Embed Size (px)
Citation preview

AEW
R
CUn
Apweiteasstuccsehecfdttsanattcvb
Kf
MtmEde
*1EAtl
Neuroscience 141 (2006) 1599–1609
0d
SPATIALLY STRUCTURED NETWORK OF INHIBITORY ANDXCITATORY CONNECTIONS DIRECTS IMPULSE TRAFFIC
ITHIN THE LATERAL AMYGDALApa(htult
vwGDantaLai1astaisita
(22sccrip2st1Mmat
. D. SAMSON AND D. PARÉ*
enter for Molecular and Behavioral Neuroscience, Rutgers Stateniversity, The State University of New Jersey, 197 University Ave-ue, Newark, NJ 07102, USA
bstract—The lateral nucleus of the amygdala is the entryoint of most sensory inputs into the amygdala. However, theay information is processed and distributed within the lat-ral nucleus still eludes us. To gain some insight into thisssue, we have examined the spatial organization of excita-ory and inhibitory connections in the lateral nucleus. To thisnd, we performed whole-cell recordings of principal lateralmygdala neurons and studied their responses to local pres-ure applications of glutamate in coronal and horizontallices of the guinea-pig amygdala. In coronal sections, glu-amate puffs performed at a distance from the recorded cellssually evoked inhibitory responses, except when the re-orded neuron was adjacent to the external capsule, in whichase excitatory responses could be evoked from ejectionites along the external capsule. In contrast, glutamate puffsvoked a mixture of excitatory and inhibitory responses inorizontal slices. Excitatory responses were generallyvoked from stimulation sites located lateral to the recordedell whereas inhibitory responses were commonly elicitedrom medial stimulation sites, irrespective of their rostrocau-al position. These findings confirm and extend previousract-tracing studies where it was found that intrinsic connec-ions within the lateral amygdala prevalently run in the dor-oventral and lateromedial directions. However, our resultslso reveal a hitherto unsuspected level of spatial heteroge-eity in the intrinsic circuit of the lateral amygdala. The prev-lence of excitatory responses in horizontal slices coupled tohe ubiquity of inhibitory responses in coronal slices suggesthat the lateral amygdala network is designed to allow asso-iative interactions within the rostrocaudal plane while pre-enting runaway excitation locally. © 2006 IBRO. Publishedy Elsevier Ltd. All rights reserved.
ey words: inhibition, glutamate, picrotoxin, microcircuit,ear, epilepsy.
any data indicate that the amygdala plays a critical role inhe acquisition, consolidation, and expression of emotionalemories (Davis, 2000; LeDoux, 2000; McGaugh, 2000;veritt et al., 2003). The basolateral complex of the amyg-ala (BLA), and particularly its lateral (LA) nucleus, hasmerged as a critical component for these functions, in
Corresponding author. Tel: �1-973-353-1080x3251; fax: �1-973-353-255.-mail address: [email protected] (D. Paré).bbreviations: BLA, basolateral amygdaloid complex; EPSP, excita-
tory postsynaptic potential; IPSP, inhibitory postsynaptic potential; LA,ateral amygdala; PV, parvalbumin.
306-4522/06$30.00�0.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2006.04.077
1599
art because it represents the main input station of themygdala for sensory inputs from the thalamus and cortexRusschen, 1986; LeDoux et al., 1990; Turner and Herken-am, 1991; McDonald, 1998). However, the transforma-ions performed by the LA on its sensory afferents remainnclear because our understanding of its intrinsic circuit is
imited. The present study was undertaken to shed light onhis issue.
There are two main types of neurons in the BLA (re-iewed in McDonald, 1992a): (1) multipolar projection cellsith spiny dendrites and highly collateralized axons and (2)ABAergic local-circuit neurons (McDonald, 1985; Mc-onald and Augustine, 1993; Paré and Smith, 1993) thatre aspiny (or sparsely spiny) and express various combi-ations of peptides (Kemppainen and Pitkänen, 2000; Ka-ona et al., 2001; McDonald and Betette, 2001; Mascagnind McDonald, 2003). Projection cells account for 80% ofA neurons (McDonald, 1992b), they use glutamate (Smithnd Paré, 1994) as a transmitter, and they contribute most
nter-nuclear projections of the amygdala (Stefanacci et al.,992; Smith and Paré, 1994). Local-circuit cells constituteheterogeneous group of neurons. According to the clas-
ification of McDonald (Mascagni and McDonald, 2003),here are at least four subtypes of BLA interneurons: parv-lbumin (PV) expressing neurons, somatostatin positive
nterneurons, large cholecystokinin interneurons, andmall bitufted cells that display various degrees of colocal-
zation of cholecystokinin, calretinin and vasoactive intes-inal polypeptide (McDonald and Pearson, 1989; Mascagnind McDonald, 2003).
Based on evidence obtained in the hippocampusFreund and Buzsáki, 1996; Somogyi and Klausberger,005) and neocortex (Markram et al., 2004; Blatow et al.,005), it is likely that these various types of interneuronstructure the activity of principal cells in distinct ways be-ause they target different postsynaptic domains and re-eive contrasting sets of inputs. For instance, PV interneu-ons of the BLA appear to be mainly involved in feedbacknhibition: they receive strong excitatory inputs from BLArojection cells but very few from the cortex (Smith et al.,000) and they form numerous inhibitory synapses on theomatic, axonal, and proximal dendritic domains of projec-ion neurons (Pitkänen and Amaral, 1993; Sorvari et al.,995; Smith et al., 1998; Kemppainen and Pitkänen, 2000;cDonald and Betette, 2001). Although their identity re-ains unclear, other types of interneurons likely gener-te feedforward inhibition in the BLA since cortical or
halamic stimulation evokes inhibitory postsynaptic po-
entials (IPSPs) in principal cells (Rainnie et al., 1991;ved.
W1ata2
iwdibtbacerwi
P
CHmEsgwadaKgm2pTb
D
WweMjjposesimttbt
pvw
Tagoarimi11
tbbsguaet
F(awgrwov
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–16091600
ashburn and Moises, 1992b; Lang and Paré, 1997a,b,998; Danober and Pape, 1998; Bissière et al., 2003)nd presumed interneurons can be orthodromically ac-ivated by such stimuli (Lang and Paré, 1998; Mahantynd Sah, 1998; Szinyei et al., 2000; Bauer and LeDoux,004).
Another outstanding question is whether the intrinsicnhibitory system of the BLA is spatially homogeneous orhether it constrains propagation of impulses in particularirections. In a previous study, we showed that feedback
nhibition is spatially heterogeneous in the LA, with feed-ack interneurons inhibiting only projection cells located in
he same coronal plane, leaving rostrocaudal interactionsetween principal cells relatively unrestricted (Samson etl., 2003). However, our previous study only examined theontribution of feedback interneurons. The present reportxtends this investigation to other types of BLA interneu-ons using local pressure applications of glutamate andhole-cell recordings of principal cells in coronal and hor-
zontal slices of the guinea-pig amygdala.
