Embed Size (px)
Citation preview

FEMS Microbiology Letters 69 (1990) 73-78 73 Published by Elsevier
FEMSLE 03984
A procedure for enrichment and isolation of mutants of the salt-tolerant yeast Debaryomyces hansenii having altered
glycerol metabolism
Carmen Morales, Lars Andr~ and Lennart Adler
Department of General and Marine Microbiology. UniL, ersJty o/G~tob~rg. Ggleborg. Sweden
Received and accepted 15 January 1990
Key words: Debaryomyces hansenii; Salt-tolerance; Osmoregulation; Glycerol production; Glycerol accumulation
1. SUMMARY
The salt-tolerant yeast Debaryomyces hansenii produces and accumulates glycerol when subjected to salt stress, whereby the buoyant density of the ceils is changed. This property allows for enrich- ment of mutants ~,ith altered glycerol metabolism by density gradient centrifugation. Colonies de- rived from cells with rapidly changing density following an osmotic shock were screened for in- creased glycerol production by observing their ability to support growth of a glycerol-requiring strain of Escherichia coli. The glycerol overproduc- ing phenotype of two isolates was confirmed by chemical analysis.
potential results in production and intraeellular accumulation of glycerol 11,2]. A substantial body of evidence indicates that glycerol is the main osmoregulatory solute used by this organism [1,3,41. Studies on its glycerol metabolism indicate that glycerol is synthesized via dihydroxyacetone phosphate, which is reduced to glycerol-3-phos- phate followed by dephosphorylation to glycerol [21. The utilization of glycerol occurs by reversal of this two-step pathway, via an alternative set of enzymes [2,51. To provide further insight in the glycerol metabolism and its regulation, a proce- dure for enrichment and isolation of mutants with altered glycerol production and accumulation was devised.
2. INTRODUCTION 3. MATERIAL AND METHODS
Exposure of the salt-tolerant yeast De- baryomyces hansenii to conditions of low water
Correspondence w: L. Adler. Department of General and Marine Microbiology. University of G~Steborg. Cad S/~Otls- bergsgata 22, S-413 19 G6teborg, Sweden. Part of this work was presented at the International Workshop on Mol~ular Biology of Stress Responses in Plants, O- bermalchtal, F.R.G.
3.1. Organisms and growth conditions Escherichia coil, strain BB20-14 [CGS 5424] [6]
was cultured in LB medium I7] supplemented with I mM glycerol. D. hansenii, strain J26 [8] and a mutant strain 26-6 [4], were grown in a defined medium composed as described by Adler and Gustafsson [1], with additional supplements as stated in the text. 1). hansenii starter cultures were prepared by inoculating 300-ml Erlenmeyer flasks
0378-1097/90/$03.50 a~ ]990 Federation of European Microbiological Societies

74
each containing 50 ml of 1% (w/v) glucose medium. These cultures were incubated for 48 h and used to inoculate 2.8-1 Fernbach flasks con- taining 500 ml of 0,5% (w/v) glucose medium. Liquid yeast cultures were incubated at 27 ° C, and E. coli cultures at 37*C, on rotary shakers. Solid media contained the defined yeast medium supp- lemented with 1% glucose and 2% Bacto (Difco) agar. This medium supported growth of E. coli BB20-14, provided glycerol was added. Growth of liquid cultures was monitored in 1-cm cuvettes at 610 nm in a Beckman B spectrophotometer. Dry weight was determined with ceils washed twice in distilled water and dried at 105 *C for 48 h, or from optical density readings using standard c u r v e s .
3.4. Gradient centrifugation Density gradients were prepared using Percoll
(Pharmacia Fine Chemicals, Sweden), Five parts of Percoll were mixed with six parts of 4.25 M sorbitol Io produce a solution slightly hyper- osmotic to the 85[ NaCI medium. A 1-ml sample from a culture concentration by a factor of 10 was layered on a 9 ml sorbitol-Percoll suspension in a 15-ml Falcon polypropylene tube. The tubes were centrifuged for 25 rain at 20000 × g in a Sorvall SM-24 fixed angle rotor, using a water cushion of 4 mi in each tube hole to prevent the tubes from cracking. To collect the cells after centrifugation, the tubes were punctured in the bottom, and 1-ml fractions were taken.
