Embed Size (px)
Citation preview

Praca poglądowa • Review Article
123
diagnostyka laboratoryjna Journal of Laboratory DiagnosticsDiagn Lab 2016; 52(2): 123-136
Actual strategies in human induced pluripotent stem cells (hiPSCs) differentiation – perspectives and challenges
Aktualne strategie różnicowania ludzkich indukowanych pluripotencjalnych komórek macierzystych – perspektywy i wyzwania
Ewelina Augustyniak1,3*, Katarzyna Kulcenty1*, Michał Lach1,3, Igor Piotrowski1, Wiktoria Maria Suchorska1,2
1Radiobiology Lab, Greater Poland Cancer Centre, Poznan, Poland2Department of Electroradiology, Poznan University of Medical Sciences, Poznan, Poland
3The Postgraduate School of Molecular Medicine, Medical University of Warsaw, Poland*The authors contributed equally to this work
SummaryThe application of stem cells (SCs) in regenerative medicine has recently become a rapidly growing field, holding promise for combat-ing a number of currently incurable disorders: including diabetes, neurodegenerative, retinal and cardiac diseases, as well as muscular dystrophy. The search for alternative approaches led to the development of human induced pluripotent stem cells (hiPSCs) which have unrestricted proliferative activity and pluripotency – the capacity to differentiation into derivatives of three germ layers (meso-, ecto – and endoderm). Because hiPSCs are developed from adult human cells throughout the forced expression of pluripotency factors, they are free from the ethical concerns associated with human embryonic stem cells (hESCs), that creation involves the destruction of human embryos. Moreover, the use of hiPSCs contributes to the development of personalized medicine that exploits patient-specific cells extremely useful in autologous grafts. In the present study the methods of hiPSCs differentiation into stem cell-derived neurons, cardiomyocytes, chondrocytes and osteocytes were summarized and evaluated having regard to their most important aspects.
StreszczenieW ostatnich latach, stosowanie komórek macierzystych w medycynie regeneracyjnej cieszy się coraz większym zainteresowaniem, ze względu na ich potencjalne wykorzystanie w stanach chorobowych, na które nie ma obecnie skutecznej metody leczenia: choroby doty-kające układ nerwowy, mięśniowy, szkieletowy. Poszukiwanie alternatywnych metod doprowadziło do rozwoju ludzkich indukowanych pluripotencjalnych komórek macierzystych (ang. human induced pluripotent stem cells, hiPSCs) z ludzkich zróżnicowanych komórek jak np. fibroblastów. Komórki te posiadają nieograniczony potencjał do samoodnowy i różnicowania w pochodne trzech listków zarodkowych (ekto-, mezo – i endodermy). Ze względu na fakt, że hiPSCs wywodzą się z ludzkich, dojrzałych komórek, ich przyszłe wykorzystanie jest wolne od kontrowersji etycznych, charakterystycznych dla ludzkich embrionalnych komórek macierzystych (ang. human embryonic stem cells, hESCs), których pozyskiwanie jest związane z niszczeniem ludzkich embrionów. Ponadto, wykorzystanie hiPSCs ma kolejną ważną zaletę, jaką jest możliwość ich wykorzystania w medycynie spersonalizowanej np. w przeszczepach autologicznych. W niniejszej pracy, zostały opisane i podsumowane aktualne metody różnicowania hiPSCs w kierunku neuronów, kardiomiocytów, chondrocytów oraz osteocytów, wraz z uwzględnieniem ich najważniejszych aspektów.
Key words: cardiomyocytes, chondrocytes, differentiation, human induced pluripotent stem cells, neurons, osteocytes, repro-gramming,
Słowa kluczowe: chondrocyty, kardiomiocyty, ludzkie indukowalne komórki pluripotencjalne, neurony, osteocyty, reprogramowanie, różnicowanie
ISSN 0867-4043
IntroductionStem cells (SCs) are primary, non-specialized cells which have abil-ity to self-renewal, reproduction and differentiation into a special-ized cells forming tissue. Those cells were firstly observed in terato-
ma – tumor arising in the ovaries of mice. Teratomas originate from germ cells, and thus have an extensive differentiation capability which also takes place during embriogenesis. The experiments on teratomas initiated a whole series of works, which in 1981 led
www.diagnostykalaboratoryjna.eu
124
to the establishment of first mouse embryonic stem cell (mESCs) lines [1, 2]. Further studies allowed to obtain the first human em-bryonic stem cells (hESCs) in 1998 [3]. These embryonic stem cells are pluripotent, what means, they are able to differentiate into all derivatives of the three primary germ layers (ecto-, meso – and endoderm). Because of their plasticity and potentially unlimited capacity for self-renewal, ES cell therapies have been proposed for regenerative medicine and tissue replacement in the case of injury or disease. However, due to the fact that they are obtained from an inner cell mass of blastocyst, which is related to its irre-versible damage, the application of SCs in therapeutic purposes raises many ethical and moral concerns. A real breakthrough in the subject of SCs was a work of Kazutoshi Takahashi and Shinya Yamanaka in 2006. They indicated the possibility of obtaining the
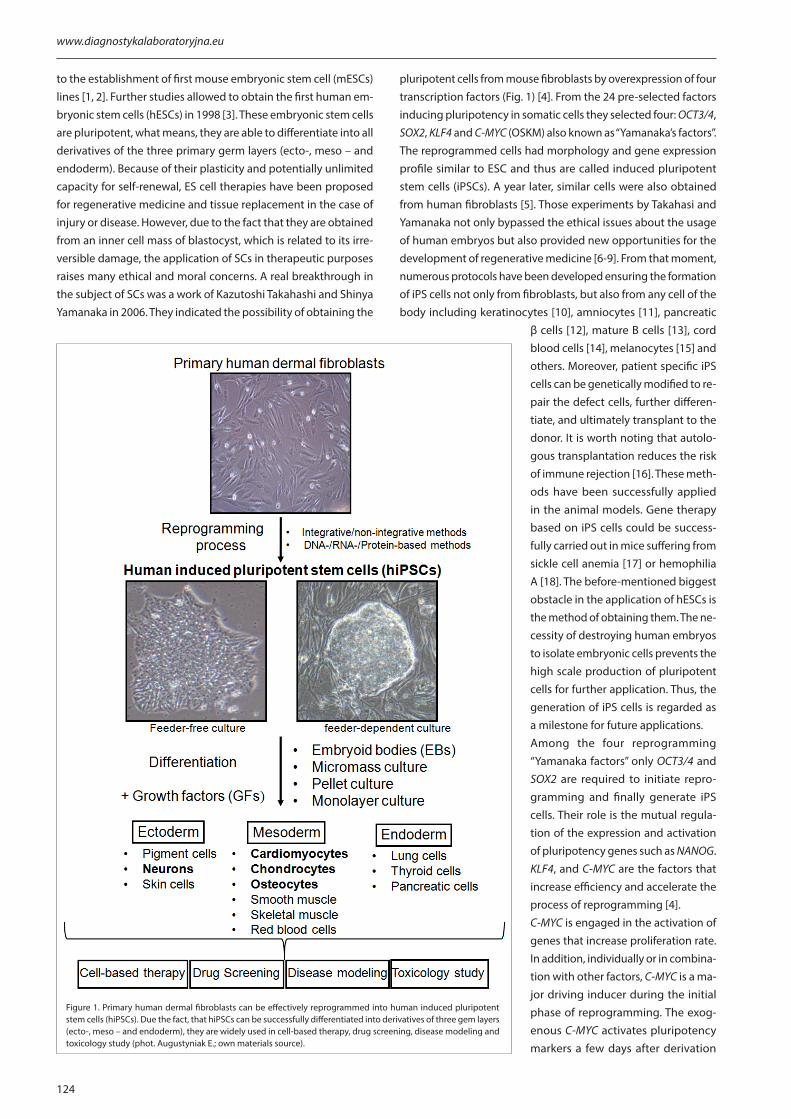
pluripotent cells from mouse fibroblasts by overexpression of four transcription factors (Fig. 1) [4]. From the 24 pre-selected factors inducing pluripotency in somatic cells they selected four: OCT3/4, SOX2, KLF4 and C-MYC (OSKM) also known as “Yamanaka’s factors”. The reprogrammed cells had morphology and gene expression profile similar to ESC and thus are called induced pluripotent stem cells (iPSCs). A year later, similar cells were also obtained from human fibroblasts [5]. Those experiments by Takahasi and Yamanaka not only bypassed the ethical issues about the usage of human embryos but also provided new opportunities for the development of regenerative medicine [6-9]. From that moment, numerous protocols have been developed ensuring the formation of iPS cells not only from fibroblasts, but also from any cell of the body including keratinocytes [10], amniocytes [11], pancreatic
β cells [12], mature B cells [13], cord blood cells [14], melanocytes [15] and others. Moreover, patient specific iPS cells can be genetically modified to re-pair the defect cells, further differen-tiate, and ultimately transplant to the donor. It is worth noting that autolo-gous transplantation reduces the risk of immune rejection [16]. These meth-ods have been successfully applied in the animal models. Gene therapy based on iPS cells could be success-fully carried out in mice suffering from sickle cell anemia [17] or hemophilia A [18]. The before-mentioned biggest obstacle in the application of hESCs is the method of obtaining them. The ne-cessity of destroying human embryos to isolate embryonic cells prevents the high scale production of pluripotent cells for further application. Thus, the generation of iPS cells is regarded as a milestone for future applications. Among the four reprogramming “Yamanaka factors” only OCT3/4 and SOX2 are required to initiate repro-gramming and finally generate iPS cells. Their role is the mutual regula-tion of the expression and activation of pluripotency genes such as NANOG. KLF4, and C-MYC are the factors that increase efficiency and accelerate the process of reprogramming [4].C-MYC is engaged in the activation of genes that increase proliferation rate. In addition, individually or in combina-tion with other factors, C-MYC is a ma-jor driving inducer during the initial phase of reprogramming. The exog-enous C-MYC activates pluripotency markers a few days after derivation
Figure 1. Primary human dermal fibroblasts can be effectively reprogrammed into human induced pluripotent stem cells (hiPSCs). Due the fact, that hiPSCs can be successfully differentiated into derivatives of three gem layers (ecto-, meso – and endoderm), they are widely used in cell-based therapy, drug screening, disease modeling and toxicology study (phot. Augustyniak E.; own materials source).