EXPERIMENTAL PROCEDURES
reparation of amygdala slices
oronal and horizontal slices of the amygdala were obtained fromartley guinea-pigs of either sex (250–300 g). Every effort wasade to minimize the number of animals used and their suffering.xperiments were done with the approval of the Rutgers Univer-ity Institutional Research Board and in accordance with the NIHuide for the care and use of laboratory animals. The animalsere deeply anesthetized with pentobarbital (60 mg/kg, i.p.), ket-mine (80 mg/kg, i.p.) and xylazine (12 mg/kg, i.p.) and thenecapitated. The brain was extracted from the skull and placed inn oxygenated solution (4 °C) containing (in mM) 126 NaCl, 2.5Cl, 1.25 NaH2PO4, 1 MgCL2, 2 CaCL2, 26 NaHCO3, and 10lucose. Brain slices (400 �m) were prepared using a vibratingicrotome and stored for one hour in an oxygenated chamber at3 °C. One slice was then transferred to a recording chambererfused with an oxygenated physiological solution (4 ml/min).he temperature of the chamber was gradually increased to 32 °Cefore the recordings began.
ata recording and analysis
hole-cell recordings were obtained with borosilicate pipettes filledith a solution containing (in mM) 130 K-gluconate, 10 N-2-hydroxy-thylpiperazine-N=-2-ethanesulfonic acid, 10 KCl, 2 MgCl2, 2 ATP-g, and 0.2 GTP-tris(hydroxy-methyl)aminomethane. pH was ad-
usted to 7.2 with KOH and osmolarity to �280 mOsm. The liquidunction potential was 10 mV with this solution and the membraneotential was corrected accordingly. The pipettes had a resistancef 4–8 M� when filled with the above solution. Recordings witheries resistance higher than 15 M� were discarded. In somexperiments, Neurobiotin (0.2%) was added to the intracellularolution to visualize the recorded neurons. Current clamp record-ngs were obtained with an Axoclamp 2B amplifier (Axon Instru-
ents, Foster City, CA, USA) under visual control using differen-ial interference contrast and infrared video microscopy. Concen-rations of drugs applied in the perfusate were (in �M) 10icuculline hydrochloride, 100 picrotoxin, 100 CGP-35348 (Mar-ina et al., 2001).
We studied the responses of principal LA neurons to localressure applications (12 p.s.i., 60 ms) of glutamate performed atarious positions with respect to the recorded cells. These tests
ere performed in coronal (Fig. 1A) and horizontal slices (Fig. 1B). phe glutamate puffs were performed via a patch pipette filled withrtificial cerebrospinal fluid containing 0.5 mM glutamate. Thelutamate concentration and stimulation parameters were chosenn the basis of previous experiments with this technique (Royer etl., 2000). Tests conducted with these stimulation parametersevealed that the effect of glutamate is extremely circumscribedn space: direct responses vanish when the ejection pipette is
oved �25–50 �m from the somata of recorded cells. Accord-ngly, the glutamate ejection pipette was initially positioned00 �m from the recorded cells and then moved in steps of00 �m toward the periphery of the LA nucleus.
When studying the synaptic responses elicited by glutamate,he membrane potential of the recorded cells was set to �65 mVy intracellular current injection and at two more negative mem-rane potentials to determine the reversal potential of evoked re-ponses. Three stimuli were applied at each membrane potential. Alutamate ejection site was considered effective only if three consec-tive stimuli elicited responses of relatively constant latencies. Syn-ptic events with amplitudes �0.5 mV were ignored. Values arexpressed as means�S.E. To determine whether response ampli-udes between cell groups differed significantly, t-tests were com-
CELCEM
BL
BM
Rh
M
V
L
D
PUOT
LA
Rec.LA BL BM
ML
R
C
V
H
EC
CORONAL SLICE
B HORIZONTAL SLICE
A
Glu
GluGlu
Glu
Glu
Glu
Rec.
ig. 1. Scheme illustrating the approach used in this study. CoronalA) or horizontal (B) slices of the guinea-pig amygdala were preparednd whole-cell patch recordings of principal LA neurons (empty circle)ere obtained under visual guidance. Local pressure applications oflutamate were performed at various sites (filled circles) around theecorded cells. Glutamate was applied via a patch pipette (Glu) filledith artificial cerebrospinal fluid containing 0.5 mM glutamate. Therientation of the slices is indicated by the crosses (D, dorsal; V,entral; L, lateral; M, medial; R, rostral; C, caudal).
uted using a fixed level of significance (P�0.05). Analyses were
pwaio
M
Wwpa(6is
Us
D
BtDprtdp
Fop
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–1609 1601
erformed off-line with the software IGOR (Wavemetrics, Lake Os-ego, OR, USA), clampfit (Axon Instruments, Union City, CA, USA),nd homemade software running on personal microcomputers. The
nput resistance (Rin) of the cells was estimated in the linear portionf current–voltage plots.
