Embed Size (px)
Citation preview

Adaptation and Constraint in the Plant Reproductive Phase
Kjell Bolmgren
Department of Botany Stockholm University
Stockholm 2004

Adaptation and constraint in the plant reproductive phase
ISBN: 91-7265-856-8, pp. 1-46 © Kjell Bolmgren 2004 Cover design by Torsten Hild (www.2hild.com) Cover photo © LSH foto Samuel Uhrdin (’Vertumnus’ by Guiseppe Arcimboldo, 1527-1593) Jannes Snabbtryck Kuvertproffset HB
2

Kjell Bolmgren
Doctoral dissertation 2004 Kjell Bolmgren Department of Botany Stockholm University SE-106 91 Stockholm Sweden Adaptation and Constraint in the Plant Reproductive Phase Abstract — Conservatism is a central theme of organismic evolution. Related species share characteristics due to their common ancestry. Some concern have been raised among evolutionary biologists, whether such conservatism is an expression of natural selection or of a constrained ability to adapt.
This thesis explores adaptations and constraints within the plant reproductive phase, particularly in relation to the evolution of fleshy fruit types (berries, drupes, etc.) and the seasonal timing of flowering and fruiting. The different studies were arranged along a hierarchy of scale, with general data sets sampled among seed plants at the global scale, through more specific analyses of character evolution within the genus Rhamnus s.l. L. (Rhamnaceae), to descriptive and experimental field studies in a local population of Frangula alnus (Rhamnaceae). Apart from the field study, this thesis is mainly based on comparative methods explicitly incorporating phylogenetic relationships. The comparative study of Rhamnus s.l. species included the reconstruc-tion of phylogenetic hypotheses based on DNA sequences.
Among geographically overlapping sister clades, biotic pollination was not corre-lated with higher species richness when compared to wind pollinated plants. Among woody plants, clades characterized by fleshy fruit types were more species rich than their dry-fruited sister clades, suggesting that the fleshy fruit is a key innovation in woody habitats. Moreover, evolution of fleshy fruits was correlated with a change to more closed (darker) habitats.
An independent contrast study within Rhamnus s.l. documented allometric rela-tions between plant and fruit size. As a phylogenetic constraint, allometric effects must be considered weak or non-existent, though, as they did not prevail among different subclades within Rhamnus s.l. Fruit size was correlated with seed size and seed num-ber in F. alnus. This thesis suggests that frugivore selection on fleshy fruit may be important by constraining the upper limits of fruit size, when a plant lineage is colo-nizing (darker) habitats where larger seed size is adaptive.
Phenological correlations with fruit set, dispersal, and seed size in F. alnus, sug-gested that the evolution of reproductive phenology is constrained by trade-offs and partial interdependences between flowering, fruiting, dispersal, and recruitment phases. Phylogenetic constraints on the evolution of phenology were indicated by a lack of correlation between flowering time and seasonal length within Rhamnus ca-thartica and F. alnus, respectively. On the other hand, flowering time was correlated with seasonal length among Rhamnus s.l. species. Phenological differences between biotically and wind pollinated angiosperms also suggested adaptive change in repro-ductive phenology.
3

Adaptation and constraint in the plant reproductive phase
Den halvfärdiga himlen av Tomas Tranströmer 1962 Modlösheten avbryter sitt lopp. Ångesten avbryter sitt lopp. Gamen avbryter sin flykt. Det ivriga ljuset rinner fram, även spökena tar sig en klunk. Och våra målningar kommer idagen, våra istidsateljéers röda djur. Allting börjar se sig omkring. Vi går i hundratals. Var människa en halvöppen dörr som leder till ett rum för alla. Den oändliga marken under oss. Vattnet lyser mellan träden.
Insjön är ett fönster mot jorden.
4

Kjell Bolmgren
Acknowledgments Jag har en svaghet. Jag misstror mina närmastes uppmuntrande ord. Som om de bara vill vara vänliga. Därför, med eftertryck: Lott, tack för att du förväntade dig något märkvärdigt av mig. Ove, tack för ditt obegripliga lugn. Det har vid upprepade tillfällen varit en förutsätt-ning för att jag inte skulle lämna skutan, liksom din ständiga beredskap med och kvali-tén i den röda pennan. Tack för att du lät mig gå min väg. Tack Bengt för att du, trots mitt eviga kvacksalvande och vänsterhandsfipplande, alltid fick konkreta saker att hända. Mikael Mildén, Heléne Fröborg, Regina Lindborg, Sara Cousins (frustrationsklubben) — ni har varit den ventil jag så ofta behövt . Tack. Åsa Eriksson, Petra Korall och Didrik Vanhoenacker — ni var räddande änglar i för mig mycket kritiska lägen. Tack. Jens Klackenberg och Thomas Karlsson vid Naturhistoriska riksmuséet öppnade en ny värld för mig. Stort tack. A special thanks to Arndt Hampe, my brother-in-Frangula, and to Peter Linder for an encouraging cooperation. Många arbetskompisar på avdelningen har varit oersättliga diskussionskamrater om allt från krig och fred till samplingsmetodik, och alla på avdelningen har varit viktiga för den hjälpsamma atmosfär som varit en förutsättning för att jag skulle kunna tänka det jag tänkt. Tack alla. Ett särskilt tack till Thord Fransson för att du tog hand om mig trots att jag var mer intresserad av fågelskiten än av fåglarna, till Patrik Lindenfors, Patrik Dinnetz och Kari Lehtilä för att ni varit generösa med att diskutera mina metodologiska bryderier, till Johan Ehrlén för att man kan lita på att du är skeptisk, till Katariina Kiviniemi-Birgersson för att man kan lita på att du tycker det är kul, till Lenn Jerling och Peo Karis för att era dörrar alltid stått öppna och till Niklas Wikström, Hans-Erik Wann-torp och Torsten Eriksson för alla lunchdiskussioner. Jenny Smedmark, Moa Holmlund, Anna Koffman, Nahid Heidari, Reija Dufva, Johan Lind, Cilla Kullberg och Martin Starvander har UV-bestrålat pollenslangar, samlat litteraturdata, översatt ryska texter, sekvenserat DNA, fångat fåglar och gjort mycket annat som jag inte haft kompetens för. Stort tack.
5

Adaptation and constraint in the plant reproductive phase
Contents
Summary 3
Acknowledgments 5 List of papers 7
Preface 9
Introduction 10 Adaptation and Constraint 11 The Plant Reproductive Phase 15 Objectives and Approach 18
Methods 19 Phylogenies and Comparative Biology 19 Data collection 23
Results 25
Discussion 32
Literature cited 38
Svensk sammanfattning 45
Appendix I—V 47
6

Kjell Bolmgren
List of papers
This thesis is based on five separate studies listed below. They will be referred to by their Roman numbers. The published papers are reprinted with the kind permission of the publishers.
I Bolmgren, K., O. Eriksson, and H. P. Linder. 2003. Contrasting flow-ering phenology and species richness in abiotically and biotically polli-nated angiosperms. Evolution 57:2001-2011.
II Bolmgren, K. and O. Eriksson. Fleshy fruits – origins, niche shifts, and diversification.
III Bolmgren, K. and B. Oxelman. in press. Generic limits in Rhamnus s.l. L. (Rhamnaceae) inferred from nuclear and chloroplast DNA sequence phylogenies. Taxon (in press).
IV Bolmgren, K. Constraints, conservatism and adaptation in the evolution of fleshy fruits and flowering phenology of Rhamnus s.l. (Rhamnaceae).
V Bolmgren, K. and O. Eriksson. Phenology and ovule fate in Frangula alnus (Rhamnaceae) – interdependence or decoupling of flowering, fruiting, dispersal, and early recruitment phases?
In all papers, I performed the studies and wrote the manuscripts. In paper I and V, I
also developed the ideas, while the study in paper II was developed together with OE.
In paper III, I developed the study, while BO guided the PAUP analyses.
7

Adaptation and constraint in the plant reproductive phase
8

Kjell Bolmgren
Preface
At the core of evolutionary biology is the discussion about adaptations and adaptabil-ity. In a way this discussion is an analogue to the philosophical discussion about free-dom and the free will, including the Nature vs. Nurture debate. To what extent are you free? What forces restrain the manifestation of the free will? How strong are these forces? One could even argue that evolutionary studies on the ability of organisms to respond and adapt to local conditions are fundamental to our understanding of freedom as a concept.
Are organisms free to adapt to the conditions they meet? To what extent does the evolutionary history constrain adaptability? How much of an organism or the cha-racteristics of an organism manifest adaptations to the local conditions under which they live their lives, and how much are due to phylogenetic constraints? Are manifes-tations of phylogenetic constraints and local adaptation mutually exclusive?
The present thesis emanated from the hypothesis that autumn migrating seed dispersing birds exert a selection pressure for fruiting time on fleshy fruited plants. A preliminary literature survey found that the evolution of phenology and fleshy fruits were considered phylogenetically constrained and that frugivores were considered too weak a selective agent to drive fleshy fruit evolution. These ideas served as stimulus to develop a couple of studies which revolved around constraints, conservatism, and adaptations.
9

Adaptation and constraint in the plant reproductive phase
Introduction
Diversity and conservatism characterize the evolution of organisms. Variation in be-haviour and form is perhaps the strongest impression of life, but conservatism is an equally important part of evolution; roses are roses, tulips are tulips, and anemones are anemones. No matter how different they are, we have generally no problem in recog-nizing related taxa. These two sides of life, diversity and conservatism, corresponds with the “two great laws — Unity of Type and the Conditions of Existence” (Darwin 1859, p. 233) that shape organisms. Unity of Type is a pre-Darwinian concept referred to Cuvier, which Darwin (1859, p. 233) transformed into his own concept unity of descent.
It is of course tempting, considering the astonishing biological diversity, to suggest that organisms attain any form (necessary) through the work of natural selec-tion; freezing bats, animal-eating plants, flying mammals, glass-eating lichens, non-swimming fishes, deep-sea-living mammals, individual fungi covering hundreds of kilometres, and plants living for hundreds of years — “I’m simply saying that … life finds a way”, as Jeff Goldblum alias Dr. Malcolm stated in Jurassic Park. Jean-Baptiste de Monet de Lamarck, who was the first to develop a coherent theory on adaptive evolution, even believed that the adaptive ability of organisms was so devel-oped that extinction was not an issue; each species develops into a new species (Bergström 1993; Brooks and McLennan 2002). Today we consider the character complex of an organism as anachronistic and, at least from a reductionist view, not at all optimally adapted to local conditions.
A previous underestimation of constraints on adaptation, particularly effects of unity of descent, and an interpretative precedence given to adaptive explanations, got the pendulum swinging during the seventies. Starting with the sociobiology debate, evolutionary biologists got involved in a development shifting the focus from local adaptations to effects related to the organisms’ phylogenetic history (Gould and Le-wontin 1979). The research field ’comparative biology’, explicitly incorporating phy-logenetic information, expanded rapidly, which lead to a critical examination of which questions that could (should?) be asked (Wanntorp 1983), important insights on statis-tical issues (Felsenstein 1985), and, in relation to this thesis, an interest in non-
10

