis) bycatch in the French tuna purse-seine fishery of the Indian Ocean. IOTC-WPEB-2008/016.
Bonfil R, R Mena, D de Anda. 1993. Biological Parameters of Commercially Exploited Silky Sharks, Carcharhinus falciformis, from the Campeche Bank, Mexico. NOAA Tech. Rep. NMFS. 115: 73-86.
Branstetter S. 1987. Age, growth and reproductive biology of the silky shark, Carcharhinus falciformis, and the scalloped hammerhead, Sphyrna lewini, from the northwestern Gulf of Mexico. Environ. Biol. Fish. 19(3): 161-173.
Compagno LJV. 1990. Shark exploitation and conservation. En: Pratt Jr, HL, SH Gruber y T Taniuchi (Eds). pp. 391-415. Elasmobranch as living resources: Advances in Biology, Ecology, Systematics and the Status of the Fisheries, NOAA Tech. Rep. NMFS 90.
Fabens AJ. 1965. Properties and fitting of the Von Bertalanffy growth curve. Growth 29: 265-289.
FAO. 1998. Noticias: tiburones y aves marinas en el programa de una reunión internacional de pesca. 22 de Octubre de 1998.
Garrick JAF. 1982. Sharks of the Genus Carcharhinus. U.S. Department of Commerce. NOAA Tech. Rep. N. M. F. S. 445 194 pp.
Hoyos-Padilla EM, BP Ceballos-Vázquez, F Galván-Magaña. 2012. Reproductive Biology of the Silky shark Carcharhinus falciformis (Chondrichthyes: Carcharhinidae) off the west coast of Baja California Sur, México. Aqua. International J. of Ichthyol. 18: 1-15.
Oshitani S, H Nakano, S Tanaka. 2003. Age and growth of the silky shark Carcharhinus falciformis from the Pacific Ocean. Fish. Sci. 69 (3): 456-464.
Sánchez-de Ita JA, C Quiñones-Velázquez, F Galván-Magaña, N Bocanegra-Castillo y F Félix-Uraga. 2011. Age and growth of the silky shark Carcharhinus falciformis from the west coast of Baja California Sur, México. J. Appl. Ichthyol. 27: 20-24.
Shoou-Jeng J, C Che-Tsung, L Hsian-Hau, L Kwang-Ming. 2008. Age, growth, and reproduction of silky sharks, Carcharhinus falciformis, in northeastern Taiwan waters. Science Direct, Fish. Res. Taiwan. 90: 78-85.
Watson JT, TE Essington, CE Lennert-Cody, MA Hall. 2008. Trade-Offs in the Design of Fishery Closures: Management of Silky Shark Bycatch in the Eastern Pacific Ocean Tuna Fishery. Conserv. Biol. 23(3): 626-625.
Palabras clave: Biocronología, pesca incidental, tiburón sedoso, crecimiento individual, planes de manejo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
185
089: cartelZONAS PRIORITARIAS PARA EL MANEJO Y CONSERVACIÓN DE TIBURONES EN EL GOLFO DE
MÉXICO Y CARIBE MEXICANO
Shasta Keyes Pulido1, Héctor Reyes Bonilla2 y Alfonso Aguilar Perera1
1Universidad Autónoma de Yucatán, Campus Ciencias Biológicas y Agropecuarias, A. P. 4-116 Col. Itzimná, Mérida, Yucatán, México. C. P. 97100. [email protected]. 2Departamento de Ciencias Marinas y Costeras. Universidad Autónoma de Baja California Sur, Apartado postal 19-B, La Paz, Baja California Sur, México. C.P. 23080. Introducción A nivel mundial, los tiburones se encuentran bajo amenaza de extinción (Dulvy et al. 2008). Su vulnerabilidad se debe a que presentan madurez sexual tardía, períodos lentos de gestación y baja fecundidad (Musick 1999). Al menos tres cuartas partes de las especies se encuentran afectados por sobrepesca a nivel mundial, lo que ha disminuido sus abundancias (Myers y Worm 2003), por pesca y degradación de hábitats de crianza y alimentación (Ferretti et al. 2010). Una de las estrategias implementadas para favorecer la recuperación de poblaciones de especies consideradas como más propensas a la extinción, es la creación de reservas marinas que propician la conectividad ecológica a nivel poblacional (Spalding et al. 2010). En México se han instituido refugios en sus principales zonas de reproducción, ya que es el sexto país con mayor volumen de captura de tiburón en el mundo (Fischer et al. 2012). Dado que estas zonas sólo cubren un estadio de vida de las especies, es necesario identificar cuáles son otras zonas prioritarias para la conservación de tiburones en el Golfo de México (GM) y Caribe Mexicano (CM), donde existen alrededor de 80 especies. El objetivo de este trabajo es determinar zonas prioritarias de conservación de tiburones en esta región con base en la identificación geográfica de áreas con mayor riqueza de especies, para apoyar las estrategias de manejo federales para la protección de tiburones.
Fig. 1. Tiburones enlistados por UICN presentes en el Golfo de México y Caribe Mexicano. DD= datos insuficientes, LC= Preocupación menor, NT= Amenazados, VU= Vulnerables, EN= En Peligro2, N= 60 especies. Material y métodos Con base en los datos de la Evaluación Mundial de Tiburones (Lucifora et al. 2009), se mapearon las especies del GM y CM dentro de cuadrantes (1º de
latitud por 1º de longitud). Las especies presentes se relacionaron y caracterizaron con base en cuatro criterios: 1) riqueza de especies; 2) categorías de riesgo de UICN; 3) reconocimiento en la Carta Nacional Pesquera (CNP, DOF 2012); y 4) índice de vulnerabilidad a la pesca de Cheung et al. (2005). Se generaron mapas identificando los pixeles donde se distribuyen las especies más susceptibles. Resultados Se analizaron 1,199 registros de 60 especies (6 órdenes y 20 familias), de las cuales 3% se encuentran en la categoría de “En Peligro (EN)”, 27% son “Vulnerables (VU)” y 20% “Amenazadas (NT)”, de acuerdo con UICN (Fig.1). De acuerdo con la CNP, 17% de las especies de la región son consideradas como “pesca objetivo”, 33% como “pesca asociada” y 50% no se encuentran enlistadas (Fig. 2).
Fig. 2. Tiburones enlistados en la Carta Nacional Pesquera presentes en el Golfo de México y Caribe Mexicano. 3 = pesca objetivo, 2 = pesca asociada y 1 = no enlistados; N = 60 especies. En el analisis espacial de la relación entre la distribución y el índice de vulnerabilidad de las especies, se encontró un valor promedio de entre 64.5 y 79 (100 indica máxima vulnerabilidad; Fig 3). Los valores más altos ocurrieronn en el centro del GM y adyacentes a la península de Yucatán. Como resultado del análisis geográfico que integra el análisis multicriterio, se encontró que los pixeles con los valores de conservación más altos se encuentran en el centro del GM, por la predominancia de especies pelágicas de gran relevancia en la conservación, porque su estado se ha deteriorado debido a la pesca (Fig. 4).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
186
Fig. 3. Valores promedio por pixel del análisis de la distribución de las especies de tiburones y el índice de vulnerabilidad en el Golfo de México y Caribe Mexicano (Cheung et al. 2005).
Fig 4. Zonas prioritarias de manejo y conservación de tiburones que se sugieren para la zona el Golfo de México y Caribe Mexicano. Discusión y conclusión De las especies de tiburones en la región, 37% se encuentran en la categoría de DD (Lista Roja UICN), no hay suficiente información para determinar tendencias poblacionales, mala posición en cuanto a conservación. La mitad de las especies registradas no están enlistadas en la CNP por que no son blanco de capturas, pero hay aprovechamiento de forma incidental. Según Salomón-Aguilar et al. (2009), 50% de los registros de pesca del GM está integrado por organismos inmaduros y hembras grávidas. Además, la falta de un sistema adecuado de registros de captura por especie, imposibilita estimar con datos certeros las tendencias demográficas de las especies de tiburones. Los valores promedio por pixel resultado del Índice de vulnerabilidad de Cheung et al. (2005), denotan en todo el GM valores arriba de 64.5, lo cual indica que en promedio la región concentra especies que deben ser manejadas con cuidado. El más alto grado de vulnerabilidad se presenta en el centro del GM y en la costa oriental de la Península de Yucatán, dponde se está proponiendo la creación de una gran reserva de la biósfera. Los datos aquí presentados apoyan dicha iniciativa. Las especies más vulnerables a la pesca en la región son Alopias vulpinus, Carcharinus longimanus, Isurus oxyrinchus y Sphyrna lewini, categorizadas como
“Vulnerables” y “En Peligro” por UICN y con alta vulnerabilidad (Cheung et al. 2005), con valores mayores a 74, y están enlistadas como “pesca objetivo” en la CNP. Se sugiere prestar atención específica para su manejo efectivo y conservación. Para determinar las zonas prioritarias de manejo y conservación de tiburones, el análisis multi-criterio indica que el centro del GM, así como la zona costera de Campeche y norte de Yucatán representan sitios prioritarios. Esto concuerda con Salomón-Aguilar (2009) y Pérez-Jiménez y Mendoza-Loeza (2015) por ser sitios de crianza. Este trabajo podría ser un apoyo importante para la toma de decisiones, y la identificación de prioridades relacionadas con el manejo sustentable de los elasmobranquios como recurso pesquero. Literatura citada Cheung WWL, TJ Pitcher, D Pauly, 2005. A fuzzy logic
expert system to estimate intrinsic extinction vulnerabilities of marine fishes to fishing. Biol. Conserv. 124:97-111
DOF 2012. Carta Nacional Pesquera. Diario Oficial de la Federación. México. 24 de agosto de 2012.
Dulvy NK, JK Baum, S Clarke, S Compagno et al. 2008. You can swim but you can’t hideμ the global status and conservation of oceanic pelagic sharks and rays. Aquat. Conserv. 18: 459–482.
Ferretti F, B Worm, GL Britten, MR Heithaus, HK Lotze. 2010. Patterns and ecosystem consequences of shark declines in the ocean. Ecology Letters 13: 1055–1071.
Fischer J, K Erikstein, B D’Offay, ε Barone, S Guggisberg. 2012. Examen de la aplicación del Plan de acción internacional para la conservación y ordenación de los tiburones. Circular de Pesca y Acuicultura FAO C1076.
Lucifora LO, VB García, B Worm. 2009. The global shark distribution database. http://www.globalshark.ca/gs_ distribution_db/data.php
Musick JA. 1999. Criteria to define extinction risk in marine fishes. Fisheries 24 (12): 6 –14.
Myers RA, B Worm. 2003. Rapid worldwide depletion of predatory fish communities. Nature 423: 280–283.
Pérez-Jiménez JC, I Méndez-Loeza. 2015. The small-scale shark fisheries in the southern Gulf of Mexico: Understanding their heterogeneity to improve their management. Fisheries Research 172: 96–104.
Salomón-Aguilar C, C Villavicencio-Garayzar, H Reyes-Bonilla. 2009. Determinación de áreas prioritarias de manejo en el Golfo de México. Mesoamericana 13 (2): 174 pp.
Spalding M, Wood LJ, Fitzgerald C, Gjerde K. 2010. The 10% target: where do we stand? In Global Ocean Protection: Present Status and Future Possibilities. Toropova C, Meliane I, et al (eds). Brest, France: IUCN WCPA; UNEP-WCMC; TNC; UNU; and WCS: Gland, Switzerland; Washington, DC and NY, USA; 25–41pp.
Palabras clave: Pesca, vulnerabilidad, tiburones, Golfo de México.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
187
090: oral COMPOSICIÓN E IMPORTANCIA ECONÓMICA DE LA PESCA RIBEREÑA EN LA LOCALIDAD DE
CHACALA EN LA COSTA CENTRO-SUR DE NAYARIT EN 2014
Sergio O. Meza Tabares, José Luis Patiño Valencia, Ma. Consepción Luna Raya, Francisco Javier de la Cruz González y Alejandro Pérez Velázquez.
Centro Regional de Investigación Pesquera de Bahía de Banderas. INAPESCA. Calle Tortuga núm.1, La Cruz de Huanacaxtle, Nayarit. Tel. 01(322)1705184. [email protected] Introducción Entre los escasos estudios que abordan la problemática de la pesca ribereña nacional, están Cruz Romero et al. (1996) en Colima y Jalisco, y López Martín et al. (1996) en Sonora, donde se describen aspectos taxonómicos, análisis de las tendencias en las capturas comerciales y el estado actual de los recursos pesqueros. Durante 2014 el Océano Pacífico aportó más de 80% del volumen de producción pesquera nacional. Nayarit sólo aporta 2% del volumen de captura total (SAGARPA 2013), pero aun así no deja de ser una actividad económica importante (Ulloa Ramírez et al. 2008), y genera alimentos, ingresos y empleos para la población costera, ya se pescan varias especies importantes como: sierra, corvina, huachinango, manta, etc. Este trabajo analizó las capturas registradas durante 2014 en la localidad de Chacala, municipio de Compostela, Nayarit. Material y métodos Se utilizó la información recabada durante el 2014 de embarcaciones ribereñas de la localidad de Chacala. Los datos utilizados se obtuvieron de las bitácoras de pesca en las que se registra la producción, gastos e ingresos de cada embarcación, nombre del pescador, recurso, precio al que se vendió el producto y el valor obtenido por la venta de la captura. Se clasificaron a nivel familia las capturas registradas. Se determinó el esfuerzo pesquero realizado y se tomó como la unidad representativa del esfuerzo al número de viajes (González-Becerril et al. 2000). Resultados Se registraron 111,808 kg de captura total y se dividieron entre 25 familias de peces y dos de crustáceos, siendo las de mayor importancia económica: Sphyranidae con 22%, Haemulidae con 16%, Scombridae con 14%, Scianidae 10% y Lutjanidae con 4% (Fig. 1). El valor total de la captura fue de $1’786,940.00 pesos. Las capturas con mayor valor económico fueron de las familias Sphyranidae (21%), Scombridae (19%), Haemulidae (17%) y Lutjanidae (14%). La variación mensual de la captura indica que marzo, abril y junio reportaron las mayores capturas,
mientras que los meses con menores capturas fueron junio, noviembre y diciembre (Fig. 2).
Fig. 1. Familias de mayor importancia en las capturas registradas de la pesca ribereña de la localidad de Chacala, Nayarit.
Fig. 2. Variación mensual de las capturas. El esfuerzo pesquero aplicado fue de 46 embarcaciones que en total realizaron 2,383 viajes de pesca. El 60% de las capturas y esfuerzo pesquero, se concentró en sólo 10 embarcaciones, por lo que cada una tuvo en promedio de 141 viajes de pesca, que equivalió a una captura de 68,913 kg, una utilidad de $86,577 pesos anuales y un costo por viaje de $193 pesos (Tabla 1). La captura por unidad esfuerzo (CPUE) como indicador de la abundancia relativa para cada una de las familias con mayor importancia económica, indicó que las capturas más abundantes fueron para la familia Sphyranidae con 50 kg/viaje en los meses de enero y febrero, y los revueltos con 50 kg/viaje en el mes de junio, seguidas de las familias Scombridae y Haemulidae con 42 y 39 kg/viaje durante marzo, respectivamente; mientras que los organismo de las familias Scianidae con 20 kg/viaje en mayo y
4
10
14
16 22
23 Lutjanidae
Scianidae
Scombridae
Haemulidae
Sphyranidae
revuelto
-
5,000
10,000
15,000
20,000
25,000
EN F M AB MY JN JL AG S O N D
Ca
ptu
ra (
kg
)
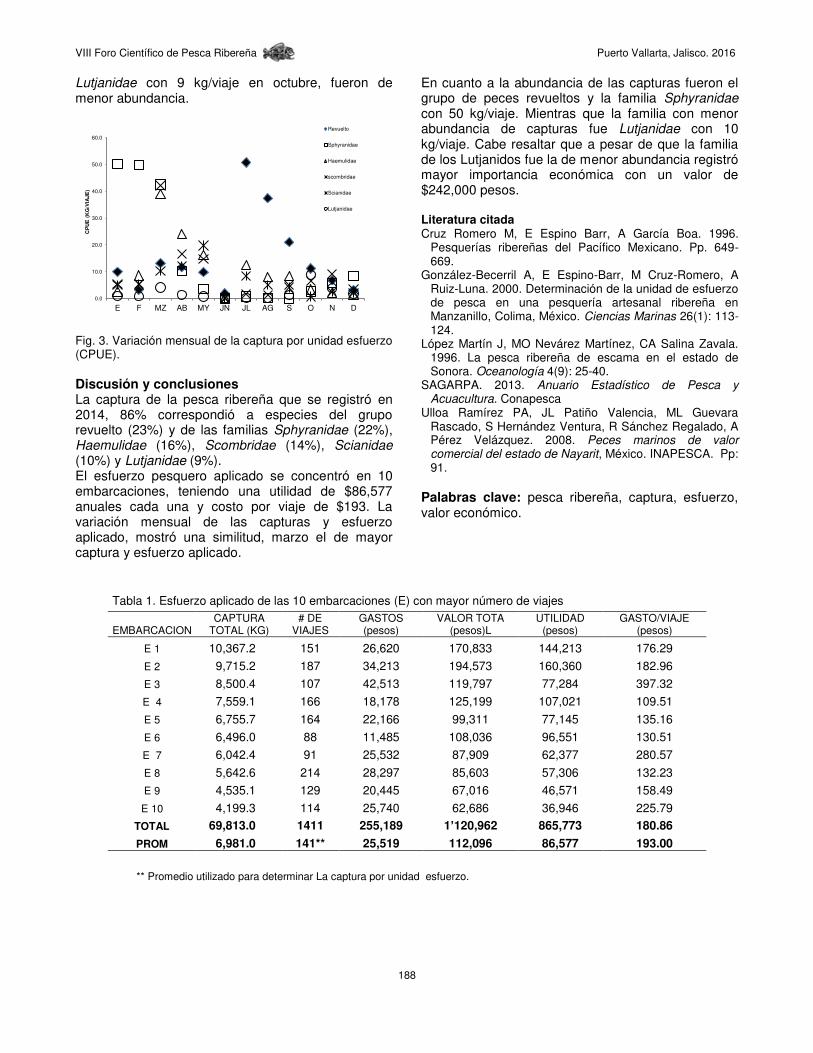
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
188
Lutjanidae con 9 kg/viaje en octubre, fueron de menor abundancia.
Fig. 3. Variación mensual de la captura por unidad esfuerzo (CPUE). Discusión y conclusiones La captura de la pesca ribereña que se registró en 2014, 86% correspondió a especies del grupo revuelto (23%) y de las familias Sphyranidae (22%), Haemulidae (16%), Scombridae (14%), Scianidae (10%) y Lutjanidae (9%). El esfuerzo pesquero aplicado se concentró en 10 embarcaciones, teniendo una utilidad de $86,577 anuales cada una y costo por viaje de $193. La variación mensual de las capturas y esfuerzo aplicado, mostró una similitud, marzo el de mayor captura y esfuerzo aplicado.
En cuanto a la abundancia de las capturas fueron el grupo de peces revueltos y la familia Sphyranidae con 50 kg/viaje. Mientras que la familia con menor abundancia de capturas fue Lutjanidae con 10 kg/viaje. Cabe resaltar que a pesar de que la familia de los Lutjanidos fue la de menor abundancia registró mayor importancia económica con un valor de $242,000 pesos. Literatura citada Cruz Romero M, E Espino Barr, A García Boa. 1996.
Pesquerías ribereñas del Pacífico Mexicano. Pp. 649-669.
González-Becerril A, E Espino-Barr, M Cruz-Romero, A Ruiz-Luna. 2000. Determinación de la unidad de esfuerzo de pesca en una pesquería artesanal ribereña en Manzanillo, Colima, México. Ciencias Marinas 26(1): 113-124.
López Martín J, MO Nevárez Martínez, CA Salina Zavala. 1996. La pesca ribereña de escama en el estado de Sonora. Oceanología 4(9): 25-40.
SAGARPA. 2013. Anuario Estadístico de Pesca y Acuacultura. Conapesca
Ulloa Ramírez PA, JL Patiño Valencia, ML Guevara Rascado, S Hernández Ventura, R Sánchez Regalado, A Pérez Velázquez. 2008. Peces marinos de valor comercial del estado de Nayarit, México. INAPESCA. Pp: 91.
Palabras clave: pesca ribereña, captura, esfuerzo, valor económico.
0.0
10.0
20.0
30.0
40.0
50.0
60.0
E F MZ AB MY JN JL AG S O N D
CP
UE
(K
G/V
IAJE
)
Revuelto
Sphyranidae
Haemulidae
scombridae
Scianidae
Lutjanidae
Tabla 1. Esfuerzo aplicado de las 10 embarcaciones (E) con mayor número de viajes
EMBARCACION CAPTURA
TOTAL (KG) # DE
VIAJES GASTOS (pesos)
VALOR TOTA (pesos)L
UTILIDAD (pesos)
GASTO/VIAJE (pesos)
E 1 10,367.2 151 26,620 170,833 144,213 176.29
E 2 9,715.2 187 34,213 194,573 160,360 182.96
E 3 8,500.4 107 42,513 119,797 77,284 397.32
E 4 7,559.1 166 18,178 125,199 107,021 109.51
E 5 6,755.7 164 22,166 99,311 77,145 135.16
E 6 6,496.0 88 11,485 108,036 96,551 130.51
E 7 6,042.4 91 25,532 87,909 62,377 280.57
E 8 5,642.6 214 28,297 85,603 57,306 132.23
E 9 4,535.1 129 20,445 67,016 46,571 158.49
E 10 4,199.3 114 25,740 62,686 36,946 225.79
TOTAL 69,813.0 1411 255,189 1’120,962 865,773 180.86
PROM 6,981.0 141** 25,519 112,096 86,577 193.00
** Promedio utilizado para determinar La captura por unidad esfuerzo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
189
091: oral DISTRIBUCIÓN Y ABUNDANCIA DE BATOIDEOS CON MAYOR INCIDENCIA EN LA FAUNA DE
ACOMPAÑAMIENTO DEL CAMARÓN EN EL GOLFO DE TEHUANTEPEC
Adriana Jazmín Alatorre Alba, A. Alejandra Lizárraga Rodríguez, Ana B. Guzmán Castellanos, Oswaldo Morales Pacheco y Marco A. Martínez Muñoz
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prolongación Playa Abierta S/N, Col. Miramar, Salina Cruz, Oaxaca. [email protected] Introducción En la pesquería de camarón la captura incidental es desechada casi en su totalidad, conocida como fauna acompañante (FAC), entre las que se encuentran batoideos que, debido a su tamaño, no representan valor comercial (Tapia-García 1997). En estudios realizados se ha observado que la distribución de los organismos costeros difiere con la etapa de vida, sexo y hora del día (Drymon et al. 2011), así mismo las especies que se distribuyen en hábitats restringidos, como ocurre con la mayoría de las rayas, aumentan el riesgo de extinción, debido a que esto reduce su capacidad de recuperación (Fischer et al. 1995). En el caso de los batoideos capturados en la FAC, la mayor incidencia ocurre en zonas cercanas a la costa, bahías y lagunas costeras poco profundas (Núñez-Orozco et al. 2013), por lo que se considera indispensable identificar las zonas de distribución de las especies con mayor incidencia en la pesquería de camarón, que permita determinar zonas prioritarias para su protección y manejo adecuado de posibles especies vulnerables. Material y métodos El Golfo de Tehuantepec (GT) se localiza al sur del Pacífico Tropical, en la zona 90 de pesca (Subzonas 91, 92, 93, 94 y 95), delimitada entre Punta Chipehua, Oaxaca y Puerto Madero, Chiapas (Cervantes-Hernández y Egremy-Valdez 2013). Las muestras fueron tomadas de 8 cruceros a bordo de dos embarcaciones camaroneras durante el 2015 (febrero, marzo, mayo, junio, julio y agosto). Se realizaron lances de 1 hora entre 14 y 128 m. Se empleó una red de arrastre de 7.6 m de longitud de nylon monofilamento con malla de 4.5 cm. Los organismos se identificaron con la clave de Fischer et al. (1995), y se agruparon en las siguientes categorías: neonatos (NN), juveniles (J), adultos (A) y hembras grávidas (HG) de acuerdo a la fase de madurez sexual utilizando la escala Stehmann (2002) por subzona. Para conocer la distribución de las especies se georreferenciaron las estaciones de muestreo mediante el programa Trip & Waypoint Manager v4 (2007) y se estimó la abundancia relativa por especie y subzona.
Resultados Los organismos analizados pertenecen a 11 especies de las cuales Urotrygon chilensis fue la más abundante con 39.2%, seguida por Narcine vermiculatus (27.1%) y U. nana (17.7%), mientras que el resto de especies (N. entemedor, Rhinobatos glaucostigma, R. leucorhynchus, Aetobatus narinari, Dasyatis longa, Zapterix xyster, Gymnura marmorata. y Rhinoptera steindachneri) representó 16%. La mayor incidencia de U. chilensis se encontró en la subzona 94 con 29.4% (Fig. 1) a 9 m de profundidad, 7% fueron hembras (H), 6% machos (M) y 3.5% HG. El mayor número de NN y J se capturaron en las subzonas 94 y 95, mientras que los A y HG frente al complejo lagunar Huave (91 y 92).
Fig.1. Distribución y abundancia de Urotrygon chilensis en
el Golfo de Tehuantepec. N. vermiculatus se capturó frente al complejo lagunar Huave con 31%. Los NN se presentaron con mayor incidencia en las subzonas 92 y 93, los J y A en la 92 y las HG en la 91 (Fig. 2). La mayor abundancia de NN y A se registró entre 8-9 m, los J y HG a los 10-11m y 24-25 m, respectivamente.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
190
Fig. 2. Distribución y abundancia de Narcine vermiculatus.
Se encontró mayor incidencia de U. nana en las 95 (60.5%) y 91 (18.6%) (Fig. 3); los NN-M se distribuyeron principalmente entre 18.2-20.1 m, mientras que las H y HG se localizaron entre los 16.4 y 20.1 m en la 95.
Fig. 3. Distribución y abundancia de Urotrygon nana. R. leuchorhyncus se encontró en la subzona 95 con el 60.5% con mayor presencia de NN, H (16.1%), M (12.9%) y HG (6.5%) a una profundidad de 28.8-30.8 m. R. steindachneri se distribuyó con mayor frecuencia en la 95 (34%) y 93 (31.6%); NN de ambos sexos (26.3%) entre 27 y 28.8 m. Discusión y conclusiones Penagos García et al. (2011) reporta 11 especies de rayas para la región Soconusco, Chiapas, encontrando mayor abundancia de R. glaucostigma y D. longa. U. chilensis presentó cambios en la distribución de acuerdo a su madurez sexual, los A de ambos sexos y HG se capturaron frente al sistema lagunar Huave, coincidiendo con la zona de mayor incidencia durante mayo (veda de tiburones y rayas en el Pacífico mexicano, DOF 2012). Los A, J, NN y HG de N. vermiculatus se capturaron frente al sistema Huave, comportamiento en su distribución que podría
deberse a etapa de vida, sexo y hora del día (Drymon et al. 2011). La mayor abundancia para U. nana se encontró frente al sistema Huave y a la desembocadura del Río Suchiate a profundidades menores a 20 m, zona donde especies del género Urotygon están mejor representadas. Así mismo, R. leucorhynchus y R. steindachneri encontraron mayor incidencia de NN en la 95, zona que está altamente influenciada por la desembocadura del sistemas lagunar Chantuto-Panzacola, lo que indica que juega un papel importante en la alimentación, protección y crianza de estas especies (Tapia-García et al. 1997). Literatura citada Cervantes-Hernández P, A Egremy-Valdez. 2013.
Variación de la captura del recurso camarón, en relación con la disposición geográfica de los sedimentos en la zona 90 de pesca. Golfo de Tehuantepec, Oaxaca, México. Hidrobiológica 23(1): 111-123.
Drymon JM, SP Powers, J Dino, B Dzwonkowski, TA Henwood. 2010. Distributions of sharks across a continental shelf in the northern gulf of Mexico. Marine and coastal fisheries: Dynamics, management, and ecosystem science. 2: 440-450.
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro oriental. Vol. 2: 745-792.
Núñez-Orozco AL, A Labastida-Che, JA Oviedo-Piamonte. 2013. Composición y abundancia de la ictiofauna en la franja sublitoral del Golfo de Tehuantepec, Oaxaca/Chiapas, México. Ciencia Pesquera 21(2): 29-40.
Penagos-García F.E., O. Lam-Gordillo, G. Rivera-Velázquez, M. Tapia Garcia y G. Juárez Hernández. 2011. Rayas (Piscis: Batoidea) de la plataforma continental de la región soconusco, Chiapas, Mexico. Lacandonia Año 5(1): 43-51.
Stehmann MFW. 2002. Proposal of a maturity stages scale for oviparous and viviparous cartilaginous fishes (Pisces, Chondrichthyes). Archive Fishery Marine Research 50(1): 23-48.
Tapia-García M. 1997. Diversidad dinámica y patrones reproductivos en la comunidad de peces demersales del Golfo de Tehuantepec. Universidad Autónoma Metropolitana. Unidad Iztapalapa Informe final SNIB-CONABIO proyecto No. B094. México, D.F.
Palabras claves: Urotrygon, rayas, Pacífico Sur, pesquería.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
191
092: cartel IMPORTANCIA DEL NOMBRE CIENTÍFICO VÁLIDO EN PESQUERÍAS
Leticia Huidobro Campos1 y Elaine Espino Barr2
1Dirección General Adjunta de Investigación Pesquera en el Atlántico. INAPESCA. Pitágoras 1320. Col. Santa Cruz Atoyac, Ciudad de México. C.P. 03310; [email protected]; 2Centro Regional de Investigaciones Pesqueras de Manzanillo. INAPESCA. Playa Ventanas s/n., Manzanillo. Colima. CP 28200. [email protected] Introducción Es común que en trabajos pesqueros o acuícolas se identifique a los recursos con nombres comunes, porque durante la obtención y registro de datos, ya sea en las oficinas de pesca o directamente con los pescadores, la información proporcionada es con nombre común. En el quehacer de los investigadores la primera tarea es asignar el nombre científico a cada recurso, para ello utilizan las claves taxonómicas del grupo o de la región de estudio. Es sabido que la taxonomía continuamente está actualizándose y las modificaciones de la nomenclatura no siempre están disponibles y al alcance de los investigadores, por lo que se suelen encontrar en publicaciones nombres que ya no son válidos o que están asignados a otros taxones, como es el caso de Gerres cinereus, que durante mucho tiempo se utilizó este nombre para la mojarra rayada (Miller 1966, Allen y Robertson 1998, Hoese y Moore 1998, McEachran y Fechhelm 2005, Amezcua 2009), incluso en ambientes estuarino-lagunares y dulceacuícolas también ha sido considerada como anfiamericana (Lozano-Vilano y Contreras-Balderas 1978, Castro-Aguirre et al. 1999, Miller et al. 2005). En trabajos pesqueros como la Guía FAO para la identificación de especies para los fines de la pesca (Fischer et al. 1995), la Carta Nacional Pesquera (DOF 2012), entre otros. El taxón que se conocía como G. cinereus fue descrito por Walbaum en 1792, en una localidad en las Bahamas. Durante mucho tiempo fue reconocido para ambos litorales, tanto en ambientes marinos como estuarinos (Regan 1906-1908, Castro-Aguirre et al. 1999, Miller et al. 2005). Sin embargo, existía la posibilidad de que fueran dos entidades taxonómicas diferentes. Ya Burnes en 2009 había identificado el problema y mediante un estudio morfométrico realizado en ejemplares tanto de la costa Atlántica como Pacífica del litoral mexicano, encontró dos poblaciones morfológicamente distintas las cuales podrían corresponder una de ellas a G. cinereus (costa Atlántica) y G. simillimus para la costa Pacífica. Sin embargo, aun cuando sus resultados corroboran la hipótesis de especies diferentes, ésta no podía considerarse como una publicación formal como el propio autor lo menciona, ya que es información inédita de una tesis de grado, y no podía ser reconocida en la literatura ictiológica.
Años después, Angulo et al. (2013) identifican a ambas especies en las costas de Costa Rica, como parte de los peces nativos de este país centroamericano, por lo que las reconocen como especies válidas. En 2014 Vergara-Solana y colaboradores en un estudio molecular y morfológico del género Diapterus, mencionan que en el grupo de los gérreidos existe una gran cantidad de taxones para los que es cuestionable su validez y que necesitan ser revisados a los que se les conoce como species inquirendae (ICZN 2016), entre ellos G. simillimus. Ambas publicaciones sustentan la validez de G. cinereus y G. simillimus en el Catálogo de Peces de la Academia de Ciencias de California (Eschmeyer et al. 2016), la referencia autorizada para los nombres taxonómicos de peces, que ofrece además una base de datos en línea. Fishbase (Froese y Pauly 2016) es otra página comúnmente utilizada para identificar especies, en la cual se reconocen ambas especies, cada una para una vertiente. Este es uno entre muchos casos de especies inquerendae (difíciles de identificar), y su problema radica en que si se consideran como una sola especie, cuando pueden ser dos o más, se ocultan o camuflajean las características propias de cada una de ellas. En cuestiones pesqueras es importante tener la seguridad de la identidad del recurso, pues se involucran aspectos biológicos clave y específicos necesarios para su manejo como la edad y longitud de primera madurez, de primera captura, la relación peso-longitud, los parámetros de crecimiento y mortalidad, etc. Aspectos de cada especie, importantes para definir los Puntos de Referencia: elementos de orden técnico que sirven para orientar el manejo, referidos ya sea a esfuerzo, captura, o tasa de mortalidad o épocas de veda, aplicada a la pesca sobre el determinado recurso (DOF 2012). Este trabajo no pretende ser una revisión taxonómica de la especie, sino intenta resaltar la importancia de utilizar el nombre válido para los distintos recursos pesqueros.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
192
Fig. 1. Gerres cinereus (Walbaum 1792), Vertiente Atlántica.
Fig. 2. Gerres simillimus Regan 1907, Vertiente Pacífica. Literatura citada Allen GR, DR Robertson. 1994. Peces del Pacífico Oriental
Tropical. CONABIO/Agrupación Sierra Madre/ CEMEX. Ciudad de México. 327p.
Amezcua LF. 2009. Peces demersales del Pacífico de México. Instituto de Ciencias del Mar y Limnología, UNAM. 281p.
Angulo A, CA Garita-Alvarado, WA Bussing, MI López. 2013. Annotated checklist of the freshwater fishes of continental and insular Costa Rica: Additions and nomenclatural revisions. Check List 9(5): 987-1019.
Burnes RLA. 2009. Estatus taxonómico de Gerres cinereus (Walbaum, 1792), (Teleostei: Gerreidae). Tesis de Maestría. CICIMAR-IPN. 98p.
Castro-Aguirre JL, H Espinosa P, JJ Schmitter-Soto. 1999. Ictiofauna estuarina, lagunar y vicaria de México. Editorial Limusa-Noriega. México.
DOF. 2012. Acuerdo por el que se da a conocer la Actualización de la Carta Nacional Pesquera. Diario Oficial de la Federación. México. 24 de agosto de 2012.
Eschmeyer WN, R Fricke, R van der Laan (eds.). 2016. Catalog of Fishes: Genera, Species, References. Versión electrónica consultada 17 junio 2016. (http://researcharchive.calacademy.org/rese arch/ichthyology/catalog/fishcatmain.asp).
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca.
Pacífico centro-oriental. Vol II. Vertebrados Parte 1: 647-1200.
Froese R, D Pauly (eds). 2016. FishBase. Publicación electrónica de la World Wide Web. www.fishbase.org, versión (01/2016).
Hoese HD, RH Moore. 1998. Fishes of the Gulf of Mexico, Texas, Louisiana, and Adjacent Waters. Texas A&M University Press. 422p.
Lozano-Vilano ML, S Contreras-Balderas. 1978. Lista zoogeográfica y ecológica de la ictiofauna continental de Chiapas, México. Southw. Nat. 32(2): 223:236.
ICZN. 2016. International Commission on Zoological Nomenclature. International Code of Zoological Nomenclature. Consultado 17 junio de 2016. http://www.nhm.ac.uk/hosted-sites/iczn/code/index.jsp
McEachran JD, JD Fechhelm. 2005. Fishes of the Gulf of México. Vol. 2: Scorpaeniformes to Tetraodontiformes. University of Texas Press, Austin. 1004p.
Miller RR. 1966. Geographical distribution of Central American freshwater fishes. Copeia (4): 773-802.
Miller RR, WL Minckley, ST Norris. 2005. Freshwater Fishes of México. University of Chicago Press. Chicago.
Regan CT. 1906-1908. Pisces. In: FD Godman y O Salvin (eds.). Biologia Centrali Americana. Vol 8: 201p.
Vergara-Solana FJ, FJ García-Rodriguez, JJ Tavera, E De Luna, J De La Cruz-Agüero. 2014. Molecular and morphometric systematics of Diapterus (Perciformes, Gerreidae). Zoologica Scripta 43(4): 338-350.
Palabras clave: Recurso pesquero, taxon, especies
inquerendae

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
193
093: oral LOS PECES SIERRA EN MÉXICO:
PASADO GLORIOSO Y FUTURO ALARMANTE DE DOS GIGANTES MARINOS
Ramón Bonfil1, Oscar Uriel Mendoza Vargas2, Melina Ricaño Soriano3, Paola Yanira Palacios Barreto4 y Nataly Bolaño Martínez4
1Océanos Vivientes A.C. Cda. Monserrat 9. La Candelaria-Coyoacán. Ciudad de México. C. P. 04380. [email protected]. 2Posgrado en Biología. UNAM. [email protected]. 3Revolución núm. 9, Sector Pajaritos, Tihuatlán, Veracruz, México. [email protected] 4Laboratorio de Genética de Organismos Acuáticos. Instituto de Ciencias del Mar y Limnología. UNAM. Circuito Exterior S/N. Ciudad Universitaria. Ciudad de México. C. P. 04510. [email protected] [email protected] Introducción Los peces sierra (Familia Pristidae) son el grupo de peces más amenazados con extinción a nivel mundial: 3 de las 5 especies que existen están clasificadas como en Peligro Crítico (CR) por la IUCN y las otras dos como en Peligro (EN) (Dulvy et al. 2014). En México las dos especies nativas, el pez sierra de dientes pequeños, Pristis pectinata y el pez sierra de dientes grandes P. pristis, están protegidas por la NOM-029-PESC-2006 y la NOM-059-SEMARNAT-2010. Ambas especies están consideradas como en Peligro Crítico (CR) por la IUCN a nivel global y como Amenazadas (A) en México por la CONANP. A la fecha no existe un solo estudio específico que indique cuáles son la distribución y abundancia de ninguna de las dos especies en México; sólo existen reportes sobre su presencia, en listas faunísticas y catálogos de especies (Castro-Aguirre y Espinosa-Pérez 1996). Por lo tanto se desconoce cuál es el estado actual de conservación de las especies.
Fig. 1. Pristis pectinata (arriba) y P. pristis (abajo) Durante 2015, Océanos Vivientes AC llevó a cabo el proyecto “Evaluación de la situación actual de los peces sierra en εéxico” (PROCER/DGOR/03/2015) cuyos objetivos fueron: a) conocer la situación actual de las poblaciones de Pristis pectinata y P. pristis en México, b) contar con información actualizada sobre
la abundancia relativa y distribución de ambas especies en México; c) identificar posibles sitios de interés para la conservación de estas especies, y d) evaluar un posible cambio de categoría de riesgo en la próxima revisión de la NOM-059-SEMARNAT-2010. Material y métodos Entre el 26 de agosto y el 20 de noviembre de 2015, se realizaron 8 viajes de campo visitando 71 localidades distribuidas en ambos litorales del país y 13 estados de la República, desde Sinaloa hasta Chiapas, y desde Tamaulipas hasta Quintana Roo. Durante el trabajo de campo se recorrieron más de 10,500 km por carretera y se entrevistaron a más de 800 personas. Usando un formato diseñado específicamente para este proyecto, se realizaron 245 encuestas a personas (principalmente pescadores) que habían visto o pescado peces sierra personalmente. Así mismo, se dieron pláticas audiovisuales de educación ambiental e información sobre el proyecto en varias de las comunidades pesqueras visitadas y se repartieron carteles informativos del proyecto llamando a reportar avistamientos de peces sierra. También se creó una página en FaceBook para difundir el proyecto, tener mayor cobertura, y captar más reportes sobre avistamientos de peces sierra. Resultados y discusión El presente trabajo constituye el primer y hasta la fecha único estudio dedicado a los peces sierra en México y establece la primera línea de base sobre la abundancia y distribución históricas de ambas especies. Los encuestados fueron en su mayoría pescadores de 50 a 75 años de edad y con edad media de 62 años. Las generaciones más jóvenes de pescadores no conocieron a los peces sierra, lo cual es un índice del deterioro de sus poblaciones desde la década de los sesenta. Los datos derivados de las encuestas indican que el pez sierra de dientes grandes P. pristis era común en todos los estados del Pacífico mexicano de Sinaloa al sur, y muy abundante en Nayarit, Colima, Michoacán y Chiapas. En el Golfo de México y Mar Caribe el pez sierra de dientes pequeños P. pectinata era de común a

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
194
abundante desde Tamaulipas hasta Quintana Roo y era especialmente abundante en Tabasco y Quintana Roo. Por su lado, en el litoral del Golfo de México y Mar Caribe, P. pristis era muy común en todo el litoral y extremadamente abundante en Veracruz, Tabasco, Campeche y Quintana Roo y específicamente en localidades como Laguna de Términos, el Río Usumacinta y sus afluentes, y la Bahía de Chetumal. En la actualidad, tanto la amplitud de distribución como la abundancia de ambas especies en todo el territorio nacional se ha colapsado alarmantemente y son iguales o muy cercanas a cero. Durante las encuestas no se recibieron reportes de la existencia de agregaciones o zonas de crianza de ninguna de las dos especies en el país. Hasta antes de 2016, el último reporte comprobado de P. pristis en el Pacífico fue en 2010 en Michoacán. Los últimos reportes comprobados de P. pectinata y P. pristis en el Golfo de México y Mar Caribe fueron en Tamaulipas entre 2005 y 2009 para la primera, y en Quintana Roo en 1998 para la segunda. Un ejemplar vivo de P. pectinata fue pescado en enero de 2016 en Barra de Cazones, Ver. y reportado al proyecto. Este es el único pez sierra vivo del que se tiene conocimiento en México hasta el momento. Una ligera mayoría de los entrevistados consideraban a los peces sierra como de poca importancia, pero todos o casi todos los desembarcaban y utilizaban. Quienes los consideraban de valor o importancia, reportaron al valor económico como la principal razón de su importancia, seguido por el uso de los rostros como trofeo, y la utilización para consumo personal. Varias personas respondieron que la carne del pez sierra era muy apreciada por ellos por su buen sabor y calidad. Nuestros datos indican que antes de su desaparición, ambas especies fueron pescadas frecuentemente de manera incidental aunque existieron pesquerías dirigidas puntuales. Las redes de enmalle de todo tipo fueron la principal causa del desplome de las poblaciones de peces sierra en México, pero ambas especies fueron y pueden ser fácilmente capturadas en todo tipo de artes de pesca, debido a su morfología y hábitos costeros. Actualmente las principales amenazas para la conservación y la recuperación de los peces sierra son en primer lugar la enorme cantidad de redes para la pesca que existen en zonas someras de la costa y principalmente en las desembocaduras de los ríos, así como en esteros, estuarios, lagunas costeras y bahías interiores, y en segundo lugar la pérdida, degradación y fragmentación de los manglares, pues estos representan hábitat esencial para la reproducción (avivamiento y crianza) de ambas especies.
Es extremadamente urgente avocar recursos suficientes de inmediato, durante varios años, para realizar la búsqueda directa de especímenes de peces sierra de dientes pequeños y de dientes grandes en zonas con mayor probabilidad de que aún tengan agregaciones pequeñas, criaderos, o individuos aislados de estas especies. Dichos esfuerzos deberán tener un fuerte componente de educación ambiental en las comunidades costeras para garantizar la supervivencia de las especies. Así mismo se debe difundir el grave problema de extinción local de estas especies entre el público en general y se debe cambiar la categoría de riesgo de ambas especies en la NOM-059-SEMARNAT-2010. Literatura citada Castro-Aguirre JL, H Espinoza-Pérez. 1996. Catálogo
sistemático de las rayas y especies afines de Mxico: Chondrichthyes: Elasmobranchii: Rajiformes: Batoideiomorpha. Listados faunísticos de εéxico. Vol. 7. Instituto de Biología. UNAε.
Dulvy NK, LNK Davidson, PM Kyne, CA Simpfendorfer, LR Harrison, JK Carlson, SV Fordham. 2014. Ghosts of the coast: global extinction risk and conservation of sawfishes. Aquatic Conserv: Mar. Freshw. Ecosyst. DOI: 10.1002/aqc.2525
Palabras Clave: Peces sierra, Pristis pectinata, Pristis pristis, extirpación, pesca incidental, conservación.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
195
094: cartel PERCEPCIÓN DE LOS PESCADORES SOBRE LA MANTA OCEÁNICA (Manta birostris)
EN YELAPA, JALISCO
Ana Cecilia González Sánchez¹, Adrián Maldonado Gasca¹, Javier Tovar Ávila² y Joshua Stewart³
¹Instituto Tecnológico Bahía de Banderas. Crucero a Punta Mita s/n, CP. 63734 La Cruz de Huanacaxtle, Nayarit. ²Centro Regional de Investigación Pesqueras de Bahía de Banderas. INAPESCA. Calle Tortuga núm. 1, La Cruz de Huanacaxtle, Nayarit. C.P. 63732. ³Scripps Institution of Oceanography 9500, Gilman Drive, La Jolla, California, USA. 92093. [email protected] Introducción Las mantas son un grupo de elasmobranquios que han adquirido un estilo de vida pelágico, basada en una alimentación por filtración similar a las ballenas y tiburón ballena. Pertenecen a la familia de las rayas águila (Myliobatidae), sin embargo forman su propia subfamilia, Mobulidae, que incluye dos Géneros Manta Bancroft, 1828 con dos especies y nueve del Género Mobula Rafinesque, 1810 (Marshall 2008). La subfamilia Mobulidae incluye a las mantas gigantes, alcanzando los siete metros de ancho de disco. Manta birostris es la especie más grande de los batoideos y uno de los peces vivientes más grandes también. Todas las especies de la subfamilia Mobulidae son filtradoras, aunque consumen también peces pequeños como presas ocasionales (Fischer et al. 1995). Los adultos de M. birostris se pueden reconocer fácilmente ya que presentan aletas pectorales largas y triangulares. A diferencia de varios mobúlidos (con boca subterminal), la boca de la manta es terminal. Es de suma importancia conocer la retrospectiva histórica de la especie Manta birostris en Yelapa, Jalisco, ya que además de ser una especie que abunda esa zona, no se tiene registro de este tipo de información y conocer la historia de una relación entre pueblo y población de mantas es primordial para entender la situación actual de estos animales. Es necesario conocer la ecología y biología básica de las especies para su adecuada conservación y manejo. Para los elasmobranquios, que son de crecimiento lento, lenta maduración y baja fecundidad (Holden 1974), es importante entender cómo las poblaciones son afectadas por impactos antropogénicos, ya que es menos probable que se recuperen de una disminución en su población (Hoenig y Gruber 1990). Material y métodos Se aplicaron encuestas durante septiembre y noviembre de 2014 en Yelapa, Jalisco; dentro del municipio de Cabo Corrientes. Los cuestionarios impresos fueron aplicados cara a cara a 25 personas de distintas edades, desde pescadores retirados hasta jóvenes que comienzan su vida como pescador. Los encuestados se dividieron en dos
grupos: pescadores frecuentes y pescadores ocasionales, donde los frecuentes se refiere a los pescadores que viven de la pesca y es su actividad diaria, laboral. Los pescadores ocasionales son aquellos que realizan otras actividades para su sustento, sin embargo la pesca forma parte de sus vidas y de alguna manera su contacto con el mar es útil para la obtención de información. Dentro de los encuestados se incluyeron pescadores, amas de casa, restauranteros, marineros y capitanes de embarcaciones. Esta encuesta tenía como objetivo conocer la perspectiva de los habitantes del pueblo sobre las mantarrayas gigantes, al mismo tiempo se pretende también conocer la situación de las poblaciones de mantas, tanto en el pasado como la actual. El formato de esta encuesta fue diseñado a partir de encuestas aplicadas por el Programa Tiburón del Centro Regional de Investigaciones Pesqueras de Bahía Banderas, enfocándose a la obtención de información de la manta gigante en Yelapa, Jalisco. Para el análisis de los resultados se realizó la prueba de χ2 para la comparación de dos variables. Se utilizó el programa Excel 2013 y PAST para su posterior comprobación. Resultados y discusión De las 25 encuestas aplicadas en Yelapa, Jalisco 22 fueron hombres y tres mujeres. Hubo diversidad en las edades de las personas, el grupo sobresaliente fue de las personas entre 20 y 30 años representado con 24%. La mayoría eran nacidos en Yelapa, le seguía Puerto Vallarta y después Chacala, Nayarit. El 60% de los encuestados dijeron que la pesca de mantas es ilegal, 24% no sabía y 16% aseguraron que es legal pescar esta especie. El 80% de encuestados aseguran que la temporada de mantas es la temporada de invierno, y los lugares donde más tránsito de mantas han visto por orden fueron: Pizota (43%), Punta del Cabo (20%), Marietas (17%), Yelapa (11%) y Quimixto (6%), mientras que 3% de personas no contestaron esta pregunta. El 56% de los pescadores dijeron que la población de mantas sí está cambiando y no precisamente para bien, lo cual es un dato preocupante, las personas que contestaron que no había cambios en la