orphological identification of recorded cells
hen recorded cells were dialyzed with neurobiotin, the slicesere removed from the chamber and fixed for 1–3 days in 0.1 Mhosphate buffer saline (pH 7.4) containing 2% paraformaldehydend 1% glutaraldehyde. Slices were then embedded in gelatin10%) and sectioned on a vibrating microtome at a thickness of0 –100 �m. Neurobiotin-filled cells were visualized by incubat-
ng the sections in the avidin– biotin– horseradish peroxidaseolution (ABC Elite Kit, Vector Laboratories, Burlingame, CA,
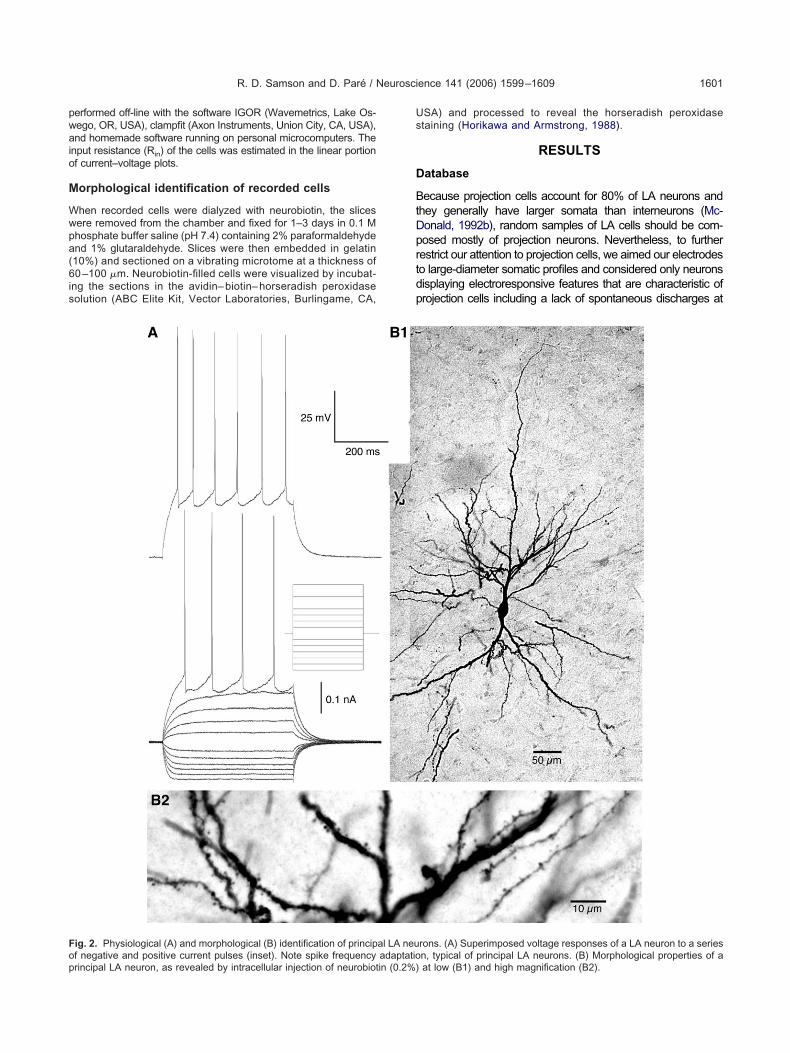
ig. 2. Physiological (A) and morphological (B) identification of princip
f negative and positive current pulses (inset). Note spike frequency adaptatiorincipal LA neuron, as revealed by intracellular injection of neurobiotin (0.2%)SA) and processed to reveal the horseradish peroxidasetaining (Horikawa and Armstrong, 1988).
RESULTS
atabase
ecause projection cells account for 80% of LA neurons andhey generally have larger somata than interneurons (Mc-onald, 1992b), random samples of LA cells should be com-osed mostly of projection neurons. Nevertheless, to furtherestrict our attention to projection cells, we aimed our electrodeso large-diameter somatic profiles and considered only neuronsisplaying electroresponsive features that are characteristic ofrojection cells including a lack of spontaneous discharges at
rons. (A) Superimposed voltage responses of a LA neuron to a series
al LA neu n, typical of principal LA neurons. (B) Morphological properties of aat low (B1) and high magnification (B2).
rcb1F
npzwnAtm2ams8
rs
Ta
Wndtpr(tp(bc
FtNcor rolonged
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–16091602
est and the generation of spike trains exhibiting frequency ac-ommodation upon membrane depolarization (Fig. 2A; Wash-urn and Moises, 1992a; Rainnie et al., 1993; Paré et al.,995; Lang and Paré, 1998; Pape and Driesang, 1998;aber et al., 2001).
Using these criteria, a total of 89 LA neurons with a Vm
egative to �60 mV and generating overshooting actionotentials were recorded (63 and 26 in coronal and hori-ontal slices, respectively). To confirm that these LA cellsere indeed principal cells, six neurons were filled witheurobiotin and their morphology was examined post hoc.s shown in the example of Fig. 2B, these cells displayed
he typical morphology of principal LA neurons includingultiple primary dendrites that branched extensively (Fig.B1) and bear a high density of spines (Fig. 2B2). Onverage, LA neurons had a resting potential of �82.2�0.7V, an input resistance of 182.3�12.9 M�, a time con-
tant of 44.1�3.2 ms, and generated action potentials of
20 mV
200 ms
5 mV
50 ms
Direct
Excitatory
Mixed
Inhibitory
RESPONSE TYPES
Dir
Ex
A B1
-100
-65
-90
-65
-90
-75
-65
ig. 3. Types of responses evoked by local pressure application of gluhe location of the ejection sites with respect to the recorded cells. Thesumbers in A indicate membrane potentials (in mV). Direct: when glutharacterized by a smooth rising phase (B). Excitatory: in some cases,f other principal neurons. Inhibitory: most commonly, glutamate puffsesponses consisted of an initial EPSP that was then followed by a p
2.0�1.1 mV lasting 1.16�0.04 ms at half amplitude. The r
esting properties of recorded cells did not vary with thelice orientation (t-tests, P�0.05).
ypes of responses evoked by local pressurepplication of glutamate
e begin by describing the types of responses evoked in LAeurons by local pressure application of glutamate. The inci-ence, reversal potential, and pharmacological sensitivity of
hese responses will be described later. Depending on theosition of the glutamate ejection pipettes with respect to theecorded cells, four types of responses could be observedFig. 3). As expected, application of glutamate in the vicinity ofhe soma or dendrites of LA neurons elicited a train of actionotentials riding on a smoothly rising depolarizing potentialFig. 3A, top). Since these responses were likely generatedy glutamate binding with receptors located on the recordedells, this type of response will hereafter be termed “direct
-50
5 mV
5 ms
2 mV
30 ms
y
Direct
Excitatory
Inhibitory
Mixed
B2
C
lutamate puffs could elicit different types of responses depending onresponse types are shown with a slow (A) and a fast (B–C) time base.s ejected directly onto the cell, a direct response was evoked. It wasmate puffs elicited a barrage of EPSPs, presumably via the activationlong IPSPs, presumably via the activation of local-circuit cells. MixedIPSP.
ect
citator
tamate. Ge variousamate wathe glutaelicited
esponse” (Fig. 3, top).

rvmpbpcpcpfssprftttem
e(bet(2ttctpmt“ItpC
Ri
BrlLKolrrptrc
tsrnpbsm
o(rs1(Asaeon
rttot5Asag
FGbaA(to
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–1609 1603
When the distance between the ejection pipette andecorded cell was increased, glutamate could evoke aariety of response types. These responses were likelyono- or disynaptic and their nature presumably de-ended on the type of LA cells that were depolarizedeyond threshold by the stimulus and their combined im-act on the recorded cell. “Excitatory responses” wereharacterized by the occurrence of a compound excitatoryostsynaptic potential (EPSP) composed of multiple dis-rete EPSPs that summated to generate an apparentlyure depolarizing response (Fig. 3, Excitatory). The lattereature was used to differentiate excitatory and direct re-ponses. “Inhibitory responses” were characterized by austained negative shift of the membrane potential with noreceding EPSPs (Fig. 3, Inhibitory). In contrast, “mixedesponses” consisted of an initial EPSP that was thenollowed by a prolonged IPSP (Fig. 3, Mixed). Note that theerms “excitatory” or “inhibitory” responses do not implyhat no IPSPs occurred during the former or EPSPs duringhe latter. These terms are only used to describe the netffect (direct or indirect) of the glutamate puffs on theembrane potential of recorded cells.