Kjell Bolmgren
adaptive interpretations of organismic features and which characteristics that constrain the adaptability of these features. Adaptation and Constraint Adaptations are features that have evolved by natural selection. That is, an adaptation is a heritable characteristic that have replaced other variants of the character in the population, as it yielded a superior increase of (inclusive) fitness. Phylogenetic defini-tions of adaptations include the somewhat controversial part that the feature should be a phylogenetic novelty. This highlights a difference between adaptation as a process and adaptation as a feature. Adaptation may be an ongoing process where a trait is conserved owing to stabilizing selection, but, according to the phylogenetic definition of adaptations, no novel features and thus no adaptations arise during such processes. Whether one should call traits that are phylogenetically conserved through stabilizing selection an adaptation or not, could just have been a matter of semantics, but the de-velopment of an intimate relationship between parsimony criteria and non-adaptive interpretations have stirred up a debate on these issues (Westoby et al. 1995).
Conserved characters have played an essential role in this debate. The explicit use of phylogenetic information in comparative biology has, as expected, confirmed that a majority of characteristics of an organism are conserved in a phylogenetic con-text. It is generally acknowledged among evolutionary biologists, that present charac-ter states express a compromise between local adaptation and phylogenetic history, but when it comes to conserved traits this balanced view has shifted in favour of phyloge-netic ‘explanations’, stating ‘no change – no process’. The rationale for such views is parsimony. Phylogenetic patterns of conserved characters do not reveal, though, whether the conserved traits are actively maintained by selection or if they have been conserved because they are fixed parts of the organism. In the special case of discrete characters, it is understandable how such a line of ‘no change–no process’ reasoning has come about. However, considering quantitative characters, for which equally strong phylogenetic conservatism is expected, the suggestion of constrained evolution becomes unreasonable (Westoby et al. 1995). For example, despite a large within-group variation, Kochmer and Handel (1986) and Hodgson and Mackey (1986), re-spectively, interpreted significant phylogenetic effects (sensu unity of descent, see Derrickson and Ricklefs 1988) as if character evolution was constrained. Such reason-
11

Adaptation and constraint in the plant reproductive phase
ing implicitly defines phylogenetic and adaptive explanations as mutually exclusive alternatives, and equalizes phylogenetic constraints with phylogenetic effects.
That phylogenetic history restrains the adaptability of an organism is generally acknowledged; the lineage-specific heritage is part of the unique conditions of exis-tence shaping each organism. Therefore, the evolutionary pathways available for each organism are also unique and limited. Conserved characters have been understood as a typical example of such phylogenetic ballasts, owing to the fact that in a world that is generally perceived to be continuously changing, conserved characters are not what is expected if adaptation is a dominant process. Taken a step further, phylogenetically conserved characters have been classified as phylogenetically constrained. In fact, constraint (historical, phylogenetic, or evolutionary) seem to have been the common term when characterizing evolution of conserved characters in the aftermath of Gould and Lewontin (1979). Once again, this could have been merely a question of seman-tics, but as phylogenetic explanations were often used as mutually exclusive, non-adaptive alternatives to adaptive explanations, the conception of conserved characters being constrained has been substantially criticized. Adaptive processes may equally well maintain conserved characters (Westoby et al. 1995), and one of these adaptive processes, phylogenetic niche conservatism (Harvey and Pagel 1991; Lord et al. 1995), will be of particular interest in this thesis.
Phylogenetic niche conservatism (PNC) is based on the idea that the selective milieu is not as dynamic as one might believe. All taxa in a local species pool will not have equal opportunities in exploiting an adjacent novel niche space. Those taxa adapted to a preceding niche space most similar to the novel conditions is expected to be the ones that successfully exploit the novel niche space. As the novel niche space is similar to preceding conditions for these taxa, the character complex of the exploiting taxa will appear generally conserved, according to the PNC hypothesis. Major direc-tional, adaptive changes will then only take place when there are major changes in the selective milieu (extinctions, island colonization, climatic change; cf punctuated equi-librium). Thus, PNC diverges from phylogenetic constraints by not invoking intrinsic, lineage-specific obstacles for adaptation, suggesting that conserved characters persist due to prevailing adaptive values (Ackerly and Donoghue 1995; Westoby et al. 1995). The potential importance of phylogenetic niche conservatism does not mean, of course, that phylogenetic constraints are unimportant, it just illustrates that phylogeny is not a constraint. To be able to separate between phylogenetic niche conservatism
12

Kjell Bolmgren
and phylogenetic constraints, claims of phylogenetic constraints must be based on more mechanistic evidence (Antonovics and van Tienderen 1991).
The common meaning of the word constrain is synonymous to limit and re-strain, and a feature is constrained when it is forced, uneasy, or unnatural (!) according to an Oxford dictionary. So, can life be unnatural? In fact, along with adaptive change and genetic drift, constraint is one of the characteristics of organismic evolution, in the sense that “yesterday’s adaptation may become tomorrow’s constraint”1. Such all encompassing use of the concept of constraint is of little value to operational studies of evolutionary constraints (Stearns 1986; Gould 1989). Further, a casual use of phyloge-netic constraints (PC) as a non-adaptive fall-back explanation has accumulated much critique of the concept (Stearns 1986; Gould 1989; Antonovics and van Tienderen 1991; Westoby et al. 1995), and during the last decade some necessary development of the concept has taken place (Antonovics and van Tienderen 1991; Edwards and Naeem 1993; McKitrick 1993; Miles and Dunham 1993; Schwenk 1995; Burt 2001; Blomberg and Garland 2002).
McKitrick suggested that a PC is “any result or component of the phylogenetic history of a lineage that prevents an anticipated course of evolution in that lineage” (McKitrick 1993). An important aspect of this definition is the explicit requirement of a specified anticipated course of evolution, meaning that claims of PC does not have an interpretative precedence over, for example, phylogenetic niche conservatism if both the character and the selective milieu are conserved (Figure 1). Miles and Dunham (1993) further emphasized that PC does not necessarily concern a conserva-tion of character states (Figure 1). If the variation of two or more traits are linked, an adaptive change in one character, may lead to a forced, non-adaptive change in an-other, linked character even if the selective milieu is conserved in relation to this char-acter (Figure 1). This is an important point, as several authors have only considered PC in relation to conserved characters (e.g. Burt 2001). Schwenk (1995) divided con-served characters into two classes based on whether (stabilizing) selection or non-selection processes are responsible for the conservatism, and suggested that “a useful conception of evolutionary constraint (PC) must exclude natural selection as a causa-tion agent” (Schwenk 1995, p. 254). Schwenk’s definition thereby implies that charac-ters that are conserved due to opposing selection pressures do not represent examples 1 This expression is used in several texts without reference to an original source. McKitrick (1993) cites a manuscript in press, which has never been published as far as I know.
13

Adaptation and constraint in the plant reproductive phase
changedconserved
EUSELECTIVE MILI
PHYLOGENT
ETICRAINTS
ICHE SM
CONS
PHYLOGE- NETIC N
CONSERVATI
ETIC RAINTS
PHYLOGEN
TCONS
ETIC RAINTS
changed
conserved
TRAIT
ON
ADAPTATI
PHYLOGEN
TCONS
Figure 1. An illustration of how phylogenetic niche conservatism, phylogenetic constraints, and adapta-tion are defined and relate to each other. A classification like this may add to the polarized view of phylogenetic versus adaptationist explanations of character evolution that is common in the literature. Therefore, I would like to stress that many characters are likely to express both phylogenetic constraints and recent adaptation (Westoby et al. 1995; Orzack and Sober 2001). (Paper IV)
of phylogenetically constrained evolution. So, which are the non-selective mechanisms that can generate phylogenetic constraints?
Allometry, trade-offs (developmental processes), and pleiotropy (a genetic process) are mechanisms generally proposed to lead to phylogenetically constrained character evolution. Lack of genetic variation is not considered a PC. It is the mecha-nism, preventing or reducing genetic or phenotypic variation, that is a PC. In conclu-sion, to claim that phylogenetic constraints are responsible for the evolutionary (or lack of) response in a character, one must show that a sufficient directional selection does not develop into an anticipated response due to a defined constraining mechanism (Antonovics and van Tienderen 1991). An additional criteria is that the anticipated adaptive response must appear in the phylogenetic sister clade lacking the suggested constraining feature (Stearns 1992). None of these definitions and criteria suggest that PC:s are unbreakable, and all of them emphasize that PC:s concern the evolution of
14

Kjell Bolmgren
characters, not organisms. This focus on characters cut off the research field on key innovations from the field of phylogenetic constraints.
Key innovations — In correspondence with the more liberal definitions of phyloge-netic constraints, key innovations have been suggested as an explanation for compara-tively high diversification rates in certain clades. Key innovations are phylogenetic novelties leading its descendents into a new niche space and thus deliberating the or-ganisms from ancestral, constraining characteristics. For example, the extraordinary species richness of angiosperms, beetles, and passerines, have risen the question whether the evolution of specific features can explain why these clades expanded more than their related clades. Regarding angiosperm radiation, biotic pollination, biotic dispersal, herbaceousness, and a short juvenile phase, are some of the features that have been suggested as such key innovations (Stebbins 1981; Crepet 1984; Tiffney 1984; Eriksson and Bremer 1992; Dodd et al. 1999; Verdu 2002). Originally, the con-cept of key innovations was used in relation to singular evolutionary radiations, as the examples above, and this approach has rendered the concept severe critique from both statistical and phylogenetic points of view (Raikow 1986). Considering this critique, the operational applications of the key innovation concept has been related more to convergent evolution (Mitter et al. 1988; Eriksson and Bremer 1992; McKitrick 1993; Miles and Dunham 1993; Dodd et al. 1999; Verdu 2002), that is, to analyse if phy-logenetically independent origins of functionally analogous features have led to com-paratively higher species richness. I use this approach here, when examining the effect of pollination syndromes and fruit types on species richness (paper I and II). The Plant Reproductive Phase The plant reproductive phase encompasses the development of flowers (incl. ovules and pollen) and fruits (incl. seeds). Along with the evolution of herbaceousness and a short juvenile phase, biotic pollination and biotic seed dispersal have been suggested to be responsible for much of angiosperm diversification (Grant 1949; Stebbins 1981; Crepet 1984; Eriksson and Bremer 1992; Ricklefs and Renner 1994; Dodd et al. 1999; Verdu 2002). Thus, many characteristics of the plant reproductive phase have been subjected to evolutionary studies. This thesis deals with adaptations and constraints in the evolution of two plant reproductive features: phenology and fleshy fruits.
15