3.2. Mutagenesis D. hansenii J26 was inoculated into 50-ml liquid
medium in a 500-ml Erlenmeyer flask and grown overnight. A portion (0.8 ml) of the cells was washed and suspended in 1.7 ml sodium phos- phate buffer, pH 8,0 and 50 p,1 ethyl methane- sulfonate (Sigma) was added (cf. [9]). The tube was shaken in a Vortex mixer and incubated without further agitation for 40 rain at 30 ° C. This treatment gave 50% survival of the population. The mutagenized culture was centrifuged and the ceils rcsuspended in 8 ml 6% sodium thiosulfate. After 10 rain the cells were washed once and transferred to a 500-ml Erlenmeyer flask contain- ing 50 ml medium, and further incubated for 48 h at 27"C.
3.3. Transfer experimeuts Five ml of the matagenized cell culture were
used to inoculate a Fernbach flask containing 500 ml medium. This culture was incubated It) reach an OD610 of about 2,0, whereafter the cells were exposed to 8% NaCI by adding an equal volume of medium containing 16% NaC, Immediately after the NaCl addition, the culture volume was ad- justed to its original value (500 ml). The culture was incubated, and at various periods of time cell samples and removed and subjected to gradient centrifugation,
3.5. Density determination The buoyant density of the sampled fractions
was determined by applying small sample volumes to a calibrated gradient of kerosene and CCI 4 in a glass cylinder ll0,11 I. The gradient was prepared in a glass cylinder using a dual-chamber gradient maker with a dense solution, composed of 200 ml CCI 4 and 50 ml kerosene, and a light solution of 100 ml CCla and 150 ml kerosene. Drops added to the gradient rapidly sank to their buoyant den- sity level and the density was determined from a calibration curve based on the observed position of solutions of known density.
3.6. Polyol determination Glycerol was determined as described by Andr6
et al. I4] and total polyols as described by Adler and Gustafsson [1].
3, 7. Determination of cell ~,olume Cell volumes were estimated by measuring the
volume of sedimented cells of a given culture volume (5 to 10 ml) in cytocrit tubes containing calibrated capillaries of 20 p.l, permitting reading within 0.5/~!, All readings were performed after centrifugation for 15 rain at 3 500 X g, in a table- top centrifuge, The number of cells were de- termined by counting in a ,Heiber counting cham- ber.
"15
4. RESULTS
4.1 Changes in cell size and glycerol contem in response to salt stress
When exponentially growing cells of D. hansenii were transfered from basal medium to medium containing 8% NaCI there was a decrease in the average cell volume from 22 fl to 14 fl, as esti- mated from cell number and sedimentation volume in cytocrit tubes. However, after an adaption period of 4 h in the saline medium, the cells had regained their original volume (cf. [12]). During this period of adaption, the cells accumulated massive amounts of glycerol, the internal levels increasing from 0.05 to 3.1/~mol/mg dry wt yeast. These observations indicated a considerable dif- ference in buoyant density between the shocked, shrunken cells and the acclimated, glycerol rich ceils.