In contrast to the biphasic inhibitory responses typicallyvoked by electrical stimulation of afferents in LA neuronsRainnie et al., 1991; Washburn and Moises, 1992b; Dano-er and Pape, 1998; Martina et al., 2001), the IPSPslicited by glutamate puffs appeared monophasic irrespec-ive of the slice orientation and position of recorded cellspeak amplitude at �65 mV of 4.2�0.31; time to peak of41.1�13.3 ms). However, measuring the reversal poten-ial of glutamate-evoked IPSPs at latencies correspondingo the peaks of electrically-evoked GABA-A and GABA-Bomponents (based on previous studies) revealed thathey were also composed of two components: an earlyhase that reversed at �79.0�1.5 mV (measured at 100s) and a late one that reversed at more negative poten-
ials (�89.1�1.6 mV, measured at 400 ms; Fig. 3A,Inhibitory”). Consistent with this, the early part of thesePSPs was sensitive to the GABA-A antagonists picro-oxin or bicuculline (n�6; Fig. 4A–B) whereas the lateortion was sensitive to the GABA-B receptor antagonistGP 35348 (68.2�7.3% reduction, n�6; Fig. 4A).
esponses evoked by pressure-applied glutamaten coronal slices
elow, we will use the terms shell and core of the LA wheneferring to the position of recording or stimulation sitesocated along the external capsule or in the center of theA, respectively. This nomenclature was introduced byrettek and Price (1978), based on a histological analysisf the cat and rat LA nucleus. We studied the effects of
ocal pressure applications of glutamate in 63 LA neuronsecorded in coronal slices (core, 52; shell, 11). Theseecordings were homogenously distributed in the differentarts of the LA, as indicated in Fig. 5A, by the Xs marking
he position of recorded cells. Overall, 45 of these 63 cellsesponded to at least one stimulation site (36 of 52 in the
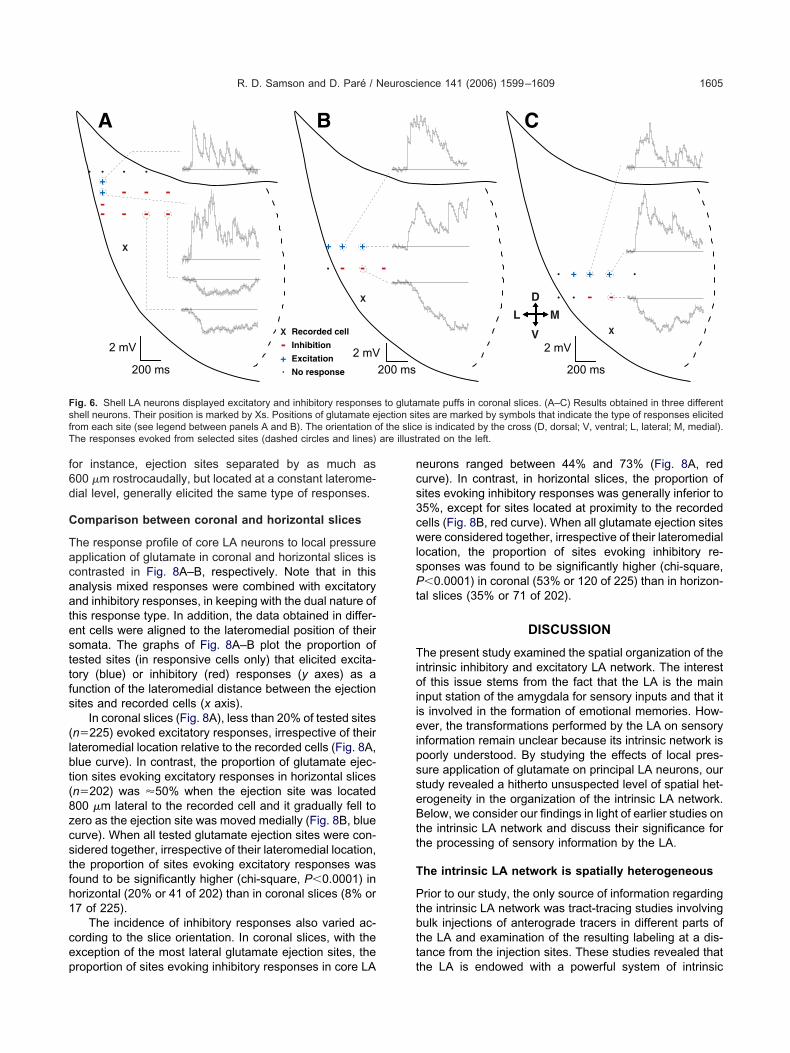
ore; nine of 11 in the shell). cIn coronal slices, we observed marked differences inhe responsiveness of principal LA neurons to local pres-ure application of glutamate depending on whether theecorded cells were located in the core or shell of the LAucleus. This point is illustrated in Figs. 5–6 where theosition of the glutamate ejection sites is marked by sym-ols that indicate the type of responses elicited from eachite (�, inhibitory response; �, excitatory response, *,ixed response; ., no response).
In the core of the LA nucleus, irrespective of the positionf the glutamate ejection sites with respect to recorded cellswhether they were medial, lateral, dorsal or ventral to theecorded cell), glutamate puffs typically evoked inhibitory re-ponses in coronal slices (87% of effective sites or 110 of27). This was true whether the cells were recorded mediallyFig. 5B–C), dorsally (Fig. 5D), or ventrally (Fig. 5E) in the LA.
few instances of excitatory (Fig. 5C, �) or mixed re-ponses (Fig. 5B–C, *) were observed, but they togetherccounted for a low proportion of responses evoked byffective sites (13%, or 17 of 127 effective sites) and werebserved in a minority of cells (excitatory, 8%; mixed, 11%;�36).