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
196
población fueron 40%, 4% de pescadores no contestó esta pregunta. Algo interesante dentro de los resultados obtenidos es que 44% no sabía el motivo del por qué la población de mantas está cambiando/disminuyendo en la zona de Yelapa. Seis personas (24%) aseguraron que las mantas son menos ahora debido al tránsito de pangas, y cuatro personas (16%) dijeron que por el uso de chinchorros. Conclusiones El pueblo en general desconoce lo que es una manta raya gigante, a pesar de estar tan cerca de ellas, no están informados sobre la especie y por este motivo les causa gran temor. Esto se debe a que anteriormente, cuando usaban embarcaciones más pequeñas tipo canoa, las mantas llegaban a hundir muchas y también arrastraban los cabos de las pangas actuales y las mangueras de las personas que bucean con compresor. En cuanto a la distribución y abundancia de M. birostris en la zona, se concluyó que enero es el mes en que se ven más, y el lugar donde más mantas se avistan en es Pizota. Inevitablemente la población de mantas está disminuyendo notablemente; un poco más de la mitad afirmó esto en las encuestas. La retrospectiva histórica de esta situación indica que debido a varios factores donde predominan el tránsito de pangas y el uso de chichorros, las mantas están en decadencia como especie. Se logró compartir la información del valor de la manta tanto ecológica como económica, sembrando en el pueblo la idea de cuidar la especie para poder realizar algún día el ecoturismo controlado y de esta manera generar un derrame de dinero para el pueblo. Se concluye que al igual que muchas otras especies, la manta gigante vale más viva que muerta. Literatura citada Fischer W, F Krupp, W Schneider, C Sommer, KE
Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca, Pacífico Centro-Oriental. Vol. II. Vertebrados, parte 1, Roma. 1200p.
Hoenig J, Gruber S. 1990. Life-history patterns in the elasmobranchs: implications for fisheries management. NOAA Tech Rep NMFS 90:1–16.
Holden MJ. 1974. Problems in the rational exploitation of elasmobranch populations and some suggested solutions. In: Jones FRH (ed) Sea fisheries research. Wiley, New York, NY, p 117–137.
IUCN. 2008. - http://www.iucnredlist.org Marshall A. 2008. Biology and Population Ecology of Manta
Birostris in Southern Mozambique. Tesis de Doctorado. University de Queensland. Australia. 306 p.
Palabras Clave: mantarraya, manta gigante, elasmobranquios, Mobulidae, Bahía de Banderas.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
197
095: cartel EL OSTIÓN DE ROCA Striostrea prismatica EN EL PACÍFICO CENTRAL MEXICANO
Karla G. Ríos González1,2, Ernesto López Uriarte1, Fernando Vega Villasante2 y Elaine Espino Barr3
1Laboratorio de Ecosistemas Marinos y Acuicultura. Departamento de Ecología. Centro Universitario de Ciencias Biológicas y Agropecuarias. U.de G. Camino Ramón Padilla Sánchez núm. 2100, Nextipac, Zapopan, Jalisco, México. C.P. 45110. [email protected] [email protected]; 2Laboratorio de Acuicultura Experimental. Centro de Investigaciones Costeras. U. de G. Av Universidad 203, Delegación Ixtapa, Puerto Vallarta, Jalisco, México. C.P. 48280. [email protected]. 3Centro Regional de Investigaciones Pesqueras de Manzanillo. INAPESCA. Playa Ventanas s/n, Manzanillo, Colima, México. C.P. 28200. [email protected] Introducción Striostrea prismatica (Gray 1825) es un molusco bivalvo que habita el litoral rocoso de la zona de entre mareas hasta el sublitoral somero. Su distribución geográfica de norte a sur, va desde La Paz, Baja California Sur (24.2°N) y Mazatlán, Sinaloa (23.2°N) en México hasta Máncora, Tumbes, Perú (4.1°S) (Keen 1971, Coan y Valentich 2012). En países como El Salvador, Costa Rica, Perú y México la densidad relativa de S. prismatica en bancos naturales oscila de 0.2 a 176 org/m2 y una estructura de longitudes que va de 2 a 15 cm de altura total (Campos y Fournier 1989, Ordinola 2010), con claros signos de sobre-explotación del recurso y uso de diversas estrategias de manejo y aprovechamiento. La veda temporal de dos años (2010-2012) a la captura de este ostión por pescadores de la costa de Jalisco, México, dio resultados ligeramente satisfactorios. El presente trabajo evalúa el estado actual de varios bancos naturales del ostión de roca en el Pacífico central mexicano. Material y métodos De abril 2014 a agosto 2015 se realizó el registro mensual de ostión de roca en bancos naturales de tres localidades del Pacífico central mexicano (PCM), mediante buceo libre y SCUBA. La abundancia fue estimada empleando el método de las distancias propuesto por Gallucci et al. (1996), que consiste en el registro de organismos en un transecto de 50 m de longitud dispuesto al azar (área de 100 m2). Para determinar la estructura de longitudes de la población se registró la altura total de la concha por organismo, utilizando un vernier al mm más cercano. Se consideró el muestreo organismos provenientes de la captura comercial de dos localidades de estudio, en las que se obtuvo el registro de la altura, longitud y anchos totales, peso total y de la parte blanda del organismo. La distribución se determinó por el método de Morisita publicado en Sakai et al. (1999). El tipo de crecimiento por medio de la fórmula PT=a*LTb. El índice de condición Kn=Ps/Pf*100.
Resultados y discusión Se obtuvieron 3,155 organismos en 3 localidades de muestreo y 2 métodos de muestreo. La densidad relativa promedio fue 0.52 ind/m² (d.e. ± 49.45); en el banco Tizate se encontró 0.77 ind/m2 (± 26.78), Victoria con 0.47 ind/m2 (± 41.8)1 y Palmito con 0.38 ind/m2 (± 12.93) (Tabla 1). La prueba múltiple de Tukey demostró diferencia en densidad entre el banco Victoria y Palmito (Fig. 1). Estas densidades fueron inferiores a lo reportado por Campos y Fournier (1989) de 134.26 org/m2 en Costa Rica con actividad extractiva incipiente, y 12 org/m2 en bancos del Perú. Tabla 1. Resumen de los organismos registrados y capturados en las localidades de estudio en el PCM.
Localidad Área (m²) y núm.
transecto
Registro in situ
Captura comercial
Organismos total
Tizate 1000 (10) 696 713 1409
Victoria 700 (7) 536 814 1350
Palmito 1000 (10) 396 0 396
Total 2700 (27) 1,628 1,527 3,155
Fig. 1. Distribución de la longitud altura total de los organismo registrados en las localidades del PCM. Por el contrario resultan mayor a lo reportado por Ordinola (2010) en su evaluación del recurso ostión en la costa de Perú, de 0.2 org/m2. La variación en la densidad de los bancos de ostión del presente estudio, están relacionadas a las diferencias en el tipo de fondo, dinámica de corrientes, grado de exposición y disponibilidad de alimento entre los factores ambientales y biológicos que interactúan en

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
198
las áreas de estudio. Mientras que en los de origen antrópicos es la presión por pesca y captura furtiva. La estructura de longitudes fue de 11 a 155 mm (d.e. ± 22.55), la longitud máxima y mínima se registró en Tizate. Las poblaciones en los tres bancos son polimodales, con dos a cinco modas (Tizate: tres modas 61-70, 71-80 y 81-90 mm; Victoria: dos modas 41-50 y 61-70 mm; Palmito: dos modas de 31-40 y 41-50 mm). La relación longitud-peso se obtuvo para 1,519 organismos, con una altura de concha de 12 a 155 mm y peso total de 0.45 a 472g. El valor de la pendiente es de un crecimiento alométrico negativo b˂3 (Fig. 2).
Fig. 2. Relación altura total y peso total de los organismos colectados en Tizate y Victoria del PCM. También hubo alometría negativa en la boca del Golfo de California y con otras especies del género Crassostrea (Kobayashi et al. 1997, Chávez-Villalba et al. 2005, Vidal-Briseño et al. 2015). El índice de condición fue máximo en abril y mayo (2014–2015), evidenciando incremento en la actividad reproductiva. Este resultado coincide con Cuevas y Martínez (1979) en la boca del Golfo de California, Páez-Osuna (1993) en Sinaloa y Meléndez-Galicia et al. (2015) en la costa de Michoacán. Los lípidos y las proteínas se encuentran en su máximo pico en abril y mayo, al igual que la madurez y la gametogénesis. Conclusiones Las tres localidades muestran densidades muy bajas comparadas con bancos sin explotación. Las longitudes son menores a la estipulada en la Carta Nacional Pesquera, lo que muestra una presión por pesca alta, por lo que se considera imperativo establecer época de veda y un programa de rotación de bancos, así como un límite de extracción anual, para permitir la recuperación de los bancos y hacer de esta actividad una pesquería sustentable. Literatura citada Campos JA, ML Fournier. 1989. El banco de Ostrea
iridescens (Pterioida: Ostreidae) de Bahía Curú, Costa Rica. Revista de Biología Tropical 38(2A): 331-333.
Chávez-Villalba J, M. López, J Mazón-Suástegui, M Robles-Mungaray. 2005. Growth of the oyster
Crassostrea corteziensis (Hertlein, 1951) in Sonora, México. Acuaculture Research 36: 1337-1334.
Coan EV, P Valentich. 2012. Bivalve seashells of tropical West America. Marine bivalve mollusks from Baja California to northern Peru. 2 vols, pp 1258.
Cuevas CA, A Martínez. 1979. Estudio gonádico de Crassostrea corteziensis Hertlein 1951, C. palmula Carpenter y S. prismatica Hanley, de San Blas, Nayarit, México (Bivalvia: Ostreidae). Anales del Instituto de Ciencias del Mar y Limnología 6: 81–98.
Gallucci VF, SB Saila, DJ Gustafson, BJ Rothschild. 1996. Stock assessment. Quantitative methods and applications for small-scale fisheries. 527 pp. CRC Press Lewis Publishers, Boca Ratón.
Keen AM. 1971. Sea Shells of Tropical WestAmerica Marine Mollusks from Baja California to Peru. Stanford University Press, California; pp, 1604
Kobayashi M, E Hoffmann, EN Powell, K Kusaka. 1997. A population dynamics model for the Japanese oyster, Crassostrea gigas. Aquaculture 1997: 285-321.
Meléndez-Galicia C, F Estrada, V Hernández, A Arellano, D Hernández. 2015. Madurez gonádica del ostión de roca Crassostrea iridescens, en la costa de Michoacán, México. Ciencia Pesquera 23(1): 25-36.
Ordinola E. 2010. La ostra Crassostrea iridescens (Hanley) en Tumbes, Perú. Primavera 2007. Repositorio Digital IMARPE Informe 37:139-150.
Páez-Osuna F, HM Zazueta, JI Osuna-López. 1993. Biochemical composition of oysters Crassostrea iridescens (Hanley) and C. corteziensis (Hertlein) in the Northwest coast of México: seasonal changes. J Expl Mar Biol and Ecol 170: 1-19.
Sakai S, K Momose, H Nagamitsu, AA Hamida, T Nakashizuka. 1999. Plant reproductive phenology over four years including an episode of general flowering in a lowland dipterocarp forest, Sarawaka, Malaysia. Am. J. Of Bot. 86: 1414-1436.
Vidal-Briseño C, RI Hernández, G Galido, P San Martín del Ángel. 2015. Estructura de la poblacional de la captura del ostión Crassostrea virginica en las lagunas de Tamiahua y Tampamochoco, Veracruz México. Hidrobiológica 25(2): 265-173.
Palabras clave: ostión, abundancia, estructura, densidad, Pacífico.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
199
096: cartel PRIMER REGISTRO DE Taractichthys steindachneri CAPTURADO FRENTE A LA PLATAFORMA
CONTINENTAL DEL SUR DE SINALOA, MÉXICO
María Candelaria Valdez Pineda2, Luis Antonio Salcido Guevara2, Jorge Saúl Ramírez Pérez2 y Rebeca Sánchez Cárdenas1
1CONACyT–2Facultad de Ciencias del Mar. Universidad Autónoma de Sinaloa. Paseo Claussen s/n, Col. Los Pinos, Mazatlán, Sinaloa, México. C. P. 82000. [email protected], [email protected], [email protected], [email protected] Introducción En los últimos 55 años, a medida que la pesca de todas las especies ha tenido rendimientos cada vez menores, las flotas comerciales están irrumpiendo en zonas cada vez más profundas de los océanos. En México, es escaso el conocimiento disponible sobre los recursos pesqueros con potencial de explotación, que habitan más allá de la plataforma continental (>200 m de profundidad). Esporádicamente, algunas especies de hábitos mesopelágicos y batipelágicos son capturadas por las flotas comerciales, dándonos un indicio de su existencia y del posible potencial pesquero en aguas profundas. Este trabajo presenta el primer reporte de Taractichthys steindachneri (Döderlein 1883) (Perciformes: Bramidae) en el borde de la plataforma continental de Sinaloa, México, así como la descripción de algunos aspectos anatómicos para comprender su presencia y parte de su historia de vida. Material y métodos Se realizó la identificación de un ejemplar de un pez desconocido capturado por los pescadores de la empresa “Pesca Centenario” a 130 kilómetros del puerto de Mazatlán. La identificación se realizó mediante las claves dicotómicas de Fischer et al. (1995) y el catálogo de peces de Eschmeyer et al. (2016). Resultados y discusión Nombre científico: Taractichthys steindachneri (Döderlein, 1883) (Fig. 1). Nombre común: Japuta forma de hoz (Sickle pomfret). Familia: Bramidae. Orden: Perciformes. Clase: Actinopterygii (peces de aletas radiadas). El individuo era un adulto, con 75.4 cm de longitud total y 8.66 kg de peso total (Fig. 1). El espécimen presentó un estómago vacío y la gónada se encontraba sin indicios de gametogénesis (en estado de reposo). Lo que indica que no se estaba alimentando, ni en proceso de reproducción al momento de su captura.
Fig. 1. Taractichthys steindachneri (Döderlein 1883). El individuo presentó un estómago alargado, intestino relativamente corto, cinco ciegos pilóricos de gran tamaño y branquiespinas muy separadas, lo que indica que es un organismo altamente carnívoro. La forma del cuerpo del pez (fusiforme-falciforme) y el otolito de pequeño tamaño sugieren que es un individuo de movimiento dinámico en la columna de agua (nadador) (Fig. 2).
Fig. 2. Otolitos sagitta de Taractichthys steindachneri Las japutas son importantes para las pesquerías de algunas regiones, donde son muy abundantes y apetecidos por su excelente carne (Puentes et al. 2001). La especie T. steindachneri se distribuye en el

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
200
Mar Rojo, en el Océano Indo-Pacífico, en el Pacífico oriental central, Islas Hawaii y en el sur de California, Estados Unidos y en el noroeste de México (Villavicencio-Garayzar y Chávez 1986, Byung-Jik et al. 2012, Eschmeyer et al. 2016). Sin embargo el conocimiento biológico de la especie es escaso. Se sabe que habita en el ambiente marino entre 50 y 1,000 metros de profundidad (hábitos batimesopelágicos). El primer reporte de esta especie en aguas mexicanas fue hecho en 1986, a partir de un ejemplar capturado en la Bahía de La Paz, Baja California Sur, el 19 de junio de 1986 (Villavicencio Garayzar y Chávez 1986). Conclusión La presencia de T. steindachneri es un indicio de la existencia de poblaciones en los ambientes batimesopelágicos frente a Sinaloa, México. También es un recordatorio de nuestro escaso conocimiento sobre la fauna mexicana de aguas profundas y una invitación a la exploración de su potencial pesquero. Literatura citada Byung-Jik K, K Jin-Koo, R Jung-Hwa, P Ji-Tae. 2012. First
reliable record of the sickle pomfret, Taractichthys steindachneri (Bramidae: Perciformes) from Korea. Korean Journal of Ichthyology 24(3): 230-233.
Eschmeyer WN, R Fricke, R van der Laan (eds). 2016. Catalog of Fishes: Genera, Species, References. URL: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp.
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico Centro-Oriental. Vol. II. Vertebrados – Parte 1. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Roma, 1200 p.
Puentes V, EA Rubio, LA Zapata. 2001. Primer registro del género Taractes (Pisces: Bramidae) en el Océano Pacífico colombiano. Bol. Invest. Mar. Cost. 30: 207-212.
Villavicencio-Garayzar CJ, H Chávez. 1986. Primer registro en aguas mexicanas de Taractichthys steindachneri (Döderlein) (Pisces: Bramidae). Inv. Mar. CICIMAR. 3(1): 127-132.
Palabras clave: Registro taxonómico, potencial pesquero, Japuta, Bramidae, pez de profundidad.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
201
097: cartel PLAN DE MANEJO PESQUERO DEL ECOSISTEMA RIBEREÑO: ALTATA-PABELLONES
Silvia Margarita Ortiz Gallarza, Tania Guadalupe Romero Leyva y Concepción Enciso Enciso
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Calzada Sábalo-Cerritos S/N. Contiguo a Estero El Yugo, Mazatlán, Sinaloa. México. C.P. 82112. Tel. 01(669)9881255 y 9881256. [email protected] Introducción Misión. Asegurar la utilización equilibrada y la protección del patrimonio natural constituido por los múltiples recursos naturales y los servicios ambientales que proporciona el ecosistema estuarino lagunar Altata-Pabellones A-P, a partir de la ejecución de acciones a través de proyectos y de programas de protección, conservación y restauración, en coordinación con todas las dependencias involucradas de los gobiernos a nivel federal, estatal y municipales; contando con la participación activa y comprometida de todos los actores sociales que están directa e indirectamente involucrados en el uso del mismo. Visión. La visión ideal compartida en consenso por todos los actores que participaron en su elaboración, tiene un enfoque social, ecosistémico y productivo: “el ecosistema se ha recuperado, la calidad ambiental ha mejorado y el aprovechamiento de los recursos se efectúa de manera ordenada. Todos los actores que participan, respetan las disposiciones de manejo establecidas en el PMPAP, -respetan las vedas, cuotas, longitudes mínimas de captura, etc.- La calidad de vida de los pescadores y de sus familias ha mejorado notablemente, existe una red de vigilancia coordinada entre la población, los pescadores y los tres niveles de gobierno, cuya acción conjunta, ha reducido sustancialmente la incidencia de la pesca ilegal. Finalmente, se ha logrado la suma de voluntades y el compromiso de todos los actores involucrados que participan en la toma de decisiones”. Material y métodos El Análisis de Recursos Múltiples ARM “es un enfoque para el análisis a la escala de Paisaje, que integra la información entre recursos naturales múltiples, incluyendo los servicios que proporcionan los ecosistemas y está diseñado para evaluar los impactos y las compensaciones entre el desarrollo y la conservación del paisaje” … [sirve] para sustentar las decisiones de los administradores de los recursos públicos (National Academies of Sciences, Engineering and Medicine, 2016). Los componentes fundamentales de un Análisis de Recursos Múltiples según “The U. S. Geological Survey” (USGS) son: (1) Línea Base: integrada por información sobre el estado actual de los recursos naturales múltiples,
incluidos los servicios del ecosistema. Caracterización. (2) Interrelaciones Funcionales: modelos que describen las interrelaciones entre los recursos coexistentes. [Ciencias geológicas, hidrológicas, biológicas y ecológicas]. Diagnóstico. (3) Escenarios: análisis de la evaluación de los impactos y de las medidas de compensación respecto a los recursos naturales en términos biofísicos y socioeconómicos. [Impactos potenciales y el planteamiento de escenarios alternativos de manejo para los tomadores de decisiones]. Pronóstico. Resultados En la región pesquera del ecosistema lagunar estuarino Altata-Pabellones destacan las actividades efectuadas por los socios activos de 17 cooperativas que operan legalmente, amparados por permisos, quienes se dedican tradicionalmente a la explotación de los recursos pesqueros locales: moluscos, crustáceos (camarones y jaibas) y peces (elasmobranquios [cazones, tiburones y rayas] y de escama). Con base en los avisos de arribo de la Subdelegación de Pesca en Navolato y Culiacán, de 2009 a 2014 los mayores ingresos locales de los pescadores de Altata-Pabellones, se obtuvieron a partir de las capturas de camarón de aguas protegidas, con un ingreso bruto de 451´714,476.69/100 M.N. Las capturas de jaibas $32´097,069.94/100 M.N., peces $17´952,257.36/100 M.N. y moluscos $8´432,073.50/100 M.N., resultaron considerablemente menos significativas en cuanto a las ganancias que generaron en el periodo indicado. En El Castillo se presenta el mayor número de habitantes (3,009) y en El Tetuán Viejo, el menor (7) entre 1990 y 2010. En la mayoría de las localidades hay más hombres que mujeres, como en Altata (+48), El Castillo (+141), El Tetuán Nuevo (+6), El Tetuán Viejo (+1) y las Puentes (+12), mientras que en Las Arenitas, las que predominan son las mujeres (+12). Entre 2000 y 2005 es notable la reducción de la población en cinco de seis localidades pesqueras de A-P y sólo en Las Puentes hubo un ligero crecimiento de 0.33. En 1995-2000 hubo una reducción en Las Arenitas y El Castillo. Entre 2005 y 2010 la población se redujo solo en El Tetuán Nuevo [-0.96]. En general el promedio de hijos nacidos vivos en 2010 varió entre 2.52 y 3.00; en cuanto al porcentaje

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
202
de procedencia de la población, la mayoría de la población ha nacido en la entidad (del 93.11% al 100%) y solo de 2.23% [Las Arenitas] a 6.07% [Las Puentes] provienen de otro sitio. Respecto a los índices y grados de marginación, predominó la calificación de bajo, medio y muy bajo, sobre la de alto; que se presentó sólo en El Tetuán Viejo y en Las Puentes en 2010. La población analfabeta de 15 años y más varió de 3.66% en Altata a 16.67% en El Tetuán Viejo, mientras que población de 15 años o más sin primaria completa varió de 21.5% a 50% en esos mismos poblados, respectivamente. Las viviendas particulares sin excusado fluctuaron de 0% a 14.6% en El Tetuán Viejo y Las Arenitas en dicho orden. Las viviendas particulares sin energía eléctrica variaron entre 0% en los dos Tetuanes y Las Puentes y 3.43% en El Castillo. En cuanto a las viviendas particulares sin agua entubada éstas fueron de 4.82% en Altata a 79.06% en Las Puentes. Las viviendas particulares con piso de tierra mostraron valores de 0% en El Tetuán Viejo a 18.14% en Las Puentes. Mientras que las viviendas particulares sin refrigerador fueron del 0 % en El Tetuán Viejo al 17.72% en Las Puentes. La derechohabiencia en servicios médicos señaló que la mayoría corresponde al Seguro Popular y al IMSS, seguido de la no derechohabiencia, el ISSSTE Federal y Estatal o de la categoría no especificada. El grado de escolaridad promedio varió de 4 en El Tetuán Viejo a 8.39 en Altata. En los poblados restantes, en general fluctuó entre 6.69 [Las Puentes y El Tetuán Nuevo] y 7.60 [El Castillo]. En todas las poblaciones hay niños y adolescentes que posponen su educación hasta una edad mayor a la infancia y adolescencia. Los indicadores de empleo y desempleo señalan que la población económicamente activa fue mayor en El Castillo [1,110], Altata [776) y Las Arenitas [674] y menor en El Tetuán Viejo [4] y El Tetuán Nuevo [125]. La población inactiva también es más alta en El Castillo, incluso mayor que la activa [1,243], donde la población ocupada es de 1,086 y la desocupada de 24. Literatura citada National Academies of Sciences, Engineering and
Medicine. 2016. Integrating Landscape Approaches and Multi-Resource Analysis into Natural Resource Management. Washington, DC: The National Academies Press. doi: 10.17226/21917.]
Palabras clave. Altata, sistema lagunar, políticas pesqueras, Plan de Manejo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
203
098: oral PESCA RIBEREÑA DENTRO DE UN ESQUEMA DE ADMINISTRACIÓN BASADA EN ECOSISTEMAS
José Ignacio Fernández Méndez
Oficinas Centrales. INAPESCA. Pitágoras 1320. 4to. Piso, Col. Sta. Cruz Atoyac, Ciudad de México. C.P. 03310. [email protected] Introducción La degradación de los recursos pesqueros ha sido una constante en las últimas décadas. Ese proceso se da de manera paralela a los cambios globales, sea el climático o en las condiciones de las sociedades humanas. La administración basada en ecosistemas puede ser un complemento importante de los enfoques actuales de gestión de pesquerías existentes, que considere el efecto de esos cambios (NOAA 1998). En aparente contraposición, muchos de los autores que han escrito sobre el tema concluyen que sólo se puede administrar las actividades humanas dentro de un ecosistema, no el ecosistema en sí (p. ej. Hilborn 2007). Sin embargo, el concepto clave es que es necesario adoptar enfoques más globales de administración de recursos. Todos los recursos biológicos utilizados por la humanidad forman parte de un conjunto anidado de ecosistemas locales, regionales y globales. A su vez, las comunidades humanas se encuentran dentro de un sistema similar de economías locales, regionales y globales. Las comunidades obtienen servicios ecosistémicos, que pueden ser definidos como las características, funciones o procesos de un ecosistema que directa (como recursos explotados) o indirectamente contribuyen al bienestar humano (Kubiszewski y Costanza 2012). A su vez, los ecosistemas reciben el impacto de las actividades humanas de explotación, emisión de desechos, modificación de hábitats, etc. Algunos autores (v.gr. Ostrom, 2009) consideran los sistemas aquí mencionados como parte de Sistemas Socio-Ecológicos, o Socioecosistemas. Esta perspectiva integra los ecosistemas con la sociedad humana. Algunos autores (v.gr. Walker y Salt 2006, Resilience Alliance 2010) postulan que el objetivo de la Administración Basada en Ecosistemas debe ser mantener la resiliencia de los subsistemas sociales y ecológicos. La resiliencia es fundamentalmente una propiedad del sistema. Se refiere a la magnitud de los cambios o perturbaciones que un sistema puede experimentar sin cambiar a un estado alternativo que tenga diferentes propiedades estructurales y funcionales que afecte negativamente los servicios ambientales. En contraste con esas visiones integradoras, actualmente la evaluación y administración pesquera de nuestro país tienen una visión monosectorial y, en la mayor parte de las veces, monoespecífica.
En este trabajo se intenta presentar un marco donde insertar la evaluación y administración de pesquerías ribereñas. Material y métodos Con base en un análisis de material bibliográfico relativo a la Administración Basada en Ecosistemas, Análisis de Resiliencia y Análisis de Actores se presenta un esquema conceptual para insertar las pesquerías ribereñas en un marco de Administración Basada en Ecosistemas. Resultados Como primer paso, se identifica y caracteriza al sistema de interés, en en este caso, la pesquería analizada y el sistema local donde opera. En cuanto al ecosistema a nivel local, esta caracterización incluye los límites del sistema, la identificación de recursos/ servicios ambientales y su importancia económica y ecológica y la identificación de procesos y relaciones de importancia ecológica y ambiental. En cuanto a los usuarios, la caracterización incluye análisis de actores (utilizando como base lineamientos como los presentados por Renard 2004). Este análisis caracteriza las relaciones del sector que nos interesa con los demás agentes sociales y económicos locales. El análisis de gobernanza (entendida como las formas en que las funciones públicas se llevan a cabo, los recursos públicos son administrados y se ejercen las facultades de regulación, siguiendo procedimientos como los propuestos en Boesen (2008) da una base para comprender las posibilidades de actuación de esos agentes dentro de marcos institucionales. El análisis de redes sociales, al interior de nuestro grupo de interés, permite (por ejemplo) evaluar la capacidad de auto-organización y desarrollo y cumplimiento de reglas comunes (Bodin y Prell 2011), aspectos importantes en el manejo sustentable de recursos naturales por comunidades (Ostrom 1990). La identificación y mapeo de impactos, así como el análisis de su causa raíz (análogo del enfoque fuerza motriz-presión-estado-impacto-respuesta, McKinney et al. 2011, IAEA 2014) permiten identificar y caracterizar los factores (naturales y antropogénicos) que resultan en impactos negativos sobre el ecosistema. Este análisis puede incluir factores no sólo en el nivel local sino también en los niveles regional y nacional/global. Dada la naturaleza diversa

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
204
de los orígenes y consecuencias de los impactos sobre el sistema, este análisis es inherentemente multidisciplinario. El análisis debe realizarse de manera espacial. El análisis de resiliencia puede considerarse una extensión o complemento del anterior. El objetivo es identificar, a nivel local, los umbrales que resultan en cambios en el equilibrio de los sistemas. Su objetivo es identificar áreas de intervención y definir puntos de referencia del sistema lo que provee bases para el monitoreo y administración (Resilience Alliance 2010). En particular, en la pesca ribereña, parece relevante la definición de resiliencia comunitaria como “La acción intencional para mejorar la capacidad individual y colectiva para responder a, e influir en el curso del cambio social y económico” que permite analizar estrategias de adaptación como modificación de patrones de pesca, incluyendo la pesca ilegal (CRRI 2013). Discusión La Administración Basada en Ecosistemas involucra forzosamente un análisis multidisciplinario y una planeación multisectorial. El andamiaje institucional para desarrollar esta última no existe todavía en nuestro país. Sin embargo, en mi opinión, el uso de algunos enfoques y técnicas mencionados en este trabajo debe ser evaluado en el contexto de la evaluación y administración de pesquerías ribereñas. Literatura citada Bodin Ö, C Prell. 2011. Social Networks and Natural
Resources Management. Uncovering the Social Fabric of Environmental Governance. Cambridge University Press.
Boesen N. 2008. Analysing and Addressing Governance in Sector Operations. Office for Official Publications of the European Communities. Luxemburg.
CRRI. 2013. Definitions of community resilience: an analysis. Community and Regional Resilience Institute. Washington.
Hilborn R. 2007. Managing fisheries is managing people: what has been learned? Fish and fisheries 8: 285-296.
IAEA. 2014. Root cause analysis following an event at a nuclear installation: reference manual. International Atomic Energy Agency-TECDOC series no. 1756. Vienna.
Kubiszewski I, R Costanza. 2012. Ecosystem Services for Sustainable Prosperity. State of the World 2012: Moving Toward Sustainable Prosperity. Worldwatch Institute. Washington.
McKinney l, W Tunnell, M Harwell, J Gentile, H Kelsey, W Dennison. 2011. Gulf of Mexico Report Card Initiative. Texas A&M University-Corpus Christi.
NOAA. 1998. Ecosystem-Based Fishery Management. A Report to Congress by the Ecosystem Principles Advisory Panel. NOAA, Washington.
Ostrom E. 1990. Governing the Commons. The Evolution of Institutions for Collective Action. Cambridge University Press.
Ostrom E. 2009. A General Framework for Analyzing Sustainability of Social-Ecological Systems. Science 325: 419-422.
Renard Y. 2004. Guidelines for stakeholder identification and analysis: A manual for Caribbean natural resource managers and planners. Caribbean Natural Resources Institute/ John D. and Catherine T. MacArthur Foundation.
Resilience Alliance. 2010. Assessing resilience in social-ecological systems: Workbook for practitioners. Version 2.0. Estocolmo.
Walker B, D Salt. 2006. Resilience thinking: Sustaining ecosystems and people in a changing World. Island Press. Washington.
Palabras clave: Administración de ecosistemas, pesquerías ribereñas, análisis de actores, análisis de redes sociales, análisis de causa raíz.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
205
099: cartel ESCAMAS EN LA EVALUACIÓN DE LA EDAD DEL ROBALO
Hania Gamboa Rivera, Elaine Espino Barr, Manuel Gallardo Cabello,
Arturo García Boa y Marcos Puente Gómez
Centro Regional de Investigación Pesquera de Manzanillo. INAPESCA. Playa Ventanas s/n, Manzanillo, Colima, México. C. P. 28200. Tel. (314)3323750. [email protected]; [email protected]; [email protected] .Introducción El robalo Centropomus nigrescens Günther 1864 es una especie carnívora, bentónico-demersal de fondos blandos. Rivas (1986) lo describe como diádromo y eurihalino. Como miembro de la familia Centropomidae muestra un fenómeno de inversión sexual, en el cual los machos se convierten en hembras cuando alcanzan longitudes mayores (Reinboth 1980). El tamaño máximo registrado es de 117 cm de longitud total y peso máximo de 23.8 kg (Bussing 1995). Las áreas de crianza de los juveniles se encuentran en los estuarios de donde entra y sale para alimentarse y resguardarse de la depredación (Froese y Pauly 2016). Se captura con línea de mano, curricán, red agallera y buceo. Tiene gran importancia económica local y regional debido a que su carne es excelente, se expende en la playa. Principalmente se comercializa eviscerado, fresco enhielado, y tiene una clasificación comercial como especie de 1ª clase alcanzando 120 pesos el kilogramo (Espino-Barr et al. 2004).
Fig. 1. Fotografía de una escama de robalo con cuatro annuli. Materiales y métodos Durante 8 días de cada mes de 2014 y 2015 se tomaron datos de longitud (Lt, cm), peso total (Pt, g) y se colectaron escamas de ejemplares de las zonas de acopio de Jalisco y Colima. Las escamas se lavaron y se montaron cuatro de cada individuo entre dos portaobjetos. Con la ayuda de un proyector de transparencias, adaptado se midieron ancho (Ea, mm), el largo (El, mm) y la distancia del foco a cada anillo (annuli).
También se analizó el incremento marginal (IM) para conocer el tiempo de formación del anillo. Las lecturas fueron realizadas por dos personas. Se realizaron regresiones entre Lt - Pt, Lt – El y Ea – El (Zar 1996). Resultados Se obtuvieron datos morfométricos de 504 individuos con longitudes de 11 a 101 cm Lt (promedio 46.4 cm ±15.44 d.e.) y pesos totales de 5.3 g a 9.8 kg Pt (promedio 1,004 g ± 1.1 kg d.e.). Las hembras fueron más grandes que los machos, aparecen a partir de los 67.5 cm (Fig. 2). La proporción entre el ancho y el largo de la escama es de 1: 0.78.
Fig. 2. Distribución de longitud total de hembras, machos y juveniles de robalo C. nigrescens.
Fig. 3. Relación peso-longitud de C. nigrescens. Los análisis de la relación peso longitud, muestran un crecimiento alométrico positivo, alcanzando mayores valores en el caso de las hembras (Fig. 3, Tabla 1). Tabla 1. Parámetros de las relaciones peso-longitud de Centropomus nigrescens
0
5
10
15
20
25
30
35
7.5 27.5 47.5 67.5 87.5 107.5
indefinidos hembras machos
Longitud total (cm)
Fre
cue
nci
a (
%)
0
2000
4000
6000
8000
10000
12000
0 20 40 60 80 100
Pt todos H M
Longitud total (cm)
Pe
so t
ota
l (g
)
Pt = 0.004 ∙ Lt 3.16
r2 = 0.989
n = 489

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
206
todos hembras machos
a 0.0042 0.0023 0.0049
b 3.161 3.326 3.136
r2 0.989 0.974 0.979
n 504 20 13 El factor de condición relativo (FC) mostró valores más altos de mayo a septiembre y los más bajos de diciembre a marzo. La relación entre el largo del pez y el ancho de la escama, es isométrica El = 1.346 · Lt 1.073. Los valores del incremento marginal (del último anillo al borde de la escama) mostraron una sola disminución en el año, durante los meses de noviembre a febrero y un aumento de marzo a octubre, lo que significa que se marca un anillo cuando el FC es más bajo, posiblemente por la reproducción. Se encontraron de 0 a 9 anillos de crecimiento (Fig. 4).
Fig. 4. Valores observados de lecturas de escamas por longitud de robalo Centropomus nigrescens. Discusión IGFA (2001) encuentra que la relación entre la longitud y el peso para C. nigrescens es Pt = 0.0141∙Lt3.0, mostrando un índice de crecimiento isométrico, menor al encontrado en el presente trabajo que es un índice de crecimiento alométrico positivo, en donde el pez crece más que peso que en longitud. El FC mostró un aumento anual, que puede estar relacionado con la temporada de mayor alimentación, como también se observa en el incremento marginal de las escamas. No existen estudios publicados de C. nigrescens por lo que estos datos son los primeros que se reportan para la determinación de la edad de esta especie. Sin
embargo, existen más estudios de C. undecimalis especie que puede considerarse gemela o críptica de C. nigrescens, aunque su hábitat se encuentre en el Océano Atlántico. Lorán-Núñez et al. (2012) encontraron para C. undecimalis longitudes de hasta 120 cm mayores que las reportadas para C. nigrescens de 101 cm. Caballero-Chávez (2009) encontró para C. undecimalis 10 grupos de edad similar en número a los reportados en este estudio. Conclusión La relación entre la longitud del pez y el ancho de la escama es isométrica. El factor de condición coincide con el valor más alto del incremento marginal. Las escamas de robalo sirven para determinar su edad. Literatura citada Bussing WA. 1995. Centropomidae. En: Guía FAO para la
identificación de especies para los fines de la pesca. Pacífico Centro Oriental. Vol. II Vertebrados – Parte 1
Caballero Chávez V. 2009. Pesquerías de robalo Centropomus undecimalis en Campeche. Ciencia Pesquera 17(2): 77-86.
Espino-Barr E, EG Cabral Solís, A Garcia Boa, M Puente Gómez. 2004. Especies marinas con valor comercial de la costa de Jalisco, México. SAGARPA – INP, México, 145 p.
Froese R, D Pauly (eds). 2016. FishBase. World Wide Web electronic publication. www.fishbase.org, version (01/2016).
IGFA. 2001. Database of IGFA angling records until 2001. IGFA, Fort Lauderdale, USA.
Lorán-Núñez RM, FR Martínez Isunza, AJ Valdez Guzmán, M Garduño Dionate, ER Martínez Lorán. 2012. Reproducción y madurez sexual de robalo prieto (Centropomus poeyi) y robalo blanco (C. undecimalis) en el sistema lagunar de Alvarado, Veracruz (2005-2007). Ciencia Pesquera 20(1): 49-64.
Reinboth R. 1980. Behavioral aspects of sex inversion in certain fishes. In: Bardach JE, JJ Magnuson, RC May, JM Reinhart (eds.). Fish behavior and its use in the capture and culture of fishes. ICLARM Conference Proceedings 5, 512 p.
Rivas LR. 1986. Systematic review of the Perciform Fishes of the genus Centropomus. Copeia 1986: 576-611.
Zar JH. 1996. Biostatistical analysis. 3rd ed. Prentice Hall. USA., 662p.
Palabras clave: escamas, edad, factor de condición, incremento marginal en escamas, Centropomus nigrescens.
0
20
40
60
80
100
0 1 2 3 4 5 6 7 8 9 10
Lon
git
ud
to
tal (
cm)
Número de anillos

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
207
100: oral ASPECTOS PESQUEROS DE Scomberomorus sierra, DE LA COSTA DE SINALOA, MÉXICO
Yolene Rosalía Osuna Peralta Hugo Aguirre Villaseñor, Vicente Hernández Covarrubias,
Ricardo Meraz Sánchez, Roberto Paredes Mellado y José Ángel Guillén Gamboa
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Avenida Camarón Sábalo S/N, Estero del Yugo, Mazatlán, Sinaloa, México. C.P. 82000. [email protected] Introducción La sierra (Scomberomorus sierra Jordan y Starks 1895) (Perciformes: Scombridae) es una especie tropical que se distribuye a lo largo de todo el litoral del Pacífico mexicano (Collette y Nauen 1983). Es una de las especies con mayor nivel de captura; en 2014 se reportó una producción de 2,170 t, con un valor de 33 millones de pesos (SAGARPA 2014). El presente trabajo tiene como objetivo generar bases técnicas sobre los indicadores de la pesquería de S. sierra capturada por la flota de pequeña escala en las costas de Sinaloa, que permita actualizar las medidas de manejo empleadas actualmente en la pesquería. Material y métodos Los datos provienen de muestreos de campo del programa de Pesca Ribereña del INAPESCA, de enero de 2014 a noviembre de 2015; y avisos de arribo de captura de la flota ribereña de los sitios de muestreo (Topolobampo, Cospita, Huitussi, Mármol, Isla de la Piedra y Teacapán). Estructura de longitudes. A partir de la distribución de longitud total (LT) se ajustó un modelo multinomial con la finalidad de ubicar las principales cohortes que son aprovechadas por la pesquería. La edad se estimó utilizando los parámetros de la curva de crecimiento de von Bertalanffy estimados para Colima (Espino-Barr et al. 2012). Longitud de Primera Captura (L50). La L50 fue estimada a partir de un análisis multimodelo. Con los datos de frecuencia absoluta se utilizaron cinco modelos sigmoidales para estimar la longitud media de primera captura. Los modelos son: Gompertz (1825), Richards (1959), Lysack (1980), White et al. (2002) y Brouwer y Griffiths (2005). Para elegir el mejor modelo se utilizó el criterio de decisión de Akaike (AIC) (Jacob-Cervantes y Aguirre-Villaseñor 2014). Tasa de Explotación. La mortalidad total (Z) se estimó a partir del método de la curva de captura linealizada, se calculó la tasa de sobrevivencia, el valor de mortalidad por pesca y la tasa de explotación.
Resultados Estructura de longitudes. Se analizaron 513 organismos. El intervalo de LT fue de 142 a 770 mm con una media de 433 mm. En el histograma de frecuencia se observan dos principales grupos modales con media en: 418 y 560 mm (Fig. 1).
Fig. 1. Frecuencia de longitud total (mm) y edades, de ambos sexos, de la sierra S. sierra capturada en Sinaloa 2014-2015. El 75% de captura de S. sierra corresponde a organismos de 3 años de edad, y esto concuerda con lo estipulado en la CNP, 2010, donde la redes de 3 y 3.5 pulgadas que se utilizan para la captura de esta especie, corresponden a organismos de 3 años de edad que están en proceso de maduración reproductiva (Fig. 1). El intervalo de peso total de los organismos muestreados fue de 190 g a 2,400 g con una media de 462 g. La función que describe la relación peso-longitud es de tipo potencial, w= 610-6(LT)2.98 (r2= 0.95, p<0.05), la pendiente no es significativamente diferente del valor teórico de alometría de 3 (p>0.05), por lo tanto la sierra S. sierra tiene un crecimiento isométrico. Longitud de Primera Captura (L50). Los modelos que mejor se ajustaron a los datos observados fueron: Lyssack, White, y Brouwer y Griffiths. De acuerdo con el criterio de inferencia multimodelo (Burnham y Anderson, 2002), por lo que estos modelos se utilizaron para estimar el modelo global (Fig. 2). La L50 calculada global (459 mm) corresponde a peces de 3 años de edad, este valor está por debajo de lo estipulado en la CNP, 2010 (500 mm).
0
20
40
60
80
100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
500
520
540
560
580
600
620
640
660
680
700
720
740
760
780
800
Fre
cuen
cia
abso
luta
(n)
IC (mm)
1 2 3 4 5 6 7
Edad (años)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
208
Fig. 2. Distribución de la longitud de primera captura (L50).
Tasa de Explotación. La ecuación de regresión calculada para obtener la pendiente es y = -1.1141 x + 8.0149, r2 = 0.76; n = 15. (Fig. 3).
Fig. 3. Longitud convertida curva de captura Tabla 1. Resumen de parámetros de S. sierra de Sinaloa y Colima, México.
Parámetros Este estudio Espino-Barr et
al. (2012) L∞ 995.4 k 0.205 t0 -0.06 Z 1.11 0.76 M 0.19 0.19 F 0.90 0.56 S 0.33 0.46 Sobrevivencia % 33.00 46.00 Mortalidad Total % 67.00 54.00 Si Z= 100 Mortalidad por pesca 83.00 74.00 Mortalidad Natural 17.00 26.00 E=F/Z 0.82 0.73 Los valores de mortalidad para edades entre 3 y 7 años fue de Z = 1.1. Los valores de la tasa de Z para S. sierra en Colima son de 0.7 (Espino-Barr et al. 2012), valor por debajo al obtenido en este estudio, sin embargo el estimado de mortalidad natural (M = 0.19) es similar en ambos estudios (Tabla 1). Los resultados infieren con una cierta probabilidad que 67% de la sierra muere anualmente en Sinaloa. A partir de estos valores, 83% muere por efecto de la pesca y 17% de forma natural, por lo tanto, la
pesquería de S. sierra está aprovechada al máximo sustentable, lo cual coincide con lo estipulado en la ficha de Sierra de la Carta Nacional Pesquera 2010 (DOF 2010). Conclusión La pesca de sierra en Sinaloa se basa principalmente en organismos de 3 años, por lo que existe una alta probabilidad de capturar organismo menores a la longitud de madurez. Se estimó una alta mortalidad por pesca, lo cual pudiera indicar cierto grado de sobrexplotación, sin embargo, para que este resultado sea concluyente es necesario evaluar la pesquería con modelos estructurados por edades. Literatura citada Burnham KP, DR Anderson. 2004. Multimodel inference.
Understanding AIC and BIC in model selection. Sociological Methods and Research 33: 261-304.
DOF. 2010. Carta Nacional Pesquera. Diario Oficial de la Federación. SAGARPA.
Collette BB, Nauen CE. 1983. Scombrids of the world. An annotated and illustrated catalogue of tunas, mackerels, bonitos and related species known to date. FAO Species Catalog. Vol. 2. FAO Fish. Synop. 125(2): 1–1337.
Espino-Barr E, M Gallardo-Cabello, EG Cabral Solís, A García-Boa, M Puente-Gómez. 2012. Aspects of Scomberomorus sierra fishery from the coast of Colima, México. Ciencia Pesquera 20(1): 77-88.
Jacob-Cervantes ML, H Aguirre-Villaseñor. 2014. Inferencia multimodelo y selección de modelos aplicados a la determinación de L50 para la sardina crinuda Opisthonema libertate del sur del Golfo de California. Ciencia Pesquera 22(1): 59-66.
SAGARPA. 2014. Base de datos de producción anuario 2014. Comisión Nacional de Acuacultura y Pesca, Secretaria de Agricultura, Ganadería, Pesca y Alimentación.
Palabras clave: Estructura de longitudes, longitud de primera captura, Tasa de explotación, Mortalidad
0
0.2
0.4
0.6
0.8
1
1.2
0 200 400 600 800 1000
Fre
cuen
cia
(# in
d.)
Longitud Total (mm)
Lyssack
White
Gompertz
Brouwer y Griffiths
Richards
y = -1.1141x + 8.0149
R² = 0.7661
0
1
2
3
4
5
6
0.00 2.00 4.00 6.00 8.00
LN (
Ca
ptu
ra/Δ
t)
Edad (años)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
209
101: oral ESTIMACION DE LA EDAD Y CRECIMIENTO DEL BOTETE Sphoeroides annulatus UTILIZANDO LA
INFERENCIA MULTIMODELO
Hugo Aguirre Villaseñor, Yolene Rosalía Osuna Peralta, Roberto Paredes Mellado, Vicente Hernández Covarrubias y José Ángel Guillen Gamboa
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Avenida Camarón Sábalo S/N, Estero del Yugo, Mazatlán, Sinaloa, México, C.P. 82000. [email protected] Introducción Sphoeroides annulatus (Jenyns 1842) es una especie altamente cotizada, presenta un filete de carne firme, de color blanco y libre de espinas, con un precio que oscila entre $80 y $100 por kg en el mercado del puerto de Mazatlán. De acuerdo a las cifras de CONAPESCA, en la última década (2003-2013) se ha capturado en promedio 160,487 toneladas métricas (TM) por las embarcaciones menores y 22,847 TM por las embarcaciones mayores (SAGARPA 2014). Existe una gran variedad de modelos para estimar la edad y crecimiento, el más utilizado es el de von Bertalanffy y otros muy comunes son el de: Gompertz, Logístico y Schnute y Richards (Katsanevakis 2006). La inferencia multimodelo postula que cuando los datos soportan evidencia de más de un modelo, en lugar de estimar los parámetros a partir de sólo “el mejor modelo”, es posible hacerlo a partir de varios e incluso todos los modelos considerados. Obtener a partir de varios modelos el valor promedio de la variable predicha, es una ventaja para alcanzar una inferencia robusta que no esté condicionada a un solo modelo (Burnham y Anderson 2002). El objetivo del presente trabajo es estimar los parámetros de crecimiento de S. annulatus a partir de cuatro modelos, y construir un modelo global utilizando la técnica de la inferencia multimodelo. Material y métodos Los datos de longitud total (LT) de cada individuo se colectaron en el litoral de Sinaloa durante 2014 y 2015. A partir del análisis de frecuencias de LT, las modas de las distribuciones de longitudes se ajustó a una distribución multinomial (Haddon 2001). Para validar el número de modas se utilizó el criterio de la teoría de información (Montgomery et al. 2010). Con las modas obtenidas se estimó el crecimiento individual mediante las ecuación de: von Bertalanffy, Schnute y Richards, Gompertz y Logístico (Katsanevakis y Maravelias 2008, Montgomery et al. 2010). Los modelos se ajustaron minimizando la diferencia cuadrática de los errores con la función Solver de Excel 2010, con el algoritmo de búsqueda directa de Newton. El error estándar (SE) y el intervalo de confianza de 95% (IC) de la longitud infinita �∞ se calculó para cada modelo (Katsanevakis 2006).
El criterio de inferencia multimodelo se utilizó para construir el modelo global (Katsanevakis 2006). Resultados y discusión Para el total de la muestra se analizaron 2,719 organismos de S. annulatus entre marzo de 2014 y diciembre de 2015. El intervalo de LT fue: 71 a 440 mm con una media de 220 mm. Se observaron siete grupos modales en el total de la muestra. El grupo 1 se presentó de julio a octubre 2015 con una moda de 150 mm LT. El grupo 2 en abril 2015 con una moda de 125 mm LT, su progresión modal se puede seguir hasta diciembre 2015 con una moda de 225 mm LT. El grupo 3 se presentó en marzo 2014 con una moda de 125 mm LT, y su progresión modal fue hasta diciembre 2015 con una moda de 225 mm LT. El grupo 4 se presentó en marzo 2014 con una moda de 225 mm LT, su progresión modal se puede seguir hasta junio de 2015 con una moda de 300 mm LT. El grupo 5 se presentó en abril 2014 con una moda de 325 mm LT, su progresión modal se puede seguir hasta junio 2015 con una moda de 375 mm LT. El grupo 6 se presentó en abril 2014 con una moda de 400 mm LT, su progresión modal se puede seguir hasta octubre 2015 con una moda de 450 mm LT. El grupo 7 se presentó en mayo 2015 con un organismo de 475 mm LT. Los datos de las modas obtenidas se ajustaron al modelo de crecimiento de von Bertalanffy (1), Schnute y Richards (2), Gompertz (3) y Logístico (4). Lt = ∗ 1 − e− . ∗ t− . 1 (1) Lt = . ∗ (1 − . ∗ e− . ∗t . ) .8E− (2) Lt = 1 ∗ e−e− . ∗ . − . (3) Lt = 1+e− . ∗ t− . (4) El modelo que presentó el valor más bajo de Akaike (AIC) fue el modelo de von Bertalanffy (265), seguido por el modelo de Gompertz (266.84), el modelo Logístico (269) y el modelo de Schnute y Richards (282). De acuerdo con el enfoque de inferencia multimodelo, para construir el modelo global sólo se tomaron en cuenta los valores de delta () menores o iguales a 7. El modelo de Schnute y Richards (=19) no se tomó en cuenta para la construcción del