4.1. Density enrichment To examine whether gradient centrifugation
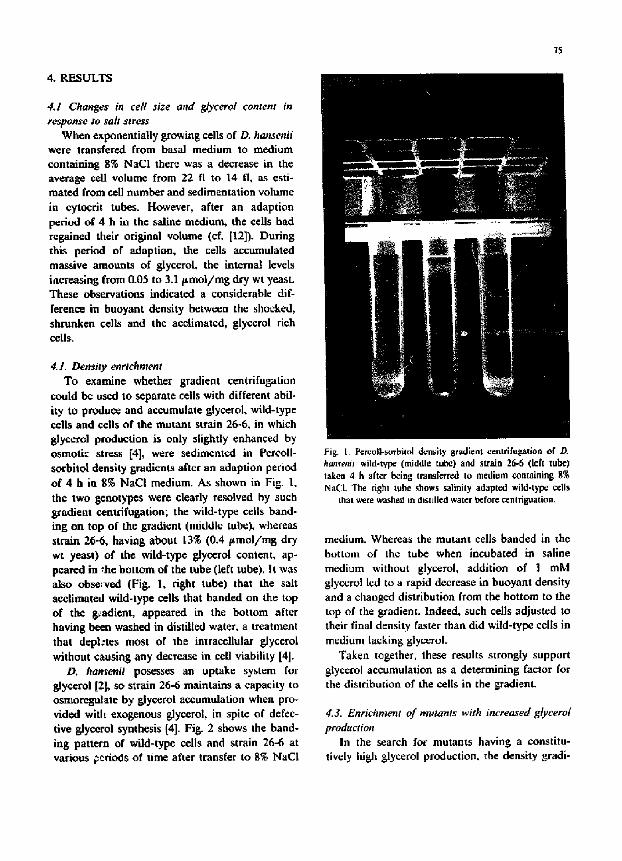
could be used to separate cells with different abil- ity to produce and accumulate glycerol, wild-type cells and cells of the mutant strain 26-6, in which glycerol production is only slightly enhanced by osmotic stress [4], were sedimented in Percoll- sorbitol density gradients after an adaption period of 4 h in 8~$ NaCi medium. As shown in Fig. 1, the two gcnotypes were clearly resolved by such gradient centdfugation; the wild-type cells band- ing on top of the gradient (middle tube), whereas strain 26-6, having about 13% (0.4 f tmol/mg dry wt yeast) of the wild-type glycerol content, ap- peared in "..he bottom of the tube (left tube). It was also observed (Fig. 1, right tube) that the salt acclimated wild-type cells that banded on the top of the g~adient, appeared in the bottom after having been washed in distilled water, a treatment that depi:tes most of the intraccllular glycerol without causing any decrease in cell viability [4].
D, hansenii posesses an uptake system for glycerol [2], so strain 26-6 maintains a capacity to osmoregulate by glycerol accumulation when pro- vided with exogenous glycerol, in spite of defec- tive glycerol synthesis [4]. Fig. 2 shows the band- ing pattern of wild-type cells and strain 26-6 at various periods of time after transfer to 8~, NaCi
Fig. 1. Pc~coll-smbitol density gradient cenlrifugatlon of D. hansemi wild-type (middle tube) and strain 26-6 (left tube) taken 4 h after being transferred to medium conlaining 8~$ NaCI÷ The right ~ube shows salinity adapted wild-type ¢¢11s
that were washed in distilled water before centriguation.
medium. Whereas the mutant cells banded in the bottom of the tube when incubated in saline medium without glycerol, addition of 1 mM glycerol led to a rapid decrease in buoyant density and a changed distribution from the bottom to the top of the gradient. Indeed, such cells adjusted to their final density faster than did wild-type cells in medium lacking glycerol.
Taken together, these results strongly support glycerol accumulation as a determining factor for the distribution of the cells in the gradient.
4.3. Enrichment Qf mutants with incregsed glycerol production
In the search fur mutants having a constitu- tively high glycerol production, the density ~adi -

76
E
Eo
+i t5
| | i j . . . . j I , * •
70 1 2 3 4 5 6 1.17 1.19 12|
Time th) Density (g/ml) Fi 8. 2. A. Distribution profile of D, hun~eniJ wild-type (A) and strain 26-6 (o 0) after eel~lrifugation in a Percoll-soebitol gradient at v~rious time points afl~ ineubalion in medium containing: ( - e) 8~ NaCI or (o) 8~ NaC| plus 1 mM glycerol. The position of the main cell band was determined visuat]y and confirmed by ¢¢11 counting. B. Density of sele¢ted
0.5 ml tractions.