The inhibitory response pattern seen in core LA neu-ons contrasted with that observed in cells located alonghe external capsule (shell of the LA). As shown in thehree examples of Fig. 6, a significantly higher proportionf glutamate ejection sites elicited excitatory responses inhese cells (shell, 30% of effective sites, or 13 of 44; core:% of effective sites, or six of 127; chi-square, P�0.05).lthough this phenomenon was seen in all responsivehell neurons, inhibitory or mixed responses could be seens well. In general, such responses were evoked fromlutamate ejection sites located at proximity of the re-
2 mV
200 ms
1 mV
200 ms
Control Picrotoxin
CGP35348
Control
Picrotoxin
A
B
ig. 4. IPSPs evoked by local glutamate application are sensitive toABA receptor antagonists. (A) Example of IPSP evoked in a LA celly local pressure glutamate application in normal aCSF (Control), afterddition of picrotoxin to the aCSF, or of picrotoxin and CGP35348. (B)nother example of glutamate-evoked IPSP recorded in normal aCSF
Control) and after addition of picrotoxin in a different LA cell. Theraces in A and B represent the averages of two to four trials, all carriedut at a membrane potential of �65 mV.
orded cells, whereas excitatory responses were triggered

f(
Ri
IaiorttzpI
abceNetolaplte
FsLas
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–16091604
rom more distant sites dorsally along the external capsuleFig. 6).
esponses evoked by pressure-applied glutamaten horizontal slices
n horizontal slices, we studied the effects of local pressurepplications of glutamate in 26 LA neurons. These record-
ngs were homogeneously distributed in the different partsf the LA nucleus (Fig. 7A). Overall, 16 of these 26 cellsesponded to at least one stimulation site. In contrast withhe results obtained in coronal slices with core LA neurons,he type of responses evoked by glutamate puffs in hori-ontal slices varied systematically as a function of theosition of ejection sites with respect to recorded cells.
-
X
----
+ -
*
X- --
- -
-
-
- - -
C EL C EM
B L B M
MV
LD
P UO T
L A
A
C D
r h
X
+
*
Position of r
Site evoking
Site evoking
Site evoking
Site evoking
-
2 mV
200 ms
XX
X
X
X
X
X
X
XX
XX
XX
X
X X
XX
X
X X
X
XX
X
X
X
X X
XX
X
XX
X
X
X
XX
X
X
X
X
X X
X
X XX
X
X
X
X
X
X
X
XXX
X XX
ig. 5. In coronal slices, core LA neurons typically display inhibitory relice of the amygdala where the position of recorded cells is marked by, lateral; M, medial). (B–E) Results obtained in four different core LAre marked by symbols that indicate the types of responses elicited froelected sites (dashed circles and lines) are illustrated on the left.
ndeed, when the ejection pipette was moved lateromedi- t
lly from the external capsule toward the recorded cell andeyond, the prevalent type of responses shifted from ex-itatory to inhibitory or mixed responses. Three strikingxamples of this phenomenon are shown in Fig. 7B and D.ote that in these three cells, the most lateral ejection siteslicited excitatory responses whereas more medial ejec-ion sites evoked either inhibitory or mixed responses. Thisbservation also applied to LA cells located close to the
ateral (Fig. 7C) or medial (Fig. 7E) margins of the LA,lthough in such cases, the proximity of the LA borderrevented us from testing the effects of stimulation sites
ocated laterally or medially to the recorded cell, respec-ively. Moreover, the exact rostrocaudal position of thejection sites with respect to the recorded cell did not seem
-X -
-
--
-
-
-
* X
-
---
-
-
----
*-*
--
E
cell
ry response
ry response
esponse
onse
following pressure application of glutamate. (A) Scheme of a coronalorientation of the slice is indicated by the cross (D, dorsal; V, ventral;
. Their position is marked by Xs. Positions of glutamate ejection sitesite (see legend between panels A and B). The responses evoked from
--
B
ecorded
inhibito
excitato
mixed r
no resp
sponsesXs. The
neuronsm each s
o affect the type of responses they evoked. In Fig. 7C–E
f6d
C
Tacaatesttfs
(lbt(8zcstfh1
cep
ncs3cwlsPt
TioiieipsseBtt
T
Ptbtt
FsfT are illust
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–1609 1605
or instance, ejection sites separated by as much as00 �m rostrocaudally, but located at a constant laterome-ial level, generally elicited the same type of responses.
omparison between coronal and horizontal slices
he response profile of core LA neurons to local pressurepplication of glutamate in coronal and horizontal slices isontrasted in Fig. 8A–B, respectively. Note that in thisnalysis mixed responses were combined with excitatorynd inhibitory responses, in keeping with the dual nature ofhis response type. In addition, the data obtained in differ-nt cells were aligned to the lateromedial position of theiromata. The graphs of Fig. 8A–B plot the proportion ofested sites (in responsive cells only) that elicited excita-ory (blue) or inhibitory (red) responses (y axes) as aunction of the lateromedial distance between the ejectionites and recorded cells (x axis).
In coronal slices (Fig. 8A), less than 20% of tested sitesn�225) evoked excitatory responses, irrespective of theirateromedial location relative to the recorded cells (Fig. 8A,lue curve). In contrast, the proportion of glutamate ejec-ion sites evoking excitatory responses in horizontal slicesn�202) was �50% when the ejection site was located00 �m lateral to the recorded cell and it gradually fell toero as the ejection site was moved medially (Fig. 8B, blueurve). When all tested glutamate ejection sites were con-idered together, irrespective of their lateromedial location,he proportion of sites evoking excitatory responses wasound to be significantly higher (chi-square, P�0.0001) inorizontal (20% or 41 of 202) than in coronal slices (8% or7 of 225).
The incidence of inhibitory responses also varied ac-ording to the slice orientation. In coronal slices, with thexception of the most lateral glutamate ejection sites, the
+
X
--
+ +
-
X
+ -
-
-
--
-+
-
A B
2 mV2 mV
200 ms
X
+
Recorded cell
Inhibition
Excitation
No response
-
ig. 6. Shell LA neurons displayed excitatory and inhibitory responsehell neurons. Their position is marked by Xs. Positions of glutamate erom each site (see legend between panels A and B). The orientation ohe responses evoked from selected sites (dashed circles and lines)
roportion of sites evoking inhibitory responses in core LA t
eurons ranged between 44% and 73% (Fig. 8A, redurve). In contrast, in horizontal slices, the proportion ofites evoking inhibitory responses was generally inferior to5%, except for sites located at proximity to the recordedells (Fig. 8B, red curve). When all glutamate ejection sitesere considered together, irrespective of their lateromedial
ocation, the proportion of sites evoking inhibitory re-ponses was found to be significantly higher (chi-square,�0.0001) in coronal (53% or 120 of 225) than in horizon-
al slices (35% or 71 of 202).
DISCUSSION
he present study examined the spatial organization of thentrinsic inhibitory and excitatory LA network. The interestf this issue stems from the fact that the LA is the main
nput station of the amygdala for sensory inputs and that its involved in the formation of emotional memories. How-ver, the transformations performed by the LA on sensory
nformation remain unclear because its intrinsic network isoorly understood. By studying the effects of local pres-ure application of glutamate on principal LA neurons, ourtudy revealed a hitherto unsuspected level of spatial het-rogeneity in the organization of the intrinsic LA network.elow, we consider our findings in light of earlier studies on
he intrinsic LA network and discuss their significance forhe processing of sensory information by the LA.
he intrinsic LA network is spatially heterogeneous
rior to our study, the only source of information regardinghe intrinsic LA network was tract-tracing studies involvingulk injections of anterograde tracers in different parts ofhe LA and examination of the resulting labeling at a dis-ance from the injection sites. These studies revealed that
+
-
X
-
+
M
V
L
D
C
+
2 mV
200 ms
mate puffs in coronal slices. (A–C) Results obtained in three differenttes are marked by symbols that indicate the type of responses elicitede is indicated by the cross (D, dorsal; V, ventral; L, lateral; M, medial).rated on the left.
