Embed Size (px)
Citation preview

Sveučilište u Zagrebu
Prirodoslovno-matematički fakultet
Biološki odsjek
Ada Zelen
Promjene u ekspresiji epitopa proteina GFAP i PSA-NCAM nakon
primjene kuprizona u miševa C57black/6
Diplomski rad
Zagreb, 2014.

ii
«Ovaj rad izrađen je na Zavodu za medicinsku biologiju Medicinskog fakulteta Sveučilišta u Osijeku, pod vodstvom Dr. sc. Marije Heffer, red. prof. sa Zavoda za
medicinsku biologiju Medicinskog fakulteta Sveučilišta u Osijeku. Rad je predan na ocjenu Biološkom odsjeku Prirodoslovno-matematičkog fakulteta Sveučilišta u
Zagrebu radi stjecanja zvanja magistra molekularne biologije.»

iii
ZAHVALA
Zahvaljujem mentorici prof. dr. sc. Mariji Heffer na srdačnom gostoprimstvu i
mnogobrojnim inspirativnim predavanjima kojima mi je probudila želju za
istraživanjem neistraženog. Ovim putem joj zahvaljujem i na velikoj pomoći i
stručnim savjetima kojima me usmjeravala tijekom mog boravka u Osijeku. Posebno
bih se htjela zahvaliti i asistentici Marti Balog koja mi je strpljivo i nesebično
pomagala u svakom koraku izrade ovog rada. Također zahvaljujem ostalim kolegama
iz laboratorija koji su me lijepo prihvatili te mi pomogli da se snađem i tih nekoliko
mjeseci osjećam kao dio tima. Zahvaljujući njima uvidjela sam kako ono naučeno
izgleda u praksi i tako zaokružila cijeli studij, a kao plod svega navedenoga proizašao
je ovaj rad. Zahvaljujem svim svojim prijateljima posebice Blanki i Vanini na
strpljenju i potpori koju su mi pružale sve ove godine.
Rad posvećujem svojoj mami koja mi je pružila najbolju moguću potporu u svakom
segmentu života i za to joj najveća hvala!

iv
TEMELJNA DOKUMENTACIJSKA KARTICA
Sveučilište u Zagrebu
Prirodoslovno-matematički fakultet
Biološki odsjek Diplomski rad
PROMJENE U IZRAŽAJU EPITOPA GFAP I PSA-NCAM NAKON PRIMJENE DEMIJELINIZIRAJUĆE TVARI KUPRIZONA KOD
MIŠEVA SOJA C57black/6
Ada Zelen
Rooseveltovtrg 6, 10000 Zagreb Tretiranjem miševa kuprizonom dolazi do masovne apoptoze te demijelinizacije neurona. Demijelinizaciju, uz propadanje oligodendrocita, prate pojava astroglije i mikroglije. Tijekom procesa demijelinizacije na aksolemi dolazi do ekspresije epitopa PSA-NCAM koji djeluje kao negativni regulator mijelinizacije, spriječavajući vezanje mijelin-stvarajućih stanica (oligodendrocita) na akson neurona. PSA-NCAM ujedino je i marker matičnih stanica i migracije. Demijelinizaciju, uz propadanje matičnih stanica prati i pojava astroglioze (porast broja reaktivnih astrocita). Astrociti mijenjaju morfologiju i ekspresiju proteina, između ostalog i proteina GFAP koji je klasični imunohistokemijski marker reaktivnih astrocita. Cilj ovog istraživanja bio je procijeniti obujam neurogeneze i gliogeneze do koje dolazi nakon primjene bakrenog kelatora kuprizona. Istraživanje je provedeno na miševima soja C57black/6 starosti 3 mjeseca, koji su hranjeni kuprizonom u trajanju od devet tjedana. Procjena demijelinizacijskog oštećenja i regeneracijskog kapaciteta načinjena je imunohistokemijskim označavanjem protutijelima GFAP i PSA-NCAM u regiji somatosenzornog korteksa. Iz ovog istraživanja možemo zaključiti da djelovanjem kuprizona dolazi do demijelinizacije neurona koja je praćena velikim izražajem PSA-NCAM epitopa i astroglioze. (34 stranice, 12 slika, 2 tablice, 75 literaturnih navoda, jezik izvornika: hrvatski) Rad je pohranjen u Središnjoj biološkoj knjižnici Ključne riječi: neurotoksin, demijelinizacija, remijelinizacija, neurogeneza,gliogeneza Voditelj: Dr. sc. Marija Heffer, red. prof. Suvoditelj: Dr. sc. Domagoj Đikić, izv. prof Ocjenitelji: Dr. sc. Maja Matulić, izv. prof., Dr. sc. Gordana Lacković-Venturin, red. prof. Rad prihvaćen: 12. rujna 2014.

v
BASIC DOCUMENTATION CARD
University of Zagreb
Faculty of Science
Department of Biology Graduation Thesis
CHANGES IN EXPRESSION OF GFAP AND PSA-NCAM EPITOPES AFTER APPLICATION OF DEMYELINATING SUBSTANCE
CUPRIZONE IN MOUSE STRAIN C57BLACK/6
Ada Zelen
Rooseveltovtrg 6, 10000 Zagreb, Croatia
Treatment of a mice with cuprizone leads to massive apoptosis and demyelination of neurons. Demyelination with declined number of oligodendrocytes is also accompanied by the emergence of astroglia and microglia. During the process of demyelination surface of axolema is expressing PSA-NCAM epitopes which act as a negative regulator of myelination, preventing the binding of myelin creating cells (oligodendrocytes) on axons of neurons. PSA-NCAM also is a marker of stem cells and migration. Demyelination, with a decadence of stem cells follows the appearance of astrogliosis (increased number of reactive astrocytes). Astrocytes change their morphology and expression of proteins, including GFAP, commonly used immunohistochemical marker of reactive astrocytes. The aim of this study was to estimate the scope of neurogenesis and gliogeneze that occurs after applying a copper chelator cuprizone. The study was conducted on mice strains C57black / 6, age 3 months, who were fed with cuprizone for a period of nine weeks. Valuation of damage and regeneration capacity was made by immunohistochemical labeling with antibodies GFAP and PSA-NCAM in the region of the somatosensory cortex. From this study it can be concluded that the cuprizone leads to demyelination of neurons and that demyelination is accompanied by overexpression of PSA-NCAM epitopes and astrogliosis.
(34 pages, 12 figures, 2 tables, 75 references, original in: Croatian)
Thesis deposited in the Central Biological library Key words: neurotoxin, demyelination, remyelination, neurogenesis, gliogenesis Supervisor: Dr. Marija Heffer, Prof. Cosupervisor: Dr. Domagoj Đikić, Assoc. Prof. Reviewers: Dr. Maja Matulić, Assoc. Prof., Dr. Gordana Lacković-Venturin, Prof. Thesis accepted: September 12, 2014

vi
SADRŽAJ
1. UVOD ............................................................................................................................. 1
1.1. Povijesni pregled ........................................................................................................................ 1
1.2. Adultna neurogeneza ................................................................................................................. 2
1.3. Gliogeneza i mijelinizacija ......................................................................................................... 2
1.4. Makroglija stanice ..................................................................................................................... 4
1.4.1. Astrociti ..................................................................................................................................... 4
1.4.2. Oligodendrociti .......................................................................................................................... 6
1.5. Mijelin i mijelinizacija ............................................................................................................... 7
1.5.1. Mijelin ....................................................................................................................................... 7
1.5.2. Mijelinizacija ............................................................................................................................. 7
1.6. GFAP (engl. glial fibrillary acidic protein) ................................................................................ 8
1.7. PSA-NCAM ................................................................................................................................ 9
1.8. Kuprizon, demijelinizacija i remijelinizacija .......................................................................... 10
1.9. Cilj istraživanja ........................................................................................................................ 13
1.9.1. Specifični ciljevi ...................................................................................................................... 13
2. MATERIJAL I METODE .......................................................................................... 14
2.1. Materijal .................................................................................................................................. 14
2.1.1. Miševi tretirani demijelinizacijskim spojevima ....................................................................... 14
2.1.2. Uvjeti uzgoja miševa ............................................................................................................... 14
2.1.3. Pokusne životinje ..................................................................................................................... 15
2.1.4. Protokol hranjenja C57BL/6 miševa kuprizonom.................................................................... 15
2.2. Imunodetekcija ........................................................................................................................ 16
2.2.1. Priprema tkiva za imunohistokemijsku analizu ....................................................................... 16
2.2.2. Imunohistikemija s protutijelima anti-PSA-NCAM I GFAP ................................................... 16
3. REZULTATI .................................................................................................................. 18
3.1. Imunohistokemijski prikaz aktiviranih astrocita kod miševa tretiranih kuprizonom ........... 18
3.2. Imunohistokemijski prikaz adultne neurogeneze kod miševa tretiranih kuprizonom ........... 21
4. RASPRAVA .................................................................................................................... 24
5. ZAKLJUČAK ................................................................................................................ 27
6. LITERATURA ............................................................................................................... 28

vii
POPIS KRATICA
ATP – adenozin trifosfat
BrdU – bromdeoksiuridin
CNS – središnji živčani sustav (prema engl. central nervous system)
GDNF – neutrofički čimbenik glija stanica (engl. glial derived neurotrophic factor)
GFAP – glijalni fibrilarni kiseli protein (engl. glial fibrillary acidic protein)
IF3 – intermedijarni filament tipa 3 (prema engl. intermediate filament type 3)
IGF3 – inzulinu sličan faktor rasta 1 (prema engl. insulin-like growth factor 1)
NCAM – neuronska adhezijska molekula (prema engl. neuronal cell adhesion
molecule)
NPC – neuronske progenitorske stanice (prema engl. neuronal precursor cells)
OPC – oligodendrocitne progenitorske stanice (prema engl. oligodendrocyte
precursor cell)
PNS – periferni živčani sustav (prema engl. peripheral nervous system)
PSA – polisijalinska kiselina (prema engl. polysialic acid)
TNF-α – čimbenik tumorske nekroze alfa (prema engl. tumor necrosis factor alpha)