ent centr i fugat ion was txied f o r mutant enrich- ment . It was ant ic ipated that mutan ts showing excessive glycerol product ion would accumulate glycerol faster than wild-type cells when subjected
to osmotic stress, and hence band in the top fractions of the gradient at an earlier stage after the osmotic shock than do wild-type cells, Accord- ingly, an exponential ly growing culture of mutage- nized cells was trans~errexl to 8% IqaCI medium and incubated t'or 30 rain, whereafter cell samples were c o n c e n t r a t e d and cen t r i fuged in the sorb i td -Perco l l gradient. Mos t of the cells banded
as expected in the bo t tom of the tube; but to search for mutants , undiluted por t ions of 0.1 ml from the pooled top fractions (3 ml) were spread on ag~tr plates, yielding about 10-30 colonies per
plate.
4.4. Identification of mutants with increased glycerol production
To identify colonies with an increased glycerol p roduc t ion a total, Of 400 colonies mere restreaked (10 per plate) on basal med ium on which the
glycerol auxotrophi¢ E, coil strain BB20-14 had been previously plated. Yeast strains with exces-
Fig. 3. Growth of the 81ycmrol requiring indicator strain E+ co// BB9,0+14 around colonies of I). ha,senit wild.type (righ~) and a mutant (left) isolated from the top fractions of the Percoll, sorbitol gradient as described in the text The two yeasl strains were streaked on agar plates coetainin 8 glycerol-tree basal
medium on 'which the E'. roll strain had been plated.
sire glycerol production were expected to leak glycerol to the medium thereby supporting visible growth of the indicator strain. The restreaking procedure was found necessary since direct plating of yeast cells on a lawn of E. coil BB20-14 re- suited in confluent bacterial growth between the yeast colonies, probably due to rapid diffusion of rdeased glycerol. About 10% of the restreaked yeast colonies were surrounded by zones of E. coli growth of varying width (Fig, 3). To verify in- creased glycerol production, five yeast colonies that supported bacterial growth were analysed for glycerol production and intracellular glycerol con- tent. Two of Ihese mutants showed clearly elevated glycerol levels when grown in basal medium (Ta- ble 1), When re-subjected to salt stress and subse- quent density gradient centrifugation the mutants
Table 1
Total glycerol production of the DL hUnSenii wild+l~t:J¢ and the toulon! strains 26+02 and 26-G10 grown in basal medmm
Strain Total 81ycerol lumol/m$ dry wt yeast
Wild-type 0.10 0,02 26-02 ~.35 0,04, 26-GI,0 0.29 0.06
V=~i~s rcpresenl mean SD ot four samp|¢s laken at a cell density of 0,3-0.6 mg dry wt/ml.

77
demonstrated a maintained ability ror what per- mitted their enrichment: a faster decrease in den- sity than the wildqype (not shown).
DISCUSSION
Mutants of Succharomyces cereoixJae that are blocked in cell surface growth or secretion b~.~come dense [13,14] and this property has been used to select mutants by density gradient centrifusation 114]. We have used the density gradient technique to enrich mutants of the yeast DebuG'oral ce.s" hansenii that have an altered glycerol metabolism.