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
210
modelo global, el modelo de von Bertalanffy aporta 69% de la información, el modelo de Gompertz 25% y el modelo Logístico 6%. El valor promedio de L∞ fue de 536 mm LT y su intervalo de confianza del 95% fue de 535 a 537 mm LT (Fig. 1).
Fig. 1. Modelo global (línea gris) calculado a partir de la inferencia multimodelo considerando cuatro modelos candidatos (línea continua): a) von Bertalanffy, b) Schnute y Richards, c) Gompertz, d) Logístico. A partir del modelo global, se estima que los organismos analizados en el presente estudio van de 9 meses (71 mm LT) a 5 años (440 mm LT). Conclusión En la investigación científica frecuentemente se requiere utilizar técnicas de simulación para comprender los fenómenos estudiados. Dichas técnicas se desarrollan tratando de representar fielmente las características de los sistemas estudiados a través de modelos estadísticos o matemáticos. Los modelos representan una abstracción de la realidad, con éstos es posible explorar sus límites mediante técnicas de simulación y evaluar el desempeño del sistema estudiado, ya sea para la prueba de hipótesis o utilizándolo para fines predictivos. La selección del modelo de crecimiento puede variar de acuerdo a la especie y a los datos disponibles. El criterio de selección del mejor modelo dependerá del nivel de ajuste de éste a los datos (Katsanevakis 2006). En virtud de lo anterior, la inferencia multimodelo es una herramienta muy poderosa en la toma de decisiones, pues permite confrontar diferentes hipótesis (modelos) e incluir las más plausibles en un modelo global incorporando la información de todos los modelos. Con base a los valores de los parámetros obtenidos en este estudio, la comparación de los modelos de crecimiento mostró una descripción similar de los datos observados, el modelo de von Bertalanffy describió mejor la información, mientras que el modelo de Schnute y Richards es el que peor se
desempeñó, el valor de =19 entre ambos modelos, se explica por la penalización que recibe Schnute y Richards por el mayor número de parámetros. Literatura citada Burnham KP y DR Anderson. 2002. Model selection and
multimodel inference: a practical information-theoretic approach. Springer. New York. 488p.
Haddon M. 2001. Modelling and quantitative methods in fisheries. Chapman & Hall, Boca Raton, 406 pp.
Katsanevakis S. 2006. Modelling fish growth: Model selection, multi-model inference and model selection uncertainty. Fisheries Research 81(2-3): 229-235.
Katsanevakis S y CD Maravelias. 2008. Modelling fish growth: multi-model inference as a better alternative to a priori using von Bertalanffy equation. Fish and Fisheries 9: 178–187.
Montgomery SS, CT Walsh, M Haddon, CL Kesby y DD Johnson. 2010. Using length data in the Schnute Model to describe growth in a metapenaeid from waters off Australia. Marine and Freshwater Research 61 1435-1445.
SAGARPA. 2014. Anuario estadístico de acuacultura y pesca 2014. Comisión Nacional de Acuacultura y Pesca, SAGARPA, México, D.F. base de datos http://www.conapesca.gob.mx/wb/cona/cona_anuario_estadistico_de_pesca
Palabras Clave: Mínimos cuadrados, Teoría de información, Akaike, intervalo de confianza.
0
100
200
300
400
500
600
0 20 40 60 80 100
LT (
mm
)
Tiempo (mes)
0
100
200
300
400
500
600
0 20 40 60 80 100
LT (
mm
)
Tiempo (mes)
0
100
200
300
400
500
600
0 20 40 60 80 100
LT (
mm
)
Tiempo (mes)
0
100
200
300
400
500
600
0 20 40 60 80 100
LT (
mm
)
Tiempo (mes)
c) d)
a) b)
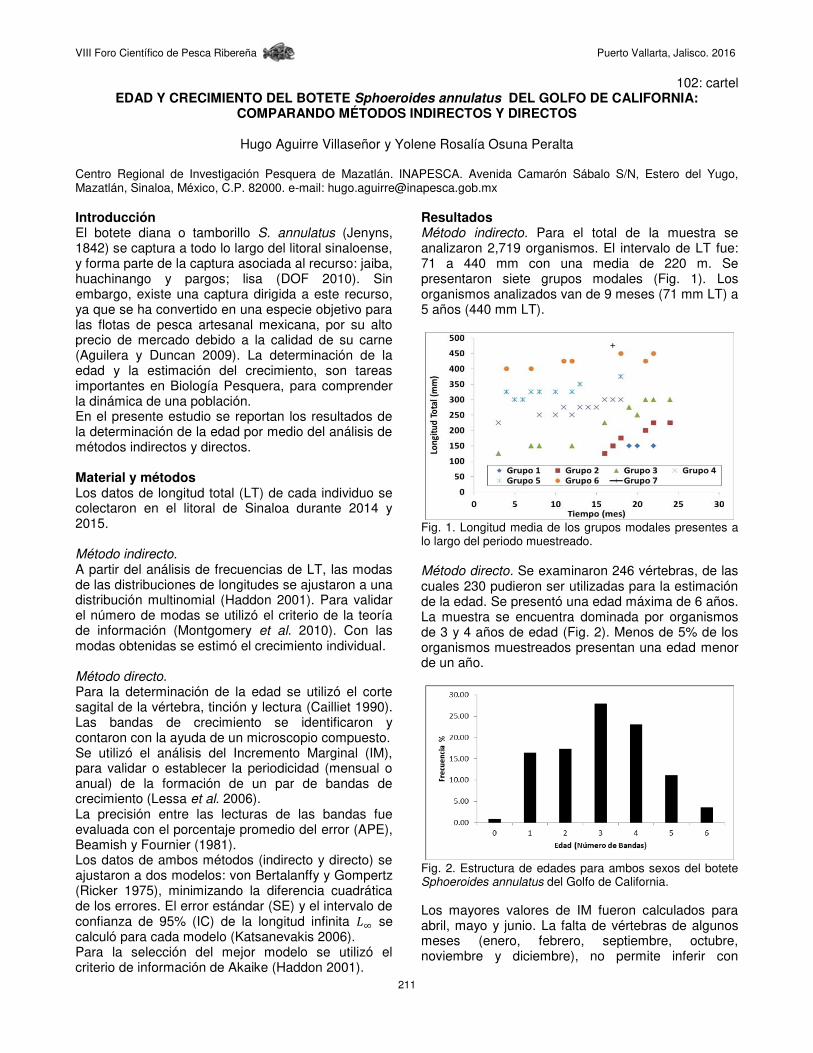
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
211
102: cartel EDAD Y CRECIMIENTO DEL BOTETE Sphoeroides annulatus DEL GOLFO DE CALIFORNIA:
COMPARANDO MÉTODOS INDIRECTOS Y DIRECTOS
Hugo Aguirre Villaseñor y Yolene Rosalía Osuna Peralta
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Avenida Camarón Sábalo S/N, Estero del Yugo, Mazatlán, Sinaloa, México, C.P. 82000. e-mail: [email protected] Introducción El botete diana o tamborillo S. annulatus (Jenyns, 1842) se captura a todo lo largo del litoral sinaloense, y forma parte de la captura asociada al recurso: jaiba, huachinango y pargos; lisa (DOF 2010). Sin embargo, existe una captura dirigida a este recurso, ya que se ha convertido en una especie objetivo para las flotas de pesca artesanal mexicana, por su alto precio de mercado debido a la calidad de su carne (Aguilera y Duncan 2009). La determinación de la edad y la estimación del crecimiento, son tareas importantes en Biología Pesquera, para comprender la dinámica de una población. En el presente estudio se reportan los resultados de la determinación de la edad por medio del análisis de métodos indirectos y directos. Material y métodos Los datos de longitud total (LT) de cada individuo se colectaron en el litoral de Sinaloa durante 2014 y 2015. Método indirecto. A partir del análisis de frecuencias de LT, las modas de las distribuciones de longitudes se ajustaron a una distribución multinomial (Haddon 2001). Para validar el número de modas se utilizó el criterio de la teoría de información (Montgomery et al. 2010). Con las modas obtenidas se estimó el crecimiento individual. Método directo. Para la determinación de la edad se utilizó el corte sagital de la vértebra, tinción y lectura (Cailliet 1990). Las bandas de crecimiento se identificaron y contaron con la ayuda de un microscopio compuesto. Se utilizó el análisis del Incremento Marginal (IM), para validar o establecer la periodicidad (mensual o anual) de la formación de un par de bandas de crecimiento (Lessa et al. 2006). La precisión entre las lecturas de las bandas fue evaluada con el porcentaje promedio del error (APE), Beamish y Fournier (1981). Los datos de ambos métodos (indirecto y directo) se ajustaron a dos modelos: von Bertalanffy y Gompertz (Ricker 1975), minimizando la diferencia cuadrática de los errores. El error estándar (SE) y el intervalo de confianza de 95% (IC) de la longitud infinita �∞ se calculó para cada modelo (Katsanevakis 2006). Para la selección del mejor modelo se utilizó el criterio de información de Akaike (Haddon 2001).
Resultados Método indirecto. Para el total de la muestra se analizaron 2,719 organismos. El intervalo de LT fue: 71 a 440 mm con una media de 220 m. Se presentaron siete grupos modales (Fig. 1). Los organismos analizados van de 9 meses (71 mm LT) a 5 años (440 mm LT).
Fig. 1. Longitud media de los grupos modales presentes a lo largo del periodo muestreado. Método directo. Se examinaron 246 vértebras, de las cuales 230 pudieron ser utilizadas para la estimación de la edad. Se presentó una edad máxima de 6 años. La muestra se encuentra dominada por organismos de 3 y 4 años de edad (Fig. 2). Menos de 5% de los organismos muestreados presentan una edad menor de un año.
Fig. 2. Estructura de edades para ambos sexos del botete Sphoeroides annulatus del Golfo de California. Los mayores valores de IM fueron calculados para abril, mayo y junio. La falta de vértebras de algunos meses (enero, febrero, septiembre, octubre, noviembre y diciembre), no permite inferir con

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
212
precisión la periodicidad de formación del par de bandas. Aun así, es posible proponer que, por analogía con especies similares, la periodicidad de las bandas encontradas es de tipo anual. Para las lecturas se obtuvo un APE de 2.49%, un porcentaje pequeño indica una mayor precisión entre las lecturas realizadas. Modelos de crecimiento. Para el método indirecto, el modelo que mejor describe el crecimiento de S. annulatus es el de von Bertalanffy (AIC=265). Para el directo fue el modelo de Gompertz (AIC=247.9) (Tabla 1). Tabla 1. Parámetros de crecimiento para S. annulatus con método directo e indirecto.
Método directo Método indirecto von
Bertanffy Gompertz von
Bertanffy Gompertz
L∞ 323.1 309.2 554.6 501.4 k 0.46 0.67 0.24 0.42 t0 -0.26 0.61 -0.21 -0.63 AIC 252 247.9 265 267
El valor de L∞ encontrado en este estudio para el método directo está por debajo de la longitud máxima encontrada para S. annulatus (440 mm LT), esta diferencia pudiera ser por la falta de análisis de vértebras de organismos con longitudes >300 mm LT, que pudo haber causado una subestimación de la longitud máxima de esta especie. Tabla 2. Comparación de la edad de S. annulatus.con métodos indirectos y directos.
Directo Indirecto Edad (año) vB G vB G 1 142 143 138 159 2 209 208 226 220 3 251 252 295 291 4 278 278 350 350 5 295 293 393 396 6 305 301 427 429
Fig. 3. Curva de crecimiento para sexos combinados con el modelo de von Bertalanffy (línea continua) y el modelo de Gompertz (línea punteada). a) Método indirecto, b) método directo. Conclusión Se considera necesario verificar el estado de salud de los stocks de estos organismos. En este sentido, son esenciales los estudios sobre edad y crecimiento
ya que con esta información es posible construir modelos demográficos cuantitativos para estimar la productividad de las especies y determinar su rendimiento máximo permisible. Los estudios de edad y crecimiento en S. annulatus son escasos, por lo que poco se sabe sobre sus características biológicas (reproducción y crecimiento, entre otras), que confieren a una especie la capacidad de resistencia a la explotación. En vista de la escasa información disponible sobre esta especie, es necesario verificar el estado de sus poblaciones contribuyendo, a la determinación de su edad, lo que permitirá en lo sucesivo estimar los parámetros poblacionales de la especie. Además de elaborar la ficha de Botete en la Carta Nacional Pesquera (DOF 2010) y el capítulo del libro de “Sustentabilidad”. Dicha información es útil y necesaria para evaluar su estatus de conservación. Literatura citada Beamish RJ, DA Fournier. 1981. A method for comparing
the precision of a set of age determinations. Can. J. Fish. Aquatic Sci. 38: 982-983.
Cailliet GM. 1990. Elasmobranch age determination and verification; an updated review. 157-165. Elasmobranchs as living resources: advances in the biology, ecology, systematics and the status of the fisheries. NOAA Tech. Rep. NMFS 90.
DOF. 2010. Carta Nacional Pesquera. SAGARPA. Diario Oficial de la Federación.
Haddon M. 2001. Modelling and Quantitative Methods in Fisheries. Chapman & Hall, Boca Raton, 406 pp.
Katsanevakis S. 2006. Modelling fish growth: Model selection, multi-model inference and model selection uncertainty. Fisheries Research 81(2-3): 229-235.
Montgomery SS, CT Walsh, M Haddon, CL Kesby y DD Johnson. 2010. Using length data in the Schnute Model to describe growth in a metapenaeid from waters off Australia. Marine and Freshwater Research 61: 1435–1445.
Ricker WE. 1975. Computation and interpretation of biological statistics of fish populations. Bull. Fish. Res. Bd Canada 191, Ottawa. 382 pp.
Palabras clave: Mínimos cuadrados, Teoría de información, Akaike, intervalo de confianza.
a) b)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
213
103: oral PESCA ARTESANAL DE TIBURÓN EN EL NORTE Y CENTRO DE VERACRUZ
Liliana Cuervo López1, Juan Fernando Márquez Farías2 y Víctor Alfonso Reyna Matezans1
1Facultad de Ciencias Biológicas y Agropecuarias. Universidad Veracruzana. 2Facultad de Ciencias del Mar. Universidad Autónoma de Sinaloa. [email protected]; [email protected]; [email protected] Introducción Se considera que las pesquerías de tiburón están aprovechadas al máximo. En Veracruz, al igual que en el resto de los estados costeros esta pesquería es fuente de alimento y empleo para las comunidades ribereñas. El Anuario Estadístico de Pesca 2014 reporta 29,435 t de producción de tiburón-cazón y 9,483 t de rayas (y similares); Veracruz y Tamaulipas son los principales estados productores en la región del Golfo de México (CONAPESCA 2014). La generación de información permite actualizar el conocimiento de las pesquerías de tiburones en el norte y centro de Veracruz, con énfasis en la composición específica en las capturas comerciales por la pesca artesanal y el comportamiento espacio–temporal de las poblaciones explotadas. Material y métodos De septiembre 2013 a abril 2014 se monitorearon las capturas comerciales de la flota artesanal en los campamentos pesqueros de Tamiahua, Tecolutla y Chachalacas, en la porción norte y centro de Veracruz. Cada organismo se examinó registrándose: longitud total extendida (LTE); sexo, y la madurez sexual. Para valorar la representatividad de la diversidad de especies se generaron curvas de acumulación sometiendo los datos a un Bootstrap de 1000 permutaciones. El esfuerzo pesquero fue el número de tiburones capturados por viaje de pesca por horas de calado. La tasa de captura se estimó utilizándose como unidad “tib/h” (tiburón/hora). Con el fin de documentar la estacionalidad del recurso, las tasas de captura se analizaron por campamento pesquero y por mes. Se evaluaron las diferencias de cada factor (campamento/mes) con un ANOVA. Resultados y discusiones Se identificaron 22 especies de tiburones de seis órdenes, ocho familias y 11 géneros. Las especies capturadas fueron mayormente representadas por longitudes menores a 1.5 m LTE. La especie más abundante en las tres zonas fue Rhizoprionodon terraenovae (38.50%), seguida de Carcharhinus limbatus (22.11%), Sphyrna lewini (10.83%), C. isodon (9.59%) y C. brevipinna (6.91%). El número de especies observadas en cada campamento pesquero fue variable (Tamiahua, 21; Tecolutla 17 y Chachalacas 14 especies). La tasa de captura de tiburón en el norte y centro de Veracruz
fue de 0.26 ± 0.02 tib/h, registrándose mayor promedio con red (0.31 ± 0.02) que con anzuelo (0.22 ± 0.03). Noviembre fue el mes con mayor promedio (0.44 ± 0.10) y el menor marzo (0.12 ± 0.02). El mes de mayor captura con anzuelo fue noviembre (0.48 ± 0.17); y con red (0.39 ± 0.06). En los tres campamentos, el uso de anzuelo registró menor tasa de captura en relación con el uso de red (0.22 ± 0.03 y 0.31 ± 0.02 tib/h, respectivamente). Las especias más capturadas con anzuelos fueron S. cubensis (1.63 ± 0.72), R. terraenovae (0.67 ± 0.28) y Mustelus norrisi (0.21 ± 0.05); con red, fueron R. terraenovae (0.51 ± 0.05), C. isodon (0.32 ± 0.08) y C. limbatus (0.28 ± 0.04). Rhizoprionodon terraenovae, C. limbatus, C. brevipinna y S. lewini fueron constantes en las capturas en el norte y centro de Veracruz durante todo el periodo; sin embargo, R. terraenovae dominó durante noviembre (1.01 ± 0.28 tib/h); las otras tres especies registraron tasas de captura más altas en diciembre (0.35 ± 0.08, 0.26 ± 0.05 y 0.24 ± 0.06, respectivamente). Con respecto al análisis de tasas de captura por mes, R. terraenovae (P = 0.135), C. isodon (P = 0.47) y C. brevipinna (P = 0.2) no mostraron diferencias significativas entre los meses en que fueron capturados; S. lewini (P = 0.01) y C. limbatus (P = 0.035) mostraron diferencias significativas entre febrero y octubre; diciembre, noviembre y marzo para cada especie, respectivamente. Por otro parte, hubo diferencias entre campamento pesqueros, ya que en cuatro de las cinco especies analizadas se encontraron diferencias significativas. S. lewini (P=0.005) y R. terraenovae (P=0.006) mostraron diferencias entre Tamiahua y Tecolutla; C. brevipinna (P=0.017) entre Tecolutla y Chachalacas; en el caso de C. limbatus las tres campamentos pesqueros tuvieron diferencias estadísticas entre sí. C. isodon (P=0.125) fue el único que no mostró diferencias significativas entre campamentos. Se enlistaron 22 especies de tiburones, riqueza biológica que difiere con otras investigaciones tales como Castillo-Géniz et al. (1998), Marín (1992) y Bonfil (1997), con 27, 28 y 33 especies respectivamente; situación que se atribuye a diferencias entre las unidades de pesca y el periodo de monitoreo. Por otro parte, el dominio de R. terraenovae, C. limbatus, C. brevipinna y S. lewini en relación con los porcentajes de captura que se reportan en este trabajo es constante en

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
214
comparación con anteriores investigaciones realizadas en el Golfo de México (Marín 1992, Rodríguez de la Cruz et al. 1996) y en Veracruz (Oviedo et al. 2008 y 2010). Un factor que influyó en la variabilidad temporal de las tasas de captura específicas fue el clima registrado durante el periodo de muestreo, resultando en una mayor congregación de cazones y tiburones juveniles (e.g. R. terraenovae, C. limbatus, C. brevipinna y C. isodon) en las “corridas” de octubre–noviembre hacia aguas más cálidas, fenómeno que es aprovechado con la captura de gran número de organismos por los pescadores de la zona; mientras que la aproximación de tiburones de mayores dimensiones (e.g. C. plumbeus, S. lewini y C. leucas) en los meses posteriores retribuyen en mayor volumen de captura pero menor cantidad de tiburones capturados. Conclusiones Durante la temporada de pesca septiembre 2014–abril 2015 las capturas artesanales de tiburón en el norte y centro de Veracruz estuvieron integradas por 22 especies de tiburones; las especies más abundantes fueron: R. terraenovae, C. limbatus, C. brevipinna, C. isodon y S. lewini. R. terraenovae fue la especie más frecuente en función de número de organismos capturados (38.50%), seguido de C. limbatus (22.11%), S. lewini (10.83%), C. isodon (9.59%), C. brevipinna (6.91%), S. cubensis (4.18%), S. tiburo (1.63%), C. leucas (1.21%), S. mokarran (1.05%), M. norrisi (0.99%) y otras 11 especies (3%). Las capturas comerciales estuvieron integradas por tiburones de longitudes menores a 1.5 m; sin embargo, el amplio intervalo de especies presentes en la zona de estudio hizo posible poder observar organismos con madurez sexual e incluso hembras preñadas de varias especies. Se percibe una disminución en las poblaciones y en las longitudes promedios de varias especies producto de los intensos regímenes de pesca a los que han sido sometidos, tales el caso de C. brevipinna, S. tiburo, C. leucas, C. acronotus, C. falciformis y G. cuvier. Las mayores tasas de captura de manera general se obtuvieron en noviembre, abril y octubre, para el uso de redes, anzuelos y ambos grupos combinados; mientras que, las especies con mayor promedio de captura fueron S. cubensis, R. terraenovae, C. isodon, C. limbatus y M. norrisi. Literatura citada Bonfil R. 1997. Status of shark resources in the Southern
Gulf of México and Caribbean: implications for management. Fish. Res. 29: 101–117
Castillo-Géniz JL, JF Márquez-Farías, MC Rodríguez de la Cruz. 1998. The Mexican artisanal shark fishery in the Gulf of Mexico: Towards a regulated fishery. Mar. Fresh. Res. 49: 611–20.
CONAPESCA. 2014. Base de datos del anuario estadístico 2014. [Actualizado al 12 de febrero de 2015]. Página electrónica: www.conapesca.gob.mx/wb/cona/anuario _2014.
Marín OR. 1992. Aspectos biológicos de los tiburones capturados en las costas de Tamaulipas y Veracruz, México. Tesis de Licenciatura. Universidad Veracruzana, Xalapa, Veracruz, México. 147 p.
Oviedo-Pérez JL, LO González, JA Pech P, LE Martínez C. 2008. Pesquería de elasmobranquios en la zona centro del estado de Veracruz, México, durante el otoño de 2007, 50–51 pp. In: Memoria del III Simposium Nacional de Tiburones y Rayas. SOMEPEC. Facultad de Ciencias, UNAM. D. F., México. 25–29 de Agosto de 2008. 252 p.
Oviedo-Pérez JL, L González–Ocaranza, LE Martínez–Cruz. 2010. Pesquería de tiburones en el estado de Veracruz durante 2008. 89–91 pp. En Díaz–Sánchez AW, CG Aguilar, OU Mendoza–Vargas (Ed.). 2010. Memorias del IV Simposium Nacional de Tiburones y Rayas. SOMEPEC. 171 pp.
Rodríguez de la Cruz MC, JL Castillo, JF Márquez. 1996. Evaluación de la pesquería de tiburón del Golfo de México. CONACyT. Informe Final del Proyecto de Investigación 116002–5–13134N–9206. 199 p.
Palabras clave: composición específica, tasas de captura, cazones, recurso pesquero.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
215
104: oral COMPOSICIÓN Y ABUNDANCIA DE LA CAPTURA DE PECES DE LA FLOTA ARTESANAL EN
MAZATLAN, SINALOA, MEXICO
Vicente Hernández Covarrubias, Hugo Aguirre Villaseñor, Yolene R. Osuna Peralta, Roberto Paredes Mellado y José Angel Guillen Gamboa
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Avenida Camarón Sábalo S/N, Estero del Yugo, Mazatlán, Sinaloa. C.P. 82000. [email protected] Introducción Las pesquerías ribereñas y en particular las de escama son un recurso tradicional en los litorales de nuestro país, por lo que es importante evaluarlas si se quiere que éstas perduren en el tiempo y continúen brindando seguridad alimentaria, ingresos y desarrollo en las comunidades costeras. Dada la gran diversidad de especies tanto demersales como pelágicas que involucran las capturas, así como la variedad de artes de pesca, existen esfuerzos importantes para encontrar métodos de análisis adecuados que incorporen las distintas dimensiones del problema (González-Becerril et al. 2006). El objetivo central de este estudio, es caracterizar la pesquería de escama del litoral de Sinaloa, mediante, la abundancia y composición de las especies que se explotan regionalmente.
Fig. 1. Área de estudio. Material y métodos Los principales lugares donde se pescan las especies de escama, son variables, en términos de distancia o tiempo de viaje. Hay lanchas que pescan en las riberas del puerto de Mazatlán, alrededor de las Tres Islas (Venado, Pájaro y Chivo) y Cerritos, mientras que otras se alejan hasta las zonas rocosas frente a Mármol, Paredones y los Caballos (Fig. 1).
Se realizaron muestreos semanales en Mazatlán de los desembarques en la centro de recepción “Lonja pesquera” de la Isla de La Piedra. A partir de los recibos de compra, se obtuvieron los datos de composición, captura (kg), y número de viajes, para estimar la captura por unidad de esfuerzo (CPUE). Para la identificación taxonómica de de las especies estudiadas, nos basamos en distintas publicaciones y catálogos de peces marinos (Fisher et al. 1995, Espino Barr et al. 2003, Amezcua Linares 2008). Resultados y Discusión Se obtuvieron datos de captura de enero a diciembre de 2014, se registraron un total de 236,126 kg, en 2,042 viajes de pesca. La captura mínima mensual fue de 494 kg en abril y la máxima 43,729 kg en diciembre (Fig. 2).
Fig. 2. Capturas totales mensuales, Lonja Pesquera Isla de la Piedra, Mazatlán en 2014. Se determinaron 81 especies pertenecientes a 28 familias. Las principales especies que componen la captura son chihuil (20.7%), sierra (16.7%), berrugata (10%), bullet (6.2%), barrilete (5.7%), manta (3.9%), chile verde (3.8%), chula (3.3%), lisa (3.3%), baqueta (3.2%), pargos (2.5%), dorado (2%) y tiburones (1.3%). El restante 16.9% incluye varias especies con capturas menores al 1% (Fig. 3).
0
5,000
10,000
15,000
20,000
25,000
30,000
35,000
40,000
45,000
50,000
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC
Ca
ptu
ra (
kg)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
216
Fig. 3. Composición de las capturas Lonja Pesquera Isla de la Piedra, Mazatlán en 2014. La CPUE mensual mínima fue de 70.7 kg/viaje en febrero, la máxima fue de 205 kg/viaje en agosto, el promedio mensual fue de 120.4 kg/viaje (Fig. 4).
Fig. 4. CPUE (kg/viaje) Lonja Pesquera Isla de la Piedra, Mazatlán en 2014. Aunque se explota una gran variedad de especies, sobresalen por sus volúmenes de captura el chihuil, la sierra y la berrugata que están en función de su abundancia, que a su vez puede estar influenciada por patrones ambientales estacionales (González-Becerril et al. 2000). Las variaciones mensuales del esfuerzo también está relacionada a la abundancia, fenómenos meteorológicos, además los factores socioeconómicos, ya que muchos pescadores ya no se dedican exclusivamente a la pesca y sólo realizan esta actividad cuando existe abundancia de especies de alto valor comercial. Se utilizó el viaje como medida del esfuerzo ya que simplifica la obtención de información necesaria para proponer medidas de manejo. Conclusión El esfuerzo de pesca varía según la presencia y abundancia de la variedad especies, muchas de las cuales aún no están reglamentadas por lo que es necesario realizar otros estudios de campo enfocados a los principales recursos de escama,
cuyos resultados contribuyan a su administración y aprovechamiento sustentable. Literatura citada Amezcua Linares F. 2008. Peces demersales del Pacífico
de México. Universidad Nacional Autónoma de México. Instituto de Ciencias del Mar y Limnología. México.
Espino Barr E, M Cruz Romero, A Garcia Boa. 2003. Peces marinos con valor comercial de la costa de Colima, México. Instituto Nacional de Pesca. CRIP Manzanillo. Comision Nacional para el Conocimiento y Uso de la Biodiversidad. México.
Fisher W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995 Guía FAO para la identificación de especies para fines de la pesca Pacífico centro-oriental. Volumen II Roma, Italia.
González Becerril A, E Espino Barr, M Cruz Romero, A Ruiz Luna. 2000. Determinación de la unidad de esfuerzo de pesca en una pesquería artesanal ribereña en Manzanillo, Colima, México. Ciencias Marinas 26(1): 113-124.
González-Becerril A, E Espino-Barr, A Ruiz-Luna, M Cruz-Romero. 2006. La pesca ribereña: Descripción, problemática y alternativas para su manejo. 611-622 pp. En: Jiménez-Quiroz MC, E Espino-Barr (Eds.). Los recursos pesqueros y acuícolas de Jalisco, Colima y Michoacán. INP, SAGARPA, 622p
Palabras clave: Composición, abundancia, recurso,
escama, CPUE.
Chihuil 20.7
Baqueta 3.2
Barrilete 5.7
Berrugata 10.5
Bullet 6.2
Pargos 2.5 Chile
verde 3.8 Chula 3.3 Lisa 3.3
Manta 3.9
Sierra 16.7
Dorado 2.0
Tiburones 1.3
Resto 16.9
0
50
100
150
200
250
ENE FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC
CP
UE
(kg
/via
je)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
217
105: oral PRESENCIA DE TIBURONES Y RAYAS EN LAS OPERACIONES DE ARRASTRE EN LA PESQUERÍA DE
CAMARÓN DEL GOLFO DE MÉXICO DURANTE 2013 Y 2014
Jorge L. Oviedo Pérez1, Heber Zea de la Cruz, Pedro C. Reyna González y Luis E. Martínez Cruz2
1Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. Av. Ejército Mexicano 106, Boca del Río, Veracruz, México. C. P. 94298. [email protected]; 2Centro Regional de Investigación Pesquera de Lerma. INAPESCA. Km 5, Carr. Campeche-Lerma, Campeche. C.P. 24500 Introducción La pesquería de arrastre de camarón es la más importante en el Golfo de México y Mar Caribe con una producción de 8,856 toneladas en peso vivo de camarón capturadas en mar abierto por una flota pesquera de 330 embarcaciones (SAGARPA 2014). La captura de tiburones y rayas asociados al arrastre ha sido poco estudiada. La relevancia que ha adquirido la explotación sustentable de elasmobranquios y la importancia de la pesquería, hacen prioritaria la determinación de su captura incidental en el arrastre camaronero. El objetivo del trabajo fue estimar el porcentaje de captura incidental de elasmobranquios en la pesquería de arrastre de camarón en el Golfo de México. Material y métodos Se realizó muestreo biológico pesquero a bordo de buques camaroneros en el Golfo de México (GM). Al inicio y fin de cada lance se registraron los datos de hora, posición geográfica, profundidad, velocidad, rumbo, tiempo de arrastre, distancia recorrida. Las capturas se clasificaron y cuantificaron en 6 grupos de especies: camarón, moluscos con valor comercial retenidos a bordo, otros crustáceos retenidos, peces óseos retenidos, elasmobranquios y descarte o fauna regresada al mar. Los elasmobranquios fueron identificados al nivel de especie, contabilizados en número de organismos, pesados por especie; de cada organismo se registró el sexo, en tiburones se tomaron longitud total (LT), longitud furcal (LF), longitud precaudal (LP); en rayas, según la especie, longitud total (LT), longitud del disco (LD) y ancho de disco (AD). La composición de la captura del sistema de arrastre (SA) fue expresada en porcentaje de contribución a la captura total en kilogramos para cada uno de los seis grupos de especies. La captura incidental de elasmobranquios en la pesquería de arrastre de camarón fue expresada en porcentaje en kilogramos de la captura total, de acuerdo con la fórmula:
100)/( te CCCI
Donde: CI = captura incidental de elasmobranquios, Ce = captura total de las especies de
elasmobranquios en kilogramos, Ct = captura total en el SA camaronero. La composición específica de elasmobranquios en el arrastre se determinó por especie en número de organismos y en peso (kg). Se analizó la estructura de longitudes de elasmobranquios por especie y sexos de las principales especies. La proporción sexual fue estimada como el cociente del número de hembras entre el número de machos. Para las principales especies de elasmobranquios se estimó el índice promedio de Captura Incidental por Unidad de Esfuerzo (CIPUE). De acuerdo con:
ii fC /CIPUEi
Donde: Ci es la captura total de todas las especies de elasmobranquios en el lance i en kilogramos; fi es el esfuerzo en horas de arrastre del lance i. El índice promedio de Captura Incidental por Unidad de Área (CIPUA) se estimó con:
ii AbC /CIPUAi
Donde: Ci es la captura en número de organismos y kg del lance i y Abi es el área barrida del lance i. Abi se estableció con:
iii ahvtiAb
Donde: ti y vi son el tiempo y la velocidad de arrastre, y ahí es la abertura operacional horizontal de la red considerada al 60 % (Klima 1976, Sparre y Venema 1997). Resultados y discusión En 2013 se monitorearon 13 cruceros y 277 lances. En 2014, 14 cruceros y 372 lances. En total se analizaron 649 lances de pesca (Fig. 1), realizados entre abril y septiembre, con un esfuerzo de 3,101 horas de arrastre, en un área barrida de 52,198 hectáreas, en profundidades de 11 a 89 m, con duración y velocidad de arrastre promedio de 4.8 horas y 2.7 nudos. La captura total fue de 290,054 kg, con camarón 61,418 kg (21.2%), moluscos retenidos a bordo 6,254 kg (2.2%), otros crustáceos retenidos 2,206 kg (0.8%), peces comerciales retenidos 24,653 kg (8.5%), elasmobranquios 9,620 kg (3.3%) y descartes 185,903 kg (64.1%) (Fig. 2).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
218
Fig. 1. Zona de muestreo / lances de pesca
Fig. 2. . Composición de la captura total.
Tabla 1. Composicion numérica y en peso RAYAS n % kg % Raja texana 5063 48 2005 21 Gymnura micrura 2957 28 3851 40 Narcine bancroftii 1100 10 647 7 Rhinobatos lentiginosus 338 3 234 2 Dasyatis americana 216 2 616 6 Dipturus olseni 51 0.5 61 1 Rhinoptera bonasus 3 0.03 27 0.3 TIBURONES n % kg % Squatina dumeril 550 5 1509 16 Squatina mexicana 167 2 540 6 Mustelus canis 54 1 114 1 Mustelus norrisi 6 0 .1 15 0.2 Sphyrna tiburo 3 0.03 3 0.03 Rhizoprionodon terraenovae
2 0.02 2 0.02
Carcharhinus acronotus 2 0.02 2 0.02 Carcharhinus brevipinna 1 0.01 2 0.02 Sphyrna lewini 1 0.01 1 0.01
TOTAL 10514 9630 Se registraron elasmobranquios en 619 de 649 lances, con una frecuencia de ocurrencia de 95%. Se contabilizaron 10,514 elasmobranquios con peso total de 9,630 kg. Las Rayas fueron 9,728 (92.5%) organismos con peso de 7,442 kg (77.3%). Los Tiburones fueron 786 (7.5%) organismos con peso de 2,188 kg (22.7%). Se identificaron 16 especies de 10 familias (Tabla 1).
La CIPUE promedio se estimó en 3.69 kg/hr y la CIPUA promedio en 0.23 kg/ha. R. texana se capturó entre 16 y 89 m profundidad, fue registrada en 518 de los 649 lances, con frecuencia de ocurrencia de 80%. La estructura de longitudes por sexos se presenta en la figura 3, el intervalo para las hembras(barras claras) fue de 9-66 cm de LT y para los machos (barras obscuras) de 9.5 a 65 cm LT. La proporción sexual hembras: machos fue de 1.14:1
Fig. 3. Estructura de longitudes de G. micrura
Conclusiones En 2013 y 2014 la captura incidental de elasmobranquios en la pesquería de camarón en el GM fue 3.3% de la captura total del SA. Los elasmobranquios fueron representados por 7 especies de rayas y 9 de tiburones. Numéricamente las rayas con 9,728 organismos representaron 92.5.8% de los elasmobranquios. En peso, con 7,442 kg representaron 77.3% de la captura. Las especies más importantes de rayas fueron R. texana. G. micrura y N. bancroftii, que en conjunto sumaron 9,120 organismos (86%) y 6,503 kg (67%). Las especies más importante de tiburones fueron S. dumeril y S. mexicana que en conjunto representaron 717 organismos (7%) y 2,049 kg (22%). Literatura citada SAGARPA-CONAPESCA. 2014. Anuario Estadístico de
Pesca y Acuacultura 2013. Mazatlán, Sinaloa, México. 225p.
Sparre P, SC Venema. 1997. Introduction to tropical fish stock assessment. Part 1. Manual. FAO Fisheries Technical Paper No. 306.1, rev. 2, Rome, FAO, 420 pp.
Palabras clave: Elasmobranquios, tiburones, rayas, incidentalidad, arrastre camaronero, Golfo de México.
Camarón 21.2%
Moluscos 2.2% Otros
crustáceos
0.8%
Peces comercial
es 8.5%
Elasmobranquios
3.3%
Descarte 64.1%
0
50
100
150
200
250
10
14
18
22
26
30
34
38
42
46
50
54
58
62
66
Nú
me
ro d
e o
rga
nis
mo
s
Longitud total (cm)
Raja texana n=3640 morfometrías

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
219
106: oral CARACTERIZACIÓN DE LA ACTIVIDAD PESQUERA EN LA LOCALIDAD DE RIACHUELOS,
MUNICIPIO TECOLUTLA, VERACRUZ, MÉXICO
Josué Martínez Trujillo, Jorge Luis Oviedo Pérez y Heber Zea de la Cruz
Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. Av. Ejército Mexicano 106, Col. Ylang, Boca del Río, Veracruz. C. P. 94298. [email protected]. Introducción La pesca ribereña tiene como características principales el empleo de una gran cantidad de fuerza de trabajo, la utilización de un amplio número de embarcaciones consideradas como flota menor (menores a 10 toneladas de arqueo neto), una tripulación muy reducida y escaso equipamiento (Flores Hernández et al. 2013). Material y métodos Entre julio y diciembre de 2015 se monitoreó la actividad pesquera. Se tomaron características técnicas de las unidades de pesca para describir embarcaciones, equipamiento y artes de pesca. Se realizó muestreo biológico pesquero de las descargas para determinar su composición en especies, longitudes y sexos. La información se procesó en hojas de cálculo electrónicas. Resultados y discusión La flota ribereña es de embarcaciones de fibra de vidrio con 7.62 m de eslora, equipadas con motores fuera de borda de hasta 75 hp y tripulación de 3 a 5 pescadores. Utilizan neveras con hielo, GPS y compás magnético. Faenan en la plataforma continental, con recorridos de hasta 30 mn y hasta 200 m de profundidad. Se censaron 13 lanchas, 4 con palangre, cimbra y red de enmalle; 4 sólo con red; 2 con palangre y cimbra, 2 sólo con palangre y 1 con palangre y red. Los palangres se usan para la pesca de especies de escama, rayas y tiburones, están armados hasta con 800 anzuelos tipo “garra de águila” de los números 8 y 9; que van sujetos sin alambrada a reinales de Poliamida Monofilamento (Pa Mono) de 1.2 mm de diámetro y 2.5 m de longitud que a su vez van sujetos a la línea madre de Pa Mono de 2 mm de diámetro. Utilizan como carnada muerta, principalmente bonito Euthynus alletteratus, chile vaquero Synodus foetens, lebrancha Mugil curema lacha Sardinella aurita, raya amarilla Gymnura micrura, raya tecolota Rhinoptera bonasus, sierra Scomberomorus maculatus y calamar. Su operación es diurna y dependiendo de la especie objetivo se calan fijas a fondo, media agua o superficie, con un tiempo de reposo de 1.5 a 2.5 horas, se hacen hasta 5 lances por faena. Las cimbras pescan tiburones, rayas y especies de escama. Están armados con hasta 400 anzuelos curvos “garra de águila” de los números 3 y 4, que
van sujetos con alambrada a reinales de Pa Mono de 2 mm de diámetro y hasta 5 m de longitud; que a su vez van sujetos a la línea madre de Pa Mono de 3 mm de diámetro. Utilizan carnada muerta principalmente morenas (Muraenidos), raya blanca D. americana, raya tecolota R. bonasus, bonito Euthynus alletteratus. Su operación es nocturna, se calan fijos a fondo con un tiempo de reposo de 24 horas. La cimbra es calada por la mañana. El resto de la faena se usa para curricanear con 3 anzuelos rectos enganchados entre sí, sujetos a una línea de Pa Mono de 0.8 a 1 mm de diámetro, con carnada de balajú Hemiramphus brasiliensis cortado por mitades. El curricán también se usa después de calar los palangres para escama. Las redes de enmalle se usan para la captura de sierra y cazones, están construidas de 15 a 25 secciones de paño de Pa Mono de 100 mallas de caída y tamaño de malla de 3”. Se usan para la pesca de sierra S. maculatus, cojinuda Caranx sp., ronco amarillo Conodon nobilis, cazón tripa Rhizoprionodon terraenovae, punterita Carcharhinus limbatus, C. brevipinna, cazón azul C. isodon, cazón limón C. acronotus, cornudita Sphyrna mokarran. Son caladas fijas a media agua y a superficie, el tiempo de reposo es de 1 hora con 5 lances por faena. Se contabilizaron 2,989 elasmobranquios. Correspondieron 2,709 a tiburones. De éstos, se identificaron 18 especies siendo la más importante R. terraenovae con 2,253 organismos (83.3%), C. limbatus con 230 (8.5%) y S. mokarran con 56 organismos (2%). El resto de las especies C. leucas, C. plumbeus, C. brevipinna, C. acronotus, C. falciformis, Mustelus norrisi, Isurus oxyrinchus, C. isodon, Galeocerdo cuvier, C. porosus, Hexanchus nakamurai, Squatina dumeril, Alopias superciliosus, S. lewini y S. tiburo con 179 individuos representaron 6.2% (Fig. 1).
Fig. 1. Composición porcentual numérica de tiburones.
83.16%
8.49%
2.06% 6.27%
R. terraenovae
C. limbatus
S. mokarran
otras especies

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
220
Con redes de enmalle se capturaron 1,557 tiburones, R. terraenovae con 1,406 organismos representó 90.3%; C. limbatus con 68 el 4.4%; S. mokarran con 35 el 2.2%. C. brevipinna con 12, C. falciformis con 13, C. isodon con 13, C. porosus con 2, C. acronotus con 7 y S. tiburo con 1 organismo representaron el 3.1%. La captura con palangre fue de 1,112 tiburones R. terraenovae con 847 organismos representó 76.2%; C. limbatus con 158 14.2%; S. mokarran con 20 1.8%; C. acronotus con 17 1.5%; las 13 especies restantes, con excepción de S. tiburo sumaron 70 tiburones que representaron 6.3%. La cimbra registró una captura de 40 tiburones, compuesta por C. leucas con 14 organismos que fue el 35%, C. plumbeus con 14 fue 35%, C. limbatus con 4 fue 10%, G. cuvier con 3 fue 7.5%, I. oxyrhinchus con 2 fue 5% y C. falciformis, S. mokarran, C. brevipinna con 1 individuo cada especie, 2.5% cada una. Para rayas se contabilizaron 280 organismos, correspondientes a 5 especies: Dasyatis americana, Gimnura micrura, Rhinoptera bonasus, Aetobatus narinari, Raja texana. Con palangre fueron capturadas 277 rayas, siendo D. americana la más importante con 194 organismos que representan el 70%; R. bonasus con 53 el 19.1%, G. micrura con 24 son 8.7% y A. narinari con 6 el 2.2%. La captura de peces óseos fue de 10,352 organismos, correspondientes a 42 especies. Con redes de enmalle se capturaron 7,750 piezas, de las que 7,048, 91% fueron sierra S. maculatus; cojinudas fueron 299, 3.9%; anchoas fueron 393, 5.1%; chopas Kyphosus sectatror fueron 10, 0.1%. La captura con palangre fue de 2,547 peces, las más importantes fueron huachinango Lutjanus campechanus con 828 organismos que representó 32.5%; pluma Calamus calamus con 302 organismos, 12%; bagre bandera Bagre marinus con 263, 10.2%; medregal Seriola dumerilli con 190, 7.5%. El resto de las especies con 964 organismos representaron el 37.8%. La captura con cimbra fue de 40 organismos, correspondieron 22, 55% a la cherna amarilla Epinephelus flavolimbatus; 11, 27.5% a la cubera Lutjanus cyanopterus y 7, 17.5% a pargo habanero Lutjanus analis
2%
70%
9%
19% A. narinari
D.
americana
G. micrura
Fig. 2. Composicion porcentual numerica de la captura de rayas con palangre Conclusión La actividad pesquera en la localidad de Riachuelos es artesanal y multiespecifica. La flota pesquera es menor y ribereña; equipada con motores fuera de borda, redes de enmalle, palangre escamero, cimbra tiburonera, curricán, neveras y GPS; opera en la plataforma continental. Se contabilizaron 13,341 organismos capturados, compuestos por 20.3% de tiburones, 2.1% de rayas y 77.6% de teleósteos. Los tiburones estuvieron representados por 18 especies, las más importantes fueron R. terraenovae, C. limbatus y S. mokarran. Las rayas fueron de 5 especies, las más importantes D. americana y R. bonasus. Los teleósteos fueron de 42 especies, las más importantes S. maculatus, L. campechanus, C. calamus, B. marinus, S. dumerilli, cojinuda y anchoa. Literatura citada Flores Hernández D, J Ramos Miranda. 2004. Las
pesquerías artesanales en el Golfo de México. El manejo costero. Universidad Autónoma de Campeche. México.
Palabras clave: Tiburones, Rayas, Tecolutla, Caracterización, Teleósteos.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
221
107: cartel TIBURONES CAPTURADOS POR LA FLOTA ARTESANAL DE PAREDÓN, CHIAPAS.
AVANCES PRELIMINARES
Cristina Celaya Castillo, Ana Bricia Guzmán Castellanos, Ana Alejandra Lizárraga Rodríguez, Adriana Jazmín Alatorre Alba y Oswaldo Morales Pacheco
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESACA. Prolongación Playa Abierta s/n, Col. Miramar Salina Cruz, Oax. C. P. 70680. [email protected] Introducción En México la captura de tiburón se realiza en zonas de pesca multiespecífica con gran variedad de longitudes y estadios de desarrollo, incluyendo hembras grávidas (Aguilar et al. 2009). En el Golfo de Tehuantepec destaca la productividad biológica (Aguilar-Castro et al. 2006) por lo que se considera importante para el uso de palangre (una sola línea de 400 a 800 anzuelos de tipo garra de águila núm. 16), redes de enmalle (de 10 m de longitud, 5 m de caída de red y luz de malla de 10 pulgadas) (Alavez-Jiménez et al. 2006). El objetivo del presente trabajo es conocer la pesquería actual de tiburones en Paredón, Chiapas, hacer un listado de las especies y sus longitudes mínima y máxima. Materiales y métodos Durante marzo y abril de 2016 se realizaron muestreos en los sitios de descarga de la flota artesanal de Paredón, Chiapas, que opera en la parte central de Golfo de Tehuantepec. Es el área de mayor captura de tiburón en comparación a otras de la misma región. Los tiburones se identificaron (Castro et al. 2008, Espinosa-Pérez et al. 2004). El sexo se determinó por la presencia o ausencia de gonopterigios, y se determinó la longitud total (LT): de la punta del hocico a la punta de la aleta caudal y longitud alternativa (LA): distancia del origen de la base de la primera aleta dorsal a la muesca precaudal. Los organismos fueron capturado por las cooperativas, permisionarios y embarcaciones libres de Paredón, Chiapas. Resultados y discusión Se identificaron 522 organismos en la pesquería de tiburón en la costa de Chiapas: 297 fueron machos y 225 hembras, pertenecientes a 4 familias, 4 géneros y 9 especies. Carcharhinus falciformis (307), Carcharhinus porosus (14), Carcharhinus limbatus (11), Carcharhinus leucas (2), Sphyrna lewini (151), Sphyrna zygaena (1), Alopias pelagicus (34), Isurus oxyrinchus (1) y Galeocerdo cuvier (1). La especie dominante fue C. falciformis, seguido de S. lewini (Fig. 1), que sostienen 88% la pesquería de Paredón.
Fig. 1. Dominancia de especies.
Este resultado es similar a lo reportado por Castillo-Geniz (2002) en Chiapas y Alejo-Plata et al. (2007) en Oaxaca (Tabla 1). Tabla 1. Longitudes por especie
Especie Longitud mín (cm)
Longitud máx (cm)
C. falciformis 71 242 C. porosus 81 123 C. limbatus 79 214 C. leucas 233 305 S. lewini 54.5 203 S. zygaena 190 A. pelagicus 190 271 I. oxyrinchus 130 G. cuvier 108
Para C. falciformis, Ortiz-Pérez (2011) reportó longitudes menores en Puerto Madero, Chiapas; Amezcua-Linares (2009) reportó C. porosus de mayor tamño, sin especificar la zona de captura. Agustín-Jiménez et al. (2006) registró para C. limbatus longitudes menores en Chipehua, Oaxaca. Cruz-Martínez et al. (2005) reportó longitudes menores en el Golfo de México. Para S. lewini se registró una longitud superior a la registrada por Bejarano-Álvarez et al. (2010) en Oaxaca y longitud máxima inferior a la registrada en este estudio. Hasta el momento se ha registrado sólo un ejemplar de S. zygaena (Tabla 1). Cruz-Jiménez et al. (2006) registraron en Oaxaca longitudes de A. pelagicus. Castellanos-Betancourt et al. (2013) tiene registros de tamaño mínimo y máximo de I. oxyrinchus, G. cuvier y S. lewini. La diferencia entre las longitudes mínimas y máximas
59%
3% 2%
0%
29%
0%
7%
0% 0%
Carcharhinus falciformis
Carcharhinus porosus
Carcharhinus limbatus
Carcharhinus leucas
Sphyrna lewini
Sphyrna zygaena
Alopias pelagicus
Isurus oxyrinchus
Galeocerdo cuvier