1
1. Uvod
1.1. Povijesni pregled
Sve do prije nekoliko desetljeća, smatralo se da u mozgu kralježnjaka prestaje
stvaranje novih neurona do rođenja ili odmah nakon njega (Zupanc i sur. 2006). Prva
istraživanja koja su ukazivala na adultnu neurogenezu započela su 1960.-tih godina
pod vodstvom Josepha Altmana. Koristeći radioaktivni 3H-timidin prvi je ukazao na
prisutnost neurogeneze kod adultnih štakora (Altman i Das, 1965; Altman, 1969).
Njegova metoda uključivala je intravenozno ubrizgavanje radioktivno obilježenog
timidina, nukleozida kojeg stanice koriste pri sintezi DNA u procesu diobe.
Radioaktivno obilježene stanice u početku bile su identificirane pomoću elektronskog
mikroskopa te kasnije radioautografijom (Kaplan, 1981; Kaplan i Hinds, 1977;
Kaplan i Bell, 1984). Paralelno s njegovim istraživanjima objavljena je studija u kojoj
je, korištenjem njegove metode detekcije, opovrgnuta pojava neurogeneze kod
adultnih majmuna te su njegova istraživanja otišla u zaborav čitavo desetljeće (Rakic,
1985).
Koristeći Altmanovu metodu detekcije, 1980.-tih godina Nottebohm objavljuje niz
znanstvenih studija kojima potvrđuje znatan broj novonastalih neurona kod adultnih
ptica, tj. adultnu neurogenezu (Goldman i Nottebohm, 1983). 1990.-tih godina
metodu detekcije pomoću 3H-timidina zamjenjuje metoda detekcije s
bromodeoksiuridinom (BrdU). BrdU također se ugrađuje u novosintetiziranu DNA, a
obilježeni neuroni detektiraju se pomoću imunohistokemije (Kuhn i sur. 1996).
Prednost ove metode je mogućnost kombinacije s drugim metodama detekcije, te se
novosintetizirane stanice uz pomoć specifičnih neuronskih markera mogu u
potpunosti identificirati kod većine sisavaca ukljućujući i ljudi (Eriksson i sur. 1998).

2
1.2. Adultna neurogeneza
Među skupinama kralježnjaka postoje razlike u područjima odraslog mozga u
kojima se događa neurogeneza odnosno proliferacija, migracija i diferencijacija
neurona. Kod sisavaca se adultna neurogeneza odvija samo u specifičnim područjima
mozga i te regije čuvaju visoku razinu izražaja polisijalinske kiseline (PSA).
Spomenuta su područja hipotalamus, hipokamus i olfaktorni bulbusi, opisana kao
mjesta velike plastičnosti (Gascon i sur, 2007). U olfaktorni bulbus nezreli neuroni
složeni poput lanca gusto poredanih stanica migriraju iz subventrikularne zone putem
nekoliko milimetara dugog tzv. rostralnog migratornog puta (Altman 1969; Luskin,
1993; Curtis i sur., 2007). Iz subgranularne zone girus dentatusa nove stanice
migriraju u zrnati sloj hipokampusa, odnosno Amonovog roga (Altman i Das, 1965;
Eriksson i sur., 1998). Te se stanice razvijaju u zrele granularne neurone (Zupanc,
2008) i uklapaju se u funkcionalne neuronske krugove. Broj novonastalih neurona
jako je malen pogotovo u usporedbi s ukupnim brojem stanica u mozgu (Zupanc,
2006).
Mnoštvo današnjih istraživanja ukazuje na to da nekoliko vrsta oštećenja
moždanog tkiva, kao što su moždani udar te oštećenja izazvana neurotoksinima, mogu
povećati razinu neurogeneze unutar onih područja adultnog mozga gdje je i inače
prisutna (Szele i Chesselet 1996; Willis i sur. 1976; Calzà i sur. 1998).
1.3. Gliogeneza i mijelinizacija
Središnji živčani sustav sastoji se od dvije temeljne vrste stanica: neurona i glije.
Glija stanice su mnogobrojnije od neurona, te čine gotovo 50% ukupnog volumena
moždanog tkiva. Za razliku od neurona, glija stanice nemaju aksone, ne stvaraju
akcijske potencijale te nemaju natrijske kanale regulirane naponom, već samo kanale
za prolaz kalijevih iona (Judaš i Kostović, 1997). Glija stanice nastaju čitavog života,
iz multipotentnih neuralnih matičnih stanica (eng. multipotent neural progenitor cells)
procesom gliogeneze (Tramotin i sur. 2003).

3
Glija stanice središnjeg živčanog sustava dijelimo na dvije glavne skupine (Judaš i
Kostović, 1997):
a) MAKROGLIJA
• astrociti
• oligodendrociti
Razvijaju se od ektoderma tj. neuroepitelnih stanica proliferacijskih zona stijenke
neuralne cijevi kao i neuroni (Judaš i Kostović, 1997).
b) MIKROGLIJA
Razvijaju se od mezoderma, a u moždano tkivo ulaze u ranom fetalnom razdoblju,
prateći urastajuće krvne žile (Judaš i Kostović, 1997). Stanice mikroglija imaju
oskudnu citoplazmu, razgranate nastavke s trnastim izdancima te su mnogobrojnije u
sivoj tvari mozga. Za razliku od tzv. „mirujućih“ mikroglija (mikroglija u zdravom
odraslom mozgu) koje predstavlju stalnoprisutne makrofage mozga, „aktivirane“
mikroglija vrše ulogu fagocitoze kod upalnih ili degenerativnih bolesti. Tijekom
upalnog procesa „mirujuća“ mikroglija se aktivira te migrira prema mjestu lezije gdje
se pretvara u fagocitnu stanicu. Nakon što se napuni debrisom, postane znatno
krupnija i citoplazma joj se ispuni brojnim vakuolama te poprimi mrežast izgled
(Judaš i Kostović, 1997).
c) OSTALE GLIJA STANICE
• Ependimske glija stanice, Bergmannova glija, Mullerova glija,
Radijalna glija, Schwannove stanice, Satelitne stanice
Schwannove stanice i satelitne stanice perifernih živčanih ganglija razvijaju se od
neuralnog grebena (Judaš i Kostović, 1997).

4
1.4. Makroglija stanice
1.4.1. Astrociti
Astrociti (eng. astrocytes) su karakteristične, zvjezdoliko oblikovane glija stanice
središnjeg živčanog sustava (CNS, prema engl. central nervous system) (Fiacco i sur.
2008), koje na temelju izgleda dijelimo u fibrozne i protoplazmatske (Judaš i
Kostović, 1997).
a) Fibrozni astrociti smješteni su poglavito u bijeloj tvari, imaju brojne
izdužene nastavke i sadrže mnogo gliofibrila.
b) Protoplazmatski astrociti imaju mnogo razgranatije, kraće nastavke, te ih
prvenstveno nalazimo u sivoj stvari CNS (Judaš i Kostović, 1997).
Slika 1. Protoplazmatski i fibrozni astrociti
One predstavljaju glavnu vrstu neuronskih stanica, odgovornih za održavanje
homeostaze mozga. Izražajem širokog spektra receptora, transportera te ionskih
kanala imaju iznimno važnu ulogu u modulaciji neuralne aktivnosti (Cornell-Bell i
sur. 1990; Wang i sur. 2006).
Izravno surađuju s neuronima na nekoliko razina ukljućujuć otpuštenje
neurotransmitera glutamata ili ATP-a na vezikularni, Ca2+ ovisan načinu (Santello i
Volterra, 2008).

5
Jedna je od uloga astroglije i biokemijska potpora endotelnih stanica koje stvaraju
krvno-moždanu barijeru (Fiacco i sur. 2008). Naime, nastavci i fibroznih i
protoplazmatskih astrocita završavaju lukovičastim proširenjima tzv. završnim
nožicama, kojima oblikuju granični sloj glije, ispod pije i oko krvnih žila, te na taj
način sudjeluju u „propusnosti“ krvno-moždane barijere (Judaš i Kostović, 2007).
Osim u održavanju krvno-moždane barijere njihova je uloga održavanje ionske
ekstracelularne razine. Naime, membrana glija stanica odabirno je propusna samo za
K+ ali ne i za Na+ ione (Çakιr i sur. 2007). To svojstvo omogućuje astrocitima da
djeluju kao „glijalni sustav za puferiranje koncentracije K+ iona u moždanom tkivu“.
Uz pomoć astroglije, kod pojačane neuronske aktivnosti suvišak K+ iona privremeno
se odstrani iz aktiviranog neuronskog žarišta sve dok se neuroni ne repolariziraju.
Ukoliko se koncentracija K+ iona poveća u većem volumenu tkiva, astrociti djeluju
kao „sifon kalijevih iona“ prebacujući preko svojih nožica višak K+ iz izvanstanične
tekućine mozga u krvotok (Judaš i Kostović, 2007).
Njihova uloga u CNS veoma je važna i uslijed ozlijeda. Na mjestu ozljede CNS
dolazi do nastanka „glijalnog ožiljka“ kojeg izgrađuju aktivirani astrociti (Rolls i sur.
2009). Oni otpuštaju mnogobrojne inhibitorne izvanstanične molekule koje
sprječavaju aksonalni rast te samim time i obnavljanje tkiva (Silver i Miller, 2004).
Slika 2. Međustanična mreža astrocita, neurona te endotelnih stanica

6
1.4.2. Oligodendrociti
Oligodendrociti (engl. oligodendrocytes) vrsta su glijalnih stanica CNS (Cameron-
Curry i sur. 1995). Za razliku od astrocita nemaju gliofibrile ni glikogena zrnca, te
sadrže velik broj mikrotubula (Judaš i Kostović, 1997).
Na temelju smještaja u određenom dijelu moždanog tkiva, dijelimo ih na :
• interfascikularne (smještene između snopića aksona, posebice u
bijeloj tvari)
• „perineuronske satelite“ (smještene uz tijela neurona) (Judaš i
Kostović, 1997)
Oligodendrociti dio su živčanog sustava i u uskom su kontaktnu s neuronima te im
poput ostalih neuroglija osiguravaju potporu (Wilkins i sur. 2003). Njihova je glavna
uloga oblikovanje mijelinskih ovojnica oko središnjih aksona. Za razliku od
perifernog živčanog sustava (PNS, prema engl. peripheral nervous system) u kojemu
Schwannova stanica sudjeluje u mijelinizaciji samo jednog perifernog aksona, svaki
oligodendrocit može sudjelovati u mijelinizaciji 30 do 40 asona CNS (Judaš i
Kostović, 1997)
Slika 3. Oligodendrocit i mijelinska ovojnica