Adaptation and constraint in the plant reproductive phase
Phenology — The temporal distribution of flowers and fruits have been considered a potential part of the reproductive isolating mechanisms leading to speciation in angio-sperms (Snow 1965; Carpenter 1976; Stiles 1977; Waser 1983). Thus, phenological niche structure, but also other aspects of phenology, have gained considerable atten-tion (Robertson 1895; Robertson 1924; Clausen et al. 1948; Slade et al. 1975; Thomp-son and Willson 1979; Primack 1980; Thomson 1980; Augspurger 1981; Rabinowitz et al. 1981; Kochmer and Handel 1986; Eriksson and Ehrlén 1991; Fuentes 1992; Pico and Retana 2000; Oberrath and Bohning-Gaese 2002; Lacey et al. 2003; Rathcke 2003; for reviews, see Primack 1985; Rathcke and Lacey 1985; Primack 1987; Oller-ton and Lack 1992; Gotelli and Graves 1996; Fenner 1998). Phenology has also been, and probably originated as, a major subject in agri- and silviculture. The economic importance of predicting phenological events (e.g. time of harvesting) and foreseeing the local match between local conditions and the phenology of commercial genotypes, have initiated a huge number of studies correlating phenology to climatic conditions. These issues have gained renewed interest, also within ecology, due to Global Change.
The phenological strategy of a plant shall potentially attract pollinators and seed dispersers, satiate predators and herbivores, effect cross-pollination, deal with varying abiotic (e.g. climatic) conditions within and between seasons, and withstand competition from co-appearing plant species. In north temperate conditions, plants must also deal with a short reproductive season.
The basic issue in evolutionary plant reproductive phenology deals with the seasonal position of a phenological event, that is, the timing of the event. For example, Linnaeus’ altruistic, pre-Darwinian suggestion was that the seasonal sequence of flow-ering has come about so that “these hundreds of plants shall be spared from causing each other harm” (Linnaeus 1760, p. 135). Today we consider all aspects of the tempo-ral distribution of flowering and/or fruiting resources as parts of a plant’s phenological strategy. That is, not only the position of the phenological curve, but also its shape is considered evolvable traits (Thomson 1980; Eriksson 1995). Such aspects of evolu-tionary phenology is further strengthened by the insight that a phenological curve is probably a compromise both between different selection pressures within the pheno-phase, as well as between selection pressures acting in different phenophases that are more or less interdependent. It seems quite obvious that the evolution of flowering and fruiting curves evolve more or less in concert (Primack 1987). These interdependences may create counteracting selection pressures on reproductive phenology traits. More-
16

Kjell Bolmgren
over, several authors have suggested that flowering and fruiting phenology may be phylogenetically constrained (Bell and Stephens 1984; Kochmer and Handel 1986; Johnson 1993; Jordano 2000), though without defining any constraining mechanism. Fleshy fruits and frugivory — The germination, seedling, and early juvenile phase is considered the most critical phase in the plant life cycle (Grubb 1977), suggesting that improved seed dispersal may have a major impact on plant fitness. The evolution of fleshy fruits have basically been attributed to three different, though not mutually exclusive, ideas. First, frugivory may facilitate dispersal away from the mother plant, reducing density-dependent offspring mortality. Second, the exploitation of animal movements suggests that seeds are dispersed in a non-random pattern, potentially to favourable recruitment sites as, for example, forest clearings and forest margins. Third, gut passage may improve germination.
Irrespective of which hypothesis that is correct, fleshy fruits must attain the right size, form, and content (nutrient, water and seeds), and appear at the right time to attract effective dispersers. Based on such hypotheses, two decades of plant-frugivore research have failed to find general support for a close match between frugivore and plant characteristics (Jordano 2000; Herrera 2002b; but see Mazer and Wheelwright 1993; Noma and Yumoto 1997; Rey et al. 1997; Wenny and Levey 1998), and there-fore fleshy fruit evolution has been considered phylogenetically constrained (Jordano 1995a). As an exception, fleshy fruit size has been considered the only fruit character that may have been adapted to local seed dispersing frugivore assemblages and thus not phylogenetically constrained (Wheelwright 1985; Mack 1993; Sallabanks 1993; Jordano 1995a; Jordano 1995b; Herrera 2002b; Lord et al. 2002).
Considering the coarse but general observation that there are correlations be-tween seed size, endozoochorous seed dispersal, and high statured vegetation (Willson et al. 1990; Hughes et al. 1994), two additional hypotheses on fleshy fruit evolution have developed. Herrera (2002a) suggested that allometric constraints may be impor-tant in governing fleshy fruit size evolution. That fleshy fruit size variation should be a result of selection on other size traits, does not explain the evolution of fleshiness per se, but, assuming that larger seeds disperse shorter distances, dispersal attributes, as fleshiness, may evolve to compensate for the dispersal loss. Such a scenario does not require a close match between frugivore and fruit characteristics. The second addi-tional hypothesis, suggests a similar line of evolution, though it is not based on devel-opmental constraints. In areas or times where dense vegetation predominates, for ex-
17

Adaptation and constraint in the plant reproductive phase
ample due to warmer climate, selection may act for both higher stature and larger seeds to cope with intensified competition for light. Under these conditions, both dis-persal away from density-dependent mortality and to light gaps may be a rationale for evolving fleshiness. This potentially climate driven evolution of fleshy fruits has sometimes been combined with an observation of conserved fruit traits (Jordano 1995a) to suggest that fleshy fruits in most taxa evolved during a limited period of global warming in the Early Tertiary (Tiffney 1984; Jordano 1987; Herrera 1992; Jordano 1995a; Eriksson et al. 2000). Objectives and Approach In this thesis I have studied adaptations and constraints in the evolution of reproduc-tive phenology and fleshy fruits. A basic objective has been to challenge previous claims that the evolution of both phenology (Kochmer and Handel 1986; Fenner 1998) and fleshy fruits (Jordano 1995a; Jordano 2000; Herrera 2002b; Herrera 2002a) are phylogenetically constrained. My approach has been to examine reproductive phenol-ogy and fleshy fruits at a hierarchy of levels, from experimental and descriptive ex-aminations of local populations, through comparative studies using reconstructed phy-logenetic hypotheses of a genus, up to comparative studies based on data sets sampled among all seed plants at the global scale. More explicitly stated, I have examined: anticipated courses of phenology evolution in relation to pollination systems (paper I) and climatological gradients (paper IV); anticipated courses of fleshy fruit evolution in relation to habitat characteristics and growth form (paper II); allometry as a phyloge-netic constraint on fleshy fruit size evolution (paper III and IV); simultaneously acting selection pressures on reproductive phenology (paper V); the strength of phylogenetic constraints (paper III and IV); and whether biotic pollination and fleshy fruit types may be considered key innovations (paper I and II).
18

Kjell Bolmgren
Methods
The hierarchical approach structuring this thesis is based on some obvious points; specific questions must be addressed at the right level; experimental studies are not possible at a general level; general claims must be supported by general samples; etc. In paper I and II, I challenged previous claims concerning flowering times of biotically pollinated plants (Kochmer and Handel 1986) and fleshy fruit evolution (Tiffney 1984; Jordano 1995a; Eriksson et al. 2000), and thus sampled among angiosperms and seed plants, respectively. High level studies like these have the potential to provide more interesting results in the sense that they suggest character complexes that are adaptive and persistent in evolutionary time. On the other hand, conclusions about processes governing the observed patterns can only be speculative at this level. In paper III and IV, I explored the phylogeny and character evolution within the monophyletic clade Rhamnus s.l. L. Such intermediate level studies allow more highly resolved phylogen-ies and a closer knowledge of the studied species, making more detailed interpretations (or speculations) of the evolutionary patterns possible. Finally, low level (local) stud-ies gave the opportunity for experiments and detailed descriptive work on different selection pressures working within and between phenological phases. At this level it was also possible to examine the interphase between phenology and fruit and seed characteristics in one fleshy fruited plant species. The drawback of these fine-grained studies are that they are sensitive to fluctuating conditions, may suggest irrelevant (evolutionary) patterns (Ehrlén 2003), or only be able to document inconsistent pat-terns (Herrera 1998). Phylogenies and Comparative Biology Interspecific comparisons is an essential tool in evolutionary biology. Incorporating explicit geneaological information into such studies have highlighted the issue of sta-tistical independence. Closely related species have a common history sharing ancestral traits, which implies that information on, for example, present character correlations in a number of species may not be founded on statistically independent observations. If phylogenetic hypotheses are considered in the sampling procedure, comparative stud-
19

Adaptation and constraint in the plant reproductive phase
ies may be based on independent observations. Further, the explicit application of phylogenetic information allowed for an examination of whether patterns of phyloge-netic constraints (i.e. allometry) prevailed at different levels in a clade.
During the last decades the production of phylogenetic hypotheses has in-creased tremendously. Irrespective of whether comparative biologists need to con-struct a composite phylogeny or make use of existing phylogenies, they have a vast open source of working material at their disposal. This does not mean that there are phylogenies available for every question, particularly not when a highly resolved phy-logeny is needed. In this thesis, I have used phylogenetic information at different lev-els of resolution. In paper I and II, where I analysed phylogenetically matched pairs (PIMPs), the samples were partly delimited by the availability of published phyloge-netic hypotheses, and in paper IV, the analyses were based on the reconstructed phy-logenies from paper III. The hands-on experience of reconstructing the phylogenetic hypotheses of the Rhamnus s.l. clade (paper III) was important to me in revealing the crucial issue of considering uncertainties in the topology of phylogenetic trees, when performing comparative studies (Donoghue and Ackerly 1996). Accordingly, the phy-logenetically independent contrasts (PICs) analyses of fruit and plant size relations in paper IV were based on a sample of phylogenetic hypotheses which we reconstructed ourselves (paper III).
Another important use of phylogenies is the possibility to polarize character evolution, that is to derive the direction of change in character evolution, by outgroup analyses. This is part of the independent contrast method used in paper IV, but was also employed in paper II so that we could divide and examine the sample based on whether fleshy fruits had evolved or been lost in the contrasted sister clades. Reconstructing phylogenetic hypotheses — The last four decades of phylogeny recon-struction rests on Hennig’s idea (1966) of using synapomorphies, that is, shared de-rived traits. Developing methods which transform this idea into quantitative analyses is a research field of its own. Methods based on the parsimony criterion were em-ployed when reconstructing the phylogenetic hypotheses on Rhamnus s.l. (paper III). Parsimony based methods evaluate the number of character state changes in a phy-logenetic hypotheses, and chooses the phylogenetic tree that is based on the smallest number of changes as the preferable phylogenetic hypothesis. When reconstructing phylogenies, one often ends up with several equally parsimonious, that is, equally preferable, phylogenetic hypotheses, meaning that other criteria have to be used to
20

Kjell Bolmgren
advocate any one of them as the most preferable hypotheses. The existence of several equally parsimonious phylogenetic hypotheses is an important issue to comparative biology (Donoghue and Ackerly 1996), which was considered in paper IV. In paper I and II, I generally used single phylogenetic (consensus) trees to generate the data points. This may be of particular concern for the data set on the temporal distribution of origin and losses of fleshy fruits in paper II. TIPS, PIMPs, and PICs — A major critique of traditional comparative studies have been the common use of species as statistically independent units. This methodology is nowadays often called TIPS, referring to the terminals or ’tips’ of a phylogeny. The major advantage of TIPS is that it does not require an explicit phylogenetic hypothesis, and thus allows for the inclusion of many more species. In this thesis (paper IV), I analysed size relations within the Rhamnus s.l. phylogeny using TIPS, sampling all species of the genera Frangula and Rhamnus s.str. The motive for this design was that the there exists no information on phylogenetic relationships with enough resolution. That is, as it is accepted to calculate phylogenetically uncorrected means when phy-logenetic information is lacking (Møller and Birkhead 1992; Wickman 1992), I ana-lyse phylogenetically uncorrected correlations when phylogenetic information is miss-ing.
The localization of phylogenetically independent events reduces the risk of confounding variables and allows for a conservative (safe) way of sampling statisti-cally independent units. When phylogenetic information was available, I applied three different comparative methods. First, the TIPS analyses in paper IV were developed by performing additional TIPS on monophyletic clades subsampled from the recon-structed phylogenies of Rhamnus s.l. Second, the analyses in paper I and II were based on phylogenetically independent matched pairs (PIMPs)2. A phylogenetically inde-pendent matched pair comprises two monophyletic sister clades separated by a phy-logenetic change in the classifying variable. PIMPs thereby only requires localization of the phylogenetic node where the trait change occurred, and knowledge about which taxa that belong to the contrasted sister clades. Quantitative values compared between the sister clades are derived without considering phylogenetic relationships within
2 I would like to dedicate this thesis to the anonymous student, who originally invented the phylogenetically independent matched pairs method; ”The suggestion that pairs of closely related organisms could be used in a way that avoided the need to know the full phylogeny was made by a student during discussion following my seminar at the Department of Genetics, University College, London. Unfortunately, I have been unable to discover her name.” (Felsenstein 1985, p. 15)
21