125
Diagn Lab 2016; 52(2): 123-136
into target cell [19]. It has been demonstrated that C-MYC togeth-er with complexes of histone acetylases induces global histone acetylation and allows for the binding of OCT3/4 and SOX2 to DNA [20]. It was revealed that KLF4 is able to inhibit the activation of NANOG and other genes characteristic for SCs. KLF4 plays a dual role in reprogramming process: it represses the function of genes characteristic for differentiated cells during the initial step and activates the pluripotency genes associated with the final stage of reprogramming (Fig. 2) [19].Despite the significant progress in generating iPS cells, many concerns about iPS cells generation must be resolved before their application on the clinical scale. The most important issue is the safety of the method used for the reprogramming and its effi-cacy. Reprogramming transgene delivery methods typically are classified to viral and non-viral ones. Both of them can be further divided to integrative and non-integrative into the host genome methods as well as to DNA-, RNA- or protein-based approaches. Non-integrating methods include the use of adenoviral vectors, plasmids, episomal vectors and Sendai virus. Direct methods of reprogramming are based on active absorption of the recom-binant proteins or mRNA by the cell. The adenoviral vectors are characterized by low efficiency of reprogramming process and the limited ability to control the level of transgene expression [21]. Also, episomal vectors oriP / EBNA-based Eppstein-Baar virus are characterized by a very low efficiency of reprogramming [22]. Although, the usage of Sendai virus guarantees relatively high efficiency of reprogramming, its removal from the forming iPS cells is problematic [23]. Plasmid vectors due to their inability to replicate in a cell, require multiple transfections during the process of de-differentiation [24]. The use of recombinant proteins allows obtaining iPS cells without introducing exogenous genetic material into a cell. However, proteins have a limited capacity
to translocate through the cell membrane. In order to provide reprogramming factors into the cell, the protein are usually com-bined with the cell membrane penetration enhancers [25, 26]. Another method of generating iPS cells is the use of recombinant mRNA. This type of reprogramming is characterized by a relatively high yield, but requires several incubations with a cocktail mRNA. A significant obstacle in potential use of this protocol on a wider scale is the fact that the introduction of a foreign RNA induces defense cell systems involving antiviral response which causes high cytotoxicity of applied methods [27]. However, safer, non-in-tegrating methods are characterized by relatively low efficiency of the reprogramming process. Integrating vectors, such as retroviral, lentiviral vectors or transposons guarantee efficient reprogram-ming of somatic cells. However, a transgene integrated into the genome of a cell, may cause genetic instability and malignant cell transformation. An important advantage of retroviral vectors is the spontaneous silencing of transgene expression. However, it can be reexpressed during cell differentiation [5]. The need to develop methods of obtaining secure, devoid of transgene expression iPS cells led to the creation of systems based on the excision of the integrated transgene. Several methods have been developed based on excision of the transgene or by regulating its expression. One of them involve the use of naturally occur-ring transposons which have the ability to translocate around the genome in a specific process – transposition. Currently, two commonly used protocols are based on transposons: piggyBAC and sleeping beauty, associated with high efficiency in reprogram-ming of somatic cells [28, 29]. Their removal from the genome is possible with the application of the enzyme transposase. Another system based on the excision of the integrated transgene is the Cre-loxP system, wherein the loxP site is positioned in the 3’LTR-region of the vector. During the integration loxP site is duplicated
Figure 2. Yamanaka’s factors (OCT3/4, SOX2, KLF4, C-MYC, OSKM) play a crucial role in reprogramming process. They are responsible for induction of pluripotency state in reprogrammed adult cells (own materials source).

www.diagnostykalaboratoryjna.eu
126
to 5’UTR region, which results in the integration of the transgene surrounded by two loxP sites. This enables subsequent excision with Cre recombinase [30]. The system of regulated transgene expression may also be based on the use of specific promoters, which activity can be pharmacologically regulated, for example a tetracycline-inducible promoters. The use of this system in such vectors depending on the requirements, enables or disables the transgene expression by supplementing the culture medium with doxycycline [31]. Another important issue concerning the application of iPS cell in the clinic is the homogeneity of generated iPS cells population. Many researchers noted, that reprogramming of mouse somatic cells into iPS cells often generates heterogeneous population with partially and fully reprogrammed iPS cells [32-34]. Mechanistic studies involving global analysis indicated how reprogramming factors regulate molecular changes during somatic to pluripotent state transition. Meissner et al firstly pointed out that DNA meth-ylation has a critical role in the restablishment of pluripotency [35]. Together with many other researchers, they pointed out, that epigenetic mechanisms involved in reprogramming process play crucial role in obtaining fully reprogrammed iPS cells which results in higher homogeneity of generated cell population [32, 36-38].The same problem of heterogenic population occurs dur-ing differentiation process. In the recent work Loh et al defined the signals controlling the binary lineage decision. Their observation enables to block differentiation toward unwanted fates and obtain the 80-90% pure human mesodermal lineage which can be used in in vivo studies [39].However many obstacles occur, first clinical trial using iPS cells started in September 2014 for treating age-related macular de-generation (AMD) [40]. The investigators at RIKEN, a research institute in Kobe, Japan, treated a single patients using retinal
pigment epithelium differentiated from autologous iPS cells de-rived from patient’s skin biopsy. Although these patients didn’t have any serious adverse effects, due to changes in regenerative medicine law, the trial was suspended in March 2015. The RIKEN group is now revising their trial and preparing new protocol for allogeneic cells application. The allogenic cells will come from CiRA, the institute which is the pioneer in developing a bank of iPS cells [41]. The use of allogeneic cells, which are haplotype matched to patient’s major human leukocyte antigens (HLA’s), can omit the problem of transplant rejection and necessity to use immunosuppression. However giving patients their own cells still remains the best solution, due to costs and time of their genera-tion, the HLA-matched bank of iPS cells seems to be beneficial. This review presents recent advances in stem cell-based therapy and summarizes data from the literature concerning hiPSC differenti-ation into neural, chondrogenic, osteogenic and cardiomyogenic lineage (Fig. 3)(Table I).
iPSC differentiation into neurons and neural progenitorsAlthough iPSC technology is a novel strategy in regenerative medicine, it shows a great promise in the therapeutic use and in disease modeling. Because, iPSC technology allows for in vitro production of neural cells and progenitors, which are difficult to obtain via alternative methods, neurological diseases are likely to be successfully treated in the near future. Development of efficient differentiation methods is especially important in the potential use of iPSC-derived cells as patient-specific grafts [42].
Differentiation of iPSCs into neuronal cell linesThere are numerous efficient methods used for iPSC differentia-tion into neuronal cells. One of the most commonly used proto-cols involve iPSC growth as spheres. Firstly, iPSCs are detached
from feeder layer and then placed in human neural progenitor growth me-dium with the addition of serum free supplement for neural cell culture – B27, basic fibroblast growth factor (bFGF), epidermal growth factor (EGF) and heparin. After two weeks, spheres are dissociated and plated on laminin coated glass coverslips. For induction of differentiation into motor neurons, spheres are cultured in neural induc-tion medium supplemented with reti-noic acid (RA), sonic hedgehog (SHH), cyclic adenosine monophosphate (cAMP), ascorbic acid (AA), brain de-rived neurotrophic factor (BDNF) and glial cell-line derived neurotrophic fac-tor (GDNF) [43]. Karumbayaram and colleagues used embryoid body (EB) formation in non-adherent conditions to differentiate hiPSCs into motor neu-rons. hiPSC-derived EBs were treated with RA and SHH pathway agonist and
Figure 3. The currently methods of hiPSC differentiation are mainly based on the formation of embryoid bodies, micromass culture, pellet culture and monolayer culture. These approaches enable obtaining stem-cell derived such as neurons, astrocytes, cardiomyocytes, chondrocytes, osteoblasts and osteocytes as well as they mimic main stages of differentiation and maturation processes taking place during embryo development (phot. Augustyniak E; own materials source).