7
Oligodendrociti nastaju iz oligodendrocitnih progenitorskih stanica (OPC, prema
engl. oligodendrocyte precursor cell) te se kao i astrociti, mogu obnavljati čitav život
(Levine i sur. 2001). Stanice subventrikularne zone migriraju i u sivu i u bijelu tvar
gdje se diferenciraju i sazrijevaju u oligodendroglije koje stvaraju mijelin. Dio tih
stanica odlazi u apoptozu, dok se dio uspije izdiferencirati (Menn i sur. 2006 ).
1.5. Mijelin i mijelinizacija
1.5.1. Mijelin
Mijelin je dielektrički materijal (Virchow, 1954), izgrađen od vode (40%) i suhe
tvari (60%). Suhu tvar mijelina čine lipidi (70 – 85%) među kojima su najzastupljeniji
kolesterol i galaktocerebrozid, te specifični proteini (15 – 30%) kao što su mijelin,
oligodendrocitni glikoprotein te mijelinski proteolipidi. Njegova je glavna uloga
smanjenje električnog kapaciteta duž stanične membrane aksona gdje služi kao
električni izolator u prijenosu impulsa duž neurona (Hartline, 2008).
1.5.2. Mijelinizacija
Mijelinizacija je proces stvaranja mijelinske ovojnice koja omata aksone neurona
središnjeg i perifernog živčanog sustava. Unutar središnjeg živčanog sustava,
mijelinsku ovojnicu izgrađuju oligodendrociti, dok u perifernom živčanom sustavu tu
ulogu imaju Schwanove stanice (Quarles i sur. 2006). Mijelinizirani aksoni izgledom
su bijele boje te predstavljaju bijelu tvar središnjeg živčanog sustava (Saher i sur.
2005).
Glavna je uloga mijelinskog omotača ubrzavanje prolaska impulsa duž
mijeliniziranog vlakna. Naime, mijelinska je ovojnica isprekidana, a na mjestima
prekida nalaze se Ranvierova suženja (zbog blagog podebljanja aksona njihov je drugi
uobičajen naziv Ranvijerovi čvorovi) na kojima je aksolema u izravnom dodiru s

8
izvanstaničnom tekućinom što je značajno za skokovito vođenje akcijskih potencijala
(Judaš i Kostović, 2007). Za razliku od nemijeliniziranih vlakana koji provode
impulse u valovima, kod mijeliniziranih vlakana akcijski potencijal „skače“od jednog
Ranvierovog suženja do drugog, ubrzavajuć prijenos impulsa (Quarles i sur. 2006).
Odsječci mijelina između dva Ranvierova suženja nazivaju se internodalnim
segmentima. Za razliku od perifernog živčanog sustava gdje svaki internodalni
segment izgrađuje jedna Schwannova stanica, te je uz jedan periferni akson potrebno
onoliko Schwannovih stanica koliko na aksonu ima internodalnih segmenata, u
središnjem živčanom sustavu jedan oligodendrocit sudjeluje u mijelinizacijij većeg
broja aksona (Quarles i sur. 2006).
1.6. GFAP (engl. glial fibrillary acidic protein)
GFAP (engl. glial fibrillary acidic protein) je protein porodice intermedijarnih
filamenata, kojeg je 1970.-tih godina otkrila skupina znanstvenika na čelu s
Lawrencome Federoffom (Fedoroff i Vernadakis, 1987).
Intermedijarni filamenti tipa 3 (IF3, prema engl. intermediate filaments), kojima
pripada i GFAP, polimeriziraju te najčešće stvaraju homopolimere koji su
karakteristični za manje diferencirane stanice kao što su glija stanice, neuronske
prekursorske stanice (NPC, prema engl. neuronal precursor cells) te zrele astrocite
(Reeves i sur. 1989). Unutar središnjeg živčanog sustava GFAP je najviše izražen na
membranama fibroznih i protoplazmatskih astrocita (Fedoroff i Vernadakis, 1987).
GFAP također može polimerizirati s drugim proteinima porodice IF3 te s
neurofilamentnim proteinima (NF-L) stvarajući heterodimere (Reeves i sur. 1990).
Iako njegova funkcija nije još u potpunosti otkrivena, vjeruje se da osim u
citoarhitekturi, ima važnu ulogu i u mnogim procesima središnjeg živčanog sustava
kao što su stanična komunikacija te funkcija krvno-moždane barijere (Tardy i sur.
1990).

9
Prilagođavajući citoskeletnu mrežu, GFAP ima važnu ulogu i u procesu mitoze.
Tijekom mitoze povećava se razina fosforiliranog GFAP kao i njegovog prostornog
„pomicanja“ na mjesto „udubljenja stanice“ stanice (Tardy i sur. 1990). Ova
specifična raspodijela GFAP, omogućuje mu pravilnu podijelu na stanice kćeri
tijekom diobe (Liedtke i sur. 1996). In vitro pokusima dokazano je da astrociti kojma
nedostaje GFAP ne stvaraju nastavke obično prisutne između neurona te da je GFAP
važan za stvaranje interakcije astrocit-neuron (Tardy i sur. 1990).
GFAP ima 8 različitih izoformi koje označavaju različite subpopulacije astrocita
kod ljudi i glodavaca. Ove izoforme uključuju: GFAP alfa, GFAP kappa, GFAP+1,
GFAP delta te je veoma česta njihova primjena u imunodetekciji astrocita
(Middeldorp i Hol, 2011).
1.7. PSA-NCAM
NCAM (prema engl. neuronal cell adhesion molecule) integrirani je membranski
protein porodice imunoglobulina (Ig) (Brummendorf i Rathjen, 1995) s jednim
polipeptidnim lancem (Rutishauser, 1984). S obzirom na relativnu molekularnu masu,
postoje tri glavne izoforme: NCAM-120, NCAM-140, NCAM-180. Izvanstanične
izoforme NCAM posjeduju dvije vrste imunoglobulinskih domena uključenih u
međusobno povezivanje (Cole i sur. 1986) te fibronektinske domene (FN III domene)
uključene u signalizaciju rasta neurona. Kada se međusobno vežu nastaje homofilno
vezanje, a kada se molekule NCAM vežu s drugim molekulama nastaje heterofilno
vezanje (Brummendorf i Rathjen, 1995). Heterofilno vezanje odnosi se na interakcije
s drugim proteinima Ig porodice te između molekule NCAM i specifičnih molekula
unutarstaničnog okoliša i izvanstaničnog matriksa.
Intracelularnim domenama NCAM veže se na različite specifične receptore važne
za stanično signaliziranje čime se nadzire stanično ponašanje kao što je metabolizam
stanice, diferencijacija, preživljavanje i proliferacija. Izvanstanična domena NCAM u
interakciji je s proteinima izvanstaničnog matriksa koji su također uključeni u
međustaničnu adheziju (GDNF protein, engl. glial derived neurotrophic factor)

10
(Gascon i sur. 2007). Zbog mogućnosti raznolikog vezanja, molekule NCAM imaju
višestruku ulogu tijekom razvoja (Rieger i sur. 2008), uključujući migraciju stanica,
proliferaciju i diferencijaciju (Crossin i Krushel, 2000). Osim embrionalne
neurogeneze, reguliraju i adultnu neurogenezu.
Funkcije NCAM modificiraju se određenim posttranslacijskim modifikacijama od
kojih je najvažnija glikosijalizacija. Molekula NCAM prolazi kroz opsežnu
glikosijalizaciju u endoplazmatskom retikulumu i Golgijevom aparatu te u tom
procesu dolazi do vezanja polisijalinske kiseline (PSA) (Nelson i sur. 1995).
Polisijalinska kiselina negativno je nabijeni lanac α-2-8-vezanih molekula sijalinskih
kiselina koji se može produžiti do dužine od 50 do 100 jedinica (Kiss i Rougon,
1997). Njenim reverzibilnim vezanjem na izvanstanične domene nastaje
postranslacijska modifikacija PSA-NCAM (Rieger i sur. 2008)
Vezanje PSA na NCAM može promijeniti funkciju molekula NCAM od adhezije
prema signalizaciji. Tijekom razvoja, anti-adhezijska funkcija PSA-NCAM
omogućava neurogenezu (Bayer i sur. 1982), migraciju neurona, (Ono i sur. 1994;
Vitry i sur. 2001) i aksonogenezu - stvaranje, rast i navođenje izdanaka aksona
(Williams, 2000). Izražaj PSA na NCAM čvrsto je reguliran i vrhunac izražaja odvija
se u ranom razvoju dok u kasnijoj fazi većina molekula NCAM u mozgu ne sadrži
PSA.
1.8. Kuprizon, demijelinizacija i remijelinizacija
Kuprizon, kelator iona već se 1960.-tih godina pokazao kao izvrsan neurotoksin za
indukciju apoptoze oligodendrocita te pojavu demijelinizacije (Skripuletz i sur. 2008).
Iako kuprizon nakon primjene dovodi do značajne redukcije koncentracije bakra u
serumu i mozgu, njegov naknadni dodatak prehrani ne dovodi do redukcije njegova
toksičnog djelovanja. Ta činjenica ukazuje na to da iako igra važnu ulogu,
deficijencija bakra nije glavni mehanizam toksičnog djelovanja kuprizona (Carlton,
1967). Ishrana većim količinama kuprizona dovodi do morfoloških promjena
mitohondrija u jetri i mozgu (Suzuki, 1969; Tandler i Hoppel, 1973; Venturini, 1973;