Adaptation and constraint in the plant reproductive phase
each sister clade (Felsenstein 1985; Møller and Birkhead 1992; Wickman 1992). In paper I, PIMPs were sampled when we could locate change to wind pollination in recently published cladograms, and in paper II, gaining or loosing fleshy fruit type was the sampling criteria. Using PIMPs one can analyse associations between characteris-tics. By adding an outgroup to the PIMPs in paper II, we analysed correlated change in fruit and habitat type. This design is related to the method of phylogenetically inde-pendent contrasts (PICs), which is the established method for analysing correlated change within phylogenies (Mazer 1998).
I employed PICs when analysing allometric effects on fruit size evolution in Rhamnus s.l. (paper IV). PICs analyses consider the difference in character states, that is, the contrast, between two related taxa as independent of previous evolution, and thus as independent in the statistical sense. Analysing PICs require detailed informa-tion on the phylogenetic relatedness for all taxa included and results therefore rests on the assumed phylogenetic hypotheses. In paper IV, I used a software package which allowed for the simultaneous analyses of several phylogenetic hypotheses (Schwilk 2001; Schwilk and Ackerly 2001). PICs further assume that character change evolve as Brownian motions. From these assumptions, PICs analyses also derive and make use of internal (‘ancestral’) contrasts. Key innovations — We examined if biotic pollination (paper I) and fleshy fruits (paper II) are correlated with enhanced species richness, using the PIMPs sampled in each study (McKitrick 1993; Miles and Dunham 1993). This means that the analyses of key innovations are based on replicates; the phylogenetically independent origins of func-tionally analogous characters (anemophily and fleshy fruits, respectively).
Key innovations are generally considered at a global scale, that is, not consider-ing confounding variables. As the occurrence of wind pollination, as well as species richness, correlates with latitude (Regal 1982; Brown and Lomolino 1998), we con-trolled for biogeography when sampling our data set (paper I). The sampling criteria used, when controlling for biogeographic autocorrelations, was that each PIMP must consist of geographically overlapping sister clades. In paper II, we controlled for growth form when analysing the effect of fleshy fruits on species richness, as we were interested in the correlated evolution of fruit type and growth habitat.
Moreover, by adding outgroups to each PIMP in the data set of paper II, it was possible to separate between effects of the evolved trait and the effect of change per se. When two sister clades are contrasted, one may suggest that the clade with con-
22

Kjell Bolmgren
served ancestral states is being detained in an existing more occupied niche space, while the other clade, evolving a novel trait, is entering a new less occupied niche space. Therefore, their potential radiation may not necessarily be comparable, as change per se could be expected to enhance radiation. In paper II, this problem was approached by separate analyses of subsamples of the PIMPs according to the direc-tion of evolutionary fruit type change. Data collection All phylogenetic comparative analyses (paper I, II and IV) were based on literature and herbarium data, while paper V was based on a field study.
In paper I, species-specific flowering onset and duration data were compiled from floras (see paper I for references). Phenological spread, which was adopted as the measure of phenological niche displacement, was derived as the standard deviation of flowering onset for each sister clade. To examine whether floristic information on flowering time was biased by pollination syndrome, phenology data taken from a re-gional flora (Weimarck 1963) was compared with data taken from herbarium sheets collected in the same region. A paired t-test revealed no biases in phenology informa-tion from floras owing to pollination syndrome.
In paper IV, flowering times for species of Rhamnus s.l. were derived from herbarium collections. This method has recently been evaluated by K. Lönnberg and K. Bolmgren (unpublished data), who found equally strong correlations between field and herbarium data sets, as between different field data sets (e.g. P-M correlations; N=26; Rfield vs. herbarium = 0.96; Rfield 1 vs. field 2 = 0.94). The species sampled in paper IV originated from different areas in North America and Europe. Comparing phenology between distantly located species, means that flowering date is not advisable as a measure, as it is thought to be correlated with local conditions (May in Sweden is not phenologically equal to May in Spain.). Therefore, in addition to comparing dates, flowering dates were transformed to temperature sums (Tuhkanen 1980; Diekmann 1996). Temperature sum is a reliable predictor of flowering events in plants (Reader 1983; White 1995; Diekmann 1996). In paper I, where we used flowering date as a measure, these issues were considered by contrasting species taken from the same flora and thereby reducing geographical effects.
23

Adaptation and constraint in the plant reproductive phase
In paper II, which examined relationships between fruit type and habitat char-acteristics, all information on growth habitats were taken from floras (see paper II for references). Based on these habitat descriptions, we made a subjective classification of each habitat according to how the light conditions was perceived in the habitat (open, intermediate, or closed). Based on these classifications, a semi-quantitative value was derived for each species, and means for each sister clade, which were then contrasted between fruit types.
We also compiled a historical distribution of changes in fruit type (paper II), based on a time-calibrated angiosperm phylogeny (Wikström et al. 2001). The data taken from Wikström et al. differs from data revealed from the fossil record (Collinson et al. 1993).
In paper IV, size measures on plant height, leaf length, and fruit diameter were taken from floras and monographs. TIPS and PICs of allometric size correlations were analysed using phylogenetic hypotheses derived in paper III.
In paper V, we analysed relations between phenology and components of re-productive success (i.e., fruit set, seed size, fruit size, dispersal, germination, juvenile size, and juvenile survival). Weekly censuses were performed of phenological changes in a Frangula alnus population, from flowering, through fruit initiation, development, ripening, and dispersal for two years. In both years, a phenological experiment was carried out, where the flowering curve was manipulated to represent early, late, peak, and asynchronous flowering. During the fruit ripening phase, every second ripe fruit was measured and collected on both untreated and treated shoots. Seeds were counted and weighed and then used in greenhouse, common garden, and field experiments on germination, juvenile development, and juvenile survival. Thus, we were able to fol-low the fate of individual ovules from flowering to second year juveniles.
As some of the processes potentially involved in the evolution of phenological strategies may be density-dependent, we analysed both 1st-order linear and 2nd-order curvilinear relations between phenology and components of reproductive success in paper V.
24

Kjell Bolmgren
Results
Phenology — PIMPs were used to examine hypothesized adaptive associations be-tween phenology and pollination syndromes (paper I). Wind pollinated trees were found to flower earlier than their biotically pollinated sister clades, but no difference in flowering onset time was found when all life forms were analysed together. Flowering onset differed up to three months between sister clades. Biotic pollination, as com-pared to wind pollination, was associated with a larger phenological spread (Figure 2). Phenological spread, which was applied as an analogue to phenological character displacement, was measured as the standard deviation of flowering onset among spe-cies within each sister clade.
BIOTIC POLLINATIONPhenological spread (# days)
Phen
olog
ical
spr
ead
(# d
ays)
WIN
D P
OLL
INA
TIO
N
0
20
40
60
80
100
120
0 20 40 60 80 100 120
Figure 2. Phylogenetically independent contrasts of phenological spread for biotically versus wind pollinated sister clades. Phenological spread was measured as the standard deviation of flowering onset. (Paper I)
Flowering time, expressed as temperature sums at flowering mode date, was
correlated with seasonal length in an interspecific data set of Rhamnus s.l. species (Figure 3). No similar correlations were found in intraspecific analyses of Rhamnus cathartica and Frangula alnus populations (paper IV). The interspecific correlation between flowering time and seasonal length translated into relatively conserved flow-ering dates. Despite a four month difference in seasonal length between localities, flowering dates were assembled within three weeks (Figure 4).
25

Adaptation and constraint in the plant reproductive phase
(ADDT +5)Total annual temperature sum
Tem
pera
ture
sum
(TS2
)fo
r flo
wer
ing
mod
e da
te
200
400
600
800
1000
800 1600 2400 3200 4000
no winterdormancy period
Figure 3. Interspecific (closed symbols) and intraspecific (open symbols) regressions of flowering mode date, expressed as temperature sum, on seasonal length, expressed as total annual temperature sum (ADDT +5). TS 2 is the temperature sum for all days above +5° C up to the date of flowering mode. The solid line illustrates the significant correlation (lin. regr.; N=5, R2=0.85, p=0.03) found in the interspeci-fic analyses of species from localities with a winter dormancy period. = R. cathartica, = R. cathar-tica MEAN, = R. crocea, = F. alnus, = F. alnus MEAN, = F. purshiana, = F. californica, and = F. caroliniana. (Paper IV)
Antwerpen
Namur
Copenhagen
Ystad
Karlshamn
Stockholm
Örnsköldsvik
St Louis
Sawyer
Julian day
degr
ee C
elsiu
s
-15
-10
-5
0
5
10
15
20
25
30
-50 0 50 100 150 200 250 300 350 400
F. car
baselinetemperature
F. cal N
F. pur
F. aln
R. cat
Figure 4. Flowering mode date for the sampled species marked (arrows) on the daily mean tempera-ture curves for the locality of collection, illustrating the narrow range of flowering dates compared to the much larger variation in seasonal length. F. car = Frangula caroliniana, F. cal N = F. californica N, F. pur = F. purshiana, F. aln = F. alnus, and R. cat = Rhamnus cathartica. (Paper IV)
26
Kjell Bolmgren
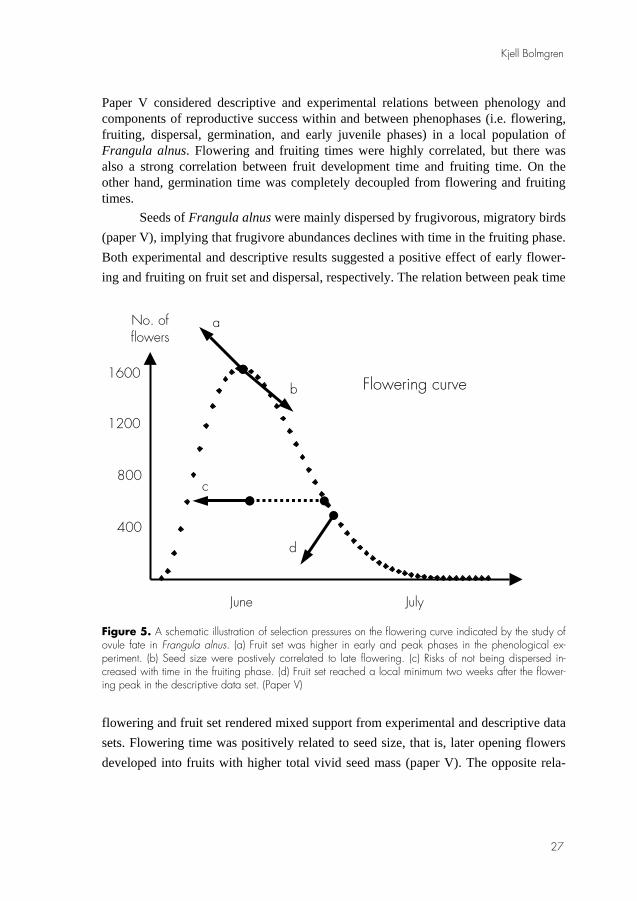
Paper V considered descriptive and experimental relations between phenology and components of reproductive success within and between phenophases (i.e. flowering, fruiting, dispersal, germination, and early juvenile phases) in a local population of Frangula alnus. Flowering and fruiting times were highly correlated, but there was also a strong correlation between fruit development time and fruiting time. On the other hand, germination time was completely decoupled from flowering and fruiting times.
Seeds of Frangula alnus were mainly dispersed by frugivorous, migratory birds (paper V), implying that frugivore abundances declines with time in the fruiting phase. Both experimental and descriptive results suggested a positive effect of early flower-ing and fruiting on fruit set and dispersal, respectively. The relation between peak time
No. of flowers
1600
1200
800
d
c
b
a
Flowering curve
400
July June
Figure 5. A schematic illustration of selection pressures on the flowering curve indicated by the study of ovule fate in Frangula alnus. (a) Fruit set was higher in early and peak phases in the phenological ex-periment. (b) Seed size were postively correlated to late flowering. (c) Risks of not being dispersed in-creased with time in the fruiting phase. (d) Fruit set reached a local minimum two weeks after the flower-ing peak in the descriptive data set. (Paper V)
flowering and fruit set rendered mixed support from experimental and descriptive data sets. Flowering time was positively related to seed size, that is, later opening flowers developed into fruits with higher total vivid seed mass (paper V). The opposite rela-
27

