Embed Size (px)
Citation preview

INTRODUCTION
The nucleolus is well known to be the site of rDNAtranscription, rRNA processing and ribosome biogenesis. Agroup of nonribosomal proteins, including nucleolin,p120/Nop2p, fibrillarin and NO38/B23, are thought toparticipate in nucleolar biogenesis in higher eukaryotes in partby serving as recruiting stations for other proteins (Tuteja andTuteja, 1998; Zatsepina et al., 1999; Dundr and Olson, 1998;Gustafson et al., 1998). Consistent with such a role, theseproteins possess several conserved domains, including polyD/E stretches and RGG repeats, that bind to nucleic acid or toother nucleolar proteins. In NO38/B23, the poly D/E isessential for binding to histone H2A and H2B (Kleinschmidtet al., 1985). In nucleolin, it may be involved in the interactionwith the protein kinase CK2α (Li et al., 1996). RGG repeatsare present in fibrillarin, p120/Nop2p and nucleolin. Innucleolin, the RGG domain mediates the interaction withseveral ribosomal proteins (Bouvet et al., 1998) and alsoparticipates in binding to nucleic acids (Heine et al., 1993;Ginisty et al., 1999). The RGG region of p120/Nop2p interactswith rRNA (Gustafson et al., 1998). Interestingly, proteins withthese conserved domains are multifunctional. Nucleolin is agood example: it is involved in numerous steps of ribosomebiogenenesis, from rRNA synthesis and processing, throughthe formation of pre-ribonucleoparticles, to nucleolar-cytoplasmic trafficking, and has additional functions unrelatedto ribosome biogenesis (Tuteja and Tuteja, 1998; Ginisty et al.,1999).
Similar domains are found in NOPP44/46, the only T. brucei
nucleolar protein characterized to date (Das et al., 1996). Itconsists of four regions: unique (U; aa 1-96), junction (J; aa97-168), acidic (A; aa 169-217) and repeat (R; aa 218-312).The R region, with its characteristic RGG repeats, is sufficientfor interaction with nucleic acids (Das et al., 1998). Themoderately acidic J region and poly D/E containing A regionare involved in the interaction with a protein kinase(s). The Aregion also interacts with the RNA binding protein p34/37(Zhang et al., 1998) to form a trimolecular complex with 5SRNA (Pitula et al., submitted). The U region is sufficient fornuclear and nucleolar localization, despite the lack of a typicalnuclear localization signal (NLS) (Das et al., 1998). It displaysweak yet significant sequence similarity with a number ofnucleolar proteins, such as nucleolar histone deacetylases andsome immunophilins (Das et al., 1998; Lusser et al., 1997),which also lack NLSs. Despite the presence of conserveddomains, NOPP44/46 does not show overall homology to anyspecific protein. The presence of multiple interaction domains,coupled with the high abundance of NOPP44/46, led us topropose that it may function as a scaffolding protein in thenucleolus, thereby facilitating the interaction of other nucleolarmolecules.
To further explore the role of NOPP44/46, we used a two-hybrid screen to identify its molecular partners. As reportedhere, among the molecules identified was a predicted G-protein, TbNOG1. TbNOG1 contains the typical motifsinvolved in GTP-binding, plus a large C-terminal extension.Closely related sequences are present in other eukaryotes andArchea. NOG sequences also have significant homology withpreviously identified Obg and DRG G-proteins, and together
173
We describe here a novel, evolutionarily conserved set ofpredicted G-proteins. The founding member of this family,TbNOG1, was identified in a two-hybrid screen as a proteinthat interacts with NOPP44/46, a nucleolar phosphoproteinof Trypanosoma brucei. The biological relevance ofthe interaction was verified by co-localization andco-immunoprecipitation. TbNOG1 localized to thetrypanosome nucleolus and interacted with domains ofNOPP44/46 that are found in several other nucleolarproteins. Genes encoding proteins highly related toTbNOG1 are present in yeast and metazoa, and related Gdomains are found in bacteria. We show that NOG1proteins in humans and Saccharomyces cerevisaeare alsonucleolar. The S. cerevisae NOG1gene is essential for cell
viability, and mutations in the predicted G motifs abrogatefunction. Together these data suggest that NOG1 may playan important role in nucleolar functions. The GTP-bindingregion of TbNOG1 is similar to those of Obg and DRGproteins, which, together with NOG, form a newlyrecognized family of G-proteins, herein named ODN. TheODN family differs significantly from other G-proteinfamilies, and shows several diagnostic sequencecharacteristics. All organisms appear to possess an ODNgene, pointing to the biological significance of this family ofG-proteins.
Key words: Human, Nucleolus, G-protein, Trypanosoma brucei,Saccharomyces cerevisiae
SUMMARY
A novel nucleolar G-protein conserved in eukaryotesJeong-Hyun Park 1,2,*, Bryan C. Jensen 1,2, Charles T. Kifer 1 and Marilyn Parsons 1,2,‡
1Seattle Biomedical Research Institute, 4 Nickerson St, Seattle, WA 98109, USA2Department of Pathobiology, School of Community Medicine and Public Health, University of Washington, Seattle, WA 98195, USA*Present address: Department of Molecular Biology and Genetics, Johns Hopkins University, Baltimore, MD 21205, USA‡Author for correspondence (e-mail: [email protected])
Accepted 19 October; published on WWW 11 December 2000Journal of Cell Science 114, 173-185 © The Company of Biologists Ltd
RESEARCH ARTICLE

174
they form a new family of G-proteins. Like TbNOG1, theSaccharomyces cerevisaeand human NOG1 proteins arenucleolar and the S. cerevisiaehomologue is essential. Thestriking conservation of amino acid sequence between distantlyrelated species and essential nature of S. cerevisae NOG1suggests an important function for NOG1 in an as yet unknownregulatory pathway in the nucleolus.
MATERIALS AND METHODS
Organisms, media and general proceduresEscherichia coliXL1-Blue was used as a plasmid host. S. cerevisiaestrains were grown in YEPD (1% yeast extract, 2% BactoPeptone and2% glucose) or synthetic complete medium (C medium) (Sherman etal., 1981) lacking appropriate supplements to select for plasmids. Foractivation of GAL1 regulated constructs, 2% galactose and 2%raffinose were used in lieu of glucose. Yeast were transformed by thelithium acetate method (Gietz et al., 1992). KAC plates (1% potassiumacetate, 0.1% yeast extract, 0.1 mg/ml adenine, and 0.1 mg/ml uracil)were used for sporulation. The strains employed include the C306diploid (MATα/MATa, trp1/trp1, ura3-52/ura3-52, leu2-3, 112/leu2-3, 112, his3D/his3D, can1/+, cyh2/+, lys5/+, ade6/+), the haploidstrain C296-21-3h1 (B. Jensen, unpublished strain), EGY191 (MATα,his3, trp1, ura3, lexA2ops-LEU2) and EGY48, which differs only bythe presence of six rather than two operators (Estojak et al., 1995).Procyclic form T. bruceiTREU667 and the transgenic 29-13 procyclicform line (Wirtz et al., 1999) were cultured in SDM-79. The lattercells co-express T7 RNA polymerase and the tetracycline repressor(TetR). COS cells were cultured in Dulbecco’s modified Eaglemedium.
All primers used, along with the relevant restriction sites, are shownin Table 1. Unless otherwise noted, for polymerase chain reaction(PCR) cloning the products were digested with the listed enzymes andligated to similarly cleaved vectors. All constructs were checked by acombination of restriction enzyme digestion and DNA sequenceanalysis.
Two-hybrid system for NOPP44/46-interacting proteins The U (aa1-96) and UJ (aa1-168) and UJA (aa1-217) regions ofNOPP44/46 were PCR amplified from T. bruceigenomic DNA using5′ primer NOPP-2S and 3′ primers NOPP-96A, NOPP-168A orNOPP-217A. The NOPP44/46 fragments cloned into pEG202. Theresulting bait plasmids encode the LexA DNA binding domain (aa1-202) fused to U, UJ, or UJA.
The yeast strain EGY191 harboring the β-galactosidase reporterplasmid p1840 (GAL1-LexA1op-lacZ and URA3gene) and the LexA-UJ bait plasmid, were transformed with a T. bruceigenomic library.This library, gift of Drs U. Goeringer and A. Souza, was generatedfrom a Sau3A partial digest of T. bruceigenomic DNA (IsTaR 1) thatwas cloned into the BglII site of pJG4-5. The libarary sequences arefused to a hemagglutinin (HA)-tagged B42 activation domain andexpression is controlled by the GAL1 promoter. Colonies expressingproteins that interacted with LexA-UJ were identified by galactose-dependent growth in medium lacking Leu and confirmed by β-galactosidase activity, and prey plasmids were rescued.
For mapping interactions, coding regions of TbNOG1 and itsfragments TbNOG1(1, 425) and TbNOG1(1, 334) were amplifiedfrom genomic DNA using 5′ primer TNOG-1S and 3′ primers TNOG-655A, TNOG-425A or TNOG-334A. XhoI-digested PCR productswere ligated with XhoI-digested pJG4-5. The PCR products were alsofor cloning into pTbmyc2 (see below). The G1 mutant GKS>AAA(aa 181-183) was constructed by site-directed mutagenesis of the fulllength TbNOG1bait plasmid using the Stratagene Quick Change site-directed mutagenesis kit and primers TNOG1-G1S and TNOG1-G1AS. Interactions were tested in strain EGY48.
DNA library screening and sequencingA portion of the original clone 4 was used as a probe to screenprocyclic and bloodstream λZap II cDNA libraries. Positive plaqueswere purified and the plasmids containing cDNAs were recoveredaccording to the manufacturer’s protocol (Stratagene). Sequences ofboth strands of two cDNAs were determined following the creationof nested deletions (Henikoff, 1987).