11
Wakabayashi i sur. 1975). Reducirane su aktivnosti mitohondrijskih enzima kao što
su monoamino oksidaze, citokrom oksidaze i superoksid dismutaze dok je aktivnost
sukcinat dehidrogenaze povećana (Venturini, 1973; Russanov i Ljutakova, 1980;
Ljutakova i Russanov, 1985; Zhang i sur. 2008). Kako su mitohondriji glavni izvor
ATP-a potrebnog za homeostazu kalcija i apoptozu, pretpostavlja se kako je promjena
mitohondrijske funkcije (tj. energetski metabolizam) glavni razlog stanične smrti
oligodendrocita (Kipp i sur. 2009). Ipak nam ta pretpostavka ne daje jasan odgovor
zbog čega su oligodendrociti posebno osjetljivi na kuprizon, s obzirom na to da ostale
stanice ostaju pošteđene (Skripuletz i sur. 2008).
Tijekom prošlosti vršila su se razna istraživanja toksičnosti kuprizona na mnogim
laboratorijskim životinjama, a miševi pokazuju posebnu sklonost razvijanju
demijelinizacije. Prvi laboratorijski miševi koji su se koristili u tu svrhu bili su Swiss
albino miševi (Carlton, 1966; Carlton, 1967; Kesterson i Carlton, 1970, 1971), no
poslijednjih nekoliko godina istraživanja remijelinizacije i demijelinizacije
uzrokovane kuprizonom vrše se na laboratorijskim miševima soja C57BL/6
(Hiremath i sur. 1998). Znanstvenik Hiremath sistematično je istraživao utjecaj
kuprizona na demijelinizaciju kod miševa soja C57BL/6 ovisno o dozama (0.1, 0.2,
0.3, 0.4, 0.5 i 0.6 %) i otkrio da je doza od 0.2% najviša prihvatljiva. Miševi hranjeni
kuprizonom koncentracije veće od 0.2% brzo su gubili na težini, postajali su
letargični te im je smrtnost bilaveća od 80%, dok kod miševa hranjenih dozama ispod
0.2%, demijelinizacija nije bila dovoljno značajna. Stoga su zaključili kako je idealna
ravnoteža između dovoljno intenzivne demijelinizacije i razmjerno niske toksičnosti
postignuta kod doze od 0.2%, koja se od tada najčešće koristi u istraživanjima na
miševima soja C57BL/6 (Hiremath i sur. 1998). Istraživanja provedena na ženskim i
muškim jedinkama soja C57BL/6 ukazala su na to da ne postoji spolna razlika u
remijelinizaciji i demijelinizaciji uzrokovanoj primjenom kuprizona (Taylor i sur.
2010), dok su studije dobnih razlika bile kontroverzne te su potrebna daljnja
istraživanja (Blakemore, 1974; Ludwin, 1978; remington i sur, 2007).
Iako točni mehanizmi djelovanja kuprizona nisu u potpunosti razjašnjeni,
histološke su analize otkrile selektivnu staničnu smrt oligodendrocita. Nakon 4 do 4,5
tjedana tretiranja kuprizonom, zreli oligodendrociti gotovo u potpunosti nestaju
(Morell i sur. 1998; Komoly, 2005; Hesse i sur. 2009). Tijekom apoptoze

12
oligodendrocita, velik broj proliferirajućih OPC stanica odlazi na mjesto masovne
apoptoze (corpus calosuma, gdje se diferenciraju u zrele oligodendrocite). Prva
pojava zrelih oligodendrocita javlja se već unutar 5 tjedana, dok broj zrelih
oligodendrocita u 6. tjednu doseže razinu >50% usporedno s netretiranom kontrolnom
skupinom (Gudi i sur. 2009). Ukoliko se ishrana s kuprizonom nastavi,
oligodendrociti poslije 6. tjedna ponovno podliježu apoptozi (Linder i sur. 2009). Te
spoznaje upućuju na to da prirodni ciklus regrutacije i diferencijacije
oligodendrocitnih prekursorskih stanica traje 5 tjedana nakon prve primjene kuprizona
te da on djeluje isključivo na „zrele“ oligodendrocite (Skripuletz i sur. 2008). Ukoliko
se kuprizon ukloni iz prehrane unutar 5. tjedna (kada dolazi do prve pojave zrelih
oligodendrocita nastalih iz OPC stanica) ujedino dolazi i do povećanja re-izražaja
mnogih mijelinskih proteina (poput MBP, CNPaze, PLP, MOG) što se očituje
spontanom remijelinizacijom (Baumann i Pham-Dinh, 2001; Maier i sur. 2008).
Uklanjanjem kuprizona iz prehrane poslije kronične demijelinizacije (ishrana
kuprizonom od 12-16 tjedana) također će dovesti do spontane remijelinizacije i
pojave oligodendrocita no za razliku od akutne remijelinizacije, pojava
oligodendrocita i ponovna remijelinizacija nastupaju kasnije (Skripuletz i sur. 2008;
Linder i sur. 2009).
Mikroglioza je uz apoptozu oligodendrocita jedna od glavnih značajki
demijelinizacije i remijelinizacije inducirane kuprizonom (McMahon i sur. 2001;
Remington i sur. 2007). Pojava aktiviranih mikroglija uočava se već nakon 2. tjedna
tretiranja miševa kuprizonom. Opseg mikroglioze raste tijekom demijelinizacije te
postiže svoj maksimum između 4. i 5. tjedna (Remington i sur. 2007; Gudi i sur.
2009). Daljnjom primjenom kuprizona, broj aktiviranih mikroglija već nakon 6.
tjedna značajno opada (Koutsoudaki i sur. 2010). Distribucija aktiviranih mikroglija
unutar korteksa nije homogena. U kortikalnim slojevima 1 – 4 tijekom čitave
demijelinizacije inducirane kuprizonom nailazimo na veoma mali broj aktiviranih
mikroglija. Distribucija aktiviranih mikroglija mijenja se iz tjedna u tjedan. Nakon 3.
tjedna indukcije demijelinizacije, najveći broj aktiviranih mikroglija nalazimo u
kortikalnom staničnom sloju 5, dok poslije 4. tjedna njihov broj raste u slojevima 5 i
6 (Skripuletz i sur. 2008). Porijeklo mikroglija stanica kod demijelinizacije inducirane
kuprizonom i dalje nije razjašnjeno. Postoje dvije najprihvatljivije pretpostavke koje
to pokušavaju razjasniti. Jedna je pretpostavka da većina mikroglija stanica regrutira

13
iz drugih dijelova mozga te lokalnom proliferacijom stvara aktivirane mikroglije
(Gudi i sur. 2009). Druga je pretpostavka da aktivirane mikroglija stanice nastaju
diferencijacijom cirkularnih monocita koji infiltracijom iz periferne krvi stižu na
mjesto leziije (McMahon i sur. 2001) . Kod inducirane demijelinizacije kuprizonom
ne dolazi do oštećenja krvno-moždane barijere te su monociti najvjerovatnije u
mogućnosti proći krvno-moždanu barijeru bez da ju oštete (Bakker i Ludwin, 1987;
Kondo i sur. 2001). Iako uloga astroglioze kod demijelinizacije i remijelinizacije
inducirane kuprizonom i dalje nije u potpunosti razjašnjena, novija istraživanja
ukazuju na to da osim fagocitozne uloge uklanjanja raspadnutog mijelina, astrociti
također utječu i na remijelinizaciju otpuštanjem raznih čimbenika rasta kao što su
IGF-1 (Mason i sur. 2001) te TNF-alfa (Arnet i sur. 2001).
1.9. Cilj istraživanja
Uz pomoć specifičnih protutijela cilj istraživanja je dokazati proces
demijelinizacije te prisutnost aktiviranih astroglija i neurogeneze kod miševa
tretiranih kuprizonom.
1.9.1. Specifični ciljevi
1. Utvrditi obujam neurogeneze i gliogeneze do kojega dolazi nakon
primjene bakrenog kelatora kuprizona.
2. Procjeniti demijelinizacijski i regeneracijski kapacitet u području
somatosenzornog korteksa.

14
2. Materijal i metode
2.1. Materijal
2.1.1. Miševi tretirani demijelinizacijskim spojevima
Bakar je esencijalni element u tragovima koji omogućuje funkcioniranje brojnih
metaloenzima kao što su: bakar-cink superoksid dizmutaza (CuZnSOD1) i
ceruloplazmin. Ukoliko se miševi divljeg tipa soja C57black/6 hrane hranom u prahu
u koju je umiješan kelator iona bakra kuprizon (N, N´-bis (cikloheksil-denamino)
etanediamid) u koncentraciji 0,2 % nastaje tkivna deficijencija bakra i
demijelinizacija. Oligodendrociti su posebno osjetljivi na male doze kuprizona dok
druge stanice ostaju pošteđene. Hranjenje miševa s kuprizonom koncentracija 0.2% i
više, dovodi do gotovo potpune demijelinizacije već nakon 6 tjedana. Tijekom
procesa demijelinizacije na aksolemi dolazi do izražaja epitopa PSA-NCAM
(polisijalinizirana neuronska adhezijska molekula) koji djeluje kao negativan
regulator mijelinizacije. Demijelinizaciju uz propadanje oligodendrocita prati i pojava
mikroglije i astroglioza (abnormalan rast broja astroglija stanica). Astroglioza
mijenja molekularni izražaj i morfologiju astrocita te pojačava izražaj mnogih
epitopa, uključujuć i GFAP. GFAP klasičan je marker reaktivne glioze te se koristi u
mnogim imunohistokemijskim istraživanjima.
2.1.2. Uvjeti uzgoja miševa
Životinje su bile pod nadzorom veterinara te su boravile u nastambi za životinje
Medicinskog fakulteta u Osijeku. Prostor je bio klimatiziran na temperaturu od
24,9°C, u kojoj se održavalo izmjenično razdoblje svjetlo-tama, 14 sati svjetla i 10
sati tame. Broj miševa u pojedinom kavezu bio je ograničena na najviše 5 životinja.