Adaptation and constraint in the plant reproductive phase
Clade age (Ma)
No.
phy
loge
netic
ally
inde
pend
ent
fruit
type
cha
nges
0
1
2
3
4
5
6
7
8
9
10
11
0-10 11-20 21-30 31-40 41-50 51-60 61-70 71-80 > 80
evolving fleshy fruit loosing fleshy fruit
Figure 6. Historical distribution of origins and losses of fleshy fruit types. (Paper II)
tions to flowering time, between fruit set and seed mass, were confirmed in regression analyses of 2nd-order polynomial (curvilinear) functions.
Seed size did affect juvenile size, but neither germination nor juvenile survival were related to seed size in both greenhouse, common garden, and field experiments (paper V).
Assuming that these different components of reproductive success (fruit set, seed mass, dispersal, germination, juvenile size, juvenile survival) translate into selec-tion pressures, Figure 5 illustrates their contributions to the selective regime moulding the flowering curve. Fleshy fruits — In paper II, we examined origins and losses of fleshy fruit types in PIMPs. The historical distribution of fleshy fruit origins was bimodal (Figure 6), peak-ing around the transitions from Paleocene to Eocene (~55 Ma) and Oligocene to Mio-cene (~25 Ma). Figure 6 was based on the oldest endpoints of the time interval sug-gested to encompass a fruit type change, and thus the distribution curve may be biased for older fruit type changes. Despite the modal peaks, Figure 6 suggests a continuous distribution of origins and losses of fleshy fruit types during the last 80 million years.
Evolutionary changes to fleshy fruit type were also correlated with habitat change (paper II), such that fleshy fruited clades colonized vegetation types that were
28

Kjell Bolmgren
1 case2 cases3 cases4 cases
Ancestral non-fleshy fruitClosedness index change
Clo
sedn
ess
inde
x ch
ange
Der
ived
fles
hy fr
uit
-1.25
-1.00
-0.75
-0.50
-0.25
0.00
0.25
0.50
0.75
1.00
1.25
-1.25 -1.00 -0.75 -0.50 -0.25 0.00 0.25 0.50 0.75 1.00 1.25
more closed
more open more closed
Figure 7. Changes in habitat light conditions, as compared to an outgroup, for clades evolving fleshy fruit type versus their sister clades retaining the ancestral non-fleshy fruit type. Habitats were classified as open, semi-open, and closed according to the prevailing vegetation type. (Paper II)
considered more closed than the habitats colonized by their dry-fruited sister clades (Figure 7).
Fleshy fruit size was studied in paper IV and V. Using the reconstructed phy-logenies from paper III, I analysed interspecific allometric relations in the Rhamnus s.l. clade, by TIPS and PICs. The independent contrast analyses (PICs) in paper IV suggested an allometric relationship between fleshy fruit size and plant mature height in all eight, topologically different, phylogenetic hypotheses of Rhamnus s.l. On the other hand, leaf length was not correlated to fruit size nor plant mature height in the PICs analyses. TIPS were analysed using different monophyletic subclades within Rhamnus s.l. Allometric correlations with fruit size revealed at the Rhamnus s.l. level, prevailed in Rhamnus s.str. but not in its sister genera Frangula (Figure 8).
In the population study of Frangula alnus (paper V), fruit diameter was posi-tively correlated with plant size (see also Table 1). Additional analyses (K. Bolmgren, unpublished data) suggested positive correlations for both fruit diameter and plant size with mean vivid seed mass per fruit (Table 1), and for individual fruits, fruit diameter was positively correlated with mean vivid seed mass and number of vivid seeds (Fig-ure 9).
29

Adaptation and constraint in the plant reproductive phase
Frangula
n.s.
N = 29
Eurhamnus-Pseudalataternus
R = 0.76**
N = 12
Alaternus
n.s
N = 3
Cervispina
R = 0.66**
N = 22
Rhamnus s.str.
R = 0.65***
N = 40
Rhamnus s.l.
R = 0.37**
N = 69
Figure 8. Fruit size and plant mature height correlations from the TIPS analyses, which were performed on different monophyletic clades within Rhamnus s.l. The hierarchical topology allowed for analyses at the supergeneric level (Rhamnus s.l.), the generic level (Rhamnus s.str. and Frangula), and the section level (Cervispina, Alaternus, and Eurhamnus-Pseuadalaternus). *P<0.05, **P<0.01, ***P<0.001. (Paper IV)
25 35
Frui
t dia
met
er (m
m)
7.5
9.0
10.
6.0
A.
123
15
Mean vivid seed mass per fruit (mg)
1 2
B.
3
No. of vivid seeds per fruit
Frui
t dia
met
er (m
m)
10
9
8
Figure 9. Fruit diameter as a function of (A) mean vivid seed mass per fruit (P-M correlations; N1 vivid seed
per fruit = 216, R1 vivid = 0.53, p<10-6; N2 vivid seeds per fruit = 335, R2 vivid = 0.59, p<10-6; N3 vivid seeds per fruit = 31, R3 vivid = 0.55, p=0.001) and (B) number of vivid seeds per fruit (ANOVA; df: 2, 581; p<10-6) in Frangula alnus 2000. (Paper V)
30

Kjell Bolmgren
Key innovations — No association between biotic pollination and higher species rich-ness was found, when compared to wind pollinated sister clades (paper I). On the other hand, evolution of fleshy fruit type was associated with more species-rich clades (pa-per II). However, this association was only valid when PIMPs characterized by woody growth forms were sampled (test 2a, Table 2), and particularly so when fleshy fruit types were the derived state (test 2b, Table 2). Among herbaceous plants, non-fleshy fruit types were associated with higher species richness (test 3a, Table 2), and the tendency was that this association was valid only when non-fleshy fruit types were the derived state (test 3c, Table 2) Change per se did not result in higher species richness (test 2c and 3b, Table 2). Table 1. Correlation coefficients for plant and fruit characteristics of Frangula alnus. *P<0.05, **P<0.01, ***P<0.001. (Paper V)
No. of annual shoots
per plant Fruit diameter
No. of vivid seeds
per fruit
Total vivid seed
mass per fruit
Mean vivid seed
mass per fruit
No. of annual shoots per plant
-
Fruit diameter 0.50* -
No. of vivid seeds per fruit
n.s. n.s. -
Total vivid seed mass per fruit
n.s. n.s. 0.76*** -
Mean vivid seed mass per fruit
0.53* 0.49* n.s. n.s. -
Table 2. Results of one-tailed sign tests of the correlation between extant species richness and fruit type. p is the cumulative p-value (Sokal and Rohlf 1995). (Paper II)
Test Growth forms Prediction p
1 All fleshy > non-fleshy 0.23
2a ‘Woody’ PICs fleshy > non-fleshy 0.01
2b Woody, with dry ancestor fleshy > non-fleshy 0.01
2c Woody, with fleshy ancestor fleshy > non-fleshy 0.34
3a ‘Herbaceous’ PICs non-fleshy > fleshy 0.02
3b Herbaceous, with dry ancestor non-fleshy > fleshy 0.19
3c Herbaceous, with fleshy ancestor non-fleshy > fleshy 0.06
31

Adaptation and constraint in the plant reproductive phase
Discussion
Phenology — In the comparison between biotically and wind pollinated plants (paper I), two anticipated phenological patterns appeared: biotic pollination was associated with wider phenological spread; and wind pollinated trees flowered earlier in the sea-son. As this study was based on PIMPs, the direction of phenological change is not known. That is, whether the biotically pollinated taxa have diverged more than random in flowering times or if the wind pollinated taxa have evolved a more narrow spread than is expected by random is not possible to conclude from this study.
Previous propositions have suggested that reproduction in wind pollinated populations is favoured by a more narrow flowering curve, as it amplifies the mass release of pollen characteristic of wind pollinated plants (Whitehead 1969; Faegri and van der Pijl 1979). If wind pollinated trees generally have a more narrow flowering curve (cf Rabinowitz et al. 1981) more species may be packed into a temporal niche space. Moreover, results by Linder and Midgley (1996) indicate that wind pollinated species may be less harmed by interspecific flowering overlap, which would further reduce phenological spread.
Wider phenological spread is one way to generate reproductive isolation be-tween related taxa, and thus part of the general conception that biotic pollination has enhanced speciation in angiosperms. Some initial suggestions of such temporal niche differentiation (e.g. Stiles 1977) were criticized on methodological grounds (for re-view, see Gotelli and Graves 1996). In paper I, phenological spread was measured as the standard deviation of flowering onset, which means that the patterns found may have been generated by microevolutionary processes, but also by ‘quantum leaps’. Plausible reasons for flowering time ‘quantum leaps’ could, for example, be changes in germination time from autumn to spring, changes in growth form, or changes in fruit type.
As with phenological spread, the study in paper I did not reveal which direc-tional change that lead to earlier flowering in wind pollinated trees. But, in contrast to the study of phenological spread, we do know the absolute seasonal position of the flowering times. Wind pollinated trees flowered early in the season which gives fur-ther support to the previously proposed adaptive and mechanistic interpretations of spring flowering times for wind pollinated trees (Whitehead 1969; Heinrich 1976).
32