Expression of epitope-tagged TbNOG1 in T. bruceiThe PCR amplified TbNOG1 fragments generated above wererestricted with XhoI and XbaI and ligated with similarly digestedpTbmyc2 (Das et al., 1998), which places a double myc tag at the Cterminus. Coding regions of TbNOG1-myc and TbNOG1(1, 334)-myc were re-amplified from these constructs using primers TNOG-1S-H and Myc-A and cloned into pLew79 (Wirtz et al., 1999),replacing the luciferase gene. The resulting plasmids express taggedTbNOG1sequences from a PARPpromoter that is regulated by theTet operator. The phleomycin resistance gene is expressed separatelyfrom a T7 promoter on the same plasmid. pLew79 derived plasmidswere linearized with NotI within the rDNA intergenic sequence andtransfected into the T. brucei29-13 cell line as described (Andersonet al., 1998) for selection with phleomycin (10 µg/ml). For theinduction, Tet was added to 10 µg/ml.
Immunoprecipitations and immunofluorescence analysisof epitope-tagged proteins in T. bruceiThe preparation of cell lysates, immunoblotting,
JOURNAL OF CELL SCIENCE 114 (1)
Table 1. PCR primersRestriction
Primer name* site‡ Sequence (5′…3′)§NOPP-2S BamHI tatgcggatcctggagggtttctacggtNOPP-96A XhoI atgctcgagttaaacagttgaacgtatccagNOPP-168A XhoI atgctcgagttacgctcaattccttcattgtNOPP-217A XhoI attatctcgagttactcagtgcgttgcactttacTNOG1-1S XhoI cgtttctctcgagatgtccacaatctacaatTNOG1-655A XhoI/ XbaI tacttactcgagttatctagagcgtctgtcgcgtcTNOG1-425A XhoI/ XbaI cccatctcgagttatctagatcccatgtcttccgc TNOG1-334A XhoI XbaI cagcgtctcgagttatctagacttgaggtcgcctaTNOG1-1S-H HindIII cgtttctaagcttatgtccacaatctacaatTNOG1-G1S gggttccccaacgtcgctgcggcaagtttcatgaacaagTNOG1-G1A cttgttcatgaaacttgccgcagcgacgttggggaacccMyc-A BamHI atcttaaactcagtagaactaggatccttacagcaggtctSNOG1-∆5 atgcaactttctggaaggatatcccactgtcgctccagc-
gccaagctcgaaattaac SNOG1-∆3 tcaacggaaatctgtcttaccgacaccacgcttaccact-
gtgaattgtaatacgactcac SNOG1-5′S cacctatgtactacctcatc SNOG1-3′A tgacgtgtacgtacatcctc SNOG1-1SE EcoRI cggaattcccgaagatgcaactttcatggaagSNOG1-647A XhoI cccgctcgagtcaacggaaatctgtcttaccgacSNOG1-1SX XhoI ctccctcctatactcgagatgcaactttcatggaagSNOG1-3′AK KpnI cggggtacctgtatacatccagtgacatac SNOG1-3′XS XhoI cccttcttctcgaggcgtggtgtcggtaagacagSNOG1-G1S ctgtggttatccaaatgtcgctgcggcctcgtttttgagat-
gcattaccaaatcagSNOG1-G1A ctgatttggtaatgcatctcaaaaacgaggccgcagcga-
catttggataaccacagSNOG1-G3S gtctgtcatggttgttatcaacgcaaccctcattatcagac-
cggaggatttagacSNOG1-G3A gtctaaatcctccggtctgataatgagggttgcgttgata-
acaaccatgacagacHNOG1-1S NotI catacggcggccgccatggcacattacaacttcHNOG1-633A ClaI accaaaatcgatctactatctcctgtcctt
*Sense primers and antisense primers are indicated by ‘S’, ‘A’,respectively. Numbers indicate the codon. Bold letters indicate nucleotideschanged in mutagenesis.
‡Restriction sites in primer used in subsequent cloning.§Restriction sites are underlined and start codons italicized.

175Nucleolar G-protein
immunoprecipitation, and immunofluorescence analysis wereperformed as described (Das et al., 1996). Primary antibodies weremouse anti-myc 9E10 (IgG1) ascites fluid and purified mouse anti-NOPP44/46 antibody ID2 (IgG2a) (Parsons et al., 1994). They wereused for immunoprecipitations at 0.75 µl/mg and 0.75 µg/mg cellularprotein, respectively, and complexes were collected with Protein G-or Protein A-Sepharose. For western blot analysis, anti-myc wasused at 1:1000 dilution and anti-NOPP44/46 was used at 1 µg/ml.Secondary antibodies were 125I-labeled goat anti-mouse IgG orProtein A. For immunofluorescence microscopy, primary antibodieswere used at 10-fold higher concentrations. They were revealed withfluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG1and Texas red (TXRD)-conjugated goat anti-mouse IgG2a at 10µg/ml. All fluorescently-tagged secondary antibodies were obtainedfrom Southern Biotechnology, Inc. Cells were counterstained with4′,6-diamidino-2-phenylindole (DAPI).
Expression of recombinant TbNOG1 and production ofantiseraThe TbNOG1gene was excised from the plasmid p413-GAL1-HA-TbNOG1(J. Park, unpublished plasmid) using SpeI and XhoI. Theresulting fragment, which encodes HA-tagged TbNOG1, was ligatedinto the NheI and XhoI sites of the pRSetA expression vector.Following transformation into BL21(DE3)pLysS cells, HA-TbNOG1synthesis was induced for two hours at 37°C with 1 mMisopropylthio-β-D-galactoside. Cell pellets were resuspended in 1×PBS, 10 mM imidazole, pH 7.4, and 100 µg/ml lysozyme, and lysedby freeze/thaw followed by sonication. HA-TbNOG1 was in theinclusion body fraction. This fraction was separated by SDS-PAGEand the appropriate band excised. The isolated material was used toimmunize rabbits (six injections of 0.5 mg, the first in completeFreund’s adjuvant and the rest in incomplete Freund’s adjuvant). Theantiserum was used at a 1/2000 dilution in immunoblot analyses,using approximately 3×106 cells per lane. As a control, the same blotswere probed with antibody to the 56 kDa isozyme ofphosphoglycerate kinase, at a 1/2000 dilution (Parker et al., 1995).
Expression of epitope-tagged human NOG1 in COS cellsHsNOG1 (accession 4191616) was PCR amplified from HeLa S3cDNA library (Clontech) using the primers HNOG1-1S and HNOG1-633A and cloned into pFLAG-CMV-2 (Kodak). The resultant plasmidencodes the full-length HsNOG1 with the FLAG tag at the N terminus.
For transfection, cells were grown as monolayers on glass coverslips.After fixation in methanol/acetic acid (1:1, v/v) for 10 minutes, cellswere permeabilized with 0.2% Triton X-100 in PBS. The nucleolus wasstained with mouse anti-NO38/B23 (1:500 dilution, gift of P. K. Chan)and revealed with TXRD-conjugated rabbit anti-mouse IgG (10 µg/ml).Tagged HsNOG1 was revealed by FITC-anti-FLAG M2 at 50 µg/ml.Cells were counterstained with DAPI.
Expression of epitope-tagged ScNOG1 in yeastThe ScNOG1open reading frame (ORF) plus 293 bp downstream ofthe stop codon was amplified usingTurbo Pfupolymerase and primersSNOG1-1SX and SNOG1-3′K. The restricted PCR product wasligated into p414-GAL1-HA-TbNOG1 (B. Jensen, unpublishedplasmid) replacing the TbNOG1gene. The resulting plasmid, pGAL-HA-ScNOG1, a double HA tagged-ScNOG1 under the control of theGAL1 promoter. The ligation reaction was directly transformed intoC306nog1∆ (see below), since it proved difficult to isolate theScNOG1gene in E. coli.
For immunofluorescence, the transformed cells were grownovernight to mid-log phase in C medium lacking Trp supplementedwith raffinose. The cells were washed with succinate buffer (95 mM,pH 6) containing 0.16% yeast nitrogen base without amino acids orammonium sulphate, and resupended in C medium lacking Trp andcontaining galactose and raffinose to induce HA-ScNOG1. After 2-2.5 hours the cells were fixed with 3.7% formaldehyde in PBS for 1
hour, and processed for immunofluorescence as described (Winey etal., 1991). The HA epitope was detected using rat anti-HA 3F10(Boehringer Mannheim) at 0.5 µg/ml and detected with TXRD-conjugated goat anti-rat IgG at 5 µg/ml. Nop1p was detected using amouse anti-Nop1p (Aris and Blobel, 1988), kindly supplied by Dr J.Aris, at 40 ng/ml and detected with FITC-goat anti-mouse IgG1 at 0.5µg/ml.
Generation and complementation of a NOG1 deletion in S.cerevisaeThe deletion disruption of ScNOG1was obtained by transformationof a PCR-synthesized marker cassette with ScNOG1 flankinghomology regions into the diploid strain C306. The KanMX marker,which specifies resistance to G418, was amplified from pBS-KanMX(B. Jensen, unpublished plasmid) with the primers SNOG1-∆5 andSNOG-∆3. The PCR product was used to directly transform the yeaststrain C306 with selection on YEPD containing 0.2 mg/ml G418.Proper integration was verified by PCR using primers SNOG-5′ andSNOG-3′. The resulting strain was referred as to C306NOG1/nog1∆.
To rescue gene disruptions, the ScNOG1ORF was amplified usingprimers SNOG1-1SE, and SNOG1-647A. The digested PCR fragmentwas ligated into p414-GAL1 (Mumberg et al., 1994) and directlytransformed into C306 NOG1/nog1∆. The resultant diploid, C306NOG1/nog1∆/pGAL1-ScNOG1, was sporulated and dissected ontomedium containing galactose or glucose to induce or repressexpression of the plasmid-encoded ScNOG1. All viable segregantswere tested for their genotype at the chromosomal NOG1 locus byplating on G418.