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
222
reportadas en este estudio a comparación de otros autores, puede deberse a las artes de pesca utilizadas, la zona y temporada de captura. Tabla 2. Longitudes (L) reportadas por otros autores para las especies registradas.
Autor Especie L mín. (cm)
L máx. (cm) Zona
Ortiz-Pérez (2011) C. falciformis 58 216 PM Chis. Amezcua-Linares (2009)
C. porosus 80 150 N/E
Agustín-Jiménez et al. (2006)
C. limbatus 130 209 Chipehua, Oax.
Cruz-Martínez et al (2005)
C. leucas 65 271 GM
Bejarano-Álvarez et al. (2010)
S. lewini 45 288 Oax.
Garza-Gisholt (2004) S. zygaena 110 283 BCS Cruz-Jiménez et al. (2006)
A. pelagicus 58.5 110 Oax.
Castellanos-Betancourt et al. (2013)
I. oxyrinchus 60 400 Ensenada, BC.
Castellanos-Betancourt et al. (2013)
G. cuvier 51 550 Pto. Chiapas
N/E = no especificado; GM = Golfo de México; PM = Puerto Madero, Chiapas
Fig. 2. A) C. falciformis, B) C. limbatus, C) C. leucas, D) C. porosus, E) S. lewini, F) S. zygaena, G) A. pelagicus, H) I. oxyrinchus, I) G. cuvier. Conclusiones Se registraron 9 especies de tiburones para esta zona (Fig. 2). Las que sostienen 88% de la pesquería son C. falciformis y S. lewini. La longitud mínima registrada es 54.5 cm y la máxima 305 cm Lt. Las artes de pesca utilizadas para la captura de estas especies de tiburones son palangre y red de enmalle. Literatura citada Aguilar CA, CJ Villavicencio-Garayzar, H Reyes-Bonilla, 2009.
Zonas y temporadas de reproducción y crianza de tiburones en el Golfo de California: Estrategia para su conservación y manejo pesquero. Ciencias Marinas 35(4): 369–388.
Aguilar Castro NA, AL Abitia Cárdenas, F Galván Magaña. 2006. Análisis trofo-dinámico del tiburón martillo Sphyrna lewini (Griffith & Smith, 1834) en el Golfo de Tehuantepec, México. Universidad Autónoma de Baja California Sur Departamento de Biología Marina.
Agustín Jiménez JA, F Galván Magaña, M Carrera Fernández. 2006. Descripción de la pesquería y aspectos reproductivos del tiburón puntas negras (Carcharhinus limbatus) y tiburón bironche (Rhizoprionodon longurio) capturados por la flota artesanal de Ensenada Chipehua, Oaxaca, durante el periodo octubre 2004 - octubre 2005.
Alávez-Jiménez C, F Galván-Magaña, M Carrera-Fernández. 2006. Distribución y abundancia de los tiburones sedoso (Carcharhinus falciformis Bibron in Müller y Henle; 1839) y pico blanco (Nasolamia velox, Gilbert in Jordan y Evermann; 1898), capturados por la flota artesanal pesquera de Ensenada Chipehua, Salina Cruz, Oaxaca.
Alejo-Plata C, JL Gómez-Márquez, S Ramos, E Herrera. 2007. Presencia de neonatos y juveniles del tiburón martillo Sphyrna lewini (Griffith y Smith 1834) y del tiburón sedoso Carcharhinus falciformis (Müller y Henle, 1839) en la costa de Oaxaca, México. Revista de Biología Marina y Oceanografía 42(3): 403 – 413.
Amezcua-Linares F. 2009. Peces demersales del Pacífico Mexicano. UNAM, ICMyL. México, D.F.
Bejarano-Álvarez M, F Galván-Magaña, RI Ochoa-Báez. 2010. Reproductive biology of the scalloped hammerhead shark Sphyrna lewini (Chondrichthyes: Sphyrnidae) off south-west Mexico. Aqua, International Journal of Ichthyology.
Castellanos-Betancourt JC, CE Ramírez Santiago, JL Castillo Géniz. 2013. Catálogo de aletas, trochos y cabezas de tiburones en el Pacífico mexicano. SAGARPA-INAPESCA.
Castillo-Geniz JL. 2002. Elasmobranquios del Golfo de Tehuantepec, litoral chiapaneco. Universidad Autónoma de Baja California. Informe técnico.
Castro JI, JL Castillo Geniz, JF Márquez Farías, R Talavera Villaseñor. 2008. Guía para la identificación de las especies de tiburones de importancia comercial en el Océano Pacífico. IV Edición.
Cruz-Jiménez CS, MC Alejo Plata, G Cerdenares Ladrón de Guevara. 2006. Estructura de tallas del tiburón zorro Alopias pelagicus (Nakamura 1935) capturado en la costa de Oaxaca, México.
Cruz-Martínez A, X Chiappa-Carrara, V Arenas-Fuentes. 2005. Age and growth of the Bull Shark, Carcharhinus leucas, from Southern Gulf of Mexico. J. Northw. Atl. Fish. Sci. 35: 367–374.
Espinosa Pérez H, JL Castro-Aguirre, L Huidobro Campos. 2004. Listados faunísticos de México. IX. Catálogo sistemático (Elamosbranchii: Selachimorpha). Instituto de Biología, UNAM.
Garza-Gisholt. 2004. Edad y crecimiento de Sphyrna zygaena (Linnaeus, 1758) en las costas de Baja California Sur, México. Tesis de licenciatura. Universidad Autónoma de Baja California Sur Área Interdisciplinaria de Ciencias del Mar Departamento de Biología Marina.
Ortiz-Pérez, 2011. Biología reproductiva del tiburón aleta de cartón. Carcharhinus falciformis, en la costa de Chiapas, México. Tesis de licenciatura. Universidad del Mar. Puerto Ángel, Oaxaca.
Palabras clave: Tiburones, Paredón, longitudes,
pesquería, listado.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
223
108: oral ASPECTOS REPRODUCTIVOS DEL MACABIL (Brycon guatemalensis)
EN LA CUENCA MEDIA DEL RIO GRIJALVA, CHIAPAS, MÉXICO
Eduardo Ramos Santiago1, Ma. Teresa Gaspar Dillanes2, Aldrin Labastida Che1, Yessica Xiomara Guzmán Camacho1, José Alfonso Oviedo Piamonte1 y Oswaldo Trejo Trejo2
1Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Pról. Playa abierta s/n. Col. Miramar. Salina Cruz, Oaxaca. C. P. 70680; [email protected]. 2Oficinas Centrales. INAPESCA. Pitágoras 1320. Col. Santa Cruz Atoyac. Del. Benito Juárez. Ciudad de México. C. P. 03310. Introducción La pesca de escama en embalses en Chiapas constituye una de las bases más importantes del desarrollo de una pesca comercial continental, pues da sustento y empleo a un gran número de familias, principalmente aquellas que ubican en comunidades indígenas. En la actualidad, la mayor producción pesquera en aguas interiores proviene de especies exóticas (tilapia, carpa y lobina); sin embargo, la diversidad de especies que albergan este tipo de ambientes hace factible que las especies nativas figuren dentro de la captura comercial. El macabil (Brycon guatemalensis) (Fig. 1) es la segunda especie más abundante y se explota comercialmente por pescadores indígenas del río Grijalva. La evaluación sugiere establecer criterios para su óptimo aprovechamiento, basado en aspectos biológicos, pesqueros, ambientales y sociales.
Fig. 1. Brycon guatemalensis Material y métodos Se realizaron muestreos mensuales de la captura comercial provenientes de las cooperativas pesqueras “Pescadores La Cuevita” y “El Indio Copainalteco” de febrero a diciembre 2014. Los organismos se midieron con un ictiómetro de 70 cm, el peso se obtuvo con una balanza digital de 5,000 g. A cada organismo se le determinó la longitud total (LT) en cm, peso total (g), sexo, peso de la gónada (g) y estadio de madurez gonádica, modificado de Nikolsky (1963), considerando sólo seis fases de madurez. Se obtuvo la estructura de longitudes, el índice gonadosomático (IG = [Pg/P]*100), la relación longitud-peso (P = a ∙ Ltb), la longitud de primera madurez sexual (Y = 1/[1+a(1-Lt/L0.5]) y el factor de condición según Vazzoler (1996).
Resultados Se analizaron 448 organismos que arrojaron un peso de 307.434 kg; 54% fueron machos y 46% hembras, una relación 0.9H:1M. La estructura de longitudes fue de 22 a 60.5 cm LT, y peso promedio de 686.23 g. El peso máximo registrado corresponde a un macho de 2934 g. La relación entre la longitud y el peso del macabil, describen un crecimiento isométrico: b no difiere estadísticamente de tres (tcal = 0.2512 < ttablas = 1.645) (Fig. 2).
Fig. 2. Relación longitud-peso. La media mensual del IG indica un incremento a partir de septiembre, con máximos de noviembre a mayo; y mínimos durante julio y agosto (Fig. 3).
0
4
8
12
FEB MAR ABR MAY JUN JUL AGO SEP OCT NOV DIC
±Std. Dev.±Std. Err.Mean
Fig. 3. Variación mensual del índice gonadosomático El análisis macroscópico de las gónadas indica mayor presencia de individuos juveniles de junio a octubre. Se observaron organismos en proceso de maduración durante todo el año, primordialmente en abril, junio, agosto y octubre. Los organismos
P = 0.0091Lt3.090 r² = 0.8012
n = 448
0
500
1000
1500
2000
2500
3000
3500
0 20 40 60 80
Pes
o (
g)
Longitud total (cm)
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
224
sexualmente maduros y en reproducción se presentaron en septiembre y de noviembre a junio. Individuos desovados se observaron en noviembre, diciembre, junio y julio (Fig. 4). El factor de condición muestra un proceso reproductivo principalmente durante los meses de noviembre a mayo (Fig. 5).
Fig. 4. Fases o estadios de madurez gonádica.
Fig. 5. Factor de condición para el macabil. El análisis de los organismos con base en la curva logística (L50), indica que esta especie alcanza una longitud de primera madurez de 37.8 cm LT (Fig.6).
Fig. 6. Longitud de primera madurez del macabil. Discusión Brycon guatemalensis es una especie de importancia comercial abundante en la cuenca media del río Grijalva durante enero, agosto y septiembre, coincidiendo con Mera-Ortiz et al. (2014). El ciclo reproductivo del macabil inicia en septiembre, con mayor intensidad de noviembre a mayo, similar con
Hernández-Portocarrero y Saborido-Rey (2009) para el Lago Nicaragua, con una temporada de desove de hembras entre julio y febrero. La longitud de primera madurez estimada en el presente estudio (L50 = 37.8 cm) fue similar al estimado por Hernández-Portocarrero y Saborido-Rey (2009) en el Lago Nicaragua (34.9 cm LT). De acuerdo con Hernández-Portocarrero y Saborido-Rey (2007), la fecundidad potencial está relacionada con la longitud, lo que indica que las hembras de mayor tamaño tienen mayor potencial reproductivo; lo que sugiere que abertura de malla del arte de pesca (red de enmalle) utilizado por los pescadores de 95.25 mm (3 ¾ plg), impactan en menor grado a los organismos con mayor capacidad reproductiva. Conclusión La longitud de primera madurez del macabil en el río Grijalva fue de 37.8 cm. El periodo reproductivo inicia en septiembre, con pulsos máximos de noviembre a mayo, por lo que las medidas de manejo en torno a esta pesqueria deberán establecer periodos de veda considerando estos meses. El arte de pesca actual es idóneo dado que la mayor captura incide en organismos con longitud por encima de la longitud de primera madurez; sin embargo, un estudio complementario de fecundidad y selectividad con redes de enmalle, sustentarían esta afirmación para las diversas especies que conforman la captura comercial en el río Grijalva. Literatura citada Hernández-Portocarrero A, F Saborido-Rey. 2007.
Abundancia relativa de los peces en la costa oriental del Lago de Nicaragua. Compendio de investigaciones de la ictiofauna de importancia comercial en Río San Juan y el Lago de Nicaragua. Volumen III. Managua: INPESCA. MARENA. 146p.
Hernández-Portocarrero A, F Saborido-Rey. 2009. Estrategia reproductiva de las hembras de Brycon guatemalensis en el Lago de Nicaragua y su importancia en la gestión pesquera. I Simposio iberoamericano de ecología reproductiva reclutamiento y pesquerías. Vigo, España. 24-28 de noviembre.
Nikolsky GV. 1963. The ecology of fishes. Academic Press. New York, EUA. 352p.
Mera-Ortiz G, E Velázquez-Velázquez, MA Peralta-Meixueiro. 2014. Aspectos biológicos pesqueros del macabil (Brycon guatemalensis) en la zona lótica del río Grijalva, municipio de Chicoasén, Chiapas, México. Memorias XIV Congreso Nacional de Ictiología. Morelia, Michoacán. México. 3 al 8 de noviembre.
Vazzoler A. 1996. Biologia da reproduçao da peixes teleosteos: teoria e prática. Maringá, EDUEM. São Paulo: Brasil. 169p.
Palabras clave: Brycon guatemalensis, ciclo reproductivo, longitud primera madurez, río Grijalva
0102030405060708090
100
feb. mar. abr may. jun. jul. ago. sep. oct. nov. dic.
% d
e M
adur
ez
Fase I Fase II Fase III Fase IV Fase V Fase VI
0
100
200
300
400
500
600
700
Nov Dic Feb Mar Abr May Jun Jul Ago Sep Oct
Fac
tor
de
Co
nd
ició
n
K
K´
0102030405060708090
100
30 32 34 36 38 40 42 44 46 48 50 52
Fre
cuen
cia
Acu
mul
ada
Longitud total (cm)
P = 1/ [1+e(20.873-0.549*Lc)]
n = 160

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
225
109: oral
RESULTADOS PRELIMINARES DEL PROGRAMA DE MARCADO CONVENCIONAL DE TIBURONES JUVENILES EN LA BAHÍA DE SEBASTIÁN VIZCAÍNO, BAJA CALIFORNIA 2010-2015
José Leonardo Castillo Geniz, Carlos Javier Godínez Padilla e Itzel Ortega Salgado
Centro Regional de Investigación Pesquera de Ensenada. INAPESCA: Carretera Tijuana-Ensenada Km 97.5, Parque Industrial Fondeport, El Sauzal de Rodríguez, Ensenada, Baja California. C.P. 22760. [email protected] Introducción En Baja California existen principalmente dos pesquerías de tiburones, la pesquería de palangre con embarcaciones de mediana altura que opera a lo largo de la costa occidental de la Península de Baja California cuyo puerto base es Ensenada (con un puerto alternativo ubicado en San Carlos, BCS), y la pesca artesanal con palangres y redes de enmalle en la que se emplean embarcaciones menores tipo “panga” y que opera en diversos campamentos pesqueros localizados en ambos litorales del estado. Las evidencias documentadas a partir de una serie de prospecciones de los desembarques de tiburones capturados por la flota artesanal que opera en Laguna Manuela (LM), campamento pesquero ubicado dentro de la Bahía Sebastián Vizcaíno (BSV), en la costa occidental de BC durante el periodo 2007-2011 (Castillo-Geniz et al. 2013), sugieren una segregación por longitudes y sexos de las principales especies de tiburones pelágicos juveniles capturados, tiburón azul, Prionace glauca y tiburón mako de aletas cortas, Isurus oxyrinchus, que sugieren que BSV puede servir como un área de refugio, nacimiento, y crianza para estas especies. Con la finalidad de conocer sus patrones de movimiento horizontal y tiempo de residencia de estas especies en BSV se inició en 2010, con el apoyo de los pescadores ribereños, un programa de marcado convencional de tiburones juveniles. Se presentan los primeros resultados del estudio. Material y métodos La etapa de campo consistió en colocar el mayor número posible de marcas convencionales tipo “espagueti” de colores en tiburones durante las actividades de pesca tradicional que realiza la flota pesquera artesanal del campamento de LM. Las embarcaciones menores fueron de fibra de vidrio de 20-25 pies de eslora con motor fuera de borda de 75-115 caballos de fuerza. Los equipos de pesca utilizados fueron palangres de superficie de 250-300 anzuelos tipo “garra de águila” (circular Mustad 0/16) usando como carnada macarela (Scomber japonicus) o lisa (Mugil cephalus). Cada tiburón que se capturó en buenas condiciones fue identificado, sexado y medido (sólo los tiburones < 100 cm LT pudieron ser pesados). De cada lance de pesca y marcado de tiburones se registró: Embarcación, fecha, número de tiburones
capturados por especie, condiciones climatológicas como nubosidad, condición del mar, corriente, temperatura superficial, y dirección viento, hora de inicio de lance y hora final de recobre, coordenadas de inicio de lance y final de recobre (GPS portátil) y tipo de carnada. Paralelamente al marcado de tiburones se llevó a cabo una campaña informativa sobre el pago de recompensas para aquellos tiburones marcados recapturados por los pescadores ribereños, por medio de carteles informativos y entrevistas. De cada marca recuperada se registró fecha de la captura, nombre del pescador, especie de tiburón, equipo de pesca empleado, y zona de pesca (coordenadas del lance de pesca). El análisis espacio-temporal de los tiburones marcados y sus recapturas hasta diciembre de 2015 se llevó a cabo mediante un análisis geográfico. Las recapturas se analizaron por sexo separado, calculándose el tiempo de libertad (en días) y la distancia recorrida en línea recta (km) de las marcas recuperadas en el periodo 2010-2015. Se elaboraron mapas donde se mostraron las trayectorias de recorrido de los tiburones recapturados empleando para ello el software ArcGIS (ESRI, 2011), y se analizaron estas trayectorias separadas por sexos. Resultados y discusión Durante los seis años del estudio de marcado-recaptura se han realizado 79 viajes de pesca y registrado en BSV 545 individuos de diez especies de tiburones y 3 individuos de dos especies de batoideos (Tabla 1). Se marcaron y liberaron en total 411 tiburones (75.4%) y todos los batoideos. El año con el mayor número de individuos marcados fue 2013 con 206 pertenecientes a 5 especies de tiburones, y 2015 con el menor número de tiburones etiquetados (24) de 7 especies de tiburones y dos de batoideos, la mayor diversidad observada en el presente estudio. El tiburón azul P. glauca y el tiburón mako de aletas cortas I. oxyrinchus fueron las principales especies que se capturaron y marcaron en BSV. Se capturaron en total 370 tiburones azules y 160 makos, etiquetándose 78.1% y 71.3%, respectivamente. La mayoría de los tiburones se capturaron en profundidades por debajo de la isobata de los 50 m (Fig. 1). Estas longitudes correspondieron a individuos juveniles y adultos de P. glauca. Todos los

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
226
tiburones mako capturados y marcados en BSV fueron juveniles y en algunos neonatos. Tabla 1. Número de tiburones capturados y marcados por especie en embarcaciones menores en Bahía Sebastián Vizcaíno, BC, durante 2010-2015
Nombre científico Nombre común
Núm. capturados*
Núm. marcados
Tiburones Prionace glauca Tiburón azul 370 289 Isurus oxyrinchus Mako 160 114 Sphyrna zygaena Cornuda
prieta 12 10
Cephaloscyllium ventriosum
Gata 7 3
Galeorhinus galeus Sulfin 6 6 Mustelus henlei Cazón
mamón 3 1
Squalus suckleyi Cazón espinoso
2 2
Alopias vulpinus Coludo pinto 2 1 Squatina californica Angelito 1 1
En total se han recapturado 73 tiburones marcados pertenecientes a 3 especies de tiburones en el periodo 2010-2015. Del tiburón azul se recuperaron en total 27 marcas recuperadas por la pesca ribereña y siete marcas colocadas en makos. Las tasas de recaptura para el tiburón azul y mako fueron de 9.3% y 4.4%, respectivamente. La distancia promedio recorrida (trayectoria lineal teórica) por hembras marcadas de P. glauca fue 249.2 ± 52.9 km con un intervalo de 13-1635.3 km; los machos recapturados recorrieron en promedio 267.33 ± 51.2 km con un intervalo de distancias de 24.1-1315.6 km. El intervalo de días de libertad de hembras de tiburón azul recapturadas fue de 9-481 días, con un promedio de 135 ± 24.5 días por hembra recapturada. Los machos recapturados tuvieron un intervalo de 12-318 días con un promedio de 113.2 ± 16 días. El intervalo de las distancias recorridas por las hembras marcadas de mako fue de 8.2-68.6 km, con un promedio de 29.1 ± 19.8 km, mientas que los machos presentaron un intervalo de 12.9-353 km con un promedio de 188.6 ± 94.2 km. Los días de libertad en promedio para las hembras de mako fue de 193.3 ± 96.8 días (con un intervalo de 80-386 días) y los machos presentaron un promedio de 147.8 ± 81 días de libertad (con un intervalo de 34-388 días).
Fig. 1. Ubicación geográfica de todos los tiburones capturados, marcados y liberados en Bahía Sebastián Vizcaíno durante 2010-2015. Las líneas continuas de color verde y rojo representan las isobatas de 20 y 50 brazas. Conclusiones El marcado convencional de tiburones neonatos y juveniles de tiburón azul y mako sugieren que ambas especies residen anualmente en BSV probablemente con la finalidad alimentarse y crecer para finalmente reclutarse a las poblaciones adultas en aguas oceánicas del Pacífico oriental. Literatura citada Castillo-Géniz JL, FJ Martínez-García, CJ Godínez-Padilla,
ME Jiménez-López, BE Velázquez-Martínez, M Serna-Sánchez, A García-Sánchez, FM Morineau-Escalante. 2013. Informe Final del Proyecto 2012 “Evaluación de la captura y esfuerzo de la pesca comercial de tiburón de Baja California y Programa piloto de marcado de tiburones en Bahía Sebastián Vizcaíno, B.C”. Centro Regional de Investigación Pesquera de Ensenada, B.C., Instituto Nacional de Pesca. Documento interno, 55 p.
ESRI. 2011. ArcGIS Desktop: Release 10.1. Redlands, CA Environmental Systems Research Institute.
Palabras clave: Tiburones, marcaje convencional, Bahía Sebastián Vizcaíno.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
227
110: oral ANÁLISIS DEL MONITOREO DE LAS CAPTURAS DE BATOIDEOS DE LA FLOTA RIBEREÑA DE
LAGUNA MANUELA, BAJA CALIFORNIA
Itzel Ortega Salgado, José Leonardo Castillo Geniz y Carlos Javier Godínez Padilla
Centro Regional de Investigación Pesquera de Ensenada. INAPESCA: Carretera Tijuana-Ensenada Km 97.5, Parque Industrial Fondeport, El Sauzal de Rodríguez, Ensenada, Baja California. C.P. 22760. [email protected] Introducción En el Pacífico mexicano la pesca artesanal de rayas (batoideos) genera valiosas fuentes de alimento, empleo y divisas. La mayor producción anual promedio de rayas se ha registrado durante 2000-2014 con 4,916 ± 211.2 t, y las mayores en Sonora y Baja California Sur (BCS). En Baja California (BC), la producción anual promedio de tiburón + cazón para el periodo 1976-2014 fue de 2,828.1 ± 156.2 t y durante 2000- 2014 la producción de rayas tuvo un promedio anual de 570.6 ± 36.7 t. Con la finalidad de caracterizar la pesca artesanal dirigida e incidental de elasmobranquios en la costa occidental de BC el Instituto Nacional de Pesca (INAPESCA) a través de su Centro Regional de Investigación Pesquera (CRIP) en Ensenada ha realizado el monitoreo sistemático de las capturas y desembarques de la pesca de lenguado y rayas con redes de enmalle de fondo de la flota artesanal de Laguna Manuela (LM), BC durante la temporada de pesca de lenguado y rayas en el periodo 2007-2015. Material y métodos Las rayas capturadas y desembarcadas por la flota de LM durante 2007−2015 fueron identificadas a nivel de especie, registrando el número de individuos y medidas morfométricas como LT (longitud total) y AD (ancho de disco) en el caso de las rayas miliobatiformes, y únicamente LT en el caso de rayas rinobatiformes. El sexado se llevó a cabo por medio de la presencia-ausencia de órganos copuladores. Se calculó una CPUE por cada 100 m de red, calculándose el total de metros de red utilizados en cada viaje de pesca por mes. Se realizaron los análisis de la relación peso-longitud por sexos y su comparación mediante pruebas de ANCOVA. La proporción sexual se analizó mediante pruebas de chi cuadrada (Castillo-Géniz et al. 2015). A partir de los lances de pesca con redes de enmalle registrados en cada mes durante el periodo de estudio se calculó un promedio aritmético de las rayas capturadas por viaje de pesca. Con los datos del número de metros de red registrados por cada viaje de pesca monitoreado se calculó la CPUE nominal numérica de rayas por cada cien metros de red. Resultados Durante 2007-2015 se han documentado 955 viajes de pesca en LM de los cuales 73.5% emplearon redes de enmalle de fondo para la captura de
lenguados, tiburones, rayas y calamares. Los muestreos de campo han registrado 29,343 individuos, de los cuales 35.7% pertenecen a rayas. En el mismo periodo se han documentado 10,487 batoideos de trece especies diferentes, de las cuales se desprende que las especies más frecuentes fueron Zapterix exasperata (42.3%), Rhinobatos productus (36.5%) y Myliobatis californica (11.7%) (Fig. 1). Estas tres especies representaron 90.5% del total de las capturas de batoideos en Laguna Manuela, el resto son mil individuos que se encuentran repartidos entre las ocho especies restantes.
Fig. 1. Composición específica de las capturas por grupos taxonómicos y especies desembarcadas de batoideos por la flota artesanal de LM (2007-2015). Desde 2012, la veda de tiburones y rayas afecta los muestreos durante los meses de mayo, junio y julio, ya que no se desembarca ningún individuo. El registro de los desembarques de las capturas de Zapterix exasperata (guitarra prieta) y Rhinobatos productus (guitarra blanca) mostró una estacionalidad definida, un primer pico en abril y otro en agosto, donde se observaron las mayores capturas de las mismas y de Myliobatis californica (Fig. 2). El comportamiento mostrado podría estar sesgado por la veda de noventa días, ya que los registros de captura se redujeron durante los meses de verano.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
228
Fig. 2. Estacionalidad mensual de las capturas (núm. individuos) de las principales especies de rayas documentadas en la flota artesanal de LM. Se examinaron 1,154 guitarras prietas, 587 fueron hembras y 567 fueron machos. La relación sexual fue equitativa 1:1 y el intervalo de longitudes de hembras fue de 15-110 cm LT con un promedio de 83.8 ± 0.4 cm LT. Los machos tuvieron un intervalo idéntico de 15-110 cm LT con un promedio de 80.0 ± 0.3 cm LT. Las capturas con redes de enmalle de esta especie permiten ubicar su población en profundidades menores a los 50 m. De la guitarra blanca R. productus se examinaron 519 individuos, que resultaron en 357 hembras y 162 machos. Las hembras presentaron un intervalo de longitudes de 32-200 cm LT con un promedio de 106.2 ± 1.0 cm LT, mientras que los machos presentaron un intervalo de longitudes de 52- 125 cm LT con una media de 101.1 ± 0.9 cm LT. La razón sexual de X2
(0.05)(1) = 73.27.
Fig. 3. Valores de captura mensual promedio por viajes de pesca y CPUE de guitarra prieta Z. exasperata (a, b) y guitarra blanca R. productus (c, d), de la flota de Laguna Manuela durante 2007- 2015. Se calculó una CPUE por cada 100 m de red, calculándose el total de metros de red utilizados en cada viaje de pesca por mes. El mayor CPUE para la guitarra prieta se dio en septiembre con 2.19 rayas por cada cien metros de red y en agosto con 1.9
rayas por cada cien metros de red en el caso de la guitarra prieta (Fig. 3). Todos los valores se muestran influenciados por el tamaño de muestra obtenido en cada uno de los meses. Conclusiones Zapterix exasperata, Rhinobatos productus y Myliobatis californica son las tres especies más importantes de batoideos en la pesca con redes de enmalle en Laguna Manuela, BC. La mayor CPUE para Zapterix exasperata, guitarra prieta, se calculó en septiembre con 2.19 rayas por cada cien metros de red. La mayor CPUE para Rhinobatos productus, guitarra blanca, se calculó para agosto con 1.9 rayas por cada cien metros de red. En 2015 destaca la aparición de Mobula thurstoni, que en años anteriores no había sido detectada en la zona. Literatura citada Castillo-Géniz JL. 2015. Estudios sobre las pesquerías de
tiburones y rayas del noroeste del Pacífico mexicano: Investigaciones aplicadas para el mejoramiento de su manejo pesquero y conservación, 2015. Informe Final POA 2015 (Documento Interno). CRIP-Ensenada. INAPESCA.
Palabras clave: Batoideos, rayas, pesca ribereña, redes de enmalle, CPUE.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
229
111: oral COMPOSICIÓN Y ANÁLISIS DE VOLUMEN DE CAPTURA DE ESPECIES COMERCIALES DE PECES EN
EL MERCADO “LA NUEVA VIGA” DE LA CIUDAD DE MÉXICO
Patricia Fuentes Mata, Gabriela Morales García, Israel Montoya Carvantes, Georgina Ramírez Soberón, José Ignacio Fernández Méndez y Miriam Nava Abarca
Oficinas Centrales. INAPESCA. Pitágoras 1320, Col. Santa Cruz Atoyac, Ciudad de México. C. P. 03310. [email protected]. Introducción En 1993 fue inaugurado el mercado de pescados y mariscos “La Nueva Viga”, en Iztacalco en la Ciudad de México, ante la necesidad de ofrecer un espacio mayor a una población en constante crecimiento para satisfacer su demanda de productos pesqueros. Aquí se genera una actividad económica importante y se comercializa entre 3% y 6% de la producción nacional de escama, moluscos y crustáceos. Se venden productos de todos los puertos de la República Mexicana y se han incrementado las importaciones, principalmente de Asia. Se consideró necesario hacer un diagnóstico general de los recursos disponibles a la venta, como punto de partida para generar políticas públicas orientadas a promover el consumo de productos pesqueros nacionales entre la población. El objetivo principal de este trabajo fue determinar la composición de especies que conforman el recurso escama, clasificándolas en: de alto valor, de precio medio y de bajo valor comercial; asimismo, analizar su volumen de captura, reconocer su procedencia por litoral y región del país e identificar la temporada de las principales especies a lo largo del ciclo anual. Materiales y métodos Se hicieron 12 muestreos durante 2015, uno por mes con las siguientes actividades: 1. Identificar la composición de las especies, su procedencia, tamaño y precio durante el ciclo anual. 2. Conservar especímenes de las principales especies comerciales para verificar su identidad taxonómica y elaborar el registro fotográfico. 3. Se diseñaron encuestas para recabar datos directos por bodega sobre procedencia de las especies por litoral y región del país; reconocer las “especies de temporada” y sus precios. 4. Sistematizar y analizar los registros de bases de datos (2010-primera mitad del 2015) de la Oficina de Pesca, empleando programas escritos (scripts) en el programa estadístico R, para calcular el volumen de captura de productos nacionales e importados, de origen marino y dulceacuícola. En el trabajo de laboratorio se revisó un lote de cada especie y se utilizaron claves taxonómicas, catálogos sistemáticos, revisiones monográficas y descripciones originales. Se consultaron: Miller y Lea (1972), Fischer et al. (1995), Eschmeyer et al. (1983), Castro-Aguirre y Espinosa-Pérez (1996), Castro
Aguirre et al. (1999), Allen y Robertson (1994), Espinosa Pérez et al. (2004). Resultados Se identificaron 122 especies de peces de 40 familias y 71 géneros. Las familias que reúnen 40% son: Carangidae (19 spp), Haemulidae (12spp), Sciaenidae (11 spp) y Serranidae- que incluye Epinephelidae (9 spp). En cuanto a la procedencia de las especies (Fig. 1), 77% proviene del Golfo de México y Caribe; 21% del Pacífico; y 2% de aguas continentales.
Fig. 1. Procedencia de las especies por litoral.
Tabla 1. Clasificación del producto por precio y tamaño. $ kg 170-200 140-169 100-139 80-99
PR
IME
RA
robalo* atún* cherna*
huachinango mero* extraviado* abadejo* negrillo*
pargos besugo cabrilla dorado* lenguado*
pámpano esmedregal constantino cobia
$ kg 50-79 40-49 29-39 15-28
SE
GU
ND
A cazón*
raya tiburón* sierra peto*
tilapia trucha-mar barrilete bagre bandera
mojarrón sargo lisa chac-chí cojinuda
jurel mojarra-mar cintilla burros jorobado
La tabla 1 muestra las especies de mayor disponibilidad en La Nueva Viga. De la 1ª calidad, hay especímenes grandes* (>50 cm). En la 2ª calidad, predominan especies de longitudes moderadas (en promedio 28 cm). Se presentan ejemplos de la disponibilidad de especies de temporada durante el ciclo anual.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
230
Tabla 2. Especies del Golfo de México y Pacífico que se comercializan en su temporada en La Nueva Viga.
Análisis del volumen de captura. Los datos registrados desde 2010 hasta 2015 (Fig. 2), señalan un mayor volumen de captura durante los meses de febrero y marzo, registrando su punto más alto en 2011, con un volumen total de 6,442 toneladas para el mes de marzo. En contraste, el menor de volumen se presenta por lo general desde septiembre-octubre hasta el final del año. La excepción a esto último fue en octubre del 2010, con 4,286 t. Los promedios anuales de volumen de captura (Fig. 3) muestran que 2014 fue el más alto, con 38,228 t, mientras que 2013, fue menor con 28,352 t.
Fig. 2. Producto pesquero (t) por mes de 2010 a2015.
Fig. 3. Promedios anuales de volumen de captura (en toneladas) en el período 2010-2015.
Discusión y conclusiones La composición de especies disponible en la mayor parte del año es de capturas ribereñas y artesanales del litoral del Golfo de México. Hay un incremento de productos de importación procedentes de la acuacultura: tilapia, basa y salmón; existe una preferencia por comercializar volúmenes grandes de ciertas especies (i.e. sierra, corvina, esmedregal y jurel) a precios competitivos. El grupo de huachinango, pargo, mero, cabrilla y robalo son especies de alta demanda y alcanzan precios elevados. Los puntos de venta del mercado señalan que los productos más ofertados son el cazón, huachinango y jureles del Golfo de México; el huachinango y el besugo son las más apreciadas a nivel nacional, por su sabor delicado, carne suave y blanca. Una variedad de peces de calidad diversa se mezclan y ofrecen al consumidor como huachinango, con tal de subir los precios. Esto es posible, pues el consumidor promedio desconoce las diferencias entre las especies y prefiere la presentación en filete, además de que tienden a adquirir el producto importado contra el nacional. El promedio anual del producto que oferta La Nueva Viga es de 35,561 t, lo cual cubre parcialmente la necesidad alimentaria de la población de la CDMX. La mejor temporada por la disponibilidad de especies ocurre desde la semana mayor hasta mayo. Los pescados y mariscos siguen siendo secundarios en la preferencia de la dieta del consumidor del área metropolitana, parte del problema es falta de infraestructura que asegure una mejor calidad del producto. Literatura citada Allen GR, DR Robertson. 1994. Fishes of the tropical
eastern Pacific. University of Hawaii Press. Hawaii, EUA. 332p.
Castro-Aguirre JL, H. Espinosa-Pérez. 1996 Catálogo sistemático de las rayas y especies afines de Méx. Listados faunísticos de México, VII. IBUNAM. 75p.
Castro-Aguirre JL, H Espinosa-Pérez, JJ Schmitter-Soto. 1999. Ictiofauna estuarino lagunar y vicaria de México. Ser. Biotec. Limusa-IPN. 711p.
Eschmeyer WN, ESHerald, H Hammann. 1983. A field guide to Pacific coast fishes of N Am. NY. 336p
Espinosa Pérez H, JL Castro-Aguirre, L Huidobro. 2004. Catálogo sistemático de tiburones y especies afines de México (Elasmobranchii: Selachimorpha). Listados faunísticos de México IX. IBUNAM. 137p.
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía para la identificación de especies para los fines de la pesca, Pacífico Centro- Oriental. Rome FAO. 1,646p.
Miller DJ, RN Lea 1972. Guide to the coastal marine fishes of California. Calif. Fish and Game 157: 235p.
Palabras Clave: Volumen de captura, peces marinos, pesquería, comercio, consumo, producción.
Especies Golfo/Caribe E F M A M J J A S O N D
BARRACUDA Sphyraena barracuda
ESMEDREGAL Seriola zonata
HUACHINANGO Lutjanus campechanus
MERO ROJO Epinephelus morio
ROBALO BLANCO Centropomus undecimalis
SIERRA Scomberomorus maculatus
Especies del Pacífico E F M A M J J A S O N D
SIERRA Scomberomorus sierra
JUREL ALETA AMARILLA Seriola lalandi
LENGUADO Paralichthys californicus
CORVINA Cynoscion parvipinnis
PARGO Lutjanus argentiventris
HUACHINANGO Lutjanus peru