127
Diagn Lab 2016; 52(2): 123-136
after 5 – 7 days generated neural progenitors that stained positive-ly for POU class 3 homeobox (BRN2), SRY (sex determining region Y)-box3 (SOX3) and Paired box 6 (PAX6). After next 8-10 days of culture with RA, SHH pathway agonists and neurotrophic factors: BDNF, ciliary neurotrophic factor (CNTF) and GDNF, EBs gave rise to motor neuron progenitors that revealed the presence of charac-teristic markers: NK6 homeobox 1 (NKX6.1) and Oligodendrocyte Lineage Transcription Factor 2 (OLIG2). Finally, after 3 – 5 weeks differentiated EBs were able to form mature motor neurons [44, 45]. Another effective approach to obtain neuronal stem cell-de-rived cells was previously used to differentiate hESCs. It involved the formation of neuroectodermal rosette-like structures through the co-culture with stromal feeder cell lines such as PA6 or MS5 [46, 47]. Cooper and colleagues (2010) successfully implemented this protocol for generation of hiPSC-derived dopaminergic (DA) neurons. Neuroectodermal induction of hiPSCs was achieved by co-culture with MS5 feeder cells using both serum replacement medium and DMEM/F-12 medium supplemented with N-2 (se-rum-free supplement based on Bottenstein’s N-1 formulation) , NOGGIN, Wingless-type MMTV integration site family, member 1 (WNT1), Fibroblast growth factor 8b (FGF8b) and RA. At day 14 neuroectodermal colonies were differentiated into medium containing N-2 supplement and with the addition of following growth factors (GFs): SHH, FGF8b, BDNF, transforming growth factor type β3 (TGF-β3), GDNF, cAMP and AA until the presence of Forkhead Box A2 (FOXA2) positive DA neurons was observed [48]. The protocols based on EB formation or co-culture with stromal cells display major drawbacks such as long duration of differentia-tion, necessity to use undefined factors like neural-inducing stro-ma and heterogeneous nature of EBs. Chambers and colleagues (2009) resigned from the formation of EBs and took advantage of direct inhibition of the SMAD pathway to obtain hiPSC-derived midbrain dopamine and spinal motor neurons after 19 days with noticeable effectiveness. hiPSCs were dissociated into single cells and transferred to the medium with ROCK-inhibitor to promote survival of single cells of iPSCs. After 3 days medium was switched to another one based on knock-out serum replacement with two SMAD inhibitors: NOGGIN and SB431542. The acquisition of do-paminergic features was achieved by sequential administration of the following GFs: SHH, BDNF, FGF8, AA, GDNF, TGFβ3 and cAMP. In order to obtain motor neuronal cells the administration of BDNF, AA, SHH and RA was applied [49]. hiPSC-derived neu-
ral progenitors obtained via NOGGIN treatment, co-culture with stromal cells and direct neural induction on laminin substrate can be maintained as neurospheres and then differentiated into fully mature neurons, glia and crest cells in the presence of EGF and bFGF [50]. hiPSC-derived neural progenitors were also suc-cessfully used for differentiation into glial cells: astrocytes and oligodendrocytes. Firstly, hiPSCs were maintained in suspension culture to form EBs. Then, attached EBs formed neuroepithelial cells that were subsequently cultured with the addition of RA and SHH for 2 months in DMEM/F12 with N1 supplement and cAMP, or with 3,3,5-Triiodo-L-thyronine (T3), platelet-derived growth factor (PDGF), insulin-like growth factor 1 (IGF-1) and neurotro-phin 1 for further astrocyte or oligodendrocyte differentiation, respectively [51]. Although, hiPSC-derived neurons can be used for high-throughput drug screening, there exist some limiting factors. The major disadvantage is the fact that hiPSC differenti-ation is cost – and time – consuming as well as the differentiated hiPSCs often form heterogeneous population consisting of cells possessing diverse properties. Hence, researchers reach for com-mercially available fully differentiated cells like iCell® neurons and iCell® astrocytes [42]. iCell® neurons are composed of pure and well-defined population of GABAergic and glutamatergic post-mitotic neurons. iCell® neurons reveal fully electrical func-tionality and show differences in neurite outgrowth depending on cell culture conditions which might be useful for body-on-a-chip (sophisticated method leading to creation of interconnected tissue-like organ structures) applications [52]. The co-culture of iCell® neurons with rat astrocytes allowed for the long-term culture of extremely stable neurons. Up to 3 month neurons maintained functional maturity and they were susceptible to addition of syn-apse agonist drugs [53]. iCell® neurons were also prone to Aβ toxicity in a similar manner to hiPSC-derived neurons [54]. These results demonstrate, that commercially available cell lines might be a useful tool in drug screening thanks to their homogeneity, long-term viability and possibility to purchase large number of cells without the need to carry out the time-consuming differ-entiation process.
The application of iPSC derived neural cells in neurological diseasesMost of currently studies involving neurological diseases are performed in vivo in animal models [55] or ex vivo on post-mor-tem tissues [56]. The iPSC technology gives a great opportunity
Table I. The most common markers characteristic for pluripotent, neural, cardiac, chondrogenic and osteogenic cell lines.
PLURIPOTENT NEURAL CARDIAC CHONDROGENIC OSTEOGENIC
POSI
TIV
E M
ARK
ERS
OCT3/4, NANOG, SSEA-3, SSEA-4, TRA-1-60,
TRA-1-81, SOX2, CD9, CD24, CD30, CD133, SALL4, STAT3,
E-CADHERIN, ALKALINE PHOSPHATASE, INTEGRIN β1, INTEGRIN α6, C-MYC,
KLF5, LIN-28, SMAD2
CD15, CD44, NESTIN, O1, O4, OLIG2, NOGGIN, NOTCH1, S100B, SOX10,
GFAP, GLAST, PAX6, MAP2, GALC, NeuN, βTUBULIN III
BMP-4, DESMIN, GATA-4, NKX2.5, TROPONIN T,
TROPONIN I, αSARCOMERIC ACTIN,
MYOSIN HEAVY CHAIN, MYOGLOBIN, CK-MB,
TBX5, ATRIAL NATRIURETIC FACTOR,
AGGRECAN, TYPE II & IV COLLAGEN, SOX5, SOX6,
SOX9, CD44, CD151, SULFATED PROTEO-
GLYCAN, COMP, NKX3.2
ALKALINE PHOSPAHATASE, TYPE I &
II COLLAGEN, OSTERIX, OSTEOCALCIN, BONE
SIALOPROTEIN 2, RUNX2, OSTEONECTIN, FIBRONECTIN, SPARC,
THROMBOPOIE-TIN, BIGLYCAN

www.diagnostykalaboratoryjna.eu
128
to use neural stem-derived cells that precisely recapitulate disease course taking place in human nervous system. Patient-specific cells that are burdened with neurological disease background can be differentiated into specific neural cells and further used to investigate disease mechanisms [57]. The ability of iPSC-derived cells to establish useful cellular models of neurological diseases constitute efficient platforms for developing new drugs [54].hiPSC-derived neuronal cells recapitulate properties observed in neuronal cells in vivo. Shi and coworkers (2014) demonstrated that differentiated hiPSCs can be used to investigate the model of human cortical development. hiPSCs were differentiated into neuroepithelial cells and then towards cortical stem and pro-genitor cells. Further differentiation in vitro mimicked processes taking place during neurogenesis in vivo, such as acquisition of mature electrophysiological properties and formation of func-tional excitatory synapses [58]. The capacity of hiPSC-derived neurons to recapitulate properties of human neurons makes them great candidates for establishing neurological disease models and drug screening platforms. Yagi and colleagues (2011) proved that hiPSCs generated from patient suffering from familial Alzheimer’s disease (FAD) can be a viable source of neurons for both disease modeling and drug screening. FAD-iPSC-derived neurons showed increased secretion of amyloid β42 (Aβ42). After treatment with a γ-secretase inhibitor and with a selective Aβ42-lowering agent, Aβ42 secretion was inhibited or modulated, as expected [59]. In another study, the hiPSC-derived cortical cells capable of recapit-ulation of Aβ-induced synaptic patomechanisms were established [60]. The immunopurification procedure allowed to generate electrophysiologically active mature hiPSC-derived neurons sus-ceptible to synaptotoxic effects of Aβ. The patient-specific hiPSC derived neurons were also used to elucidate disease mechanisms in Huntington’s disease (HD). Mattis et al. (2012) used fibroblasts derived from patients with different numbers of CAG repeats
to generate hiPSC lines. The comparison of gene expression profile between control and HD hiPSCs revealed that these two cell lines are characterized by different properties. Some of the observations contributed to the discovery of the novel altered pathways involving matrix metalloproteinases or developmental genes. HD hiPSCs that had been differentiated into neural pro-genitor cells through formation of spheres displayed decreased cell-cell adhesion properties and reduced energy metabolism as well as they showed significant changes in actin cytoskeleton organization. Differentiated HD neural stem cells (NSCs) with higher number of CAG repeats were more prone to cell death and were more sensitive to the absence of BDNF [43, 61]. An and co-workers (2012) proved that hiPSCs generated from patients with HD can be genetically corrected in vitro by the replacement of the expanded CAG region with a normal repeat region using Flp-FRT recombination. Both corrected HD-hiPSCs and NSCs de-rived from them showed no further HD progression. Moreover, NSCs derived from corrected HD-hiPSCs were transplanted into the striatum of mouse affected by HD, wherein they have the capacity to populate the striatum and further differentiate into neurons and glial cells [62]. hiPSCs were also used as a source of cells for autologous transplantation in cynomolgus mokey (CM) Parkinson’s disease (PD) model. CM-iPSCs were differentiated into dopamine neurons using co-culture with stromal feeder cells [63]. The monkeys subjected to the autologous transplantation of do-pamine neurons into putamen revealed significant improvement of motor function and activity. Cells derived from iPSCs were also used in the transplantation during therapy of spinal cord injury (SCI) in the common marmoset. In this case, secondary and tetriary neurospeheres differentiated from hiPSCs, were transplanted into non-human primate model of SCI. Cell transplantation reduced the severe demyelination, increased axonal survival and regrowth as well as improved angiogenesis in the lesion epicenter. Fur-
Table II. The summary of the most important applications of iPSC derived neural cells in neurological diseases
Author [number of reference]
iPSC origin Cell type into which iPSC differentiate
Model of neurological disease
The most important conclusions
Yagi T et. al [49]Fibroblasts from FAD
patientsNeurons
Familial Alzheimer’s disease
Neurons from FAD iPSCs shown increased Aβ42 secretion
Nieweg K et. al [50]Commercially available
human iPSC lines (8/25 and DF6–9–9T.B)
GABAergic and glutamatergic neurons
Alzheimer’s diseaseNeurons shown Aβ-induced
patomechanisms
Mattis VB et. al [51] Fibroblasts from HD patientsNeural progenitor cells,
neural stem cells, striatal neurons
Huntington’s diseaseHD associated changes in HD neurons correlate with number of CAG repeats
An MC et. al [52] Fibroblasts from HD patients Neural stem cells Huntington’s diseaseNeurons from genetically corrected
iPSCs populated striatum and differentiated after transplantation
Hallett PJ et. al [53]Fibroblasts from cynomolgus
monkeyMidbrain dopamine
neuronsParkinson’s disease
Neural cell transplantation improved motor functions and activity
Kobayashi Y et. al [54]Commercially available
human iPSC lines (201B7)Secondary and tertiary
neurospheresSpinal cord injury
Neurosphere transplantation improved angiogenesis, axonal survival,
myelination and functional recovery
Abbreviations: iPSC – induced pluripotent stem cells; FAD – familial Alzheimer’s disease; GABA – γ-aminobutyric acid; Aβ – amyloid-β; HD – Huntoington’s disease

129
Diagn Lab 2016; 52(2): 123-136
thermore, the animals after transplantation demonstrated better functional recovery after therapy of SCI [64].The iPSC technology could serve as a new tool to study neuro-logical disease mechanisms in vitro and to screen for new drugs. The ability to differentiate patient derived iPSCs into neuronal cells could provide researchers with new data regarding neurode-generative diseases. iPSC-derived cells not only allow to establish disease models but also could be used as a source of cells for autol-ogous transplantation in neurological disease treatment (Table II).