Generation and functional analysis of mutations in theGTP-binding domain of ScNOG1The transcriptional terminator of ScNOG1 was amplified from strainC296-21-3h1 with the primers SNOG-3′SX and SNOG-3′AK andcloned into p414-GAL1 to yield p414-GAL1-NOG1term. XbaI andXhoI were used to cleave the plasmid between the promoter andterminator. DNA fragments containing mutations in either the G1 or G3motifs of ScNOG1 were generated using the Turbo-Pfu (Stratagene) institching PCR. For the GKS to AAA mutation at residues 179-181 (G1motif), the M13 reverse primer and the mutagenic SNOG1-G1S primerwere used to generate a fragment spanning the GAL1 promoter regionthrough the mutated residues using pGAL-HA-ScNOG1 as template.Similarly, primers SNOG1-G1A and SNOG-3′AK were used togenerate a fragment spanning the mutated residues through theterminator region. PCR products were mixed together to serve astemplate for a final round of PCR using the M13 reverse primer andSNOG-3′AK primers. The mutation KTD to ATL at residues 279-281was similarly performed using the mutagenic primers SNOG1-G3S andSNOG-G3A. Following cotransformation into yeast with the digestedp414-GAL1-NOG1term DNA, the plasmid-encoded NOG1 mutantgenes were amplified and sequenced.
The C306 NOG1/nog1∆ diploids harboring the mutant or wild-typeHA-ScNOG1 plasmids or vector, p414-GAL1-NOG1term, weresporulated. The progeny were plated on galactose plus raffinose Cmedium lacking Arg and Trp and containing canavanine (60 µg/ml,Sigma) and cycloheximide (10 µg/ml, Sigma) to select for haploidsegregants. To test for the presence of the nog1::KAN allele,individual segregants were replica plated to galactose plus raffinosemedium lacking Trp and to the same medium containing 400 µg/mlG418.
RESULTS
Two-hybrid screening with the UJ region ofNOPP44/46To detect potential molecular partners of NOPP44/46, we

176
conducted an interaction screen using the yeast two-hybridsystem. We screened a genomic library since T. bruceigenesare densely packed and rarely contain introns. To minimize falsepositives, the bait plasmid contained the U to J region (aa1-168)of NOPP44/46 (Das et al., 1998), but lacked the highly chargedA and R domains. Additionally, we used a stringent system,where only two operators were present next to the selectablemarker (LEU2) (Estojak et al., 1995). From 2 millionindependent transformants (equivalent of approximately 40haploid genomes), we identified 34 clones that activated thereporters in combination with LexA-UJ but not with LexAalone or LexA-lamin. The plasmids mapped into fivemajor groups. Some features of each predicted protein ascharacterized to date are summarized in Table 2. Analysis ofthe predicted protein sequence of partial DNA sequences ofclones 1 and 2 revealed no obvious similarities with sequencesin the databases. Clone 3 displayed a high degree of homologywith a member of the variant surface glycoprotein family (majorplasma membrane proteins in T. brucei). It was thereforedeemed a non-physiological positive. Clone 5 was related to apredicted protein of unknown function in Arabadopsis thaliana.Clone 4 displayed a high degree of sequence similaritywith hypothetical proteins in Saccharomyces cerevisiae,Caenorhabditis elegans, and humans.
cDNA, structure and interaction map of clone 4 To obtain a complete clone 4 cDNA, we screened bothbloodstream and procyclic cDNA libraries and chose thelongest clone from each library for sequencing. The combinedsequence comprised 3934 nt plus a poly(A) tail (Fig. 1a). Thepresence of the spliced leader (the sequence trans-spliced toform the 5′ end of all nuclearly-encoded mRNAs in T. brucei)(Parsons et al., 1984), together with the poly(A) tail, indicatesthat the sequence represents the full-length mRNA.Interestingly, structural heterogeneity was found at the 3′ end:relative to the stop codon, the poly(A) tail was attached at nt64 in the procyclic clone and at nt 1711 in the bloodstreamstage clone. Whether this represents a stage-regulateddifference is unclear.
The ORF encodes a polypeptide of 655 aa residues with acalculated molecular mass of 74.8 kDa (Fig. 1b). The predictedprotein has a pI of 9.8, with 16.3% basic and 12.5% acidicresidues. PSORT (Nakai and Horton, 1999), identifies abipartite NLS bipartite at residues 131-144 and two predictedNLSs at residues 395-401 and 511-518 (Fig. 1b).
Codons 176-293 of the ORF contain the three consensus
motifs, designated here as G1, G2 and G3, that define a G-protein (Fig. 1c). The G1 motif, GXXXXGK(S/T), is found inmost proteins that bind purine nucleoside triphosphates,including G-proteins, protein kinases and ATPases (Walker etal., 1982). The G2 motif, DXXG, is situated close to the G1motif in the tertiary structure and is thought to be involved in theconformational change between the GTP- and GDP-boundforms (Bourne et al., 1991; Seeburg et al., 1984). The G3 motif,NKXD, determines the specificity for guanine. Based on thepresence of well-characterized GTP-binding motifs in therequired order, we predict that the clone 4 gene encodes a G-protein. Since the N-terminal region (aa1-334) contains theGTP-binding motifs, it is referred as to the G-domain. As shownbelow, the protein is localized to the nucleolus and the gene istherefore named TbNOG1(T. bruceinucleolar G-protein 1).
To map the interaction of TbNOG1 and NOPP44/46, wetested of various regions of TbNOG1 and the UJA, UJ, and Udomains of NOPP44/46 in the two-hybrid system (Fig. 2). Theoriginal two-hybrid clone encodes TbNOG1(426, 655), whichinteracted with the UJA and UJ regions of NOPP44/46 butnot with the U region. Interestingly, TbNOG1(1, 425) andTbNOG1(1, 334), which do not overlap with TbNOG1(426,655), interacted with all of the NOPP44/46 constructs,including the U region. The same results were obtained withthe full-length TbNOG1. Together, these data indicate that theN-terminal 334 aa, which contains the G domain, also containsa binding site for the U region of NOPP44/46, while the C-terminal 230 aa contains a binding site for the J region. Theseexperiments do not rule out additional sites of interaction.
We also tested the ability of the NOPP44/46 fragments tointeract with TbNOG1 that contained mutations in the G1motif. Side chains of residues within this motif are involvedbinding the β-phosphate of GTP and coordination with themagnesium ion (Kjeldgaard et al., 1996). In this experiment,the conserved G1 residues GKS were mutated to AAA,residues that would be incapable of mediating theaforementioned interactions. The mutant protein was stillcapable of interacting with U, UJ, and UJA fragments.
JOURNAL OF CELL SCIENCE 114 (1)
Table 2. Clones isolated from the yeast two-hybrid screenCoding
Clone Interaction* Homology Motifs‡ region
1 UJ None NLS ND2 U and UJ None None ~430 aa3 UJ VSG ND ND4 UJ and U S. cerevisiae, C. elegans,NLSs, GTP- 655 aa
human ORFs, binding motifsbacterial proteins
5 U and UJ A. thalianaORF NLS ~750 aa
*Assessed as described in the legend of Fig. 2. ‡Identified by PSORT (Nakai and Horton, 1999) and Scanprosite
(http://www.expasy.ch/prosite). ND, not determined.
poly A
500 bp
G1: GFPNVGKS
E: YAFTT
G2: DTPG G3: NKSD
100aa
(a)
(b)
Fig. 1.The clone 4 (TbNOG1) cDNA and predicted protein.(a) cDNA. The spliced leader is shown as an open circle and the openreading frame as a cylinder. The first polyadenylation site is markedby an arrow. (b) Structure of the predicted protein. Location ofpredicted NLSs are shown as dotted boxes. The black regiondelineates the area containing typical GTP-binding motifs. Theenlargement shows the location (hatched) and sequence of individualmotifs within this region.

177Nucleolar G-protein
Expression of myc-tagged TbNOG1 protein in T.bruceiWe were unable to isolate stable T. brucei transfectantsconstitutively expressing myc-tagged full-length TbNOG1, orits truncated versions (aa 1-425, aa 1-334 and aa 255-655). Wetherefore turned to the Tet-inducible system for the expressionof myc-tagged TbNOG1 (Wirtz et al., 1999). Sequencesencoding myc-tagged TbNOG1 were cloned into pLEW79,where expression is driven by the PARP promoter andcontrolled by an adjacent Tet operator (Wirtz et al., 1999).Following electroporation into T. bruceiprocyclic forms, stabletransfectants were isolated in the absence of Tet. In induced,but not uninduced, cells western blot analysis with anti-myc detected a single protein with the molecular mass ofapproximately 83 kDa (Fig. 3a). This size approximates thatexpected for the 74.8 kDa protein plus 3.4 kDa tag. Inductionof TbNOG1-myc did not alter the expression of NOPP44/46(Fig. 3a).
TbNOG1-myc is nucleolar and interacts withNOPP44/46 in T. brucei To investigate the subcellular localization of TbNOG1, thetransfectants were processed for immunofluorescence. Fig. 3bshows the fluorescence patterns of cells grown in the absenceor presence of Tet and then stained with anti-myc to disclosethe location of TbNOG1-myc. Staining with DAPI and anti-NOPP44/46 revealed the nucleus and nucleolus respectively. Inuninduced cells, no signal was observed with anti-myc. Uponaddition of Tet to induce TbNOG1-myc, anti-myc detected aregion within the nucleus that is indistinguishable from theanti-NOPP44/46 stained nucleolus (see Fig. 3b, overlay). Thus,TbNOG1 is a nucleolar protein in T. brucei.
To provide biochemical evidence that TbNOG1 interactswith NOPP44/46 in T. brucei, we carried out co-immunoprecipitation experiments (Fig. 3c). In cells expressingTbNOG1-myc, the tagged protein was immunoprecipitatedwith anti-myc. As detected by western blot analysis with anti-NOPP44/46, the precipitates also contained NOPP44/46. As
expected, anti-myc immunoprecipitates from uninduced cellsdid not contain NOPP44/46, demonstrating that co-precipitation is mediated by TbNOG1. Conversely, anti-NOPP44/46 immunoprecipitates from induced cells containedTbNOG1-myc as detected with anti-myc antibodies. Thecolocalization and coprecipitation studies confirm that theinteraction observed in yeast reflects a biologically relevantinteraction in the parasite.