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
231
112: oral EFECTO DE LOS DIFERENTES TIPOS DE ANZUELO UTILIZADOS POR LA FLOTA ARTESANAL, SOBRE LA COMPOSICIÓN DE ESPECIES Y LONGITUDES DEL MERO ROJO (Epinephelus morio), EN LA ZONA
COSTERA DEL ESTADO DE YUCATÁN
Carmen Monroy, Humberto Medina y Mariana Uribe
Centro Regional de Investigaciones Pesqueras de Yucalpetén, INAPESCA. [email protected] Introducción La plataforma de la Península de Yucatán es una región de gran importancia pesquera por ser hábitat natural de numerosas especies comerciales, entre las que destaca por su importancia económica la pesquería de mero que durante muchos años fue sostenida por una sola especie el mero rojo (Epinephelus morio). Esta pesquería, desde el punto de vista socio-económico, ha sido una de las actividades de gran relevancia para las comunidades costeras del Estado. En esta pesquería participan dos flotas con diferente poder de pesca (mediana altura y artesanal y/o ribereña), ambas operan en la Plataforma de la Península de Yucatán bajo un marco de pesquería secuencial con interacciones tecnológicas, que explotan diferentes componentes de la población de E. morio (Monroy 1998, Seijo 2007). La flota artesanal incide sobre la fracción juvenil de la población y opera en aguas someras a lo largo del litoral yucateco, desplazándose desde el Suroeste de Celestún y siguiendo el contorno de la isobata de las 20 brazas, hasta llegar al extremo este (Contoy). La flota artesanal, en su actividad pesquera, utiliza diferentes sistemas de captura. El anzuelo (línea o palangre de mano) es el principal método de pesca; se utilizan dos tipos de anzuelos de diferentes tamaños: el circular o tipo huachinanguero y el recto o tipo J con dos modalidades: pata larga (PL) y pata corta (PC). Los meros presentan ciertas características en su estrategia de vida que los hacen más susceptibles a la sobreexplotación (Coleman et al. 1996). Actualmente, el stock de E. morio del Banco de Campeche está considerado sobreexplotado y uno de los factores que han contribuido a la situación actual del recurso es la sobre-pesca de juveniles (DOF 2012, Monroy et al. 2014). El objetivo de este trabajo fue analizar el efecto de los diferentes tamaños y tipos de anzuelo, en la composición de longitudes de E. morio capturadas por la flota artesanal, para evaluar el cumplimiento de la longitud mínima de captura de esta especie (NOM-065-SAG/PESC-2014: DOF 14/07/2015). Material y método Se analizó información de 457 viajes realizados por la flota artesanal, entre 2013 y 2015. Los viajes
fueron de ida y vuelta el mismo día y se realizaron entre las 1.5 y 31.5 brazas de profundidad. En cada viaje se registró: la zona de pesca, la profundidad y las características del arte de pesca (tipo y tamaño de anzuelo). Asimismo, se registró la captura por especie desembarcada y todos los organismos capturados de mero rojo fueron medidos (longitud total, LT y longitud furcal, LF; cm) y pesados (peso eviscerado, PE; g). Se analizaron los tres tipos de anzuelos utilizados por la flota artesanal, pero sólo se consideraron los tamaños que son más utilizados por los pescadores: anzuelo pata larga y pata corta 8/0 y 9/0 y los anzuelos circulares del 8/0, 9/0, 10/0, 11/0 y 12/0. Los anzuelos circulares fueron utilizados principalmente con el palangre de mano de 120 a 600 anzuelos y los rectos o tipo “J” con el cordel o línea de mano. Se estimó por zona de pesca y por tipo y tamaño de anzuelo (recto y circular), el perfil de la captura y la captura por unidad de esfuerzo (CPUE), total y de la captura de mero rojo. Las comunidades pesqueras, fueron divididas en cuatro zonas de pesca: poniente (Celestún y Sisal); centro (Progreso, Chuburna y Techac); Dzilam de Bravo y Oriente (San Felipe y Rio Lagartos). La distribución de frecuencia de longitudes de los diferentes tipos y tamaños de anzuelo, se comparó con la prueba no-paramétrica de Kolmogorov-Smirnov, ya que los datos no cumplían con los supuestos de normalidad y homogeneidad de varianza. Con la distribución de frecuencia de longitudes por tipo y tamaño de anzuelo, se estimó la proporción de organismos capturados de mero rojo, por abajo de la longitud mínima de captura (TMC), establecida en la NOM 065-SAG/PESC-2014 y con una longitud menor a la longitud en la que el 50% de las hembras son maduras (L50 =50.9 LF), reportada para la especie por Brulé et al. (1999). La TMC fue convertida a longitud furcal (TMC =31.5 LF). Resultados y discusión La actividad pesquera de la flota artesanal en la zona costera de Yucatán se ha centrado principalmente en la explotación de peces demersales. Si bien, el esfuerzo de pesca se ha dirigido a la captura del mero rojo (E. morio), actualmente esta pesquería es multi-específica.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
232
El análisis de la información muestra que la captura estuvo conformada de gran diversidad de especies. Los más representados fueron cinco especies de dos familias: Serranidae (E. morio), Lutjanidae (Ocyurus chrysurus, Lutjanus synagris, L. analis y L. griseus). El tipo de anzuelo y la zona de pesca, son factores importantes que definen la composición de especies en la captura. Por ejemplo, los pescadores de la zona oriente utilizan con mayor frecuencia el anzuelo circular con el palangre de mano y 81.3% de la captura fue mero rojo. En cambio, los pescadores de la zona poniente utilizan principalmente el anzuelo recto, donde sólo 22.5% de la captura, fue de mero rojo. En este caso, el decremento de E. morio, ha sido sustituido con el incremento en la captura de rubia (L. synagris) y canané (O. chrysurus), pero también se registraron otras especies de importancia, para el consumo de subsistencia como: el chac chi (Haemulon sp.) y las mojarras (Calamus sp.). Un total de 4,885 meros de E. morio fueron capturados con los diferentes tipos y tamaños de anzuelo; el anzuelo recto PL y circular fueron los que registraron mayor número de mero capturados (Tabla 1). Se registraron diferentes valores de la longitud media (LF), con los diferentes tipos y tamaños de anzuelo. Sin embargo, con los anzuelos rectos PL y PC, no se registró un incremento conforme al tamaño del anzuelo (Tabla 1). Tabla 1. Características estadísticas de la longitud furcal (LF), de los meros de E. morio, capturados por la flota artesanal, con diferentes tipos y tamaños de anzuelos.
Anzuelo N Media Std.Dev. Std.Err. Mediana
Recto PL 8 1,970 37.0 5.9 0.13 36.0 9 1,461 36.3 5.6 0.15 35.0
Recto PC 8 392 37.3 5.6 0.28 36.0 9 152 36.0 6.1 0.49 34.5
Circular 9 482 39.9 7.4 0.34 38
10 2,093 40.7 7.2 0.16 39 11 1,749 40.6 7.3 0.17 39 12 561 42.7 5.2 0.22 42
La distribución de frecuencia de longitudes de la captura, registrada con los anzuelos rectos (8 vs 9), presentaron diferencia significativas (prueba de Kolmogorov-Smirnov: p < 0.001). En la combinación de los anzuelos circulares 10 vs 11, no se mostró diferencia significativa (Kolmogorov-Smirnov: p = 0.261). El porcentaje de meros (E. morio), capturados con una longitud inferior a la TMC, con los diferentes tipos y tamaños de anzuelos rectos, fue alto (43.7% - 56.6%). Este porcentaje de meros pequeños en la
captura disminuyo con los anzuelos circulares (6.1% - 28.2%). Si se considera la longitud donde el 50% de las hembras son maduras (L50), se incrementó el porcentaje de organismos capturados (89.8-98.0), con todos los tipos y tamaños de anzuelos. Literatura citada Coleman FC, CC Koenig, LA Collins. 1996. Reproductive
styles of shallow-water grouper (Pisces: Serranidae) in the eastern Gulf of Mexico and the consequences of fishing spawning aggregations. Environmental Biology of Fishes 47: 129-141.
DOF. 2012. Carta Nacional Pesquera. Diario Oficial de la Federación. México. 24 de agosto de 2012.
DOF. 2015. Norma Oficial Mexicana NOM 065-SAG/PESC-2014, Para regular el aprovechamiento de las especies de mero y especies asociadas, en aguas de jurisdicción federal del litoral del Golfo de México y Mar Caribe. Diario Oficial de la Federación, 14 de julio.
Monroy C. 1998. Análisis bio-económico de la pesquería del mero (Epinephelus morio) en el Banco de Campeche bajo condiciones de riesgo incertidumbre. Tesis de Maestría, CINVESTAV, Unidad Mérida, Yucatán, México.
Monroy C, A Hernández, G Galindo. 2014. La pesquería de mero Epinephelus morio en la Península de Yucatán. En: MA Cisneros-Mata, LF Beléndez-Moreno, E Zárate-Becerra, MT Gaspar-Dillanes, LC López-González, C Saucedo-Ruíz, J Tovar-Ávila (eds.). Sustentabilidad y pesca responsable en México. Evaluación y manejo 1999-2000. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, pp: 587-614.
Seijo JC. 2007. Estimación de riesgo de exceder puntos de referencia límite por sobrecapacidad del esfuerzo en pesquerías secuenciales: la pesquería de mero (Epinephelus morio) de la plataforma continental de Yucatán. En: Agüero M (ed.). Capacidad de pesca y manejo pesquero en América Latina y el Caribe. FAO Doc. Tec. Pesca. Roma. 461, 403p.
Palabras clave: Epinephelus morio, anzuelo, longitud mínima de captura, composición de la captura.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
233
113: oral TENDENCIA DE LA PRODUCCIÓN DE RAYAS (BATOIDEA) EN LA COSTA DE TABASCO: CON ÉNFASIS
EN Dasyatis americana
Raúl E. Lara Mendoza, Ángel R. Galeana Cortazar y Ana G. Díaz Álvarez
Centro Regional de Investigaciones Pesqueras de Ciudad del Carmen. INAPESCA. Av. Héroes del 21 de Abril, No 26, Col. Playa Norte. Ciudad del Carmen, Campeche. C.P. 24100. e-mail: [email protected] Introducción Al igual que otras pesquerías ribereñas del país, la pesquería artesanal de tiburones y rayas es una actividad de suma relevancia socioeconómica en la región del Golfo de México (GDM) y mar Caribe (MC), debido a que representan una alternativa de subsistencia para las comunidades pesqueras, ya que es una fuente generadora de empleo y alimento (Castillo-Géniz 1992). A diferencia de otros recursos pesqueros que soportan una intensa explotación comercial, como la sardina y el camarón, las poblaciones de elasmobranquios pueden ser abatidas más fácilmente, inclusive con niveles modestos de mortalidad por pesca, debido a que poseen un bajo potencial reproductivo, resultado de características biológicas que incluye un crecimiento lento, alta longevidad, una madurez sexual tardía, baja fecundidad y una estrecha relación denso-dependiente entre reclutas y reproductores (Bonfil 1994, Musick 1999, Cortés 2007). La pesquería de tiburones, rayas y especies relativas en el GDM y MC son muy importantes en términos de producción, ya que en los últimos 15 años se han reportado cifras de hasta 10,351 toneladas (durante el 2000), representando 23.5% del total nacional (producción de 2000-2014) (CONAPESCA 2014). Históricamente, la producción pesquera de estas especies se ha llevado a cabo en el noroeste del GDM (Veracruz y Tamaulipas), aunque en los últimos cuatro años la producción de los estados de Tabasco y Campeche han rebasado la producción a nivel regional, debido a la disminución de las capturas en los estados del noroeste del GDM. En particular, la pesquería rayas (principalmente balá, Dasyatis americana) en el estado de Tabasco representa un fuerte impacto social y económico para la región, debido a que es una importante fuente de alimento y empleos directos e indirectos a lo largo del año. Debido a lo anterior, el objetivo del presente estudio es describir y dimensionar la magnitud y tendencia de la captura de rayas (Batoidea), en específico de la raya denominada balá, Dasyatis americana, la cual es una de las principales especies que es capturada de forma dirigida e incidental por la pesquería ribereña de palangre y que sirva como un primer acercamiento para estimar el estado de explotación de esta especie de la que se carece información biológica y pesquera.
Material y métodos Se recopiló la información pesquera contenida en los avisos de arribo de la Subdelegación de Pesca del estado de Tabasco para los años 2006-2012; así como, las libretas de pesca de los principales centros de acopio a lo largo de la costa de Tabasco. La información recabada fue analizada separando al grupo de cazones, tiburones, las rayas y en específico al balá (D. americana) para hacer los análisis mensuales e interanuales de su captura. Información adicional de las operaciones de pesca de estas especies se recopilaron in situ en los principales sitios de desembarque. Para determinar la existencia de diferencias significativas entre los años evaluados, se aplicó un estadístico no paramétrico Kruskal-Wallis (H) debido a que estos no presentaron una distribución normal. Resultados y discusión La producción de rayas en el GDM y MC tiene sus inicios con los primeros reportes en 1986 con 636 t, mientras que, la producción para el estado de Tabasco en 1990, con su máximo histórico en el 2011 aportando 1,697 t (Fig. 1).
Fig. 1. Serie histórica de producción de elasmobranquios (todas las especies) y rayas a nivel nacional, Golfo de México y mar Caribe (GDMyMC). En la costa del estado de Tabasco, la producción la pesca de rayas esta soportada principalmente por la especie comúnmente denominada balá, D. americana, aunque también son capturadas incidentalmente y en menor intensidad otras especies como la chucha pinta (Aetobatus narinari), el chucho (Rhinoptera bonasus), la raya cola de rata o mariposa (Gymnura micrura) y la guitarra (Rhinobatos lentiginosus). D. americana es la especie con mayor producción, reportándose entre
0
5
10
15
20
25
30
35
40
1970 1975 1980 1985 1990 1995 2000 2005 2010 2015 2020
Ton
elad
as (x
100
0)
Elasmobranquios total Elasmobranquios GDMyMC
Rayas total Rayas GDMyMC
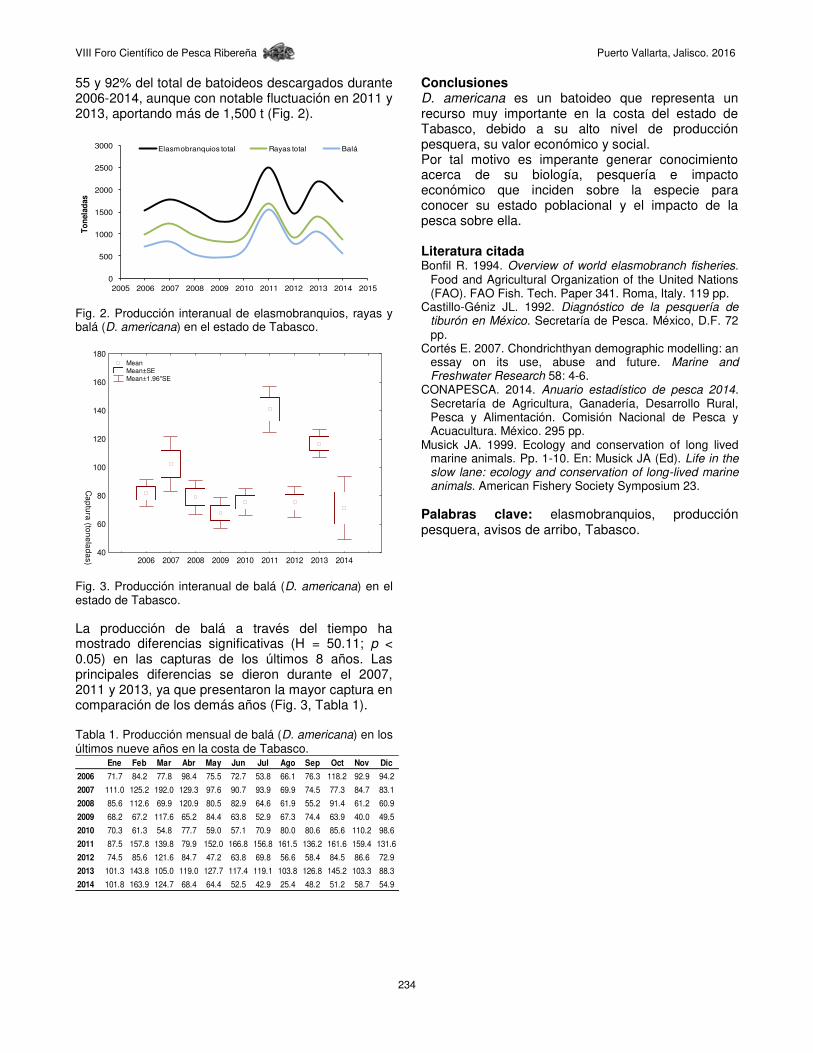
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
234
55 y 92% del total de batoideos descargados durante 2006-2014, aunque con notable fluctuación en 2011 y 2013, aportando más de 1,500 t (Fig. 2).
Fig. 2. Producción interanual de elasmobranquios, rayas y balá (D. americana) en el estado de Tabasco.
Fig. 3. Producción interanual de balá (D. americana) en el estado de Tabasco. La producción de balá a través del tiempo ha mostrado diferencias significativas (H = 50.11; p < 0.05) en las capturas de los últimos 8 años. Las principales diferencias se dieron durante el 2007, 2011 y 2013, ya que presentaron la mayor captura en comparación de los demás años (Fig. 3, Tabla 1). Tabla 1. Producción mensual de balá (D. americana) en los últimos nueve años en la costa de Tabasco.
Conclusiones D. americana es un batoideo que representa un recurso muy importante en la costa del estado de Tabasco, debido a su alto nivel de producción pesquera, su valor económico y social. Por tal motivo es imperante generar conocimiento acerca de su biología, pesquería e impacto económico que inciden sobre la especie para conocer su estado poblacional y el impacto de la pesca sobre ella. Literatura citada Bonfil R. 1994. Overview of world elasmobranch fisheries.
Food and Agricultural Organization of the United Nations (FAO). FAO Fish. Tech. Paper 341. Roma, Italy. 119 pp.
Castillo-Géniz JL. 1992. Diagnóstico de la pesquería de tiburón en México. Secretaría de Pesca. México, D.F. 72 pp.
Cortés E. 2007. Chondrichthyan demographic modelling: an essay on its use, abuse and future. Marine and Freshwater Research 58: 4-6.
CONAPESCA. 2014. Anuario estadístico de pesca 2014. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Comisión Nacional de Pesca y Acuacultura. México. 295 pp.
Musick JA. 1999. Ecology and conservation of long lived marine animals. Pp. 1-10. En: Musick JA (Ed). Life in the slow lane: ecology and conservation of long-lived marine animals. American Fishery Society Symposium 23.
Palabras clave: elasmobranquios, producción pesquera, avisos de arribo, Tabasco.
0
500
1000
1500
2000
2500
3000
2005 2006 2007 2008 2009 2010 2011 2012 2013 2014 2015
Tone
lada
s
Elasmobranquios total Rayas total Balá
2006 2007 2008 2009 2010 2011 2012 2013 201440
60
80
100
120
140
160
180
Ca
ptu
ra (to
ne
lad
as)
Mean Mean±SE Mean±1.96*SE
Ene Feb Mar Abr May Jun Jul Ago Sep Oct Nov Dic
2006 71.7 84.2 77.8 98.4 75.5 72.7 53.8 66.1 76.3 118.2 92.9 94.2
2007 111.0 125.2 192.0 129.3 97.6 90.7 93.9 69.9 74.5 77.3 84.7 83.1
2008 85.6 112.6 69.9 120.9 80.5 82.9 64.6 61.9 55.2 91.4 61.2 60.9
2009 68.2 67.2 117.6 65.2 84.4 63.8 52.9 67.3 74.4 63.9 40.0 49.5
2010 70.3 61.3 54.8 77.7 59.0 57.1 70.9 80.0 80.6 85.6 110.2 98.6
2011 87.5 157.8 139.8 79.9 152.0 166.8 156.8 161.5 136.2 161.6 159.4 131.6
2012 74.5 85.6 121.6 84.7 47.2 63.8 69.8 56.6 58.4 84.5 86.6 72.9
2013 101.3 143.8 105.0 119.0 127.7 117.4 119.1 103.8 126.8 145.2 103.3 88.3
2014 101.8 163.9 124.7 68.4 64.4 52.5 42.9 25.4 48.2 51.2 58.7 54.9
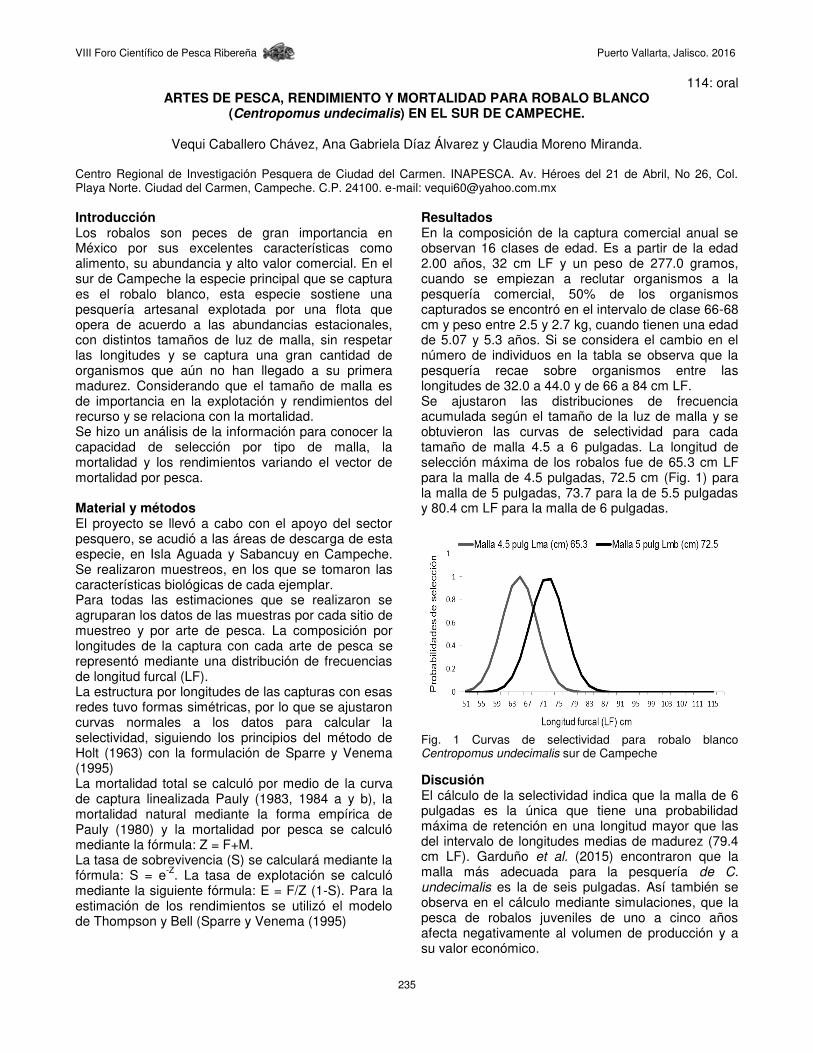
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
235
114: oral ARTES DE PESCA, RENDIMIENTO Y MORTALIDAD PARA ROBALO BLANCO
(Centropomus undecimalis) EN EL SUR DE CAMPECHE.
Vequi Caballero Chávez, Ana Gabriela Díaz Álvarez y Claudia Moreno Miranda.
Centro Regional de Investigación Pesquera de Ciudad del Carmen. INAPESCA. Av. Héroes del 21 de Abril, No 26, Col. Playa Norte. Ciudad del Carmen, Campeche. C.P. 24100. e-mail: [email protected] Introducción Los robalos son peces de gran importancia en México por sus excelentes características como alimento, su abundancia y alto valor comercial. En el sur de Campeche la especie principal que se captura es el robalo blanco, esta especie sostiene una pesquería artesanal explotada por una flota que opera de acuerdo a las abundancias estacionales, con distintos tamaños de luz de malla, sin respetar las longitudes y se captura una gran cantidad de organismos que aún no han llegado a su primera madurez. Considerando que el tamaño de malla es de importancia en la explotación y rendimientos del recurso y se relaciona con la mortalidad. Se hizo un análisis de la información para conocer la capacidad de selección por tipo de malla, la mortalidad y los rendimientos variando el vector de mortalidad por pesca. Material y métodos El proyecto se llevó a cabo con el apoyo del sector pesquero, se acudió a las áreas de descarga de esta especie, en Isla Aguada y Sabancuy en Campeche. Se realizaron muestreos, en los que se tomaron las características biológicas de cada ejemplar. Para todas las estimaciones que se realizaron se agruparan los datos de las muestras por cada sitio de muestreo y por arte de pesca. La composición por longitudes de la captura con cada arte de pesca se representó mediante una distribución de frecuencias de longitud furcal (LF). La estructura por longitudes de las capturas con esas redes tuvo formas simétricas, por lo que se ajustaron curvas normales a los datos para calcular la selectividad, siguiendo los principios del método de Holt (1963) con la formulación de Sparre y Venema (1995) La mortalidad total se calculó por medio de la curva de captura linealizada Pauly (1983, 1984 a y b), la mortalidad natural mediante la forma empírica de Pauly (1980) y la mortalidad por pesca se calculó mediante la fórmula: Z = F+M. La tasa de sobrevivencia (S) se calculará mediante la fórmula: S = e-Z. La tasa de explotación se calculó mediante la siguiente fórmula: E = F/Z (1-S). Para la estimación de los rendimientos se utilizó el modelo de Thompson y Bell (Sparre y Venema (1995)
Resultados En la composición de la captura comercial anual se observan 16 clases de edad. Es a partir de la edad 2.00 años, 32 cm LF y un peso de 277.0 gramos, cuando se empiezan a reclutar organismos a la pesquería comercial, 50% de los organismos capturados se encontró en el intervalo de clase 66-68 cm y peso entre 2.5 y 2.7 kg, cuando tienen una edad de 5.07 y 5.3 años. Si se considera el cambio en el número de individuos en la tabla se observa que la pesquería recae sobre organismos entre las longitudes de 32.0 a 44.0 y de 66 a 84 cm LF. Se ajustaron las distribuciones de frecuencia acumulada según el tamaño de la luz de malla y se obtuvieron las curvas de selectividad para cada tamaño de malla 4.5 a 6 pulgadas. La longitud de selección máxima de los robalos fue de 65.3 cm LF para la malla de 4.5 pulgadas, 72.5 cm (Fig. 1) para la malla de 5 pulgadas, 73.7 para la de 5.5 pulgadas y 80.4 cm LF para la malla de 6 pulgadas.
Fig. 1 Curvas de selectividad para robalo blanco Centropomus undecimalis sur de Campeche Discusión El cálculo de la selectividad indica que la malla de 6 pulgadas es la única que tiene una probabilidad máxima de retención en una longitud mayor que las del intervalo de longitudes medias de madurez (79.4 cm LF). Garduño et al. (2015) encontraron que la malla más adecuada para la pesquería de C. undecimalis es la de seis pulgadas. Así también se observa en el cálculo mediante simulaciones, que la pesca de robalos juveniles de uno a cinco años afecta negativamente al volumen de producción y a su valor económico.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
236
Todas las longitudes de máxima retención calculadas en el presente trabajo cumplen con este requisito, ya que corresponden a organismos de entre 6 y 8 años de edad, pero la máxima retención son las de las redes con mallas de seis pulgadas o mayor (80.4 cm LF). Esto la hace recomendable para esta pesquería. Conclusiones Dejar de capturar organismos juveniles, baja los índices de mortalidad, por lo tanto se debe utilizar un arte de pesca con la malla óptima para la captura de individuos adultos, malla de seis pulgadas. Sin que se incremente el esfuerzo, ya que en esta porción de la población se encuentra el mayor número de reproductores. Literatura citada Garduño-Dionate M, FA Aguilar-Salazar, C Aguilar
Cardozo, JD Martínez-Aguilar, R Figueroa-Paz, F Figueroa-Paz, PA Pérez-Velázquez. 2015. Selectividad de redes de enmalle en la pesquería de robalo blanco, Centropomus undecimalis en la Laguna de Yalahau, Quintana Roo. Instituto Nacional de Pesca. Ciencia Pesquera 23 (1): 37-45pp.
Holt SJ. 1963. A method for determining gear selectivity and its application. ICNAF Spec. public. 5: 106-115.
Pauly D. 1980. On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J. Cons. CIEM, 39 (2): 175-192.
Pauly D. 1983. Algunos métodos simples para la evaluación de recursos pesqueros tropicales. FAO Doc Téc. Pesca (234): 49 pp.
Pauly D. 1984a. Length-converted catch curves. A powerful tool for fisheries research in the tropics. (Part II). ICLARM Fishbyte 2 (1): 17-19
Pauly D. 1984b. Length-converted catch curves. A powerful tool for fisheries research in the tropics. (III: Conclusion). ICLARM Fishbyte 2 (3): 9-10
Sparre P, SC Venema 1995. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1- Manual. FAO Documento técnico de pesca 306/1 Rev.1. (Chile), 1995.
Palabras clave: Selección, explotación, pesca artesanal, sobrevivencia.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
237
115: oral DISTRIBUCION DE LA JAIBA Callinectes spp. EN LA LAGUNA DE TÉRMINOS,
CIUDAD DEL CARMEN, CAMPECHE
Mitzy Torres Blanco, Vequi Caballero y Ana Gabriela Díaz Álvarez
Centro Regional de Investigación Pesquera de Ciudad del Carmen. INAPESCA. Av. Héroes del 21 de Abril, No 26, Col. Playa Norte. Ciudad del Carmen, Campeche. C.P. 24100. Introducción La Laguna de Términos es uno de los tres sistemas estuarios de mayor extensión en el sureste del Golfo de México, la heterogeneidad ambiental de esta laguna determinada por su salinidad, tipo de vegetación, textura del sedimento, favorece el reclutamiento y establecimiento de diversos grupos faunísticos, entre los cuales los macro- crustáceos ocupan un lugar preponderante (Escobar 1984). Esta laguna sustenta la pesquería de jaiba del género Callinectes spp. que se captura y se comercializa sin diferenciación específica. Las jaibas del género Callinectes son especies macro plantónicas con distribución ontogénica diferencial: fase larva pelágica y fase bentónica en lagunas costeras y estuarios, donde crecen y se reproducen (Hendrickx 1984, Hernández y Arreola-Lizárraga 2007, Ramos-Cruz 2008). Son crustáceos que habitan la zona de ribera marina y en los sistemas lagunares costeros durante su fase adulta. Con el reflujo de la marea, las larvas zoeas de C. sapidus se alejan de las zonas estuarinas y permanecen de 30 a 45 días en la ribera adyacente hasta su metamorfosis al estadio megalopa, que tiene capacidad de migración vertical diel (por la noche están en la superficie y de día se sumergen). El estadio megalopa dura de seis a 20 días; es en esta etapa que las larvas son transportadas a cuerpos de agua estuarinos, donde se protegen entre la vegetación hasta convertirse en el primer estadio juvenil (Fischer y Wolff 2006). Esta pesquería es de gran importancia comercial ya que sustenta plantas procesadoras que permiten su procesamiento y exportación generando empleos adicionales y divisas, a pesar de ser como lo define la FAO (1985): “pesquerías de pequeña escala, de intensa labor y conducidos por obreros artesanales”. No sólo es importante conocer la biología, longitudes, y estadios de madurez, se requiere saber el stock susceptible de explotación; es por ello, que este trabajo pretende conocer la distribución de las zonas de pesca de Callinectes spp. de la Laguna de Términos de Cd. del Carmen, para contribuir a un manejo sustentable del recurso. Material y métodos Se realizó un análisis cualitativo que consistió en tres muestreos mensuales de mayo a diciembre 2015, basados en la captura de los sitios de desembarco y
en entrevistas a los pescadores al finalizar sus viajes de pesca. Las muestras se conformaron tomando de manera aleatoria ejemplares de jaiba de las capturas del día. Se contó el número total de jaibas de cada especie en la captura. Los datos del esfuerzo aplicado se obtuvieron mediante entrevista, preguntando al pescador el nombre del sitio de pesca, el número de aros o de trampas utilizadas y la hora aproximada de inicio y de finalización del viaje, profundidad y gastos de avituallamiento. Para la identificación del sexo se siguió el criterio descrito por Tabares (2002). Resultados y discusiones Se identificaron 15 zonas de captura en la Laguna de Términos (Fig. 1).
Fig. 1. Mapa con puntos de pesca de Callinectes spp. en la Laguna de Términos (mayo a diciembre 2015). Tabla 1. Organismos machos y hembras por especie de Callinectes en la Laguna de Términos (mayo-diciembre 2015)
Especie total machos hembras C. sapidus 4,653 2,896 1,857 C. rathbunae 165 137 47 C. similis 7 5 2 C. bocourti 3 3 3 C. danae 2 1 1
Se muestrearon 4,832 organismos (Tabla 1), se identificaron 5 especies: Callinectes sapidus, C. similis, C. rathbunae, C. bocourti, C. danae. La C. sapidus tuvo mayor presencia en 12 sitios de la laguna, C. similis y C. rathbunae en 4 sitios, C. danae y C. bocourti en sólo 1 sitio.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
238
Las longitudes mínimas y máximas que se presentaron se muestran en la Tabla 2. Tabla 2. Longitudes máximas, mínimas y promedio de Callinectes spp en la Laguna de Términos.
Especie Mín (mm)
Máx (mm)
Promedio (mm)
C. sapidus 86.69 167.90 138.45 C. rathbunae 80.67 128.00 102.40 C. similis 76.33 99.67 87.10 C. bocourti 73.30 90.00 92.40 C. danae 140.00 140.00 140.00
C. sapidus se captura en las zonas de la Laguna de menor salinidad lo que coincide con Cisneros-Mata et al. (2014); esta especie prefiere las zonas costeras poco profundas del complejo estuarino, con fondos fangosos y fango-arcilloso, en las zonas de mayor inundación. Presenta una longitud máxima mayor a las demás especies. Es abundante en la parte oriental de la laguna: boca de Pargos y esteros de Sabancuy, así como en el litoral con temperaturas de 28.5 a 30°C y salinidades de 15 a 33‰, y a 1.6 m de profundidad promedio. C. similis se encuentra en el sector oeste, norcentral y noreste, abundante en la parte occidental de la Laguna de Términos a temperaturas de 28 a 31°C y salinidades de 5 a 33‰, a una profundidad de 2 metros promedio. C. rathbunae se encontró en el sector oeste y sector sur de la laguna, abundante en el área de inundación de la península de Atasta, a una profundidad de 1.5 m. C. bocourti se encuentra en la zona oriental y en el sur de la laguna, abundante en las áreas de inundación de Sabancuy a 1.2 m de profundidad. La Laguna de Términos presenta mayor abundancia de C. sapidus, es el recurso más explotado en la zona, por lo es necesario continuar con el monitoreo de la pesquería para determinar el flujo de esta especie, series históricas anuales completas que sirvan para diagnosticar la pesquería y definir medidas de manejo actualizadas. Literatura citada Hendrickx ME. 1984. Estudio de la fauna marina y costera
del sur de Sinaloa, México. III. Clave de identificación de los cangrejos de la Familia Portunidae (Crustacea: Decapoda). Anales del Instituto de Ciencias del Mar y Limnología.
Hernández L, JA Arreola-Lizárraga. 2007. Estructura de tallas y crecimiento de los cangrejos Callinectes arcuatus y C. bellicosus (Decapoda: Portunidae) en la laguna costera Las Guásimas, México. Rev. Biol. Trop. 55(1): 225-233.
Fischer S, M Wolff. 2006. Fisheries assessment of Callinectes arcuatus (Brachyura, Portunidae) in the Gulf of Nicoya, Costa Rica. Fisheries Research 77: 301311
Ramos-Cruz S. 2008. Estructura y parámetros poblacionales de Callinectes arcuatus Ordway, 1863. (Decapoda: Portunidae), en el sistema lagunar La Joya-
Buenavista, Chiapas, México. Julio a diciembre de 2001. Pan-American Journal of Aquatic Sciences 3(3): 259-268.
Palabras clave: Jaiba, distribución, Laguna de Términos, pesquería, Campeche.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
239
116: oral RELACIÓN LONGITUD-PESO DE LA CHABELITA Peprilus medius
(PERCIFORMES, STROMATEIDAE)
Rebeca Sánchez Cárdenas1, María de los Ángeles Maldonado Amparo2, Jorge Saúl Ramírez Pérez2, Luis Antonio Salcido Guevara2 y Ramón Enrique Morán Angulo2.
1CONACyT-2Facultad de Ciencias del Mar. Universidad Autónoma de Sinaloa. Paseo Claussen s/n, Col. Los Pinos Mazatlán, Sinaloa, México. C.P. 82000. e-mail: [email protected] Introducción La chabelita Peprilus medius (Peters 1869) pertenece a la familia Stromateidae y se distribuye del Golfo de California al Norte de Chile (Palacios-Salgado et al. 2012). Hoy en día, P. medius es un recurso pesquero apreciado y su importancia comercial va creciendo, por su calidad, bajo precio y disponibilidad durante todo el año. La captura se destina para el consumo humano local, como producto de exportación, para elaborar alimento para peces (Inga et al. 2008, Martínez-Muñoz 2012, Moreira-Arcentales 2013), y como carnada. Cualquiera que sea el uso, la pesquería americana de P. medius beneficia al sistema socioeconómico de diversas comunidades pesqueras, por ejemplo, en Perú, Ecuador, Colombia y México. La captura de P. medius no cuenta con ninguna regulación y monitoreo en la mayoría de las costas donde se realiza. Una excepción es Perú, donde se ejercen medidas de manejo específicas: una longitud mínima legal de captura (23 cm de longitud total), monitoreo y un programa de investigación constante (El Peruano 2007). El monitoreo y el conocimiento biológico de especies como P. medius es fundamental para implementar y dar eficiencia a la administración pesquera. Pero además, esta especie podría ser utilizada como un bioindicador, dado que puede responder a cambios en el ambiente, por ejemplo, su ocurrencia fuera de su área de distribución está relacionada con eventos cálidos (Sielfeld et al. 2010). En ese contexto es necesario establecer puntos de referencia biológicos, que faciliten la interpretación e integración del conocimiento a las decisiones de manejo pesquero. En ese sentido, el presente trabajo presenta una revisión de la relación longitud total-peso total de P. medius y estimaciones inéditas. Material y métodos La revisión de la relación longitud total-peso total (RLP) de P. medius se realizó a través de documentos publicados (tesis, artículos, e informes). Además se realizaron estimaciones a partir de datos de longitud total (Lt, cm) y peso total (Pt, g) obtenidos de la captura comercial del sureste del Golfo de California, durante los periodos diciembre 2011 a octubre 2014 (P1), y de septiembre 2014 a noviembre 2015 (P2). La RLP se estimó utilizando la
función potencial: �� = ����, y los parámetros a y b fueron obtenidos a través de la función linealizada (Ricker 1975, Froese 2006). El parámetro b fue comparado con b = 3 a través de una prueba t Student (Sokal y Rolf 1979), para definir el tipo de crecimiento (A+, alométrico positivo; IS, isométrico; A-, alométrico negativo). La tendencia latitudinal de b fue explorada mediante el coeficiente de correlación por rangos de Spearman (R, Spearman 1904) y la similitud de R con R = 0 se evaluó mediante una prueba t Student (Sokal y Rolf 1979). Resultados y discusiones La revisión mostró que la RLP se ha estimado en stocks de P. medius que habitan las costas de Colombia (a = 0.1024, b = 2962, r2 = 0.8228; Zapata-Padilla 2011), El Salvador (a = 0.0315, b = 2.6771, r2
= 0.9501; Anónimo 2009), Baja California Sur, México (a = 0.025, b = 3.073, r2 = 0.937, A+; Rodríguez-Romero et al. 2009) y en el sureste del Golfo de California (a = 0.008, b = 3.089, r2 = 0.882, A+; Nieto-Navarro et al. 2010). Vera et al. (2006) reportaron que P. medius presentan un crecimiento alométrico negativo en la costa de Tumbes, Perú, pero no presentaron los parámetros de la RLP. Las RLP ajustadas para el sureste del Golfo de California, presentaron los siguientes parámetros: a = 0.0515, b = 2.5420, A-, r2=0.9524 (P1); y a = 0.0302, b = 2.7239, A-, r2 = 0.9501 (P2). Los valores de b mostraron una tendencia positiva en función de la latitud, aunque sin significancia estadística (r = 0.4928; t6-2 = 1.1327; P = 0.3206; Fig. 1). La evaluación de la tendencia requiere la incorporación de más información, y de confirmarse se establecería que el tipo de crecimiento de P. medius tiende a la alometría negativa en latitudes tropicales y a la alometría positiva en latitudes subtropicales. Además este comportamiento sería coherente con la regla de Bergman, dado que los individuos tenderían a ser más robustos en la región subtropical que en la tropical (Meiri 2011). Las variantes latitudinales de la RLP de P. medius se recomienda sean consideradas para propósitos de manejo pesquero, a manera de indicadores de salud de los stocks.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
240
Fig.1. Tendencia latitudinal de coeficiente b de la RLP de Peprilus medius. Conclusión P. medius presenta crecimiento alométrico positivo y alométrico negativo a lo largo de su distribución geográfica. Las variantes latitudinales de la RLP son coherentes con la regla de Bergman. Literatura citada Anónimo. 2009. Informe de la campaña de investigación
pesquera El Salvador 2009: Del 2 al 14 de diciembre, B/O Miguel Oliver. Instituto Español de Oceanografía, Gobierno de España - Centro de Desarrollo de la Pesca y la Acuicultura, El Salvador. 99 p.
El Peruano. 2007. Resolución ministerial No. 371-2007-PRODUCE. Sistema Peruano de Información Jurídica. Publicado en Lima, Perú, el 5 de diciembre de 2007.
Froese R. 2006. Cube law, condition factor and weight-length relationships: history, meta analysis and recommendations. J. Appl. Ichthyol. 22: 241-253.
Inga-Barreto C, J Rujel-Mena, E Gómez-Sulca, E Ordinola-Zapata. 2008. El chiri, Peprilus medius (Peters) en Tumbes, Perú. Parámetros biológico pesqueros y talla mínima de captura. Inf. Inst. Mar Perú 35 (3): 209–214.
Martínez-Muñoz MA. 2012. Estructura y distribución de la comunidad íctica acompañante en la pesca del camarón (Golfo de Tehuantepec, Pacífico Oriental, México). Tesis doctoral. Universidad de Barcelona. España. 604 p.
Meiri S. 2011. Bergmann´s rule – what´s in a name?. Global Ecol. Biogeogr. 20: 203–207.
Moreira-Arcentales EL. 2013. Evaluación del estado actual de las concesiones de áreas de manglar entregadas a comunidades ancestrales en la provincia del Guayas. Tesis de Maestría. Universidad de Guayaquil. Ecuador. 94 p.
Nieto-Navarro JT, M Zetina-Rejón, F Arreguín-Sánchez, NE Arcos-Huitrón, E Peña-Messina. 2010. Length-weight relationship of demersal fish from the Eastern coast of the mouth of the Gulf of California. J. Fish. Aquat. Sci. 5(6): 494-502.
Palacios-Salgado DS, XG Moreno-Sánchez, LA Abitia-Cárdenas, FJ Gutiérrez-Sánchez, J Rodríguez-Romero. 2012. Ichthyodiversity of San Jose, San Francisco, and El Pardito Islands in the southwestern Gulf of California, México. Acta Ichthyol. Piscat, 42(3): 177–191.
Ricker WE. 1975. Computation and interpretation of biological statistics of fish populations. B. Fish. Res. Board Can. 191: 300-382.
Rodríguez-Romero J, DS Palacios-Salgado, J López-Martínez, S Hernández-Vázquez, JI Velázquez-Abunader. 2009. The length-weight relationship
parameters of demersal fish species off the western coast of Baja California Sur, Mexico. J. Appl. Ichthyol. 25: 14-116.
Sielfeld W, J Laudien, M Vargas, M Villegas. 2010. El Niño induced changes of the coastal fish fauna off northern Chile and implications for ichthyogeography. Rev. Biol. Mar. Oceanogr. 45: 705-722.
Sokal R, J Rohlf. 1979: Biometry: The principles and practice of statistics in biological research. Blume Editions, San Francisco.
Spearman C. 1904. The proof and measurement of association between two things. Am. J. Psychol. 15(1): 72-101.
Vera M, C Inga, E Ordinola, S Alemán, J Llanos. 2006. Parámetros poblacionales de “chiri” Peprilus medius (Perciformes: Stromatidae) en la Región de Tumbes, durante el 2006.
Zapata-Padilla LA. 2011. Aspectos ecológicos y pesqueros de la ictiofauna demersal asociada a la plataforma continental del Pacífico colombiano. Tesis de Maestría. Universidad del Valle. Colombia. 193 p.
Palabras clave: alometría negativa, alometría positiva, isometría, tipo de crecimiento, tendencia latitudinal
Latitud norte
0 5 10 15 20 25 30
Pen
dien
te (
b)
0.0
2.0
2.5
3.0
3.5
Latitud versus b TendenciaRs = 0.4928

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
241
117: oral LONGITUD DE MADUREZ DE CURVINA GOLFINA Cynoscion othonopterus
EN EL ALTO GOLFO DE CALIFORNIA
Jaime Edzael Mendivil Mendoza y Eugenio Alberto Aragón Noriega
Centro de Investigaciones Biológicas del Noroeste, Unidad Sonora. Km 2.35 camino al Tular, Estero Bacochibampo, Guaymas, Sonora, México. C.P. 85454. [email protected]; [email protected] Introducción La curvina golfina Cynoscion othonopterus (Gilbert y Jordan 1882) es un pez diádromo endémico del Alto Golfo de California (AGC), que pertenece a la familia Sciaenidae comúnmente llamados curvinas o roncadores. Este organismo es un pez migratorio y anualmente de febrero a mayo se dirige hacia la Reserva de la Biosfera del Alto Golfo de California y Delta del Río Colorado (RBAGCyDRC) con fines reproductivos (Román-Rodríguez 2000, Acosta-Valenzuela 2010). C. othonopterus es una especie de alto valor pesquero por el volumen que representa y debido a que se pesca durante la cuaresma. Los registros oficiales de pesca indican que en 2002 se registró la máxima producción histórica con 5,952 t. Desde ese año, las capturas han fluctuado entre 1,500 y 3,700 t. Con el fin de aprovechar el recurso sin afectar su capacidad de renovación, en 2005 entra en vigor la NOM-063-PESC-2005 donde se establece como longitud mínima de captura 65 cm de longitud total, red agallera 5¾ pulgadas de luz de malla y cuotas de capturas para cada temporada por mencionar algunos términos y condiciones. Actualmente, el recurso curvina representa la pesquería más importante en el AGC, ya que es la única especie con permiso de explotación pesquera dentro AGCDRC, por decreto del Gobierno de México, publicado en el DOF (10/04/2015). Por esta razón el objetivo del presente estudio fue identificar los organismos maduros mediante el criterio macroscópico para establecer la longitud de madurez de Cynoscion othonopterus durante cuatro temporadas reproductivas (2008, 2013, 2014 y 2015). Material y métodos Los datos analizados en el presente estudio, provienen de muestreos de campo que se realizaron durante los meses de febrero a abril de 2008, 2013, 2014 y 2015 en el AGC. Los muestreos consistieron en la captura de curvina golfina, a las cuales se les registraron los datos biométricos (peso total, longitud total). El criterio utilizado para designar un organismo maduro e inmaduro, se realizó mediante el análisis macroscópico adoptado y utilizado por Nikolsky (1963) para C. othonopterus. El cálculo de la longitud de madurez (L50), se realizó obteniendo el porcentaje de individuos maduros por clase de longitudes cada 50 mm, y posteriormente,
ajustando estos valores a un modelo logístico a través de la siguiente ecuación: Pi = 11+exp− Li−L /a´
Donde: Pi = porcentaje de madurez a la longitud i, Li = intervalo de longitud; L50 = longitud de 50% de madurez poblacional: a = constante de ajuste. Mediante iteraciones, los parámetros del modelo fueron ajustados maximizando el algoritmo de log normal de máxima verosimilitud de la función binomial (Brouwer y Griffiths 2005):
Donde: n = número de total de organismos de la clase i, m = número de organismos maduros en la clase i. Se obtuvieron los intervalos de confianza de la longitud de madurez a 95% con base en los perfiles de verosimilitud y la distribución chi-cuadrada (Venzon y Moolgavkar 1988). El intervalo de confianza fue definido como todos los valores de θ que satisfacen la siguiente desigualdad.
1,1
2))((((2 xYLYl best Donde: )( bestYL = log normal de máxima
verosimilitud del valor más probable de θ y 1,1
2x = valor de la distribución Chi-cuadrada cuando d = 1(3.84). Resultados y discusión Se analizaron 920 organismos, los cuales 526 se encontraban maduros. La longitud mínima registrada de madurez fue 350 mm y la máxima de 950 mm. La distribución de longitudes osciló entre 150 y 950 mm, estando más representado por organismos de 600 a 850 mm. Estos resultados concuerdan con los reportados por Román-Rodríguez (2000) y Acosta-Valenzuela (2010). La longitud de madurez al 50% (L50) para los años 2008, 2013, 2014 y 2015 fue de 502 mm, 559 mm, 740 mm y 460 mm, respectivamente (Fig. 1). Estos resultados de L50 muestran una disminución de la longitud de madurez con respecto a las investigaciones previas realizadas por Román-
,ln)1ln(*1
ln*1
i
i
nmPn
P
PmL ii
i
i
i
n
i

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
242
Rodríguez (2000) y Acosta-Valenzuela (2010). Los perfiles de verosimilitud para la longitud de madurez se muestran en la figura 2. Se observa que el intervalo de confianza para 2008 fue de 490 a 514 mm, para 2013 de 540 a 576 mm, 2014 de 728 a 752 mm y 2015 de 430 490 mm.
Fig. 1. Curvas de la longitud de madurez de curvina golfina Cynoscion othonopterus en el Alto Golfo de California. Los círculos son datos observados, la línea es la curva ajustada.
Fig. 2. Perfiles de verosimilitud para la longitud de madurez de Cynoscion othonopterus estimada con la curva logística. Conclusiones La disminución de la L50 se debe a la alta presión por pesca a la cual se encuentra sometida C. othonopterus, por lo que el organismo responde de manera estratégica para madurar a bajas longitudes con el fin de preservar la especie.
Literatura citada Acosta-Valenzuela Y. 2010. Ecología y potencial
reproductivo de la curvina golfina Cynoscion othonopterus (Jordan y Gilber, 1882) en el Alto Golfo de California y Delta del Río Colorado. Tesis de Maestría, Instituto Tecnológico de Guaymas, Sonora. 122 p.
Brouwer SL, MH Griffiths. 2005. Reproductive biology of carpenter sea-bream (Argyrozona argyrozona) (Pisces: Sparidae) in a marine protected area. Fisheries Bulletin 103(2): 258-269.
Nikolsky GV. 1963. The ecology of fish. Academic Press, New York. 234p.
Román-Rodríguez MJ. 2000. Estudio poblacional del chano norteño, Micropogonias megalops y la curvina Golfina Cynoscion othonopterus (Gilbert) (Pisces: Sciaenidae), especies endémicas del Alto Golfo de California, México. Instituto del Medio Ambiente y Desarrollo Sustentable del Estado de Sonora. Informe final SNIB-CONABIO. Proyecto No. L298. México D. F.
Venzon DJ, SH Moolgavkar. 1988. A method for computing profile-likelihood-based confidence intervals. Applied Statistics 37(1): 87-94.
Palabras clave: Cynoscion othonopterus, longitud de madurez, máxima verosimilitud, Alto Golfo de California, México.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
243
118: oral ACADJA, ALTERNATIVA PARA EL AUMENTO DEL POTENCIAL PESQUERO EN
BARRA DEL TORDO, TAMAULIPAS
Leobardo García Solorio1, Víctor Martin Zárate Noble1, María de la Luz Juárez Villalobos2
1Centro Regional de Investigación Pesquera de Tampico, INAPESCA. 2Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. [email protected] Introducción La pesca en parques de ramas (acadja) es una forma tradicional de acuicultura de poca tecnología que se practica en aguas continentales y salobres de muchas partes del mundo. Las acadjas consisten en un núcleo interior o círculo concéntrico de ramas de árboles u otros materiales puestos muy apretados y rodeados de una estructura exterior de madera más robusta, que se pesca periódicamente, generalmente mediante el cerco. Se prefieren montarlas en aguas someras tranquilas de no más de 1.5 m de profundidad. Éstas sirven de refugio, pero su función más importante es como una fuente abundante de alimentos. Esta práctica acuícola se tomó como prototipo en la región del Tordo en Tamaulipas, considerando al ostión como un recurso base dentro de un ecosistema complejo y dado que la densidad de los bancos de la zona sur de Tamaulipas tienen tendencia a la baja. Se analiza la posibilidad de tomar como una alternativa de recuperación ecosistemático a la “acadja”.
Fig. 1. Ubicación de acadja en rio Carrizales, Barra de Tordo, Tamaulipas. Material y métodos En la zona de Barra de Tordo municipio de Aldama Tamaulipas, se trabajó en conjunto con la cooperativa “Pescadores Unidos de Barra del Tordo” y se optó por instalar una acadja en la zona del rio donde es un área de rápido desarrollo para la parte de ostricultura, así como ser un punto de congruencia del río con el mar (Fig. 1). El diseño de la acadja para las zonas de estudio consistió en tres triángulos conectados en el centro, dichos encierros fueron formados por red tipo camaronera de 5 mm de abertura de malla, encabalgada a cabo, fijada al fondo con estacas (para evitar el uso de ramas).
En los encierros triangulares, para modo de comparación se colocaron arpillas de ostión vivo en una, en otro arpilla de concha de ostión y en el tercer encierro no se colocó nada (Fig. 2).
Fig. 2. Diseño modular de sistema tipo acadja, para la recuperación y creación de hábitat por agregación: a) fondo con arpillas de ostión vivo, b) fondo con arpilla de concha de ostión, c) testigo sin elementos de fondo (diseño del arte: Zárate 1999). Se evaluó la capacidad de agregación de biomasa por medio de la diversidad y abundancia de la infauna como indicador de impacto, el muestreo fue efectuado con un nucleador de PVC que abarca un área de muestreo de 0.00385 m2 con una profundidad de penetración de 10 cm. Se tomaron 5 nucleadores para representar una muestra de 0.01925 m2 de cada sección de la acadja (vacía, con concha de ostión y con ostión vivo) y del área exterior adyacente para calcular posteriormente los valores a 1 m2. Las muestras fueron pasadas por un tamiz con luz de malla de 2 mm, la muestra fue preservada con formalina al 5% en agua de mar. Las muestras fueron revisadas con lupa de 4x para búsqueda de macroinvertebrados, identificándolos a nivel de filo. Se tomaron dos muestras cada dos meses en transectos radiales a los 5, 10, 20 y 40 metros a partir del centro la acadja. La comparación entre los valores de diversidad y abundancia entre los tipos de acadjas y los sitios de colocación se compararon estadísticamente para verificar la existencia de diferencias significativas por estos efectos, tomando indicadores ecológicos de asociación. Resultados Se evaluó mensualmente el comportamiento que se presenta en cuanto a la atracción de especies en la

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
244
zona externa de la acadja, esto realizando corraleos tanto en la acadja como lejos de él, con esto se observó que la acadja en la zona es punto de atracción, aunque la presencia más clara de peces principalmente se dio hasta el cuarto mes y las principales especies capturadas en los alrededores fueron lisas, pargo, mojarritas. Análisis de agregación de especies (Infauna). Aun existiendo condiciones adversas para el asentamiento de infauna debido a la velocidad de la corriente, se logró observar el incremento significativo de organismos dentro de las secciones de las acadjas aun utilizando solamente la abundancia relativa como un indicador y tan sólo a nivel de filo. De 4,107 organismos, se pudieron observar en la sección de ostiones vivos, para el caso de moluscos, al menos cuatro especies predominantes, dos almejas y dos caracoles. En el caso de los artrópodos se trató al parecer de una sola especie de crustáceo y en los anélidos parecerían 4 especies (Fig. 3). Sin embargo, no se tuvo tiempo de identificarlos para hablar sobre diversidad. Para el efecto de este experimento sólo se quería evidenciar la importancia del ostión como formador de hábitat.
Fig. 3. Representación gráfica del índice de abundancia por filo en las diferentes zonas de la acadja. En un análisis de comparación entre zonas, se observa en el dendrograma de abundancia relativa de especies por zona, que existe una relación estrecha entre la zona externa y la región que no tiene ostión dentro de la acadja, así como una relación entre la zona de ostión vivo y concha de ostión (Fig. 4 y 5).
Fig. 4. Representación gráfica del análisis de conglomerados entre las áreas de la acadja.
Conclusiones La acadja como sistema de agregación de biomasa, es funcional con diferentes efectos por la presencia o ausencia de ostión vivo o concha de ostión se observa en las imágenes. En la zona con ostión vivo se observan los peces como especies principales de agregación, en la zona de conchas de ostión creció una especie de pasto marino, además de agregación de peces y crustáceos. En la zona testigo se observan sólo peces en mucha menor cantidad que las otras dos zonas. Sin embargo, la simple presencia de especies de escama dentro de la zona testigo demuestra la capacidad de agregación que representa una acadja. Es importante reconocer la importancia que juegan los sistemas de agregación como una herramienta de desarrollo ecológico y del mismo modo pesquero acuícola. Sin embargo, estos sistemas de recuperación no pueden evaluarse en corto tiempo, ya que los resultados son de mediano a largo plazo por lo que se requiere tener continuidad en los estudios.
Fig. 5. Diferencias entre las áreas de la acadja, Barra del Tordo, Tamaulipas. Literatura citada Zarate NVM. 1999. The use infaunal analysis over
Seagrass Beds as possible indicator for Coastal Rehabilitation Plans. Thesis of Master in science. University of Wales, Bangor.
Palabras clave: Acadja, Agregación, ostión, potencial pesquero.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
245
119: oral AGREGACIÓN ESTACIONAL DE HEMBRAS GRÁVIDAS DE TIBURÓN AZUL (Prionace glauca) EN EL
OCÉANO PACÍFICO MEXICANO E IMPLICACIONES PARA EL MANEJO SUSTENTABLE
Luis Vicente González Ania1 y José Leonardo Castillo Géniz2
1Oficinas Centrales. INAPESCA. Pitágoras 1320, Col. Santa Cruz Atoyac, Ciudad de México. C.P. 03310. [email protected]; 2Centro Regional de Investigación Pesquera de Ensenada. INAPESCA. Carr. Tijuana-Ensenada km 97.5, El Sauzal de Rodríguez, Ensenada, Baja California, México. C. P. 22760. [email protected] Introducción La evaluación poblacional más reciente del tiburón azul Prionace glauca (Linnaeus 1758) en el Océano Pacífico norte (Takahashi et al. 2014), mostró que la condición actual del stock es saludable, con una biomasa superior a la estimada para el rendimiento máximo sostenible. En el Pacífico mexicano las capturas de tiburón azul han ido en aumento, desde menos de 500 t promedio en los 1970, hasta alrededor de 4,000 t en la segunda mitad de los 2000. Estas capturas se realizan por flotas artesanales y de mediana altura, estando integradas en gran medida por ejemplares juveniles (Sosa-Nishizaki 2013). En cuanto al ciclo de vida del tiburón azul, Vögler et al. (2012) encontraron evidencia de que el Pacífico tropical de México es clave en el proceso reproductivo de esta especie, al registrar agregaciones de juveniles y hembras gestantes cercanas a las costas de la Península de Baja California y Colima, pero sin cuantificar las distancias. Estas observaciones confirmaron resultados obtenidos por Carrera-Fernández et al. (2010) en Baja California Sur, siendo consistentes en que las mayores capturas de hembras grávidas se presentan en otoño. Para el manejo del recurso tiburón del Pacífico, la veda establecida de mayo a julio trata de proteger etapas reproductivas críticas de las principales especies pesqueras, procurando un compromiso entre sus distintas dinámicas reproductivas, las cuales coinciden parcialmente en el periodo que cubre la veda. Por lo anterior, el objetivo del trabajo es contribuir a identificar y confirmar estadísticamente la presencia estacional de estructuras de agregación de hembras grávidas de tiburón azul a distancias definidas de la costa y discutir su consistencia con la veda vigente. Material y métodos Se analizaron los datos validados de 1,914 lances de palangre utilizados originalmente para dictaminar la veda de tiburones del Pacífico, realizados en la Zona Económica Exclusiva del noroeste y centro del Pacífico mexicano, en 2006, 2007, 2008 y 2010, principalmente por las flotas de mediana altura con base en Ensenada, B.C. y Mazatlán, Sin. Los datos
provienen del Programa de Observadores de Tiburón (INAPESCA-FIDEMAR). Se utilizó un modelo aditivo generalizado (GAM por sus siglas en inglés), para analizar las capturas por lance de hembras gestantes de tiburón azul, en función del trimestre (estación) y la distancia de la costa (Wood 2006). La distancia (millas náuticas) de los lances al punto más cercano de la costa continental, se calculó con el método de Sparre y Venema (1997). Resultados y discusión En el trimestre de invierno no se encontró relación (p = 0.15) entre la captura por lance de hembras grávidas (hg) de tiburón azul y la distancia de los lances a la costa (Fig. 1).
Fig. 1. Relación entre distancia a la costa y captura de hembras grávidas de tiburón azul por lance (modelo GAM). El área sombreada representa el intervalo de 95% de confianza. Nótense las distintas escalas verticales. En cambio, en primavera se observó una relación altamente significativa (p = 9.94 ∙ e-10), con agregación marcada de hg a distancias de 100 a 300 mn de la costa. Este periodo (abril-junio), actualmente está cubierto en su mayoría (mayo y

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
246
junio) por la veda. Sin embargo, la captura por lance máxima fue relativamente baja (<1 hg/lance). En verano hubo indicios de agregación de 100 a 200 mn de la costa, con captura cercana a 1 hg/lance, pero la incertidumbre de la estimación es alta y la relación es sólo marginalmente significativa (p = 0.04). En este periodo (julio-septiembre), la veda solamente está vigente en julio. El otoño por otra parte fue la estación más notable, tanto por mostrar una relación estadísticamente muy significativa (p = 3.49 ∙ e-06), como por la agregación observada entre 200 y 300 mn de la costa, con capturas medias cercanas a 5 hg/lance, es decir, al menos 5 veces superiores a las máximas observadas durante los meses que actualmente protege la veda. A más de 300 mn de la costa la incertidumbre aumenta progresivamente, debido a que, al igual que en verano, se realizaron menos lances a esas distancias. La principal zona geográfica de agregación de hembras grávidas en otoño (octubre-diciembre), es claramente visible al suroeste de Baja California Sur (Fig. 2).
Fig. 2. Distribución espacio-temporal de lances de palangre con captura de hembras grávidas de tiburón azul. Leyenda: los números representan meses y los colores trimestres. Estos resultados son totalmente consistentes con los obtenidos por Vögler et al. (2012) y Carrera-Fernández et al. (2010), en cuanto a las mayores capturas de hembras gestantes en otoño. Asimismo, confirman las agregaciones identificadas por estos autores, en zonas cercanas a la Península de Baja California, especialmente en Baja California Sur. Los resultados además son explícitos acerca de las distancias mínimas de la costa a las que se encuentran agregaciones comprobables y las correspondientes abundancias relativas de hembras grávidas de tiburón azul. Conclusiones En otoño se encontró una agregación de alta abundancia, con un máximo de 5 hembras grávidas de tiburón azul por lance, a 200-300 mn de la costa occidental de Baja California Sur. Si bien la población de este recurso está en buenas condiciones, el aumento registrado en las capturas
hace pertinente conocer a tiempo este tipo de eventos de especial vulnerabilidad a la pesca, fuera del periodo de veda. Lo mismo resulta de interés en el caso de otras especies. Literatura citada Carrera-Fernández M, F Galván-Magaña, BP Ceballos-
Vázquez. 2010. Reproductive biology of the blue shark Prionace glauca (Chondrichthyes: Carcharhinidae) off Baja California Sur, México. Aqua, International Journal of Ichthyology 16: 101–110.
Sparre P, SC Venema. 1997. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1. Manual. FAO Documento Técnico de Pesca No. 306. 1 Rev. 2: 420 p.
Takahashi N, M Kanaiwa, S Ohshimo, T Sippel, K Yokawa. 2014. Stock assessment and future projections of blue shark in the North Pacific Ocean by Bayesian Surplus Production Model using Revised Data. ISC/14/SHARKWG-2/01. Working document submitted to the ISC Shark Working Group Workshop, 03‐10 June 2014, National Taiwan Ocean University, Keelung, Taiwan.
Sosa-Nishizaki O. 2013. Unofficial blue shark catch estimations for the Mexican Pacific (1976-2011). ISC/13/SHARKWG-2/INFO-1. Working document submitted to the ISC Shark Working Group Workshop, 16-24 April 2013, Shizuoka, Japan.
Vögler R, E Beier, S Ortega-García, H Santana-Hernández, JJ Valdez-Flores. 2012. Ecological patterns, distribution and population structure of Prionace glauca (Chondrichthyes: Carcharhinidae) in the tropical-subtropical transition zone of the north-eastern Pacific. Mar. Environ. Res. 73: 37–52.
Wood SN. 2006. Generalized additive models: An introduction with R. Chapman and Hall/CRC Press.: 384 p.
Palabras clave: Veda de tiburones, palangre pelágico, modelos GAM, Baja California, pesca de mediana altura.
120° 115° 110° 105° 100° 95° 90°
15°
20°
25°
30° 1 2 3 4 5 6 7 8 9 10 11 12

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
247
120: oral COMPOSICIÓN DE LA CAPTURA Y SELECTIVIDAD DEL PALANGRE EXPERIMENTAL PARA LA
CAPTURA DE PELÁGICOS MAYORES EN LA COSTA DE OAXACA
Aldrin Labastida Che, Ada Lisbeth Núñez Orozco, Eduardo Ramos Santiago y José Alfonso Oviedo Piamonte
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prolongación Playa Abierta s/n. Col. Miramar, Salina Cruz, Oax. C. P. 70680. Tels: 01 (971) 7145003 y 7140386. [email protected], [email protected], [email protected], [email protected] Introducción En México, la NOM-017-PESC-1994 regula la pesca deportiva y reserva el aprovechamiento de especies a embarcaciones deportivas que operen dentro de una franja costera de 50 millas. Esta restricción no se aplica en otros países y ha propiciado iniciativas para modificar la norma, que no evita la captura ilegal con fines comerciales de estas especies en los distintos puertos pesqueros de México. En el Pacífico sur de México, el dorado Coryphaena hippurus Linnaeus 1758 es la especie objetivo, tanto por la pesca deportiva como por la comercial (pesca artesanal). Los principales artes de pesca son: palangres (o cimbras) y redes de deriva. El palangre se comenzó a utilizar en México a principios de los cincuentas con el objeto de capturar atunes. En la actualidad, se ha adaptado para la pesca de tiburones con variantes y con incidencia de especies pelágicas migratorias como pez vela, marlin, pez espada, atún y dorado, en función de la temporada de abundancia, hábitos alimenticios y distribución geográfica (Macías-Zamora et al. 1999). Material y métodos Se realizaron muestreos de pesca experimental durante 2014 en la costa de Oaxaca, con la operación de un palangre para la captura de pelágicos mayores, similar a los que ocupan los pescadores: línea madre de 6,000 m, en 3 bloques; cada uno de 50 anzuelos circulares, de tres tamaños diferentes: C13/0, C14/0 y C15/0, respectivamente, con un orinque de 4 m de profundidad, distancia entre reinales de 40 m, y boyas a cada 3 reinales. Los datos obtenidos fueron: tiempo de operación, composición de la captura, tamaño de anzuelo y longitudes de los organismos. Se hicieron comparaciones básicas en donde los anzuelos circulares se combinaron de manera intercalada, con el fin de que el experimento se considerara completamente aleatorio, de acuerdo con el diagrama de la figura 1. Se usaron dos tipos de carnada: barrilete (Euthynnus lineatus) y ojotón (Selar crumenophtalmus). Composición de la captura: Se determinó a través de los datos de captura por cada tipo de anzuelo contra el número total de anzuelos calados, se estandarizó el esfuerzo al número promedio de anzuelos
utilizados por la flota de menor escala (150 anzuelos). Selectividad: La expresión matemática más simple utilizada para describir la curva de selección es la llamada “curva logística”. Se establece que existe una estrecha correspondencia entre S1 y S2 y L25%, L50% y L75%, longitudes a las cuales, respectivamente, el 25%, 50% y 75% de los peces son retenidos por el arte de pesca. El intervalo de longitudes entre L25% y L75%, que es simétrico en torno a L50%, se llama "rango de selección". El análisis de regresión se realiza en un intervalo de logitudes comprendido entre una retención equivalente a cero (0) y otra de retención total (1), no considerando los intervalos de longitudes inferiores, donde no se registran peces retenidos, o superiores a la primera longitud a la cual se obtiene la retención total. En otras palabras, en el análisis se utilizan solamente el intervalo de longitudes comprendidas entre valores de retención 0 y 1 (Sparre y Venema, 1995). El procedimiento fue aplicado para la especie capturada con mayor frecuencia y para cada tamaño de anzuelo utilizado.
Fig. 1. Diagrama del palangre experimental para comparar tres tamaños de anzuelos tipo circular en la pesca artesanal de la costa de Oaxaca. Resultados Efectividad de los anzuelos C13/0, C14/0 y C15/0. Durante el experimento se realizaron 23 lances, de 138 horas, 3,450 anzuelos calados y el número de organismos capturados por tamaño de anzuelo fue: C13/0 = 150, C14/0 = 152 y C15/0 = 185. La captura fue de Coryphaena hippurus (dorado), Istiophorus platypterus (pez vela), Carcharhinus porosus (cazón), Carcharhinus falciformis (tiburón sedoso), Alopias pelagicus (tiburón zorro), Thunnus