The use of differentiation process in obtaining human cardi-omyocytes-like cellshiPSC-derived cardiomyocytes (hiPSC-CMs) are considered as candidates for the emerging field – regenerative medicine in-volving personalized medicine (e.g. autologous cardiomyocytes) and patient/disease-specific in vitro models of human cardiac tissue. The capacity to differentiate hiPSCs into cardiomyocytes was advanced by the observation that three-dimensional ESC ag-gregates named EBs could give rise to spontaneously contracting cells, which possess cardiomyocyte-specific markers [65]. There are several effective methods of obtaining hiPSC-CMs that will be summarized in this part of study. HiPSCs were differentiated into cardiomyocytes throughout EBs for 10 days. This process proceeded spontaneously for 30-50 days on gelatin-coated plates [66]. One of the less complicated and cost-effective method of hiPSCs differentiation into cardiomyocytes is the formation of EBs and their further culture in suspension for two weeks. This approach eliminates the addition of expensive exogenous GFs and the need for transfer of EBs on gelatin-coated plates as well as this protocol is not time-consuming. Zwi and co-workers (2009) proved that hiPSCs have the ability to differentiate into cardiomy-ocytes via EBs without the additional GFs. They demonstrated that during the first stage of differentiation, the level of pluripotency markers such as OCT4, NANOG abruptly decreased as well as primitive streak and cardiomesoderm markers (Brachyury, mes-oderm posterior bHLH transcription factor: 1-MESP1) increased [67]. Then, the presence of cardiac-specific transcription factors and finally cardiac-specific structural genes (sarcomeric – related proteins and ion channels) was observed. These cells possess characteristics assigned to mature cardiomyocytes such as the presence of homeobox protein NKX2.5 (NKX2.5), GATA binding protein 4 (GATA4), atrial natriuretic peptide as well as they can be used in in vitro pharmacological tests, particularly cardiac elec-trophysioloic studies and drug screening [68]. EBs formation and culture them in EB20 medium (DMEM containing 20% FBS, 1 mM non-essential amino acids, 1 mM l-glutamine, 0.1 mMβ-mercap-toethanol, 100 units/ml penicillin, 100 μg/ml streptomycin) is the most common method of obtaining hiPSC-CMs [69, 70]. Another approach of hiPSCs differentiation into cardiomyocytes was rep-resented by Liang et al. (2013), who obtained hiPSC-CMs via EBs in suspension culture [71]. They implemented sequential adminis-tration of following GFs: BMP-4, bFGF, activin A, Dickkopf-related protein 1 (DKK1) and vascular endothelial growth factor (VEGF) for generation of hiPSC-MCs from healthy subjects and patients with hereditary long QT syndrome, hypertrophic cardiomyopathy as
well as dilated cardiomyopathy patients. In that study, hiPSC-MCs were used in evaluation of drug-induced cardiotoxicity. Healthy and diseased individuals reveal different susceptibilities to cardi-otoxic drugs what confirms that disease-specific hiPSC-CMs may be used in effective prediction of adverse drug response. Kim and collaborators (2015) established the following protocol for obtaining hiPSC-derived cardiomyocytes via EBs in suspension culture: days 0–1, BMP4 (5 ng/ml); days 1–4, BMP4 (10 ng/ml), bFGF (5 ng/ml) and Activin (1.5 ng/ml); days 4–20, DKK1 (150 ng/ml) and VEGF (10 ng/ml) for the investigation of mechanism responsible for spontaneous contractions of hiPSC-CMs [72]. Car-diac differentiation through attached on 0.1% gelatin EBs, can be strongly enhanced by the addition of 100 µM ascorbic acid during the whole differentiation process, which lasted approximately 20 days. These cells were successfully utilized in the investigation of hypertrophic cardiomyopathy characteristic for Friedreich’s ataxia (FRDA) disease [73]. Another approach involves the creation of nearly pure hiPSC-CTMs population. The hiPSC-CMs with high cardiomyocyte purity can be selected throughout the expression of blasticidin resistance under the control of the cardiac myosin heavy chain 6 (MYH6) promoter. The hiPSC-CMs with high purity display cellular electrophysiological properties that are similar to mature cardiomyocytes. In addition, these cells can be easily expanded suggesting the susceptibility to high-throughput assays as well as they can be effectively cryopreserved [74]. Relatively small number of protocols involve differentiation into cardiomy-ogenic lineage through monolayer culture. Jung and colleagues (2016) established efficient method for obtaining hiPSC-CMs via complex monolayer differentiation protocol on Matrigel-coated plates. They characterized the ontogeny of downstream pathways of β-AR signaling in hiPSC-CMs. In those cells cardiac functionality matured significantly earlier than features involving cardiac re-modeling and toxicity. Those data contribute to conclusion that hiPSC-CMs should be studied and investigated when components of β-AR signaling reflecting cardiac remodeling and toxicity are fully mature [75]. Another direct differentiation protocol included application of activin A and BMP-4 under serum-free, monolayer culture conditions. Furthermore, the use of T3 promoted matura-tion of hiPSC-CMs: they exhibited a larger cells size, longer sarcom-eres length, enhanced calcium handling properties and increased maximal mitochondrial respiration capacity [76]. Currently, there is reduced demand for differentiation of hiPSCs into hiPSC-CMs on a laboratory scale. The autonomous differentiating process is time – and cost-consuming. Furthermore, the utility of obtained cells should be validated: the increase of desirable ectodermal markers, decrease of pluripotency markers, functionality in in vitro and in vivo models. Thus, researchers reach for fully defined and available cell lines, e.g. Guo and co-workers (2011) used iCell® car-diomyocytes in estimating the risk of drug-induced proarrhtyhmia [77]. Unlike primary culture systems of human cardiomyocytes, iCell® cardiomyocytes express cardiac contractile proteins, ma-jor functional ion channels and reflect spontaneous mechanical and electrical activities assigned to mature cardiomyocytes and are very useful in human-based cardiotoxicity testing models in vitro [78] . iCell® cardiomyocytes have not only ion channels

www.diagnostykalaboratoryjna.eu
130
activities that are remarkably similar to adult cardiomyocytes, but also have many features required for advanced studies of cardiac ion transporters (Na/K pump, Na/Ca exchange currents, Na/H exchange and Cl/OH exchange). Fine and colleagues (2013) demonstrated that these cells relatively easily undergo molecular manipulations like overexpression and knockdown of transporters and regulatory proteins [79]. This characteristic makes them very attractive culture system for studies involving regulatory proteins and long-term regulatory pathways influencing the expression and function of membrane transporters.
The most relevant properties of hiPSC-CMsCardiomyocytes differentiated from hiPSCs exhibit features char-acteristic for in vivo cardiac myocytes: functional ion channels, receptors, transporters, syncytial and contractile activities as well as electrophysiological and biochemical responses upon expo-sure to environmental stimuli. Furthermore, Rana and colleagues (2012) revealed that hiPSC-derived cardiomyocytes use anaerobic or aerobic respiration depending on available carbon source. They closely resemble adult heart tissue when they are cultured with galactose or galactose and fatty acids at the same time [80]. Zwi-Dantsis and collaborators (2013) proved that hiPSCs-CMs derived from heart failure patients show the ability to functionally integrate with pre-existing cardiac tissue in vitro and to engraft, survive and finally integrate with the rat myocardium following in vivo transplantation [81]. Thus, not only young and healthy indi-viduals can provide functional hiPSCS-CMs by EBs differentiation on gelatin-coated plates but also patients suffering from heart failure and clasiffied for autologous myocardial cell-replacement procedure can provide effective hiPSCs-CMs. HiPSC-CMs obtained from patients with arrhythmogenic right cardiomyopathy (ARVC) mimic key features of this disease phenotype: reduced cell surface localization of desmosomal proteins and better adipogenic phe-notype in comparison with cardiomyocytes from control hiPSCs. Thus, patient-specific hiPSC-CMs are likely to be a helpful tool in study of ARVC [70]. Latest literature data indicate that application of patient specific hiPSC-CMs obtained via co-culture of hiPSCs with inactivated mouse visceral endoderm-like (END-2) cells, reca-pitulate clinical observations gained from patients with LQTS and provide further evidence that these cells can be used in in vitro drug assays to investigate their vulnerability to arrhythmia [82]. Ivashchenko and collaborators (2012) demonstrated that after differentiation process, the immature cardiomyocyte-like from hiPSCs are obtained [83]. During the culture period, they acquire more functionally mature phenotype. The experiment at different time points may give varying results and lead to inconsistent and incorrect conclusions concerning the functionality of hiPSC-CMs. Therefore, there is a strong need to monitor phenotype of these cells during the duration of experiment. Similar results were ob-tained by Kamakura et al. (2013) who reported that long-term culture (180 – and 360 – days) of hiPSC-MCs obtained via EBs differentiation leads to maturation of differentiated cells. On the contrary, short – and mid-term culture of hiPSC-CMs favor the less mature cells than those of adult hearts [84]. A comprehensive research was done by Citro and co-workers (2014) who proved
that hiPSCs-CMs are equivalent to human mesenchymal stem cells (hMSCs) in improving cardiac function following myocardial infarction in a rat model. Moreover, hiPSCs-CMs revealed supe-rior characteristic in attenuating fibrosis to MSCs. Transplanted hiPSC-CMs proper integrated with the host myocardium and ex-pressed mature cardiomyocyte markers suggesting electrome-chanical coupling of engrafted cells [85]. Zhang and colleagues (2009) compared the CMs obtained from hiPSCs and hESCs via EBs differentiated in suspension and following monolayer culture [86]. hiPSC – and hESC – derived cardiomyocytes revealed similar cardiac gene expression patterns, indistinguishable sarcomeric organizations, capacity to differentiate into nodal-, atrial – and ventricular phenotypes as well as responsiveness to β-adrenergic stimulation. However, the authors indicate that hiPSCs retain re-sidual transgene expression because of blunted downregulation of pluripotency genes (OCT4 and NANOG) in hiPSCs compared to hESCs during differentiation. iPSC-CMs represent a novel tool that bridge gaps between animal-based models and adult human cardiomyocytes. However, it is important to note that iPSC-CMs constitute a single-cell model of cardiac disease and lack the complexity of native cardiac tissue. Thus, iPSC-CMs may not reflect the complexity of organ-level or systemic interaction that could influence cardiac disorders [87].