The two-hybrid interaction mapping (Fig. 2) demonstratedthat the G-domain of TbNOG1 interacts with the U region ofNOPP44/46. We also expressed a double myc-tagged versionof this fragment in T. bruceiusing the Tet-inducible system.Immunofluorescence analysis demonstrated that TbNOG1(1,334)-myc is nucleolar (Fig. 4a). As shown in Fig. 4b,NOPP44/46 was coprecipitated with TbNOG1(1, 334)-myc ininduced cells. In the reciprocal experiment, the taggedG domain coimmunoprecipitated with NOPP44/46.Phosphorimaging analysis showed that about three-fold moreNOPP44/46 was associated with full-length NOG1-myc thanwith the G-myc domain (normalized to the amount ofimmunoprecipitated myc-tagged protein).
TbNOG1 is expressed in bloodstream and procyclicstage T. brucei To assess the expression of TbNOG1 in the parasite life cycle,HA-tagged TbNOG1 was expressed in E. coli, purified, andused to raise antisera. The antiserum detected a protein ofapparent molecular mass of 81 kDa in immunoblot analyses(Fig. 5). The protein was observed in the actively dividingprocyclic (insect midgut) and mammalian bloodstream slenderforms, as well as in the non-dividing stumpy bloodstreamforms. Normalization of of the signals to those seen with theconstitutively expressed 56 kDa phosphoglycerate kinase(PGK-A) indicates that the levels of NOG1 are not highlyregulated during parasite development. Immunofluorescenceanalysis using anti-TbNOG1 revealed a nucleolar distribution(data not shown).
TbNOG1 is a member of a novel family of G-proteinsA BLASTp search (Altschul et al., 1997) of the predictedprotein data banks showed that sequences highly related toTbNOG1 are found in diverse eukaryotes, including Homosapiensand S. cerevisiae(Fig. 6). The G domain (aa 1-334)displayed a high degree of sequence homology (62-71%identity), while the C-terminal half was much less conserved(25-31% identity). Interestingly, the G-domain also showedover 50% similarity with proteins in members of the Archaea.In the archeal proteins, the C-terminal extension is missing.All the proteins shown in Fig. 6 were found fortuitously bygenomic or cDNA cloning and thus are functionallyuncharacterized. Apart from the GTP-binding region,TbNOG1 displayed no obvious sequence similarities withthe well-characterized G-proteins of the Ras, EF-2 andheterotrimeric G protein families, indicating that it is a memberof a distinct family of G-proteins.
The effector (E) motif, also called Switch I, is not involvedin GTP-binding but is thought to participate in the interactionwith effector molecules. Unlike the G1-G3 motifs, which areshared among almost all G-protein families, this motif isfamily-specific. The E motif is located between the G1 and G2motifs, and contains a conserved threonine (Sprang et al.,
UJA UJ UTbNOG1NOPP44/46
Full length + + +
1, 425 + + +
1, 334 + + +
426, 655 + + -
pJG4-5 vector - - -
G1 mut+ + +*
Fig. 2.TbNOG1 contains multiple binding sites for NOPP44/46.TbNOG1, its deletions, and the G1 mutant were cloned into pJG4-5and tested for interaction with the UJA, UJ and U regions ofNOPP44/46 (expressed from the bait plasmid). The GTP-bindingregion (position 176-293) of TbNOG1 is marked by the black box.Plus symbols indicate that the transformed yeast EGY48 cells grewin galactose medium lacking Leu, revealing an interaction betweenbait and prey.

178
1997). In the sequences closely related to NOG1, a conservedthreonine containing motif is found in an appropriate position.This motif, YAFTT, occurs in TbNOG1 at position 199-203(Fig. 1b). To identify the constellation of proteins related to theNOG1 G-domain, we scanned the protein databases with thepattern (G(X)4GKS(X)15YXFTT) corresponding to the G1 andE motifs. The screen identified 42 sequences, all of which alsocontain G2 and G3 motifs, indicating they are all predicted G-
proteins. Analysis of the regions containing GTP-bindingmotifs (a subset is shown in Fig. 7a) identified several residuesthat are conserved within these proteins but not with other G-proteins. These include Pro residues at position 3 in the G1 andG2 motifs, and Leu or Ile at position 5 in the G2 motif.
Alignments of the regions spanning the G motifs were used
JOURNAL OF CELL SCIENCE 114 (1)
Fig. 4. Coimmunoprecipitation and colocalization of TbNOG1(1,334)-myc and NOPP44/46. Stable transfectants containing inducibleTbNOG1(1, 334)-myc were examined as in Fig. 3b,c.(a) Colocalization. (b) Coimmunoprecipitation.
Fig. 3. Expression, colocalization and coimmunoprecipitation ofTbNOG1 and NOPP44/46 in T. brucei. (a) Western analysis ofTbNOG1-myc and NOPP44/46 expression. Cells containing Tet-regulated TbNOG1-myc were incubated for 12 hours in the presence(+) or absence (−) of Tet. Total cell lysates were examined byimmunoblot analysis with anti-myc and anti-Nopp44/46. (b)Colocalization. Transfectants were stained with anti-myc (IgG1) andanti-NOPP44/46 (IgG2a), followed by FITC-conjugated anti-IgG1and TXRD-conjugated anti-IgG2a. The nucleus and mitochondrialDNA (small dot) were stained with DAPI. Green and red colors wereoverlaid. Bar, 5 µM. (c) Coimmunoprecipitation. Anti-myc and anti-NOPP44/46 (as marked above the blot) were used toimmunoprecipitate (IP) the relevant molecules from total cell lysates.The resultant immunoprecipitates were analyzed for the presence ofTbNOG1-myc or NOPP44/46 by immunoblot analysis with theantibodies indicated below each panel.

179Nucleolar G-protein
to calculate a phylogenetic tree. As shown in Fig. 7b, the 42proteins clustered into three groups, one of which containedall the NOG1 related proteins. Within this cluster, thephylogenetic tree was very similar to that seen when the treewas built with the entire sequences of NOG1 proteins (data notshown). The other two clusters contain the known Obg andDRG proteins (Okamoto et al., 1997; Maddock et al., 1997;Sazuka et al., 1992), as well as some unrecognizedhomologues. This analysis shows that NOG1, Obg and DRGG regions are closely related to each other, but only distantlyrelated to the Ras family of G-proteins. EF-2 andheterotrimeric G-proteins were even less related. The closeevolutionary relatedness, and the striking conservation offamily-specific E motif and other residues clearly indicate thatObg, DRG and NOG1 are sister groups, forming a novel, largefamily of G-proteins. We propose this family be designated asan ODN for its prototype subfamily constituents.
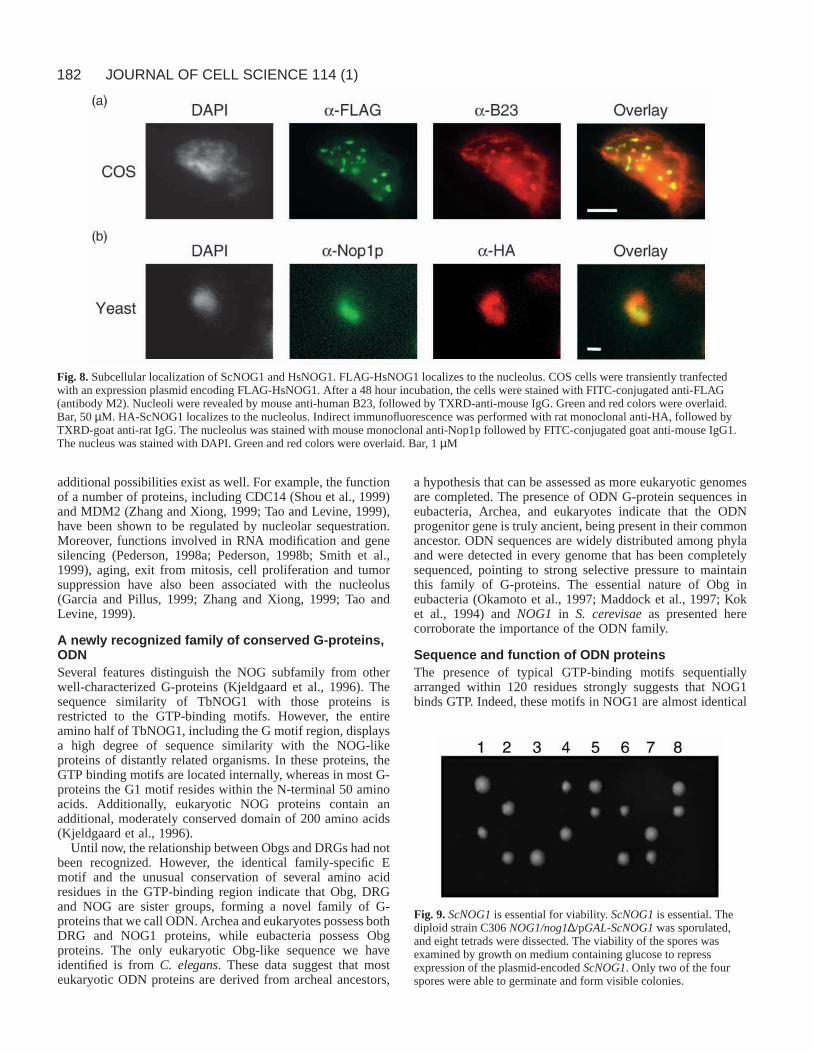
Human and yeast NOG1 homologues localize to thenucleolus Humans possess two predicted proteins with overall homologyto NOG1, whileS. cerevisaeencodes a NOG1-related proteinat the YPL093w locus (named herein ScNOG1). To determinewhether nucleolar localization is a common feature of
Fig. 5. NOG1 expression duringT. bruceidevelopment.Immunoblot analysis wasperformed using lysates preparedfrom procyclic forms (PC),slender bloodstream forms (SL)and stumpy bloodstream forms(ST). The same blots weresuccessively incubated withrabbit anti-TbNOG1 (a) and withrabbit anti-56PGK (b). Controlblots with the preimmune serumshowed no reactivity at similardilutions.