Kjell Bolmgren
The comparison of Rhamnus s.l. species resulted in an anticipated correlation between seasonal length and flowering times (paper IV). Still, flowering dates were relatively conserved, gathered within a week despite a four month difference in sea-sonal lengths. Assuming that the temperature sum based correlations indeed are local adaptations, the conserved flowering dates are puzzling. Of course, it may just be a coincident, but considering the different selection pressures acting on flowering phenology in an insect-pollinated, bird-dispersed, temperate plant (cf Figure 5), stabi-lizing selection may also be an alternative. Most phenological strategies are probably moulded by several, differently directed selection pressures (Brody 1997; Pico and Retana 2000; Lacey et al. 2003). Apart from the effects of the mutualistic interactions (pollination and frugivory), phenology was also correlated to seed size in the local study of Frangula alnus (paper V). This seed size correlation suggests that recruitment processes may be part of reproductive phenology evolution in a more complex way than has generally been considered (Ollerton and Diaz 1999; Lacey et al. 2003).
Fleshy fruits — Functionally analogous fleshy ‘fruits’ may be considered the plesio-morphic fruit type for seed plants (Herrera 2002b), but among angiosperms several independent evolutions of fleshy fruits have occurred (e.g. paper II; Herrera 1989). The sudden rise in abundance of fleshy fruited taxa at the Late Cretaceous–Early Pa-leocene boundary has been an issue in itself (Tiffney 1984; Eriksson et al. 2000), but it has also been used as an explanation for conserved fleshy fruits (Tiffney 1984; Jor-dano 1987; Eriksson and Bremer 1992; Jordano 1995a; Eriksson et al. 2000). That is, particular conditions, for example a unique period of global warming and/or the radia-tion of passerines, changed the adaptive landscape profoundly, giving frugivorous seed dispersal fitness advantages during this particular period in angiosperm history. Paper II supported the correlation between fleshy fruit evolution and habitat change, but also a continuous distribution of fleshy fruit origins during the last 80 My. The historical distribution of origins and losses of fleshy fruit types was also bimodal with one peak more or less coinciding with the previously suggested peak in the Global warming period 50 Ma (Janis 2003). Previous studies have concluded that abundances of fleshy fruited plants peaked in Paleocene–Eocene (Eriksson et al. 2000), and paper II further suggest that independent origins of fleshy fruits peaked 55 Ma (Figure 6). This con-nection between abundance and evolutionary origins of fleshy fruited plants was fur-ther manifested by the correlation between fleshy fruit types and higher species rich-ness among woody plants (paper II). So, not only was the evolution of fleshy fruits
33

Adaptation and constraint in the plant reproductive phase
correlated with more closed vegetation types, fleshy fruits also seems to be a key in-novation under conditions related to these habitats (cf Tiffney and Mazer 1995).
The correlations between fleshy fruit types, diversification effects, and habitat types, suggest that selection for/on fleshy fruits may be stronger under these habitat conditions. Few studies exist explicitly analysing demographic effects of fleshy fruit characteristics and fleshy fruit dispersal in different habitats. Piper (1986) found mixed responses to different light conditions, and interpreted large seeded species as more shade tolerant. Herrera et al. (1994) found varying dispersal and recruitment patterns for the same species in different habitats, while Wenny and Levey (1998) is the only study I know of that have shown a positive effect of directed dispersal to favourable recruitment sites by a frugivorous bird. Increased seed size affects several recruitment processes positively, and may be comparatively more important in shady habitats (Eriksson and Jakobsson 1998; Ehrlén and Eriksson 2000; Leishman et al. 2000; Ja-kobsson and Eriksson 2002). At a general level, vertebrate dispersed fruits have larger seeds, though the seed size range and overlap with other fruit types is large (Hughes et al. 1994), and fleshy fruited plants have been associated with communities character-ized by higher stature (Willson et al. 1990; Guitian and Sanchez 1992; Jakobsson and Eriksson 2002). In Frangula alnus (Figure 9) fruit size was correlated both with seed size and seed number, implying that the selective regime of seed driven fruit size evo-lution may incorporate the frugivores. Frugivores drop larger fruits more often (Jor-dano 2000) and can change feeding behaviour from swallowing to pecking, and thus may not disperse seeds (Rey et al. 1997). Larger fruits also face a reduced frugivore guild (Wheelwright 1985). Moreover, the larger of the main seed dispersing frugivores of F. alnus fruits were the earliest autumn migrators potentially placing an additional pressure on the evolution of larger seeds. On the other hand, paper V did not suggest any selection pressure for larger seeds in F. alnus. Seed size in F. alnus affected juve-nile size, but neither seed size nor juvenile size had any general effects in the early recruitment phase (paper V).
Fruit size variation may also be an allometric effect. Herrera (2002a) found such relations between leaf and fruit sizes among Iberian fleshy fruited plants. Simi-larly, a positive correlation between plant height and fruit diameter was found among species in Rhamnus s.l. (paper IV) and within species in F. alnus (paper V), but no interspecific allometric relations to fruit size was found within Frangula (paper IV). Allometric relations may, apart from being a phylogenetic constraint (further discussed below), express simultaneous selection pressures due to abiotic conditions. For exam-
34

Kjell Bolmgren
ple, Herrera (2002a) suggested that water deficiency could lead to an evolutionary parallel reduction in both leaf size and fruit size.
Constraints — A key innovation is a feature that yields comparatively rapid radiation within a phylogenetic lineage, releasing the lineage from ancestral diversification constraints. To be able to test hypotheses on key innovations, the present thesis have adopted the additional criteria that key innovations must be phylogenetically repeated (Mitter et al. 1988; McKitrick 1993; Miles and Dunham 1993).
In contrast to previous studies (Eriksson and Bremer 1992; Ricklefs and Renner 1994; Dodd et al. 1999), biotic pollination was not associated with higher species richness when compared to wind pollinated clades. The main reason for this, perhaps, unexpected result may have been that the PIMPs sampled had to be geographically overlapping. Thereby the question at stake was merely if wind and biotic pollination differ in effects on diversification, and not whether biotic pollination is associated with angiosperm diversity. Considering the present results and that wind pollination be-comes more common when moving polewards (Regal 1982), the reason why biotic pollination may be associated with angiosperm radiation is that biotic pollination dominates in the species-rich tropical regions.
Fleshy fruits were not found to be a global key innovation (paper II), which is a result congruent with previous studies (Eriksson and Bremer 1992; Ricklefs and Renner 1994; Tiffney and Mazer 1995). The result changed, though, when growth form was controlled for, as fleshy fruits were associated with higher species richness in woody plants. Fleshy fruits are expensive fruits reducing the total number of seeds produced, and may therefore not pay-off for smaller (herbaceous) plants (Tiffney and Mazer 1995).
Paper I, II, and IV examined and found anticipated courses of evolution for flowering phenology in relation to pollination syndromes and seasonal conditions, and for fleshy fruits in relation to habitat change. However, these studies do not completely rule out the existence of phylogenetic constraints. They simply suggest that natural selection for adaptive change may overpower phylogenetic constraints to the extent that we can detect the anticipated outcomes. Based on the results from paper I, where phenology patterns supported adaptive hypotheses at a global scale, niche related phenology evolution must be considered more important than just a ‘fine-tuning’ proc-ess (Kochmer and Handel 1986, p. 319). Further, the differences in flowering times between sister clades indicate that previous interpretations of conserved flowering
35

Adaptation and constraint in the plant reproductive phase
time rank orders (Kochmer and Handel 1986) may have been unnecessarily biased towards phylogenetic constraints. Such patterns may equally well be interpreted as an outcome of phylogenetic niche conservatism (Harvey and Pagel 1991; Lord et al. 1995).
Ecotypic differentiation of flowering times to variation in seasonal conditions, for example, along a latitude or altitude transects, is a much supported pattern (e.g. Clausen et al. 1948; Reader 1983), and the adaptability of the seasonal sequence of phenological phases to make them fit into local conditions may be an important con-straint in migration processes. No such correlation between local seasonal length and flowering time, measured as temperature sums, was found in the two intraspecific data sets of Frangula alnus and Rhamnus cathartica (paper IV). This unexpected outcome may be interpreted as an indication of phylogenetic constraints, although such a con-clusion of phylogenetic constraints may be unwarranted if the preconditions (fitness differences, selection pressure) have not been examined (Antonovics and van Tienderen 1991). Similarly, Jordano (1995a) used a lack of an anticipated outcome to suggest that the evolution of fleshy fruits has been phylogenetically constrained. The underlying assumption in Jordano’s study was that a close match in the fruit–frugivore interaction would support an adaptive process. Thus, the lack of such a close match falsified the generality of tight interactions. Along the same line of reasoning, Jordano (1995a) suggested that frugivores are generally week selective agents on fleshy fruit characteristics, except for seed size. A lot of fruit character variation was attributed to phylogenetic affinities, thus confirming phylogenetic effects, but as long as no fitness differences and no constraining mechanisms are presented, general claims of phyloge-netically constrained character evolution must be considered with caution.
Allometry was found to be a potential mechanism underlying phylogenetically constrained fleshy fruit size evolution in Rhamnus s.l., as fruit size covaried with plant mature height (paper IV). Allometric effects have recently been proposed as an addi-tional explanation of fleshy fruit size variation (Herrera 2002a). As a phylogenetic constraint, the allometric mechanism within Rhamnus s.l. seems rather week, consider-ing that the allometric relation vanished within Frangula, while it prevailed in its mo-nophyletic sister clade Rhamnus s.str.
The final paper of this thesis (paper V) considered functional constraints, that is counteracting selection pressures, which Schwenk (1995) argued should not be re-garded as a phylogenetic constraint at all. When disentangling the different selective and constraining forces moulding the evolution of a character, a ‘liberal’ view of the
36

Kjell Bolmgren
constraint concept might be useful, at least when approaching the study system. Ex-perimental and descriptive examination of the relation between phenology and com-ponents of reproductive success indicated a complex pattern of selection pressures on the flowering curve of Frangula alnus (Figure 5). In general the early part of the curve favours fruit set and dispersal, while increasing seed size is favoured after the flower-ing peak. Thus, fruit set, and thereby seed number, and seed size were inversely corre-lated to flowering time in F. alnus. Even if seed size variation is just a result of the trade-off between seed number and seed size (Shipley and Dion 1992; Jakobsson and Eriksson 2000), selection for a phenological strategy leading to increased fruit set may be counteracted by selection for larger seeds. For example, within the population of F. alnus examined in paper V, a retained seed size range may be accomplished by a se-lection for a wider flowering curve. In the special case of F. alnus, however, seed size was only found to affect juvenile size in the recruitment phase, while neither germina-tion nor juvenile survival was affected by seed size under the conditions employed.
37