Induced pluripotent stem cells in cartilage and bone repair.The another application of SC based therapies involves the treat-ment of muscle-skeletal diseases concerning bones and joints. In the near future, currently incurable diseases such as osteoarthritis, osteoporosis will constitute a great burden, particularly in well developed countries [88, 89]. Cartilage has reduced regeneration capacity, due to its avascular nature and small number of func-tional cells associated with the dense extracellular matrix (ECM). Thus, the treatment of diseases related to the joint degeneration is extremely challenging. The current techniques of cartilage re-generation such as autologous chondrocyte implantation (ACI) or microfracture very often result in the formation of undesirable fibrocartilage instead of characteristic hyaline cartilage [90, 91].In turn, bones have the capacity to regenerate itself, when the damages or depletion are relatively small. In patients suffering from huge trauma such as bone tumor resection, osteoporosis, bone infections, the self-regenerative process is not sufficient and thus, commercial available scaffolds are necessary to improve recovery of patients [89, 92, 93]. The capacity to regenerate frac-tures in elder’s people limbs may be decreased because of the presence of osteoporosis, which is responsible for the reduction of natural bone scaffold. Those issues constitute a challenging area of regenerative medicine, because currently there is no direct treat-ment reflecting natural microenvironment of tissue. MSCs seems to be a suitable source of cells for SC based therapies, due to their increased proliferation capacity and ability to differentiate into chondrocytes or osteoblast. However, their potential application is often strongly limited because of the age and condition of patients that may result in significant decreased number of MSCs [94, 95]. Thus, the pluripotent SCs are an interesting and promising tool in treatment of degenerative diseases related to the bones and

131
Diagn Lab 2016; 52(2): 123-136
cartilage [96]. They are an unlimited source of cells with self-re-newal properties, which enable to cultivate them to appropriate amount of cells and subsequently differentiate them into desired lineage [97]. There are many studies involving chondrogenic and osteogenic differentiation of pluripotent SCs. There are many issues, which need to be verified before future application of stem cell-derived chondrocytes and/or osteoblasts. Functionality similar to donor tissues, complete integration to the donor site, recovery of three dimensional structure of implemented cells is required. Moreover, these cells should be obtained with the use of efficient, cost-effective and not time-consuming protocols. Thus, further research involving differentiated chondro – or osteocytes from SCs is needed.
Cartilage and bone developmentDuring embryo development three germ layers: mesoderm, endoderm and ectoderm are present. The musculoskeletal and hematopoietic tissues have an origin in mesenchyme, which is mature form of mesoderm. Because bone and cartilage have the common origin, the currently differentiation protocols are mostly based on receiving mesodermal population of cells [98]. During the first stage of chondro – and osteogenesis, the cells originated from condensed mesenchymal cells express the N-cadherin, neuronal adhesion molecule 1 (NCAM1) Tenascin-C (TNC), SRY (sex deter-mining region Y)-box 9 (SOX9) and type II collagen (COL2A1) [99]. This process enables the formation of cartilage-like nodules. Then, the deposition of proteoglycans caused by increased expression of aggrecan (ACAN) proceeds [100]. The maturation of chondrocytes is accompanied by hypertrophy: chondrocytes growth, enlarge and stop producing ECM proteins characteristic for hyaline carti-lage (first of all type II collagen). The type I collagen (COL1A2), type X collagen (COL10A1), Runt-related transcription factor 2 (RUNX2) and metalloproteinase 13 (MMP13) expression is also increased, what results in the final maturation of chondrocytes [99, 101]. After the mineralization of the cartilage, ECM serves as suitable scaffold for formation of the bones in the process called endochon-dral ossification. The remaining ECM is infiltrated by blood vessels and populated by bone marrow mesenchymal cells. RUNX2 and Osterix are one of the crucial genes responsible for differentiation of mesenchyme into osteoblasts and further osteoclasts [102]. The increased level of RUNX2 contributes to synthesis of type I collagen – one of component of bones, osteopontin (OPN) and osteocalcin (OCN). Differentiation and maturation of osteoblasts is maintained by high expression of OPN and bone sialoprotein (BSP). Further deposition of inorganic and organic component leads to the formation of osteocytes [103-105].
iPSC differentiation into chondrocyte-like cellsFor cartilage treatment, many approaches involving different pop-ulations of SCs: MSCs and hESCs were established. They include SC differentiation protocols based on micromasses, high density pellets, monolayer, EBs or combined various techniques, i.e. mon-olayer with high density pellets or EBs combined with monolayer and micromasses. In the case of iPSC, the confirmation of effi-ciency and real usefulness of established protocols is still widely
investigated. Above-mentioned techniques of differentiation are crucial, because they reflect and mimic biology of formation of cartilage tissue during embryo development and reconstruct three dimensional (3D) structures of tissues. It was revealed, that 3D systems of in vitro culture of chondrocytes improves their prop-erties and decrease likelihood of undesirable dedifferentiation process characterized by decreased production of type II colla-gen, aggrecan with simultaneous increased production of type I, X collagen and the presence of hypertrophic phenotype [106]. The basic pro-chondrogenic medium consists of insulin – trans-ferrin – selenium (ITS), dexamethasone, L-ascorbic acid (both re-sponsible for increased expression of ECM in cartilage), L-proline (basic component of cartilage cells), sodium pyruvate, non-es-sential amino acids. Furthermore, medium is supplemented with defined pro-chondrogenic GFs, that mostly are members of TGF-β and Bone Morphogeneic Proteins (BMP) family [96]. In order to confirm the presence of chondrocyte-like cells af-ter differentiation, the gene expression of chondrocyte-related genes must be evaluated. SOX-trio (SOX9, SOX6, SOX5), COL2A1, ACAN genes are a key factors in the regulation of ECM cartilage. The cartilage function strongly depends on the appropriate deposition of ECM compounds, therefore immunohistochem-istry techniques are very useful to detect specific compounds of fibrous components. For cartilage, the alcian blue (intense blue), Safranin-O (intense red), toluidine blue (intense purple) stains are routinely performed. They enable the detection of polysaccharides responsible for the appropriate biomechanical functions of cartilaginous tissues.Yang and co-workers (2012) implemented differentiation protocol of pluripotent SCs derived from keratinocytes (KiPSCs), which had been previously established by Oldershaw (2010) on hESC lines (HUES1, HUES8 and HUES9) [107, 108]. That protocol was executed onto matrigel – coated plates in the presence of growth defined medium mTeSR daily supplemented with other GFs. KiPSCs were differentiated for 14 days and the whole chondrogenic process was divided into two stages. Firstly, the mesoendoderm was ob-tained by culture of cells with Wingless-type MMTV integration site family member 3A (WNT3A), Activin A, BMP-4, fibroblast growth factor 2 (FGF-2) and follistatin. During the second stage, the chon-drogenesis was induced by growth/differentiation factor 5 (GDF5) belonging to the TGF-β family. It resulted in the high expression of SOX9 and COL2A1. Although, the expression of pluripotency markers had lower level in comparison with pluripotent SCs, their presence was clearly noticeable [108]. Nejadnik and colleagues (2015) induced mesenchymal-like cells from hiPSCs using 4 week-monolayer culture. HiPSC-derived mesenchymal cells were differentiated into chondrocyte-like cells for 3 weeks in pellet culture using chondrogenic medium supplemented with TGF-β3 (10 ng/ml). That approach resulted in the formation of chondrocyte – like cells, what was confirmed by immunohistochemistry and gene expression analysis of mark-ers characteristic for chondrocytes. The appropriate long culture period ensured significant decrease of pluripotency markers and resulted in non-teratoma regeneration of full-thickness of carti-lage in rat knee [109].