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
248
albacares (atún aleta amarilla) y Tetrapturus audax (marlin rayado). El mayor porcentaje de la captura en los tres tamaños de anzuelo fue de dorado, con C13/0 fue de 91%, 88% en el C14/0 y 90% con C15/0 (Fig.2).
Fig. 2. Composición de especies capturadas con los tres tamaños de anzuelos circulares 13/0, 14/0 y 15/0. Selectividad de los tres tamaños de anzuelos C13/0, C14/0 y C15/0 para dorado. El análisis fue sobre la especie que presentó mayor frecuencia en las capturas: el dorado C. hippurus. El intervalo de longitudes del dorado osciló de 53 y 126 cm de longitud furcal (Lf), en las capturas del anzuelo C13/0, de 50 a 123 cm Lf con C14/0 y 51 a 145 cm con el anzuelo C15/0. En la figura 3 se muestran las curvas logísticas de selectividad, para los tres tamaños de anzuelos. La selección al 50% para el dorado con el anzuelo C13/0 es de 84.05 cm Lf, con el anzuelo C14/0 es 89.29 cm Lf y para el anzuelo C15/0 es de 93.06 cm Lf.
Fig. 3. Curvas de selectividad de los tres tamaños de anzuelos C 13/0, C 14/0 y C 15/0 en la captura de Coryphaena hippurus. Discusiones Los porcentajes de captura de dorado del presente trabajo coinciden con los de Rodríguez-Valencia et al. (2008), donde evaluaron anzuelos circulares para reducir la captura incidental en las operaciones pesqueras de los palangreros ribereños de Sinaloa; mediante 53 lances obtuvieron 936 organismos; de ellos 94.6% fueron dorado. En el presente estudio se realizaron 23 lances y se obtuvieron 487 organismos y 91% fueron dorado.
Pero difiere con los resultados del experimento para determinar la eficiencia y selectividad del palangre de deriva con tres tipos de anzuelos y dos tipos de carnada realizado por Santana-Hernández et al. (2008), en el cual el porcentaje de dorado en las capturas fue de 4.43% afuera de las 50 mn. Difiere de los resultados obtenidos por Sarmiento Náfate et al. (2010) en la evaluación tecnológica de la pesquería de pequeña escala de atún aleta amarilla (Thunnus albacares) y otros pelágicos mayores realizados en la misma zona de estudio que el presente trabajo. Éstos realizaron 57 lances y obtuvieron 293 organismos de los cuales 54% fue de dorado. Conclusiones Se determinó que el dorado fue la especie que se capturó con mayor frecuencia con los tres tamaños de anzuelo. La selectividad de los tres tamaños de anzuelos circulares fue el siguiente: con C13/0 de 84.05 cm Lf, el C14/0 obtuvo ejemplares de 89.29 cm Lf y con el C15/0 de 93.06 cm Lf. Literatura citada Macías ZR, AL Vidaurri S, H Santana H, JJ Valdéz F. 1999.
Análisis descriptivo de las capturas obtenidas por las embarcaciones ribereñas palangreras en el Estado de Colima .CRIP - Manzanillo, Colima. Instituto Nacional de la Pesca. México. 11 p. Inédito.
Rodríguez-Valencia JA, MA Cisneros-Mata, H Ortega-Casillas, I Castro-Leal, G Rodríguez-Domínguez, A Chávez-Castro, LG Rodríguez-Delgado. 2008. Anzuelos circulares como opción para reducir la captura incidental en las operaciones pesqueras de los palangreros ribereños de Sinaloa (México). Ciencia Pesquera 16: 67-78.
Santana-Hernández H, IM Gómez-Humarán, JJ Valdez-Flores, MC Jiménez-Quiroz. 2008. Experimento para determinar la selectividad y eficiencia del palangre de deriva con tres tipos de anzuelo y dos tipos de carnada en la pesca de tiburón con embarcaciones de mediana altura en el Pacifico Central mexicano. Ciencia Pesquera 16: 57-66.
Sarmiento-Náfate S, H Santana-Hernández, J Villalobos-Toledo, HA Gil-López, JA Agustín-Jiménez, C Pineda-García, M Martínez-Aquino, A Labastida-Che. 2010. La pesca ribereña del atún aleta amarilla (Thunnus albacares Bonaterre, 1788) mediante el uso de palangres en la costa de Oaxaca. Informe técnico. INAPESCA-CRIPSCO. 44p.
Sparre P, SC Venema. 1995. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1. Manual. Documento Técnico de pesca, FAO 306/1. Valparaiso, Chile. 420 pp.
Palabras clave: Palangre, capturas, eficiencia, selectividad, anzuelos

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
249
121: oral ALGUNOS ASPECTOS BIOLÓGICOS DEL DORADO (Coryphaena hippurus)
EN LA COSTA DE OAXACA Y CHIAPAS
Ada Lisbeth Núñez Orozco, Aldrin Labastida Che, José Alfonso Oviedo Piamonte y Eduardo Ramos Santiago
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prolongación Playa Abierta s/n. Col. Miramar, Salina Cruz, Oax. C. P. 70680. Tels: 01(971)7145003 y 7140386. [email protected], [email protected], [email protected], [email protected] Introducción El dorado (Coryphaena spp.) es un recurso conformado por especies circumtropicales oceánicas epipelágicas, restringidas a aguas cálidas con temperaturas superiores a 20°C (Gibbs y Collete 1959, Collete 1995), de importancia pesquera a escala mundial, y aprovechada en ambos litorales de México. Se trata de peces altamente migratorios, que alcanzan longitudes cercanas a los 2 m y pesos de hasta 40 kg. Son muy apreciados en la pesca deportiva y capturados de manera incidental por la pesca artesanal. Por la excelente calidad de su carne se convierte en una especie con amplia demanda en términos comerciales. La conjunción de conocimientos sobre la biología de las especies para la zona de estudio y la generación de información en aquellas áreas en las que no se ha realizado investigación al respecto, posibilita un mejor conocimiento de la dinámica del recurso y así mismo aporta información necesaria para implementar las medidas de manejo pertinentes. Material y métodos Se analizaron los datos de febrero a diciembre de 2014 obtenidos de los muestreos biométricos de la captura comercial de Puerto Escondido y Huatulco, Oaxaca; Paredón y Puerto Madero, Chis. Se registraron datos de longitud furcal (Lf), peso total (P), sexo y madurez sexual de los organismos. Se determinó la relación morfométrica longitud-peso por medio de la función potencial alométrica y se aplicó la prueba t Student, para comprobar la significancia estadística del valor de b (Zar 1996). Se determinó la proporción macho-hembra y se aplicó la prueba X2 para comprobar una proporción 1:1. Se estimó la longitud de primera madurez al 50% de la población (L50) mediante el ajuste de la proporción de individuos maduros en cada intervalo de longitudes a una función logística. Los parámetros de crecimiento fueron estimados mediante un análisis de progresión modal (Sparre y Venema 1995) y modelados con la ecuación de crecimiento de von Bertalanffy (1938). Resultados Se obtuvo una muestra total de 1,306 organismos, con una longitud promedio de 81.25 cm Lf y peso promedio de 5.3 kg. Presentó un intervalo de
longitudes de 44 a 145 cm Lf, con una moda en 61 cm Lf (Fig.1).
Fig. 1. Distribución de longitudes de Coryphaena hippurus. El modelo potencial de la relación longitud furcal-peso total para ambos sexos fue P = 0.000009 Lf2.97 (r2 = 0.952), lo cual indica que el crecimiento es de tipo isométrico (t(1305,2) = 0.0116; p = 0.99), es decir, el crecimiento en peso es proporcional al incremento en longitud. La relación longitud-peso para machos y hembras no presentó diferencias significativas (b = 2.99 para machos y b = 2.92 para hembras; p>0.05). La relación por sexos presentó una proporción 1.3:1, con un porcentaje mayor de machos (57%) que de hembras (43%) (X2 = 22.39; p = 0.000002). De acuerdo a los registros mensuales de las fases de madurez gonádica se observaron organismos maduros durante todo el periodo de estudio, lo cual puede indicar que esta especie se reproduce durante todo el año; sin embargo presenta un periodo reproductivo más acentuado de junio a septiembre (Fig. 2).
Fig. 2. Comportamiento de los estadios de madurez gonádica de las hembras de Coryphaena hippurus.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
250
El análisis de los organismos con base en la curva sigmoidea indica que esta especie alcanza la longitud de primera madurez a los 76 cm Lf (Fig. 3).
Fig. 3. Longitud de primera madurez de Coryphaena hippurus.
La ecuación de crecimiento presentó una longitud infinita L = 156 cm Lf, una tasa de crecimiento k = 0.78 año-1, y una longitud cero to = -0.1306; la edad relativa en la cual los dorados alcanzan el crecimiento máximo asintótico es a los 3.8 años (Fig. 4).
Fig. 4. Curva de crecimiento para Coryphaena hippurus.
Conclusiones La población de dorado presentó un intervalo de longitudes de 44 a 145 cm Lf. La relación longitud-peso, presenta un crecimiento isométrico. La fase de madurez gonádica III y IV se presentaron durante todo el año, con mayor incidencia en los meses de junio a septiembre. La longitud de primera madurez se determinó en 76 cm Lf. El dorado es una especie de crecimiento rápido k = 0.78.
Literatura citada Collete BB. 1995. Coryphaenidae. En: Fisher W, F Krupp,
W Schneider, C Sommer, KE Carpenter & VH Niem (eds). Guía FAO para la identificación de especies para los fines de la Pesca. Pacífico Centro-Oriental, volumen II. Vertebrados - Parte 1, pp. 1036-1038. FAO, Roma.
Gibbs RH, BB Collette. 1959. On the identification, distribution and biology of the dolphins, Coryphaena hippurus and C. equiselis. Bull. Mar. Sci. Gulf Carib. 9: 117-152.
Sparre P, SC Venema, 1995. Introducción a la evaluación de recursos pesqueros tropicales. Parte 1 - Manual. FAO Doc. Tec. de Pesca 306/1, Roma, 420p.
von Bertalanffy L. 1938. A quantitative theory of organic growth. Hum. Biol. 10: 181-213.
Zar JH. 1996. Biostatistical analysis. Prentice-Hall, Inc. New Jersey, United States.
Palabras clave: longitudes, madurez gonádica, longitud de primera madurez, crecimiento.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
251
122: cartel ESTRUCTURA DE LONGITUD ASOCIADA A LA CAPTURA DE BATOIDEOS
EN EL SUR DEL PACÍFICO MEXICANO
Ana Bricia Guzmán Castellanos1, Genoveva Cerdenares Ladrón de Guevara2, Alejandra Lizárraga Rodríguez1, Adriana Alatorre Alba1 y Oswaldo Morales Pacheco1
1Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prolongación Playa Abierta, Col. Miramar S/N, Salina Cruz, Oaxaca. 2Universidad del Mar. Carretera a Zipolite, S/N. Puerto Ángel, Oaxaca. [email protected] Introducción En años recientes la captura de elasmobranquios a nivel mundial ha ido en aumento (Bizzarro et al. 2007) comercializándose sólo 25% de las especies y el resto descartadas en su gran mayoría batoideos (rayas) (Agnew et al. 1999). En el sur del Pacífico mexicano las rayas se capturan incidentalmente o son fauna de acompañamiento (FAC) dentro de la pesquería de camarón que se realiza en la zona de pesca número 90, Golfo de Tehuantepec (GT) (INAPESCA 2004), donde se utilizan redes de arrastre de fondo poco selectivas, de 10 a 114 m de profundidad (6 a 62 brazas) (Núñez-Orozco et al. 2013), y donde se capturan todas las longitudes. A pesar de que los batoideos en el GT han sido reportados como especies recurrentes y abundantes se desconocen las generaciones (cohortes) por especie capturadas. Identificar cohortes es importante porque permite evidenciar e identificar la correlación entre el tamaño (longitud) de las especies y su vulnerabilidad ante una presión de pesca. Materia y métodos Los organismos fueron obtenidos como FAC en la pesca de arrastre de camarón en el GT (14°10' N, 92°15' O - 16°13' N, 95°55' O) (Fig. 1) de enero-agosto 2015. Cada espécimen fue medido con un vernier electrónico con precisión de 0.1 mm para organismos pequeños y con una cinta métrica graduada en centímetros para organismos grandes. Cada individuo se identificó a nivel de especie (Fischer et al. 1995). La estructura de longitudes de cada especie se analizó a partir de la longitud total (Lt, cm), con objeto de identificar el número de cohortes susceptibles a la pesca de arrastre, a través de una distribución multinomial que identificó el número de cohortes dada una función de máxima verosimilitud (Morales-Bojórquez y Pacheco-Bedoya 2016) para machos y hembras de manera conjunta. La bondad de ajuste de esta función es sensible en la identificación de valores modales asociados a una distribución normal, la cual proporciona el número de salidas específicas del valor promedio de la cohorte en longitud total y de la varianza asociada (Haddon 2001). Los datos fueron ajustados cuando el logaritmo de verosimilitud –��� fue minimizado mediante el
algoritmo de búsqueda directa de Newton (Neter et al. 1996).
Fig. 1. Zona de pesca 90, para la captura de camarón. Resultados y discusión Se identificaron 867 rayas y 11 especies recurrentes: Urotrygon chilensis (Günther 1872), U. nana Miyake y McEachran 1988, U. rogersi (Jordan y Starks 1895), U. aspidura (Jordan y Gilbert 1882), Rhinoptera steindachneri Evermann y Jenkins 1891, Rhinobatos leucorhynchus Günther 1867, R. glaucostigma Jordan y Gilbert 1883, Aetobatus narinari (Euphrasen 1790), Dasyatis longa (Garman 1880), Zapteryx xyster Jordan y Evermann 1896, Narcine entemedor Jordan y Starks 1895 y N. vermiculatus Breder 1928. De acuerdo a la Unión Internacional para la conservación de la Naturaleza (IUCN) la gran mayoría de estas especies se encuentra sin ninguna categoría debido a que los datos sobre su biología son insuficientes; además se consideran vulnerables ante la presión de pesca por ser estrategas K (Stevens et al. 2000). Por el número de total de organismos U. chilensis representó 31.1% de las especies capturadas, seguida por N. vermiculatus 23.58%, U. nana 14.98%, U. rogersi 7.67% y U. aspidura 7.55%, el resto de las especies (R. steindachneri 4.53%, R. leucorhynchus 3.60%, A. narinari 2.9%, D. longa 1.39%, Z xyster 1.28%, N. entemedor 0.35% y R. glaucostigma 0.35%); las dos últimas no fueron consideradas en los análisis por el bajo número de organismos. Tapia-García y García-Abad (1998) y Medina-Baustista (2011) reportan estas especies como 50%

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
252
de la abundancia de elasmobranquios que se capturan en el GT. Con el análisis de distribución de frecuencias de Lt (cm) U. nana (n = 129) tuvo 5 cohortes, con longitudes entre 8 y 26 cm Lt (Fig. 2a). U. rogersi (n = 66) presentó 4 cohortes con una longitud mínima de captura de 16 cm Lt (Fig. 2b). Se estimaron 3 cohortes: N. vermiculatus (n = 203) (Fig. 2c), R. leucorhynchus (n = 31) (Fig. 2d), A. narinari (n = 25) (Fig. 2e), D. longa (n = 12) (Fig. 2f) y G. marmorata (n = 13) (Fig. 2g). U. chilensis (n = 267) (Fig. 2h), U. aspidura (n = 65) (Fig. 2i) y Z. xyster (n = 11) (Fig. 2j) estimaron dos cohortes y R. steindachneri (n = 39) presentó una cohorte (Fig. 2k). De algunas especies se capturan todas las longitudes, lo que es un impacto en sus poblaciones, incluidas hembras grávidas, con un efecto negativo sobre el rendimiento reproductivo de las poblaciones (Braccini et al. 2006).
Fig. 2. Estimación del número de cohortes para rayas del Golfo de Tehuantepec, calculado desde una función multinomial a partir de Longitud total (cm). Los datos de las once especies de rayas no fueron separados por sexos debido a la presencia/ausencia de machos o hembras que se registraron en algunas especies, sin embargo, hay una diferencia entre la frecuencia de longitud respecto a sexos por características asociadas a la segregación sexual, época reproductiva y eventos de migración y competencia (Klimley 1987). El análisis por sexos para cada especie permitira identificar las diferencias en el número de cohortes a través del tiempo y estimar la distribución en la estructura de longitudes que son más susceptibles a la pesca de arrastre en el GT. Literatura citada Agnew DJ, CP Nolan, J Pompert. 1999. Management of the
Falkland Islands skate and ray fishery. En: R Shotton (ed.). Case studies of the management of Elasmobranch fisheries. FAO Fisheries Technical Paper, 378: 268-284.
Bizzarro JJ, WD Smith, JF Márquez–Farías, RE Hueter. 2007. Artisanal fisheries and reproductive biology of the golden cownose ray, Rhinoptera steindachneri Evermann and Jenkins, 1891, in the northern Mexican Pacific. Fisheries Research. 84: 137– 146.
Braccini JM, BM Gillanders, TI Walker. 2006. Determining reproductive parameters for population assessments of chondrichthyan species with asynchronous ovulation and parturition: piked spurdog (Squalus megalops) as a case study. Marine and Freshwater Research. 57:105–119.
Fischer W, F Krupp, W Schneider, C Sommer, KE Carpenter, VH Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. FAO. Vol. II: 648-746.
Haddon M. 2001. Modelling and quantitative methods in fisheries. Chapman and Hall, CRC Press, Boca Raton.
Klimley AP. 1987. The determinants of sexual segregation in the scalloped hammerhead shark Sphyrna lewini. Environmental Biology of Fishes 18(1): 27–40.
Medina-Baustista J. 2011. Elasmobranquios capturados por la pesquería de arrastre camaronero en el Golfo de Tehuantepec. Tesis de Licenciatura. UMAR. Oaxaca. México. 82 p.
Morales-Bojórquez E, JL Pacheco-Bedoya. 2016. Population dynamics of jumbo squid Dosidicus gigas in Pacific Ecuadorian Waters. Journal of Shellfish Research 35: 211-224.
Neter J, MH Kutner, W Wasserman, J Nachtschien. 1996. Applied linear statistical models. McGraw-Hill/Irwin. Chicago. 1408 p.
Nuñez-Orozco AL, A Labastida-Che, JA Oviedo-Piamonte. 2013. Composición y abundancia de la íctiofauna en la franja sublitoral del Golfo de Tehuantepec, Oaxaca-Chiapas. Ciencia Pesquera 21: 29-40.
Stevens JD, R Bonfil, NK Dulvy, PA Walker. 2000. The effects of fishing on sharks, rays and chimaeras (Chondrichthyans) and the implications for marine ecosystems. ICES Journal of Marine Science. 57:476-494.
Tapia-García M, MC García-Abad. 1998. Los peces acompañantes del camarón y su potencial como recurso en las costas de Oaxaca y Chiapas. Universidad Autónoma Metropolitana. México, D.F. 970-654 p.
Palabras clave: Golfo de Tehuantepec, rayas, pesca de arrastre, captura incidental.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
253
123: oral ESTRUCTURA POBLACIONAL DE Carcharhinus falciformis Y Sphyrna lewini
EN EL GOLFO DE TEHUANTEPEC
Ana Alejandra Lizárraga Rodríguez, Adriana J. Alatorre Alba, Ana B. Guzmán Castellanos, J. L. Hernández Corona, Oswaldo Morales Pacheco y Marco A. Martínez Muñoz
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prolongación Playa Abierta, Col. Miramar S/N, Salina Cruz, Oaxaca. [email protected] Introducción La actividad pesquera de tiburón en el Golfo de Tehuantepec (GT) se caracteriza por ser multiespecífica y multiartes. Se explota por la pesca artesanal e industrial local que opera tanto en la zona costera como oceánica, además de capturarse como fauna de acompañamiento en las pesquerías dirigidas a escama y camarón (Alejo-Plata et al. 2006). La flota pesquera dirigida a tiburones opera principalmente con palangre de superficie; la cual captura un gran volumen y riqueza de especies, de las que se han documentado 21, siendo Carcharhinus falciformis (Bibron in Müller & Henle 1839) (35.7%) y Sphyrna lewini (Griffith & Smith 1834) (22.7%) las de mayor abundancia (Soriano-Velásquez et al. 2006). Estas especies tienen importancia ecológica y económica, lo que hace importante establecer la información biológico-pesquera con el fin de aprovechar de manera sustentable el recurso y evitar el colapso de esta pesquería. Material y métodos Los datos analizados provienen de la flota artesanal de tiburón (palangre), escama (redes agalleras) y camaronera (red de arrastre) del Golfo de Tehuantepec durante 2013 (de junio a noviembre), 2014 (de junio a diciembre) y 2015 (marzo a diciembre). Se registró la captura por especie mediante la utilización de las claves de identificación de Compagno et al. (1995) y la guía de campo de Martínez-Ortiz (2010). El sexo fue determinado por la presencia (machos) y ausencia (hembras) de gonopterigios, agrupándose en tres categorías, en función de la fase de madurez, utilizando la escala propuesta por Stehman (2002): neonatos (donde además se consideró a los juveniles de S. lewini de 75 cm LT máxima y C. falciformis de 79 cm LT máxima), juveniles (en maduración) y adultos (Alejo-Plata et al. 2006), verificando la proporción de sexos mediante una prueba de X2 entre los valores observados y esperados. La estructura de longitudes (LT) fue analizada en histogramas de frecuencias para cada sexo en STATGRAPHICS versión 16.1, y se aplicó una prueba t para comparar las longitudes entre sexos.
Resultados Se registraron 3,346 organismos pertenecientes a 10 especies, de las cuales C. falciformis con 68% y S. lewini con 29% sostienen la pesquería. El 3% se distribuyó entre ocho especies: Rhizorpionodon longurio y C. limbatus (2%), el resto está representado por C. leucas, Alopias pelagicus, Galeocerdo cuvier, Nasolamia Velox, Triaenodon obesus y Ginglymostoma cirratum. Carcharhinus falciformis (aleta de cartón) registró 2,285 organismos, con una distribución de longitudes entre 47 y 281 cm LT (promedio = 113, d.e. = 36.42), 53% fueron machos y 47% hembras (Fig. 1), ambos sexos con mayor incidencia en agosto (27%).
Fig. 1. Estructura de longitudes por sexo de Carcharhinus falciformis. No se presentaron diferencias significativas entre el promedio de las longitudes por sexo (t = 1.78; p> 0.07). Las capturas se componen principalmente por juveniles (73%) con longitudes de 80 a 171 cm LT y una proporción de sexos 0.83:1 (H: M), los neonatos presentaron una incidencia de 19% con longitudes entre 47 y 79 cm LT y una proporción de 0.94:1, mientras que 8% restante correspondió a organismos adultos entre 172 y 281 cm LT con una proporción de 1:1.11. La prueba X2 mostró tanto para neonatos (X2
= 0.333, p<0.05) como para adultos (X2 = 0.537, p<0.05), que no hay diferencias significativas entre sexos, mientras que para los juveniles hubo diferencia significativa (X2 = 13.67, p<0.05).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
254
Sphyrna lewini (tiburón martillo) registró 960 ejemplares; 48% fueron hembras y 52% machos, las longitudes oscilaron de 33.5-383 cm LT (promedio = 59.8, d.e. = 22.8) con una proporción de sexos de 1:1 (Fig. 2). Agosto presentó el mayor número de organismos (52.7%). En el análisis de frecuencia de longitudes se excluyeron 11 organismos con longitudes mayores a 170 cm LT que no son representadas en la distribución.
Fig. 2. Estructura de longitudes por sexo de Sphyrna lewini. Las longitudes por sexo no presentaron diferencias significativas en sus medias (t= 0.4; p= 0.63). De la captura de S. lewini, 93% está conformada por neonatos con longitudes de 33.5-75 cm LT y una proporción de sexos de 0.92:1. El 6% fueron organismos juveniles de longitudes 75-144.5 cm LT con una proporción de sexos 1:1; el 1% restante corresponde a los organismos adultos de longitudes de 171 a 383 cm LT, 0.22:1. La prueba X2 mostró que en los neonatos y juveniles no existe diferencias significativas entre sexos (X2=1.38 p<0.05; X2=0.0 p<0.05); mientras en los adultos sí hubo diferencia (X2 = 4.45, p<0.05). Discusión Se observó que C. falciformis se compone principalmente por juveniles en la pesca dirigida a tiburón, empleando palangre de superficie. La proporción de sexos resultó de 1:1 durante todo el año, lo cual coincide con lo reportado por Alejo-Plata et al. (2006) para Salina Cruz. La mayor parte de los organismos de S. lewini proviene de la pesca incidental de camarón y escama; por lo que en su mayoría (96%) está constituida por neonatos, puesto que se encuentran en zonas cercanas a la costa (Puerto Escondido y Huatulco), las cuales coincidieron con las reporta Alejo-Plata et al. (2006). Adicionalmente, la mayor presencia de los organismos se encuentra estrechamente relacionada con las boca barras de Tonalá, Boca del Cielo y Río Suchiate, lo que los hace susceptibles a su captura con el uso de redes, corroborando la importancia que
tienen estas áreas tanto de crianza como de alimentación para ambas especies. Literatura citada Alejo-Plata MC, S Ramos-Carrillo, JL Cruz-Ruiz. 2006. La
pesquería artesanal del tiburón en Salina Cruz, Oaxaca, México. Ciencia y Mar. 10(30): 37-51.
Compagno LJV, F Krupp, W Schneider. 1995. Tiburones. in: Fisher W, F Krupp, W Schneider, C Somer, KE Carpenter y VH Niem (eds). Guía FAO para la identificación de especies para los fines de la pesca. Pacífico Centro-Oriental, II, Vertebrados, parte 1, Roma.
Martínez-Ortiz J. 2010. Guía de campo para la identificación de los principales tiburones del Océano Pacífico Oriental. Proyecto APECFWG01/2001 T. Primera Edición. Guayaquil, Ecuador. 20 pp.
Soriano-Velásquez SR, DE Acal Sánchez, JL Castillo-Geniz, N Vázquez-Gómez, CE Ramírez Santiago. 2006. Tiburón del Golfo de Tehuantepec. En: Sustentabilidad y Pesca Responsable en México. Instituto Nacional de la Pesca. 323-366 pp.
Stehmann MFW. 2002. Proposal of a maturity stages scale for oviparous and viviparous cartilaginous fishes (Pisces, Chondrichthyes). Archive Fishery Marine Research. 50(1): 23-48.
Palabras clave: Pesquería, Tiburón, Neonatos, Estructura de longitudes.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
255
124: cartel DISTRIBUCIÓN, ABUNDANCIA Y DIVERSIDAD ESPACIAL DE LA ICTIOFAUNA DESCARTADA EN LA
PESCA DEL CAMARÓN EN EL GOLFO DE TEHUANTEPEC
Marco A. Martínez Muñoz1; Adolfo Gracia Gasca1; Oswaldo Morales Pacheco2; Saúl Sarmiento Náfate2; Ricardo Ramírez Murillo3; Adriana J. Alatorre Alba2 y Ana Alejandra Lizárraga Rodriguez2
1Laboratorio de Ecología Pesquera de Crustáceos. Instituto de Ciencias del Mar y Limnología. UNAM. A.P. 04510, Ciudad de México. C.P. 70-305. [email protected]; [email protected]; 2Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Prol. Playa Abierta S/N. Colonia Miramar. Salina Cruz, Oaxaca. C.P. 70680. [email protected]; [email protected], [email protected]. 3Instituto de Educación Media Superior del D.F. Plantel Tlalpan I. Av. San Lorenzo #290, Col. del Valle Sur, Ciudad de México. C.P. 03100 [email protected]. Introducción La preocupación por las elevadas capturas de especies de peces no objetivo en la pesquería de camarón, ha estado focalizada en especies de importancia económica que son el objetivo de otras pesquerías comerciales, o en especies vulnerables (Stobutzki et al. 2001). Un número significativo de especies no comerciales se capturan incidentalmente y se descartan al mar, con excepción de algunos grandes peces que se comercializan. El trabajo tiene como objetivo caracterizar los patrones espaciales de la distribución de la composición, abundancia, y de la diversidad de la ictiofauna asociada a la captura del camarón, en la plataforma continental del Golfo de Tehuantepec. Materia y métodos Los muestreos de pesca se llevaron a cabo durante la época de veda del camarón en el Golfo de Tehuantepec, entre abril-agosto de 2003-2004-2005, en 62 estaciones de operaciones de pesca (Fig. 1).
Fig.1. Área de estudio, subzonas y red de estaciones. El arte de pesca utilizado fue una red camaronera, con abertura de la boca de 24.4 m, en operación dos redes. La duración por lance, fue de una hora. Se realizaron entre 12 y 70 m, a una velocidad de 2 nudos tanto de día como de noche. Se realizaron 321 arrastres. Se tomaron 15 kg de muestra. La identificación taxonómica se determinó con las claves de Robertson y Allen (2002).
Los datos cuantitativos de la abundancia se expresaron en términos de densidad = ind/ha y biomasa = kg/ha, para esto fue necesario calcular el área de barrido. La diversidad se estimó con la ecuación de Shannon-Weaver. La abundancia, y diversidad fueron analizadas por crucero, subzona, región y estrato de plataforma. Resultados La distribución de la abundancia presentó variaciones en la densidad: las mayores (512 ind/ha) se detectaron en la región norte (subzona 91) en aguas costeras del sistema lagunar Huave y frente al sistema lagunar Mar Muerto. Mientras que al sur del sistema estuarino Chantuto-Panzacola, se localizaron densidades más bajas. La biomasa presentó un patrón similar, con valores máximos (29 kg/ha) en la zona norte que disminuyen gradualmente hacia el sur. Las mínimas (3 kg/ha) se localizaron hacia el sur de Carretas-Pereyra (subzona 95). La distribución espacial de la densidad presentó los valores más altos entre Carretas y Chantuto. De manera similar la distribución de la biomasa fue alta frente al sistema lagunar Huave disminuyendo hacia la región sur. La mayor diversidad se detectó en la región sur (Carretas y Chantuto) y la mínima se localizó frente al sistema Huave (Fig. 2). Las familias Haemulidae y Paralichthyidae, presentaron la máxima densidad en la subzona 91, disminuyendo gradualmente en las siguientes zonas. La biomasa mostró variaciones, resaltan Paralichthyidae, Haemulidae y Carangidae en la 91 disminuyendo hasta la 94. Se observó un patrón inverso en Ariidae y Polynemidae (Fig. 3). Ocho especies presentaron amplia distribución (>40%) y abundancia (>77%): destacan Haemulopsis axillaris, Selene peruviana, Diapterus brevirostris y Urotrygon chilensis.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
256
Fig. 2. Distribución espacial de la densidad (a), biomasa (b) y diversidad (c) de la ictiofauna demersal.
Fig. 3. Variación espacial de la abundancia de la ictiofauna a) Densidad y b) Biomasa por subzona y crucero. La variación de la diversidad fue en ascenso de norte a sur, ubicando la menor cerca del sistema Huave. El máximo se alcanza en el sur del sistema Chantuto. La diversidad mostró una tendencia a disminuir con la profundidad. Discusión Los trabajos de Acal y Arias (1990) y de Tapia-García (1998) constatan altos valores de abundancia y
biomasa frente a los sistemas Huave y Mar Muerto por abajo de 50 m, con los máximos en primavera-verano y mínimos en invierno, y un patrón similar al observado aquí. Esta tendencia se asocia con la productividad primaria de las aguas someras. La distribución y abundancia espacial de los peces siguen un patrón controlado por la dinámica climática (épocas de vientos, precipitación y de sequía), que a su vez se asocia al aumento de la productividad causada por afloramientos, en la región norte del golfo.
Fig. 4. Variación de la diversidad batimétrica y entre subzonas de la ictiofauna demersal. Conclusión Se encontraron diferencias en la heterogeneidad espacial y ambiental entre subzonas y profundidades. La plataforma interna está caracterizada por una mayor abundancia; la media presenta una alta diversidad, mientras que la externa, sólo hay pocas especies dominantes, debido a su estrategia adaptativa a la hipoxia. Literatura citada Acal ED, A Arias. 1990. Evaluación de los recursos
demerso-pelágicos vulnerables a redes de arrastre de fondo en el sur del Pacífico de México. Ciencias Marinas 16: 93-129.
Robertson DR, GR Allen. 2002. Shore fishes of the Tropical Eastern Pacific: an information system. Smithsonian Tropical Research Institute, Balboa, Panamá.
Stobutzki IC, MJ Miller, P Jones, JP Salini. 2001. Bycatch diversity and variation in a tropical Australian Penaeid fishery; the implications for monitoring. Fish. Res. 53: 283-301.
Tapia-García M. 1998. Evaluación ecológica de la fauna demersal. p. 129-148. In: Tapia-García M (ed.). El Golfo de Tehuantepec: el ecosistema y sus recursos. UAM-I, México, D. F.
Palabras clave: Abundancia, diversidad, distribución, ictiofauna, Golfo de Tehuantepec.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
257
125: oral LA PESCA RIBEREÑA DE ESCAMA MARINA EN TAMAULIPAS Y NORTE DE VERACRUZ
Ma. Guadalupe Gómez Ortiz, Hortensia López Navarrete, Brenda Yanet Rangel Arteaga y
Diana Guadalupe Herrera Gómez
Centro Regional de Investigación Pesquera de Tampico. INAPESACA. Prolong. Altamira s/n, Col. Isleta Pérez. Zona Centro, Tampico, Tamaulipas. C.P. 89000. [email protected] Introducción Las especies de escama marina que conforman diversas pesquerías multiespecíficas en el Golfo de México y Mar Caribe representan alrededor de 60% de la producción pesquera total (DOF 2012), Esta actividad está compuesta de una gran variedad de especies de peces en Tamaulipas, contribuye con 2.92% al PIB nacional y ocupa 13º lugar a nivel nacional por su producción pesquera de 30,744 t, que representa 1.76% respecto a la producción nacional. Veracruz contribuye con 4.65% al PIB nacional y ocupa el 5º lugar a nivel nacional por su producción pesquera con 69,631 t, que representa 3.99% respecto a la producción nacional (SAGARPA 2013). La actividad pesquera en ambas zonas es importante en la estructura social y de generación de ingresos de un gran número de pescadores que habitan las comunidades aledañas a las zonas de costa, siendo toda pesca de pequeña escala o artesanal. Se utiliza una gran diversidad de artes de pesca que varían en sus materiales de construcción, dimensiones y sistemas de operación según sea el recurso objeto de la pesca. Con el propósito de regionalizar el manejo del grupo multiespecífico “escama marina”, el objetivo de este proyecto que inició desde 2010 en Tamaulipas y 2012 en Veracruz, va dirigido a establecer las bases para el manejo regionalizado en el Golfo de México y Mar Caribe, permitiendo dar un seguimiento apropiado al desarrollo de las pesquerías y dictaminar las medidas de manejo que faciliten su sustentabilidad. Material y métodos Los resultados de este trabajo son producto de muestreos realizados en 2014 en la zona centro y sur de Tamaulipas y en el norte de Veracruz, directamente en las zonas de descarga de las embarcaciones, apoyados con entrevistas al pescador sobre métodos de pesca, equipos y artes, jornada de trabajo, así como captura por viaje. Los muestreos se realizaron de las principales especies comerciales registrando datos morfométricos de longitud y peso, y en lo posible sexo y madurez gonadal. Por zona de pesca se caracterizó la diversidad de especies presentes con su identificación taxonómica y se construyó la estructura de las poblaciones en longitud así como la caracterización de las artes y métodos de pesca.
Resultados Se monitorearon 53 especies de peces de las cuales 23 fueron en Tamaulipas representando 43% y 52 en el norte de Veracruz representando 98%. En Tamaulipas las especies se agruparon en 11 familias donde la Lutjanidae representó 64% con 6 especies, el huachinango Lutjanus campechanus el más abundante con 74%; la familia Scombridae representó 20% e incluye 2 especies, la sierra Scomberomorus maculatus (81%) y peto S. cavalla (19%) (Fig. 1).
Fig. 1. Proporción de la captura de escama por familia y zona costera de Tamaulipas y norte de Veracruz, 2014. En el norte de Veracruz las especies se integraron en 21 familias donde la familia Arridae representó 46% con 2 especies de bagre, el bandera Bagre marinus (52%) y el banderilla Auriopsis felis (48%); la familia Scombridae representó 14% con 3 especies de las cuales la sierra representó 84%; 13% lo representó la familia Lutjanidae con 8 especies donde el huachinango (41%) y besugo Rhomboplites aurorubens (37%) fueron los más abundante (Fig. 1). Del total de especies económicamente importantes y presentes en los muestreos de las capturas comerciales por zona, fueron 19 que incidieron en ambas zonas de captura, donde el huachinango, villajaiba Lutjanus synagris, cubera L. cyanopterus y cochino Balistes capriscus fueron más abundantes en la zona centro-sur de Tamaulipas, mientras que sierra, peto, boquilla Haemulon plumieri, medregal Seriola zonata, cabrilla Hyporthodus adscencionis, negrilla Mycteroperca rubra y pargo mulato L. griseus fueron más abundantes en el norte de Veracruz. El besugo se presentó en la misma proporción en ambas zonas. El bagre bandera, banderilla, sargo,
0 10 20 30 40 50 60 70
ArridaeScombridae
LutjanidaeHaemulidaeCarangidae
ScianidaeTetraodontidae
GerreidaeSerranidae
CentropomidaeSparidae
BalistidaeSphyraenidaeSynodontidae
BrotulidaeCoryphaemidae
LabridaeEphippidae
OphichthidaeElopidae
Frecuencia relativa
Norte de Veracruz
Tamaulipas

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
258
ratón Menticirrhus saxatilis, jurel aleta amarilla Caranx hippos, bonito Euthynnus alleteratus y ronco amarillo sólo se presentaron en el norte de Veracruz (Fig. 2).
Fig.2. Proporción especies importantes económicamente en la zona costera de Tamaulipas y norte de Veracruz, 2014. Dada la característica multiespecífica de la pesca hay una gran variedad de especies y longitudes en los muestreos, debido a que las flotas ribereñas utilizan diversos artes que van desde líneas o cordeles simples hasta palangres y redes. Por viaje de pesca, los pescadores, dependiendo de la especie objetivo, pueden obtener de 1 a 8 especies y en promedio 4.
Fig.3. Frecuencia de uso de artes de pesca en la zona costera de Tamaulipas y norte de Veracruz, 2014. En Tamaulipas las redes agalleras utilizadas son exclusivamente para la pesca de sierra, mientras que en el norte de Veracruz además usan la robalera y la palotera (truchas). Las líneas de pesca en Tamaulipas son los cordeles, las ristras, los palangres y las cimbras, mientras que en el norte de Veracruz ristras, palangres y cimbras (Fig. 3). Discusión y conclusiones La distribución regional de los recursos de escama en el Golfo de México trasciende límites estatales e incluso nacionales, y presenta variaciones espacio-temporales de distribución y abundancia, que dan lugar a diversas asociaciones y ensambles de especies, según la zona, la temporada y los sistemas de pesca, generando pesquerías multiespecíficas.
El enfoque monoespecífico para las regulaciones resulta poco objetivo y por lo tanto con escaso nivel de resultados en el desarrollo sustentable de las pesquerías. A través del programa de coordinación regional se tienen avances en cuanto a la implementación de una base de datos con estructura única para los estudios de peces costeros, donde ya se revisaron las tablas y su contenido, mínimas necesarias para el acopio de todos los datos generados tanto de los muestreos como la información estadística y el diseño conceptual para que todas las variables puedan ser analizadas de forma integral (Quiroga-Brahms et al. 2014). Literatura citada DOF. 2012. Carta Nacional Pesquera. SAGARPA. Diario
Oficial de la Federación 24 de agosto de 2012. Quiroga-Brahms C, E Romero-Hernández, MG Gómez-
Ortiz, V Caballero-Chávez, B Álvarez-López, A Wakida-Kusunoki, A Hernández-Flores, J Villanueva-Fortanelli, K Ramírez-López, H López-Navarrete, L Pantoja-González, G Pantoja-Yépez, AG Díaz-Álvarez, C Monroy-García, J Santos-Valencia, G Galindo-Cortez. 2014. Programa de coordinación regional de la investigación sobre peces óseos del Golfo de México y Caribe. SAGARPA-INAPESCA. Informe de Investigación 2013.
SAGARPA. 2013. Anuario estadístico de acuacultura y pesca. Dirección de Estadística y registros pesqueros. SAGARPA-CONAPESCA. México.
Palabras clave: especies, multiespecífica, líneas, agalleras.
0
20
40
60
80
100
120
Fre
cu
en
cia
re
lati
va
Centro-Sur Tamaulipas
Norte de Veracruz
2%
67%
1
27%
3%
Norte de Veracruz
14%
62%
14%6%
5%
TamaulipasCORDEL
RISTRAS
PALANGRES
CIMBRAS
RED SIERRERA

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
259
126: cartel FAUNA ASOCIADA A LA CAPTURA DE CAMARÓN EN LA RIBERA ADYACENTE A LA BOCA SUR DEL
SISTEMA SANTA MARIA, SINALOA
Lorenia Guadalupe Gómez Medina, Darío Chávez Herrera y Yolene Rosalía Osuna Peralta
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Avenida Camarón Sábalo S/N, Estero del Yugo, Mazatlán, Sinaloa. C.P. 82000. e-mail: [email protected] Introducción La pesca de camarón por arrastre es señalada como destructiva debido al volumen considerable de fauna asociada de camarón (FAC). La Organización de Naciones Unidas para la Agricultura y la Alimentación, está promoviendo estudios con vistas a disminuir los daños que pueden producir en el ambiente marino y determinar las medidas más adecuadas para lograrlo. Esas medidas deberán servir para subsanar las deficiencias de las prácticas de pesca (Covantes-Rodríguez et al. 2011). El objetivo de este trabajo es estimar la estructura y dinámica de las comunidades de los grupos que componen la FAC en la ribera centro de Sinaloa. Material y métodos El área de estudio se localiza en Santa María, Sinaloa. Se consideró la información de los muestreos del año 2015. Las muestras se separaron e identificaron utilizando diversas claves, se tomó el peso total (PT, g), la longitud total (LT, mm) y el sexo. La distribución de longitudes se ajustó con un modelo multinomial. Se calculó la proporción entre hembras y machos. Se calcularon 3 índices, abundancia relativa (Rodríguez-Romero et al. 2008), valor biológico Sanders (1960) e importancia relativa (Pinkas et al. 1971). Resultados y discusión En total se capturaron 2,363 kg de FAC por 138.7 kg de camarón lo que hace una proporción de 17:1, valores por encima de los encontrados en el 2014 en el mismo sistema (Gómez-Medina et al. 2015). En la variación mensual de FAC el valor mínimo fue en noviembre (82.5%) y el más alto en diciembre (98.4%). El intervalo de longitudes para el total de peces fue de 53 a 415 mm de LT y las longitudes donde los organismos fueron más abundantes (85.9%) estuvo entre los 80 y 170 mm de LT (Fig. 1), esto es parecido con el sistema lagunar bahía Magdalena-Almejas, BCS (Rábago-Quiroz y García-Borbón 2015). En su mayoría las especies estaban en estado juvenil.
Fig. 1. Distribución de longitudes de las 55 especies de peces de la fauna asociada a la captura de camarón durante el 2015. La profundidad donde se encontró la mayor ocurrencia de peces fue a las 12 brazas. Para Balistes polylepis fue a las 5 brazas con longitudes de 72 a 112 mm LT. Se encontraron dos grupos modales para hembras con media en: 100 y 125 mm LT y para los indeterminados un grupo modal con media en 100 mm LT (Fig. 2).
Fig. 2. Distribución de longitud-frecuencia (mm) de Balistes polylepis, durante el 2015.
020406080
100120140160180200220240
50 70 90 110
130
150
170
190
210
230
250
270
290
310
330
350
370
390
410
430
450
Ab
un
dan
cia
(Nú
m. d
e O
rgan
ism
os)
Longitud Total (mm)
n = 1,018

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
260
Fig. 3. Distribución de longitud-frecuencia (mm) de Sphoeroides annulatus, durante el 2015. Para la especie Sphoeroides annulatus la mayor ocurrencia fue a las 12 brazas con longitudes de 95 a 310 mm LT, se encontraron dos grupos modales para machos y hembras con media en: 225 y 275 mm; 125 y 250 mm LT, respectivamente (Fig.3). La mayor ocurrencia de Lutjanus guttatus fue a las 12 brazas con longitudes de 72 a 310 mm LT. Se encontró un grupo modal para los indeterminados con media en: 125 mm LT (Fig. 4).
Fig. 4. Relación entre longitud total (mm) y peso total (g) de Lutjanus guttatus, durante el 2015. Tabla 1. Valores de los Índices de IAR, IVB e IIR de las especies de mayor abundancia en la FAC. Familia Especie IAR IVB IIR Carangidae Selene peruviana A 45 2078 Gerreidae Eucinostomus
argenteus A 22 1555
Paralichthyidae Cyclopsetta querna A 34 1720 Etropus crossotus A 45 1717
Tetradontidae Sphoeroides annulatus
A 10 2162
En la tabla 1 se muestra que de acuerdo al IIR las 4 primeras especies fueron las más abundantes con bajo valor comercial a excepción de S. annulatus, coincidiendo con los resultados encontrados en los muestreos del 2014 para el mismo sistema (Gómez-Medina et al. 2015).
Conclusión Es necesario dar continuidad a los muestreos y un seguimiento de la FAC para establecer los posibles impactos sobre las comunidades que integran cada uno de los grupos. Literatura citada Covantes-Rodriguez C, JL Beraud-Lozano. 2011. Pesca de
camarón de altamar en Mazatlán, Sinaloa. Aprovechamiento sostenible y conservación de los recursos vivos de altamar. 1 ed. Universidad Autónoma de Sinaloa, Culiacán, México. 235 pp.
Gómez-Medina LG, D Chávez-Herrera, YR Osuna-Peralta, MA Osuna-Zamora. 2014. Fauna asociada a la captura de camarón en la plataforma continental adyacente a la boca Sur del sistema Santa María La Reforma Sinaloa. Informe Técnico. INAPESCA.
Pinkas L, MS Oliphant, LK Iverson. 1971. Food habits of albacore, bluefin tuna and bonito in California waters. Fish Bulletin 152: 1-105.
Rábago-Quiroz CH, JA García-Borbón. 2015. Caracterización de la fauna de acompañamiento del camarón durante el período de veda 2014, en el sistema lagunar Bahía Magdalena-Almejas, B.C.S. Informe Técnico. INAPESCA.
Rodríguez-Romero J, DS Palacios-Salgado, J López-Martínez, S Hernández-Vázquez, G Ponce-Díaz. 2008. Composición y zoogeografía de los peces demersales de la costa occidental de Baja California Sur, México. Rev. Biología Tropical 56: 1765-1783.
Sanders HL. 1960. Benthic studies in Buzzards Bay III. The structure of soft-bottom community. 5: 138-153.
Palabras clave: FAC, Estructura de longitudes, Sinaloa.
0
1
2
3
4
5
6
7
8
9
75 100 125 150 175 200 225 250 275 300 325 350 375 400 425
Fre
cuen
cia
abso
luta
(n)
Longitud Total (mm)
MachosHembrasMachos calculadasHembras calulada

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
261
127: oral RELACION ENTRE LA CAPTURA DEL PALANGRE TIBURONERO RIBEREÑO Y LA RED DE ARRASTRE
CAMARONERO: ESPECIES COMPARTIDAS CON VALOR COMERCIAL
Jorge L. Oviedo Pérez1, Heber Zea de la Cruz1, Luis E. Martínez Cruz1, Leticia González Ocaranza2 1Centro Regional de Investigación Pesquera de Veracruz. INAPESCA. Av. Ejército Mexicano 106, Boca del Río, Veracruz, México. C.P. 94298. [email protected]; 2
Centro Regional de Investigación Pesquera de Lerma. INAPESCA. Km 5, Carr. Campeche-Lerma, Campeche. C. P. 24500; 3Universidad Veracruzana. Introducción La pesquería ribereña de tiburones y rayas con palangres y la pesquería de arrastre de camarón tienen varios aspectos en común: 1) Se realizan sobre la plataforma continental, la de tiburones hasta 150 m de profundidad y la de camarón de 9 a 86 m. 2) Los equipos de pesca operan sobre el fondo marino, el palangre en reposo y la red de arrastre en movimiento. 3) La captura es multiespecífica, la de palangres reporta 99 especies y la de arrastre 146. 4) Se capturan elasmobranquios y teleósteos, en el palangre, que representan respectivamente 71% y 29% en número de organismos; mientras que en el arrastre representan el 4 y 11% del peso. Este trabajo contribuye a explicar la relación entre los peces con valor comercial asociados al palangre tiburonero y a la red de arrastre camaronero. Material y métodos Considerando que el INAPESCA realiza monitoreo anual de la composición de la captura de ambas pesquerías en el Golfo de México, se examinaron los resultados obtenidos en el CRIP Veracruz durante las campañas de investigación 2001 a 2015 sobre tiburones y rayas y 2004 a 2015 sobre la captura incidental en el arrastre camaronero. En el monitoreo de la flota palangrera tiburonera se contabilizaron e identificaron a nivel de especie todos los organismos capturados. En el arrastre camaronero el monitoreo de la captura se realizó a bordo; la producción fue clasificada en seis grupos de especies (camarones, peces retenidos a bordo, otros crustáceos retenidos, moluscos retenidos, elasmobranquios y descartes) determinando el peso en kilogramos para cada uno de los grupos en cubierta. Adicionalmente para los peces retenidos y elasmobranquios se determinaron número de organismos, especies, longitudes y peso por especie. Los resultados de las dos pesquerías se compararon y se determinaron las especies compartidas que tienen valor comercial. Resultados y discusión En el palangre tiburonero, en número de organismos, 26 especies de tiburones representan 58% de la captura, 7 de rayas 13% y 67 de teleósteos 29%. En la pesquería de arrastre, en cuanto a peso, 8 especies de tiburones y 8 de rayas representan 4% de la captura y 65 especies de teleósteos retenidos a bordo representan el 11% (Fig. 1).
Fig. 1. Composición de la captura del arrastre camaronero en el Golfo de México. Las pesquerías comparten 12 especies de elasmobranquios. A tiburones corresponden 7 especies: Rizoprionodon terraenovae, Carcharhinus brevipinna, Mustelus cannis, M. norrisi, Squatina dumeril, S. mexicana, Squalus cubensis. Las especies más importantes del palangre son R. terraenovae y C. brevipinna, pero en el arrastre su presencia es irrelevante. Las especies más importantes en el arrastre son S. dumeril y S. mexicana, pero en el palangre su contribución es irrelevante. Las rayas corresponden a 5 especies: Dasyatis americana, Gymnura micrura, Rhinobatus lentiginosus, Rhinoptera bonasus, Aetobatus narinari. En ambas pesquerías son importantes D. americana y G. micrura. En el arrastre es importante R. lentiginosus, pero es irrelevante en el palangre. La importancia de cada una de estas especies se muestra en la tabla 1, expresada en contribución porcentual en número de organismos en la captura de elasmobranquios del palangre y la red. El grupo de teleósteos retenidos a bordo en el arrastre camaronero está formado por 27 familias y 65 especies. Los chiles Saurida normani, Synodus foetens y Trachinocephalus myops; los chivos Upeneus parvus y Mullus auratus; los lenguados Cyclopsetta chittendeni, Syacium gunteri y Syacium microrum son las 8 especies más importantes, representan 60.7% del peso de este grupo de especies y no se registran en la captura del palangre tiburonero. En 29.3% restante de los teleósteos retenidos se han encontrado 31 especies comunes para ambas pesquerías. En la tabla 2 se muestran las especies comunes; en el palangre tiburonero la importancia de cada una es expresada como su
n = 200,694 kg kg