Tb 1 MSTIYNFKTVTVVPSYKDFIDIVLSKTQRKTPTVVHKGYHISRIRQFYMRKVKFTQKTINEKLTHILTEFPRMDDIHPFYGDLMHVLYDRSc 1 --MQLSWKDIPTVAPANDLLDIVLNRTQRKTPTVIRPGFKITRIRAFYMRKVKYTGEGFVEKFEDILKGFPNINDVHPFHRDLMDTLYEKHs1 1 -MAHYNFKKITVVPSAKDFIDLTLSKTQRKTPTVIHKHYQIHRIRHFYMRKVKFTQQNYHDRLSQILTDFPKLDDIHPFYADLMNILYDKHs2 1 -----------------DFIDLTLSKTQRKTPTVIHKHYQIHRIRHFYMRKVKFTQQNYHDRLSQILTDFPKLDDIHPFYADLMNILYDK
Tb 91 DHYKVALGQVGAVRHMVDNVGRDYVRLLKYGDSLYRCKQLKRAALGRMATACKKLTSALVYLEKVRQHMSRLPSIDPNARTLLITGFPNVSc 89 NHYKISLAAISRAKSLVEQVARDYVRLLKFGQSLFQCKQLKRAALGRMATIVKKLRDPLAYLEQVRQHIGRLPSIDPNTRTLLICGYPNVHs1 90 DHYKLALGQINIAKNLVDNVAKDYVRLMKYGDSLYRCKQLKRAALGRMCTVIKRQKQSLEYLEQVRQHLSRLPTIDPNTRTLLLCGYPNVHs2 74 DHYKLALGQINIAKNLVDNVAKDYVRLMKYGDSLYRCKQLKRAALGRMCTIIKRQKQSLEYLEQVRQHLSRLPTIDPNTRTLLLCGYPNV
Tb 181 GKSSFMNKVTRADVEVQPYAFTTKSLFVGHTDFKYASWQVIDTPGILDHSLEERNVIEMQAITALAHLRACILFFMDLSTQCGHSIAQQVSc 179 GKSSFLRCITKSDVDVQPYAFTTKSLYVGHFDYKYLRFQAIDTPGILDRPTEEMNNIEMQSIYAIAHLRSCVLYFMDLSEQCGFTIEAQVHs1 180 GKSSFINKVTRADVDVQPYAFTTKSLFVGHMDYKYLRWQVVDTPGILDHPLEDRNTIEMQAITALAHLRAAVLYVMDLSEQCGHGLREQLHs2 164 GKSSFINKVTRADVDVQPYAFTTKSLFVGHMDYKYLRWQVVDTPGILDHPLEDRNTIEMQAITALAHLRAAVLYVMDLSEQCGHGLKEQL
Tb 271 SLFKSIGPLFTGKPVIVVFNKSDLCTFDDITAEEQSLVMTAIEECGAKWITTSTLTDAGVGDLKTLACETLLAHRSEQKEGS-GRFQAIQSc 269 KLFHSIKPLFANKSVMVVINKTDIIRPEDLDEERAQLLESVKEVPGVEIMTSSCQLEENVMEVRNKACEKLLASRIENKLKSQSRINNVLHs1 270 ELFQNIRPLFINKPLIVVANKCDVKRIAELSEDDQ-KIFTDLQSEGFPVIETSTLTEEGVIKVKTEACDRLLAHRVETKMKG-NKVNEVLHs2 254 ELFQNIRPLFINKPLIVVANKCDVKRITELSEEDQ-KIFTDLQAEGFPVIETSTLTEEGVIQVKTEACDRLLAHRVETKMKG-NKVNEVL
Tb 360 NRLYCAVPQKRDNV-------------------------------------------------------ERPAYVPPSVEEIRQRLASGESc 359 NKIHVAQPQARDDV-------------------------------------------------------KRTPFIPESVKNLKKYDPEDPHs1 358 NRLHLAIPTRRDDK-------------------------------------------------------ERPPFIPEGVVARRKRMETEEHs2 342 NRLHLAVPNKRDDKPMEVFVFVFLRQGFSVALEAVLELALIDQAGLKITEICLPLPPSAGTKGVHHHLPERPPFIPEGVVARRKRMEIVE
Tb 395 PVRERRKTERDYEWENGGPGQYQPNERKTWDLENPEWVDDIIPDIMDGHNIYDNVDPDIHERLMELEAEEDARLQE-----LELEASKKRSc 394 ---NRRKLARDIEAENGGAGVFNVNLKDKYLLEDDEWKNDIMPEILDGKNVYDFLDPEIAAKLQALEEEEEKLENEGFYNSDDEEEIYDGHs1 393 ---SRKKRERDLELEMG--DDYILDLQKYWDLMNLSEKHDKIPEIWEGHNIADYIDPAIMKKLEELEKEEELRTAA-----GEYDSVSESHs2 432 ---PRKKRERDLELEMG--DDYILDLQKYWDLMNSSEKYDKIPEIWEGHNVADYIDPAIMK-----------------------------
Tb 480 PQYELHDSTVEAVRFIRDKIKVIKMERAMKNPSLRRTRRQSVAIEKFNKRTGSQVDRSATPGCSSETTNSSAKRGRSLSAAQEAVMRDRSSc 481 FEASEVDDIKEKAAWIRNRQKTMIAEARNRK-SLKNKAIMPRSKLTKSFGKMEEHMSTLGHDMSALQDKQNRAARKN--R-----YVERGHs1 473 -EDEEMLEIRQLAKQIREKKKLKILESKEKN---TQGPRMPRTAKKVQRTVLEKEMRSLGVDMDDKDDAHYAVQAR----------RSRSHs2 488 ------------AKQIREKKKLKILQSKEKN---TQGPRMPRTAKKVQRADLENEMRSLGVDMDDKDNAHYAVQAR----------RSRS
Tb 570 SSHMSTKTTRGISSHSATRDRSLSVNRGEGYRDVNEKLRAVKLSKVVARPRNLQAKKGEGDRAIPNLRPKHLFTGKVKSNGRRDRRSc 563 SDVVFGDQDALTASTENGVKLRQTDRLLDGVADGSMRSKADRMAKMERRERNRHAKQGESDRHNAVSLSKHLFSGKRG-VGKTDFRHs1 549 ITRKRKREDSAPPSSVARSGSCSRTPRDVSGLRDVKMVKKAKTMKNAQKKMNRLGKKGEADRHVFDMKPKHLLSGKRK-AGKKDRRHs2 553 VTRKRKREES----------------------------------------------------------------------------
G1
G2E
G3
Fig. 6.Alignment of TbNOG1 with predicted eukaryotic homologues. Proteins shown are from human (Hs, accession no. Hs1, 4191616 andHs2, 3153873), and S. cerevisiae(Sc, 2132184). Gaps are indicated by hyphens. Resides identical or conserved in the proteins are marked byblack or grey boxes respectively. The conserved motifs, G1, E, G2 and G3, which comprise the GTP-binding region, are overlined. TheTbNOG1sequence data are available from GenBank/EMBL/DDBJ under accession number AAF01061.

180
eukaryotic NOG proteins, we expressed epitope-taggedversions of ScNOG1 and one of the human proteins (named
herein HsNOG1). FLAG-tagged HsNOG1 was expressedtransiently in COS cells (Fig. 8a). Anti-FLAG staining revealed
JOURNAL OF CELL SCIENCE 114 (1)
Fig. 7.Relationship of NOG, Obg, and DRG families of G-proteins. (a) Multiple alignment of the regions containing GTP-binding motifs.Shading marks where 5/6 residues are conserved or identical in the Obg, DRG, and NOG proteins. Shading was then extended to the Rasprotein where the residues matched. (b) Phylogenetic tree. The regions containing GTP-binding motifs of proteins containing the consensuspattern (G(X)4GKS(X)15YXFTT) were aligned using ClustalX (Thompson et al., 1994) and from this alignment phylogenetic trees werecalculated using the PHYLIP Phylogeny Inference Package version 3.5c (http://evolution.genetics.washington.edu/phylip.html). The unrootedphylogeny shown is the most parsimonious (PROTPARS on default settings with sequence order jumbled ten times). Distance trees using theprogram PRODIST showed similar topology. The tree was drawn with the DRAWTREE program. For clarity, some closely related proteinswere omitted from the tree. Numbers adjacent to selected nodes are bootstrap values, calculated from 100 random replica using the PAUPprogram. The previously known members of Obg and DRG subfamilies are marked by asterisks. The names of the organisms are abbreviated asfollows; Aa, Aquifex aeolicus; Af, A. fulgidus; Bb, Borrelia burgdorferi; Bs, Bacillus subtilis; Cc, Caulobacter crescentus; Ce, C. elegans; Ct,Chlamydia trachomatis; Dm, Drosophila melanogaster; Hc, Halobacterium cutirubrum; Hi, Haemophilus influenza; Hs, Homo sapiens; Mj, M.jannaschii; Mm, Mus musculus; Mt, M. thermoautotrophicum; Mu, Mycobacterium tuberculosis; Ph, Pyrococcus horikoshii; Sg, Streptomycesgriseus; Sm, Streptomyces coelicolor; Ss, Synechocystis sp.; Xl, Xenopus laevis. Accession numbers for Obg proteins shown are: Aa, O67849;Bb, O51722; Bs P20964; Cc, 2555098; Ce, O45691; Ct, O84423; Hi, 1176187; Mu, P71909; Sc, P95722; Sg, P95758; Ss, P72931; Tp,O83724. Accession numbers for DRG proteins are: Af, 2648391; Dm, 416555; Hc,141353; Hs-1, 4127988; Hs-2, 1706518; Mj, 1591967; Mm,346685; Mt, 2622747; Sc-1, 1723727; Sc-2, 731276; Xl, 1169421. Accession numbers for NOG1 proteins are: Ce, 2702365; Mt, 2621950; Mj,2127948; Ph, 3257743; and AF, O29821.

181Nucleolar G-protein
that tagged HsNOG1 was found in several discrete patcheswithin the nucelus. These regions corresponded to the regionsstaining strongly for the nucleolar protein B23. Because B23shuttles between the nucleolus and cytoplasm (Finch et al.,1993), some cytoplasmic staining is also observed. This resultindicates that HsNOG1, like TbNOG1, localizes to thenucleolus.