Adaptation and constraint in the plant reproductive phase
Literature cited Ackerly, D. D., and M. J. Donoghue. 1995. Phylogeny and ecology reconsidered.
Journal of Ecology 83:730-733. Antonovics, J., and P. H. van Tienderen. 1991. Ontoecogenophyloconstraints - the
chaos of constraint terminology. Trends in Ecology and Evolution 6:166-168.
Augspurger, C. K. 1981. Reproductive synchrony of a tropical shrub: experimental studies on effects of pollinators and seed predators on Hybanthus prunifolius (Violaceae). Ecology 62:775-788.
Bell, D. T., and L. J. Stephens. 1984. Seasonality and phenology of Kwongan spe-cies. Pp. 205-226 in J. S. Pate and J. S. Beard, eds. Kwongan: Plant Life of the Sandplain. University of Australia Press, Nedlands.
Bergström, J. 1993. Lamarckism. Nationalencyklopedin. Bra Böcker, Band 12, Hö-ganäs.
Blomberg, S. P., and T. Garland. 2002. Tempo and mode in evolution: phylogenetic inertia, adaptation and comparative methods. Journal of Evolutionary Biology 15:899-910.
Brody, A. K. 1997. Effects of pollinators, herbivores, and seed predators on flower-ing phenology. Ecology 78:1624-1631.
Brooks, D. R., and D. A. McLennan. 2002. The Nature of Diversity. An Evolution-ary Voyage of Discovery. University of Chicago Press, Chicago.
Brown, J. H., and M. V. Lomolino. 1998. Biogeography. Sinauer Associates, Inc., Sunderland.
Burt, D. B. 2001. Evolutionary stasis, constraint and other terminology describing evolutionary patterns. Biological Journal of the Linnean Society 72:509-517.
Carpenter, F. L. 1976. Plant-pollinator interactions in Hawaii: pollination energetics of Metrosideros collina (Myrtaceae). Ecology 57:1125-1144.
Clausen, J., D. D. Keck, and W. M. Hiesey. 1948. Experimental Studies on the Na-ture of Species. III. Environmental Responses of Climatic Races of Achillea. Carnegie Institution of Washington, Washington, D.C.
Collinson, M. E., M. C. Boulter, and P. L. Holmes. 1993. Magnoliophyta ('Angio-spermae'). Pp. 809-841 in M. J. Benton, ed. The Fossil Record 2. Chapman & Hall, London.
Crepet, W. L. 1984. Advanced (constant) insect pollination mechanisms: Patterns of evolution and implication vis-a-vis angiosperm diversity. Annals of the Missouri Botanical Garden 71:607-30.
Darwin, C. 1859. The Origin of Species by Means of Natural Selection. Penguin Books, London.
38

Kjell Bolmgren
Derrickson, E. M., and R. E. Ricklefs. 1988. Taxon-dependent diversification of life-history traits and the perception of phylogenetic constraints. Func-tional Ecology 2:417-423.
Diekmann, M. 1996. Relationship between flowering phenology of perennial herbs and meteorological data in deciduous forests of Sweden. Canadian Journal of Botany 74:528-537.
Dodd, M. E., J. Silvertown, and M. W. Chase. 1999. Phylogenetic analysis of trait evolution and species diversity variation among angiosperm families. Evolution 53:732-744.
Donoghue, M. J., and D. D. Ackerly. 1996. Phylogenetic uncertainties and sensitivity analyses in comparative biology. Philosophical Transactions of the Royal Society of London. Series B, Biolological Sciences 351:1241-1249.
Edwards, S. V., and S. Naeem. 1993. The phylogenetic component of cooperative breeding in perching birds. American Naturalist 141:754-789.
Ehrlén, J. 2003. Fitness components versus total demographic effects: evaluating herbivore impacts on a perennial herb. American Naturalist 162:796-810.
Ehrlén, J., and O. Eriksson. 2000. Dispersal limitation and patch occupancy. Ecology 81:1667-1674.
Eriksson, O. 1995. Asynchronous flowering reduces seed predation in the perennial forest herb Actaea spicata. Acta Oecologica 16:195-203.
Eriksson, O., and B. Bremer. 1992. Pollination systems, dispersal modes, life forms, and diversification rates in angiosperm families. Evolution 46:258-266.
Eriksson, O., and J. Ehrlén. 1991. Phenological variation in fruit characteristics in vertebrate-dispersed plants. Oecologia 86:463-470.
Eriksson, O., E. M. Friis, and P. Löfgren. 2000. Seed size, fruit size, and dispersal systems in angiosperms from the Early Cretaceous to the Late Terti-ary. American Naturalist 156:47-58.
Eriksson, O., and A. Jakobsson. 1998. Abundance, distribution and life histories of grassland plants: a comparative study of 81 species. Journal of Ecol-ogy 86:922-933.
Faegri, K., and L. van der Pijl. 1979. The Principles of Pollination Ecology. Perga-mon Press, Oxford.
Felsenstein, J. 1985. Phylogenies and the comparative method. American Naturalist 125:1-15.
Fenner, M. 1998. The phenology of growth and reproduction in plants. Perspectives in Plant Ecology, Evolution and Systematics 1:78-91.
Fuentes, M. 1992. Latitudinal and elevational variation in fruiting phenology among western European bird-dispersed plants. Ecography 15:177-83.
Gotelli, N. J., and G. R. Graves. 1996. The temporal niche. Pp. 95-111 in N. J. Gotelli and G. R. Graves, eds. Null Models in Ecology. Smithsonian Institution, Washington D.C.
39

Adaptation and constraint in the plant reproductive phase
Gould, S. J. 1989. A developmental constraint in Cerion, with commments on the definition and interpretation of constraint in evolution. Evolution 43:516-539.
Gould, S. J., and R. C. Lewontin. 1979. The spandrels of San Marco and the Panglos-sian paradigm: a critique of the adaptationist programme. Proceedings of the Royal Society of London. Series B, Biological sciences 205:581-598.
Grant, V. 1949. Pollination systems as isolating mechansims in angiosperms. Evolu-tion 3:82-97.
Grubb, P. J. 1977. The maintenance of species-richness in plant communities: the importance of the regeneration niche. Biological Review 52:107-145.
Guitian, J., and J. M. Sanchez. 1992. Seed dispersal spectra of plant-communities in the Iberian Peninsula. Vegetatio 98:157-164.
Harvey, P. H., and M. D. Pagel. 1991. The Comparative Method in Evolutionary Biology. Oxford University Press, Oxford.
Heinrich, B. 1976. Flowering phenologies: bog, woodland, and disturbed habitats. Ecology 57:890-899.
Hennig, W. 1966. Phylogenetic Systematics. University of Illinois Press, Urbana. Herrera, C. M. 1989. Seed dispersal by animals: a role in angiosperm diversification?
American Naturalist 133:309-322. Herrera, C. M. 1992. Historical effects and sorting processes as explanations for
contemporary ecological patterns: character syndromes in Mediterra-nean woody plants. American Naturalist 140:421-446.
Herrera, C. M. 1998. Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits: a 12-year study. Ecological Monographs 68:511-538.
Herrera, C. M. 2002a. Correlated evolution of fruit and leaf size in bird-dispersed plants: species-level variance in fruit traits explained a bit further? Oi-kos 97:426-432.
Herrera, C. M. 2002b. Seed dispersal by vertebrates. Pp. 185-208 in C. M. Herrera and O. Pellmyr, eds. Plant-Animal Interactions. An Evolutionary Ap-proach. Blackwell Science.
Herrera, C. M., P. Jordano, L. López-Soria, and J. A. Amat. 1994. Recruitment of a mast-fruiting, bird-dispersed tree: bridging frugivory activity and seedling establishment. Ecological Monographs 64:315-344.
Hodgson, J. G., and J. M. L. Mackey. 1986. The ecological specialization of dicote-lydonous families within a local flora: some factors constraining op-timization of seed size and their possible evolutionary significance. New Phytolologist 104:497-515.
Hughes, L., M. Dunlop, K. French, M. R. Leishman, B. Rice, L. Rodgerson, and M. Westoby. 1994. Predicting dispersal spectra - a minimal set of hy-potheses based on plant attributes. Journal of Ecology. 82:933-950.
Jakobsson, A., and O. Eriksson. 2000. A comparative study of seed number, seed size, seedling size and recruitment in grassland plants. Oikos 88:494-502.
40

Kjell Bolmgren
Jakobsson, A., and O. Eriksson. 2002. Seed size and frequency patterns of understory plants in Swedish deciduous forests. Ecoscience 9:74-78.
Janis, C. M. 2003. Tectonics, climate change, and the evolution of mammalian eco-systems. Pp. 319-338 in L. J. Rotschild and A. M. Lister, eds. Evolu-tion on Planet Earth. Academic Press, London.
Johnson, S. D. 1993. Climatic and phylogenetic determinants of flowering seasonal-ity in the Cape flora. Journal of Ecology. 81:567-572.
Jordano, P. 1987. Patterns of mutualistic interactions in pollination and seed disper-sal: connectance, dependence, asymmetries, and coevolution. Ameri-can Naturalist 129:657-677.
Jordano, P. 1995a. Angiosperm fleshy fruits and seed dispersers - a comparative analysis of adaptation and constraints in plant-animal interactions. American Naturalist 145:163-191.
Jordano, P. 1995b. Frugivore-mediated selection on fruit and seed size - birds and St Lucies cherry, Prunus mahaleb. Ecology 76:2627-2639.
Jordano, P. 2000. Fruits and frugivory. Pp. 125-165 in M. Fenner, ed. Seeds: the Ecology of Regeneration in Plant Communities. CABI, Wallingford.
Kochmer, J. P., and S. N. Handel. 1986. Constraints and competition in the evolution of flowering phenology. Ecological Monographs 56:303-325.
Lacey, E. P., D. A. Roach, D. Herr, S. Kincaid, and R. Perrott. 2003. Multigenera-tional effects of flowering and fruiting phenology in Plantago lanceo-lata. Ecology 84:2462-2475.
Leishman, M. R., I. J. Wright, A. T. Moles, and M. Westoby. 2000. The evolutionary ecology of seed size. Pp. 31-57 in M. Fenner, ed. Seeds: the Ecology of Regeneration in Plant Communities. CABI, Wallingford.
Linder, H. P., and J. M. Midgley. 1996. Anemophilous plants select pollen from their own species from the air. Oecologia 108:85-87.
Linnaeus, C. 1760. Politia naturae - naturens styrelseskick (translated to Swedish, 1978) in G. Broberg and A. Piltz, eds. Om jämvikten i naturen. Bok-förlaget Carmina, Stockholm.
Lord, J., M. Westoby, and M. Leishman. 1995. Seed size and phylogeny in 6 temper-ate floras - constraints, niche conservatism, and adaptation. American Naturalist 146:349-364.
Lord, J. M., A. S. Markey, and J. Marshall. 2002. Have frugivores influenced the evolution of fruit traits in New Zealand? Pp. 55-68 in D. J. Levey, W. R. Silva and M. Galetti, eds. Seed Dispersal and Frugivory: Ecology, Evolution and Conservation. CABI Publishing, Wallingford.
Mack, A. L. 1993. The sizes of vertebrate-dispersed fruits - a Neotropical-Paleotropical comparison. American Naturalist 142:840-856.
Mazer, S. J. 1998. Alternative approaches to the analysis of comparative data: com-pare and contrast. American Journal of Botany 85:1194-99.
Mazer, S. J., and N. T. Wheelwright. 1993. Fruit size and shape - allometry at differ-ent taxonomic levels in bird-dispersed plants. Evolutionary Ecology 7:556-575.
41