15
2.1.3. Pokusne životinje
Pokus se provodio na 6 miševa soja C57BL/6, starosti 3 mjeseca koji su kroz 9
tjedana hranjeni hranom u prahu u koju je umiješan kelator bakra, kuprizon [N, N´-bis
(cikloheksil-denamino) etanediamid] (Sigma-Aldrich St. Louis, MO, SAD) u
koncentraciji od 0,2 %. Kontrolnu skupinu činili su miševi istog soja koji su bili
podvrgnuti jednakim uvjetima kao i miševi tretirani kuprizonom.
2.1.4. Protokol hranjenja C57BL/6 miševa kuprizonom
Tablica 1. Ishrana miševa soja C57BL/6 kuprizonom
Datum Ishrana
14.01. – 02.02.2014. Ishrana s kuprizonom (konc. 0.2%)
03.02. – 05.02.2014. Isključena hrana s kuprizonom
06.02. – 11.02.2014. Ishrana s kuprizonom (konc. 0.1 %)
12.02. – 18.03.2014. Naizmjenična ishrana miševa hranom s
kuprizonom konc 0.1% (5 dana) i
hranom bez kuprizona (2 dana)
19.03. – 20.03.2014. Žrtvovanje kuprizonom tretiranih miševa
24.03.2014. Žrtvovanje WT miševa
* Dvadesetog dana isključena je hrana s 0,2 % kuprizonom i uvedena hrana s 0,1 %
kuprizonom kako bi se zadržala krivulja hipomijelinizacije od 33%

16
2.2. Imunodetekcija
2.2.1. Priprema tkiva za imunohistokemijsku analizu
Pomoću inhalacijskog anestetika sevofurana (Baxter, Deerfield, IL, USA),
životinje su prije žrtvovanja duboko anestezirane. Učinjena im je kardijalna perfuzija,
s 0,1 M otopinom PBS-a (fosfatni pufer, engl. phosphate buffer saline), a zatim s 4%-
tnim paraformaldehidom (Sigma-Aldrich, St. Louis, MO) otopljenim u 0,1 M otopini
PBS-a (podešenepH vrijednosti 7,4). Nakon perfuzije svi uzorci mozga fiksirani su
24h pri temperaturi od +4°C u 4%-tnom paraformaldehidu (Sigma-Aldrich, St. Louis,
MO) u 0,1 M PBS-u, pH 7,4.
Nakon fiksacije, uzorci su krioprotektirani. Krioprotekcijom uzorci se dehidriraju
kako bi se izbjegla gruba kristalizacija citoplazmatske vode čime bi se u procesu
smrzavanja moglo oštetiti tkivo. Krioprotekcija učinjena je na +4°C tako da su uzorci
uronjeni u otopini 10% saharoze u 0,1M PBS kroz 24 sata, a potom u 20%
puferiranoj saharozi 48 sati. Nakon kriopreotekcije uslijedilo je smrzavanje uzoraka
kroz nekoliko sekundi u podhlađenom isopentanu (Sigma-Aldrich, St.Louis, MO,
USA) na -80°C, te su se na navedenoj temperaturi uzorci čuvali do daljnje analize.
Uzorci mozgova namijenjeni imunohistokemijskoj metodi izrezani su na kriostatu
(Leica, CM3050S, Germany) u koronarnom i sagitalnom smjeru, na debljini od 35
μm. Rezovi su skupljani u označene polistirenske jažice napunjene 0,1M PBS-om .
Pomoću mišjeg atlasa („The Mouse Brain“) izolirani su rezovi koji su sadržavali
Barrel field područje te se na tim rezovima vršila imunohistokemija.
2.2.2. Imunohistikemija s protutijelima anti-PSA-NCAM I GFAP
Imunohistokemija je rađena na slobodno plutajućim rezovima koji su pretretirani u
1% otopini vodikovog peroksida (Kemika, Zagreb) u 0,1M PBS-u kako bi se uklonila
aktivnost endogenih peroksidaza.
Nespecifično vezanje protutijela blokirano je otopinom 1% goveđeg serumskog
albumina (Sigma-Aldrich, St.Louis, MO, USA) i 5% kozjeg seruma (Gibco,

17
Invitrogen Auckland, NZ) u 0,1M PBS-u tijekom 2 sata na +4°C uz neprekidno
treskanje. Inkubacija u primarnom protutijelu trajala je preko noći na +4°C uz
neprekidno treskanje.
Sva primarna protutijela pripremljena su u otopini za blokiranje, a korištena su u
slijedećim omjerima: anti-GFAP, 1:4000; anti-PSA-NCAM, 1:500. Protutijelo anti-
GFAP (DAKO, Z033Y, 00019620) prepoznaje aktivirane glija stanice, dok anti-
PSA-NCAM protutijelo (Chemicon, Temecula, CA, USA) prepoznaje polisijalinsku
kiselinu vezanu na NCAM molekulu (tehnički podatci dobiveni uz komercijalno
protutijelo).
Nakon inkubacije u primarnom protutijelu rezovi su tri puta isprani u pothlađenom
0,1M PBS-u te inkubirani u sekundarnom protutijelu kroz 4 sata na +4°C uz
neprekidno treskanje. Ovisno o primarnom protutijelu korištena su dva sekundarna
protutijela, biotinilirani kozji anti-zečji IgG (VectorLaboratories, Burlingame, CA,
USA) uz anti-GFAP i kozji anti-mišji IgM (Jackson Immunoresearch lab., West
Grove, PA, USA) uz anti-PSA-NCAM, oba u razrjeđenju 1:500. Sekundarno
protutijelo također je uklonjeno ispranjem tri puta po 10 minuta u pothlađenom 0,1M
PBS. Uzorci su potom prebačeni u tercijarni kompleks, Vector Elite kit (Vector
Laboratories, Burlingame, CA, USA), pripremljen u skladu s uputama proizvođača.
Tercijarni kompleks sastoji se od avidina i biotinilirane peroksidaze te se zbog toga
prije upotrebe treba prekonjugirati u otopini za blokiranje u trajanju od oko pola sata.
U tercijarnom protutijelu rezovi su inkubirani 2 sata na +4°C uz neprekidno treskanje.
Rezovi su ponovno isprani, a potom su razvijeni precipitirajućim supstratom Vector
DAB Substrate Kit (Vector Laboratories, Burlingame,CA, USA) pripremljen u skladu
s uputama proizvođača. Pozitivna reakcija uočava se kao smeđe do sivo obojenje na
mjestima gdje je došlo do vezanja primarnog protutijela.
Preparati su još jednom isprani s 0,1M PBS-om, a potom su iz destilirane vode
navučeni na silanizirana predmetna stakalca, osušeni, skenirani skenerom
(NikonScan) te slikani mikroskopom (Carl Zeiss, Germany). Slike su obrađene i
pripremljene za tisak programima Photoshop i Image J.

18
3. Rezultati
3.1. Imunohistokemijski prikaz aktiviranih astrocita kod miševa
tretiranih kuprizonom
Imunohistokemijom sa specifičnim protutijelom anti-GFAP prikazana je
raspodijela aktiviranih astroglija stanica unutar Barrel field područja miševa tretiranih
kuprizonom (Slika 4 i 5) te wt miševa (Slika 6 i 7). Zbog kvantifikacijskih rezultata,
imunohistokemija je učinjena na koronarnim i sagitalnim rezovima mozga.
A B
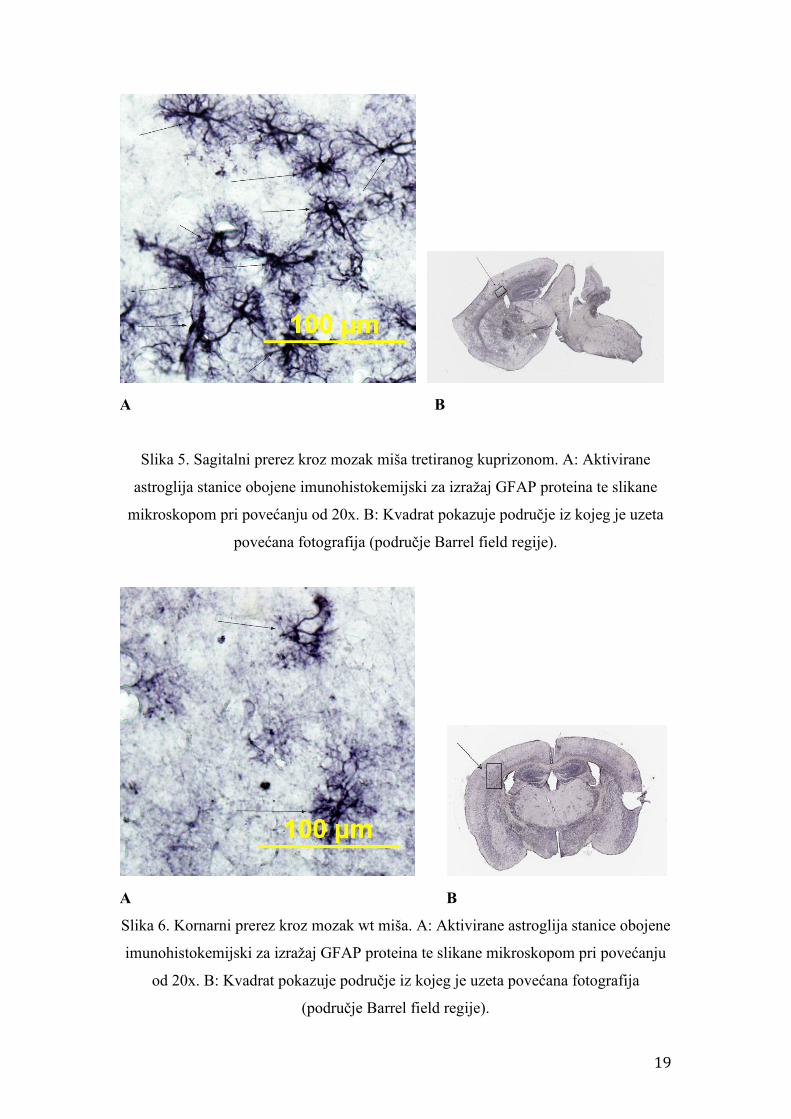
Slika 4. Koronarni prerez kroz mozak miša tretiranog kuprizonom. A: Aktivirane
astroglija stanice obojene imunohistokemijski za izražaj GFAP proteina te slikane
mikroskopom pri povećanju od 20x. B: Kvadrat pokazuje područje iz kojeg je uzeta
povećana fotografija (područje Barrel field regije).
19
A B
Slika 5. Sagitalni prerez kroz mozak miša tretiranog kuprizonom. A: Aktivirane
astroglija stanice obojene imunohistokemijski za izražaj GFAP proteina te slikane
mikroskopom pri povećanju od 20x. B: Kvadrat pokazuje područje iz kojeg je uzeta
povećana fotografija (područje Barrel field regije).
A B
Slika 6. Kornarni prerez kroz mozak wt miša. A: Aktivirane astroglija stanice obojene
imunohistokemijski za izražaj GFAP proteina te slikane mikroskopom pri povećanju
od 20x. B: Kvadrat pokazuje područje iz kojeg je uzeta povećana fotografija
(područje Barrel field regije).