www.diagnostykalaboratoryjna.eu
132
Ko and colleagues (2014) established protocol that effectiveness was confirmed onto rat in vivo model. Firstly, the hiPSC cells were differentiated for 10 days via EBs. After that, EBs were dissociated and differentiated in micromass in chondrogenic medium sup-plemented with TGF-β3 (10 ng/ml) for 21 days. Those obtained cells revealed the characteristic features of hyaline cartilage. The gene expression of SOX9 and COL2A1 was elevated in differenti-ated cells demonstrating the obtaining of chondrocyte-like cells [23]. Moreover, the expression of genes (COL10A1, COL1A2 and RUNX2) responsible for hypertrophic cartilage was at very low level [110, 111].
iPSC differentiation into osteoblast-like cellsAlthough, osteoblast and osteocytes significantly differ from chondrocytes, the formation of these cell populations in vitro from SCs is based on very similar protocols. The combination of EBs formation and monolayer culture of pluripotent SCs are the most often used methods. The obtaining induced pluripotent stem cells derived mesenchymal stem cells (iPSC-MSC), is closely related to the first stages (e.g. formation of mesenchyme) of chon-dro – and osteogenesis naturally occurring in vivo during embryo development. In the majority of protocols, medium used for os-teogenic differentiation is relatively similar to chondrogenic one. The well-defined osteogenic medium is consisted of L-ascorbic acid, dexamethasone (both cause synthesis of type II collagen, but concentration of dexamethasone is higher in osteogenic medium, than in chondrogenic one) and β-glycerophosphate (activates alkaline phosphatase, triggering the deposition of minerals into the bones). However, in some protocols the addition of 1,25 – di-hydroxyvitamin D or GFs such as TGF-β and BMP family members remarkably improved osteogenic process in vitro [112-116]. In the osteoblast – like cells, the RUNX2 and Osterix – crucial regu-lators of mesenchyme differentiation into osteoblasts are widely evaluated. The presence of COL1A1, due to its high amount in the bone environment, is also commonly analysed to confirm the obtaining of osteoblast-like cells. Finally, in order to prove the maturation of osteoblasts accompanied by the organic and inorganic components, the alkaline phosphatase (ALP), OCN, BSP, and OPN gene expression are assessed. To demonstrate the de-sirable properties of obtained stem cell-derived osteoblasts and osteocytes, the immunohistochemistry analysis, such as staining alkaline phosphatase activity (intense dark blue colour observed in highly ALP active cells) and Alizarin red staining (intense red) is required. The interesting results were obtained by Yamamoto et al. (2015) who carried out the direct induction of fibroblast cells into osteo-blasts by transduction with the combination of four factors (ROXL): RUNX2, Osterix (both characteristic for osteogenic cells) OCT4 and L-MYC. That approach enabled avoiding the additional steps such as derivation of pluripotent SCs by transduction with Yamanaka factors and then differentiation them into desired cell lineage and this assures the direct receiving of osteoblast – cell like cells. The gene expression analysis (microarray analysis) and Alizarin Red S staining indicated high correlations between ROXL-derived and primary osteoblasts indicating the close similarity of these two
cell populations. Finally, the use of osteoblast-like cells resulted in complete repair of femoral diaphysis in NOC/SCID mice. [117].Tang and collaborators (2014) received osteoblasts from repro-grammed adult bone marrow CD34+ cells to hiPSCs. Then, the formed EB were differentiated into mesenchymal stem cell like lineage. For induction of osteogenic differentiation, the osteo-genic medium with addition of 1,25 – dihydroxyvitamin was ap-plied. During differentiation of hiPSC-MSC, the calcium phosphate cements (CPC) as a scaffold was also used. After differentiation, the cells cultivated on CPC in osteogenic medium indicated high deposition of minerals and the ALP, RUNX2, COL1A2, OCN high expression level. [114, 115]. Another approach involved receiving the osteoblast-like cells from 7 days EBs and subsequent 2 weeks monolayer culture. The exposition of cells to TGF-β, IGF-1 and FGF-2 improved osteogenic differentiation of hiPSCs. The selection of pure population of os-teogenic cells based on expression of tissue-nonspecific alkaline phosphatase (TNAP) led to obtaining high amount of cells with an osteogenic potential. Further propagation of those derived cells in osteogenic medium for 40 days led to increased expression of genes related to osteoblasts such as OSX, RUNX2, COL1A1, OCN and BSP. Alizarin red staining and alkaline phosphatase activity confirmed osteogenic potential of obtaining cells [118].
ConclusionsThe application of stem cells (SCs) in regenerative medicine has grown rapidly in recent years due to the enormous potential of these cells to treat a variety of disorders. Although numerous types of SCs exhibit differentiation potential, the development of human induced pluripotent stem cells (hiPSCs) is considered a turning point in tissue engineering. They have capacity to un-restricted self-renewal and capacity to develop into derivatives of all three primary germ layer (pluripotency). These properties make them a promising candidate for obtaining patient-specific neurons, cardiomyo-, chondro – and osteocytes. Nevertheless, the hiPSC – based therapy is a novel strategy and thus the further and intense research is required.
Funding for this work was supported by the National Science Centre[grant number 2012/07/E/NZ3/01819]
References1. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured
in medium conditioned by teratocarcinoma stem cells. Proceedings of the Na-
tional Academy of Sciences of the United States of America. 1981; 78: 7634-7638.
2. Evans MJ, Kaufman MH. Es lishment in culture of pluripotential cells from mouse
embryos. Nature. 1981; 292: 154-156.
3. Thomson JA, Itskovitz-Eldor J, Shapiro SS et al. Embryonic stem cell lines derived
from human blastocysts. Science. 1998; 282: 1145-1147.
4. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse em-
bryonic and adult fibroblast cultures by defined factors. Cell. 2006; 126: 663-676.
5. Takahashi K, Tanabe K, Ohnuki M et al. Induction of pluripotent stem cells from
adult human fibroblasts by defined factors. Cell. 2007; 131: 861-872.
6. Wu Z, Luby-Phelps K, Bugde A et al. Capacity for stochastic self-renewal and
differentiation in mammalian spermatogonial stem cells. The Journal of cell
biology. 2009; 187: 513-524.

133
Diagn Lab 2016; 52(2): 123-136
7. Cantz T, Martin U. Induced pluripotent stem cells: characteristics and perspec-
tives. Advances in biochemical engineering/biotechnology. 2010; 123: 107-126.
8. Chen C, Xiao SF. Induced pluripotent stem cells and neurodegenerative diseases.
Neuroscience bulletin. 2011; 27: 107-114.
9. Lengerke C, Daley GQ. Disease models from pluripotent stem cells. Annals of
the New York Academy of Sciences. 2009; 1176: 191-196.
10. Maherali N, Ahfeldt T, Rigamonti A et al. A high-efficiency system for the gen-
eration and study of human induced pluripotent stem cells. Cell stem cell.
2008; 3: 340-345.
11. Anchan RM, Quaas P, Gerami-Naini B et al. Amniocytes can serve a dual function
as a source of iPS cells and feeder layers. Human molecular genetics. 2011;
20: 962-974.
12. Stadtfeld M, Brennand K, Hochedlinger K. Reprogramming of pancreatic beta
cells into induced pluripotent stem cells. Current biology : CB. 2008; 18: 890-894.
13. Hanna J, Markoulaki S, Schorderet P et al. Direct reprogramming of terminally
differentiated mature B lymphocytes to pluripotency. Cell. 2008; 133: 250-264.
14. Su RJ, Baylink DJ, Neises A et al. Efficient generation of integration-free ips cells
from human adult peripheral blood using BCL-XL together with Yamanaka
factors. PloS one. 2013; 8: e64496.
15. Utikal J, Maherali N, Kulalert W et al. Sox2 is dispensable for the reprogramming
of melanocytes and melanoma cells into induced pluripotent stem cells. Journal
of cell science. 2009; 122: 3502-3510.
16. Cherry AB, Daley GQ. Reprogrammed cells for disease modeling and regener-
ative medicine. Annual review of medicine. 2013; 64: 277-290.
17. Hanna J, Wernig M, Markoulaki S et al. Treatment of sickle cell anemia mouse mod-
el with iPS cells generated from autologous skin. Science. 2007; 318: 1920-1923.
18. Xu D, Alipio Z, Fink LM et al. Phenotypic correction of murine hemophilia A using
an iPS cell-based therapy. Proceedings of the National Academy of Sciences of
the United States of America. 2009; 106: 808-813.
19. Polo JM, Anderssen E, Walsh RM et al. A molecular roadmap of reprogramming
somatic cells into iPS cells. Cell. 2012; 151: 1617-1632.
20. Soufi A, Donahue G, Zaret KS. Facilitators and impediments of the pluripotency
reprogramming factors’ initial engagement with the genome. Cell. 2012; 151:
994-1004.
21. Zhou W, Freed CR. Adenoviral gene delivery can reprogram human fibroblasts
to induced pluripotent stem cells. Stem cells. 2009; 27: 2667-2674.
22. Su RJ, Neises A, Zhang XB. Generation of iPS Cells from Human Peripheral Blood
Mononuclear Cells Using Episomal Vectors. Methods in molecular biology.
2016; 1357: 57-69.
23. Nishimura K, Sano M, Ohtaka M et al. Development of defective and persistent
Sendai virus vector: a unique gene delivery/expression system ideal for cell
reprogramming. The Journal of biological chemistry. 2011; 286: 4760-4771.
24. Okita K, Hong H, Takahashi K et al. Generation of mouse-induced pluripotent
stem cells with plasmid vectors. Nature protocols. 2010; 5: 418-428.
25. Kim D, Kim CH, Moon JI et al. Generation of human induced pluripotent stem cells
by direct delivery of reprogramming proteins. Cell stem cell. 2009; 4: 472-476.
26. Zhou H, Wu S, Joo JY et al. Generation of induced pluripotent stem cells using
recombinant proteins. Cell stem cell. 2009; 4: 381-384.
27. Warren L, Manos PD, Ahfeldt T et al. Highly efficient reprogramming to pluri-
potency and directed differentiation of human cells with synthetic modified
mRNA. Cell stem cell. 2010; 7: 618-630.
28. Kaji K, Norrby K, Paca A et al. Virus-free induction of pluripotency and subse-
quent excision of reprogramming factors. Nature. 2009; 458: 771-775.