Due to a high water permeability of the plasma membrane, the size and density of cells are af- fected by changes in medium osmolarity. On transfer to low water potential the cells shrink and become smaller and denser. For walled cells, ilke yeasts and Gram-negative bacteria, the elasticity of the cell wall is considerab!e. On, osmotically induced water loss, the cell wall shrinks together with the protoplast over large volume changes [15-17], in order to regain celt volume and /o r turgot in response to an osmotic stress, cells possess adaptive mechanisms which regulate the number of osmotically active molecules within the cells. By accumulating an osmoregulatory solute water re-enters the cell, the cell s~vells and be- comes less dense, In O. hansenii glycerol serves as the principal solute used for osmoregulation [1,~,1. In the present work, a Percotl gradient adjusted to high osmolarity was shown to separate osmotically adapted yeast ceils from cells having a higher density due to a low intraceilular glycerol content (Fig. 1 and 2). Based on these findings, a method was devised to enrich mutants with enhanced glycerol production (Fig. 3). Slight modification of this density separation procedure would make feasible the enrichment of other types of mutants. such as glycerol transport mutants and mutants with reduced ahillty to produce and accumulate glycerol. Since glycerol accumulaF, on appears to be a common response for yeasts exposed to osmotic stress [18 -20], the method should be gen-
erally applicable for enrichment of yeast mutants with defective glycerol metabolism.
ACKNOWLEDGEM ENTS
We arc most grateful to Hhkan Perotft for kindly helping us with density determinations and to Anders Btomberg for helpful discussions. This work was supported by grants from Mag:,n. Bergva|ls Stiftets och As-son Johnsons stiftelse,
REFERENCES
I l l Adler, L. and r,.3uslafsso., L. ~19BO) Arcn. M~crohiol. 124, 123- ] 30.
[2] Adler. L.. Blombcrg. A, and Nilsson, A. (1985) J. Bacleriol. IM. 300-306.
[3] Reed. RH. Chudek. J.A., Fosler, R. and (;add. GM. (19871 Appl. Envinm. MicrobJol, .~3. 2119 2123.
141 AndrE. L. Nilsson, A. and Adlar, L. (19881 J. Gen. Mbzro- biol. 134. 669 677.
[51 Nils,~)n, A., Thomson. K-g, and Adler. h. (1989) Biochirn. nlopby~. Acla 9gl. 296-302.
16l Cronan. J,E. Jr. and Bell, RM. (1974) J. Bacteriol. 118, 59~ 605.
[7] Davies, K,W,, Botslein, D. and Ro,,h. J.R, (lqF, O) A Man- u~) /or OCrlel~c Engin¢cr~.ng. Advar~=ed Bac~:erial Genetics. CSH. New York.
[81 Norkran!,, P,. (1065) Arch. Microbiol, 54. 374-396. 19] Fink, G. (19701 The biochemical genetics of yeast, pp.
59-78. in (Tabor, H. attd Tabor, C.W., eds.~, Mettzods in En~ymology. V,,l XV~IA.
[101 Osler. G. and Yilmumolo. ~,t. {1963) Ch~m. Roy. 63. 257-268.
[111 Tengvar. Ch., R)rxsen, M., Hultslr~m. D., OIsson. Y. Pertofl, H. and Pe;terss0n. A. 0982) Acta Neuropalhol. 57. 143-130.
ll2l Norkrans. B, and Kylin, A. (19691 J, BaCW.TioL 100. 836 845,
[13] HenD', S.A,, Atkm~on, K.D.. KolaL A.I. and Culbertson. M.R. (10771 J. Bacterlol. 130+ 472-484.
[]4] Novick, P,. Fidd, C and Sehekman, R. (19801 Celt 21, 20:5-215.
[15} Kogh. A,L, 0984) J. Bactenol. 'i59, 919-924. ll6] Baldy, in. W., Shcu, M.J.-T.. Bankston. P.W. and
WoLdringh, C.L. (1988) I. ~acterioL 170. 452-455. [171 Morris. G.J., Wmter~, L, C0ulson G.E, and Clarke. K.J,
(1986} J. Gen MicrobioL 132. 2023~2034. [18] Brown. A.D (19781 Adv. Microbial, Physiol. 19. t81~242. 119] Bellin~.er, Y. and Lahrcr. F. 119881 Can. J. Microbiol, 34,
605-612.
[201 Reed, R.H., Chudek, J.A.. Fo~ler. R. and Gadd, G.M. (1987) Appt. Environ. Mlcrobiol. 53, 2119-2123.





