20
A B
Slika 7. Sagitalni prerez kroz mozak wt miša. A: Aktivirane astroglija stanice obojene
imunohistokemijski za izražaj GFAP proteina te slikane mikroskopom pri povećanju
od 20x. B: Kvadrat pokazuje područje iz kojeg je uzeta povećana fotografija
(područje Barrel field regije).
Pomoću tri različita polja Barrel field područja koronarnih i sagitalnih rezova
napravljana je kvantifikacija aktiviranih astroglija stanica kod miševa tretiranih
kuprizonom i wt miševa (Tablica 2). Omjer aktiviranih astroglija kod kuprizon
tretiranih i wt miševa prikazan je na Slici 8.
Tablica 2. Broj aktiviranih astroglija stanica prisutnih unutar tri različita polja Barrel
field područja kod miševa tretiranih kuprizonom te kontrolne skupine
Koronarni presjek Sagitalni presjek
CUP 35 45
WT 16 7
CUP – skupina tretirana kuprizonom
WT – kontrolna skupina

21
Slika 8. Prikaz omjera aktiviranih astroglija stanica unutar Barrel field područja
kuprizon tretiranih miševa i kontrolne skupine (koronarnih i sagitalnih prereza)
3.2. Imunohistokemijski prikaz adultne neurogeneze kod miševa
tretiranih kuprizonom
Imunohistokemijom sa specifičnim protutijelom anti-PSA-NCAM prikazana je
raspodijela ovog epitopa unutar Barrel field područja miševa tretiranih kuprizonom
(Slika 9 i 10) te wt miševa (Slika 11 i 12). Imunohistokemija je rađena na koronarnim
i sagitalnim rezovima mozga.
WT
Kuprizon
0
5
10
15
20
25
30
35
40
45
Koronarni prerez
Sagitalni prerez
WT
Kuprizon
22
A B
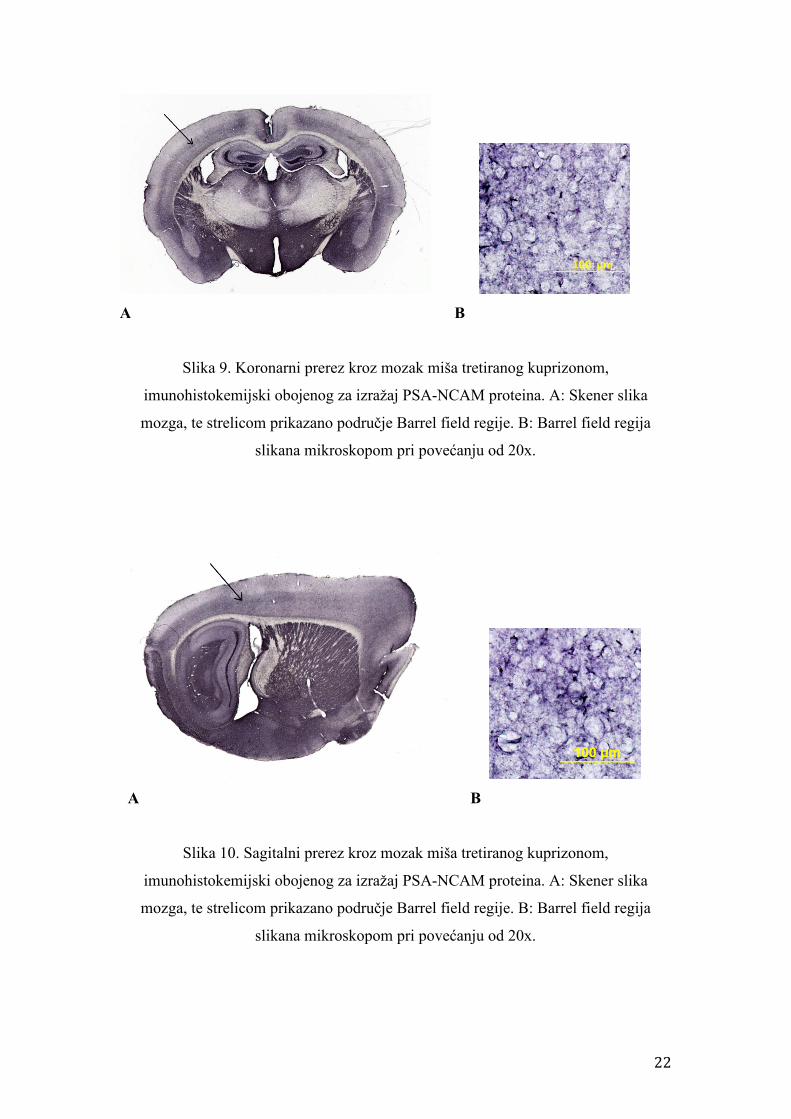
Slika 9. Koronarni prerez kroz mozak miša tretiranog kuprizonom,
imunohistokemijski obojenog za izražaj PSA-NCAM proteina. A: Skener slika
mozga, te strelicom prikazano područje Barrel field regije. B: Barrel field regija
slikana mikroskopom pri povećanju od 20x.
A B
Slika 10. Sagitalni prerez kroz mozak miša tretiranog kuprizonom,
imunohistokemijski obojenog za izražaj PSA-NCAM proteina. A: Skener slika
mozga, te strelicom prikazano područje Barrel field regije. B: Barrel field regija
slikana mikroskopom pri povećanju od 20x.

23
A B
Slika 11. Koronarni prerez kroz mozak wt miša imunohistokemijski obojenog za
izražaj PSA-NCAM protein. A: Skener slika mozga, te strelicom prikazano područje
Barrel field regije. B: Barrel field regija slikana mikroskopom pri povećanju od 20x.
A B
Slika 12. Sagitalni prerez kroz mozak wt miša imunohistokemijski obojenog za
izražaj PSA-NCAM protein. A: Skener slika mozga, te strelicom prikazano područje
Barrel field regije. B: Barrel field regija slikana mikroskopom pri povećanju od 20x.

24
4. Rasprava
Istraživanje opisuje izražaj proteina GFAP i PSA-NCAM unutar Barrel field
područja, primjenom imunohistokemijske metode na moždanom tkivu miševa
tretiranih kuprizonom. Tijekom procesa demijelinizacije na aksolemi se povećava
izražaj epitopa PSA-NCAM koji djeluje kao negativni regulator mijelinizacije,
spriječavajući vezanje oligodendrocita na akson neurona (Charles i sur. 2000).
Rezultati dobiveni istraživanjem jasno ukazuju na pojačani izražaj epitopa unutar
Barrel field područja kod miševa tretiranih kuprizonom.
PSA-NCAM ujedno je i marker matičnih stanica i migracije (Bonfanti i sur. 2006).
Obim neurogeneze dobiven u ovom radu kvalitativno smo prikazali Slikama
9,10,11,12 koje ukazuju na razliku u izražaju epitopa između wt miševa te miševa
tretiranih demijelinizirajućim neurotoksinom. Iz slika jasno se vidi migracija neurona
kod miševa tretiranih kuprizonom unutar Barrel field područja, dok je pojava
neurogeneze kod kontrolne skupine smanjena.
Demijelinizaciju, uz propadanje matičnih stanica, prati i pojava astroglioze (porast
broja reaktivnih astrocita). Astrociti mijenjaju morfologiju i izražaj proteina, između
ostalog i proteina GFAP koji je klasični imunohistokemijski marker reaktivnih
astrocita (Guo i sur. 2013).
Iz rezultata u ovom radu može se jasno uočiti promjena izražaja proteina GFAP tj.
broj aktiviranih astroglija stanica unutar Barrel field područja kod miševa tretiranih
kuprizonom i kontrolne skupine (Slike 4,5,6,7). Obojenje jasno prikazuje zvjezdaste
aktivirane astroglija stanice koje su brojnije kod skupine tretirane kuprizonom. Obim
aktiviranih astroglija stanica kvalitativno je prikazan na slikama u povećanju od 20x,
te kvantitativno u Tablici 2., koristeći broj aktiviranih astroglija stanica izbrojenih
unutar 3 različita polja Barrel field područja kontrolne skupine te skupine tretirane
kuprizonom.
Današnja istraživanja na tom području mnogobrojna su. Istraživanje provedeno
pod vodstvom Matsushima i sur. (2011) ukazuje na to da upravo model
demijelinizacije inducirane kuprizonom može odgovoriti zbog čega su baš

25
oligodendrociti „osjetljivi“ pri procesu demijelinizacije. Svojim radom dokazali su
kako su mitohondrijska oštećenja uzrok masovne apoptoze oligodendroglija stanica te
posljedične demijelinizacije. Osim toga dokazali su da tijekom procesa
demijelinizacije dolazi i do otpuštanja niza citokina i čimbenika rasta koji iniciraju
regrutaciju i proliferaciju mikroglija stanica (astrogliozu), dokazanu i u ovom radu.
Također smatraju da mikroglija stanice potiču astrocite na otpuštanje čimbenika koji
utječu na remijelinizaciju, tj. regrutaciju velikog broja progenitornih stanica koje će
kasnije djelovati na remijelinizaciju. Pojava regrutacije progenitornih stanica, tj.
neurogeneze također je dokazana i u ovom radu (imunohistokemijskim bojanjem s
anti-PSA-NCAM proteina).
Drugo istraživanje provedeno pod vodstvom Koutsoudaki i sur. (2010) ukazuje na
negativan utjecaj PSA-NCAM molekule unutar demijelinizirajućih područja kod
bolesnika s multiplom sklerozom. Iako se izražaj molekule PSA povezuje s mnogim
fundamentalnim procesima kao što su regeneracija, plastičnost te razvoj središnjeg
živčanog sustava (Rutishauseri sur.), na njegov negativan utjecaj na mijelinizaciju
također je ukazao članak i Charles i sur. (2007). Znanstveno je dokazan izražaj
molekula PSA-NCAM na demijeliniziranim aksonima neurona kod bolesnika
multiple skleroze (MS) što ukazuje na njenu inhibitornu ulogu u remijelinizaciji
(Charles i sur.).
Koutsoudaki i sur. (2010) istraživanje su vršili na St8SiaIV (polisialtransferaza)
deficijentim miševima te dokazali da su miševi koji su imali smanjen izražaj PSA-
NCAM pokazivali bržu remijelinizaciju poslije demijelinizacije inducirane toksinom.
Rezultati dobiveni u ovom radu također potvrđuju te spoznaje. Iz Slike 12 jasno se
uočava da kod miševa tretiranim kuprizonom dolazi do većeg izražaja proteina PSA-
NCAM u usporedbi s kontrolnom skupinom. Istraživanje opisano u ovom radu dio je
veće studije kojom se procjenjuje opseg demijelinizacijskog oštećenja i
regeneracijskog kapaciteta na miševima s isključenim genima za sintezu glikolipida
kako bi se spoznali molekularni mehanizmi uključeni u procese demijelinizacije i
remijelinizacije.
Velika prednost rada na modelima tretiranim kuprizonom, osim veoma lako
inducirane i uočljive demijelinizacije, jest i „zaobilaženje“ krvno-moždane barijere, te
taj pristup omogućuje istraživanje mehanizama demijelinizacije i remijelinizacije bez