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
262
contribución porcentual en número de organismos en la captura de teleósteos (Fig. 2), mientras que en la red de arrastre la importancia es expresada como su contribución porcentual en peso de la captura de teleósteos retenidos. Tabla 1. Tiburones y rayas compartidos
TIBURONES
Palangre % # elasmobranquios
n = 43,636
Arrastre % # elasmobranquios
n = 13,767 R. teraenovae 54% 0.01 % C. brevipinna 1.4% 0.01% M. cannis 0.02% 0.02% M. norris 0.01% 0.06% S. dumeril 0.01% 0.01% S. mexicana ---------- 1.44% S. cubensis 0.8% 0.02%
RAYAS D. americana 13.3% 3.8% G. micrura 3.4% 24.3% R.lentiginosus 0.1% 14.43% R. bonasus 0.3% 0.01% A. narinari 0.3% 0.04%
Tabla 2. Teleósteos compartidos
TELEOSTEOS “otros” = 19% del resto de especies
del palangre (Fig. 2)
Palangre % #
Teleósteos n=18,229 org
Arrastre % peso Teleósteos
n=22,513 kg Caranx hippos Caranx latus
17 0.11%
Scomberomorus cavalla 15 0.10% Bagre spp. 9 0.06% Lutjanus cyanoptera 9 0.001% Lutjanus analis Lutjanus griseus
8 0.011%
Lutjanus campechanus 7 1.7% Seriola dumerili 5 0.09% Rachycentron canadum 2 0.02% Conodon nobilis otros 6.03% Lagocephalus laevigatus otros 4.73% Balistes capriscus otros 4.35% Menticirrhus americanus otros 1.98% Priacanthus arenatus otros 1.80% Scorpaena plumieri otros 1.37% Calamus calamus otros 1.12% Aluterus heudeloti Aluterus scriptus
otros 0.89%
Rhomboplites aurorubens otros 0.63% Brotula barbata otros 0.53% Lutjanus synagris otros 0.12% Kyphosus sectatror otros 0.02% Trachinotus carolinus otros 0.01% Haemulon spp. otros 0.0% Caranx crysos otros 0.01% Sphyraena barracuda otros 0.01% Epinephelus flavolimbatus E. drummondhayi
otros 0.011%
Archosargus probatocephalus otros 0.001% Centropomus undecimalis otros 0.001% Scomberomorus maculatus Sin registro 0.03%
Fig. 2. Composición porcentual de la captura de teleósteos del palangre en número de organismos. Conclusiones La pesquería de tiburón con palangre y la pesquería de camarón con red de arrastre son multiespecíficas. Comparten zonas de pesca con fondos suaves en la plataforma continental, en profundidades de 9 a 86 m. Las especies comunes para ambas pesquerías son 43, de éstas corresponden 7 a tiburones, 5 a rayas y 31 a teleósteos. La mayoría de especies comunes más importantes en una pesquería, son irrelevantes o ausentes en la otra. ólo 3 especies son importantes en ambas pesquerías: Lutjanus campechanus. D. americana y G. micrura. En otras 7 se consideran con un nivel de importancia menor: C. nobilis, L. laevigatus, B. capriscus, M. americanus, P. arenatus, S. plumieri, C. calamus. Literatura citada CRIP-Veracruz. 2001–2015. Sobre pesquería de
elasmobranquios 2004-2015. Sobre captura incidental del arrastre camaronero. Ineditos. Informes Técnicos Documentos Internos.
Palabras clave: Palangre, arrastre camaronero, especies compartidas, incidentalidad.
n=18,229 organismos

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
263
128: cartel EXPLORACIÓN DE INFORMACIÓN OFICIAL DE LAS PRINCIPALES ESPECIES COMERCIALES DE
ESCAMA MARINA EN EL ESTADO DE BAJACALIFORNIA SUR
Gabriel Antonio Jiménez Llanos, Marcela Selene Zúñiga Flores, Armando Vega Velázquez y Jose Carlos Monroy Hernández
Centro Regional de Investigación Pesquera de La Paz. INAPESCA. Carretera a Pichilingue km 1 s/n, Col. Esterito, La Paz, Baja California Sur. C. P. 23020. [email protected] Introducción En Baja California Sur se practica mayoritariamente la pesca de pequeña escala, sobre todo en las zonas cercanas a la costa, debido a la limitada autonomía de las embarcaciones, además de que no requiere una fuerte inversión. Exhibe diversos grados de desarrollo en infraestructura, equipamiento y organización social para la producción, procesamiento y comercialización de las capturas. La capacidad de producción está sujeta al número y tamaño de las embarcaciones, así como a la eficiencia de las artes de pesca (Cortés et al. 2006). Existe una gran variedad de especies de escama marina, que de acuerdo al valor comercial que constituyen, se agrupan y aprovechan pasa su venta o autoconsumo. Material y métodos Se hizo el análisis de los resúmenes mensuales de la información oficial de avisos de arribo, proporcionados por la CONAPESCA del periodo de enero de 2004 a diciembre de 2015 esta información contiene: Oficina de Pesca donde se arribó, mes, año, especie y su presentación, peso desembarcado y peso vivo ambos en kilogramo, así como el valor del producto y valor por kilogramo en pesos mexicanos en el estado de Baja California Sur. Se clasificó la información por Oficina de Pesca para escama marina, mantas y rayas. Con esto se obtuvo el peso desembarcado en toneladas (kg/1000). Asimismo la producción por Oficina de Pesca se calculó mediante el peso desembarcado promedio en toneladas en el mismo periodo de tiempo. Se analizó el peso desembarcado por especie en toneladas del 2015, se realizó mediante porcentajes ((peso desembarcado (t) por especie X 100)/ peso desembarcado (t) de escama marina 2015 de BCS) para observar las especies de escama marina con mayor representación en los volúmenes de captura de la última temporada. Resultados La producción de escama marina para el estado de BCS se encuentra entre las 14,307 toneladas anuales, las mantas y rayas aportan cerca de 1,048 toneladas anuales (Fig. 1). Las Oficinas de Pesca que tienen mayor registro de volúmenes son Cd. Constitución, Santa Rosalía, Puerto San Carlos, La Paz y Abreojos (en conjunto 85%).
Cd Constitución con 5,530 t anuales aporta 32% de la producción de escama marina en BCS (Fig. 2).
Fig. 1. Producción histórica de escama marina (línea punteada), escama marina incluyendo mantas y rayas (línea solida) en el estado de Baja California Sur.
Fig. 2. Producción promedio de escama marina (t) por Oficina de Pesca (2004 a 2015), en el estado de Baja California Sur. La figura 3 muestra las especies en un orden de categorización en porcentaje. En orden de importancia en 2015 están el verdillo, jurel, huachinango, pierna, corvina, cochi, lisa, lenguado, manta, sierra, cabrilla, blanco, cardenal, mojarra, botete, cabaicucho, ratón, pargo, roncacho, pámpano el conjunto de estas especies representa 14,630 toneladas para 2015 y 86% de la producción de BCS, el 14 % restante equivalente a 2,400 toneladas

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
264
anuales son el conjunto de las demás especies arribadas.
Fig. 3. Porcentaje de captura (barras azules) y porcentaje acumulado (línea punteada) de las principales especies escama marina en B.C.S. en 2015. Discusión y conclusiones El estado de Baja California Sur cuenta con pesquerías de alto valor comercial como la de Langosta. Vega-Velázquez (2006) menciona que 83% de la población de langosta se encuentra en la península de Baja California, sin embargo no todas las unidades económicas pesqueras cuentan con permiso o concesión de este recurso pesquero. Sin embargo la mayoría de los productores cuentan con permisos comerciales de escama marina, este permiso se ampara la pesca de mantas y rayas estos se encuentra protegido bajo NOM-029-PESC-2006, (DOF 2015) lo cual se limita a una restricción pesquera durante cierta época del año. La principales localidades de producción en volumen de escama marina en B.C.S son Cd. Constitución, Santa Rosalía, Pto. San Carlos y La Paz, probablemente por el gran número de organizaciones que se dedican a la pesca en esas localidades. La Zona Pacífico Norte que lo conforma Punta Abreojos, Bahía Asunción, Bahía Tortugas son ocho organizaciones pesqueras que se encuentran más organizadas para aportar un valor agregado a las capturas de escama. La mayor producción pesquera de escama marina lo aporta el verdillo (P. nebulifer) cerca de 16%, para el cual se está haciendo un plan de manejo para su sustentabilidad como pesquería. En segundo lugar el jurel (Seriola lalandi), con porcentaje por arriba de 10% de la producción en B.C.S., este recurso incluso se le da un valor agregado en algunas sociedades cooperativas, en donde se envasa en diferentes presentaciones, en filete para venta local, nacional e internacional. La información analizada da una idea de cómo se encuentra distribuido el recurso escama marina y la
importancia de cada especie para el estado y por localidad. Sin embargo es necesario precisar que el análisis de la información oficial debe ser complementado con datos biológicos, sociales, ambientales y económicos, todo esto para la obtención de conclusiones y hacer una toma de decisiones en función del mejor manejo integral de una pesquería. Literatura citada DOF. 2015. PROYECTO de Modificación a la Norma Oficial
Mexicana NOM-029-PESC-2006, Pesca responsable de tiburones y rayas. Especificaciones para su aprovechamiento. Diario Oficial de la Federación: 11/02/2015.
Vega-Velázquez A. 2006. Langosta de la península de Baja California. En: Arreguín Sánchez et al. Sustentabilidad y pesca responsable en México: evaluación y manejo 2001-2005. Instituto Nacional de la Pesca/SAGARPA. México. P. 155-210.
Palabras clave: avisos de arribo, producción, oficina de pesca, escama marina.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
265
129: oral CAPTURA INCIDENTAL EN LA PESQUERIA DE LANGOSTA EN BAJA CALIFORNIA SUR
José Carlos Monroy Hernández, Armando Vega Velásquez, Gabriel Antonio Jiménez Llanos,
Marcela Selene Zúñiga Flores y Edgardo Camacho Bareño
Centro Regional de Investigación Pesquera de La Paz. INAPESCA. Carretera a Pichilingue km 1 s/n, Col. Esterito, La Paz, Baja California Sur. C. P. 23020. tiburó[email protected] Introducción Una de las principales pesquerías con alto valor económico en México, es la langosta. La península de Baja California (PBC) cuenta con los mayores volúmenes de captura en el país. La pesquería está compuesta por tres especies: roja (Panulirus interruptus), azul (P. inflatus) y verde (P. gracilis). P. interruptus es de afinidad templada y es el principal recurso en valor y volumen, alcanzando las mayores producciones en la zona centro de la PBC. Las otras dos especies langosta azul (P. inflatus) y la langosta verde (P. gracilis) son más abundante en la zona sur de la PBC, ya que son de afinidad tropical. El Instituto Nacional de Pesca desde la década de los setenta mantiene un programa de investigación y monitoreo de manera continua hasta la actualidad, donde se tiene actualizada una gran cantidad de información científica, sobre su ciclo de vida y sus principales procesos biológicos, aspectos pesqueros, económicos y sociales. Ello permite evaluar las estrategias para su administración y manejo, y coadyuvar a su conservación y aprovechamiento sostenido y sustentable (Vega-Velazquez 2006). La pesquería de langosta en la PBC se realiza mediante trampas de alambre con sus características descritas y normadas en el Diario Oficial de la Federación con base en la NOM-006-PESC-1993 (DOF 31-12-1993), a diferencia de otras partes del país donde su captura se realiza mediante redes de enmalle y buceo. Una de las principales características de las trampas langosteras es que cuenta con un dispositivo de escape (excluidor) que libera aquellas langostas juveniles y longitudes no comerciales, así mismo también sirve para la exclusión de la fauna de acompañamiento (captura incidental). Los organismos de longitudes más grandes que la del diámetro del excluidor quedan atrapadas y son liberadas hasta que las trampas son revisadas. En algunos casos la captura incidental cuenta con especies de valor comercial como pueden ser crustáceos, peces, tiburones y moluscos. Material y métodos La información que se utilizó para realizar el análisis de captura incidental en las trampas langosteras en la zona de Bahía Asunción, La Bocana, y Punta Abreojos, BCS (Fig. 1), fue recopilada de las bitácoras de pesca que son enviadas al CRIP-La Paz, por las Cooperativas de la zona y llenadas por
los mismos pescadores de cada día de pesca, reportando el número de langostas de longitud legal y de longitud sublegal, así como las especies de captura incidental y el número de organismos por cada especie. La información de la captura incidental proviene de varios campos pesqueros de la zona como son San Roque, Punta Prieta, San Hipólito, La Bocanita y Punta Abreojos.
Fig.1. Zona de pesca de langosta para Bahía Asunción, La Bocana y Punta Abreojos B.C.S. Resultados Se recopiló información de las bitácoras de pesca diarias de la pesquería de langosta, de septiembre a febrero 2014-2015, de la zona de Bahía Asunción, La Bocana y Punta Abreojos BCS. De 120,901 trampas revisadas en los diferentes campos pesqueros de la zona, se encontró que la especie con mayor abundancia o incidencia en las trampas langosteras, fue la jaiba azul (Callinectes bellicosus) con 65%, seguido por el grupo de los peces (16%), donde las viejas (Semicossyphus pulcher) fueron los peces más abundantes. Los pulpos (Octopus bimaculatus) representaron 9%. En el caso de los tiburones se tiene 7% y el restante lo complementan anguilas (Muraena helena) y un mínimo de caracoles ermitaños (Dardanus arrosor) (Fig. 2 y Tabla 1).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
266
Fig. 2. Principales especies capturadas en las trampas langosteras para la zona de Bahía Asunción, La Bocana y Punta Abreojos 2014-2015. Tabla.1. Número de organismos capturados por trampa.
Especies Número de organismos
Frecuencia relativa (%)
jaibas 3,481 2.88 peces 838 0.69 pulpos 463 0.38 tiburones 396 0.33 caracoles 128 0.11 anguilas 47 0.04
Discusión y conclusión La captura incidental o fauna de acompañamiento en las trampas langosteras es relativamente bajo, a diferencia de otras pesquerías, como la de camarón o de escama marina. En la región donde se obtuvieron los datos, da una idea de la distribución y abundancia de qué tipo de especies de crustáceos, peces, tiburones y moluscos, se encuentran en las zonas de pesca analizadas. Gran número de organismos de la captura incidental son aprovechados por el pescador para venta local o autoconsumo. Otras especies que se registran no tienen un valor comercial pero se utilizan como carnada para las mismas trampas langosteras o para las líneas de mano. Hay una mínima cantidad de fauna, que no se utiliza ni para consumo, ni para carnada y es liberada nuevamente al agua, principalmente los tiburones, como el tiburón perro (Heterodontus mexicanus) y el gata (Ginglymostoma cirratum). Literatutara citada DOF. 1993. Norma Oficial Mexicana NOM-006-PESC-
1993, Para regular el aprovechamiento de todas las especies de langosta en las aguas de jurisdicción federal del Golfo de México y Mar Caribe, así como del Océano Pacífico incluyendo el Golfo de California. Diario Oficial de la Federación. 1ª sec. p. 50.
Vega-Velázquez A. 2006. Langosta de la península de Baja California. En: Arreguín Sánchez F., (eds.) Sustentabilidad y pesca responsable en México: evaluación y manejo 2001-2005. Instituto Nacional de la Pesca, SAGARPA. México. P. 155-210.
Palabras clave: Captura incidental,Pesquería de langosta, excluidor.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
267
130: oral FUNCIÓN DE VEROSIMILITUD CONJUNTA BASADA EN DISTRIBUCIONES DE PROBABILIDAD NORMAL
Y MULTINOMIAL PARA ANALIZAR LA VARIABILIDAD FENOTÍPICA EN EL CRECIMIENTO DE ALMEJA DE SIFÓN Panopea globosa
Marlene Anaid Luquin Covarrubias1, Enrique Morales Bojórquez1
, Sergio Scarry González Peláez1, 2 y Daniel Bernardo Lluch Cota1
1Centro de Investigaciones Biológicas del Noroeste. Instituto Politécnico Nacional 195. Col. Playa Palo de Santa Rita Sur. La Paz, B.C.S., México. C.P. 23096. 2 Universidad Autónoma de Baja California Sur, Carretera al Sur km 5.5, La Paz, Baja California Sur. México. C. P. 23080. [email protected], [email protected], [email protected], [email protected] Introducción Las almejas del género Panopea han mostrado una alta variabilidad en la composición de longitud de concha-edad, sin embargo, los métodos usados para modelar el crecimiento individual han sido comúnmente ajustados a trayectorias que ignoran la variabilidad biológica intrínseca en la población. Bajo este contexto, es importante considerar dos perspectivas: si la variabilidad en la longitud de concha-edad disminuye con el tiempo o la edad el fenómeno es conocido como crecimiento compensatorio; por el contrario, si la variabilidad incrementa con la edad, se denomina crecimiento depensatorio (Gurney y Veitch 2007). En el presente estudio, un nuevo enfoque para analizar la variabilidad individual en el crecimiento de Panopea globosa fue desarrollado, incorporando datos de distribución de frecuencias de longitud de estadios tempranos y datos de longitud de concha-edad de individuos adultos mediante una función negativa de verosimilitud conjunta, suponiendo que un efecto depensatorio actúa sobre la población de Bahía Magdalena, BCS. Material y métodos Los datos de longitud de concha-edad �� fueron analizados asumiendo que los residuales son normalmente distribuidos, la función objetivo fue expresada como sigue:
La distribución de frecuencias de longitud dentro de cada grupo de edad a, fueron descritas usando la siguiente función de densidad probabilística:
Seis modelos de crecimiento candidatos 1) von Bertalanffy, (2) Gompertz, (3) Johnson, (4) Logístico, (5) generalizado von Bertalanffy (GVB) y (6) Richards, fueron ajustados a ambas fuentes de datos. Los parámetros i (ϒ, ω, L∞, k, t0, σ
2L∞, σ2
ϒ, α,
δ, λa1, λa2) fueron estimados minimizando una función negativa de verosimilitud conjunta (LJ ), mediante el algoritmo de Newton.
La varianza (σi
2) para cada edad observada i en los datos de longitud-edad fue estimada usando el siguiente criterio estadístico (Tabla 1). Tabla 1. Función matemática de varianza ��
para cada
edad i de acuerdo a los datos de longitud de concha-edad de P. globosa
Funciones matemáticas similares para cada modelo de crecimiento candidato fueron desarrolladas para estimar a y σa
2 para cada grupo de edad a observado en los estadios tempranos. El ajuste de los seis modelos de crecimiento candidatos a ambas fuentes de datos fue analizado mediante un enfoque de inferencia multi-modelo (IMM). Resultados Los parámetros de crecimiento fueron optimizados conjuntamente para cada modelo de crecimiento. La estimación de ��∞ y �Υ mostró que el modelo de VB (77.73 y 25.46, respectivamente) y el modelo Logístico (72.47 y 45.40, respectivamente) tuvieron los valores más altos (Fig.1). El efecto de ��∞ y �Υ influyó en la estimación de ��
donde las estimaciones de varianza para cada edad i fueron mayores en los modelos de VB y Logístico. El resto de los modelos de crecimiento mostraron

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
268
estimaciones similares con valores de σi2 = 20 (3
años) y σi2 = 60 (47 años).
Fig. 1. Comparación de la varianza ��∞ para individuos viejos y �Υ individuos jóvenes para cada modelo de crecimiento candidato analizado.
Fig. 2. Trayectorias de varianza ��
para cada edad i de
acuerdo a cada modelo de crecimiento candidato analizado.
El enfoque de inferencia multi-modelo basado en AICc mostró que el mejor ajuste a ambas fuentes de datos (frecuencia de longitud de concha y longitud de concha-edad) fue descrito por el modelo de crecimiento de Johnson (AICc = 6612.02, Δi = 0.00, wi = 0.47) (Fig. 2). Discusión En el presente estudio fue posible estimar la varianza para cada edad observada (��
) en la población de P. globosa de Bahía Magdalena, BCS. Esta característica no había sido previamente modelada para las especies Panopea, permitiendo analizar el incremento de longitud para individuos juveniles y adultos. Este nuevo enfoque basado sobre variabilidad fenotípica en el crecimiento describió satisfactoriamente estimaciones de t0 para seis modelos de crecimiento candidatos y una mejor interpretación biológica, mediante la inclusión de distribución de frecuencias de longitud asociadas a estadios tempranos. Finalmente, este análisis permite incluir más fuentes de datos en la función de verosimilitud conjunta (ej.
observaciones de marcado y recaptura), lo cual podría representar el ciclo de vida ontogénico de la especie, mejorando los parámetros. Además podría ser usado en modelos de evaluación de stock (ej. máximo rendimiento por recluta), el cual sería útil en el establecimiento de directrices de gestión para las especies Panopea en el Pacífico mexicano (Aragón-Noriega et al. 2012). Conclusiones El uso de una función negativa de verosimilitud conjunta mostró un alto desempeño estadístico para analizar explícitamente la variación en la estructura de edades, mejorando la estimación de los parámetros de crecimiento de P. globosa. Adicionalmente, mejoró la estimación del parámetro t0 y las trayectorias de crecimiento en el origen. Evitado valores de 0, así como valores negativos y altamente desviados. Se demostró un patrón de crecimiento depensatorio en P. globosa. La interacción de dos fuentes de datos mostró ser más informativa para estimar la trayectoria de crecimiento esperado con una mejor interpretación biológica. Este enfoque metodológico basado sobre variabilidad fenotípica en crecimiento de P. globosa se logró desarrollar de manera exitosa y original en 5 modelos de crecimiento, incluyendo 2 de tipo generalizado, y la rutina de selección de modelos. Literatura citada Aragón-Noriega EA, E Alcántara-Razo, LE Calderón-
Aguilera, R Sánchez-Fourcade. 2012. Status of geoduck clam fisheries in Mexico. Journal of Shellfish Research 31(3): 733–738.
Gurney WSC, AR Veitch. 2007. The dynamics of size-at-age variability. Bulletin of Mathematical Biology 69(3): 861–885.
Palabras clave: Variabilidad individual, longitud-edad, crecimiento, función de verosimilitud, AIC.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
269
131: oral CICLO REPRODUCTIVO DE LA JAIBA VERDE GUERRERA Callinectes bellicosus EN
BAJA CALIFORNIA SUR, MÉXICO
Verónica Castañeda Fernández de Lara1, Carlos Gómez Rojo1, Saúl González Romero1 y Juan Carlos Castro Salgado2
Centro Regional de Investigación Pesquera de La Paz. INAPESCA. Carretera a Pichilingue km 1 s/n, Col. El Esterito, La Paz, Baja California Sur, México. C. P. 23020. [email protected] [email protected], [email protected]; 2CICIMAR. IPN. Av. Instituto Politécnico Nacional s/n, Col. Playa Palo de Santa Rita. A.P. 592. La Paz, Baja California Sur, México. C.P. 23096. [email protected] Introducción La jaiba guerrera Callinectes bellicosus (Stimpson 1859) se distribuye desde el sur de California, EEUU hasta Perú. Es una especie meroplanctónica con distribución ontogenética diferencial: fase larval pelágica y fase bentónica en lagunas costeras y estuarios, donde crecen y se reproducen (Hernández y Arreola-Lizárraga 2007). En Baja California Sur C. bellicosus es la especie dominante de las jaibas y completa su ciclo de vida dentro de los cuerpos lagunares o bahías del estado (Castro-Salgado 2013). La jaiba en Bahía Magdalena se reproduce principalmente de julio a agosto y sus larvas son más abundantes en agosto, cuando la temperatura del agua es mayor, aunque se observó otro pico menor en primavera (4% del total de larvas encontradas en marzo y abril, Sánchez-Ortiz y Gómez-Gutiérrez 1992). Las postlarvas se observaron en mayor abundancia en septiembre 2009 utilizando colectores de superficie (Castañeda, Obser. Pers). Los juveniles regresan a la bahía, con un mayor número en la boca de la bahía. La tasa de crecimiento, densidad, tamaño, madurez y longitud de madurez depende de sus condiciones ambientales como temperatura y salinidad (Guillory et al. 2001). Las oscilaciones interanuales en las capturas podrían estar relacionadas con los fenómenos El Niño, La Niña (Huato-Soberanis et al. 2006). La comprensión del ciclo reproductivo de las especies y del período de máxima reproducción es escencial cuando se requiere emplear una veda como medida de manejo en una pesquería. En el caso de C. bellicosus existen dos periodos de veda en Sonora y Sinaloa. A) Del 1 de mayo al 30 de junio, y B) Del 1 al 9 de julio para hembras (DOF 2014). En BCS no se ha establecido un periodo de veda, no se capturan hembras y se respeta la longitud mínima legal. Sin embargo, las vedas de Sonora y Sinaloa podrían incentivar que pescadores o compradores vengan y promueven la captura de hembras y no respeto a la longitud mínima. Por lo tanto se planteó la pregunta: ¿Las vedas establecidas en Sonora y Sinaloa protegerían el pico reproductivo de C. bellicosus en BCS?
Material y métodos Se realizó la biometría mensual de 1,364 jaibas repartidas en Puerto San Carlos (PSC), Bahía Magdalena (BM) y La Laguna Ojo de Liebre (LOL), Guerrero Negro (GN), BCS durante 2013-2015. Se colectaron al azar 30 machos y 30 hembras en la captura comercial tomados a bordo de la embarcación de un pescador. Se registró la longitud, peso y madurez de todos los ejemplares colectados. Los dos sitios tienen establecida la pesquería de jaiba y difieren por 3 grados de latitud (Fig. 1), lo que posiblemente implique diferencias en los picos reproductivos.
Fig. 1. Cuerpos lagunares donde se captura la jaiba C. bellicosus en Baja California Sur, México. Se utilizó la escala de 6 estadios de madurez interna para hembras y 5 para machos: Inactivo (H1, M1), Gametogénesis inicial (H2, M2), Desarrollo (H3, M3), Madurez (H4, M4) y Ovígera (H5) y Re-absorción (R) (Castañeda-Fernández de Lara et al. 2015). Resultados Los machos de ambas zonas se reproducen práctimanete todo el año, con un máximo de primavera a verano. El pico reproductivo en BM fue en agosto. En 2015 en el caso de las jaibas de LOL, el periodo reproductivo de hembras inició en mayo y hubo mayor número de hembras con huevos en octubre, aunque se encontraron hembras en re-absorción hasta diciembre, que indica que la

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
270
actividad reproductiva continuó. Durante 2015 las jaibas machos de BM no presentaron un marcado pico reproductivo con un aumento en verano-otoño, es decir, desde julio hasta noviembre (Fig. 2a) y en hembras se observan estadios avanzados de madurez con huevos H5 y R iniciando en julio continuando incluso hasta diciembre 2015 (Fig. 2b).
Fig. 2. Madurez gonádica en a) machos y b) hembras de la jaiba de Puerto San Carlos, Bahía Magdalena (junio 2013-agosto 2014).
Fig. 3. Madurez gonádica de la jaiba de Puerto San Carlos, Bahía Magdalena (marzo a diciembre 2015), a) Machos, b) Hembras. El pico reproductivo de C. bellicosus en GN continuó hasta diciembre, pero inició marcadamente en octubre, desfasado del pico observado en BM (Fig. 3a, b y Fig. 4a, b). Discusión El pico reproductivo de C. bellicosus de BM durante 2013-2014, mostró el máximo en agosto. Durante 2015 hubo continuidad de junio a diciembre, diferente a lo observado previamente, lo que puede estar relacionado con las temperaturas cálidas del agua de mar (El Niño del 2015). El pico reproductivo de las hembras de LOL, GN fue en mayo-junio y octubre-diciembre; sin embargo,
dadas las condiciones ambientales de El Niño, se desconoce si este pico es normal o muestra desfasamiento.
Fig. 4. Madurez gonádica de la jaiba Callinectes bellicosus de Guerrero Negro, BCS de febrero a diciembre 2015 a)
Machos, b) Hembras. Conclusiones El periodo de veda establecido para Sonora y Sinaloa no es adecuado para proteger el pico reproductivo de C. bellicosus en las condiciones ambientales de calentamiento resultado del fenómeno de El Niño, durante 2013-2015. Literatura citada Castañeda-Fernández de Lara V, L Carvalho-Saucedo, JA García-
Borbón, C Gómez-Rojo, JC Castro-Salgado. 2015. Validación histológica de una escala morfocromática de maduración gonadal para la jaiba verde, Callinectes bellicosus. Ciencia Pesquera, número especial 23: 43-52
Castro-Salgado JC. 2013. Tendencias de producción pesquera de jaiba en la costa occidental de Baja California Sur, México (1998–2010). Tesis de Licenciatura. UABCS. 70p.
DOF. 2014. Acuerdo para modificar periodo de veda de jaiba frente al litoral de los estados de Sonora y Sinaloa. Diario Oficial de la Federación México. 13 de junio de 2014.
Guillory V, HM Perry, S VanderKooy (eds.). 2001. The blue crab fishery of the Gulf of Mexico, United States: A regional management plan. Gulf States Marine Fisheries Commission, Publication 96, Ocean Springs, Mississippi. 304p.
Hernández L, JA Arreola-Lizárraga. 2007. Estructura de longituds y crecimiento de los cangrejos Callinectes arcuatus y C. bellicosus (Decapoda: Portunidae) en la laguna costera Las Guásimas, México. Rev. de Biología Tropical 55(1): 225–233.
Huato-Soberanis L, M.J. Haro-Gardy, E Ramírez-Félix, LC López-González. 2007. Estudio socio-económico de la pesquería de jaiba en Sinaloa y Sonora. En: JB Santinelli. Situación del sector pesquero en México. Centro de Estudios para el Desarrollo Rural Sustentable y la Soberanía Alimentaria, pp: 50-93.
Sánchez-Ortiz CA, J Gómez-Gutiérrez. 1992. Distribución y abundancia de los estadios planctónicos de las jaibas Callinectes bellicosus (Decapoda: Portunidae) en el complejo lagunar Bahía Magdalena, BCS, México. Revista de Investigaciones Científicas de la Universidad Autónoma de Baja California Sur (Serie Ciencias Marinas) 3: 47–60.
Palabras clave: Jaiba, Callinectes bellicosus, reproducción, Baja California Sur.
0%
20%
40%
60%
80%
100%
RM4M3M2M1
0%
20%
40%
60%
80%
100%
RH5H4H3H2H1
b)
0%
20%
40%
60%
80%
100%
R
M4
M3
M2
M1
0%
20%
40%
60%
80%
100%
R
H5
H4
H3
H2
H1
b)
0%
20%
40%
60%
80%
100%
R
M4
M3
M2
M1
a)
0%
20%
40%
60%
80%
100%
R
H5
H4
H3
H2
H1
b)
a)
))
a)
))

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
271
132: oral EVALUACION Y MANEJO DE LA CURVINA GOLFINA Cynoscion othonopterus, EN
EL ALTO GOLFO DE CALIFORNIA
Concepción Enciso Enciso y Gabriela Galindo Cortez
Centro Regional de Investigación Pesquera de Mazatlán. INAPESCA. Calzada Sábalo-Cerritos S/N. Contiguo a Estero El Yugo, Mazatlán, Sinaloa. C. P. 82112. Tel. 6699881255 y 9881256. [email protected] Introducción La curvina golfina es la segunda pesquería de escama de mayor importancia social y económica en el Alto Golfo de California (AGC), es considerada endémica de la región y su captura se realiza principalmente en la Reserva de la Biosfera del Alto Golfo de California y Delta del Río Colorado (DOF 2007). Por estar sometida a una alta presión pesquera, es posible que su población pueda verse afectada, por lo que se hace necesario implementar estudios biológico-pesqueros que den los elementos para proponer medidas de manejo fundamentadas técnicamente, que permitan alcanzar la maximización del rendimiento sin poner en riesgo a la población. El estudio presenta una evaluación del stock en el cual se integran factores biológicos y económicos, permitiendo valorar diferentes escenarios para el establecimiento de una cuota de captura, como una medida de manejo. Material y métodos Se utilizaron datos de capturas de las temporadas de pesca de 1993 a 2015 reportadas en las comunidades pesqueras de San Felipe, Golfo de Santa Clara y Pueblo Indígena Cucapá. Para evaluar el recurso se utilizó el Modelo Bioeconómico de Schaefer-Gordon, incorporando parámetros biológicos del recurso y económicos de la pesquería. Para validar el modelo se utilizó la prueba no paramétrica de Kolmogorov-Smirnov para probar la hipótesis nula de que la distribución de los valores de captura observadas son iguales a aquellos simulados por el modelo. Se realizó un análisis de sensibilidad sobre aquellos parámetros con mayor incertidumbre en el modelo, como son: r, K, q, el costo por unidad de esfuerzo y el precio de la curvina, con relación a la captura al máximo rendimiento económico CMRE (Anderson y Seijo 2010). Se incorporó un análisis de riesgo evaluando la respuesta de la biomasa y la rentabilidad económica (BMRE y BEBE) ante diferentes escenarios de manejo (cuotas de captura). Resultados El modelo bioeconómico fue validado de manera exitosa respecto a los valores de captura observados y simulados (p = 0.983) (Fig. 1).
Fig. 1. Valores observados y simulados para la captura el periodo de 1995 a 2015. Los niveles de biomasa total se observan estables durante el periodo de estudio con un valor promedio cercano a 26,185 t (CV = 8.9 %), observándose una tendencia decreciente de 1994 a 2002, y en el 2006 vuelve a incrementarse sin alcanzar los niveles iniciales de la serie (Fig. 2).
Fig. 2. Biomasa del stock de curvina producido por el modelo para el periodo de 1995 a 2015. Se detectó que el valor de la capacidad de carga K y la capturabilidad q, son los parámetros más sensibles que explican el modelo en 71.51% de la variabilidad. La captura al máximo rendimiento económico (CMRE) obtenida fue de 3,606 toneladas en promedio de peso eviscerado, con un intervalo del 95% de confianza entre 3,602 y 3,610 toneladas (Fig. 3).
20,000
22,000
24,000
26,000
28,000
30,000
32,000
19
94
19
95
19
96
19
97
19
98
19
99
20
00
20
01
20
02
20
03
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
12
20
13
20
14
20
15
Bio
ma
sa
(t)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
272
Fig. 3. Distribución de la captura al máximo rendimiento económico. Promedio en línea negra punteada y percentiles 25 y 75% en líneas rojas punteadas. En el análisis de riesgo (Fig. 4) se proyectó la CMRE promedio obtenida (3606 t), encontrando que la probabilidad de que la biomasa de la población se mantenga superior a la BMRE es de 70.7%, por debajo de la BMRE es de 19.8% y por debajo de la BEBE es de 9.5%.
Fig. 4. Escenarios de riesgo simulado al año 2020 obtenidos con 100,000 simulaciones del modelo bioeconómico de Schaefer-Gordon. Conclusiones El modelo bioeconómico fue validado de manera exitosa respecto a los valores de captura observados y simulados. Los parámetros con mayor incertidumbre en la variable de interés, captura al máximo rendimiento económico fueron el valor de K y q. Desde un punto de vista precautorio, la cuota de captura para la temporada 2016 al máximo rendimiento económico CMRE, no deberá ser superior a 4,343 t de peso entero, que es equivalente a 3,606 t de peso eviscerado y un intervalo al 95% de confianza entre 3,602 t y 3,611 t. Literatura citada Anderson LG, JC Seijo. 2010. Bioeconomics of fisheries
management. Edition first published 2010. DOF. 2007. Norma Oficial Mexicana NOM-063-PESC-
2005, pesca responsable de curvina golfina (Cynoscion othonopterus) en aguas de jurisdicción federal del Alto
Golfo de California y Delta del Río Colorado. Especificaciones para su aprovechamiento. Pub. DOF, 16 de agosto de 2007. 1ra. Sección, pp. 30-35.
Palabras clave: Curvina, Modelo bioeconómico, Alto Golfo de California.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0
2000
4000
6000
8000
10000
12000
14000
10
00
14
00
18
00
22
00
26
00
30
00
34
00
38
00
42
00
46
00
50
00
54
00
58
00
62
00
Pro
ba
bil
ida
d a
cu
mu
lad
a
Fre
cu
en
cia
CMRE (100,000 iteraciones)
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
273
133: oral LA PESCA ARTESANAL DE PELÁGICOS MENORES EN BAJA CALIFORNIA SUR, MÉXICO
Carlos Gómez Rojo, Verónica Castañeda Fernández de Lara y Saúl Sánchez Romero
Centro Regional de Investigación Pesquera de La Paz. INAPESCA. Carretera a Pichilingue Km 1 s/n, Col. Esterito, La Paz Baja California Sur, México. C.P. 23020. [email protected], [email protected], [email protected] Introducción La pesquería de pelágicos menores es por su volumen de producción, la más importante en México (primer lugar de producción pesquera y quinto por valor). Se desarrolla principalmente en el noroeste del país y en los últimos años ha representado 48% de la producción pesquera nacional (CONAPESCA 2014). Son fuente importante de proteína para consumo humano directo y para la producción de alimento balanceado para la industria avícola, acuícola y porcina, así como carnada para pesca comercial, deportiva y artesanal, entre otros usos. La regulación de esta pesquería se establece en la NOM-PESQ-003-1993 (DOF 1993). Esta pesquería es multiespecífica, incide sobre sardina monterrey (Sardinops sagax), sardina crinuda, tres especies (Opisthonema libertate, O. bulleri y O. medirastre), macarela (Scomber japonicus), anchoveta norteña (Engraulis mordax), bocona (Cetengraulis mysticetus), sardina japonesa (Etrumeus teres), charrito (Trachurus symmetricus) y piña (Oligoplites altus, O. refulgens y O. saurus). La captura comercial de estas especies se realiza principalmente por embarcaciones pesqueras mayores (DOF 2012). En Baja California Sur también opera una flota de embarcaciones menores (menos de 15 m de eslora) dedicada a la explotación de pelágicos menores que utilizan como arte de pesca la red lámpara, similar a una red de cerco, pero de menores dimensiones, la captura se destina principalmente a abastecer de carnada a pesquerías de muy alta importancia económica en la región, como son: langosta, jaiba, pulpo, verdillo, tiburón, etc., principalmente en la costa occidental del Estado y pesca deportiva en Loreto, La Paz y Los Cabos. Las especies que aprovecha la flota menor son: sardina monterrey, macarela y sardina crinuda. El objetivo del presente trabajo es describir las características de esa flota así como la distribución temporal de la captura y esfuerzo. Material y métodos La información de captura y esfuerzo de pesca ejercido por la flota mayor y menor se obtuvo de las Oficinas Federales de Pesca, y complementada con datos proporcionados por los pescadores de la flota menor. Además, se recopiló información de las condiciones ambientales.
Resultados La flota menor que participó en la captura de pelágicos menores en la costa occidental del Estado en 2015 estuvo conformada por 19 embarcaciones menores, registradas en 5 unidades económicas, 2 del sector privado y 3 cooperativas. Las zonas de captura son cercanas a la costa, a no más de 2 km. Con respecto al año anterior, hubo una disminución de 50% tanto en el número de embarcaciones, como de unidades de producción dedicadas a esta actividad. Los puertos de descarga principales son: Bahía Tortugas, Bahía Asunción y Puerto San Carlos, en Bahía Magdalena (Fig.1).
Fig. 1. Puertos de desembarque de la flota menor sardinera en BCS. Se estima se realizaron 423 viajes de pesca de menos de 1 día de duración, y se descargaron 878.7 toneladas, lo cual es una fuerte disminución de la captura, pues sólo se capturó 30% de lo obtenido en 2014 (2,852.8 t). La composición de las capturas fue similar a la obtenida en 2014, 93.8% correspondió a sardina monterrey y 6.2% a macarela (Fig. 2).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
274
Fig. 2. Captura (t) de sardina monterrey y macarela capturadas por la flota menor en BCS durante 2015 La captura por unidad de esfuerzo promedio (toneladas /viaje de pesca) de la flota menor en BCS fue de 1.0 t/viaje, con los valores más altos de enero a febrero y de agosto a septiembre (Fig.3).
Fig. 3. Número de viajes y CPUE (captura (t) por viaje) de la flota menor que captura sardina en BCS en 2015 Discusión Las capturas de pelágicos menores en BCS durante 2015 se han visto afectadas por las condiciones ambientales que han prevalecido en el estado desde 2013, las cuales se han caracterizado por un incremento de la temperatura superficial del mar, y una disminución de la productividad primaria. Estas condiciones se han asociado a una serie de fenómenos oceánicos y atmosféricos, como son el debilitamiento de los vientos alisios, elevación de la temperatura en la Corriente de California, cambio en el periodo de PDO (oscilación decadal del Pacífico), así como el desarrollo de un evento de El Niño a partir del tercer trimestre de 2014. Esta situación también se reflejó en las capturas de la flota menor, la cual por su área de operación tuvo un acceso limitado a las existencias de sardina monterrey cercanas a la costa. En las capturas de la flota menor predominó la sardina monterrey (93.8%) y en las obtenidas la flota
mayor esta especie aportó 35.5% de la captura y la sardina crinuda 55.9%. Esta diferencia refleja la distribución de las especies a lo largo de la costa y las diferentes zonas de operación de cada flota. Conclusión La pesquería de pelágicos menores en Baja California Sur, se realiza por dos flotas de muy distinta composición que comparten la explotación de las mismas especies, pero no en las mismas zonas costeras, y el destino de las capturas de ambas flotas es diferente. Los instrumentos normativos actuales no reflejan esta complejidad, lo cual es una fuente potencial de conflictos entre ambas flotas, por lo que es necesario se realicen los estudios pertinentes que permitan incluir las actividades de la flota artesanal de pelágicos menores en la regulación vigente. Literatura citada CONAPESCA. 2014. Anuario estadístico de acuacultura y
pesca 2013. SAGARPA. Comisión Nacional de Acuacultura y Pesca Mazatlán, Sin. 385 p.
DOF. 1993. Norma Oficial Mexicana NOM-003-PESC-1993, Para regular el aprovechamiento de las especies de sardina Monterrey, piña, crinuda, bocona, japonesa y de las especies anchoveta y macarela, con embarcaciones de cerco, en aguas de Jurisdicción Federal del Océano Pacífico, incluyendo el Golfo de California. Diario Oficial de la Federación. México. 31 de diciembre de 1993
DOF. 2012. Acuerdo mediante el cual se da a conocer el Plan de Manejo Pesquero para la Pesquería de Pelágicos Menores (sardinas, anchovetas, macarela y afines) del Noroeste de México. Diario Oficial de la Federación. México. 8 de noviembre de 2012
Palabras clave: sardinas, pesca artesanal, Baja California Sur

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
275
134: oral ASENTAMIENTO DE POSTLARVAS DE LANGOSTA ROJA Panulirus interruptus Y SU RELACION CON FACTORES OCEANOGRAFICOS EN BAHIA ASUNCION, BAJA CALIFORNIA SUR.
Armando Vega Bolaños, Fernando López Salas, Armando Vega Velázquez, Gabriel Jiménez Llanos,
Edgardo Camacho Bareño y José Carlos Monroy Hernández
Centro Regional de Investigación Pesquera de La Paz. INAPESCA. Carretera a Pichilingue Km 1 s/n, Col. Esterito, La Paz Baja California Sur, México. C. P. 23020. [email protected] Introducción En la dinámica poblacional de las langostas espinosas, la abundancia de la población adulta sujeta a explotación depende del reclutamiento en las fases tempranas del ciclo de vida (larvas y postlarvas), el cual a su vez está fuertemente influenciado por cambios ambientales (Koslow et al. 2012). La importancia de este tipo de estudio se justifica por sus aplicaciones prácticas en pesquerías hacia pronóstico de capturas, una vez que se tengan series de más de 8-10 años, además de la caracterización de áreas de crianza, evaluación de los niveles del stock reproductor y la disponibilidad de “semilla” para semicultivo de langosta. Este conocimiento que ayudará a fortalecer las herramientas de evaluación y manejo del recurso. Se describe en este trabajo los resultados del asentamiento de postlarvas (puerulus) y juveniles de P. interruptus (Randall 1840) en la zona costera de Bahía Asunción, y su relación con algunos factores oceanográficos durante el período junio 2010 a junio 2015. Material y métodos El asentamiento fue monitoreado mediante colectores artificiales tipo Phillips (Phillips y Booth 1994), que son estructuras metálicas triangulares con tres placas de PVC que llevan mechones de filástica que simulan a los pastos marinos, colocadas en áreas costeras de crianza. Se determinó un índice mensual de asentamiento con base al número de puerulus por colector, como indicativo de la abundancia relativa. Paralelamente se registró la temperatura superficial del mar (TSM) in situ con termómetro de inmersión (°C), diariamente en el muelle de Bahía Asunción, BCS, estimándose el promedio mensual y a partir de éste se detectaron las anomalías térmicas. También se obtuvieron índices mensuales de surgencias (IS) y las anomalías del cuadrante 27° Norte y 116° Oeste en el Pacífico Nororiental de la base de datos del Pacific Fisheries Environmental Laboratory (*NOAA/NMFS/SWFSC). A partir de la información obtenida se describe el patrón mensual de asentamiento, observado bajo condiciones asociadas al evento océano-atmosférico La Niña 2010-11, relacionándolo con la TSM y el índice de surgencias. Resultados y discusión
Los mayores índices de asentamiento de postlarvas ocurrieron principalmente entre primavera-verano, seguido por un periodo menor en invierno (Fig. 1).
Fig.1. Índice de asentamiento por estación de 2010 a 2015. ANDEVA para las medias del IA. El pico del asentamiento coincide cuando se debilitan las surgencias durante el periodo más cálido del año (Fig. 2). Sin embargo, el pico de asentamiento varía entre años influenciado por los eventos ENSO. Por ejemplo, bajo condiciones de un enfriamiento anómalo durante 2011 el índice de asentamiento se mantuvo escaso, lo cual probablemente resultará en bajas capturas 5 años después (Arteaga-Ríos et al. 2007). Posteriormente, cuando las condiciones neutrales comienzan a hacerse presente en 2012-13, el asentamiento incrementó hacia valores promedio, mientras que en condiciones cálidas hacia la transición de El Niño 2014 se observaron índices por arriba del promedio en primavera (Fig. 2 y 3).

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
276
Fig. 2. Índice de asentamiento (IA) de P. interruptus (barras), TSM in situ Bahía Asunción, BCS (arriba) e índice de surgencias cuadrante 27°N (abajo), de 2010 a 2015.
Fig. 3. ANDEVA para la media anual del IA. Literatura citada Arteaga-Ríos LD, J Carrillo-Laguna, J Belmar-Pérez, S
Guzmán-del Próo. 2007. Postlarval settlement of California spiny lobster Panulirus interruptus in Bahía Tortugas, Baja California and its relationship to the commercial catch. Fish. Res. 88: 51-55
Koslow JA, L Rogers-Bennett, DJ Neilson. 2012. A time series California spiny lobster (Panulirus interruptus) phyllosoma from 1951 to 2008 links abundance to warm oceanographic conditions in southern California. CalCOFI Rep. 53: 132-139
Phillips BF, JD Booth. 1994. Design, use and effectiveness of collectors for catching the puerulus stage of spiny lobster. Rev. Fish. Sci. 2: 255-289.
*http://www.pfel.noaa.gov/products/PFEL/modeled/indices/upwelling/NA/data_download.html
Palabras clave: Langosta roja, Bahía Asunción, ENSO

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
277
135: oral MIGRACIÓN REPRODUCTIVA DEL CARACOL CHINO ROSA (Hexaplex erythrostomus) EN LA BAHÍA DE
BACOCHIBAMPO, COSTA CENTRO DE SONORA
Estefani Larios Castro, Leonardo Tolano Rodríguez y Raúl Ulloa Herrera
Centro Regional de Investigación Pesquera de Guaymas. INAPESCA. Calle 20 Sur No. 605, Col. La Cantera, Guaymas, Sonora. [email protected] Introducción La pesca como actividad del sector productivo primario requiere de la generación de conocimiento como soporte a la administración del uso de los recursos pesqueros, y como política general, se define como objetivo el uso sustentable de los mismos (Arreguín-Sánchez y Arcos-Huitrón 2011). Hexaplex erythrostomus conocido vermicularmente como caracol chino rosa se distribuye en el Golfo de California y algunas bahías y esteros de la costa occidental de Baja California Sur. Es de hábitos alimenticios carnívoros y se asocia a bancos de almejas, principalmente de almeja chocolata (Megapitaria squalida) (Arellano et al. 2006). Aunque la pesca de esta especie es por medio de trampas y en forma manual por medio de buceo semi autónomo, la pesquería ha tenido un auge en los últimos años en el noroeste de México, por lo que se requiere conocer el comportamiento del recurso en las costas del estado de Sonora. El presente estudio tiene como finalidad conocer la distribución y la variación del recurso caracol chino rosa en la Bahía de Bacochibampo y aportar información de la dinámica poblacional de este recurso. Material y métodos El área de estudio se encuentra en Bahía Bacochibampo parte central de la costa del estado de Sonora, en el municipio de Guaymas (Fig. 1). En el área se realizaron muestreos mensuales por medio de trampas cebadas tipo Chesapeake (60x60x25 cm) desde enero a diciembre del 2015, en la Bahía de Bacochibampo, con el fin de identificar el proceso de migración y agregación de este recurso pesquero. Además en agosto y septiembre la captura fue por medio de buceo semiautónomo, para determinar la dinámica poblacional y la distribución del caracol chino rosa en la zona.
Fig. 1. Ubicación del área de estudio. Resultados Se capturaron en total 4,854 organismos, los individuos presentaron una longitud mínima de 4.68 cm y un máximo de 14.5 cm de longitud de concha mientras que la amplitud de concha presentó un valor mínimo de 2.5 cm y un máximo de 10.2 cm, los valores promedio se situaron en 9.06 y 5.94 cm respectivamente. Los valores de longitud total a lo largo del año no presentan tendencia clara, como se observa en la figura 2a, sin embargo los valores de peso desconchado muestran una clara tendencia en decremento entre enero y diciembre (Fig. 2b). En la figura 3 se observa la variación espacial y temporal en la distribución del número de caracoles capturados, donde se puede evidenciar cómo el número de caracoles varía en forma estacional migrando hacia la desembocadura del Estero El Soldado. Sólo se presenta la distribución espacial de los organismos hasta el mes de septiembre, ya que el número de organismos capturados en los siguientes meses no fue significativo.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
278
a)
b)
Fig. 2. a) Longitud promedio y b) Peso desconchado promedio de caracol chino. Discusión y conclusión De acuerdo a lo observado, H. erythrostomus presenta agregaciones con el fin de proteger la masa ovígera en época reproductiva (mayo-agosto), periodo en el cual no se alimenta, al igual que el caracol chino negro (H. nigritus), según estudios realizados por Cudney-Bueno et al. (2008). Esta agregación reproductiva se da en la zona de la desembocadura del estero El Soldado, y se infiere que esto sea con el fin de que los futuros caracoles en su estado planctónico tengan alimento accesible hasta llegar a la etapa juvenil, como ha sido descrito para otros organismos planctónicos (Inda-Díaz 2013). Literatura citada Arellano MM, AM Quiñones, VBP Ceballos, FM Villalejo.
2006. Reproductive pattern of the squalid callista Megapitaria squalida from Northwestern Mexico. J. Shellfish Res. 25: 849-855.
Arreguín-Sánchez F, E Arcos-Huitrón. 2011. La pesca en México: estado de la explotación y uso de los ecosistemas. Hidrobiológica 21(3): 431-462.
Inda-Díaz EA, D Domínguez-Ojeda, CA Romero-Bañuelos, JT Nieto-Navarro, J Cepeda-Morales. 2013. El zooplancton marino de la costa de Nayarit, México. Revista fuente nueva época 4(14). Julio-Septiembre 2013. ISSN 2007-0713.
Fig. 3. Varición espacial y temporal de la distribución del caracol chino rosa.
Palabras clave: caracol chino, agregaciones, migración, distribución espacial.