-
200 ms
s to glutajection sif the slic
he LA is endowed with a powerful system of intrinsic

clraPlb1d1ogo
aol
cnscpdsnjbp
eh(s
Fcios d sites (d
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–16091606
onnections that prevalently runs in the dorsoventral andateromedial directions, with prominent divergence in theostrocaudal axis (Krettek and Price, 1978; Stefanacci etl., 1992; Smith and Paré, 1994; Pitkänen et al., 1995;itkänen and Amaral, 1998). At the electron microscopic
evel, anterogradely labeled axons terminals were found toe enriched in glutamate but not GABA (Smith and Paré,994) and to form asymmetrical synapses, typically withendritic spines (Stefanacci et al., 1992; Smith and Paré,994). Since the dendrites of local-circuit cells are aspinyr sparsely spiny (McDonald, 1992a), these results sug-ested that the prevalent target of LA projection cells werether projection neurons.
However, a limitation of such studies is that they onlyllow examination of monosynaptic connections. More-ver, because the tracer injections are relatively large, only
X
-
-
--
-
-
---
-
-
--
-
--
+ + -
+ + + +
+ +
L A
B L B MML
R
C
V
H
E C
X
+
*Recorded cell
Inhibition
Excitation
Mixed response
No response-Direct Response
A
C D
400
X
XXXX
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
X
ig. 7. In horizontal slices, the responses of LA neurons to glutamateell in the lateromedial axis. (A) Horizontal slice of the amygdala wherndicated by the cross (R, rostral; C, caudal; L, lateral; M, medial). (B–Ef them are illustrated in panel B). Positions of glutamate ejection siteite (see legend below panel A). The responses evoked from selecte
ong-range connections could be examined, with the likely n
onsequence of overlooking the contribution of local-circuiteurons. By using local pressure of glutamate at variousites with respect to the recorded cells, we were able toircumvent these difficulties and examine the emergentroperties of the intrinsic LA network. With the exception ofirect responses, local glutamate application evoked re-ponses that were likely mono- or disynaptic and theirature presumably depended on the type of LA cells (pro-
ection cells and/or interneurons) that were depolarizedeyond threshold by the stimulus and their combined im-act on the recorded cell.
The most important observation to emerge from ourxperiments is that the intrinsic LA network is spatiallyeterogeneous, yet organized (Fig. 9). In coronal slicesFig. 9A), marked differences in the responsiveness ofhell vs. core LA neurons were observed. Whereas shell
X
-+
+ -
X
++ **-
*
X
+
-+
+
+
X
--
- --
B
E
V
pended on the position of the ejection pipette relative to the recordedition of recorded cells is marked by Xs. The orientation of the slice isobtained in five different neurons. Their position is marked by Xs (two
rked by symbols that indicate the type of responses elicited from thatashed circles and lines) are illustrated on the left.
-
-
4 m
ms
puffs dee the pos) Resultss are ma
eurons could often be excited by glutamate puffs per-

fntwMdtet
dpfssnsoaidicisowtp
rreTaob
bbswonfov
I
Octttnd2sp
thmsarhTor(de
Fsro
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–1609 1607
ormed at shell sites dorsal to the recorded cells, coreeurons were typically inhibited, irrespective of the location of
he glutamate ejection site (this, of course, excludes casesere glutamate was applied directly onto the recorded cells).oreover, in horizontal slices, the responses of LA neuronsepended on the position the glutamate ejection sites relative
he recorded cell, with lateral ejection sites generally evokingxcitatory responses (Fig. 9B1), whereas more medial onesypically triggered IPSPs (Fig. 9B2).
This contrasting responsiveness of core LA neuronsepending on the slice orientation is consistent with arevious report that focused on the spatial organization of
eedback inhibition in the LA (Samson et al., 2003). Thistudy took advantage of the fact that principal LA neuronsend a largely unidirectional projection to the basomedialucleus. By backfiring principal LA neurons from the ba-omedial nucleus and analyzing evoked responses inther LA projection cells, the targets of their intranuclearxons (projection cells vs. feedback interneurons) could be
nferred. As was seen here with glutamate puffs, basome-ial stimuli evoked markedly different synaptic responses
n LA neurons depending on the orientation of the slices. Inoronal slices, the typical response of LA neurons was annhibition. In horizontal slices, excitation prevailed (Sam-on et al., 2003). This suggested that the prevalent targetf the intrinsic axon collaterals of projection cells variesith the rostrocaudal position of target neurons relative to
he parent cell body: feedback interneurons at rostrocaudalroximity vs. other projection cells at a distance.
Although glutamate puffs also evoked more excitatoryesponses in horizontal than in coronal slices, inhibitoryesponses could be observed in horizontal slices when thejection sites were located medially to the recorded cells.he difference between the present and earlier (Samson etl., 2003) study likely results from the selective recruitmentf feedback interneurons by electrical basomedial stimuli,
0.8
0.6
0.4
0.2
0.0
0.2
0.4
0.6
-800 -400 0 400
CORONAL
Pro
port
ion
of te
sted
site
s
Lateromedial distanLateral Med
EPSPs
IPSPs
A
ig. 8. Contrasting response profile of core LA neurons in coronal (A)ites that evoked excitatory (blue) or inhibitory (red) responses (y axisecorded cells. On average, each data point is derived from 28.1 and 2f tested sites was 225 in coronal slices and 202 in horizontal slices.
y opposition to the recruitment of all types of interneurons V
y glutamate puffs in the LA. Nevertheless, it is clear fromoth sets of experiments that impulse propagation is notubjected to as much inhibition longitudinally comparedith the transverse plane. Moreover, the inhibition that isbserved in horizontal slices is not distributed homoge-eously. Excitation of lateral sites within the LA evokesewer inhibitory responses than that of medial sites. Thisbservation implies that the intrinsic inhibitory network fa-ors lateromedial propagation of excitatory impulses.
mplication for sensory processing within the LA
ur findings indicate that the connectivity of the shell andore of the LA nucleus differs: glutamate puffs in the shellend to excite shell neurons located ventral to the stimula-ion site whereas core LA neurons typically display inhibi-ory responses irrespective of the stimulation site in coro-al slices. Since inhibition was reported to gate activity-ependent synaptic plasticity in the LA (Bissière et al.,003), the prevalence of excitatory interactions in the shelluggests that this region of the LA nucleus may play arominent role in associative neuronal interactions.