The coding region of ScNOG1was tagged at its 5′ end withsequences encoding two HA epitopes and the resulting plasmid(pGAL1-HA-ScNOG1) was transformed into diploid yeast.Following galactose induction for two hours, the fluorescencepattern of HA-ScNOG1 shows the characteristic nucleolarcrescent shape at the edge of the nucleus (Fig. 8b). The stainingoverlaps that of the nucleolar marker Nop1p (Aris and Blobel,1988), albeit being fainter and perhaps slightly more diffuse.Upon prolonged induction, HA-ScNOG1 was also found in thenucleus and, eventually, cytoplasm. No staining was seenwithout induction. These data indicate that ScNOG1 isnucleolar when expressed at low levels.
ScNOG1 is an essential geneWe examined the effect of disruption of the NOG1gene in S.cerevisae,an organism more amenable to genetic manipulationthan trypanosomes. One copy of ScNOG1in the diploid strainC306 was replaced with the KanMX marker module, whichconfers resistance to G418, to generate the strain C306NOG1/nog1∆. A plasmid expressing ScNOG1 under thecontrol of a GAL1 promoter was then transformed into thisstrain. The transformants were sporulated, and dissected ontomedium containing glucose to repress expression of plasmid-encoded ScNOG1. Subsequent analysis of the dissected tetradsgrown on glucose showed a 2:2 segregation of viable tononviable spores (Fig. 9). Microscopy revealed that the deadspores were able to germinate and divide only once. Thehaploid colonies failed to grow on medium containing G418indicating that they received the wild-type gene and that thedead spores must have received the nog1::KanMx deletionallele. These data are consistent with a recently releasedgenome-based analysis that suggested that ScNOG1is essential(Winzeler et al., 1999). When tetrads were dissected onmedium containing galactose to induce expression of plasmidborne ScNOG1, the tetrads yielded either three or four viablecolonies. This rescue indicates the loss of viability was causedby disruption of ScNOG1gene. Thus ScNOG1is an essentialgene.
The predicted G motifs are required for ScNOG1functionTo assess the functional significance of the signature GTP-binding motifs, we tested the ability of ScNOG1 mutated ineither the G1 or G3 motifs to complement the ScNOG1deletion. In the G1 mutant, the conserved residues GKS (aa179-181) were mutated to AAA. In the G3 mutant, theconserved residues KTD (279 to 281) were mutated to ATL.The plasmids were transformed into the diploid C306NOG1/nog1∆ and the expression of the wild-type and mutantHA-tagged proteins was verified by immunoblot analysis usinganti-HA (Fig. 10a). The wild-type tagged ScNOG1 and G1mutant were expressed at similar levels, while expression of theG3 mutant protein was somewhat lower. Immunolocalizationusing anti-HA demonstrated that, like wild-type ScNOG1, the
G1 mutant protein resided in the nucleolus (Fig. 10b). Thus themutant protein was functional with respect to localization.
The diploids containing the mutated ScNOG1genes weresporulated and individual haploid segregants were replicaplated onto medium plus and minus G418. Only segregantswith the chromosomal nog1 deletion (nog1::KanMx) cansurvive the drug. To survive, cells must also have a (plasmid-encoded) functional NOG1gene. As shown in Table 3, colonieswith the genotype C306 nog1∆ that contained the wild-typeHA-ScNOG1plasmids were viable, but no colonies wereobtained when we attempted to cover the deletion with theG1 (Scnog1-g1) or G3 (Scnog1-g3) mutant constructs. Thusmutation of the G1 or G3 motifs abrogates NOG1 function,strongly suggesting that GTP/GDP binding is a requiredfunction of this molecule. The recovery of colonies bearing thewild-type construct was lower than theoretically expected,suggesting that the plasmid borne HA-ScNOG1 may not becompletely equivalent to the wild-type chromosomal copy.Such a finding could be due to the presence of the tag or toaltered expression levels resulting from the use of the GAL1promoter.
DISCUSSION
Interaction of NOPP44/46 and a nucleolar G-protein,NOG1T. bruceiNOPP44/46 does not show overall homology to anyknown protein, yet each of its domains has significantsimilarity with modules that participate in intermolecularinteractions in other nonribosomal nucleolar proteins (Tutejaand Tuteja, 1998; Zatsepina et al., 1999; Dundr and Olson,1998; Gustafson et al., 1998). Due to the presence of thesedomains and the relatively high abundance of NOPP44/46, wepreviously hypothesized that this protein provides a dockingsite that brings other nucleolar proteins into proximity. To gaininsight into this possibility, we searched for proteins thatdirectly interact with NOPP44/46 using the two-hybrid system.One of the proteins thus identified was TbNOG1, a nucleolarpredicted G-protein. A search of the literature indicates oneother potential nucleolar G-protein. This protein, encoded bythe human Ngp-1 gene, has the typical G1 and G2 motifs(Racevskis et al., 1996). However, the identified G3 motif isamino to the G1 motif, a highly unusual situation.
Using the two-hybrid system and coimmunoprecipitationstudies, we demonstrated that sites within the G-domain andC-terminal extension of TbNOG1 interact with the U and Jregions of NOPP44/46 respectively. As noted above, sequencesrelated to the U and J regions are present in a number ofnucleolar proteins. We speculate that proteins with U- or J-likedomains may interact with their corresponding NOG1homologues. The multiple binding sites for conserved domainsof nucleolar proteins raise the possibility that some but not allinteractions with NOG1 are modulated by the GDP/GTP ratio.Our two-hybrid analysis of the Tbnog1-g1 mutant proteinsuggests that the interaction with NOPP44/46 is not fullyrestricted to the GTP-bound form of NOG1. This findingsuggests that the interaction between these two proteins maybe of a constitutive nature or may be more subtly altered bythe ratio of GTP to GDP. NOG1’s interactions with otherproteins may be directly related to ribosome biogenesis, but
182
additional possibilities exist as well. For example, the functionof a number of proteins, including CDC14 (Shou et al., 1999)and MDM2 (Zhang and Xiong, 1999; Tao and Levine, 1999),have been shown to be regulated by nucleolar sequestration.Moreover, functions involved in RNA modification and genesilencing (Pederson, 1998a; Pederson, 1998b; Smith et al.,1999), aging, exit from mitosis, cell proliferation and tumorsuppression have also been associated with the nucleolus(Garcia and Pillus, 1999; Zhang and Xiong, 1999; Tao andLevine, 1999).
A newly recognized family of conserved G-proteins,ODNSeveral features distinguish the NOG subfamily from otherwell-characterized G-proteins (Kjeldgaard et al., 1996). Thesequence similarity of TbNOG1 with those proteins isrestricted to the GTP-binding motifs. However, the entireamino half of TbNOG1, including the G motif region, displaysa high degree of sequence similarity with the NOG-likeproteins of distantly related organisms. In these proteins, theGTP binding motifs are located internally, whereas in most G-proteins the G1 motif resides within the N-terminal 50 aminoacids. Additionally, eukaryotic NOG proteins contain anadditional, moderately conserved domain of 200 amino acids(Kjeldgaard et al., 1996).
Until now, the relationship between Obgs and DRGs had notbeen recognized. However, the identical family-specific Emotif and the unusual conservation of several amino acidresidues in the GTP-binding region indicate that Obg, DRGand NOG are sister groups, forming a novel family of G-proteins that we call ODN. Archea and eukaryotes possess bothDRG and NOG1 proteins, while eubacteria possess Obgproteins. The only eukaryotic Obg-like sequence we haveidentified is from C. elegans. These data suggest that mosteukaryotic ODN proteins are derived from archeal ancestors,
a hypothesis that can be assessed as more eukaryotic genomesare completed. The presence of ODN G-protein sequences ineubacteria, Archea, and eukaryotes indicate that the ODNprogenitor gene is truly ancient, being present in their commonancestor. ODN sequences are widely distributed among phylaand were detected in every genome that has been completelysequenced, pointing to strong selective pressure to maintainthis family of G-proteins. The essential nature of Obg ineubacteria (Okamoto et al., 1997; Maddock et al., 1997; Koket al., 1994) and NOG1 in S. cerevisaeas presented herecorroborate the importance of the ODN family.
Sequence and function of ODN proteinsThe presence of typical GTP-binding motifs sequentiallyarranged within 120 residues strongly suggests that NOG1binds GTP. Indeed, these motifs in NOG1 are almost identical
JOURNAL OF CELL SCIENCE 114 (1)
Fig. 8.Subcellular localization of ScNOG1 and HsNOG1. FLAG-HsNOG1 localizes to the nucleolus. COS cells were transiently tranfectedwith an expression plasmid encoding FLAG-HsNOG1. After a 48 hour incubation, the cells were stained with FITC-conjugated anti-FLAG(antibody M2). Nucleoli were revealed by mouse anti-human B23, followed by TXRD-anti-mouse IgG. Green and red colors were overlaid.Bar, 50 µM. HA-ScNOG1 localizes to the nucleolus. Indirect immunofluorescence was performed with rat monoclonal anti-HA, followed byTXRD-goat anti-rat IgG. The nucleolus was stained with mouse monoclonal anti-Nop1p followed by FITC-conjugated goat anti-mouse IgG1.The nucleus was stained with DAPI. Green and red colors were overlaid. Bar, 1 µM
Fig. 9. ScNOG1is essential for viability. ScNOG1is essential. Thediploid strain C306 NOG1/nog1∆/pGAL-ScNOG1was sporulated,and eight tetrads were dissected. The viability of the spores wasexamined by growth on medium containing glucose to repressexpression of the plasmid-encoded ScNOG1. Only two of the fourspores were able to germinate and form visible colonies.

183Nucleolar G-protein
to those in Bacillus subtilis (Fig. 7) and CaulobactercrescentusObg-like proteins, the only ODN family membersthat have been shown biochemically to bind GTP (Lin et al.,1999; Welsh et al., 1994). Attempts to express soluble NOG1in E. coli under a variety of conditions were unsuccessful,precluding detailed biochemical studies and [32P]GTP geloverlay assays are unsuitable for larger G-proteins (Huber andPeter, 1994). However, our analysis of mutants in the G1 andG3 motifs of ScNOG1 shows that these motifs are essential forfunctional complementation, buttressing the argument thatNOG1 function involves GTP/GDP binding.