26
utjecaja složenog imunološkog sustava (Skripuletz i sur. 2008). Iako se spoznaje
dobivene istraživanjem na tom modelu često koriste kao okosnica za otkrivanje
mehanizama demijelinizacije i remijelinizacije kod multiple skleroze (MS), valja
imati na umu kako nije moguće istraživati patofiziologiju ljudske bolesti kao što je
MS na jednom animalnom modelu. Daljnja istraživanja moraju se temeljiti na više
animalnih modela od kojih će se svaki koristiti za otkrivanje drugih mehanizama
demijelinizacije i remijelinizacije (Skripuletz, 2008).

27
5. Zaključak
1. Tretiranjem miševa soja C57black/6 kuprizonom, u trajanju od 9 tjedana nastaje
masovna apoptoza te demijelinizacija neurona u području somatosenzornog
korteksa.
2. Iz ovog i nekoliko prijašnjih istraživanja možemo zaključiti da tijekom procesa
demijelinizacije na aksolemi dolazi do pojačanog izražaja epitopa PSA-NCAM
koji djeluje kao negativni regulator mijelinizacije te ujedino predstavlja i marker
matičnih stanica i migracije.
3. Demijelinizaciju uz propadanje matičnih stanica prati i pojava astroglioze (porast
broja reaktivnih astrocita). Astrociti mijenjaju morfologiju i izražaj mnogih
proteina, između ostalog i proteina GFAP, koji služi kao imunomarker
aktiviranih astroglija.

28
6. Literatura
1. Altman J. 1969. Autoradiographic and histological studies of postnatal
neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with
special reference to persisting neurogenesis in the olfactory bulb. J. Comp.
Neurol. 137, 433–457.
2. Altman J., Das G. D. 1965. Autoradiographic and histological evidence of
postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 124, 319–335.
3. Arnett H. A., Mason J., Marino M., Suzuki K., Matsushima G. K., Ting J. P.
2001. TNF alpha promotes proliferation of oligodendrocyte progenitors and
remyelination. Nat. Neurosci. 4, 1116-1122.
4. Bakker D. A. and Ludwin S. K. 1987. Blood-brain barrier permeability during
Cuprizone-induced demyelination. Implications for the pathogenesis of immune-
mediated demyelinating diseases. J. Neurol. Sci. 78, 125-137.
5. Baumann N. and Pham-Dinh D. 2001. Biology of oligodendrocyte and myelin in
the mammalian central nervous system. Physiol. Rev. 81, 871-927
6. Bayer S., Yackel J., Puri P. 1982. Neurons in the rat dentate gyrus granular layer
substantially increase during juvenile and adult life. Science 216, 890-892.
7. Brümmendorf T., Rathjen F. 1995. Cell adhesion molecules 1: immunoglobulin
superfamily. Protein profile 2, 963-1108.
8. Çakιr T., Alsan S., Saybaşιlι H., Akιn A., Ülgen K. Ö. 2007. Reconstruction and
flux analysis of coupling between metabolic pathways of astrocytes and neurons:
application to cerebral hypoxia. Theoretical Biology and Medical Modelling4,
48.
9. Calzà L., Giardino L., Pozza M., Bettelli C., Micera A., Aloe L. 1998.
Proliferation and phenotype regulation in the subventricular zone during
experimental allergic encephalomyelitis: in vivo evidence of a role for nerve
growth factor. Proc. Natl. Acad. Sci (USA) 95, 3209–14.
10. Cameron-Curry P., Le Douarin N. M. 1995. Oligodendrocyte precursors originate
from both the dorsal and the ventral parts of the spinal cord. Neuron 15(6), 1299-
310.

29
11. Carlton W. W. 1966. Response of mice to the chelating agents sodium
diethyldithiocarbamate, alpha-benzoinoxime, and biscyclohexanone
oxaldihydrazone. Toxicol. Appl. Pharmacol. 8, 512-521.
12. Carlton W. W. 1967. Studies on the induction of hydrocephalus and spongy
degeneration by cuprizone feeding and attempts to antidote the toxicity. Life Sci.
6, 11-19
13. Cole G., Loewy A., Cross N., Akeson R., Glaser L. 1986. Topographic
localization of the heparin-binding domain of the neural cell adhesion molecule
n-cam. J cell biol 103, 1739-1744.
14. Cornell-Bell A. H., Finkbeiner S. M., Cooper M. S., Smith S. J. 1990. Glutamate
induces calcium waves in cultured astrocytes: long-range glial
signaling.Science 247, 470–473.
15. Crossin K. L., Krushel L. A. 2000. Cellular signaling by neural cell adhesion
molecules of the immunoglobulin superfamily. Dev. dyn. 218, 260-279.
16. Curtis M. A., Kam M., Nannmark U., Anderson M. F., Axell M. Z., Wikkelso C.,
Holtas S, van Roon-Mom W. M. C., Bjork-Eriksson T., Nordborg C., Frisen J.,
Dragunow M., Faull R. L. M., Eriksson P. S. 2007. Human neuroblasts migrate to
the olfactory bulb via a lateral ventricular extension. Science 315, 1243-1249.
17. dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor
18. Eriksson P. S., Perfilieva E., Björk-Eriksson T., Alborn A. M., Nordborg C.,
Peterson D. A., and Gage F. H. 1998. Neurogenesis in the adult human
hippocampus. Nat. Med.4, 1313–1317.
19. Fedoroff S., Vernadakis A., Cellular Neurobiology (1986): Astrocites ,
Development, Morphology, and Regional Specialization of Astrocytes. Academic
Press, Orlando
20. Fiacco T. A., Agulhon C., McCarthy K. D. 2008. Sorting out Astrocyte
Physiology from Pharmacology.Annu. Rev. Pharmacol. Toxicol. 49 , 151–74.
21. Gascon E., Vutskits L., Kiss J. Z. 2007. Polysialic acid-neural cell adhesion
molecule in brain plasticity: from synapses to integration of new neurons. Brain
Res. Rev. 56, 101-118.
22. Gascon E., Vutskits L., Kiss J. Z. 2007. Polysialic acid-neural cell adhesion
molecule in brain plasticity: from synapses to integration of new neurons. Brain
res. Rev. 56, 101-118.

30
23. Goldman S. A., Nottebohm F. 1983. Neuronal production, migration, and
differentiation in a vocal control nucleus of the adult female canary brain. Proc.
Natl Acad. Sci. USA 80, 2390–2394.
24. Gudi V., Moharregh-Khiabani D., Skripuletz T., Koutsoudaki P. N., Kotsiari A.,
Skuljec J., Trebst C., Stangel M. 2009. Regional differences between grey and
white matter in cuprizone induced demyelination. Brain Res. 1283, 127-138.
25. Hiremath M. M., Saito Y., Knapp G. W., Ting J. P., Suzuki K., Matsushima G.
K. 1998. Microglial/macrophage accumulation during cuprizone-induced
demyelination in C57BL/6 mice. J.Neuroimmunol. 92, 38-49
26. Judaš, M., Kostović, I. (1997) : TEMELJI NEUROZNANOSTI. 1. izd. MD,
Zagreb.
27. Kaplan M. S. 1981. Neurogenesis in the 3-month-old rat visual cortex. J. Comp.
Neurol. 195, 323–338.
28. Kaplan M. S., Bell D. H. 1984. Mitotic neuroblasts in the 9-day-old and 11-
month-old rodent hippocampus. J. Neurosci. 4, 1429–1441.
29. Kaplan M. S., Hinds J. W. 1977. Neurogenesis in the adult rat: electron
microscopic analysis of light radioautographs. Science 197, 1092–1094.
30. Kesterson J. W. and Carlton W. W. 1970. Aqueductal stenosis as the cause of
hydrocephalus in mice fed the substituted hydrazine, cuprizone. Exp. Mol.
Pathol. 13, 281-294.
31. Kesterson J. W. and Carlton W. W. 1971. Histopathologic and enzyme
histochemical observations of the cuprizone-induced brain edema. Exp. Mol.
Pathol. 15, 82-96
32. Kipp M., Clarner T., Dang J., Copray S. and Beyer C. 2009. The cuprizone
animal model: new insights into an old story. Acta Neuropathol. 118, 723-736
33. Kiss J., Rougon G. 1997. Cell biology of polysialic acid. Current opinion in
neurobiology 7, 640-646
34. Kondo A., Nakano T., Suzuki K. 1987. Blood-brain barrier permeability to
horseradish peroxidase in twitcher and cuprizoneintoxicated mice. Brain Res.
425, 186-190.
35. Koutsoudaki P. N., Hildebrandt H., Gudi V., Skripuletz T., Skuljec J., Stangel M.
2010. Remyelination after cuprizone induced demyelination is accelerated in
mice deficient in the polysialic acid synthesizing enzyme St8siaIV. Neuroscience
171, 235-244.