In the core of the LA nucleus, the asymmetric distribu-ion of sites evoking excitatory and inhibitory responses inorizontal slices implies that sensory afferents terminatingedially in the core of the LA can block transmission of
ignals ending more laterally in the LA. Conversely, iffferent signals reach the lateral aspect of the LA first, theesponsiveness of more medial LA neurons might be en-anced by summation of the two excitatory responses.hus, depending on the relative timing and termination sitef sensory afferents in the LA, neuronal and behavioralesponses should vary. Interestingly, in cats and monkeysbut not in rats), cortical inputs conveying sensory inputs ofifferent modalities were observed to end at different lat-romedial levels in the LA (reviewed in McDonald, 1998).
0.8
0.6
0.4
0.2
-800 -400 0 400
HORIZONTAL
0.0
0.2
0.4
0.6
recorded cell (µm)Lateral Medial
EPSPs
IPSPs
zontal (B) slices. The two graphs plot the proportion of tested ejectionction of the lateromedial distance between the ejection sites and the
d sites in coronal and horizontal slices, respectively. The total number
Propo
rtio
n of
test
ed s
ites
ce fromial
B
and hori) as a fun5.3 teste
isual and auditory inputs end laterally whereas visceral,

adiontr
Abb
B
B
B
D
D
E
F
F
H
K
K
K
L
L
L
L
L
M
Fsr(tTfiietneatvptrfitsiiamediolateral axes.
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–16091608
nd gustatory inputs end medially. Our findings thus pre-ict that in these species, animals would be more likely to
gnore visual or auditory inputs when preceded by visceral,r gustatory stimuli. In other words, inscribed in the LAetwork, would be a predisposition to preferentially attendo stimuli that affect the individual directly compared withemote stimuli.
cknowledgments—This material is based upon work supportedy the National Science Foundation under grant No. 0208712 andy NIMH grant 066856.
REFERENCES
auer EP, LeDoux JE (2004) Heterosynaptic long-term potentiation ofinhibitory interneurons in the lateral amygdala. J Neurosci 24:9507–9512.
issière S, Humeau Y, Lüthi A (2003) Dopamine gates LTP inductionin lateral amygdala by suppressing feedforward inhibition. NatNeurosci 6:587–592.
latow M, Caputi A, Monyer H (2005) Molecular diversity of neocorticalGABAergic interneurones. J Physiol 562:99–105.
anober L, Pape HC (1998) Mechanisms and functional significanceof a slow inhibitory potential in neurons of the lateral amygdala. EurJ Neurosci 10:853–867.
avis M (2000) The role of the amygdala in conditioned and uncon-ditioned fear and anxiety. In: The amygdala: a functional analysis(Aggleton JP, ed), pp 213–287. Oxford: Oxford University Press.
veritt BJ, Cardinal RN, Parkinson JA, Robbins TW (2003) Appetitivebehavior: impact of amygdala-dependent mechanisms of emo-tional learning. Ann N Y Acad Sci 985:233–250.
aber E, Callister RJ, Sah P (2001) Morphological and electrophysi-ological properties of principal neurons in the rat lateral amygdalain vitro. J Neurophysiol 85:714–723.
reund TF, Buzsáki G (1996) Interneurons of the hippocampus. Hip-pocampus 6:347–470.
orikawa K, Armstrong WE (1988) A versatile means of intracellularlabeling: injection of biocytin and its detection with avidin conju-gates. J Neurosci Methods 25:1–11.
atona IN, Rancz EA, Acsády L, Ledent C, Mackie K, Hájos N, FreundTF (2001) Distribution of CB1 cannabinoid receptors in the amyg-dala and their role in the control of GABAergic transmission. J Neu-rosci 21:9506–9518.
emppainen S, Pitkänen A (2000) Distribution of parvalbumin, calretinin,and calbindin-D28k immunoreactivity in the rat amygdaloid complexand colocalization with gamma-aminobutyric acid. J Comp Neurol426:441–467.
rettek JE, Price JL (1978) A description of the amygdaloid complex inthe rat and cat with observations on intra-amygdaloid axonal con-nections. J Comp Neurol 178:255–280.
ang EJ, Paré D (1997a) Similar inhibitory processes dominate theresponses of cat lateral amygdaloid projection neurons to theirvarious afferents. J Neurophysiol 77:341–352.
ang EJ, Paré D (1997b) Synaptic and synaptically activated intrinsicconductances underlie inhibitory potentials in cat lateral amygda-loid projection neurons in vivo. J Neurophysiol 77:353–363.
ang EJ, Paré D (1998) Synaptic responsiveness of interneurons ofthe cat lateral amygdaloid nucleus. Neuroscience 83:877–889.
eDoux JE (2000) Emotion circuits in the brain. Annu Rev Neurosci23:155–184.
eDoux JE, Cicchetti P, Xagoraris A, Romanski LM (1990) The lateralamygdaloid nucleus: sensory interface of the amygdala in fearconditioning. J Neurosci 10:1062–1069.
ahanty NK, Sah P (1998) Calcium-permeable AMPA receptors me-diate long-term potentiation in interneurons in the amygdala. Na-
Medial
Rostral
CaudalLateralMedial
Dorsal
VentralLateral
A1
A2
B1
B2
Glutamatergic
GABAergic
Coronal Slice Horizontal Slice
Rostral Caudal
Dorsal
Ventral
Medial
Lateral
C
ig. 9. Pattern of intrinsic LA connections emerging from the presenttudy and earlier tract tracing investigations. (A, B) Schemes summa-izing the intrinsic connections observed in coronal (A) and horizontalB) slices. For clarity, excitatory (empty arrows, panels 1) and inhibi-ory (filled arrows, panels 2) connections are represented separately.he orientation of the slices is indicated by the cross at the top of thegures. In coronal slices, a different pattern of connections was seenn the core vs. shell of the lateral nucleus. In the shell, along thexternal capsule, evidence of excitatory (A1) and inhibitory connec-ions (A2) was obtained but inhibitory effects seemed to affect onlyeurons located at proximity of the glutamate ejection site whereasxcitatory effects could be elicited from more distant sites. In the core,lthough tract-tracing studies indicate that principal LA neurons con-ribute massive excitatory connections running from dorsolateral toentromedial (A1), local pressure application of glutamate evokedrevalently inhibitory responses irrespective of the relative position ofhe ejection sites with respect to the recorded cell. This presumablyesults from the presence of dense divergent GABAergic projectionsrom local-circuit neurons (A2). In horizontal slices, excitatory (B1) andnhibitory (B2) connections appeared to be oriented in opposite direc-ions (lateromedially vs. mediolaterally). However both showed exten-ive rostrocaudal divergence. (C) Three-dimensional rendering of thentrinsic inhibitory circuit of the core of the lateral nucleus. Gray arrowsndicate inhibitory axons fanning in a single coronal plane. Blackrrows represent inhibitory axons extending in the rostrocaudal or
ture 394:683–687.