The ODN family of G-proteins shows specific, uniquesequence characteristics. All members possess a Pro residuewithin the G1 motif (Fig. 7a), corresponding to Gly-12 in H-Ras. Gly-12 of Ras is mutated in several types of humancancers (Hamer et al., 1991) and interestingly, proline is theonly substitution that is not oncogenic (Seeburg et al., 1984).Conversely, in Streptomyces coelicolor, an Obg with a Pro to
Val mutation does not support cell viability (Okamoto andOchi, 1998). The substitution of Gln-61 of the G2 motif withLeu in Ras family of G-proteins (e.g. RanQ69L and RhoQ63L)abolishes intrinsic GTPase activity (Bischoff et al., 1994;Renshaw et al., 1996). All the members of the ODN familycontain Leu or Ile at this position, implying that low GTPaseactivity seen in the Obgs of B. subtilisand C. crescentus(Welshet al., 1994; Lin et al., 1999) may be a characteristic of thisfamily. These two Obgs also have low affinity for GTPcompared to Ras-like G proteins (micromolar versusnanomolar) and exchange is several logs faster.
Recently, Obg has been shown to be required for stressactivation of σB, a transcription factor that enables expressionof theB. subtilis stress regulon (Scott and Haldenwang, 1999)and to associate with ribosomes in this organism (Scott et al.,2000). Additionally, Obg is necessary for the transition fromvegetative growth to sporulation in B. subtilisand S. coelicolor(Okamoto et al., 1997; Kok et al., 1994). Since fluctuations incellular GTP content are thought to be the primary initiator ofmorphological development in these cells (Okamoto et al.,1997; Kok et al., 1994), it has been proposed that Obg acts asa sensor for intracellular GTP/GDP ratio and thereby for cellgrowth status set by nutrient availability. This hypothesis iscompatible with the biochemical analysis of Obg, whichindicates that the molecule has characteristics expected for adirect sensor: fast exchange, similar micromolar affinities forGTP and GDP, and a low rate of hydrolysis. Intracellular GTPlevels are linked to growth in multiple ways. For example, inbacteria, promoter activity is the rate-limiting step in ribosomesynthesis (Nomura et al., 1984; Gaal et al., 1997) and efficientinitiation requires a high concentration of GTP (Gaal et al.,1997). In eukaryotes, the cellular GTP levels have also beenthought to be responsible for the regulation of rRNA synthesis(Grummt and Grummt, 1976). Additionally, localization ofNO38/B23 into the nucleolus is dependent on the cellular GTPlevels (Finch et al., 1993). These observations raise intriguingpossibilities for the role of a nucleolar G protein such asNOG1, which could sense the cellular energy status and usethat information to aid in the integration of nucleolar functions.It is interesting to recall that the class II transcriptionaltransactivator protein CIITA of mammals is also a GTP-binding protein with low hydrolysis rates (Harton et al., 1999).
Trypanosome, yeast and human NOG1 proteins are
Fig. 10. Expression and localization of mutant ScNOG1 proteins.Immunoblot analysis. Lysates from equivalent numbers of uninducedcells or cells induced overnight in galactose plus raffinose mediumwere examined by immunoblot analysis using monoclonal anti-HA.Bound antibodies were revealed with goat anti-mouse IgG usingchemiluminescence (NEN). The strains contain empty vector (Vec)or plasmids specifying HA-tagged wild-type ScNOG1 (WT),Scnog1-g1 (G1) or Scnog1-g3 (G3). The Scnog1-g1 mutant proteinlocalizes to the nucleolus. Cells containing plasmids encoding HA-tagged wild-type ScNOG1 and Scnog1-g1 were induced for 2.5hours on galactose plus raffinose. The tagged protein was revealed byimmunofluorescence using anti-HA plus FITC-conjugated goat anti-mouse IgG. Cells were counterstained with DAPI. Bar, 1 µM.
Table 3. ScNOG1genes with mutations in the G-motifs failto complementColonies Growth on
Plasmid tested G418 P*
ScNOG1 527 34I.‡ Scnog1-g1 287 0II. Scnog1-g1 228 0
Total 515 0 <0.0002
I. Scnog1-g3 288 0II. Scnog1-g3 228 0
Total 516 0 <0.0002
Vector 527 0 <0.0002
*The probability that the proportion of colonies growing on G418 isdifferent from that for the wild-type gene.
‡I and II are independent sets of transformants for each plasmid.

184
localized to the nucleolus of their respective organisms, clearlyimplying that nucleolar localization is a common characteristicof these proteins in eukaryotes. The findings that NOG1proteins are evolutionarily conserved and that ScNOG1 isessential for viability suggest that these proteins may play acrucial role in nucleolar functions. A global survey of yeastmRNA expression shows that NOG1transcript levels decreasedramatically as the cells switch from glycolytic to fermentativemetabolism (DeRisi et al., 1997). This shift is accompanied bya major decrease in transcription of genes involved in ribosomebiogenesis (Neuman-Silberberg et al., 1995). We are currentlyexploring the effects of NOG1 on rDNA transcription andrRNA processing in S. cerevisae.
We thank Drs E. Wirtz and G. Cross for providing pLew79 and 29-13 cells, Dr R. Brent for providing two-hybrid vectors and strains, andDrs U. Goeringer and A. Souza for providing the T. bruceigenomictwo-hybrid library. We are grateful to Dr Q. Wang for testing theinteractions of Tbnog1-g1. We also thank Drs J. Aris and P. Chan forantibodies, Drs S. Yarfitz and B. Hedlund for assistance inbioinformatics, and Dr Nicole Doria-Rose for assistance with COScell transformation. This work was supported in part by NIH AI31077and NIH S10 RR11865-01. B.J. was supported by NIH 1T32AI07509-01.
REFERENCES
Altschul, S. F., Madden, T. L., Schaffer, A. S., Zhang, J., Zhang, Z., Miller,W. and Lipman, D. J. (1997). Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucl. Acids Res. 25, 3389-3402.
Anderson, S. A., Carter, V., Hagen, C. B. and Parsons, M. (1998).Molecular cloning of the glycosomal malate dehydrogenase ofTrypanosoma brucei. Mol. Biochem. Parasitol. 96, 185-189.
Aris, J. P. and Blobel, G. (1988). Identification and characterization of a yeastnucleolar protein that is similar to a rat liver nucleolar protein. J. Cell Biol107, 17-31.
Bischoff, F. R., Klebe, C., Kretschmer, J., Wittinghofer, A. and Ponstingl,H. (1994). RanGAP1 induces GTPase activity of nuclear Ras-related Ran.Proc. Nat. Acad. Sci. USA 91, 2587-2591.
Bourne, H. R., Sanders, D. A. and McCormick, F. (1991). The GTPasesuperfamily: conserved structure and molecular mechanism. Nature 349,117-127.
Bouvet, P., Diaz, J. J., Kindbeiter, K., Madjar, J. J. and Amalric, F. (1998).Nucleolin interacts with several ribosomal proteins through its RGGdomain. J. Biol Chem. 273, 19025-19029.
Das, A., Peterson, G. C., Kanner, S. B., Frevert, U. and Parsons, M. (1996).A major tyrosine-phosphorylated protein of Trypanosoma bruceiis anucleolar RNA-binding protein. J. Biol. Chem. 271, 15675-15681.
Das, A., Park, J. -H., Hagen, C. B. and Parsons, M. (1998). Distinct domainsof a nucleolar protein mediate protein kinase binding, interaction withnucleic acids and nucleolar localization. J. Cell Sci. 111, 2615-2623.
DeRisi, J. L., Iyer, V. R. and Brown, P. O. (1997). Exploring the metabolicand genetic control of gene expression on a genomic scale. Science 278,680-686.
Dundr, M. and Olson, M. O. (1998). Partially processed pre-rRNA ispreserved in association with processing components in nucleolus-derivedfoci during mitosis. Mol. Biol. Cell 9, 2407-2422.
Estojak, J., Brent, R. and Golemis, E. A. (1995). Correlation of two-hybridaffinity data with in vitro measurements. Mol. Cell. Biol. 15, 5820-5829.
Finch, R. A., Revankar, G. R. and Chan, P. K. (1993). Nucleolar localizationof nucleophosmin/B23 requires GTP. J. Biol Chem. 268, 5823-5827.
Gaal, T., Bartlett, M. S., Ross, W., Turnbough, C. L. J. and Gourse, R. L.(1997). Transcription regulation by initiating NTP concentration: rRNAsynthesis in bacteria [see comments]. Science 278, 2092-2097.
Garcia, S. N. and Pillus, L. (1999). Net results of nucleolar dynamics. Cell97, 825-828.
Gietz, D., St, Woods, R. A. and Schiestl, R. H. (1992). Improved method for
high efficiency transformation of intact yeast cells. Nucl. Acids Res. 20,1425-1429.
Ginisty, H., Sicard, H., Roger, B. and Bouvet, P. (1999). Structure andfunctions of nucleolin. J. Cell Sci. 112, 761-772.
Grummt, I. and Grummt, F. (1976). Control of nucleolar RNA synthesis bythe intracellular pool sizes of ATP and GTP. Cell 7, 447-453.
Gustafson, W. C., Taylor, C. W., Valdez, B. C., Henning, D., Phippard, A.,Ren, Y., Busch, H. and Durban, E. (1998). Nucleolar protein p120contains an arginine-rich domain that binds to ribosomal RNA. Biochem J.331, 387-393.
Hamer, P. J., La Vecchio, J., Ng, S., DeLellis, R., Wolfe, H. and Carney,W. P. (1991). Activated Val-12 ras p21 in cell culture fluids and mouseplasma. Oncogene 6, 1609-1615.
Harton, J. A., Cressman, D. E., Chin, K. C., Der, C. J. and Ting, J. P.(1999). GTP binding by class II transactivator: role in nuclear import.Science 285, 1402-1405.