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
279
136: cartel CARACTERIZACIÓN DE LA PESQUERÍA DE ESCAMA EN LA CUENCA MEDIA DEL RIO GRIJALVA,
CHIAPAS, MÉXICO
Eduardo Ramos Santiago1, Ma. Teresa Gaspar Dillanes2, Aldrin Labastida Che1, Yessica Xiomara Guzmán Camacho1, José Alfonso Oviedo Piamonte1 y Oswaldo Trejo Trejo2
Centro Regional de Investigación Pesquera de Salina Cruz. INAPESCA. Pról. Playa abierta s/n. Col. Miramar. Salina Cruz, Oaxaca. C.P. 70680. [email protected]. Oficina Centrales. INAPESCA. Pitágoras 1320. Col. Santa Cruz Atoyac. Del. Benito Juárez. Ciudad de México. C.P. 03310. Introducción El estado de Chiapas se caracteriza por poseer un extenso potencial hidrológico, aprovechado por la construcción de diversas presas, para la generación de energía eléctrica y prevención de inundaciones e irrigación de diversos cultivos (CFE 1976, Pérez-Castañeda 2012). Las presas poseen organismos acuáticos de gran valor nutricional que aportan una pesca comercial local. El presente análisis es para evaluar en términos biológico-pesquero, ambiental y social, la actividad pesquera en la cuenca media del río Grijalva, aguas abajo de la Presa Chicoasén (Fig. 1). Ello permitió conocer la problemática del sector asociada a la explotación de recursos y caracterizar la pesca de escama con base en la conjunción de factores y determinar puntos de referencia que conlleven a la adecuada administración de los recursos pesqueros.
Fig. 1. Área de estudio. Cuenca media río Grijalva, Chiapas. Material y métodos Se realizaron muestreos mensuales provenientes de la captura comercial de febrero a diciembre del 2014. Las especies se determinaron taxonómicamente utilizando literatura especializada. Se elaboraron fichas, que incluyen algunos aspectos biológicos para las especies de importancia comercial. Mediante censos, se describieron las especificaciones técnicas de cada uno de los artes identificados y por medio de
entrevistas se obtuvo información sobre los principales aspectos sociales, económicos y pesqueros de la población. Resultados Se analizaron 1,464 organismos de las cooperativas “Pescadores La Cuevita” y El Indio Copainalteco”. Se determinaron 14 especies de peces, agrupadas en 13 géneros y ocho familias. La familia Cichlidae fue la mejor representada con siete especies; el resto cuentan con un solo representante (Fig. 2).
Fig. 2. Número de especie por grupo taxonómico. Siete especies fueron determinadas como importantes en términos comerciales, tres son de autoconsumo y el resto sin importancia (Tabla 1). Tabla 1. Especies de importancia comercial
Nombre Cientifico Nombre común
Longitud (cm) Peso (g) Origen
Petenia splendida tenhuayaca 21-34 120-590 Nativa Oreochromis niloticus tilapia 21-39.5 175-1,121 Exótica Paraneetroplus hartewgi mojarra negra 13.5-32.5 42-695 Nativa Cichlasoma pearsei zacatera 17-29 88-483 Nativa Brycon guatemalensis Macabil 22-60.5 98-2,934 Nativa Aplodinotus grunniens mojarra blanca 27.45 210-1,354 Naiva Ictalurus meridionalis Bagre 32-77 264-6,120 Nativa
Fig. 3. Abundancia de las especies de importancia comercial.
0 2 4 6 8
CLUPEIDAE
CHARACIDAE
ICTALURIDAE
HEPTAPTERIDAE
BELONIDAE
GERREIDAE
SCIAENIDAE
CICHLIDAE
50%
31%
5%
7% 4% Negra
Macabil
Zacatera
Tilapia

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
280
Abundancia específica: La mojarra negra (Paraneetroplus hartwegi) fue la más abundante, representa 50% de la captura, la segunda especie en importancia fue el macabil (Brycon guatemalensis) con 31%, seguida por tilapia (Oreochromis niloticus) con 7%, la zacatera (Cichlasoma pearsei) 5% y el bagre o bobo (Ictalurus meridionalis) constituye 4%. El resto de la captura (3%) engloba a la tenguayaca (Petenia splendida), sabalote (Dorosoma anale) y mojarra blanca (Aplodinotus grunniens) (Fig. 3). Producción Pesquera: Los registros de producción pesquera del 2014 indican que en promedio, los meses de mayor producción pesquera fueron enero, abril-mayo y agosto-septiembre, con capturas mínimas en febrero-marzo y junio-julio (Fig. 4).
Fig. 4. Producción pesquera mensual 2014.
Artes y equipos de pesca. La unidad de pesca consta de embarcaciones menores tipo cayuco de fibra de vidrio entre 3.70 y 7.0 m de eslora, con motor fuera de borda entre 2 y 15 HP, las que no cuentan con motor utilizan remos de madera. En la captura se emplean: 1) red de enmalle de longitud variable entre 50 y 120 m, altura entre 3 y 5 m y luz de malla de 76.2 mm hasta 119.3 mm. El tipo de hilo de la malla es de monofilamento, calibre 0.20 y 0.25; 2) atarrayas de monofiliamento entre 2.5 y 5 m de longitud y luz de malla de 76.2 mm hasta 101.6 mm; 3) palangre de cabo de polipropileno, entre 60 y 130 m de largo y de 14 a 30 anzuelos tipo "J" del número 7 y 8. Generalmente participan en la faena dos pescadores por panga, los cuales incluyen mujeres esposas de pescadores. Aspectos demográficos. Los indicadores muestran un bajo nivel de escolaridad en la población, y rezagos en los servicios médicos. La falta de acceso a servicios, las condiciones inadecuadas de vivienda, los bajos ingresos, entre otros, son factores que se reflejan en un alto grado de marginación. La localidad de Chicoasén presentó un grado de marginación medio durante el 2005, y en 2010 se clasifica como alto. Todas las demás localidades analizadas no presentan cambios en cuanto al índice de marginación, pero destacan Tres Picos y Nuevo Vergel (municipio de Copainalá) con un índice de marginación muy alto.
Aspectos Socioeconómicos. Se hicieron 51 encuestas, 48 a los pescadores y 3 a esposas de éstos que acompañan a realizar la actividad pesquera. De los pescadores de “La Cuevita”, Chicoasén, 30% tiene primaria y 34% secundaria. Lo anterior contrasta con los pescadores de la cooperativa “El Indio Copainalteco”, ya que 40% manifestó no haber asistido a la escuela y 60% sabe leer y escribir. La pesca se realiza todo el año, las especies capturadas son nativas, excepto la tilapia, se comercializan directamente, ya sea al consumidor, o en los restaurantes y mercados locales, obteniendo buenas ganancias. Todos los pescadores cuentan con embarcaciones y artes de pesca. Discusión La composición de especies está asociada con aspectos biológicos y ecológicos, ya que su distrubucion y abundancia está directamente relacionada con la dinamica del río, clasificado como un sistema lótico, influenciado por el cierre y apertura de las turbinas de desfogue de la presa Chicoasén. La pesca es la principal actividad de donde los pescadores obtienen sus ingresos por lo que puede considerarse una actividad productiva, además de tener una tradición de más de diez años. Los pescadores de la SC La Cuevita obtienen mayores ingresos derivados de la pesca, ya que los comercializan a mejores precios. Los pescadores de la SC El Indio Copainalteco tienen mayores dificultades para realizar la actividad, desde la transportación hasta la comercialización de su producto. Conclusión Es una pesquería muy importante para la economía del sector local y principal fuente de alimento en la región; sin embargo es necesario que se realicen estudios para determinar las épocas de reproducción de las especies. La SC Cuevita comercializa mejor su producto en comparacion con la SC El Indio Copainalteco, el cual muestra niveles muy altos de marginación (eduación, servicios médicos y vivienda). La comercialización es fresco-eviscerado en mercados y restaurantes locales. Literatura citada Comisión Federal de Electricidad. 1976. México construye:
Proyecto hidroeléctrico Chicoasén y la Angostura. México.
Pérez-Castañeda JW. 2012. Composición de peces en la pesquería de la Presa Hidroeléctrica Nezahualcóyotl, Chiapas. Tesis de Licenciatura. Facultad de Ciencias Biológicas. Universidad de Ciencias y Artes de Chiapas. 69p.
Palabras clave: río Grijalva, Chicoasén, Copainalá Chiapas, Cichlidae, especies nativas.
0
5000
10000
ENE MAR MAY JUL SEP NOV
Cap
tura
(kg
)
Promedio Maximo Minimo

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
281
137: oral CRECIMIENTO DE POSTLARVAS DE LAS LANGOSTAS Panulirus inflatus Y P. gracilis EN CAJAS
OSTRÍCOLAS EN SINALOA, MÉXICO
Jesús Audomar Landeros Armenta, Raúl Pérez González, Guillermo Rodríguez Domínguez, Martín I. Borrego y Luis M. Valadez Manzano
Universidad Autónoma de Sinaloa. Paseo Claussen S/N, Col. Centro, Mazatlán, Sinaloa. C.P. 82000. [email protected] Introducción Las langostas espinosas de la familia Palinuridae son un recurso muy importante dentro de los crustáceos, ya que presentan un gran valor económico en el mercado nacional e internacional y su pesquería es una actividad muy importante. Panulirus inflatus y P. gracilis contribuyen anualmente a la producción nacional anual con un promedio de 153 t (CONAPESCA 2011). En los últimos años se ha incrementado el interés por la acuicultura de langostas espinosas, como consecuencia de las mínimas capturas y además por la continua demanda de este recurso pesquero. Desde 1995 esta actividad de cultivo solamente se ha realizado en Vietnam con P. ornatus (Jones 2010), con resultados satisfactorios. Material y métodos Se capturaron postlarvas de langosta en la bahía de Mazatlán (23º12’32.4” N-106º25’33.2” O) y en el estero Cospita, Sinaloa (24°05’54.77” N-107°09’12.41” O), utilizando colectores tipo Sandwich (Montgomery y Craig 1997) para introducirlas en módulos de crecimiento (cajas ostrícolas). Se realizaron mediciones de la longitud del cefalotórax (LC) y del peso (PT). Los muestreos fueron mensuales de febrero a septiembre 2015 en el estero Cospita y de junio 2015 a febrero 2016 en la bahía de Mazatlán. Se utilizó el modelo de Schnute (1981) modificado por Montgomery et al. (2010) para aplicarlo al seguimiento de longitudes medias por intervalos de tiempo y los dos tipos de errores aditivo y multiplicativo, así como el índice de Akaike (1981) para seleccionar el mejor modelo. Además se realizaron relaciones entre LC y PT para determinar el tipo de crecimiento que presentan las postlarvas. Para conocer el tipo de crecimiento relativo, se utilizó la prueba t-Student con un nivel de confianza de 95%. Resultados En la bahía de Mazatlán fue seleccionado el caso 1 del modelo de Schnute con error aditivo y se obtuvieron valores para K = 5.7 y una ɛ = 24.9 mm para P. inflatus. En la curva de crecimiento se observa un incremento de 9.5 (0.8 g) a 24.1 mm de LC (12 g) durante 9 meses (Fig. 1).
Fig. 1. Curva de crecimiento usando los parámetros calculados del caso 1 del modelo Schnute (1981), para las postlarvas de la langosta Panulirus inflatus en la bahía de Mazatlán, Sinaloa. En el estero Cospita el modelo que mejor se ajustó fue el caso 2 para las postlarvas de P. inflatus y P. gracilis, con valores de K = 3.4 y ɛ = 44.6 mm y K= 3.8 y ɛ = 44.2 mm, respectivamente (Fig. 2).
Fig. 2. Curva de crecimiento usando los parámetros calculados del caso 2 del modelo Schnute (1981), para las postlarvas de las langostas Panulirus inflatus y P. graclis en el estero Cospita, Sinaloa.
0
5
10
15
20
25
30
1 1.5 2 2.5
Lo
ng
itu
d d
e ce
falo
tóra
x
(mm
)
Edad (años)
20
25
30
35
40
45
1.5 2 2.5 3
Lo
ng
itu
d d
el
cefa
lotó
rax (
mm
)
Edad (años)
5
15
25
35
45
1.2 1.4 1.6 1.8 2 2.2
Lo
ng
itu
d d
el
cefa
lotó
rax (
mm
)
Edad (años)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
282
Fig. 3. Relación entre la longitud del cefalotórax y el peso total de las postlarvas de las langostas Panulirus inflatus en la bahía de Mazatlán, Sinaloa.
a)
b) Fig. 4. Relación entre la longitud del cefalotórax y el peso total de las postlarvas de las langostas a) Panulirus inflatus y b) P. gracilis en el estero Cospita, Sinaloa. Discusión Los datos analizados en la bahía de Mazatlán y en el estero Cospita muestran que las postlarvas de P. inflatus y P. gracilis presentan un crecimiento acelerado en esta etapa de su ciclo de vida: K = 5.7 (Mazatlán) y K = 3. 4 para P. inflatus y K =3.8 para P. gracilis en el estero Cospita. Estos resultados son similares a los descritos por García-Paredes (2013) con K = 5.16 (para el caso 5 del modelo de Schnute) e indica que probablemente este elevado parámetro podría deberse a que en esta etapa de vida las postlarvas tienen un crecimiento rápido con respecto a la de adulto, que de acuerdo a Flores-Reyes (2003) y Pérez-González (2008) es de 0.30 a 0.38. Las postlarvas presentaron un crecimiento de tipo isométrico, crecen proporcionalmente en sus dimensiones. Rodríguez-Negrete (2006) obtuvo un crecimiento isométrico en juveniles de P. inflatus y P. gracilis; sin embargo, no lo demuestra
estadísticamente, y Arzola-González (2001) en adultos. Literatura citada Akaike H. 1981. Likelihood of a model and information
criteria. Journal of Econometrics 16: 3-14. Arzola-González JF. 2001. Biometría y crecimiento de las
langostas Panulirus inflatus (Bouvier 1895) y P. gracilis Streets 1871, en el sur de Sinaloa. Tesis de Maestría, Universidad Autónoma de Sinaloa. 72 p.
CONAPESCA 2011. Anuario estadístico de acuacultura y pesca 2011. Comisión Nacional de Acuacultura y Pesca CONAPESCA, SAGARPA, México. 305p.
Jones CM. 2010. Tropical spiny lobster aquaculture development in Vietnam, Indonesia and Australia. J. Mar. Biol. Ass. India 52(2): 304–315.
Flores-Reyes A. 2003. Análisis de la estructura poblacional y el crecimiento de las langosta Panulirus inflatus (Bouvier, 1895) y Panulirus gracilis Streets 1871, en las costas de Nayarit. Tesis de Licenciatura, Universidad Autónoma de Sinaloa. 60 p.
García-Paredes JR. 2013. Densidad y estructura de tallas de los juveniles de la langosta Panulirus inflatus en la bahía de Mazatlán, Sinaloa. Tesis de Maestría, Facultad de Ciencias del Mar. Universidad Autónoma de Sinaloa. 79 p.
Montgomery S, C Walsh, M Haddon, C Kesby, D Johnson. 2010. Using length data in the Schnute Model to describe growth in a metapenaeid from water off Australia. Marine and Freshwater Research 61: 1435-1445.
Montgomery SS, JR Craig. 1997. A strategy for measuring the relative abundance of pueruli of spiny lobster J. verreauxi. Pp. 574-578. En: Hancock DA, DC Smith. Fisheries Resources. The state of science and Management. 2nd World Fisheries Congress Proceedings. Collingwood, Australia CSIRO Publishing.
Pérez-González R. 2008. Biología y pesquería de la langosta Panulirus gracilis Street 1871 en el sur de Sinaloa, México. Tesis Doctoral. Facultad de Ciencias, Universidad Nacional Autónoma de México. 180 p.
Rodríguez-Negrete RE. 2006. Densidad y descripción del hábitat de las langostas Panulirus inflatus (Bouvier 1895) P. gracilis Streets 1871, en la bahía de Mazatlán, Sinaloa. Tesis de Licenciatura, Facultad de Ciencias del Mar. Universidad Autónoma de Sinaloa. 78 p.
Schnute J. 1981. A versatile growth model with statistically stable parameters. Canadian Journal of Fisheries and Aquatic Science 38: 1128-1140.
Palabras clave: Crecimiento, postlarvas, Panulirus
spp., cautiverio, Sinaloa.
PT = 0.0006 LC3.142
r² = 0.993
0
5
10
15
20
0 5 10 15 20 25 30
Pes
o (
g)
Longitud del cefalotórax (mm)
PT = 0.0007LC3.0466
r² = 0.9657
0
20
40
60
80
100
0 10 20 30 40 50
Pes
o (
g)
Longitud del Cefalotórax (mm)
PT = 0.0007LC3.0368
r² = 0.9975
0
10
20
30
40
50
60
70
0 10 20 30 40 50
Pes
o (g
)
Longitud del cefalotórax (mm)
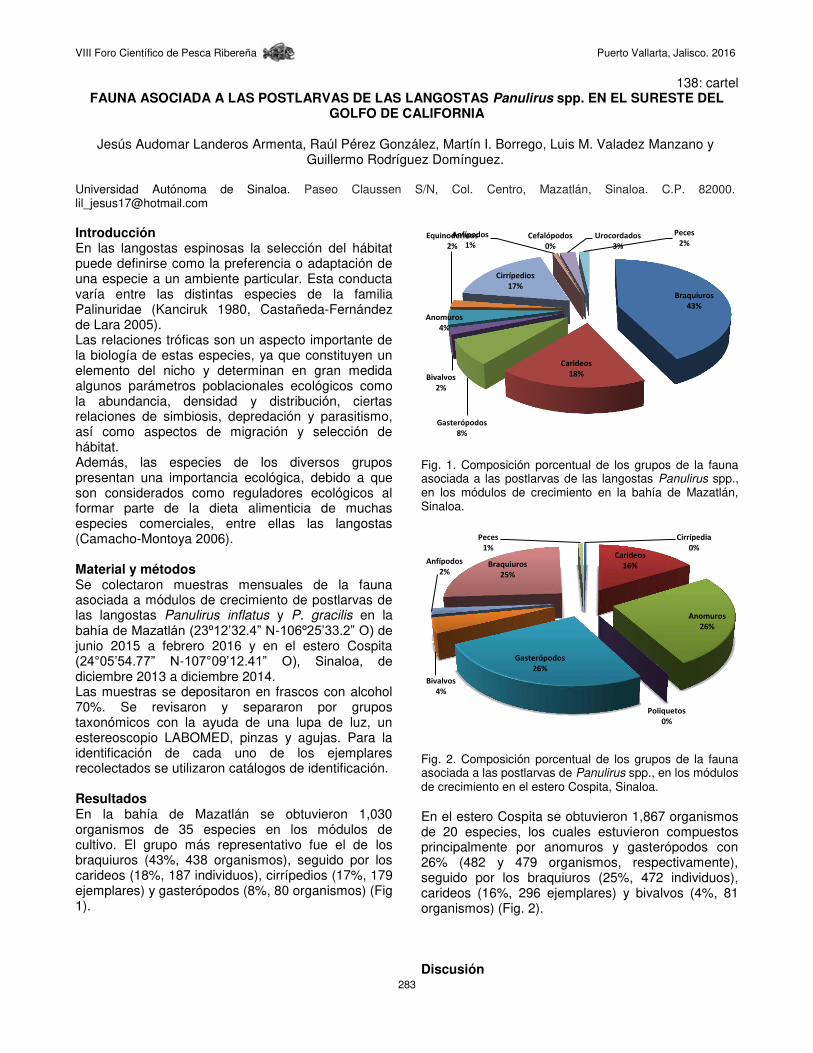
VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
283
138: cartel FAUNA ASOCIADA A LAS POSTLARVAS DE LAS LANGOSTAS Panulirus spp. EN EL SURESTE DEL
GOLFO DE CALIFORNIA
Jesús Audomar Landeros Armenta, Raúl Pérez González, Martín I. Borrego, Luis M. Valadez Manzano y Guillermo Rodríguez Domínguez.
Universidad Autónoma de Sinaloa. Paseo Claussen S/N, Col. Centro, Mazatlán, Sinaloa. C.P. 82000. [email protected] Introducción En las langostas espinosas la selección del hábitat puede definirse como la preferencia o adaptación de una especie a un ambiente particular. Esta conducta varía entre las distintas especies de la familia Palinuridae (Kanciruk 1980, Castañeda-Fernández de Lara 2005). Las relaciones tróficas son un aspecto importante de la biología de estas especies, ya que constituyen un elemento del nicho y determinan en gran medida algunos parámetros poblacionales ecológicos como la abundancia, densidad y distribución, ciertas relaciones de simbiosis, depredación y parasitismo, así como aspectos de migración y selección de hábitat. Además, las especies de los diversos grupos presentan una importancia ecológica, debido a que son considerados como reguladores ecológicos al formar parte de la dieta alimenticia de muchas especies comerciales, entre ellas las langostas (Camacho-Montoya 2006). Material y métodos Se colectaron muestras mensuales de la fauna asociada a módulos de crecimiento de postlarvas de las langostas Panulirus inflatus y P. gracilis en la bahía de Mazatlán (23º12’32.4” N-106º25’33.2” O) de junio 2015 a febrero 2016 y en el estero Cospita (24°05’54.77” N-107°09’12.41” O), Sinaloa, de diciembre 2013 a diciembre 2014. Las muestras se depositaron en frascos con alcohol 70%. Se revisaron y separaron por grupos taxonómicos con la ayuda de una lupa de luz, un estereoscopio LABOMED, pinzas y agujas. Para la identificación de cada uno de los ejemplares recolectados se utilizaron catálogos de identificación. Resultados En la bahía de Mazatlán se obtuvieron 1,030 organismos de 35 especies en los módulos de cultivo. El grupo más representativo fue el de los braquiuros (43%, 438 organismos), seguido por los carideos (18%, 187 individuos), cirrípedios (17%, 179 ejemplares) y gasterópodos (8%, 80 organismos) (Fig 1).
Fig. 1. Composición porcentual de los grupos de la fauna asociada a las postlarvas de las langostas Panulirus spp., en los módulos de crecimiento en la bahía de Mazatlán, Sinaloa.
Fig. 2. Composición porcentual de los grupos de la fauna asociada a las postlarvas de Panulirus spp., en los módulos de crecimiento en el estero Cospita, Sinaloa. En el estero Cospita se obtuvieron 1,867 organismos de 20 especies, los cuales estuvieron compuestos principalmente por anomuros y gasterópodos con 26% (482 y 479 organismos, respectivamente), seguido por los braquiuros (25%, 472 individuos), carideos (16%, 296 ejemplares) y bivalvos (4%, 81 organismos) (Fig. 2). Discusión
Braquiuros
43%
Carideos
18%
Gasterópodos
8%
Bivalvos
2%
Anomuros
4%
Equinodermos
2%
Cirrípedios
17%
Anfípodos
1% Cefalópodos
0%
Urocordados
3%
Peces
2%
Cirrípedia
0% Carideos
16%
Anomuros
26%
Poliquetos
0%
Gasterópodos
26%
Bivalvos
4%
Anfípodos
2% Braquiuros
25%
Peces
1%

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
284
En las dos zonas de muestreo los grupos más representativos fueron los crustáceos (braquiuros principalmente) y los moluscos (gasterópodos y bivalvos), los cuales se ha demostrado mediante análisis de contenido estomacal que forman parte de la dieta principal de las postlarvas y adultos de la langosta (Lopeztegui-Castillo y Capetillo-Piñar 2006, Castañeda-Fernández et al. 2005). Por otra parte, Lopeztegui-Castillo et al. (2015) trabajaron con fauna asociada a postlarvas de P. argus y obtuvieron un total de 10 grupos zoológicos, siendo Brachyura y Osteichthyes los más representativos. Las investigaciones realizadas sobre alimentación de langostas indican que en general los crustáceos son un grupo primordial en la alimentación de estos organismos (Lalana y Ortiz 1991, Lopeztegui-Castillo y Capetillo-Piñar 2006). Literatura citada Camacho-Montoya C. 2006. Alimentación de juveniles de
las langostas Panulirus inflatus (Bouvier, 1895) y P. gracilis Streets 1871 y su relación con el bentos en la bahía de Mazatlán, Sinaloa. Tesis de Licenciatura. Facultad de Ciencias del Mar, Universidad Autónoma de Sinaloa. 154 p.
Castañeda-Fernández de Lara V. 2005. Caracterización del hábitat de juveniles de la langosta roja Panulirus interruptus (Randall 1839) en dos sitios ubicados en su zona de mayor abundancia en Baja California Sur, México. Tesis de Doctorado. Centro de Investigaciones Biológicas del Noroeste, S. C. 131 p.
Kanciruk P. 1980. Ecology of juvenile and adult Palinuridae (spiny lobsters). En: Cobb SJ, BF Phillips (eds.). The biology and management of lobsters. Academic Press, Inc. New York, USA. 59-92.
Lalana R, M Ortiz, 1991. Contenido estomacal de puérulos y post-puérulos de la langosta P. argus en el Archipiélago de Los Canarreos, Cuba. Revista de Investigaciones Marinas 12(1-3): 107-116.
Lopeztegui-Castillo A, R Cuellar-Peña, U Amador-Marrero. 2015. Fauna asociada a colectores artificiales de puerulos de langosta Panulirus argus (Decapoda: Palinuridae) en Cuba. Boletín del Centro de Investigaciones Biológicas 49(1): 55–72.
Lopeztegui-Castillo A, N Capetillo-Piñar. 2006. Alimentación natural de los estadios postlarvales de la langosta Panulirus argus en la zona sur de cayo Matías, Archipiélago de Los Canarreos, Cuba. Revista de Investigaciones Marinas 27 (2): 153-158.
Palabras clave: Fauna, postlarvas, Panulirus spp.,
Sinaloa

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
285
INDICE DE AUTORES
Autores - núm. de página (núm. de trabajo) A Acosta Quintana JD 75 (34) Águila y Elvira AJ 121 (57) Aguilar Betancourt C 123 (58) Aguilar Ibarra A 57 (25) Aguilar Palomino B 129 (61) Aguilar Perera A 185 (89) Aguirre Villaseñor H 15(4), 207(100), 209 (101),
211 (102), 215 (104) Alatorre Alba AJ 189 (91), 221 (107),
251 (122), 253 (123), 255 (124)
Alfaro Gómez K 133 (63) Álvarez J 37 (15) Aragón Noriega EA 31 (12), 41 (17), 241 (117) Arellano Torres A 133 (63), 163 (78) Arredondo Vargas E 159 (76), 161 (77) Arroyo Salazar SA 125 (59) Avendaño CJ 159 (76), 161 (77) Aviña Romero MJ 73 (33) B Balan Che LI 43 (18), 45 (19) Barrera Melo FN 105 (49) Bartoleño Sánchez A 69 (31), 71 (32) Bolaño Martínez N 193 (93) Bonfil R 1 (magistral), 193 (93) Borrego MI 281 (137), 283 (138) Braccini JM 59 (26) Bravo Olivas ML 151 (72), 153 (73) Briones Hernández SA 129 (61) C Caballero Chávez V 115 (54), 235 (114),
237 (115) Cabrera Mancilla E 117 (55), 119 (56) Camacho Bareño E 265 (129), 275 (134) Can González MJ 121 (57) Carbajal Arana VR 169 (81) Carrasco Águila MA 69 (31), 71 (32), 177 (85) Carrillo Colín LD 47 (20) Carrillo Nolasco I 145 (69)
Castañeda Fernández de Lara V 269 (131), 273 (133)
Castañeda Lomas N 31 (12), 89 (41) Castillo Geniz JL 113 (53), 171 (82), 225 (109),
227 (110), 245 (119) Castro Moreno PN 173 (83) Castro Salgado JC 269 (131) Cavieses Núñez RA 83 (38) Celaya Castillo C 221 (107) Cerdenares Ladrón de Guevara G 95 (44),
251 (122) Cervantes Gutiérrez F 183 (88) Cervera Cervera K 85 (39), 137 (65) Cervera Paul ED 137 (65) Chan Cab A 131 (62) Chávez Arrenquín DA 93 (43), 97 (45), 147 (70) Chávez Dagostino RM 151 (72), 153 (73) Chávez EA 9 (1) Chávez Herrera D 97 (45), 99 (46), 259 (126) Cob Pech EF 85 (39), 179 (86) Concha Toro FJ 5 (magistral) Cruz Fonseca EO 13 (3) Cruz Sánchez JL 145 (69), 179 (86) Cuervo López L 213 (103) Cupido Castellanos K 131 (62) D Danemann GD 173 (83) De Anda Fuentes DE 137 (65) De la Cruz González FJ 101 (47), 187 (90) Denton Castillo J 127 (60) Díaz Álvarez AG 115 (54), 233 (113), 235 (114),
237 (115) Díaz Quijano R 85 (39) Díaz Torres E 125 (59) E Enciso Enciso C 201(97), 271(132) Escalona Maurice MJ 57 (25) Escobar Sánchez O 109 (51) Esparza Leal HM 49 (21) Espino Barr E 15 (4), 53 (23), 63 (28),
197 (95), 205 (99)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
286
Espinosa Romero MJ 37 (15), 39 (16), 61 (27), 81 (37)
Espinoza Méndez JC 43 (18), 85 (39), 137 (65), 179 (86)
Espinoza Rodríguez IJ 79 (36) Estrada Navarrete FD 163 (78) F Fernández Méndez JI 203(98), 229(111) Fernández Ordóñez YM 57 (25) Fernández Rivera Melo F 29 (11), 81 (37), 61 (27) Flores Irigollen A 83 (38) Flores Ramírez LA 91 (42) Franco Marrujo LP 89 (41) Frisch Jordán A 79 (36) Fuentes Mata P 229 (111) Fuerte Velázquez Y 159 (76), 161 (77) Fulton S 39 (16), 81 (37) G Galeana Cortazar AR 233 (113) Galindo Cortez G 271 (132) Gallardo Cabello M 205 (99) Galván Magaña F 25 (9), 77 (35), 105 (49),
107(50), 113 (53), 129(61), 175(84), 183(88)
Galván Piña VH 129 (61) Gamboa Rivera H 91 (42), 205 (99) Garcés García KC 93 (43) García Boa A 15 (4), 63 (28), 205 (99) García Solorio L 243 (118) García Urquídez D 49 (21) Garduño Dionate M 11 (2) Gaspar Dillanes MT 223 (108), 279 (136) Gastelum E 81 (37) Godínez Padilla CJ 171 (82), 225 (109),
227 (110) Goldman N 61 (27), 81 (37) Gómez A 81 (37) Gómez García MJ 123 (58) Gómez Gauna CG 157 (75) Gómez Gómez A 29 (11) Gómez Medina LG 259 (126) Gómez Ortiz MG 257 (125) Gómez Rojo C 273 (133), 269 (131) Gonzáles Armas R 105 (49) González Ania LV 245 (119)
González Cárdenas JJ 133 (63) González Cedano J 153 (73) González López WA 121 (57) González Medina G 259 (126) González Ocaranza L 165 (79), 261 (127) González Peláez SS 267 (130) González Romero S 269 (131) González Sánchez AC 195 (94) González Sansón G 123 (58), 169 (81) Gracia Gasca A 255 (124) Gracia Olea JA 157 (75) Guevara Cruz C 55 (24), 131 (62) Guevara Rascado ML 101 (47) Guillén Gamboa JA 207 (100), 207 (100),
215 (104), Gutiérrez González JL 111 (52) Gutiérrez Pérez C 33 (14) Gutiérrez Zavala RM 117 (55), 119 (56) Guzmán Camacho YX 223 (108), 279 (136) Guzmán Castellanos AB 189 (91), 221 (107),
251 (122), 253 (123) H Hau Pacheco DA 179 (86) Hernández A 105 (49) Hernández Corona JL 103 (48), 253 (123) Hernández Covarrubias V 101 (47), 163 (78),
207 (100), 209 (101), 215 (104)
Hernández Herrera A 113 (53) Hernández Juárez M 57 (25) Hernández Montaño D 163 (78) Hernández Morales PD 175 (84) Hernández Ventura S 67 (30) Herrera Gómez DG 257 (125) Hoyos Padilla M 113 (53) Hudson Weaver A 143 (68), 167 (80) Huidobro Campos L 191 (92) J Jiménez Llanos GA 263 (128), 265 (129),
275 (134) Juárez Villalobos ML 243 (118) K Keyes Pulido S 185 (89)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
287
L Labastida Che A 223 (108), 247 (120),
249 (121), 279 (136) Landeros Armenta JA 281 (137), 283 (138) Lara Mendoza RE 47 (20), 233 (113) Larios Castro E 277 (135) Lejbowicz A 29 (11) Lizárraga Rodríguez AA 189 (91), 221 (107),
251 (122), 253 (123), 255 (124)
Lluch Cota DB 267 (130) López Álvarez KY 153 (73) López López LE 147 (70) López Navarrete H 257 (125) López Salas F 275 (134) López Terán C 91 (42) López Uriarte E 197 (95) Lorán Núñez RM 11 (2), 53 (23) Lucano Ramírez G 123 (58), 169 (81) Luna Raya MC 101 (47), 187 (90) Lupio Rodríguez AM 67 (30) Luquín Covarrubias MA 267 (130) M Macías Zamora R 69 (31), 71 (32) Madrid Vera J 31 (12) Magadán Revelo LD 57 (25) Maldonado Amparo MA 35 (14), 239 (116) Maldonado Coyac JA 109 (51) Maldonado Gasca A 195 (94) Maldonado Ibarra OA 151 (72) Mancha MM 37 (15) Márquez Farías JF 47 (20), 87 (40), 213 (103) Marrón Espitia JM 143 (68) Martínez Cruz LE 43 (18), 45 (19), 165 (79), 217 (105), 261 (127) Martínez Isunza FR 11 (2) Martínez Muñoz MA 253 (123), 255 (124) Martínez Trujillo J 219 (106) Mata Martínez D 135 (64) Medina H 231 (112) Medina Quijano HA 145 (69) Meléndez Galicia C 133 (63), 163 (78) Melo Barrera FN 183 (88) Mendivil Mendoza JE 241 (117) Mendoza Vargas OU 193 (93) Meraz Sánchez R 207 (100) Meza Tabares SO 187 (90)
Mondragón Sánchez LF 59 (26), 73 (33), 75 (34) Monroy C 231 (112) Monroy Hernández JC 263 (128), 265 (129),
275 (134) Montoya Carvantes I 229 (111) Morales Bojórquez E 15 (4), 15 (4) Morales García G 229 (111) Morales Martínez RG 149 (71) Morales Pacheco O 189 (91), 221 (107), 251 (122),
253 (123), 255 (124) Morales Parra LG 43 (18), 45 (19) Morán Angulo RE 239 (116) Moreno Herrero A 61 (27), 81 (37) Moreno Miranda C 115 (54), 235 (114) Muñoz Rubí HA 97 (45), 99 (46) N Núñez Díaz A 27(10) Núñez Márquez G 13 (3) Núñez Orozco AL 247 (120), 249 (121) O Ochoa Báez RI 25 (9), 77 (35) Oddone MC 25 (9) Ojeda Ruiz de la Peña MA 83 (38) Ortega Lizárraga GG 31 (12), 41 (17) Ortega Ortiz CD 125 (59) Ortega Salgado I 171 (82), 225 (109),
227 (110) Ortiz Gallarza SM 201 (97) Osuna Paredes C 159 (76), 161 (77) Osuna Peralta YR 207 (100), 209 (101),
211 (102), 215 (104), 259 (126)
Oviedo Perez JL 43 (18), 45 (19), 141 (67), 165 (79), 217 (105), 261 (127)
Oviedo Piamonte JA 223 (108), 247 (120), 249 (121), 279 (136)
P Páez Osuna FN 87 (40) Palacios Barreto PY 193 (93) Palma Barrón E 21 (7) Pantoja González LM 13 (3)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
288
Paredes Mellado R 207 (100), 209 (101), 215 (104) Parra Alaniz JC 181 (87) Patiño Valencia JL 101 (47), 187 (90) Pérez González R 31 (12), 281 (137),
283 (138) Pérez Juárez A 159 (76), 161 (77) Pérez Olivares I 17 (5), 19 (6), 21 (7),
23 (8) Perez Toledo A 169 (81) Pérez Velázquez PA 11 (2), 187 (90) Plomozo Lugo T 167 (80) Puente Gómez M 15 (4), 63 (28), 205 (99) R Ramírez Montes De Oca S 177 (85) Ramírez Murillo R 255 (124) Ramírez Pérez JS 35 (14), 109 (51), 127 (60),
199 (96), 239 (116) Ramírez Rodríguez M 93 (43), 181 (87) Ramírez Soberón G 229 (111) Ramírez Zavala JR 65 (29) Ramos Carrillo E 91 (42), 119 (56) Ramos Carrillo S 95 (44) Ramos Santiago E 223 (108), 247 (120),
249 (121), 279 (136) Rangel Arteaga BY 257 (125) Rentería Bravo J 103 (48) Reyes Bonilla H 185 (89) Reyna González PC 141 (67), 165 (79),
217 (105) Reyna Matezans VA 213 (103) Ricaño Soriano M 193 (93) Ríos González KG 197 (95) Ríos Lara GV 137 (65) Rodríguez Domínguez G 31 (12), 41 (17),
155 (74), 281 (137), 283 (138)
Rodríguez Franco IL 49 (21) Rodríguez Madrigal JA 51 (22) Rodríguez Preciado JA 67 (30) Rodríguez Quiroz G 49 (21) Rodríguez Tirado A 89 (41) Rodríguez Van Dyck S 143 (68) Rojas González RI 145 (69) Rojo M 81 (37) Romero Leyva TG 201 (97) Ruiz Ramírez S 123 (58), 169 (81)
S Salas Maldonado M 69 (31), 71 (32), 177 (85) Salas S 55 (24) Salcido Guevara LA 27(10), 35 (14), 127 (60),
155 (74), 199 (96), 239 (116)
Salgado Ugarte IH 17 (5), 19 (6), 21 (7), 23 (8), 135 (64)
Sánchez Cárdenas R 27(10), 35 (14), 87 (40), 111 (52), 127 (60), 155 (74), 199 (96), 239 (116)
Sánchez Cota JA 107 (50) Sánchez Cota JB 105 (49) Sánchez González A 113 (53) Sánchez Reyes NA 173 (83) Sánchez Romero S 273 (133) Santana LD 145 (69) Santana Hernández H 51 (22), 73 (33) Santos Guzmán S 65 (29) Santos Valencia J 121 (57), 149 (71), 179 (86) Sarmiento Náfate S 255 (124) Simental Anguiano MR 107 (50) Soberano Palomares IM 155 (74) Soto López K 25 (9), 77 (35) Stewart J 195 (94) Suárez Castillo A 37 (15), 39 (16) T Tamburin E 113 (53) Tolano Rodríguez L 277 (135) Toro Ramírez A 145 (69) Torre J 37 (15), 81 (37) Torres Blanco M 237 (115) Tovar Ávila J 51 (22), 59 (26), 73 (33),
75 (34), 77 (35), 93 (43), 101 (47), 103 (48), 105 (49), 147 (70), 175 (84), 183 (88), 195 (94)
Tovar Juárez E 175 (84) Trejo Trejo O 223 (108), 279 (136) U Ulloa Herrera R 277 (135) Ulloa Jara M 65 (29) Uribe Cuevas M 137 (65), 231 (112)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
289
V Valadez Manzano LM 281 (137), 283 (138) Valdez Flores JJ 51 (22), 73 (33) Valdez Ledón P 15 (4) Valdez Pineda MC 27(10), 111 (52), 127 (60), 199 (96) Vargas García M 63 (28) Vázquez Garcés M 143 (68) Vega Bolaños A 275 (134) Vega Velásquez A 263 (128), 265 (129),
275 (134) Vega Villasante F 197 (95) Velásquez Polanco RI 95 (44) Velázquez Abunader JI 27 (10) Villanueva Fortanelli JJ 13 (3) Villegas Calzada J 139 (66) Villegas Hernández F 99 (46) Vital Rodríguez B 159 (76), 161 (77) W Wakida Kusunoki AT 145 (69) Z Zárate Becerra ME 75 (34) Zárate Noble VM 243 (118) Zavala Valdez JD 111 (52) Zea De la Cruz H 45 (19), 141 (67), 165 (79), 217 (105), 219 (106),
261 (127) Zetina Moguel CE 137 (65) Zúñiga Flores MS 157 (75), 263 (128),
265 (129) Zúñiga Pacheco LC 157 (75)

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
290

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
291
INDICE ANALITICO CON PALABRAS CLAVE Palabra núm. de página A Abundancia 100, 118, 138, 142,
198, 216, 256 Abundancia relativa 52 Acadja 244 Achirus mazatlanus 124 Administración de ecosistemas 204 Agalleras 258 Agregación 244 Agregaciones 278 AIC 268 Akaike 16, 76, 210, 212 Alometría negativa 240 Alometría positiva 240 Altata 202 Alto Golfo de California 242, 272 Alvarado, Veracruz 14 Amplitud trófica 108 Análisis de actores 204 Análisis de causa raíz 204 Análisis de contenido estomacal 130, 136 Análisis de incremento marginal 106 Análisis de redes sociales 204 Análisis de riesgo 32 Análisis fotográfico 126 Ancho de disco 26 ANMUCOVA 20, 24, 136 Anomalías 32 Anzuelo 232 Anzuelos 248 Áreas de crianza 124 Ariidae 110 Arrastre camaronero 142, 218, 262 Arrecife Alacranes 138 Arte de pesca 174, 182 Avisos de arribo 234, 264 B Bahía Asunción 276 Bahía de Banderas 196 Bahía Sebastián Vizcaíno 226 Baja California 42, 246 Baja California Sur 270, 274 Balance ecológico 154 Ballena jorobada 126 Ballenas 80 Banco de Campeche 44 Banda de crecimiento 176 Bandas 76 Bandas de crecimiento 48 Batoideos 88, 148, 228 Biocapacidad 154
Biocronología 184 Borde opaco 176 Bramidae 200 Brycon guatemalensis 224 C Calidad de vida 50 Callinectes 10, 14 Callinectes bellicosus 270 Camarón 90, 154 Camarón azul 100 Camarón blanco 98 Camarón rosado 146 Cambio climático 28, 38 Campeche 146, 238 Cápsula ovígera 26 Captura 34, 52, 64, 188 Captura comercial 120 Captura incidental 252, 266 Capturas 248 Caracol chino 278 Caracterización 30, 178, 220 Caranx caballus 24 CASA 32 Cautiverio 92, 282 Cazón tutzúm 46 Cazones 214 Cefalópoda 102 Centropomus nigrescens 206 Certificación 62 Certificaciones 82 Chicoasén 280 Chihuil karateca 128 Cichlidae 280 Ciclo reproductivo 158, 224 Co-manejo 82 Comercialización 140 Comercio 230 Comercio Justo 90 Composición 216 Composición de la captura 232 Composición específica 214 Comunidades 66 Comunidades pesqueras 152 Comunidades ribereñas 82 Conectividad 38 Conservación 80, 194 Consumo 230 Contenido estomacal 94, 132 Cooperativas 152 Cooperativismo 58 Copainalá, Chiapas 280

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
292
Corredor San Cosme-Punta Coyote 168 Cortes histológicos 164 Coryphaena hippurus 130 Costa de Jalisco 170 CPUE 44, 64, 118,
172, 216 228 Crassostrea iridescens 164 Crecimiento 12, 22, 42,
122, 160,162, 250, 268, 282
Crecimiento individual 184 Crecimiento poblacional 60 Criterio de información de Akaike 106 Crustáceo 32 Cuasi renta 150 Curvina 272 Cynoscion othonopterus 242 D Densidad 112, 138, 198 Depredadores secundarios 100 Depredador-presa 110 Desarrollo económico y social 90 Desarrollo turístico 152 Descripción de crecimiento 106 Determinación de edad 106 Determinación estadística 52 Dieta 94, 110 Dimorfismo sexual 96 Dinámica de flotas 182 Dinámica poblacional 76 Distribución 44, 180, 238, 256 Distribución espacial 278 Diversidad 108, 256 Dorado 70 E Ecología pesquera 10 Ecología trófica 110, 114 Edad 22, 176, 206 Edad y crecimiento 76 EDK's 136 Edks circulares 18 Eficiencia 248 Elasmobranquios 134,196, 218, 234 Enmallamientos 80 ENSO 276 Epinefélidos 28 Epinephelus morio 232 Época de desove 36, 128 Época de incubación 128 Escama 174, 206, 216
Escama marina 264 Esfuerzo 64, 188 Esfuerzo pesquero 52, 134, 144 Especies 258 Especies bentónicas 138 Especies compartidas 262 Especies dominantes 120 Especies inquerendae 192 Especies nativas 280 Especies objetivo 182 Espectro Trófico 136 Estándares 62 Estiletes 122 Estilo reproductivo 88 Estructura 198 Estructura de pesos 112 Estructura de tallas 68, 98, 100, 208,
254, 260 Evaluaciones pesqueras 168 Excluidor 266 Éxito reproductivo 88 Explotación 236 Extirpación 194 F FAC 260 Factor de condición 92, 206 Factor de conversión 86 Familia 34 Fauna 284 Fauna de acompañamiento 68, 94, 148 Fecundidad 46, 78 Fotoperiodo 156 Función de verosimilitud 268 G Gaviota 56 Generación de información pesquera 168 Glándula oviducal 26 Golfo de California 48 Golfo de México 186, 218 Golfo de Tehuantepec 252, 256 Guerrero 118 H Hábitat 54, 180 Hábitos alimenticios 104, 130, 132 Haemulon flavigutattum 20 Historia de vida oportunista 36 Huachinango 118

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
293
I Ictiofauna 256 IGS 20, 24 IHS 20, 24 Impacto 132 Incertidumbre 56 Incidentalidad 218, 262 Incremento en peso 92 Incremento marginal en escamas 206 Incrementos 122 Índice de importancia relativa 94, 104 Índice gonadosomático 18, 124, 156, 158,
170 Intermediarios 140 Intervalo de confianza 16, 210, 212 Intervalos de confianza 96 Investigación 10 Isometría 240 Isótopos Estables 114 J Jaiba 32, 238, 270 Jaibas 10 Japuta 206 K K 20, 24 L Laguna de Términos 238 Lagunas de Cruz de Loreto 64 Langosta 62 Langosta roja 276 Líneas 258 Listado 222 Literatura de acceso libre 132 Longevidad 176 Longitud 104 Longitud de madurez 124, 170, 242 Longitud de primera madurez 36, 224, 250 Longitud media de madurez 78 Longitud mínima de captura 232 Longitud primera captura 208 Longitud ojo-furca 72 Longitud-edad 268 Longitudes 222, 250 Lutjanus inermis 22 M Macrobrachium 54 Macrobrachium tenellum 12
Madurez 98 Madurez gonádica 102, 164, 250 Madurez temprana 12 Manejo 82 Manejo pesquero 30, 182 Manta gigante 196 Mantarraya 196 Marcaje convencional 226 Marginación 50 Marginalidad 140 Máxima verosimilitud 16, 242 Medida de manejo 156 México 30, 42, 130, 242 Michoacán 134, 164 Migración 278 Mínimos cuadrados 210, 212 Mobulidae 196 Modelo bioeconómico 272 Modelo de reservas marinas 40 Modelo multinomial 100 Modelos 84, 96 Modelos de crecimiento 106 Modelos GAM 246 Monitoreo pesquero 168 Mortalidad 12, 60, 116, 208 MRS 32 MSC 62 Mujeres 34 Multiespecífica 258 Multimodelo 42 N Neonatos 254 Nivel trófico 94, 154 Normatividad 54 O Océano Pacífico Oriental 130 Octopoda 102 Octopus maya 180 Oficina de pesca 264 Olmstead-Tukey 120 Oreochromis spp 160, 162 Ostión 198, 244 Ostión de roca 164 Otolitos 22 Ovario 26, 78 P Pacífico 198 Pacífico Sur 190 Palangre 52,166, 172,
248, 262

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
294
Palangre pelágico 246 Pandalus platyceros 42 Panulirus spp 282, 284 Parámetros biológicos 28 Parámetros de crecimiento 102 Paredón 222 Participación 34 Participación social 50 Peces marinos 230 Peces sierra 194 Pepino de mar 86, 112 Percepción 50 Periodicidad 48 Periodos térmicos 120 Pesca 44, 186 Pesca artesanal 28, 30, 56, 78,
170, 236, 274 Pesca de arrastre 252 Pesca de mediana altura 246 Pesca deportiva 72 Pesca deportiva recreativa 178 Pesca ilegal 14 Pesca incidental 80, 184, 194 Pesca ribereña 38, 40, 76, 118, 126, 140, 144,
146, 150, 166, 172, 174, 188, 228
Pescadores 66 Peso 72, 86 Peso promedio 70 Pesquería 10, 190, 222,
230, 238, 254 Pesquería artesanal 134 Pesquería de escama 84 Pesquería de langosta 266 Pesquería multiespecífica 182 Pesquerías ribereñas 58, 204 Pez de profundidad 200 Pez vela 72, 178 Plan de Manejo 202 Planeación espacial marina 38 Planes de manejo 184 Pleuronectiformes 124 Políticas pesqueras 202 Políticas públicas (PP) 50 Postlarvas 282, 284 Potencial pesquero 200, 244 Potencial reproductivo 112 Precipitaciones 156 Presa El Gallo 160 Presa La Yesca 162 Presas principales 108 Prevención 80 Primera zoae 12 Pristis pectinata 194 Pristis pristis 194
Problemática de la pesquería 54 Problemáticas 144 Proceso de cooperación 56 Producción 230, 264 Producción doméstica 34 Producción pesquera 234 Productividad biológica 60 Productividad primaria 154 Profundidad 142, 166 Proporción sexual 124, 136, 170 Puerto San Carlos 58 Pulpo 122 Pulpo rojo 180 R R. steindachneri 48 Rayas 134, 142, 190, 218, 220, 228, 252 Recurso 216 Recurso pesquero 192, 214 Recursos pesqueros 118, 174 Redes de enmalle 228 Redes neuronales artificiales 84 Refugio pesquero 144 Regionalización 58 Registro taxonómico 200 Reglamentación pesquera 36 Relación Longitud-Peso 68, 102 Remuestreo 16 Rendimiento 86, 160, 162 Rendimientos 116, 150 Reproducción 18, 46, 156, 164, 270 Reproducción asincrónica 74 Reservas marinas 38 Resiliencia 60 Retrocálculo 74 Ribereña 66 Riesgo 126 Río Grijalva 224, 280 Riqueza específica de peces 138 Ronco blanco 76 S Sardinas 274 Satisfacción 152 Scianidae 170 Selección 236 Selectividad 116, 248 Series de tiempo 70, 72 Seriola lalandi 30 SIG 58 Sinaloa 28, 260, 282, 284 Sinergia 152

VIII Foro Científico de Pesca Ribereña Puerto Vallarta, Jalisco. 2016
295
Sistema estuarino lagunar 68 Sistema lagunar 202 Sobrepesca 66 Sobrevivencia 116, 236 Socioeconómico 174 Sociología 58 Soluciones 144 Subsidios 66 Suripera 90 Sustentabilidad 62, 82 T Tabasco 234 Tasa de explotación 116, 160, 162, 208 Tasas de captura 214 Taxon 192 Tecolutla 220 Teleósteos 166, 220 Temperatura superficial del mar (TSM) 120 Tendencia 70 Tendencia latitudinal 240 Teoría de información 1 6, 212, 212 Testículo 26 Tiburón 46, 78, 254 Tiburón blanco 114 Tiburón mako 114 Tiburón sedoso 184 Tiburones 44, 132, 134, 142 172, 186, 218, 220, 222, 226 Tilapia 160, 162 Tinción 164 Tipo de crecimiento 240 Tipologías 58 Trampas 14 Traslape ecológico 110 Traslape 108 U Umbrina xanti 170 Urotrygon 190 V Valor económico 188 Variabilidad individual 268 Veda 46 Veda de tiburones 246 Veda reproductiva 128 Veracruz 166 Verdillo 158 Verosimilitud 96 Vértebra 176
Vértebras 48, 74, 106, 148 Viajes en lancha 178 Volumen de captura 230 von Bertalanffy 74, 104 Vulnerabilidad 88, 140, 186 Y Yucatán 122, 146, 180 Z Zona norte de Campeche 150 Zonas de refugio pesquero 40





