M
M
M
M
M
M
M
M
M
M
M
P
P
P
P
P
P
R
R
R
R
S
S
S
S
S
S
S
S
T
W
W
R. D. Samson and D. Paré / Neuroscience 141 (2006) 1599–1609 1609
arkram H, Toledo-Rodriguez M, Wang Y, Gupta A, Silberberg G, WuC (2004) Interneurons of the neocortical inhibitory system. Nat RevNeurosci 5:793–807.
artina M, Royer S, Paré D (2001) Cell-type-specific GABA responsesand chloride homeostasis in the cortex and amygdala. J Neuro-physiol 86:2887–2895.
ascagni F, McDonald AJ (2003) Immunohistochemical characteriza-tion of cholecystokinin containing neurons in the rat basolateralamygdala. Brain Res 976:171–184.
cDonald AJ (1985) Immunohistochemical identification of gamma-aminobutyric acid-containing neurons in the rat basolateral amyg-dala. Neurosci Lett 53:203–207.
cDonald AJ (1992a) Cell types and intrinsic connections of theamygdala. In: The amygdala: Neurobiological aspects of emotion,memory, and mental dysfunction (Aggleton JP, ed), pp 67–96. NewYork: Wiley-Liss.
cDonald AJ (1992b) Projection neurons of the basolateral amygdala:A correlative Golgi and retrograde tract tracing study. Brain ResBull 28:179–185.
cDonald AJ (1998) Cortical pathways to the mammalian amygdala.Prog Neurobiol 55:257–332.
cDonald AJ, Pearson JC (1989) Coexistence of GABA and peptideimmunoreactivity in non-pyramidal neurons of the basolateralamygdala. Neurosci Lett 100:53–58.
cDonald AJ, Augustine JR (1993) Localization of GABA-like immu-noreactivity in the monkey amygdala. Neuroscience 52:281–294.
cDonald AJ, Betette RL (2001) Parvalbumin-containing neurons inthe rat basolateral amygdala: morphology and co-localization ofcalbindin-D(28k). Neuroscience 102:413–425.
cGaugh JL (2000) Memory: a century of consolidation. Science287:248–251.
ape HC, Driesang RB (1998) Ionic mechanisms of intrinsic oscilla-tions in neurons of the basolateral amygdaloid complex. J Neuro-physiol 79:217–226.
aré D, Smith Y (1993) Distribution of GABA immunoreactivity in theamygdaloid complex of the cat. Neuroscience 57:1061–1076.
aré D, Pape HC, Dong J (1995) Physiological properties of catbasolateral amygdaloid neurons: Intracellular recordings in barbi-turate-anesthetized cats. J Neurophysiol 74:1179–1191.
itkänen A, Stefanacci L, Farb CR, Go GG, LeDoux JE, Amaral DG(1995) Intrinsic connections of the rat amygdaloid complex: Pro-jections originating in the lateral nucleus. J Comp Neurol 356:288–310.
itkänen A, Amaral DG (1993) Distribution of parvalbumin-immunore-active cells and fibers in the monkey temporal lobe: The amygda-loid complex. J Comp Neurol 331:14–36.
itkänen A, Amaral DG (1998) Organization of the intrinsic connec-tions of the monkey amygdaloid complex: Projections originating in
the lateral nucleus. J Comp Neurol 398:431–458.ainnie DG, Asprodini EK, Shinnick-Gallagher P (1991) Inhibitorytransmission in the basolateral amygdala. J Neurophysiol 66:999–1009.
ainnie DG, Asprodini EK, Shinnick GP (1993) Intracellular recordingsfrom morphologically identified neurons of the basolateral amyg-dala. J Neurophysiol 69:1350–1362.
oyer S, Martina M, Paré D (2000) Polarized synaptic interactionsbetween intercalated neurons of the amygdala. J Neurophysiol 83:3509–3518.
usschen FT (1986) Cortical and subcortical afferents of the amyg-daloid complex. In: Excitatory amino acids and epilepsy (SchwarzR, Ben-Ari Y, eds), pp 35–52. New York: Plenum.
amson RD, Dumont ÉC, Paré D (2003) Feedback inhibition definestransverse processing modules in the lateral amygdala. J Neurosci23:1966–1973.
mith Y, Paré D (1994) Intra-amygdaloid projections of the lateralnucleus in the cat: PHA-L anterograde labeling combined withpost-embedding GABA and glutamate immunocytochemistry.J Comp Neurol 342:232–248.
mith Y, Paré JF, Paré D (1998) Cat intraamygdaloid inhibitorynetwork: Ultrastructural organization of parvalbumin-immunoreac-tive elements. J Comp Neurol 391:164–179.
mith Y, Paré JF, Paré D (2000) Differential innervation of parvalbu-min-immunoreactive interneurons of the basolateral amygdaloidcomplex by cortical and intrinsic inputs. J Comp Neurol 416:496–508.
omogyi P, Klausberger T (2005) Defined types of cortical interneu-rone structure space and spike timing in the hippocampus.J Physiol 562:9–26.
orvari H, Soininen H, Paljärvi L, Karkola K, Pitkänen A (1995) Dis-tribution of parvalbumin-immunoreactive cells and fibers in thehuman amygdaloid complex. J Comp Neurol 360:185–212.
tefanacci L, Farb CR, Pitkänen A, Go G, LeDoux JE, Amaral DG(1992) Projections from the lateral nucleus to the basal nucleus ofthe amygdala. J Comp Neurol 323:586–601.
zinyei C, Heinbockel T, Montagne J, Pape HC (2000) Putative cor-tical and thalamic inputs elicit convergent excitation in a populationof GABAergic interneurons of the lateral amygdala. J Neurosci20:8909–8915.
urner BH, Herkenham M (1991) Thalamoamygdaloid projections inthe rat: A test of the amygdala’s role in sensory processing.J Comp Neurol 313:295–325.
ashburn MS, Moises HC (1992a) Electrophysiological and morpho-logical properties of rat basolateral amygdaloid neurons in vitro.J Neurosci 12:4066–4079.
ashburn MS, Moises HC (1992b) Inhibitory responses of rat baso-lateral amygdaloid neurons recorded in vitro. Neuroscience 50:
811–830.(Accepted 28 April 2006)(Available online 6 June 2006)





