31
36. Kuhn H. G., Dickinson-Anson H.,Gage F. H. 1996. Neurogenesis in the
37. Levine J. M., Reynolds R., Fawcett J. W. 2001. The oligodendrocyte precursor
cell in health and disease. Volume 24, 39-47
38. Liedtke W., Edelmann W., Bieri P. L. 1996. GFAP is necessary for the integrity
of CNS white matter architecture and long-term maintenance of myelination.
Neuron17(4), 607-15.
39. Lindner M., Fokuhl J., Linsmeier F., Trebst C. and Stangel M. 2009. Chronic
toxic demyelination in the central nervous system leads to axonal damage despite
remyelination. Neurosci. Lett. 453, 120-125.
40. Luskin M. B., 1993. Restricted proliferation and migration of postnatally
generated neurons derived from the forebrain subventricular zone. Neuron 11,
173-189.
41. Maier O., Hoekstra D. and Baron W. 2008. Polarity development in
oligodendrocytes: sorting and trafficking of myelin components. J. Mol.
Neurosci. 35, 35-53
42. Mason J. L., Suzuki K., Chaplin D. D., Matsushima G. K. 2001b. Interleukin-
1beta promotes repair of the CNS. J. Neurosci. 21, 7046-7052.
43. Matsushima G. K. and Morell P. 2001. The neurotoxicant, cuprizone, asamodel
to study demyelination and remyelination in the central nervous system. Brain
Pathol. 11, 107-116.
44. McMahon E.J., Suzuki K. and Matsushima G.K. 2002. Peripheral macrophage
recruitment in cuprizone-induced CNS demyelination despite an intact blood-
brain barrier. J. Neuroimmunol. 130, 32-45.
45. Menn B., Garcia-Verdugo J. M., Yaschine C., Gonzalez-Perez O., Rowitch D.,
Alvarez-Buylla A. 2006. Origin of Oligodendrocytes in the Subventricular Zone
of the Adult Brain. J.Neurosci. 26(30), 7907-7918.
46. Nelson R. W., Bates P.A., Rutishauser U. 1995. Protein determinants for specific
polysialylation of the neural cell adhesion molecule. J. biol. chem 270, 17171-
17179.
47. Ono K., Tomasiewicz H., Magnuson T., Rutishauser U. 1994. N-cam mutation
inhibits tangential neuronal migration and is phenocopied by enzymatic removal
of polysialyc acid. Neuron 13, 595-609.
48. Rakic P. 1985. Limits of neurogenesis in primates. Science 227, 1054–1056.

32
49. Reeves S. A., Helman L. J., Allison A., Israel M. A. 1989. Molecular cloning
and primary structure of human glial fibrillary acidic protein. Proc. Nat.l Acad.
Sci. USA 86(13), 5178–5182.
50. Remington L. T., Babcock A. A., Zehntner S. P., Owens T. 2007. Microglial
recruitment, activation, and proliferation in response to primary demyelination.
Am. J. Pathol. 170, 1713-1724.
51. Rieger S., Volkmann K., Köster R. W. 2008. Polysialyltransferase expression is
linked to neuronal migration in the developing and adult zebrafish. Dev. dyn.
237, 276-285.
52. Rolls A., Shechter R., Schwartz M. 2009. The bright side of the glial scar in CNS
repair Rolls. Nat Rev Neuroscience10(3), 235-41.
53. Russanov E. M. and Ljutakova S. G. 1980. Effect of cuprizone on copper
exchange and superoxide dismutase activity in rat liver. Gen.Pharmacol. 11, 535-
538.
54. Rutishauser U. 1984. Developmental biology of a neural cell adhesion molecule.
Nature 310, 549-554.
55. Saher G., Brügger B., Lappe-Siefke C., Möbius W., Tozawa R., Wehr M.
C., Wieland F., Ishibashi S., Nave K. A. 2005. High cholesterol level is essential
for myelin membrane growth. Nat. Neurosci.8, 463-75
56. Santello M., Volterra A. 2008. Synaptic modulation by astrocytes via Ca(2+)-
dependent glutamate release. Neuroscience158(1), 253-9.
57. Silver J., Miller J. H. 2004. Regeneration beyond the glial scar. Nature Reviews
Neuroscience 5, 146-156.
58. Skripuletz T., Lindner M., Kotsiari A., Garde N., Fokuhl J., Linsmeier F.,Trebst
C. and Stangel M. 2008. Cortical demyelination is prominent in the murine
cuprizone model and is strain-dependent. Am. J.Pathol. 172, 1053-1061
59. Suzuki K. 1969. Giant hepatic mitochondria: production in mice fed with
cuprizone. Science 163, 81-82
60. Szele F. G., Chesselet M., 1996. Cortical lesions induce an increase in cell
number and PSA-NCAM expression in the subventricular zone of adult rats. J
Comp Neurol 368, 439–54.
61. Tandler B. and Hoppel C. L. 1973. Division of giant mitochondria during
recovery from cuprizone intoxication. J. Cell Biol. 56, 266-272.

33
62. Tardy M., Fages C., Le Prince G., Rolland B. 1990. Regulation of the glial
fibrillary acidic protein (GFAP) and of its encoding mRNA in the developing
brain and in cultured astrocytes. 1990. Adv. Exp. Med. Biol. 265, 41-52.
63. Taylor L. C., Gilmore W., Ting J. P. and Matsushima G. K. 2010. Cuprizone
induces similar demyelination in male and female C57BL/6 mice and results in
disruption of the estrous cycle. J.Neurosci. Res. 88, 391-402.
64. Tramontin A. D., Garcia-Verdugo J. M. G., Lim D.A., Alvarez-Buylla A. 2003.
Postnatal development of radial glia and the ventricular zone: A continuum of the
neural stem cell compartment. Cereb. Cortex 13, 580–587.
65. Venturini G. 1973. Enzymic activities and sodium, potassium and copper
concentrations in mouse brain and liver after cuprizone treatment in vivo. J.
Neurochem. 21, 1147-1151
66. Virchow R. 1854. Über das ausgebreitete Vorkommen einer dem Nervenmark
analogen Substanz in den tierischen Geweben. Virchows Arch. Pathol. Anat. 6,
562-572
67. Wakabayashi T., Asano M., Kurono C. 1975. Mechanism of the formation of
megamitochondria induced by copper-chelating agents. I. On the formation
process of megamitochondria in cuprizonetreated mouse liver. Acta Pathol. Jpn.
25, 15-37
68. Wang X., Lou N., Xu Q., Tian G. F., Peng W. G., Han X., Kang J., Takano T.,
Nedergaard M. 2006. Astrocytic Ca2+ signaling evoked by sensory stimulation in
vivo. Nat Neurosci. 9, 816–823.
69. Wilkins A., Majed H., Layfield R., Compston A., Chandran S. 2003.
Oligodendrocytes Promote Neuronal Survival and Axonal Length by Distinct
Intracellular Mechanisms: A Novel Role for Oligodendrocyte-Derived Glial Cell
Line-Derived Neurotrophic Factor. J. Neurosci. 23(12), 4967– 4974.
70. Williams R. W. 2000. Mapping genes that modulate mouse brain development: a
quantitative genetic approach. Results probl. cell differ. 30, 21-49.
71. Willis P., Berry M., Riches A. C. 1976. Effects of trauma on cell production in
the subependymal layer of the rat neocortex. Neuropath Appl Neurobiol 2, 377-
88.
72. Zhang Y., Xu H., Jiang W., Xiao L., Yan B., He J., Wang Y., Bi X., Li X.,Kong
J. and Li X. M. 2008. Quetiapine alleviates the cuprizoneinduced white matter
pathology in the brain of C57BL/6 mouse. Schizophr. Res. 106, 182-191.

34
73. Zupanc G. K. 2001a. A comparative approach towards the understanding of adult
neurogenesis. Brain behav. evol 58, 246-249.
74. Zupanc G. K., 2008. Adult neurogenesis and neuronal regeneration in the brain of
teleost fish. Journal of physiology-paris 102, 357-373.

Životopis
Ada Zelen rođena je u Splitu, 7.5.1988. godine. Osnovnu školu “Split 3” završava u
Splitu, nakon čega upisuje II. gimnaziju u Splitu. Tijekom dvije godine
srednjoškolskog obrazovanja završava tečaj talijanskog jezika u Riminiju (Rimini,
Italija). Godine 2007. upisuje Preddiplomski studij biologije na Prirodoslovno-
matematičkom fakultetu Sveučilišta u Zagrebu. Nakon završetka preddiplomskog
studija, 2011. godine upisuje Diplomski studij molekularne biologije na istom
fakultetu. Tijekom 2011. godine boravi u SAD-u kao polaznik „Work and travel“
programa na kojem dodatno usavršava engleski jezik te stječe radno iskustvo. Godine
2013. dobiva ERASMUS stipendiju te odlazi u ljetnom semestru na Sveučilište u
Potsdamu (Potsdam, Njemačka) gdje polaže engleski (C1, prema CEF stupnjevanju),
njemački (A1 prema CEF stupnjevanju), te talijanski jezik (B2, prema CEF
stupnjevanju). Godine 2013. odlazi na medicinski fakultet Sveučilišta u Osijeku gdje
radi diplomski rad na Zavodu za medicinsku biologiju pod mentorstvom prof. dr. sc.
Marije Heffer. 2014. godine volontira na projektu „Tjedan mozga” u Osijeku te
polaže radionicu „Glow brain project”-a na Hrvatskom institutu za istraživanje
mozga na Medicinskom fakultetu u Zagrebu. Prisustvuje te polaže radionicu na
međunarodnom kongresu „Bridges in Life Sciences 9th Conference” u Splitu. Od
svibnja 2014. volontira na Zavodu za embriologiju i histologiju na medicinskom
fakultetu Sveučilišta u Splitu. Koristi se Microsoft Office programskim paketom, te
Windows operativnim sustavima.

![PLANIRANJE RADA HIDROELEKTRANA ZA KRATKOROČNI … · 2019-01-26 · kvalitetno satno planiranje proizvodnje i potrošnje električne energije.[5] U takvom okruženju do izražaja](https://img.dokumen.tips/doc/110x75/5e5a3f3f192e0b65c13537a5/planiranje-rada-hidroelektrana-za-kratkorooeni-2019-01-26-kvalitetno-satno-planiranje.jpg)



