29. Woltjen K, Michael IP, Mohseni P et al. piggyBac transposition reprograms fibro-
blasts to induced pluripotent stem cells. Nature. 2009; 458: 766-770.
30. Sommer CA, Sommer AG, Longmire TA et al. Excision of reprogramming trans-
genes improves the differentiation potential of iPS cells generated with a single
excisable vector. Stem cells. 2010; 28: 64-74.
31. Gossen M, Bujard H. Tight control of gene expression in mammalian cells by
tetracycline-responsive promoters. Proceedings of the National Academy of
Sciences of the United States of America. 1992; 89: 5547-5551.
32. Xu X, Smorag L, Nakamura T et al. Dppa3 expression is critical for generation of
fully reprogrammed iPS cells and maintenance of Dlk1-Dio3 imprinting. Nature
communications. 2015; 6: 6008.
33. Chan EM, Ratanasirintrawoot S, Park IH et al. Live cell imaging distinguishes bona
fide human iPS cells from partially reprogrammed cells. Nature biotechnology.
2009; 27: 1033-1037.
34. Silva J, Barrandon O, Nichols J et al. Promotion of reprogramming to ground
state pluripotency by signal inhibition. PLoS biology. 2008; 6: e253.
35. Mikkelsen TS, Hanna J, Zhang X et al. Dissecting direct reprogramming through
integrative genomic analysis. Nature. 2008; 454: 49-55.
36. Chen J, Liu H, Liu J et al. H3K9 methylation is a barrier during somatic cell
reprogramming into iPSCs. Nature genetics. 2013; 45: 34-42.
37. Liu L, Luo GZ, Yang W et al. Activation of the imprinted Dlk1-Dio3 region cor-
relates with pluripotency levels of mouse stem cells. The Journal of biological
chemistry. 2010; 285: 19483-19490.
38. Mattout A, Biran A, Meshorer E. Global epigenetic changes during somatic cell
reprogramming to iPS cells. Journal of molecular cell biology. 2011; 3: 341-350.
39. Loh KM, Chen A, Koh PW et al. Mapping the Pairwise Choices Leading from
Pluripotency to Human Bone, Heart, and Other Mesoderm Cell Types. Cell.
2016; 166: 451-467.
40. Ilic D, Devito L, Miere C et al. Human embryonic and induced pluripotent stem
cells in clinical trials. British medical bulletin. 2015; 116: 19-27.
41. Garber K. RIKEN suspends first clinical trial involving induced pluripotent stem
cells. Nature biotechnology. 2015; 33: 890-891.
42. Cunningham JJ, Ulbright TM, Pera MF et al. Lessons from human teratomas
to guide development of safe stem cell therapies. Nature biotechnology. 2012;
30: 849-857.
43. Ebert AD, Yu J, Rose FF, Jr. et al. Induced pluripotent stem cells from a spinal
muscular atrophy patient. Nature. 2009; 457: 277-280.
44. Karumbayaram S, Novitch BG, Patterson M et al. Directed differentiation of
human-induced pluripotent stem cells generates active motor neurons. Stem
cells. 2009; 27: 806-811.
45. Lowry WE, Richter L, Yachechko R et al. Generation of human induced pluripo-
tent stem cells from dermal fibroblasts. Proceedings of the National Academy
of Sciences of the United States of America. 2008; 105: 2883-2888.
46. Kawasaki H, Mizuseki K, Nishikawa S et al. Induction of midbrain dopaminergic neu-
rons from ES cells by stromal cell-derived inducing activity. Neuron. 2000; 28: 31-40.
47. Sonntag KC, Pruszak J, Yoshizaki T et al. Enhanced yield of neuroepithelial pre-
cursors and midbrain-like dopaminergic neurons from human embryonic stem
cells using the bone morphogenic protein antagonist noggin. Stem cells. 2007;
25: 411-418.
48. Cooper O, Hargus G, Deleidi M et al. Differentiation of human ES and Parkinson’s
disease iPS cells into ventral midbrain dopaminergic neurons requires a high
activity form of SHH, FGF8a and specific regionalization by retinoic acid. Mo-
lecular and cellular neurosciences. 2010; 45: 258-266.
49. Chambers SM, Fasano CA, Papapetrou EP et al. Highly efficient neural conver-
sion of human ES and iPS cells by dual inhibition of SMAD signaling. Nature
biotechnology. 2009; 27: 275-280.
50. Denham M, Dottori M. Neural differentiation of induced pluripotent stem cells.
Methods in molecular biology. 2011; 793: 99-110.
51. Hu BY, Weick JP, Yu J et al. Neural differentiation of human induced pluripotent
stem cells follows developmental principles but with variable potency. Pro-
ceedings of the National Academy of Sciences of the United States of America.
2010; 107: 4335-4340.
52. Berry BJ, Akanda N, Smith AS et al. Morphological and functional characteriza-
tion of human induced pluripotent stem cell-derived neurons (iCell Neurons) in
defined culture systems. Biotechnology progress. 2015; 31: 1613-1622.
53. Odawara A, Saitoh Y, Alhebshi AH et al. Long-term electrophysiological activity
and pharmacological response of a human induced pluripotent stem cell-de-
rived neuron and astrocyte co-culture. Biochemical and biophysical research
communications. 2014; 443: 1176-1181.
54. Xu X, Lei Y, Luo J et al. Prevention of beta-amyloid induced toxicity in human iPS
cell-derived neurons by inhibition of Cyclin-dependent kinases and associated
cell cycle events. Stem cell research. 2013; 10: 213-227.
55. Hargis KE, Blalock EM. Transcriptional signatures of brain aging and Alzheimer’s
disease: What are our rodent models telling us? Behavioural brain research.
2016.
56. Gomez-Nicola D, Boche D. Post-mortem analysis of neuroinflammatory changes
in human Alzheimer’s disease. Alzheimer’s research & therapy. 2015; 7: 42.
57. Szlachcic WJ, Switonski PM, Krzyzosiak WJ et al. Huntington disease iPSCs show
early molecular changes in intracellular signaling, the expression of oxidative
stress proteins and the p53 pathway. Disease models & mechanisms. 2015; 8:
1047-1057.

www.diagnostykalaboratoryjna.eu
134
58. Shi Y, Kirwan P, Smith J et al. Human cerebral cortex development from pluri-
potent stem cells to functional excitatory synapses. Nature neuroscience. 2012;
15: 477-486, S471.
59. Yagi T, Ito D, Okada Y et al. Modeling familial Alzheimer’s disease with induced
pluripotent stem cells. Human molecular genetics. 2011; 20: 4530-4539.
60. Nieweg K, Andreyeva A, van Stegen B et al. Alzheimer’s disease-related amy-
loid-beta induces synaptotoxicity in human iPS cell-derived neurons. Cell death
& disease. 2015; 6: e1709.
61. Consortium HDi. Induced pluripotent stem cells from patients with Hunting-
ton’s disease show CAG-repeat-expansion-associated phenotypes. Cell stem
cell. 2012; 11: 264-278.
62. An MC, Zhang N, Scott G et al. Genetic correction of Huntington’s disease phe-
notypes in induced pluripotent stem cells. Cell stem cell. 2012; 11: 253-263.
63. Tanzi RE, Bertram L. Twenty years of the Alzheimer’s disease amyloid hypothesis:
a genetic perspective. Cell. 2005; 120: 545-555.
64. Kobayashi Y, Okada Y, Itakura G et al. Pre-evaluated safe human iPSC-derived
neural stem cells promote functional recovery after spinal cord injury in com-
mon marmoset without tumorigenicity. PloS one. 2012; 7: e52787.
65. Josowitz R, Carvajal-Vergara X, Lemischka IR et al. Induced pluripotent stem
cell-derived cardiomyocytes as models for genetic cardiovascular disorders.
Current opinion in cardiology. 2011; 26: 223-229.
66. Itzhaki I, Rapoport S, Huber I et al. Calcium handling in human induced pluripo-
tent stem cell derived cardiomyocytes. PloS one. 2011; 6: e18037.
67. Zwi L, Caspi O, Arbel G et al. Cardiomyocyte differentiation of human induced
pluripotent stem cells. Circulation. 2009; 120: 1513-1523.
68. Tanaka T, Tohyama S, Murata M et al. In vitro pharmacologic testing using hu-
man induced pluripotent stem cell-derived cardiomyocytes. Biochemical and
biophysical research communications. 2009; 385: 497-502.
69. Di Pasquale E, Song B, Condorelli G. Generation of human cardiomyocytes:
a differentiation protocol from feeder-free human induced pluripotent stem
cells. Journal of visualized experiments : JoVE. 2013.
70. Ma D, Wei H, Lu J et al. Generation of patient-specific induced pluripotent stem
cell-derived cardiomyocytes as a cellular model of arrhythmogenic right ven-
tricular cardiomyopathy. European heart journal. 2013; 34: 1122-1133.
71. Liang P, Lan F, Lee AS et al. Drug screening using a library of human induced
pluripotent stem cell-derived cardiomyocytes reveals disease-specific patterns
of cardiotoxicity. Circulation. 2013; 127: 1677-1691.
72. Kim JJ, Yang L, Lin B et al. Mechanism of automaticity in cardiomyocytes derived
from human induced pluripotent stem cells. Journal of molecular and cellular
cardiology. 2015; 81: 81-93.
73. Hick A, Wattenhofer-Donze M, Chintawar S et al. Neurons and cardiomyocytes
derived from induced pluripotent stem cells as a model for mitochondrial
defects in Friedreich’s ataxia. Disease models & mechanisms. 2013; 6: 608-621.
74. Ma J, Guo L, Fiene SJ et al. High purity human-induced pluripotent stem cell-de-
rived cardiomyocytes: electrophysiological properties of action potentials and
ionic currents. American journal of physiology Heart and circulatory physiology.
2011; 301: H2006-2017.