Heine, M. A., Rankin, M. L. and DiMario, P. J. (1993). The gly/arg-rich(GAR) domain of Xenopusnucleolin facilitates in vitro nucleic acid bindingand in vivo nucleolar localization. Mol. Biol. Cell 4, 1189-1204.
Henikoff, S. (1987). Unidirectional digestion with exonuclease III in DNAsequence analysis. Meth. Enzymol. 155, 156-165.
Huber, L. A. and Peter, M. E. (1994). Mapping small GTP-binding proteinson high-resolution two-dimensional gels by a combination of GTP bindingand labeling with in situ periodate-oxidized GTP. Electrophoresis 15, 283-288.
Kjeldgaard, M., Nyborg, J. and Clark, B. F. (1996). The GTP binding motif:variations on a theme. FASEB J. 10, 1347-1368.
Kleinschmidt, J. A., Fortkamp, E., Krohne, G., Zentgraf, H. and Franke,W. W. (1985). Co-existence of two different types of soluble histonecomplexes in nuclei of Xenopus laevisoocytes. J. Biol Chem. 260, 1166-1176.
Kok, J., Trach, K. A. and Hoch, J. A. (1994). Effects on Bacillus subtilisofa conditional lethal mutation in the essential GTP-binding protein Obg. J.Bacteriol. 176, 7155-7160.
Li, D., Dobrowolska, G. and Krebs, E. G. (1996). The physical associationof casein kinase 2 with nucleolin. J. Biol. Chem. 271, 15662-15668.
Lin, B., Covalle, K. L. and Maddock, J. R. (1999). The CaulobactercrescentusCgtA protein displays unusual guanine nucleotide binding andexchange properties. J. Bacteriol. 181, 5825-5832.
Lusser, A., Brosch, G., Loidl, A., Haas, H. and Loidl, P. (1997).Identification of maize histone deacetylase HD2 as an acidic nucleolarphosphoprotein. Science 277, 88-91.
Maddock, J., Bhatt, A., Koch, M. and Skidmore, J. (1997). Identificationof an essential Caulobacter crescentusgene encoding a member of the Obgfamily of GTP-binding proteins. J. Bacteriol. 179, 6426-6431.
Mumberg, D., Muller, R. and Funk, M. (1994). Regulatable promoters ofSaccharomyces cerevisiae: comparison of transcriptional activity and theiruse for heterologous expression. Nucl. Acids Res. 22, 5767-5768.
Nakai, K. and Horton, P. (1999). PSORT: a program for detecting sortingsignals in proteins and predicting their subcellular localization. TrendsBiochem Sci. 24, 34-36.
Neuman-Silberberg, F. S., Bhattacharya, S. and Broach, J. R. (1995).Nutrient availability and the RAS/cyclic AMP pathway both induceexpression of ribosomal protein genes in Saccharomyces cerevisiaebut bydifferent mechanisms. Mol. Cell. Biol. 15, 3187-3196.
Nomura, M., Gourse, R. and Baughman, G. (1984). Regulation of thesynthesis of ribosomes and ribosomal components. Annu. Rev. Biochem53:75-117, 75-117.
Okamoto, S., Itoh, M. and Ochi, K. (1997). Molecular cloning andcharacterization of the obg gene of Streptomyces griseusin relation to theonset of morphological differentiation. J. Bacteriol. 179, 170-179.
Okamoto, S. and Ochi, K. (1998). An essential GTP-binding proteinfunctions as a regulator for differentiation in Streptomyces coelicolor. MolMicrobiol. 30, 107-119.
Parker, W. L., Hill, T., Alexander, K., Murphy, N. B., Fish, W. R. andParsons, M. (1995). Three genes and two isozymes: gene conversion andthe compartmentalization and expression of the phosphoglycerate kinasesof Trypanosoma (Nannomonas) congolense. Mol. Biochem. Parasitol. 69,269-279.
Parsons, M., Nelson, R. G., Watkins, K. P. and Agabian, N. (1984).Trypanosome mRNAs share a common 5′ spliced leader sequence. Cell 38,309-316.
Parsons, M., Ledbetter, J. A., Schieven, G. L., Nel, A. E. and Kanner, S.B. (1994). Developmental regulation of pp44/46, tyrosine-phosphorylated
JOURNAL OF CELL SCIENCE 114 (1)

185Nucleolar G-protein
proteins associated with tyrosine/serine kinase activity in Trypanosomabrucei. Mol. Biochem. Parasitol. 63, 69-78.
Pederson, T. (1998a). Growth factors in the nucleolus? J. Cell Biol 143, 279-281.
Pederson, T. (1998b). The plurifunctional nucleolus. Nucl. Acids Res. 26,3871-3876.
Racevskis, J., Dill, A., Stockert, R. and Fineberg, S. (1996). Cloning of anovel nucleolar guanosine-5′-triphosphate binding protein autoantigen froma breast tumor. Cell Growth Differ. 7, 271-280.
Renshaw, M. W., Toksoz, D. and Schwartz, M. A. (1996). Involvement ofthe small GTPase rho in integrin-mediated activation of mitogen-activatedprotein kinase. J. Biol. Chem. 271, 21691-21694.
Sazuka, T., Tomooka, Y., Ikawa, Y., Noda, M. and Kumar, S. (1992). DRG:a novel developmentally regulated GTP-binding protein. Biochem. Biophys.Res. Commun. 189, 363-370.
Scott, J. M. and Haldenwang, W. G. (1999). Obg, an essential GTP bindingprotein of Bacillus subtilis, is necessary for stress activation of transcriptionfactor σB. J. Bacteriol. 181, 4653-4660.
Scott, J. M., Ju, J., Mitchell, T. and Haldenwang, W. G. (2000). The Bacillussubtilis GTP binding protein Obg and regulators of the σB stress responsetranscription factor cofractionate with ribosomes. J. Bacteriol. 182, 2771-2777.
Seeburg, P. H., Colby, W. W., Capon, D. J., Goeddel, D. V. and Levinson,A. D. (1984). Biological properties of human c-Ha-ras1 genes mutated atcodon 12. Nature 312, 71-75.
Sherman, F., Fink, G. R. and Hicks, J. B. (1981). Methods in Yeast Genetics.Cold Spring Harbor Laboratory Press, Cold Spring Harbor.
Shou, W., Seol, J. H., Shevchenko, A., Baskerville, C., Moazed, D., Chen,Z. W., Jang, J., Charbonneau, H. and Deshaies, R. J. (1999). Exit frommitosis is triggered by Tem1-dependent release of the protein phosphataseCdc14 from nucleolar RENT complex P19(ARF) stabilizes p53 by blockingnucleo-cytoplasmic shuttling of Mdm2. Cell 97, 233-244.
Smith, J. S., Caputo, E. and Boeke, J. D. (1999). A genetic screen forribosomal DNA silencing defects identifies multiple DNA replication andchromatin-modulating factors. Mol. Cell. Biol. 19, 3184-3197.
Sprang, S. R. (1997). G protein mechanisms: insights from structural analysis.Annu. Rev. Biochem. 66, 639-678.
Tao, W. and Levine, A. J. (1999). P19(ARF) stabilizes p53 by blocking
nucleo-cytoplasmic shuttling of Mdm2. Proc. Nat. Acad. Sci. USA 96, 6937-6941.
Thompson, J. D., Higgins, D. G. and Gibson, T. J. (1994). CLUSTAL W:improving the sensitivity of progressive multiple sequence alignmentthrough sequence weighting, position-specific gap penalties and weightmatrix choice. Nucl. Acids Res. 22, 4673-4680.
Tuteja, R. and Tuteja, N. (1998). Nucleolin: a multifunctional majornucleolar phosphoprotein. Crit. Rev. Biochem. Mol. Biol. 33, 407-436.
Walker, J. E., Saraste, M., Runswick, M. J. and Gay, N. J. (1982). Distantlyrelated sequences in the alpha- and beta-subunits of ATP synthase, myosin,kinases and other ATP-requiring enzymes and a common nucleotide bindingfold. EMBO J. 1, 945-951.
Welsh, K. M., Trach, K. A., Folger, C. and Hoch, J. A. (1994). Biochemicalcharacterization of the essential GTP-binding protein Obg of Bacillussubtilis. J. Bacteriol. 176, 7161-7168.
Winey, M., Goetsch, L., Baum, P. and Byers, B. (1991). MPS1and MPS2:novel yeast genes defining distinct steps of spindle pole body duplication.J. Cell Biol. 114, 745-754.
Winzeler, E. A., Shoemaker, D. D., Astromoff, A., Liang, H., Anderson,K., Andre, B., Bangham, R., Benito, R., Boeke, J. D., Bussey, H., Chu,A. M., Connelly, C., Davis, K., Dietrich, F., Dow, S. W., El Bakkoury,M., Foury, F., Friend, S. H., Gentalen, E., Giaever, G., Hegemann, J. H.,Jones, T., Laub, M., Liao, H. and Davis, R. W. (1999). Functionalcharacterization of the S. cerevisiaegenome by gene deletion and parallelanalysis. Science 285, 901-906.
Wirtz, E., Leal, S., Ochatt, C. and Cross, G. A. (1999). A tightly regulatedinducible expression system for conditional gene knock-outs and dominant-negative genetics in Trypanosoma brucei. Mol. Biochem. Parasitol. 99, 89-101.
Zatsepina, O. V., Rousselet, A., Chan, P. K., Olson, M. O., Jordan, E. G.and Bornens, M. (1999). The nucleolar phosphoprotein B23 redistributesin part to the spindle poles during mitosis. J. Cell Sci. 112, 455-466.
Zhang, J., Ruyechan, W. and Williams, N.(1998). Developmental regulationof two nuclear RNA binding proteins, p34 and p37, from Trypanosomabrucei. Mol. Biochem. Parasitol. 92, 79-88.
Zhang, Y. and Xiong, Y. (1999). Mutations in human ARF exon 2 disrupt itsnucleolar localization and impair its ability to block nuclear export ofMDM2 and p53. Mol. Cell 3, 579-591.





