Adaptation and constraint in the plant reproductive phase
McKitrick, M. C. 1993. Phylogenetic constraint in evolutionary theory - has it any explanatory power. Annual Review of Ecology and Systematics 24:307-330.
Miles, D. B., and A. E. Dunham. 1993. Historical perspectives in ecology and evolu-tionary biology - the use of phylogenetic comparative analyses. An-nual Review of Ecology and Systematics 24:587-619.
Mitter, C., B. Farrell, and B. Wiegmann. 1988. The phylogenetic study of adaptive zones: has phytophagy promoted insect diversification? American Naturalist 132:107-128.
Møller, A. P., and T. R. Birkhead. 1992. A pairwise comparative method as illus-trated by copulation frequency in birds. American Naturalist 139:644-656.
Noma, N., and T. Yumoto. 1997. Fruiting phenology of animal-dispersed plants in response to winter migration of frugivores in a warm temperate forest on Yakushima Island, Japan. Ecological Research 12:119-129.
Oberrath, R., and K. Bohning-Gaese. 2002. Phenological adaptation of ant-dispersed plants to seasonal variation in ant activity. Ecology 83:1412-1420.
Ollerton, J., and A. Diaz. 1999. Evidence for stabilising selection acting on flowering time in Arum maculatum (Araceae): the influence of phylogeny on adaptation. Oecologia 119:340-348.
Ollerton, J., and A. J. Lack. 1992. Flowering phenology - an example of relaxation of natural selection. Trends in Ecology and Evolution 7:274-276.
Orzack, S. H., and E. Sober. 2001. Adaptation, phylogenetic inertia, and the method of controlled comparisons. Pp. 45-63 in S. H. Orzack and E. Sober, eds. Adaptationism and optimality. Cambridge University Press, Cambridge.
Pico, F. X., and J. Retana. 2000. Temporal variation in the female components of reproductive success over the extended flowering season of a Mediter-ranean perennial herb. Oikos 89:485-492.
Piper, J. K. 1986. Germination and growth of bird-dispersed plants: effects of seed size and light on seedling vigor and biomass allocation. American Journal of Botany 73:959-965.
Primack, R. B. 1980. Variations in the phenology of montane shrubs in New Zealand. Journal of Ecology. 68:849-62.
Primack, R. B. 1985. Patterns of flowering phenology in communities, populations, individuals and single flowers. Pp. 571-593 in J. E. White, ed. The Population Structure of Vegetation. Junk, Dordrecht.
Primack, R. B. 1987. Relationships among flowers, fruits, and seeds. Annual Review of Ecology and Systematics 18:409-430.
Rabinowitz, D., J. K. Rapp, V. L. Sork, B. J. Rathcke, G. A. Reese, and J. C. Weaver. 1981. Phenological properties of wind and insect pollinated prairie plants. Ecology 62:49-56.
Raikow, R. J. 1986. Why are there so many kinds of passerine birds? Systematic Zoology 35:255-259.
42

Kjell Bolmgren
Rathcke, B. J. 2003. Floral longevity and reproductive assurance: seasonal patterns and an experimental test with Kalmia latifolia (Ericaceae). American Journal of Botany 90:1328-1332.
Rathcke, B. J., and E. P. Lacey. 1985. Phenological patterns of terrestrial plants. Annual Review of Ecology and Systematics 16:179-214.
Reader, R. J. 1983. Using heat sum models to account for geographic variation in the floral phenology of two ericaceous shrubs. Journal of Biogeography 10:47-64.
Regal, P. J. 1982. Pollination by wind and animals: ecology of geographic patterns. Annual Review of Ecology and Systematics 13:497-524.
Rey, P. J., J. E. Gutiérrez, J. Alcántara, and F. Valera. 1997. Fruit size in wild olives: implications for avian dispersal. Functional Ecology 11:611-618.
Ricklefs, R. E., and S. S. Renner. 1994. Species richness within families of flowering plants. Evolution 48:1619-1636.
Robertson, C. 1895. The philosophy of flower seasons, and the phaenological rela-tions of the entomophilous flora and the anthophilous insect fauna. American Naturalist 29:97-117.
Robertson, C. 1924. Phenology of entomophilous flowers. Ecology 5:393-407. Sallabanks, R. 1993. Hierarchical mechanisms of fruit selection by an avian
frugivore. Ecology 74:1326-1336. Schwenk, K. 1995. A utilitarian approach to evolutionary constraint. Zoology
98:251-262. Schwilk, D. W. 2001. CACTUS 1.13. http://www.pricklysoft.org. Schwilk, D. W., and D. D. Ackerly. 2001. Flammability and serotiny as strategies:
correlated evolution in pines. Oikos 94:326-336. Shipley, B., and J. Dion. 1992. The allometry of seed production in herbaceous an-
giosperms. American Naturalist 139:467-83. Slade, N. A., J. S. Horton, and H. A. Mooney. 1975. Yearly variation in the phenol-
ogy of California annuals. American Midland Naturalist 94:209-214. Snow, D. W. 1965. A possible selective factor in the evolution of fruiting seasons in
tropical forests. Oikos 15:274-281. Sokal, R. R., and F. J. Rohlf. 1995. Biometry, 3rd ed. W. H. Freeman and Company,
New York. Stearns, S. C. 1986. Natural selection and fitness, adaptation and constraint. Pp. 23-
44 in D. M. Raup and D. Jablonski, eds. Patterns and Processes in the History of Life. Springer, New York.
Stearns, S. C. 1992. The evolution of life histories. Oxford University Press, Oxford. Stebbins, G. L. 1981. Why are there so many species of flowering plants? Bioscience
31:573-77 Stiles, F. G. 1977. Coadapted competitors: the flowering seasons of hummingbird-
pollinated plants in a tropical forest. Science 198:1177-1178. Thompson, J. N., and M. F. Willson. 1979. Evolution of temperate fruit/bird interac-
tions: phenological strategies. Evolution 33:973-982.
43

Adaptation and constraint in the plant reproductive phase
Thomson, J. D. 1980. Skewed flowering distributions and pollinator attraction. Ecol-ogy 61:572-579.
Tiffney, B. H. 1984. Seed size, dispersal syndromes, and the rise of the angiosperms: evidence and hypothesis. Annals of the Missouri Botanical Garden 71:551-576.
Tiffney, B. H., and S. J. Mazer. 1995. Angiosperm growth habit, dispersal and diver-sification reconsidered. Evolutionary Ecology 9:93-117.
Tuhkanen, S. 1980. Climatic parameters and indices in plant geography. Acta Phyto-geographica Suecica 67:1-105.
Wanntorp, H.-E. 1983. Historical constraints in adaptation theory: traits and non-traits. Oikos 41:157-160.
Waser, N. 1983. Competition for pollination and floral character differences among sympatric plant species: a review of evidence. Pp. 277-293 in C. S. Jones and R. J. Little, eds. Handbook of Experimental Pollination Bi-ology. Van Nostrand-Reinhold, New York.
Weimarck, H. 1963. Skånes flora. Bokförlaget Corona AB, Malmö. Wenny, D. G., and D. J. Levey. 1998. Directed seed dispersal by bellbirds in a tropi-
cal cloud forest. Proceedings of the National Academy of Sciences of the United States of America 95:6204-6207.
Verdu, M. 2002. Age at maturity and diversification in woody angiosperms. Evolu-tion 56:1352-1361.
Westoby, M., M. R. Leishman, and J. M. Lord. 1995. On misinterpreting the phy-logenetic correction. Journal of Ecology. 83:531-534.
Wheelwright, N. T. 1985. Fruit size, gape width, and the diets of fruit-eating birds. Ecology 66:808-818.
White, L. M. 1995. Predicting flowering of 130 plants at 8 locations with temperature and daylength. Journal of Range Management 48:108-114.
Whitehead, D. R. 1969. Wind pollination in the angiosperms: evolutionary and envi-ronmental considerations. Evolution 23:28-35.
Wickman, P.-O. 1992. Sexual selection and butterfly design - a comparative study. Evolution 46:1525-1536.
Wikström, N., V. Savolainen, and M. W. Chase. 2001. Evolution of the angiosperms: calibrating the family tree. Proceedings of the Royal Society of Lon-don Series B-Biological Sciences 268:2211-2220.
Willson, M. F., B. L. Rice, and M. Westoby. 1990. Seed dispersal spectra: a compari-son of temperate plant communities. Journal of Vegetation Science 1:547-562.
44

Kjell Bolmgren
Svensk sammanfattning Äpplet faller inte långt från trädet. Men varför? Den biologiska mångfalden präglas i stor utsträckning av fylogenetiskt bevarade karaktärsdrag; närbesläktade arter är lika. Det pågår en diskussion bland evolutionsbiologer om i vilken utsträckning denna kon-servatism är ett resultat av naturlig selektion eller av en begränsad anpassningsförmå-ga.
Denna avhandling diskuterar begreppet evolutionära begränsningar i relation till den reproduktiva fasen hos växter. I fokus ligger särskilt evolutionen av bärliknan-de (endozoochora) frukter respektive evolutionen av säsongsmässiga mönster (fenolo-gi) för blomning och fruktsättning. Avhandlingen är hierarkiskt organiserad så att olika delstudier gjordes på olika skalnivåer: fenologi- och fruktevolution analyserades för fröväxter respektive gömfröiga växter; inom brakvedssläktena Rhamnus och Frangula (Rhamnaceae); samt för en lokal population av brakved (Frangula alnus). Populationsstudien baserades på såväl experimentella som deskriptiva data, medan övriga studier i huvudsak genomfördes med fylogenetisk komparativ metodik baserade på litteraturdata. Som en del av de komparativa studierna rekonstruerades fylogenetis-ka hypoteser för Rhamnus s.l. utifrån DNA-sekvenser (ITS, trnL-F), vilka gav stöd för att Frangula och Rhamnus är monofyletiska systersläkten.
I en biogeografiskt kontrollerad fylogenetisk kontrastanalys upptäcktes inga skillnader i artrikedom mellan djur- och vindpollinerade växter. Bärliknande frukter verkar dock vara en betydelsefull karaktär (key innovation) i skogsmiljöer. För det första var fylogenetiska klader med bärliknande frukter mer artrika än systerklader med torra frukter. Dessutom var uppkomster av bärliknande frukt korrelerad till habi-tat med mer sluten vegetation.
En fylogenetisk kontrastanalys av allometriska effekter visade på en positiv korrelation mellan växtindividens och fruktens storlek inom Rhamnus s.l. Upprepade analyser av dessa allometriska samband i olika monofyletiska subklader inom Rham-nus s.l. indikerar dock att allometri, såsom evolutionär begränsning, är svag. I popula-tionsstudien av F. alnus var fruktstorlek positivt korrelerad till såväl fröantal som frövikt. En generell hypotes utifrån denna avhandling blir därför att frugivorer (de djur som äter bären och därmed sprider växtens frön) utövar ett starkt selektionstryck på växter som koloniserar ett habitat där fröstorlek är adaptivt, eftersom fruktstorlek på-verkar frugivorernas (ssk. fåglars) sätt att hantera frukten.
45

Adaptation and constraint in the plant reproductive phase
Reproduktiv fenologi var korrelerad till fruktsättning, fröspridning och fröstor-lek hos F. alnus. Detta stödjer tanken att evolutionen av blomnings- och fruktsätt-ningstider begränsas av avvägningar (trade-offs) och partiella beroenden mellan blom-ning-, fruktsättnings-, spridnings och rekryteringsfaserna. Inomartsvariation i blom-ningstid för F. alnus respektive getapel (Rhamnus cathartica) över en nordeuropeisk latitudinell transekt var inte korrelerad till säsongslängd, vilket antyder att utveckling-en av blomningstider är evolutionärt begränsad. Å andra sidan påvisades en sådan korrelation mellan blomningstid och säsongslängd i en mellanartsstudie inom Rham-nus s.l. Den fylogenetiska kontrastanalysen mellan djur- och vindpollinerade växter visade också på adaptiva skillnader i fenologi. Blomningstiderna inom en klad med biotiskt pollinerade arter var mer åtskilda än inom den vindpollinerade systerkladen, och den välkända iakttagelsen att vindpollinerade träd blommor tidigare på säsongen fick även stöd i ett fylogenetiskt komparativt perspektiv.
46





