75. Jung G, Fajardo G, Ribeiro AJ et al. Time-dependent evolution of functional vs.
remodeling signaling in induced pluripotent stem cell-derived cardiomyocytes
and induced maturation with biomechanical stimulation. FASEB journal : official
publication of the Federation of American Societies for Experimental Biology.
2016; 30: 1464-1479.
76. Yang X, Rodriguez M, Pabon L et al. Tri-iodo-l-thyronine promotes the matura-
tion of human cardiomyocytes-derived from induced pluripotent stem cells.
Journal of molecular and cellular cardiology. 2014; 72: 296-304.
77. Guo L, Abrams RM, Babiarz JE et al. Estimating the risk of drug-induced proar-
rhythmia using human induced pluripotent stem cell-derived cardiomyocytes.
Toxicological sciences : an official journal of the Society of Toxicology. 2011;
123: 281-289.
78. Lee S, Lee HA, Choi SW et al. Evaluation of nefazodone-induced cardiotoxicity
in human induced pluripotent stem cell-derived cardiomyocytes. Toxicology
and applied pharmacology. 2016; 296: 42-53.
79. Fine M, Lu FM, Lin MJ et al. Human-induced pluripotent stem cell-derived
cardiomyocytes for studies of cardiac ion transporters. American journal of
physiology Cell physiology. 2013; 305: C481-491.
80. Rana P, Anson B, Engle S et al. Characterization of human-induced pluripo-
tent stem cell-derived cardiomyocytes: bioenergetics and utilization in safety
screening. Toxicological sciences : an official journal of the Society of Toxicology.
2012; 130: 117-131.
81. Zwi-Dantsis L, Huber I, Habib M et al. Derivation and cardiomyocyte differen-
tiation of induced pluripotent stem cells from heart failure patients. European
heart journal. 2013; 34: 1575-1586.
82. Kuusela J, Kujala VJ, Kiviaho A et al. Effects of cardioactive drugs on human
induced pluripotent stem cell derived long QT syndrome cardiomyocytes.
SpringerPlus. 2016; 5: 234.
83. Ivashchenko CY, Pipes GC, Lozinskaya IM et al. Human-induced pluripotent stem
cell-derived cardiomyocytes exhibit temporal changes in phenotype. American
journal of physiology Heart and circulatory physiology. 2013; 305: H913-922.
84. Kamakura T, Makiyama T, Sasaki K et al. Ultrastructural maturation of human-in-
duced pluripotent stem cell-derived cardiomyocytes in a long-term culture.
Circulation journal : official journal of the Japanese Circulation Society. 2013;
77: 1307-1314.
85. Citro L, Naidu S, Hassan F et al. Comparison of human induced pluripotent stem-
cell derived cardiomyocytes with human mesenchymal stem cells following
acute myocardial infarction. PloS one. 2014; 9: e116281.
86. Zhang J, Wilson GF, Soerens AG et al. Functional cardiomyocytes derived from
human induced pluripotent stem cells. Circulation research. 2009; 104: e30-41.
87. Sallam K, Li Y, Sager PT et al. Finding the rhythm of sudden cardiac death: new
opportunities using induced pluripotent stem cell-derived cardiomyocytes.
Circulation research. 2015; 116: 1989-2004.
88. Garstang SV, Stitik TP. Osteoarthritis: epidemiology, risk factors, and pathophys-
iology. American journal of physical medicine & rehabilitation / Association of
Academic Physiatrists. 2006; 85: S2-11; quiz S12-14.
89. Lou X. Induced Pluripotent Stem Cells as a new Strategy for Osteogenesis and
Bone Regeneration. Stem cell reviews. 2015; 11: 645-651.
90. Minas T, Gomoll AH, Rosenberger R et al. Increased failure rate of autologous
chondrocyte implantation after previous treatment with marrow stimulation
techniques. The American journal of sports medicine. 2009; 37: 902-908.
91. Mobasheri A, Kalamegam G, Musumeci G et al. Chondrocyte and mesenchy-
mal stem cell-based therapies for cartilage repair in osteoarthritis and related
orthopaedic conditions. Maturitas. 2014; 78: 188-198.
92. Dimitriou R, Jones E, McGonagle D et al. Bone regeneration: current concepts
and future directions. BMC medicine. 2011; 9: 66.
93. Oryan A, Alidadi S, Moshiri A et al. Bone regenerative medicine: classic options,
novel strategies, and future directions. Journal of orthopaedic surgery and
research. 2014; 9: 18.
94. Kanawa M, Igarashi A, Ronald VS et al. Age-dependent decrease in the chondro-
genic potential of human bone marrow mesenchymal stromal cells expanded
with fibroblast growth factor-2. Cytotherapy. 2013; 15: 1062-1072.
95. Si YL, Zhao YL, Hao HJ et al. MSCs: Biological characteristics, clinical applications
and their outstanding concerns. Ageing research reviews. 2011; 10: 93-103.
96. Lietman SA. Induced pluripotent stem cells in cartilage repair. World journal of
orthopedics. 2016; 7: 149-155.
97. Tsumaki N, Okada M, Yamashita A. iPS cell technologies and cartilage regener-
ation. Bone. 2015; 70: 48-54.
98. Mahmoudifar N, Doran PM. Chondrogenesis and cartilage tissue engineering:
the longer road to technology development. Trends in biotechnology. 2012;
30: 166-176.
99. Zuscik MJ, Hilton MJ, Zhang X et al. Regulation of chondrogenesis and chon-
drocyte differentiation by stress. The Journal of clinical investigation. 2008;
118: 429-438.
100. Goldring MB. Chondrogenesis, chondrocyte differentiation, and articular car-
tilage me olism in health and osteoarthritis. Therapeutic advances in muscu-
loskeletal disease. 2012; 4: 269-285.
101. Csaki C, Matis U, Mobasheri A et al. Chondrogenesis, osteogenesis and adi-
pogenesis of canine mesenchymal stem cells: a biochemical, morphological
and ultrastructural study. Histochemistry and cell biology. 2007; 128: 507-520.
102. Mackie EJ, Ahmed YA, Tatarczuch L et al. Endochondral ossification: how carti-
lage is converted into bone in the developing skeleton. The international journal
of biochemistry & cell biology. 2008; 40: 46-62.
103. Heino TJ, Hentunen TA. Differentiation of osteoblasts and osteocytes from
mesenchymal stem cells. Current stem cell research & therapy. 2008; 3: 131-145.
104. Chen G, Deng C, Li YP. TGF-beta and BMP signaling in osteoblast differenti-
ation and bone formation. International journal of biological sciences. 2012;
8: 272-288.

135
Diagn Lab 2016; 52(2): 123-136
105. Bruderer M, Richards RG, Alini M et al. Role and regulation of RUNX2 in osteo-
genesis. European cells & materials. 2014; 28: 269-286.
106. Foldager CB, Nielsen AB, Munir S et al. Combined 3D and hypoxic culture im-
proves cartilage-specific gene expression in human chondrocytes. Acta ortho-
paedica. 2011; 82: 234-240.
107. Yang SL, Harnish E, Leeuw T et al. Compound screening platform using human
induced pluripotent stem cells to identify small molecules that promote chon-
drogenesis. Protein & cell. 2012; 3: 934-942.
108. Oldershaw RA, Baxter MA, Lowe ET et al. Directed differentiation of human
embryonic stem cells toward chondrocytes. Nature biotechnology. 2010; 28:
1187-1194.
109. Nejadnik H, Diecke S, Lenkov OD et al. Improved approach for chondrogenic
differentiation of human induced pluripotent stem cells. Stem cell reviews.
2015; 11: 242-253.
110. Ko JY, Kim KI, Park S et al. In vitro chondrogenesis and in vivo repair of oste-
ochondral defect with human induced pluripotent stem cells. Biomaterials.
2014; 35: 3571-3581.
111. Sun MM, Beier F. Chondrocyte hypertrophy in skeletal development, growth,
and disease. Birth defects research Part C, Embryo today : reviews. 2014; 102:
74-82.
112. Kato H, Ochiai-Shino H, Onodera S et al. Promoting effect of 1,25(OH)2 vitamin
D3 in osteogenic differentiation from induced pluripotent stem cells to osteo-
cyte-like cells. Open biology. 2015; 5: 140201.
113. Ko JY, Im GI. Chondrogenic and Osteogenic Induction from iPS Cells. Methods
in molecular biology. 2016; 1357: 441-450.
114. Tang M, Chen W, Liu J et al. Human induced pluripotent stem cell-derived
mesenchymal stem cell seeding on calcium phosphate scaffold for bone re-
generation. Tissue engineering Part A. 2014; 20: 1295-1305.
115. Wang P, Zhao L, Chen W et al. Stem Cells and Calcium Phosphate Cement Scaf-
folds for Bone Regeneration. Journal of dental research. 2014; 93: 618-625.
116. Illich DJ, Demir N, Stojkovic M et al. Concise review: induced pluripotent stem
cells and lineage reprogramming: prospects for bone regeneration. Stem cells.
2011; 29: 555-563.
117. Yamamoto K, Kishida T, Sato Y et al. Direct conversion of human fibroblasts into
functional osteoblasts by defined factors. Proceedings of the National Academy
of Sciences of the United States of America. 2015; 112: 6152-6157.
118. Ochiai-Shino H, Kato H, Sawada T et al. A novel strategy for enrichment and
isolation of osteoprogenitor cells from induced pluripotent stem cells based
on surface marker combination. PloS one. 2014; 9: e99534.
Corresponding author:mgr inż. Ewelina Augustyniak Greater Poland Cancer Centre Radiobiology Lab 61-866 Poznan, Garbary 15th Street tel: +48 61 8850474e-mail: [email protected]
Otrzymano: 9.06.2016Akceptacja do druku: 30.06.2016
Konflikt interesów: nie zgłoszono






























